
Submitted:
13 May 2025
Posted:
14 May 2025
You are already at the latest version
Abstract
The growing demand for efficient sustainable biocatalysts for industrial applications has driven the exploration of extremozymes from extremophiles, particularly those thriving in geothermal environments. This study aimed to isolate and characterize extracellular amylase-producing thermophilic bacteria from the Nelumwewa geothermal spring in Sri Lanka, an underexplored ecosystem. Among the isolated thermophilic bacterial strains, NW2 isolates exhibited a prominent extracellular amylase activity. Molecular characterization by 16S rRNA gene sequencing confirmed the close phylogenetic relationship between NW2 and Thermaerobacillus caldiproteolyticus which is well-known for thermostable proteases. Biochemical assays revealed optimal amylase activity of NW2 isolate at 60 ° C and pH 8.0, with a crude enzyme activity of 0.85 U/mL. The enzyme demonstrated efficient hydrolysis of raw cassava starch, highlighting its potential for industrial applications in food, biofuel, and detergent industries. This study reports the first T. caldiproteolyticus-like strain from Sri Lanka with significant extracellular amylase activity, emphasizing the biotechnological potential of geothermal springs as sources of novel extremozymes. These findings contribute to the growing repository of thermostable enzymes highlighting the need for further exploration of Sri Lanka’s geothermal microbial diversity for industrial biocatalysts.
Keywords:
thermophilic bacteria
; thermostable amylase
; geothermal springs
; Nelumwewa geothermal spring
; Thermaerobacillus caldiproteolyticus
1. Introduction
Many industrial processes use large amounts of expensive and hazardous synthetic chemical catalysts, which pose serious risks to the environment and human health. In response, biocatalysts are receiving a greater attention as sustainable and eco-friendly alternatives with superior selectivity, efficiency, and cost-effectiveness [1,2]. Microbial extremozymes are a unique class of biocatalysts that exhibited remarkable stability and catalytic efficiency under extreme environmental conditions, such as elevated temperatures, extreme pH, and high salinity [3,4,5]. Among the various extremozymes, thermozymes produced by thermophilic bacteria are of particular interest in modern biotechnological industries due to their high thermostability, which is essential for maintaining enzymatic activity under the harsh conditions of industrial processes [6]. Thermophilic bacteria are broadly classified based on their optimal growth temperatures into moderate (optimal growth 45–80 °C) and extreme (optimal growth > 80 °C) types [7,8]. Bacterial thermozymes, such as amylases, proteases, lipases, polymerases, cellulases, and chitinases have gained significant industrial importance due to their efficient activity under extreme conditions [6].
Among microbial extremozymes, thermostable amylases are of commercial significance in various industries representing approximately 25% of the global enzyme market. These amylases hydrolyze starch at elevated temperatures, eliminating the need for energy-intensive cooling systems contributing to substantial reductions in processing costs [9]. They offer promising operational advantages across a range of industrial sectors, including food processing, pharmaceutical, pulp and paper, textile manufacturing, detergent, biofuel production, bioremediation, and waste management [6,10,11]. Thermostable amylases are primarily produced for industrial applications by several well-characterized bacterial genera, such as Bacillus [12], and Anoxybacillus [13,14,15] due to their high enzymatic yield, remarkable thermostability, and scalability. Moreover, numerous studies have been conducted on the isolation and characterization of bacterial species that produce thermostable amylases from diverse ecological niches. Thermostable amylases have been identified in species, such as Streptomyces erumpens, Thermobifida fusca, and Bacillus strains isolated from soil environments [16,17,18,19]. In the ongoing search for excellent thermostable amylases for industrial applications, several researchers have successfully isolated and characterized thermostable amylases from thermostable bacteria thrived in thermal spring environments [15,20,21,22,23,24].
Despite significant advancements in identification of navel extremophiles for industrial applications, the geothermal ecosystems of Sri Lanka remain largely underexplored. A study has been reported on exploring the microbial diversity of a geothermal spring of Sri Lankan [25], However, this study did not report the presence of well-known thermostable amylase producing bacteria. This highlights a significant gap in the understanding of Sri Lanka’s geothermal microbial diversity in the context of industrially important extremozymes. Therefore, the present study was conducted with the objective of isolation and molecular characterization of thermophilic bacteria producing extracellular amylases from Sri Lankan geothermal springs, and assessing their potential for industrial applications.
2. Materials and Methods
2.1. Water Sample Collection
Water samples were collected into sterile vacuum flasks from Nelumwewa geothermal spring (7°53′28.0″N 81°11′58.2″E) in Polonnaruwa district of Sri Lanka (Figure 1). Water temperature and pH of the hot water spring were recorded. The water samples were immediately transferred to the laboratory for thermophilic bacterial isolation.
2.2. Isolation of Thermophilic Bacteria from Water Samples
An aliquot of 2 mL from each water sample was inoculated separately into 100 mL of nutrient broth (NB) and incubated overnight at 55 °C with shaking at 150 rpm for enrichment. Then, the incubated samples were serially diluted and 100 µl from each sample, was spread on nutrient agar plates separately. The bacterial colonies with different morphology were carefully selected and pure cultures were isolated on nutrient agar plates incubating at 55 °C overnight. The pure cultures were prepared by streaking and subculturing and maintained on nutrient agar plates at 4 °C. For the long-term storage, 20% glycerol stocks were prepared from each pure culture and stored at -80 °C. Gram’s staining was conducted to differentiate Gram-positive and Gram-negative bacterial isolates.
2.3. Catalase Test
To identify the catalase producing bacteria, small amount of each bacterial isolate was place on clean glass slides, separately. A drop of 3% hydrogen peroxide solution was added to each bacterial sample and formation of bubbles was observed. The bacterial isolates that gave bubbles were identified to be catalase producing bacteria [27].
2.4. Screening of Thermophilic Bacterial Isolates for Extracellular Amylase Production
Extracellular amylase producing bacterial isolates were screened by growing pure isolates on starch agar plates (Peptone 5 g/L, NaCl 5 g/L, yeast extract 1.5 g/L, beef extract 1.5 g/L, soluble starch 2 g/L, agar15 g/L, pH 7.5) at 55 °C overnight. Then the plates were flooded with iodine solution (2% I2 and 0.2% KI) for 5 minutes and iodine solution was decanted. The isolates that gave clear zone around the colonies were selected and sub-cultured to maintain pure cultures and they were identified as extracellular amylase producing thermophilic bacteria [28].
2.5. Screening of Thermophilic Bacterial Isolates for Extracellular Protease Production
All the bacterial isolates were screened for extracellular protease by growing pure isolates on skim milk agar plates (5.0 g/L peptone, 3.0 g/L yeast extract, 1.0 g/L skim milk powder, pH 7.5) at 55 °C overnight. The thermophilic bacterial isolates that produce extracellular protease were identified by observing the clear zone around the bacterial colonies due to the proteolytic activity.
