Submitted:
12 May 2025
Posted:
13 May 2025
You are already at the latest version
Abstract
Understanding the genetic basis of muscle development and meat quality traits in divergent pig breeds is crucial for advancing precision breeding strategies. This study aimed to investigate transcriptome differences in the longissimus dorsi muscle between Chinese Congjiang Xiang (CX) and Landrace (LAN) pigs. RNA sequencing was performed on muscle tissues from ten individuals of each breed, with an average mapping rate of 89.3% to the pig reference genome. Transcriptional profiling revealed 785 genes exclusively expressed in CX pigs and 457 genes unique to LAN pigs, with 7,099 co-expressed genes shared by both breeds. Principal component analysis demonstrated clear separation between the breeds. Differential expression analysis identified 2,459 differentially expressed genes, with 1,745 up-regulated and 714 down-regulated in CX pigs. Notably, genes associated with meat flavor formation (ELOVL5/6, FASN, DGAT2, ALDH1A3, PPAR-γ) were significantly up-regulated in CX pigs. Functional enrichment analysis showed that up-regulated genes in CX pigs were primarily enriched in immune responses, while down-regulated genes were associated with myosin complex and PPAR signaling pathway. These findings provide novel insights into the molecular mechanisms underlying the distinctive meat characteristics of indigenous Chinese and commercial European pig breeds, offering valuable genetic resources for future breeding programs aimed at enhancing pork quality.
Keywords:
transcriptome
; pig
; longissimus dorsi
; meat quality
; PPAR signaling
1. Introduction
Pork stands as one of the most extensively consumed meat proteins worldwide, accounting for approximately 36% of global meat consumption [1]. This popularity stems from its nutritional profile, which includes high-quality proteins, essential amino acids, and various micronutrients crucial for human health and development [2]. In recent years, meat characteristics, particularly muscle quality traits such as intramuscular fat content, tenderness, and water-holding capacity, have emerged as significant factors influencing consumer purchasing decisions [3]. The longissimus dorsi, commonly known as the loin muscle, is especially valued for its tenderness and flavor attributes, making it one of the most economically important cuts in pork production [4]. These quality traits are predominantly influenced by three major factors: genetics, nutrition, and environmental conditions [5]. Among these, genetic factors play a fundamental role in determining muscle development, fat deposition patterns, and ultimately, meat quality characteristics, contributing to approximately 30-45% of the variation observed in key traits such as intramuscular fat content [6].
With the advancement of modern molecular breeding techniques, the improvement of pork quality has entered a new era that extends beyond traditional selective breeding methods. Contemporary approaches integrate conventional breeding with molecular biology, bioinformatics, and computational technologies to achieve more precise and efficient genetic improvements. Molecular marker-assisted selection, genomic selection, and gene editing technologies have revolutionized pig breeding by enabling the identification and utilization of genes directly associated with economically important traits [7,8]. The screening and validation of functional genes have become crucial components of molecular breeding programs aimed at enhancing meat quality characteristics [9]. Particularly, the genetic mechanisms governing fat deposition in muscle tissues involve intricate and highly synchronized gene expression programs that regulate adipogenesis, lipid metabolism, and fatty acid composition. These complex biological processes ultimately determine the sensory qualities of pork, including juiciness, tenderness, and flavor intensity, making them priority targets for genetic improvement efforts in pig breeding [10].
Transcriptomics has emerged as a pivotal molecular technology in biological research, offering comprehensive insights into gene expression patterns under various physiological and developmental conditions. The rapid advancement of RNA sequencing (RNA-Seq) technology has significantly expanded its application in the livestock industry, providing unprecedented resolution in transcriptome profiling [11]. RNA-Seq analyses enable the identification of differentially expressed genes, novel transcripts, alternative splicing events, and regulatory networks across different species, breeds, and tissues. In pig research specifically, RNA-Seq has been extensively employed to investigate transcriptional variations associated with muscle development, fat deposition, and meat quality traits [12,13]. These studies have identified numerous candidate genes and molecular pathways involved in muscle fiber formation, adipogenesis, and lipid metabolism, contributing substantially to our understanding of the genetic basis of pork quality. The application of transcriptomics is particularly valuable for comparing divergent pig breeds with distinct phenotypic characteristics, as it can reveal the underlying genetic mechanisms responsible for their phenotypic differences [14].
The Chinese Congjiang Xiang pig, indigenous to Congjiang County in Guizhou Province, represents one of China’s valuable native pig genetic resources. This breed has evolved over centuries under local environmental conditions and selective pressures, developing unique adaptations to the mountainous terrain and subtropical climate of southwestern China. Congjiang Xiang pigs are characterized by their small body size, black coat color, and exceptional adaptability to extensive farming systems with limited resources. Their notable resilience to diseases, stress tolerance, and reproductive efficiency highlight their evolutionary advantages as a local breed [15]. Most significantly, Congjiang Xiang pigs exhibit distinctive meat quality characteristics, including a higher intramuscular fat content (typically 3-5%), finer muscle fibers, and a distinctive flavor profile compared to commercial breeds [16]. These attributes contribute to the exceptional tenderness, juiciness, and unique taste of their meat, which is highly valued in traditional Chinese cuisine and local gastronomy. The Landrace pig, originating from Denmark and subsequently improved in various European countries, represents one of the most widely utilized commercial pig breeds globally. They demonstrate superior feed conversion efficiency and lean meat percentage (typically 58-62%) compared to most indigenous Chinese breeds [17]. However, the intensive selection for production efficiency in Landrace pigs has been associated with reduced intramuscular fat content (generally 1.5-2.5%), which may affect certain aspects of meat quality such as flavor intensity and juiciness [18]. The comparative analysis of muscle traits between Congjiang Xiang and Landrace pigs reveals distinct differences in muscle fiber composition, fat distribution patterns, and metabolic characteristics that directly influence their respective meat quality profiles.
The selection of Congjiang Xiang and Landrace pigs as research models provides an exceptional opportunity to investigate the genetic basis of divergent muscle development and quality traits. These breeds represent distinct evolutionary paths in pig domestication and improvement: Congjiang Xiang embodies centuries of natural selection and adaptation to local conditions with minimal human intervention, while Landrace exemplifies intensive artificial selection for production efficiency and carcass traits [19]. This genetic divergence has resulted in pronounced phenotypic differences in muscle characteristics, particularly in intramuscular fat content, muscle fiber composition, and metabolic properties. Comparing these genetically distant breeds enables the identification of key genes, regulatory elements, and biological pathways associated with critical muscle quality traits [20]. Additionally, the contrasting fat deposition patterns between these breeds—with Congjiang Xiang demonstrating superior marbling capacity despite lower overall growth rates—presents a valuable model for understanding the molecular mechanisms governing intramuscular adipogenesis [21]. The knowledge derived from studying these contrasting breeds has significant potential applications for precision breeding strategies aimed at improving meat quality while maintaining production efficiency in commercial pig lines.