2.6. Molecular Characterization of Isolated Thermophilic Bacteria by 16S rRNA Gene Sequencing and Phylogenetic Analysis
For 16S rRNA gene bidirectional sequencing with 27F (5′-AGTTTGATCMTGGCTCAG-3′) and 1492R (5′- TACGGCTACCTTGTTACGACTT -3′) universal primer pair, all the six pure isolates were sent to Macrogen, Inc. South Korea through Genetech Sri Lanka Pvt. Ltd. Colombo. The consensus sequences were generated by BioEdit version 7. To identify the thermophilic bacterial isolates, all the sequences were blasted against NCBI rRNA/ITS databases (https://blast.ncbi.nlm.nih.gov/Blast.cgi). The partial 16S rRNA gene sequences of isolated thermophilic bacteria were deposited in NCBI GenBank under the accession numbers PV489066, PV489057, and PV489038. Phylogenetic analysis was conducted using MEGA software version 12 [29]. The phylogenetic tree was constructed using neighbor joining method with bootstrap value of 1000 and the tree was visualized with modifications using tvBOT [30].
2.7. Determination of the Optimum Growth Temperature for the NW2 Isolate
The NW2 and NW4 isolates were cultured in 20 mL of NB (Beef extract 3 g/L, peptone 5 g/L, and NaCl 5 g/L, pH 7.5) at 55 °C by shaking at 150 rpm until the OD600 value reaches to 0.5. Then, 1 mL of culture was inoculated into 60 mL of NB and incubated at different temperatures (40 °C, 45 °C, 50 °C, 55 °C, 60 °C, and 65 °C) by shaking at 150 rpm. At one hour interval, OD600 values of each culture were measured using spectrophotometer and growth curves were plotted. The exponential phase of each growth curve was selected and corresponding OD600 values were converted to Natural Log values and growth rates were determined by linear regression analysis by Microsoft Excel® version 2019. Each experiment was performed in triplicates.
2.8. Isolation of Crude Extracellular Amylase Enzyme from the NW2 Isolate
The crude extracellular amylase enzyme was produced and isolated from overnight culture of NW2 strain incubated at 55 °C in 50 mL of NB (Peptone 5 g/L, NaCl 5 g/L, yeast extract 1.5 g/L, beef extract 1.5 g/L, soluble starch 2 g/L, pH 7.5) shaking at 150 rpm. Then, the culture was centrifuged at 10000 rpm for 10 minutes at 4 °C. The supernatant was filtered through a bacterial filter (0.22 μm) and stored at 4 °C for conducting amylase activity assay using 3,5-Dinitrosalicylic acid (DNSA) colorimetric assay [31].
2.9. Extracellular Amylase Activity Analysis by DNSA Assay
The extracellular amylase activity was analyzed using DNSA colorimetric assay as described previously [23,31] with slight modifications. Briefly, 0.1 mL of crude enzyme was mixed with 0.9 mL of 1% (w/v) soluble starch solution dissolved in 0.05 M phosphate buffer (pH 8) and incubated at 60 °C for 20 minutes. Then, 1 mL of DNSA reagent was added and heated the mixture in a boiling water bath for 5 minutes. After cooling the reaction mixture at room temperature, 8 mL of distilled water was added to the mixture and absorbance was measured at 540 nm using a spectrophotometer. A solution without crude enzyme was used as the control. The amount of maltose (µmol) produced was determined from a maltose standard curve. All experiments were conducted in triplicates. All experiments were conducted in triplicates and results are expressed as mean ± SD. One unit (U) of crude amylase activity was defined as the amount of crude enzyme that liberates 1 µmol of reducing sugars (as maltose) per minute under assay conditions and crude extracellular amylase enzyme activity was expressed as U/mL. The crude extracellular amylase activity was calculated using the following formula [32].
2.10. Determination of the Optimum Temperature for the Extracellular Amylase Activity
The optimum temperature for crude extracellular amylase activity was determined by reacting 0.1 mL of crude enzyme with 0.9 mL of 1% soluble starch solution dissolved in 0.05 M phosphate buffer (pH 7.5) for 20 minutes at different temperatures (40 °C, 45 °C, 50 °C, 55 °C, 60 °C, and 65 °C). Then the activity was analyzed by DNSA assay as described above. All experiments were conducted in triplicates and results are expressed as mean ± SD.
2.11. Determination of Optimum pH for the Extracellular Amylase Activity
For determining optimum pH for crude extracellular amylase activity, 0.05 M sodium phosphate buffer (pH 5-8) and 0.05 M Tris-HCl buffer (pH 7-9) were used. 0.1 mL of crude enzyme was mixed with 0.9 mL of 1% soluble starch solution dissolved in different buffer solutions for 20 minutes at 55 °C. The crude enzyme activity of each reaction was analyzed by DNSA assay. All experiments were conducted in triplicates and results are expressed as mean ± SD.
2.12. Hydrolysis of Raw Cassava Starch by Crude Extracellular Amylase
Cassava starch was purchased from local market. Different concentrations of casava starch (0.5%, 1%, 2%, 3%, and 4%,) suspension were prepared by dissolving in 0.05 M sodium phosphate buffer (pH 8.0). 0.1 mL of crude enzyme was mixed with 0.9 mL of each different casava starch suspension separately and incubated for 4 hours at 60 °C. Then, 0.1 mL of iodine solution (2% I2 and 0.2% KI) was added into each reaction mixture and mixed well to observe the color differences. A control reaction prepared by 1% starch solution without the addition of crude enzyme was included for comparison.
3. Results
3.1. Isolation, Morphological and Biochemical Characterization of Thermophilic Bacterial Strains
In this study, we isolated six thermophilic bacterial strains (NW1, NW2, NW3, NW4, NW6, and MD2) from the Nelumwewa geothermal spring of Sri Lanka. The thermal spring exhibited an average pH of 7.5 indicating slightly alkaline conditions while average water temperature ranged from 48 °C and 55 °C across different sampling points. All isolates showed robust growth at 55 °C in NB as well as on nutrient agar plates at pH 7.5. The morphological and biochemical characteristics of the isolates are summarized in the Table 1. Only NW2 and NW4 exhibited prominent extracellular amylase activity whereas, NW6, and MD2 showed marginal activity. The isolate NW1 and NW3 did not show extracellular amylase activity at all (Figure 2). Moreover, NW2 and NW4 isolates indicated an extracellular protease activity as well (Figure 3). Light microscopy revealed that the NW2 isolate is a Gram-positive, rod-shaped bacterium (Figure 4).
3.2. Identification of Thermophilic Bacterial Isolates That Produce Extracellular Amylase and Protease
Among the six bacterial isolates, only NW2 and NW4 exhibited prominent clear zones around their colonies (Figure 3), with an average diameter of 25 mm, indicating a relatively high potential for extracellular amylase production. In contrast, isolates NW6 and MD2 demonstrated only weak amylase activity, with average zone diameters of approximately 5 mm. The remaining isolates showed no detectable extracellular amylase activity. Additionally, slight extracellular protease activity was observed only in NW2 and NW4, suggesting potential for extracellular protease production (Figure 4). Based on molecular characterization results, NW2 and NW4 were identified to be the identical isolates (Table 2). Therefore, Due to their pronounced extracellular amylase activity, NW2 was selected for further and detailed characterization of their amylase enzymes.
3.3. Molecular Characterization of Thermophilic Bacterial Isolates
Based on 16S rRNA gene sequence analysis of the thermophilic bacterial isolates, isolates NW2 and NW4 identified to be the identical isolates and both exhibited 99.3% of identity to the Thermaerobacillus caldiproteolyticus strain SF03 (NR_115200.1). Whereas, NW6, and MD2 isolates were also identical to each other and showed 99.71% identity to Bacillus licheniformis strain ATCC 14580 (NR_074923.1). Moreover, NW1, and NW3 were also identical isolates and showed 96.85% of identity to Anoxybacillus gonensis strain G2 (NR_025667.1) (Table 2).
The phylogenetic analysis of the thermophilic bacterial isolates revealed that NW2, NW6, and NW3 are clustered into three separate groups A, B, and C, respectively (Figure 5). The NW2 isolate is grouped into cluster A showing higher similarity to T. caldiproteolyticus species. The cluster B is mainly consisting with B. licheniformis strains and the NW6 isolate is grouped into cluster B exhibiting a close phylogenetic relationship to B. licheniformis. Moreover, NW3 isolate is grouped into cluster C with a higher similarity to A. gonensis.
3.4. Optimum Growth Temperature of the NW2 Isolate
The NW2 isolate exhibited the optimal growth characteristics at 55 °C (Figure 6). The lag phase of the growth curve lasted for about 5 hours and then reached to exponential phase. The bacterial growth rates at exponential phases were determined by linear regression analysis (R2 > 0.95). The bacterial growth reached to the stationary phase after about 11 hours indicating its rapid growth characteristics at its optimum temperature. The NW2 isolate exhibited suboptimal growth at all other temperatures tested (Figure 6). Moreover, the maximum growth rate of NW2 isolate exhibited at 55 °C (Figure 7). These results indicated that NW2 isolate is a moderate thermophile.
3.5. Characterization of Crude Extracellular Amylase Produced by NW2 Isolate
3.5.1. Determination of Optimal Temperature for the Enzyme Activity
To determine the optimal temperature of extracellular amylase produced by NW2 isolate, activity of crude amylase was analyzed by DNSA assay at different temperatures ranging from 40 °C to 80 °C. According to the results, the optimum temperature was determined to be 60 °C. However, the enzyme exhibited substantial activity up to 75 °C and declined drastically thereafter (Figure 8). These results indicate that the extracellular amylase produced by NW2 isolates is thermostable in a wide range of temperature from 55 °C to 75 °C.
3.5.2. Determination of Optimal pH Enzyme Activity
Moreover, the amylase enzyme exhibited optimal activity at pH 8.0 in both sodium phosphate and tris-HCl buffer systems (Figure 9). Furthermore, it remains substantially active within a pH range of 7.5 to 8.5. These findings suggest that the extracellular amylase functions effectively at slightly alkaline conditions.
3.6. Extracellular Crude Amylase Activity of NW2 Isolate Under Optimum Temperature and pH
The crude extracellular amylase was harvested from overnight culture of NM2 isolate cultured in NB (60 °C and pH 8). By DNS Assy, the crude extracellular amylase activity was quantified revealing a mean activity of 0.85 U/mL under the optimal assay conditions (60 °C and pH 8).
3.7. Hydrolysis of Raw Cassava Starch by NW2 Crude Extracellular Amylase
Under the optimum conditions, (pH 8.0 and temperature 55 °C), the crude amylase isolated from the NW2 isolate exhibited hydrolyzing of raw cassava starch (Figure 10) indicating its potential commercial application in food, textile, and detergent industries.
4. Discussion
Extremophiles are increasingly recognized as valuable sources of industrially relevant enzymes due to their ability to produce extremozymes that remain active under extreme environmental conditions. Many thermally adapted enzymes or thermozymes, such as thermostable amylases, proteases, cellulases, and lipases, used in diverse industrial applications have been extensively derived from thermophilic and other extremophilic microorganisms [6]. Owing to their remarkable resilience and efficiency, extremozymes have enormous economic potential in biotechnology sector. Among the microbial extremozymes, thermally stable amylases are of significant interest in all starch-related industries [6,33] while these enzymes account for nearly 25% of the world enzyme market [9]. As a result, thermozymes have received a greater attention for their suitability in high-temperature industrial processes, where conventional enzymes often fail to perform optimally [19].
In the present study, three thermophilic bacterial strains were successfully isolated and molecularly characterized from the Nelumwewa geothermal springs in Sri Lanka. These isolates exhibited robust growth at 55 °C under slightly alkaline conditions (pH 7.5). However, among them, only NW2 isolate exhibited prominent extracellular amylase activity as well as extracellular protease activity indicating it as a promising candidate to produce thermostable industrial enzymes. Molecular characterization of NW2 isolate revealed that it shared 99.3% 16S rRNA gene sequence identity with T. caldiproteolyticus strain SF03, previously classified as Anoxybacillus caldiproteolyticus or Geobacillus caldoproteolyticus [34,35], a well-documented thermophilic bacterium known for its thermostable enzyme production [36,37]. Phylogenetic analysis grouped the three isolates into three distinct clusters. The NW2 isolate clustered with T. caldiproteolyticus strains (Cluster A), indicating a close evolutionary relationship, while NW6 and NW3 were grouped into Clusters B and C, respectively, showing phylogenetic affiliation with Bacillus licheniformis and A. gonensis. A recent study on bacterial diversity analysis of Mahapelessa geothermal spring of Sri Lanka has identified some bacterial species such as, Pannonibactor spp, Gulbenkiania spp, Pigmentiphaga spp, Enterobacter spp, Klebsiella spp, and Bacillus spp [25]. However, they did not have isolated any Thermaerobacillus spp, Anoxybacillus spp. or B. licheniformis underscoring a wide bacterial diversity across different geothermal springs of Sri Lanka.
Under the optimal growth conditions (55 °C and pH 7.5), NW2 isolate reached to the stationary phase after about 11 hours in NB medium indicating its rapid growth characteristics. Some reports have suggested that bacterial amylase production is growth dependent [38,39]. Moreover, NW2 extracellular amylase exhibited optimal activity at 60 °C and pH 8.0 which are higher than the those of optimal bacterial growth. This is a common trait found in some thermostable enzymes due to their structural rigidity [40] suggesting evolutionary adaptation to seasonal fluctuation of temperature and pH of geothermal springs. The optimal temperature and pH of the extracellular amylase activity of NW2 isolate are almost consistent with already reported amylase activity of some Anoxybacillus spp. [22,23]. Under these optimal conditions for the enzyme activity, the crude extracellular amylase unit activity of the NW2 was found to be 0.85 U/mL. The unit activity of this enzyme is somewhat consistent with previous reports on crude amylase activity from thermophilic Anoxybacillus species [23].
Numerous studies have demonstrated that extracellular amylases isolated from various bacterial species possess significant potential for industrial-scale applications. Amylases derived from thermophilic bacteria B. licheniformis exhibits excellent amylolytic properties and has been successfully applied in the biodegradation of food waste [41]. Moreover, amylases from different bacterial sources display remarkably high thermal optima, including 135 °C for B. subtilis [42] and 85 °C for Rhodothermus marinus [43]. Other thermophilic bacteria, such as Bacillus caldolyticus [44] and Geobacillus thermoleovorans [45] exhibit peak enzymatic activity at around 70 °C highlighting the wide range of thermostability among bacterial amylase. The thermostability and broad pH tolerance of these enzymes reduce the need for cooling steps and pH adjustments, improving process efficiency and cost-effectiveness.