In the present study, we employed RNA-Seq technology to analyze differential gene expression patterns in the longissimus dorsi muscle of Chinese Congjiang Xiang and Landrace pigs. This comprehensive transcriptomic approach allowed us to identify and characterize differentially expressed genes (DEGs) between these breeds with distinct muscle development and quality characteristics. The identified DEGs were subjected to detailed bioinformatic analyses, including functional annotation, pathway enrichment, and protein-protein interaction network construction, to elucidate the biological processes and molecular mechanisms underlying the observed phenotypic variations. Our findings reveal novel functional genes and regulatory networks that may serve as potential molecular markers or targets for genetic improvement programs focusing on enhancing pork quality while maintaining production efficiency. Furthermore, this study contributes to the broader understanding of the genetic architecture governing muscle development in divergent pig breeds and provides valuable resources for future molecular breeding strategies aimed at optimizing meat quality traits in pigs.
2. Materials and Methods
2.1. Animals and Sample Collection
Ten male pigs each from Congjiang Xiang (CX) and Landrace (LAN) breeds were used in this study. The pigs were raised under the same similar altitudes and shared similar natural climatic conditions in Congjiang County, Qiandongnan Miao and Dong Autonomous Prefecture, Guizhou Province. All animals were slaughtered at 12 months of age with an average body weight of 74.95±18.81 kg (CX, mean±SD) and 189.95±40.95 kg (LAN, mean±SD). The longissimus dorsi (LD) muscle tissues were immediately collected from the carcasses, snap-frozen in liquid nitrogen, and stored at -80°C until RNA extraction. All the animal experiments in the present study strictly complied with the relevant regulations regarding the care and use of experimental animals issued by the Academic Committee of Kaili University (Approval ID: 202402).
2.2. RNA Extraction, Library Construction, and Sequencing
Total RNA was isolated using TRIzol reagent (Invitrogen, USA) according to the manufacturer’s protocol. RNA purity and concentration were assessed using a NanoDrop ND-2000 spectrophotometer (NanoDrop Technologies, USA), and RNA integrity was checked by 1% agarose gel electrophoresis. The RNA integrity number (RIN) was determined using an Agilent 2100 Bioanalyzer (Agilent Technologies, USA), and only samples with RIN ≥ 7.5 were used for library construction.
Sequencing libraries were generated using the TruSeq RNA Sample Preparation Kit (Illumina, USA) following the manufacturer’s instructions. Briefly, mRNA was purified from total RNA using oligo(dT) magnetic beads. The mRNA was then fragmented and used for first-strand cDNA synthesis with random hexamer primers and reverse transcriptase. Second-strand cDNA synthesis was subsequently performed using DNA polymerase I and RNase H. The double-stranded cDNA was purified and subjected to end repair, A-tailing and adapter ligation. After size selection using AMPure XP beads (Beckman Coulter, USA), the libraries were PCR-amplified and purified. Library quality was assessed on the Agilent Bioanalyzer 2100 system. The clustering of the index-coded samples was performed on a cBot Cluster Generation System using TruSeq PE Cluster Kit v3-cBot-HS (Illumina, USA). The libraries were sequenced on an Illumina NovaSeq 6000 platform, and 150-bp paired-end reads were generated.
2.3. Read Mapping and Differential Expression Analysis
Raw reads in FASTQ format were first processed using Trimmomatic (v0.36) to remove adapter sequences, low-quality reads (Q < 20), and short reads (< 50 bp). The cleaned reads were then aligned to the pig reference genome (Sscrofa 11.1) using HISAT2 (v2.1.0) with default parameters. StringTie (v1.3.5) was used to calculate the fragments per kilobase of transcript per million mapped reads (FPKM) value for each gene. Differential expression analysis between CX and LAN pigs was performed using the DESeq2 R package (v1.24.0). Genes with |log2(fold change)| ≥ 1 and adjusted p-value < 0.05 were considered as differentially expressed genes (DEGs).
2.4. Functional Enrichment and Pathway Analysis
Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses of DEGs were performed using the clusterProfiler R package (v3.12.0). GO terms and KEGG pathways with adjusted p-value < 0.05 were considered significantly enriched.
2.5. Protein–Protein Interaction (PPI) Network Analysis
The PPI network for DEGs was constructed using the STRING database (v11.0) with a confidence score ≥ 0.9. Cytoscape (v3.7.2) was used to visualize the PPI network.
2.6. Validation by Quantitative Real-Time PCR (qRT-PCR)
Ten DEGs (five up-regulated and five down-regulated in CX pigs) were randomly selected for qRT-PCR validation. The qRT-PCR primers (Table S1) were designed using Primer Premier 5.0 software. The qRT-PCR analysis was performed on a LightCycler 480 II Real-time PCR System (Roche, Switzerland) using TB Green Premix Ex Taq II (Takara, Japan) according to the manufacturer’s instructions. The porcine GAPDH gene was used as an internal control. The relative expression levels of the selected DEGs were calculated using the 2−ΔΔCt method. Statistical analysis was performed using Student’s t-test in SPSS 22.0 software (IBM, USA), and p-value < 0.05 was considered statistically significant.
3. Results
3.1. Overview of RNA Sequencing Data
To systematically investigate the transcriptome differences between Congjiang Xiang (CX) and Landrace (LAN) pigs, we performed RNA sequencing on longissimus dorsi (LD) muscle tissues from ten individuals of each breed. A total of 874.5 million raw reads were generated, with an average of 43.7 million reads per sample. After quality control, 846.0 million clean reads were obtained and mapped to the pig reference genome (Sscrofa 11.1), with an average mapping rate of 89.3% (Table S2). The co-expression Venn diagram analysis revealed distinct transcriptional profiles between the two porcine breeds, with 785 genes being exclusively expressed in CX pigs and 457 genes uniquely expressed in LAN pigs. Notably, the intersection region demonstrated a core set of 7,099 co-expressed genes in the LD muscle shared by both breeds (Figure 1A). Principal component analysis (PCA) based on the expression profiles showed a clear separation between CX and LAN pigs (Figure 1B), indicating distinct transcriptome landscapes in the LD muscle between the two breeds.
3.2. Identification of Differentially Expressed Genes
Differential expression analysis identified a total of 2459 differentially expressed genes (DEGs) between CX and LAN pigs, including 1745 up-regulated and 714 down-regulated genes in CX pigs (Figure 2A). Hierarchical clustering analysis revealed that the DEGs could clearly distinguish the two breeds (Figure 2B), suggesting their potential roles in driving the phenotypic differences. The details of the top 10 up- and down-regulated DEGs are shown in Table 1 and Table 2, respectively. Notably, regulatory factors known to be closely associated with meat flavor formation, such as ELOVL5/6, FASN, DGAT2, ALDH1A3, MGST1, MSTN, THRSP, PPAR-γ, PFKFB3, and ACSL4/5, were significantly up-regulated in CX pigs (Figure 2C), implying enhanced meat flavor characteristics in this breed.