The bacterial species T. caldiproteolyticus (A. caldiproteolyticus) is pridominantly known for its production of extracellular proteases rather than amylase [36]. To date, only a single study has reported extracellular amylase production by A. caldiproteolyticus [46] suggesting further studies to investigate the potential of producing extracellular amylase by this species. To the best of our knowledge, this is the first report of a T. caldiproteolyticus-like strain from Sri Lankan geothermal springs exhibiting a remarkable extracellular amylase activity. The NW2 isolate is a gram-positive, rod shaped and catalase positive bacterium. These morphological and biochemical characteristics are commonly associated with well-known amylase producing bacteria [47]. In contrast, NW6 and MD2 isolates exhibited a very close phylogenetic relationship with 99.71% 16S rRNA gene sequence identity to B. licheniformis, a well-established industrial bacterial species recognized for its thermostable amylase production [48,49,50,51]. However, neither NW6 nor MD2 exhibited significant extracellular amylase activity under the tested conditions, suggesting potential strain-level variability or previously undocumented functional diversity within the species.
Cassava starch is extensively utilized across diverse industries, including food processing, textiles, and bioethanol production. However, its hydrolysis is often associated with high operational costs, primarily due to the energy-intensive nature of conventional processing methods. Recent studies have demonstrated the successful application of microbial amylases in the production of bioethanol from raw cassava starch [52]. In this context, the extracellular amylase produced by the NW2 isolate in the present study exhibited strong hydrolytic activity toward raw cassava starch, highlighting its potential for use in both the food and biofuel industries, where efficient and rapid starch degradation is essential. Furthermore, the enzyme’s stability under moderately alkaline conditions enhances its applicability in the detergent industry, where alkaline-tolerant amylases are required.
Overall, the findings of the present study not only reveal the biotechnological potential of the bacteria isolates from the Nelumwewa geothermal spring but also highlight the geothermal springs of Sri Lanka as a promising niche environment for exploring thermophilic enzyme producing microbes.
5. Conclusions
This study highlights the biotechnological potential of NW2, a T. caldiproteolyticus-like strain from the Nelumwewa geothermal springs in Sri Lanka, a previously unexplored thermal ecosystem. The NW2 isolate emerged as a promising candidate for industrial applications because of its prominent extracellular amylase production with notable thermostability and activity in alkaline environments. Furthermore, the extracellular amylase of NW2 demonstrated effective hydrolysis of raw cassava starch, suggesting its direct applicability in food, bioethanol, and detergent industries where robust and efficient enzymatic performance under high temperature and pH is critical. This marks the first report of a T. caldiproteolyticus-like strain from Sri Lanka with such enzymatic activity, expanding the known functional diversity of this species and underscoring the potential for strain-level variation. The findings of the present study reinforce the value of geothermal environments as reservoirs of novel extremozymes suggesting further exploration to support the development of sustainable, high-performance biocatalysts for industrial applications.
Author Contributions
Conceptualization, S.B., P.J. and A.W.; methodology, S.B., L.P., B.D., P.J. and A.W.; validation, L.P. and S.B.; formal analysis, L.P., B.D. and S.B.; investigation, S.B.; data curation, L.P., B.D. and S.B.; writing—original draft preparation, B.D. and L.P.; writing—review and editing, S.B., A.W.; supervision, S.B. and A.W.; project administration, S.B.; funding acquisition, S.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Science and Technology Human Resource Development (STHRD) Project, Ministry of Education, Sri Lanka, under the patronage of Asian Development Bank (ADB). Grant No. CRG/R3/SB1. (https://sthrd.gov.lk/index.php).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study.
Abbreviations
The following abbreviations are used in this manuscript:
| NB | Nutrient broth |
| DNSA | 3,5-Dinitrosalicylic acid |
References
- Raddadi, N.; Cherif, A.; Daffonchio, D.; Neifar, M.; Fava, F. Biotechnological Applications of Extremophiles, Extremozymes and Extremolytes. Appl Microbiol Biotechnol 2015, 99, 7907–7913. [Google Scholar] [CrossRef]
- Cabrera, M.Á.; Blamey, J.M. Biotechnological Applications of Archaeal Enzymes from Extreme Environments. Biological Research 2018, 51, 1–15. [Google Scholar] [CrossRef]
- Marzban, G.; Tesei, D. The Extremophiles: Adaptation Mechanisms and Biotechnological Applications. Biology 2025, 14, 412. [Google Scholar] [CrossRef]
- Elleuche, S.; Schröder, C.; Sahm, K.; Antranikian, G. Extremozymes-Biocatalysts with Unique Properties from Extremophilic Microorganisms. Curr Opin Biotechnol 2014, 29, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R.; Siddiqui, K.S.; Andrews, D.; Sowers, K.R. Low-Temperature Extremophiles and Their Applications. Curr Opin Biotechnol 2002, 13, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Ashaolu, T.J.; Malik, T.; Soni, R.; Prieto, M.A.; Jafari, S.M. Extremophilic Microorganisms as a Source of Emerging Enzymes for the Food Industry: A Review. Food Sci Nutr 2025, 13. [Google Scholar] [CrossRef]
- Al-Batayneh, K.M.; Jacob, J.H.; Hussein, E. Isolation and Molecular Identification of New Thermophilic Bacterial Strains of Geobacillus Pallidus and Anoxybacillus Flavithermus. 2011; Vol. 11. [Google Scholar]
- Wang, Q.; Cen, Z.; Zhao, J. The Survival Mechanisms of Thermophiles at High Temperatures: An Angle of Omics. Physiology 2015, 30, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Kikani, B.A.; Singh, S.P. Amylases from Thermophilic Bacteria: Structure and Function Relationship. Crit Rev Biotechnol 2022, 42, 325–341. [Google Scholar] [CrossRef]
- Arbab, S.; Ullah, H.; Khan, M.I.U.; Khattak, M.N.K.; Zhang, J.; Li, K.; Hassan, I.U. Diversity and Distribution of Thermophilic Microorganisms and Their Applications in Biotechnology. J Basic Microbiol 2022, 62, 95–108. [Google Scholar] [CrossRef]