3.3. Functional Enrichment Analysis of DEGs
To gain insights into the biological functions of the DEGs, we performed Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses. GO analysis revealed that the up-regulated genes in CX pigs were significantly enriched in immune response, defense response, regulation of immune system process, positive regulation of immune system process, and response to external stimulus (Figure 3A–C). In contrast, the down-regulated genes were mainly involved in myosin complex, proton-transporting V-type ATPase complex, and actin cytoskeleton. KEGG pathway analysis showed that the up-regulated genes were highly enriched in pathways related to viral protein interaction with cytokine and cytokine receptor, hematopoietic cell lineage, chemokine signaling pathway, cytokine-cytokine receptor interaction, and cell adhesion molecules (Figure 2B). The down-regulated genes were significantly associated with PPAR signaling pathway and glycerolipid metabolism (Figure 3B). These findings may contribute to the superior meat flavor characteristics in CX pigs compared to LAN pigs.
3.4. PPI Network Analysis of DEGs
To further explore the functional interactions among the DEGs, we constructed a protein-protein interaction (PPI) network using the STRING database. The PPI network contained 1306 nodes and 138 edges (Figure 4A). PPAR-γ was identified as hub genes in the network (Figure 4B), indicating their central regulatory roles in meat flavor formation. Moreover, several key genes (e.g., ACTG1, ACTB, IGF1) were also found to be closely connected in the network, suggesting coordinated regulation of these processes.
3.5. Validation of DEGs by qRT-PCR
All designed primers demonstrated good amplification efficiency and specificity. The qRT-PCR validation results revealed that the expression patterns of all ten selected genes were highly consistent with the RNA-Seq analysis findings, with consistency rates ranging from 95.9% to 99.6% (Table 3). In CX pigs, AP1S2, FCGR3A, SLA-DQA, CD37, and EMP2 genes were significantly up-regulated, while VPS72, UBL5, GDE1, AQP7, and CUL1 genes were significantly down-regulated (p<0.001). These results confirm the reliability of our transcriptome analysis findings.
4. Discussion
In the present study, we performed RNA-Seq analysis to identify differentially expressed genes in the longissimus dorsi muscle between Chinese Congjiang Xiang and Landrace pig breeds. Our results revealed substantial transcriptional differences between these breeds, with 2459 differentially expressed genes (DEGs) identified, including 1745 up-regulated and 714 down-regulated genes in CX pigs compared to LAN pigs. Principal component analysis and hierarchical clustering analysis demonstrated distinct transcriptional profiles that clearly separated the two breeds, highlighting the significant genetic divergence between indigenous Chinese and commercial European pig breeds. We observed breed-specific gene expression patterns, with 785 genes exclusively expressed in CX pigs and 457 genes unique to LAN pigs, suggesting their potential contributions to phenotypic differences. Notably, several regulatory factors associated with meat flavor development, including ELOVL5/6, FASN, DGAT2, ALDH1A3, MGST1, MSTN, THRSP, PFKFB3, and ACSL4/5, were significantly up-regulated in CX pigs, which may explain the enhanced flavor characteristics typically associated with this indigenous breed.
The distinct transcriptional profiles observed between CX and LAN pigs provide valuable insights into the molecular basis of breed-specific characteristics. The 785 genes exclusively expressed in CX pigs may represent a unique genetic signature that contributes to the distinctive traits of this indigenous Chinese breed. Indigenous pig breeds have evolved under diverse environmental conditions and selective pressures that differ from those experienced by commercial breeds, leading to genetic adaptations reflected in their transcriptional landscapes. Similar breed-specific expression patterns have been reported in previous studies comparing indigenous and commercial pig breeds. For instance, Yang et al. (2024) identified unique expression patterns in Tibetan pigs that contributed to their adaptation to high-altitude environments [22], while Li et al. (2019) reported breed-specific transcriptional signatures in Meishan pigs associated with their superior fertility traits [23]. Our findings add to this growing body of evidence suggesting that breed-specific gene expression patterns underlie the phenotypic divergence between indigenous and commercial pig breeds, highlighting the value of indigenous genetic resources for understanding the molecular basis of economically important traits.
The up-regulation of flavor-related genes in CX pigs represents one of the most significant findings of our study. Genes involved in lipid metabolism, including ELOVL5/6, FASN, and DGAT2, play crucial roles in the biosynthesis and modification of fatty acids that contribute to meat flavor and tenderness [24]. ELOVL5/6 encode elongases that catalyze the elongation of fatty acids, while FASN is a key enzyme in de novo fatty acid synthesis [25]. DGAT2 catalyzes the final step in triglyceride synthesis, contributing to intramuscular fat deposition [26]. The elevated expression of these genes in CX pigs suggests enhanced lipid metabolism and fat deposition capabilities, which align with the higher intramuscular fat content typically observed in indigenous Chinese breeds. Previous studies have demonstrated that increased intramuscular fat content positively correlates with meat tenderness, juiciness, and flavor [27]. Similarly, the up-regulation of ALDH1A3, which participates in the metabolism of aldehydes derived from lipid oxidation, and MGST1, involved in glutathione metabolism and oxidative stress response, may contribute to the development of flavor compounds during meat aging and cooking [28]. These findings are consistent with those reported by Wu et al. (2023), who observed enhanced expression of lipid metabolism genes in Jinhua pigs compared to intensive pig breeds, suggesting a common molecular mechanism underlying the superior meat quality of indigenous Chinese breeds [29].
The enrichment of immune-related functions among up-regulated genes in CX pigs represents an intriguing finding that warrants further investigation. Our GO analysis revealed significant enrichment of terms related to immune response, defense response, regulation of immune system process, and response to external stimulus among the up-regulated genes in CX pigs. This enhanced expression of immune-related genes may reflect the adaptation of indigenous breeds to more diverse and challenging environmental conditions compared to commercial breeds raised in controlled environments. Several studies have suggested potential connections between immune functions and meat quality traits. For instance, Pothakam et al. (2021) reported that inflammatory responses influence muscle fiber characteristics and intramuscular fat deposition in pigs [30]. The elevated expression of immune-related genes in CX pigs may contribute to their distinctive muscle fiber composition and fat distribution patterns, indirectly affecting meat quality. Additionally, indigenous breeds like CX pigs have been subjected to natural selection for disease resistance over centuries, which may have shaped their immune gene expression profiles. Gu et al. (2023) observed similar patterns of immune gene up-regulation in Tibetan pigs, suggesting that enhanced immune functions may be a common feature of indigenous pig breeds that have evolved under less controlled environmental conditions than commercial breeds [31].
The identification of PPAR-γ as a hub gene in our protein-protein interaction network analysis highlights its central role in regulating the transcriptional differences between CX and LAN pigs. PPAR-γ is a master regulator of adipocyte differentiation and lipid metabolism, influencing the expression of numerous genes involved in fatty acid uptake, transport, and storage [32,33]. Interestingly, while PPAR-γ itself was up-regulated in CX pigs, our KEGG pathway analysis revealed that the PPAR signaling pathway was significantly associated with down-regulated genes. This apparent contradiction may reflect complex regulatory mechanisms involving negative feedback loops or differential regulation of specific components within the pathway. The increased expression of PPAR-γ in CX pigs likely contributes to their enhanced intramuscular fat deposition and subsequent flavor development. Previous studies have demonstrated that PPAR-γ activation promotes intramuscular adipogenesis and improves meat quality in pigs [34,35]. Our findings suggest that differential regulation of PPAR-γ and its downstream targets may be a key mechanism underlying the meat quality differences between indigenous and commercial pig breeds.