- Ali, Z.; Abdullah, M.; Yasin, M.T.; Amanat, K.; Sultan, M.; Rahim, A.; Sarwar, F. Recent Trends in Production and Potential Applications of Microbial Amylases: A Comprehensive Review. Protein Expr Purif 2025, 227, 106640. [Google Scholar] [CrossRef]
- Prakash, O.; Jaiswal, N. α-Amylase: An Ideal Representative of Thermostable Enzymes. Appl Biochem Biotechnol 2010, 160, 2401–2414. [Google Scholar] [CrossRef] [PubMed]
- Slavić, M.Š.; Kojić, M.; Margetić, A.; Stanisavljević, N.; Gardijan, L.; Božić, N.; Vujčić, Z. Highly Stable and Versatile α-Amylase from Anoxybacillus Vranjensis ST4 Suitable for Various Applications. Int J Biol Macromol 2023, 249, 126055. [Google Scholar] [CrossRef]
- Kikani, B.A.; Singh, S.P. The Stability and Thermodynamic Parameters of a Very Thermostable and Calcium-Independent α-Amylase from a Newly Isolated Bacterium, Anoxybacillus Beppuensis TSSC-1. Process Biochemistry 2012, 47, 1791–1798. [Google Scholar] [CrossRef]
- Agüloǧlu Fincan, S.; Enez, B.; Özdemir, S.; Matpan Bekler, F. Purification and Characterization of Thermostable α-Amylase from Thermophilic Anoxybacillus Flavithermus. Carbohydr Polym 2014, 102, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Ammar, Y.B.; Matsubara, T.; Ito, K.; Iizuka, M.; Limpaseni, T.; Pongsawasdi, P.; Minamiura, N. New Action Pattern of a Maltose-Forming α-Amylase from Streptomyces Sp. and Its Possible Application in Bakery. BMB Rep 2002, 35, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.H.; Liu, W.H. Purification and Properties of a Maltotriose-Producing α-Amylase from Thermobifida Fusca. Enzyme Microb Technol 2004, 35, 254–260. [Google Scholar] [CrossRef]
- Burhan, A.; Nisa, U.; Gökhan, C.; Ömer, C.; Ashabil, A.; Osman, G. Enzymatic Properties of a Novel Thermostable, Thermophilic, Alkaline and Chelator Resistant Amylase from an Alkaliphilic Bacillus Sp. Isolate ANT-6. Process Biochemistry 2003, 38, 1397–1403. [Google Scholar] [CrossRef]
- Ullah, I.; Khan, M.S.; Khan, S.S.; Ahmad, W.; Zheng, L.; Shah, S.U.A.; Ullah, M.; Iqbal, A. Identification and Characterization of Thermophilic Amylase Producing Bacterial Isolates from the Brick Kiln Soil. Saudi J Biol Sci 2021, 28, 970–979. [Google Scholar] [CrossRef]
- Silva-Salinas, A.; Rodríguez-Delgado, M.; Gómez-Treviño, J.; López-Chuken, U.; Olvera-Carranza, C.; Blanco-Gámez, E.A. Novel Thermotolerant Amylase from Bacillus Licheniformis Strain Lb04: Purification, Characterization and Agar-Agarose. Microorganisms 2021, 9, 1857. [Google Scholar] [CrossRef]
- Deljou, A.; Arezi, I. Production of Thermostable Extracellular α-Amylase by a Moderate Thermophilic Bacillus Licheniformis-AZ2 Isolated from Qinarje Hot Spring (Ardebil Prov. of Iran). Period Biol 2016, 118, 405–416. [Google Scholar] [CrossRef]
- Acer, Ö.; Pirinççioʇlu, H.; Matpan Bekler, F.; Gül-Güven, R.; Güven, K. Anoxybacillus Sp. AH1, an α-Amylase-Producing Thermophilic Bacterium Isolated from Dargeçit Hot Spring. Biologia (Poland) 2015, 70, 853–862. [Google Scholar] [CrossRef]
- Sharif, S.; Shah, A.H.; Fariq, A.; Jannat, S.; Rasheed, S.; Yasmin, A. Optimization of Amylase Production Using Response Surface Methodology from Newly Isolated Thermophilic Bacteria. Heliyon 2023, 9. [Google Scholar] [CrossRef]
- Al-Harthy, O.D.; Hozzein, W.N.; Abdelkader, H.S.; Alharbi, S.A.A. Thermostable Amylase from Cytobacillus Firmus: Characterization, Optimization, and Implications in Starch Hydrolysis. Egypt J Microbiol 2024. [Google Scholar] [CrossRef]
- Samarasinghe, S.N.; Wanigatunge, R.P.; Magana-Arachchi, D.N. Bacterial Diversity in a Sri Lankan Geothermal Spring Assessed by Culture-Dependent and Culture-Independent Approaches. Curr Microbiol 2021, 78, 3439–3452. [Google Scholar] [CrossRef] [PubMed]
- Patabendigedara, S.; Kumara, S.M.P.G.S.; Dharmagunawardhane, H.A. GEOSTRUCTURAL MODEL FOR THE NELUMWEWA THERMAL SPRING; NORTH CENTRAL PROVINCE, SRI LANKA A GEOSTRUCTURAL MODEL FOR THE NELUMWEWA THERMAL SPRING: NORTH CENTRAL PROVINCE, SRI LANKA, 2014; Vol. 16. [Google Scholar]
- Reiner, K. Catalase Test Protocol. - References - Scientific Research Publishing. 2012. Available online: https://www.scirp.org/reference/referencespapers?referenceid=1367750 (accessed on 8 May 2025).
- Hols, P.; Ferain, T.; Garmyn, D.; Bernard, N.; Delcour, J. Use of Homologous Expression-Secretion Signals and Vector-Free Stable Chromosomal Integration in Engineering of Lactobacillus Plantarum for Alpha-Amylase and Levanase Expression. Appl Environ Microbiol 1994, 60, 1401. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.G. Building Phylogenetic Trees from Molecular Data with MEGA. Mol Biol Evol 2013, 30, 1229–1235. [Google Scholar] [CrossRef]
- Xie, J.; Chen, Y.; Cai, G.; Cai, R.; Hu, Z.; Wang, H. Tree Visualization by One Table (TvBOT): A Web Application for Visualizing, Modifying and Annotating Phylogenetic Trees. Nucleic Acids Res 2023, 51, W587–W592. [Google Scholar] [CrossRef]
- Bernfeld, P. [17] Amylases, α and β. Methods Enzymol 1955, 1, 149–158. [Google Scholar] [CrossRef]
- Soy, S.; Nigam, V.K.; Sharma, S.R. Enhanced Production and Biochemical Characterization of a Thermostable Amylase from Thermophilic Bacterium Geobacillus Icigianus BITSNS038. Journal of Taibah University for Science 2021, 15, 730–745. [Google Scholar] [CrossRef]
- Yadav, A.N. Biodiversity and Bioprospecting of Extremophilic Microbiomes for Agro-Environmental Sustainability. J Appl Biol Biotechnol 2021, 9, 1–6. [Google Scholar]
- Patel, I.; Bello, S.; Gupta, R.S. Phylogenomic and Molecular Marker Based Studies to Clarify the Evolutionary Relationships amongst Anoxybacillus Species and Demarcation of the Family Anoxybacillaceae and Some of Its Constituent Genera. Int J Syst Evol Microbiol 2024, 74. [Google Scholar] [CrossRef] [PubMed]
- Coorevits, A.; Dinsdale, A.E.; Halket, G.; Lebbe, L.; de Vos, P.; van Landschoot, A.; Logan, N.A. Taxonomic Revision of the Genus Geobacillus: Emendation of Geobacillus, G. Stearothermophilus, G. Jurassicus, G. Toebii, G. Thermodenitrificans and G. Thermoglucosidans (Nom. Corrig., Formerly ’Thermoglucosidasius’); Transfer of Bacillus Thermantarcticus to the Genus as G. Thermantarcticus Comb. Nov.; Proposal of Caldibacillus Debilis Gen. Nov., Comb. Nov. Int J Syst Evol Microbiol 2012, 62, 1470–1485. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.H.; Wang, Y.; Zhang, X.Y.; Sun, M.L.; Zhang, X.; Song, X.Y.; Zhang, Y.Z.; Zhang, Y.; Chen, X.L. Characterization and Diversity Analysis of the Extracellular Proteases of Thermophilic Anoxybacillus Caldiproteolyticus 1A02591 From Deep-Sea Hydrothermal Vent Sediment. Front Microbiol 2021, 12. [Google Scholar] [CrossRef]