The enrichment of cytokine and chemokine signaling pathways among up-regulated genes in CX pigs suggests potential roles of inflammatory processes in muscle development and meat quality. Our KEGG pathway analysis revealed significant enrichment of pathways related to viral protein interaction with cytokine and cytokine receptor, cytokine-cytokine receptor interaction, and chemokine signaling pathway. Cytokines and chemokines are signaling molecules that regulate inflammatory responses, but they also participate in various physiological processes, including tissue development and remodeling [36]. In muscle tissue, inflammatory signaling can influence satellite cell activation, myoblast proliferation and differentiation, and extracellular matrix remodeling, all of which affect muscle growth and meat quality [37]. The enhanced expression of genes involved in these pathways in CX pigs may contribute to their distinctive muscle fiber characteristics and intramuscular fat distribution. Ren et al. (2024) reported similar enrichment of inflammatory pathways in the muscle transcriptome of Wuzhishan pigs, suggesting that modulation of inflammatory processes might be a common feature of indigenous pig breeds [38]. Furthermore, certain inflammatory mediators have been shown to influence lipid metabolism, potentially contributing to the development of flavor precursors [39]. These findings highlight the complex interplay between inflammatory signaling, muscle development, and meat quality traits in different pig breeds.
While our study provides valuable insights into the transcriptional differences between CX and LAN pigs, several limitations should be acknowledged. First, our analysis was based on a relatively small sample size of ten individuals per breed, which may limit the statistical power to detect subtle expression differences. Second, we focused exclusively on the longissimus dorsi muscle, while transcriptional differences in other tissues might also contribute to breed-specific characteristics. Different muscles exhibit distinct fiber composition and metabolic profiles, which could result in tissue-specific gene expression patterns [40]. Third, our study represents a snapshot of gene expression at a single time point, while temporal changes in gene expression during development might provide additional insights into breed differences. Finally, environmental and developmental factors that might influence gene expression were not fully controlled in our study design. Despite these limitations, our study provides a robust foundation for understanding the molecular basis of meat quality differences between indigenous and commercial pig breeds.
In conclusion, our RNA-Seq analysis revealed significant transcriptional differences in the longissimus dorsi muscle between Chinese Congjiang Xiang and Landrace pig breeds. The identified DEGs, particularly those involved in lipid metabolism, immune response, and inflammatory signaling, provide insights into the molecular mechanisms underlying the superior meat flavor characteristics associated with indigenous Chinese breeds. The up-regulation of flavor-related genes, including ELOVL5/6, FASN, DGAT2, and PPAR-γ, in CX pigs suggests enhanced lipid metabolism and fat deposition capabilities that contribute to their distinctive meat quality traits. Our findings highlight the value of indigenous genetic resources for understanding the molecular basis of economically important traits and improving commercial pig breeds. The comprehensive transcriptional profile provided by our study could guide future breeding programs aimed at enhancing meat quality while maintaining production efficiency, ultimately benefiting both producers and consumers in the pork industry.
5. Conclusions
In conclusion, our RNA-Seq analysis revealed significant transcriptional differences between the longissimus dorsi muscles of Chinese Congjiang Xiang and Landrace pig breeds, providing molecular insights into their distinctive meat quality characteristics. The identification of 2,459 differentially expressed genes, with 1,745 up-regulated and 714 down-regulated in Congjiang Xiang pigs, highlights the complex genetic architecture underlying breed-specific traits. Particularly noteworthy is the up-regulation of key flavor-associated genes (ELOVL5/6, FASN, DGAT2, ALDH1A3,) in Congjiang Xiang pigs, which likely contributes to their enhanced meat flavor profiles. The functional enrichment patterns—immune and defense responses in up-regulated genes and structural muscle components in down-regulated genes—suggest a potential trade-off between flavor development and muscle structure that differentiates these breeds. The identification of PPAR-γ as a hub gene further emphasizes its central regulatory role in coordinating lipid metabolism pathways critical for meat quality formation. These findings not only enhance our understanding of the molecular basis for meat quality differences between indigenous Chinese and commercial European pig breeds but also provide valuable candidate genes for marker-assisted selection programs aimed at improving pork quality while preserving the unique genetic resources represented by local breeds like Congjiang Xiang pigs.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, K.Y. and J.Y.; methodology, K.Y. and J.Y.; software, J.Y. and Q.T.; validation, K.Y. and J.Y.; formal analysis, J.Y. and Q.T.; investigation, C.S., Q.L. and X.L.; resources, K.Y.; data curation, K.Y. and J.Y.; writing—original draft preparation, J.Y. and K.Y.; writing—review and editing, K.Y.; visualization, J.Y. and L.H.; supervision, K.Y.; project administration, K.Y.; funding acquisition, K.Y., L.H. and Y.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Youth Science and Technology Talent Growth Project of Guizhou Provincial Department of Education (No. Qian Jiaoji [2024]225), Kaili University Doctoral Research Special Project (BS20240215), Kaili University Doctoral Development Special Research Project (BSFZ202302), the Guizhou Provincial Basic Research Program (No. QHKJC [2020]1Z028), and the Youth Science and Technology Talent Growth Project of Guizhou Provincial Department of Education (No. Qian Jiaohe KY Zi [2022]076).
Institutional Review Board Statement
All the animal experiments in the present study strictly complied with the relevant regulations regarding the care and use of experimental animals issued by the Academic Committee of Kaili University (Approval ID: 202402).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kim, S.W.; Gormley, A.; Jang, K.; Duarte, M. Current status of global pig production: an overview and research trends. Animal Bioscience 2023, 37, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Zhang, P.; Yin, J.; Zhang, X. New insights into the effects of dietary amino acid composition on meat quality in pigs: A review. Meat science 2024, 221, 109721. [Google Scholar] [CrossRef] [PubMed]
- Aboah, J.; Lees, N. Consumers use of quality cues for meat purchase: Research trends and future pathways. Meat science 2020, 166, 108142. [Google Scholar] [CrossRef]
- Chen, G.; Cai, Y.; Su, Y.; Wang, D.; Pan, X.; Zhi, X. Study of meat quality and flavour in different cuts of Duroc-Bamei binary hybrid pigs. Veterinary Medicine and Science 2020, 7, 724–734. [Google Scholar] [CrossRef]
- Fu, Q.-Q.; Shi, H.; Hu, D.; Cheng, J.; Chen, S.; Ben, A. Pork longissimus dorsi marinated with edible mushroom powders: Evaluation of quality traits, microstructure, and protein degradation. Food research international 2022, 158, 111503. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.T.; Zheng, Y.; Xu, J.; Zhao, Z.; Zhang, Q.; Zhang, Y.; Li, M.; Zou, H.; Azeem, R.-M.; Sun, W.-S.; et al. Transcriptome and metabolome insights into key genes regulating fat deposition and meat quality in pig breeds. Animals 2024, 14, 3560. [Google Scholar] [CrossRef]
- Sharma, P.; Doultani, S.; Hadiya, K.; George, L.; Highland, H. Overview of marker-assisted selection in animal breeding. Journal of Advances in Biology & Biotechnology 2024, 27, 303–318. [Google Scholar] [CrossRef]
- Meuwissen, T.; Hayes, B.; Goddard, M. Genomic selection: A paradigm shift in animal breeding. Animal Frontiers 2016, 6, 6–14. [Google Scholar] [CrossRef]
- Šimon, M.; Bogićević, S.; Kaić, A.; Luštrek, B.; Potočnik, K. Exploring genetic influences on equine meat quality: A bioinformatics approach. Foods 2025, 14, 533. [Google Scholar] [CrossRef]
- Cesar, A.; Regitano, L.; Reecy, J.; Poleti, M.; Oliveira, P.; De Oliveira, G.; Moreira, G.; Mudadu, M.; Tizioto, P.; Koltes, J.; et al. Identification of putative regulatory regions and transcription factors associated with intramuscular fat content traits. BMC Genomics 2018, 19, 499. [Google Scholar] [CrossRef]
- Hasan, M.; Feugang, J.; Liao, S. A nutrigenomics approach using RNA sequencing technology to study nutrient–gene interactions in agricultural animals. Current Developments in Nutrition 2019, 3, nzz082. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.-Z.; Wu, J.; Zhou, J.; Zhang, Y.; Qiao, M.; Sun, H.; Li, Z.; Li, L.; Chen, N.; Oyelami, F.; et al. Integration of ATAC-seq and RNA-seq analysis identifies key genes affecting intramuscular fat content in pigs. Frontiers in Nutrition 2022, 9, 1016956. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, A.; Óvilo, C.; Núñez, Y.; Benítez, R.; López-García, A.; García, F.; Félix, M.; Laranjo, M.; Charneca, R.; Martins, J. Transcriptomic profiling of skeletal muscle reveals candidate genes influencing muscle growth and associated lipid composition in portuguese local pig breeds. Animals 2021, 11, 1423. [Google Scholar] [CrossRef]
- Ropka-Molik, K.; Żukowski, K.; Eckert, R.; Gurgul, A.; Piórkowska, K.; Oczkowicz, M. Comprehensive analysis of the whole transcriptomes from two different pig breeds using RNA-Seq method. Animal genetics 2014, 45, 674–684. [Google Scholar] [CrossRef]
- Liu, W.; Gong, T.; Xu, Y. The co-expression of steroidogenic enzymes with T1R3 during testicular development in the Congjiang Xiang pig. Animal reproduction science 2023, 251, 107216. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Chen, Z.; Ruan, Y.; Xu, H. Differential regulatory roles of microRNAs during intramuscular adipogenesis in Chinese Guizhou Congjiang Xiang pigs. Epigenetics 2022, 17, 1800–1819. [Google Scholar] [CrossRef]
- Guo, J.; Shan, T.; Wu, T.; Zhu, L.; Ren, Y.; An, S.; Wang, Y. Comparisons of different muscle metabolic enzymes and muscle fiber types in Jinhua and Landrace pigs. Journal of animal science 2011, 89 1, 185–191. [Google Scholar] [CrossRef]
- Kasprzyk, A.; Tyra, M.; Babicz, M. Fatty acid profile of pork from a local and a commercial breed. Archives Animal Breeding 2015, 58, 379–385. [Google Scholar] [CrossRef]
- Vidal, O.; Noguera, J.; Amills, M.; Varona, L.; Gil, M.; Jiménez, N.; Dávalos, G.; Folch, J.; Sánchez, A. Identification of carcass and meat quality quantitative trait loci in a Landrace pig population selected for growth and leanness. Journal of animal science 2005, 83, 293–300. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, Z.-Y.; Liu, Q.-X. Gene coexpression networks reveal key drivers of phenotypic divergence in porcine muscle. BMC Genomics 2015, 16, 50. [Google Scholar] [CrossRef]
- Tan, L.; Chen, Z.; Teng, M.; Chen, B.; Xu, H. Genome-wide analysis of mRNAs, lncRNAs, and circRNAs during intramuscular adipogenesis in Chinese Guizhou Congjiang pigs. PLoS ONE 2022, 17, e0261293. [Google Scholar] [CrossRef]