- Chen, X.G.; Stabnikova, O.; Tay, J.H.; Wang, J.Y.; Tay, S.T.L. Thermoactive Extracellular Proteases of Geobacillus Caldoproteolyticus, Sp. Nov., from Sewage Sludge. Extremophiles 2004, 8, 489–498. [Google Scholar] [CrossRef]
- Simair, A.A.; Khushk, I.; Qureshi, A.S.; Bhutto, M.A.; Chaudhry, H.A.; Ansari, K.A.; Lu, C. Amylase Production from Thermophilic Bacillus Sp. BCC 021-50 Isolated from a Marine Environment. Fermentation 2017, 3, 25. [Google Scholar] [CrossRef]
- Hmidet, N.; El Hadj Ali, N.; Zouari-Fakhfakh, N.; Haddar, A.; Nasri, M.; Sellemi-Kamoun, A. Chicken Feathers: A Complex Substrate for the Co-Production of α-Amylase and Proteases by B. Licheniformis NH1. J Ind Microbiol Biotechnol 2010, 37, 983–990. [Google Scholar] [CrossRef]
- Hmidet, N.; Bayoudh, A.; Berrin, J.G.; Kanoun, S.; Juge, N.; Nasri, M. Purification and Biochemical Characterization of a Novel α-Amylase from Bacillus Licheniformis NH1. Cloning, Nucleotide Sequence and Expression of AmyN Gene in Escherichia Coli. Process Biochemistry 2008, 43, 499–510. [Google Scholar] [CrossRef]
- Msarah, M.J.; Ibrahim, I.; Hamid, A.A.; Aqma, W.S. Optimisation and Production of Alpha Amylase from Thermophilic Bacillus Spp. and Its Application in Food Waste Biodegradation. Heliyon 2020, 6, e04183. [Google Scholar] [CrossRef] [PubMed]
- Konsoula, Z.; Liakopoulou-Kyriakides, M. Co-Production of α-Amylase and β-Galactosidase by Bacillus Subtilis in Complex Organic Substrates. Bioresour Technol 2007, 98, 150–157. [Google Scholar] [CrossRef]
- Gomes, I.; Gomes, J.; Steiner, W. Highly Thermostable Amylase and Pullulanase of the Extreme Thermophilic Eubacterium Rhodothermus Marinus: Production and Partial Characterization. Bioresour Technol 2003, 90, 207–214. [Google Scholar] [CrossRef]
- Schwab, K.; Bader, J.; Brokamp, C.; Popović, M.K.; Bajpai, R.; Berovič, M. Dual Feeding Strategy for the Production of α-Amylase by Bacillus Caldolyticus Using Complex Media. N Biotechnol 2009, 26, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Uma Maheswar Rao, J.L.; Satyanarayana, T. Improving Production of Hyperthermostable and High Maltose-Forming α-Amylase by an Extreme Thermophile Geobacillus Thermoleovorans Using Response Surface Methodology and Its Applications. Bioresour Technol 2007, 98, 345–352. [Google Scholar] [CrossRef]
- CALISKAN OZDEMIR, S.; COLERI CIHAN, A.; KILIC, T.; COKMUS, C. OPTIMIZATION OF THERMOSTABLE ALPHA-AMYLASE PRODUCTION FROM GEOBACILLUS SP. D413. Journal of microbiology, biotechnology and food sciences 2016, 6, 689–694. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, W.J.; Ryu, J.; Yerefu, Y.; Tesfaw, A. Amylase Production by the New Strains of Kocuria Rosea and Micrococcus Endophyticus Isolated from Soil in the Guassa Community Conservation Area. Fermentation 2025, 11, 211. [Google Scholar] [CrossRef]
- Aras, S.; Altıntas, R.; Aygun, E.; Goren, G.; Sisecioglu, M. Purification of α-Amylase from Thermophilic Bacillus Licheniformis SO5 by Using a Novel Method, Alternating Current Magnetic-Field Assisted Three-Phase Partitioning: Molecular Docking and Bread Quality Improvement. Food Chem 2025, 484. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudnia, F. Comparison of the Synthesis of the Alpha-Amylase Enzyme by the Native Strain Bacillus Licheniformis in Immobilized and Immersed Cells. Iran J Microbiol 2024, 16, 827. [Google Scholar] [CrossRef]
- Suthar, S.; Joshi, D.; Patel, H.; Patel, D.; Kikani, B.A. Optimization and Purification of a Novel Calcium-Independent Thermostable, α-Amylase Produced by Bacillus Licheniformis UDS-5. World J Microbiol Biotechnol 2024, 40. [Google Scholar] [CrossRef] [PubMed]
- Divakaran, D.; Chandran, A.; Pratap Chandran, R. Comparative Study on Production of A-Amylase from Bacillus Licheniformis Strains. Braz J Microbiol 2011, 42, 1397–1404. [Google Scholar] [CrossRef]
- Roble, N.D.; Ogbonna, J.; Tanaka, H. Simultaneous Amylase Production, Raw Cassava Starch Hydrolysis and Ethanol Production by Immobilized Aspergillus Awamori and Saccharomyces Cerevisiae in a Novel Alternating Liquid Phase–Air Phase System. Process Biochemistry 2020, 95, 115–121. [Google Scholar] [CrossRef]
Figure 1.
Locations of geothermal springs across different geological zones of Sri Lanka [26].
Figure 1.
Locations of geothermal springs across different geological zones of Sri Lanka [26].
Figure 2.
Screening of extracellular amylase producing thermophilic bacterial isolates. Extracellular amylase producing isolates exhibit clear zone around colonies. (a) NW1; (b) NW2; (c) NW3; (d) NW4; (e) NW6; (f) MD2 isolates.
Figure 2.
Screening of extracellular amylase producing thermophilic bacterial isolates. Extracellular amylase producing isolates exhibit clear zone around colonies. (a) NW1; (b) NW2; (c) NW3; (d) NW4; (e) NW6; (f) MD2 isolates.

Figure 3.
Screening of extracellular protease producing thermophilic bacterial isolates. Extracellular protease producing isolates exhibit clear zone around colonies. (a) NW2; (b) NW4 isolates.
Figure 3.
Screening of extracellular protease producing thermophilic bacterial isolates. Extracellular protease producing isolates exhibit clear zone around colonies. (a) NW2; (b) NW4 isolates.

Figure 4.
Microscopic visualization of Gram stained NW2 isolate under light microscope.
Figure 5.
The phylogenetic analysis of thermophilic bacterial isolates using 16 S rRNA gene sequences. The tree was constructed using Neighbor joining method with MEGA 12 and visualized with modifications using tvBOT [30].
Figure 5.
The phylogenetic analysis of thermophilic bacterial isolates using 16 S rRNA gene sequences. The tree was constructed using Neighbor joining method with MEGA 12 and visualized with modifications using tvBOT [30].
Figure 6.
Growth curves of NW2 isolate at different temperatures from 40 °C to 65 °C in NB (pH 7.5). The graph show data from triplicate experiments (mean ± SD).
Figure 6.
Growth curves of NW2 isolate at different temperatures from 40 °C to 65 °C in NB (pH 7.5). The graph show data from triplicate experiments (mean ± SD).
Figure 7.
Growth rate of NW2 isolate at different temperatures from 40 °C to 65 °C in NB (pH 7.5). The graph show data from triplicate experiments (mean ± SD).
Figure 7.
Growth rate of NW2 isolate at different temperatures from 40 °C to 65 °C in NB (pH 7.5). The graph show data from triplicate experiments (mean ± SD).