- Yang, Y.; Yuan, H.; Yao, B.; Zhao, S.; Wang, X.; Xu, L.; Zhang, L. Genetic adaptations of the Tibetan pig to high-altitude hypoxia on the Qinghai–Tibet plateau. International Journal of Molecular Sciences 2024, 25, 11303. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Yang, Y.; Liu, Y.; Chen, J.; Yang, B.; Tang, Z. High serum reproductive hormone levels at mid-pregnancy support Meishan pig prolificacy. Journal of Integrative Agriculture 2023, 22, 3489–3499. [Google Scholar] [CrossRef]
- Renaville, B.; Bacciu, N.; Lanzoni, M.; Mossa, F.; Piasentier, E. Association of single nucleotide polymorphisms in fat metabolism candidate genes with fatty acid profiles of muscle and subcutaneous fat in heavy pigs. Meat science 2018, 139, 220–227. [Google Scholar] [CrossRef]
- Jakobsson, A.; Westerberg, R.; Jacobsson, A. Fatty acid elongases in mammals: their regulation and roles in metabolism. Progress in lipid research 2006, 45, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Chitraju, C.; Walther, T.; Farese, R. The triglyceride synthesis enzymes DGAT1 and DGAT2 have distinct and overlapping functions in adipocytes[S]. Journal of Lipid Research 2019, 60, 1112–1120. [Google Scholar] [CrossRef]
- Yu, T.-Y.; Tian, X.; Li, D.; He, Y.; Yang, P.; Cheng, Y.; Zhao, X.; Sun, J.; Yang, G. Transcriptome, proteome and metabolome analysis provide insights on fat deposition and meat quality in pig. Food research international 2023, 166, 112550. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Cao, S.; Yang, L.; Li, Z. Flavor formation based on lipid in meat and meat products: A review. Journal of food biochemistry 2022, 46, 14439. [Google Scholar] [CrossRef]
- Wu, F.; Chen, Z.; Zhang, Z.; Wang, Z.; Zhang, Z.; Wang, Q.; Pan, Y. The role of SOCS3 in regulating meat quality in Jinhua pigs. International Journal of Molecular Sciences 2023, 24, 10593. [Google Scholar] [CrossRef]
- Pothakam, N.; Supakankul, P.; Norseeda, W.; Liu, G.; Teltathum, T.; Naraballobh, W.; Khamlor, T.; Sringarm, K.; Mekchay, S. Association of adipocytokine IL-1A and IL-6 genes with intramuscular fat content and fatty acid composition in pigs. Meat science 2021, 179, 108554. [Google Scholar] [CrossRef]
- Gu, X.; Chang, X.; Yang, L.; Chamba, Y.; Geng, F. Quantitative proteomic analysis of Tibetan pig livers at different altitudes. Molecules 2023, 28, 1694. [Google Scholar] [CrossRef]
- Qiu, Y.; Gan, M.; Wang, X.; Liao, T.; Chen, Q.; Lei, Y.; Chen, L.; Wang, J.; Zhao, Y.; Niu, L.; et al. The global perspective on peroxisome proliferator-activated receptor γ (PPARγ) in ectopic fat deposition: A review. International journal of biological macromolecules 2023, 127042. [Google Scholar] [CrossRef]
- Janani, C.; Kumari, R. PPAR gamma gene--A review. Diabetes & metabolic syndrome 2015, 9, 46–50. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.-L.; Wu, L.; Guo, Y.; Yang, G.; Li, X.; Shi, X.e. Rosiglitazone-induced PPARγ activation promotes intramuscular adipocyte adipogenesis of pig. Animal Biotechnology 2023, 34, 3708–3717. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Zhou, Y.; Yang, J.; Li, J.; Peng, Y.; Zhang, X.; Miao, Y.; Jiang, W.; Bu, G.; Hou, L.; et al. Targeted overexpression of PPARγ in skeletal muscle by random insertion and CRISPR/Cas9 transgenic pig cloning enhances oxidative fiber formation and intramuscular fat deposition. The FASEB Journal 2021, 35, e21308. [Google Scholar] [CrossRef] [PubMed]
- Commins, S.; Borish, L.; Steinke, J. Immunologic messenger molecules: cytokines, interferons, and chemokines. The Journal of allergy and clinical immunology 2010, 125, S53–S72. [Google Scholar] [CrossRef] [PubMed]
- Blanc, R.; Kallenbach, J.; Bachman, J.; Mitchell, A.; Paris, N.; Chakkalakal, J. Inhibition of inflammatory CCR2 signaling promotes aged muscle regeneration and strength recovery after injury. Nature Communications 2020, 11, 4167. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, F.; Sun, R.; Zheng, X.; Liu, Y.; Lin, Y.; Hong, L.; Huang, X.; Chao, Z. The genetic selection of HSPD1 and HSPE1 reduce inflammation of liver and spleen while restraining the growth and development of skeletal muscle in Wuzhishan pigs. Animals 2024, 14, 174. [Google Scholar] [CrossRef]
- Nara, H.; Watanabe, R. Anti-inflammatory effect of muscle-derived interleukin-6 and its involvement in lipid metabolism. International Journal of Molecular Sciences 2021, 22, 9889. [Google Scholar] [CrossRef]
- Wang, W.; Li, W.; Liu, W.; Wang, Z.; Xie, B.; Yang, X.; Tang, Z. Exploring multi-tissue alternative splicing and skeletal muscle metabolism regulation in obese- and lean-type pigs. Genes 2024, 15, 196. [Google Scholar] [CrossRef]
Figure 1.
Transcriptome Comparison of CX and LAN Pigs: (A) Co-Expressed and Breed-Specific Genes in LD Muscle, (B) PCA-Based Separation of Transcriptional Profiles.
Figure 1.
Transcriptome Comparison of CX and LAN Pigs: (A) Co-Expressed and Breed-Specific Genes in LD Muscle, (B) PCA-Based Separation of Transcriptional Profiles.
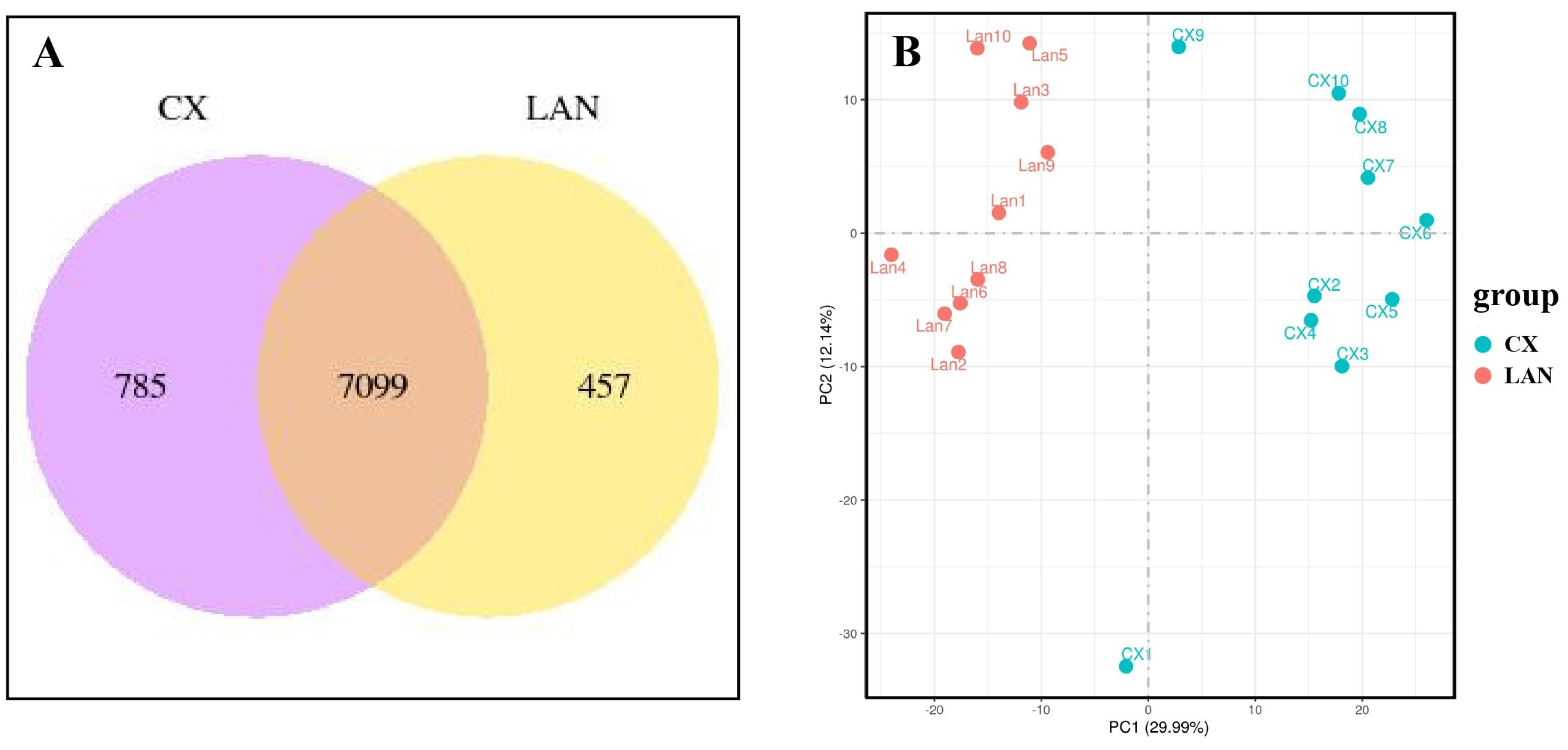
Figure 2.
Differentially Expressed Genes (DEGs) Between CX and LAN Pigs: (A) Volcano Plot of DEGs, (B) Hierarchical Clustering, (C) Upregulated Flavor-Related Genes in CX Pigs.
Figure 2.
Differentially Expressed Genes (DEGs) Between CX and LAN Pigs: (A) Volcano Plot of DEGs, (B) Hierarchical Clustering, (C) Upregulated Flavor-Related Genes in CX Pigs.
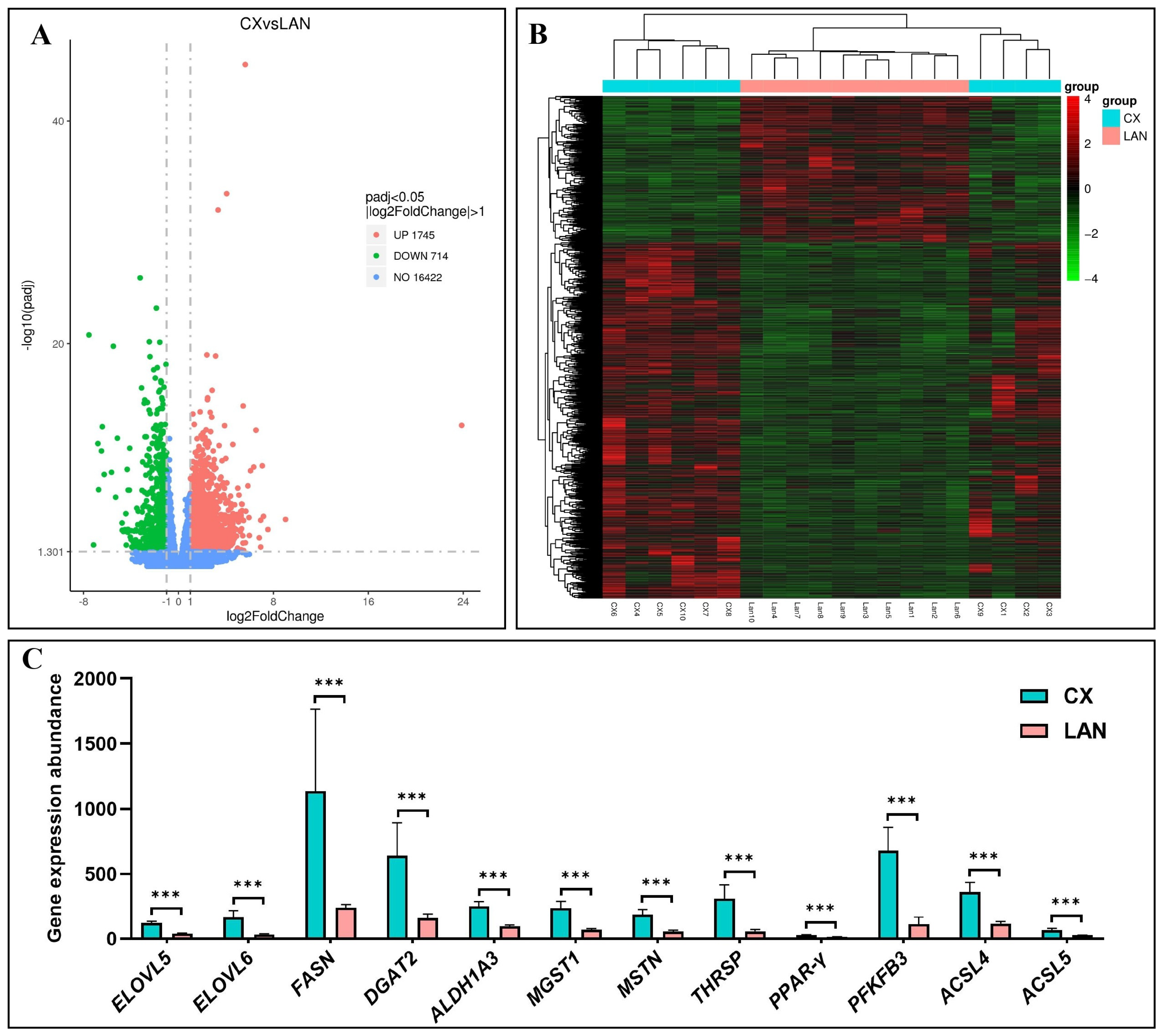
Figure 3.
Functional Enrichment Analysis of DEGs: (A) BP (GO), (B) MF (GO), (C) CC (GO), and (D) KEGG pathways.
Figure 3.
Functional Enrichment Analysis of DEGs: (A) BP (GO), (B) MF (GO), (C) CC (GO), and (D) KEGG pathways.
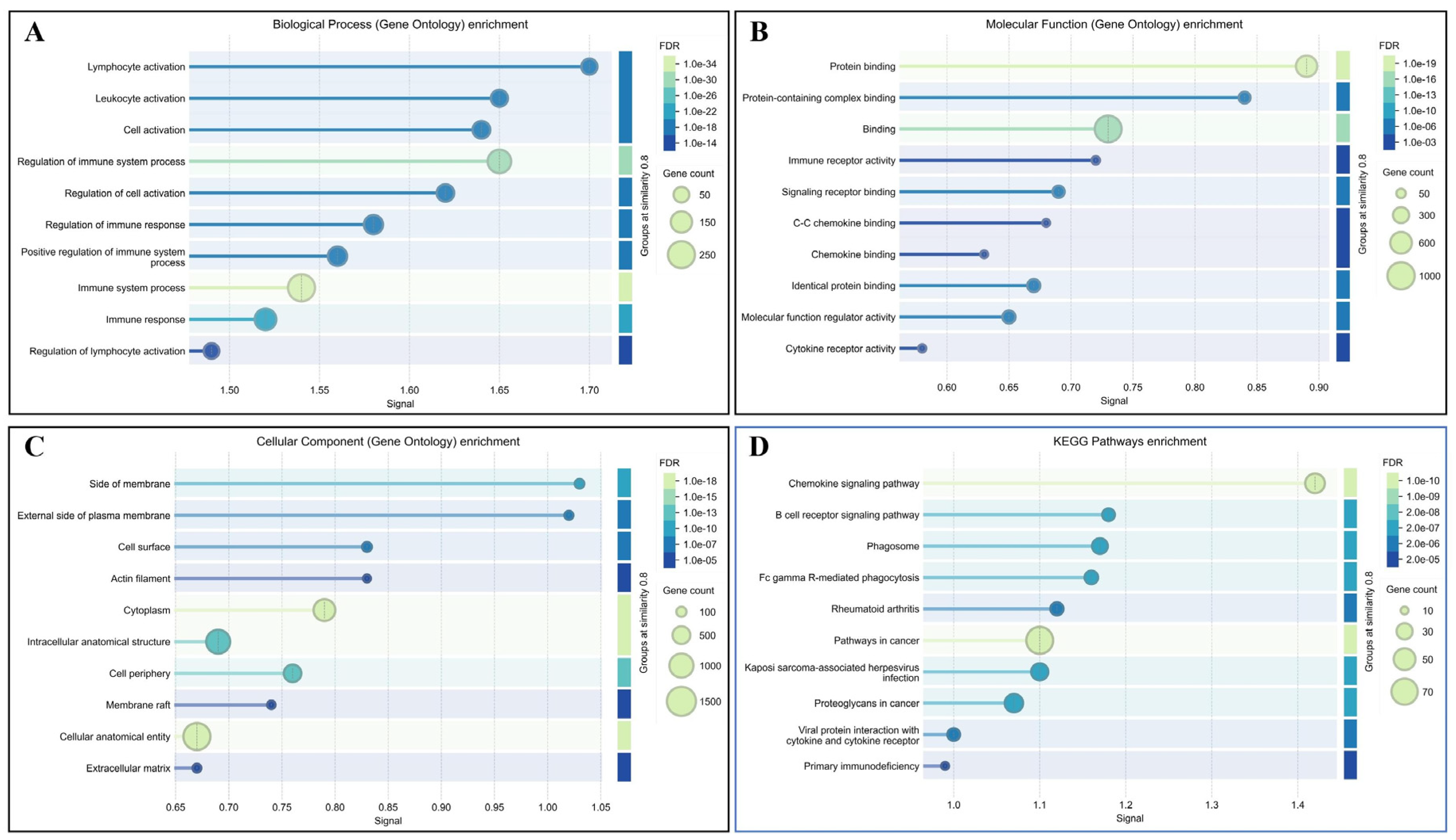
Figure 4.