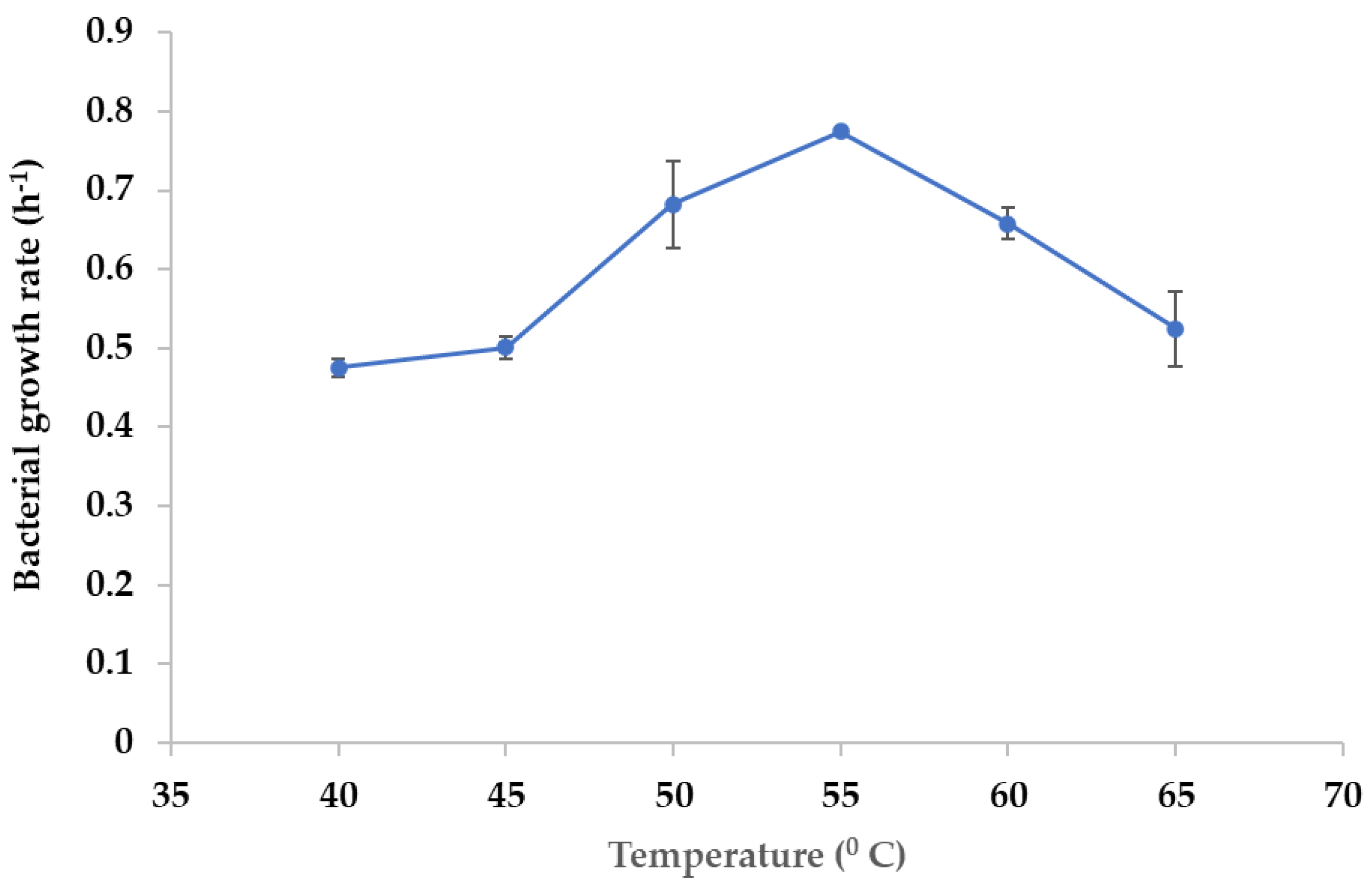
Figure 8.
Optimum temperature for the extracellular amylase activity. The graph show data from triplicate experiments (mean ± SD).
Figure 8.
Optimum temperature for the extracellular amylase activity. The graph show data from triplicate experiments (mean ± SD).
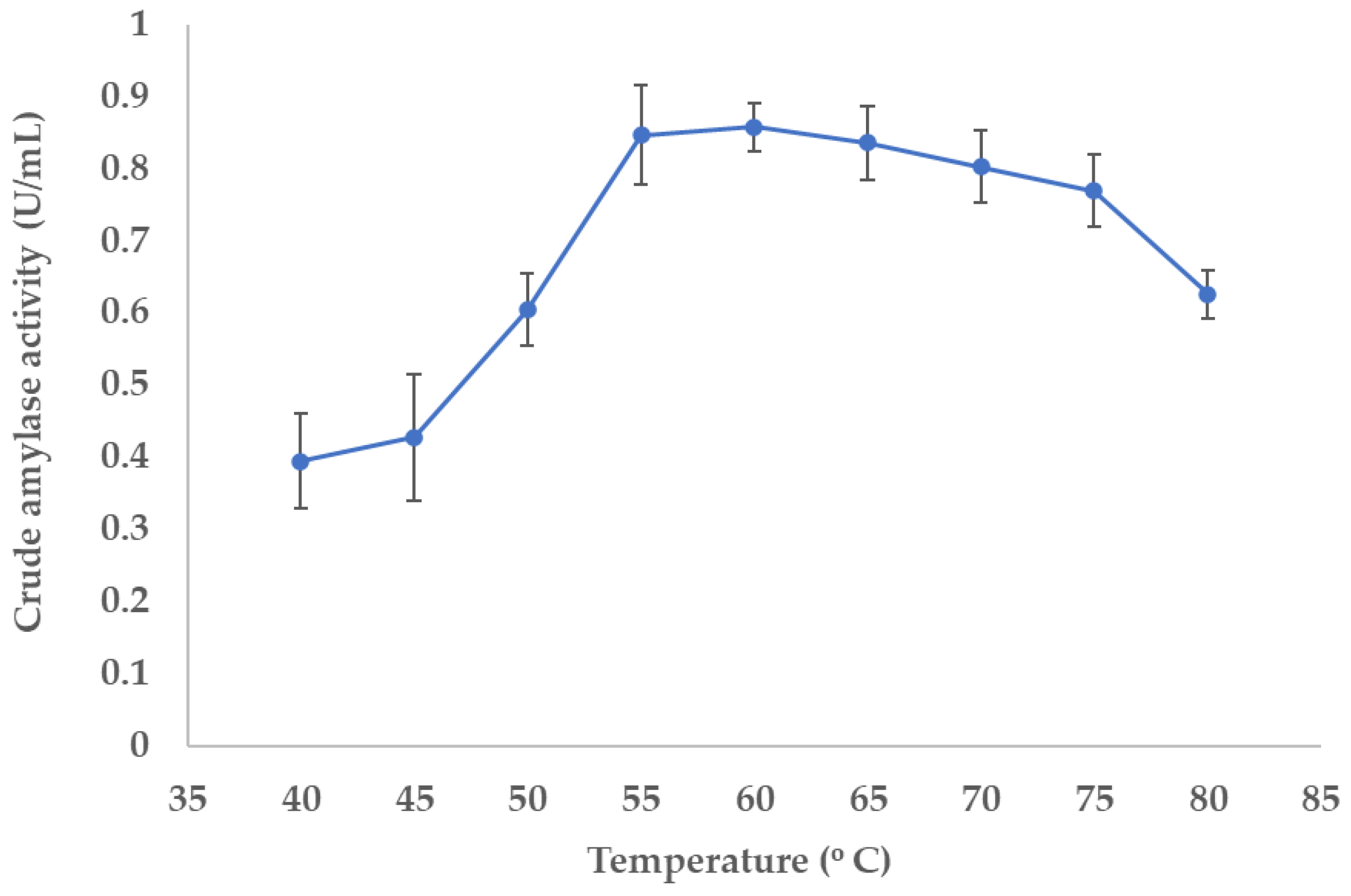
Figure 9.
Optimum pH for the extracellular amylase activity. The graph show data from triplicate experiments (mean ± SD).
Figure 9.
Optimum pH for the extracellular amylase activity. The graph show data from triplicate experiments (mean ± SD).
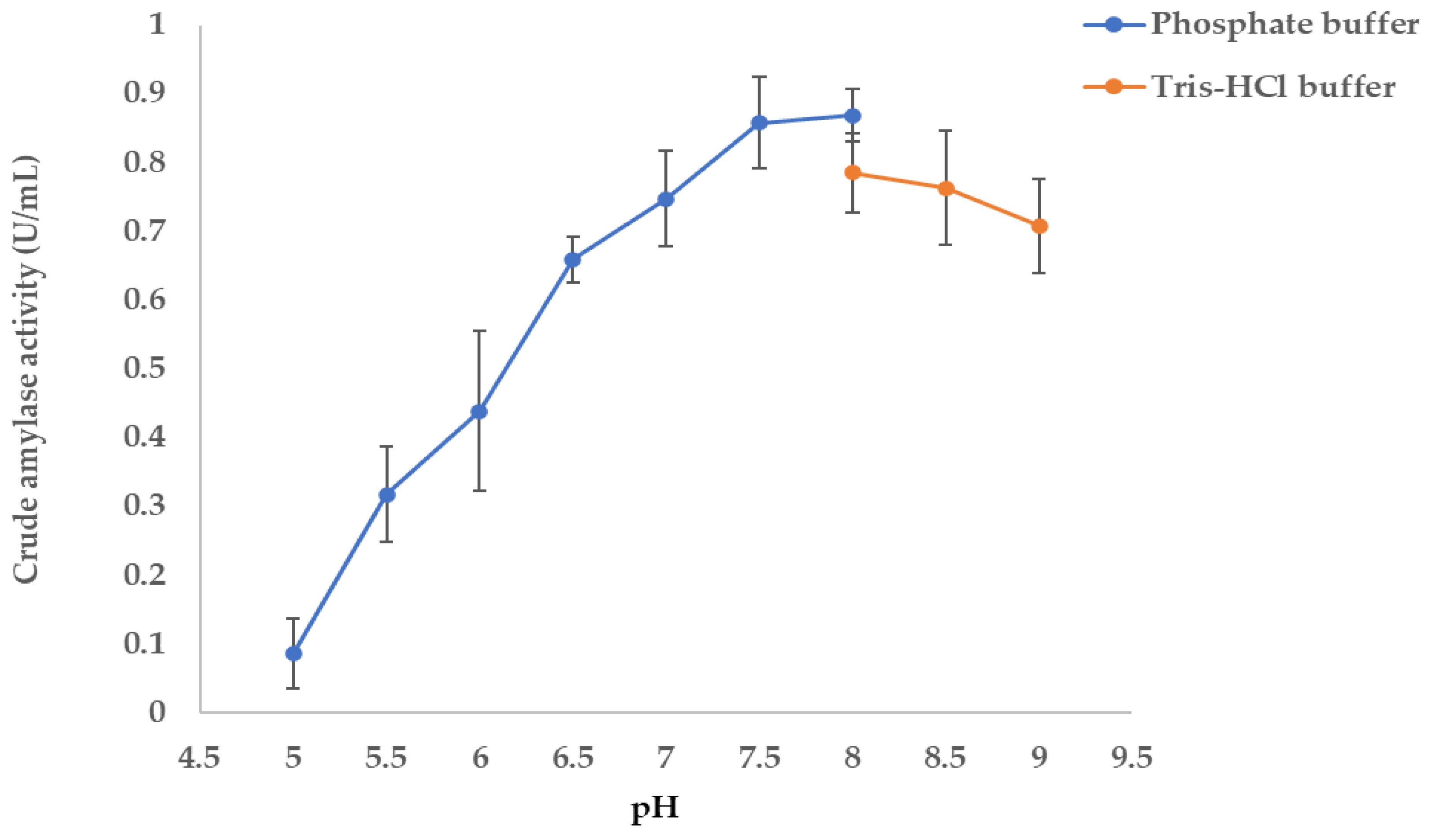
Figure 10.
Hydrolysis of different concentration of cassava starch, (a) 0.5%; (b) 1%; (c) 2%; (d) 3%; (e) 4%, by crude amylase from NW2 isolate under optimum conditions (pH 8.0 and temperature 55 °C); (f) control (1% casava starch with no crude enzyme).
Figure 10.
Hydrolysis of different concentration of cassava starch, (a) 0.5%; (b) 1%; (c) 2%; (d) 3%; (e) 4%, by crude amylase from NW2 isolate under optimum conditions (pH 8.0 and temperature 55 °C); (f) control (1% casava starch with no crude enzyme).


Table 1.
Morphological and biochemical characteristics of the isolated thermophilic bacteria. (+ and – signs indicate positive and negative biochemical tests results, respectively).
Table 1.
Morphological and biochemical characteristics of the isolated thermophilic bacteria. (+ and – signs indicate positive and negative biochemical tests results, respectively).
| Bacterial Isolate | NW1 | NW2 | NW3 | NW4 | NW6 | MD2 |
|---|---|---|---|---|---|---|
| Colony morphology | ||||||
| Shape | Circular | Circular | Circular | Circular | Irregular | Irregular |
| Margin | Entire | Entire | Entire | Entire | Undulate | Undulate |
| Elevation | Convex | Convex | Convex | Convex | Flat | Flat |
| Surface | Smooth | Smooth | Smooth | Smooth | Wrinkled | Wrinkled |
| Color | Off- white | Creamy white | Creamy white | Creamy white | Off- white |
Off- white |
| Cell shape | Rod | Rod | Rod | Rod | Rod | Rod |
| Catalase activity | + | + | + | + | + | + |
| Extracellular enzymes activity | ||||||
| Amylase (Average diameter of clear zone in mm) | 0 | 25 | 0 | 25 | 5 | 5 |
| Protease | - | + | - | + | - | - |
Table 2.
Molecular characterization of thermophilic bacterial isolates by 16S rRNA gene sequencing.
| Isolates | Accession number 1 | Closest relative bacterial species 2 | Accession number 3 | Identity 4 |
|---|---|---|---|---|
| NW2/NW4 | PV489057 | Thermaerobacillus caldiproteolyticus strain SF03 | NR_115200.1 | 99.30% |
| NW1/NW3 | PV489066 | Anoxybacillus gonensis strain G2 | NR_025667.1 | 96.85% |
| NW6/MD2 | PV489038 | Bacillus licheniformis strain ATCC 14580 | NR_074923.1 | 99.71% |
1 NCBI gene bank accession number of thermophilic bacteria isolated in this study. 2 The closest bacterial species with the highest percentage identity match available in NCBI gene bank. 3 NCBI gene bank accession number of the closest bacterial species. 4 BLAST search identity of the closest bacterial species available in NCBI gene bank.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