Prediction of protein interaction between genes related to meat flavor formation and candidate genes.
Figure 4.
Prediction of protein interaction between genes related to meat flavor formation and candidate genes.
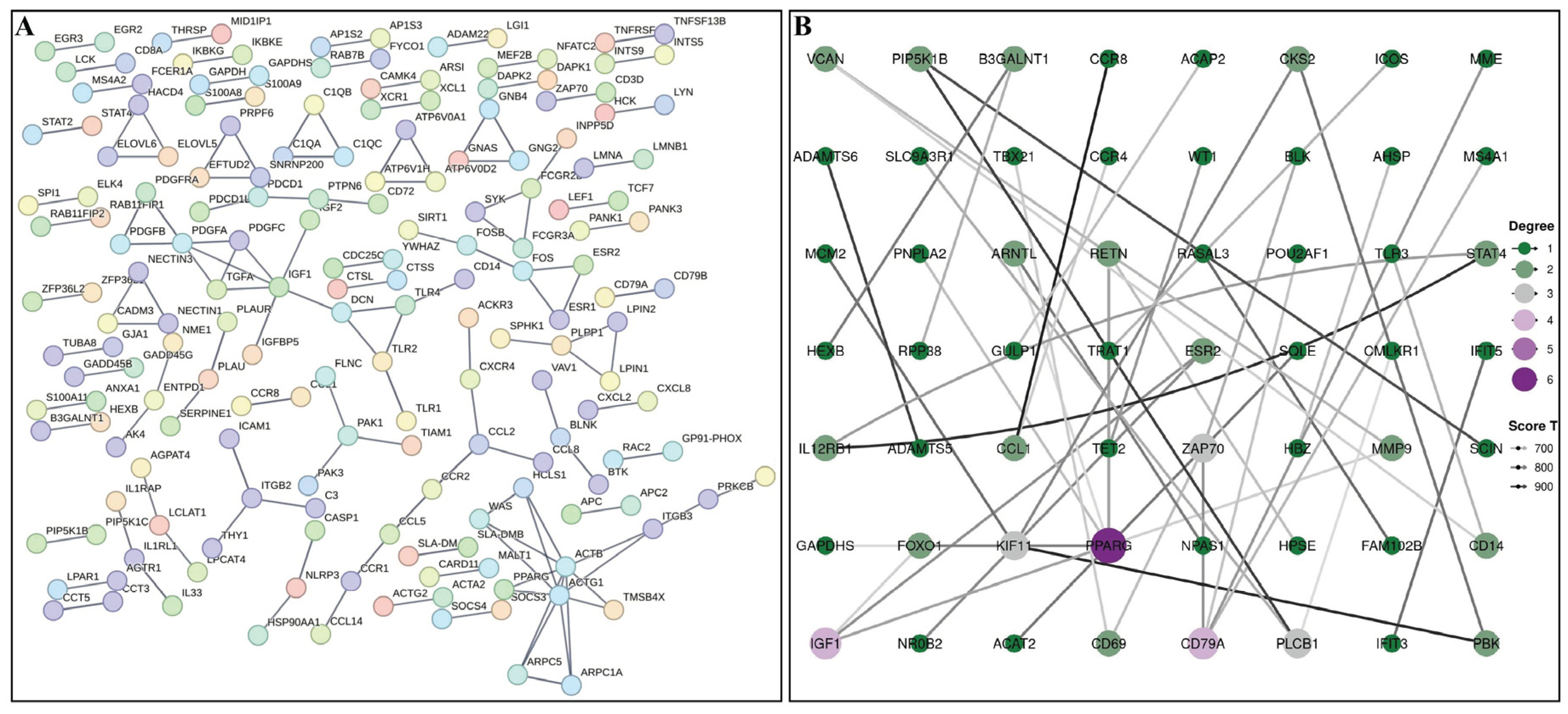

Table 1.
The top 10 up-regulated DEGs in Longissimus dorsi in CX and LAN pigs.
| Gene_name | Gene_id | CX | LAN | Log2FC | Adjusted p-value |
| AP1S2 | ENSSSCG00045031746 | 414.59 | 41.03 | 3.33 | 1.00e-32 |
| FCGR3A | ENSSSCG00045036350 | 264.93 | 50.60 | 2.39 | 1.05e-19 |
| SLA-DQA | ENSSSCG00045011704 | 749.70 | 105.10 | 2.83 | 1.59e-16 |
| CD37 | ENSSSCG00045013461 | 47.82 | 8.73 | 2.45 | 7.50e-16 |
| EMP2 | ENSSSCG00045000984 | 477.11 | 134.64 | 1.82 | 1.40e-14 |
| ARMCX2 | ENSSSCG00045011080 | 47.47 | 7.32 | 2.67 | 1.55e-14 |
| CHPT1 | ENSSSCG00045032049 | 2151.92 | 928.29 | 1.21 | 2.16e-14 |
| C5AR1 | ENSSSCG00045034745 | 102.66 | 14.72 | 2.80 | 4.14e-14 |
| DNASE1L3 | ENSSSCG00045035470 | 49.55 | 3.38 | 3.88 | 2.36e-13 |
| CD53 | ENSSSCG00045035517 | 183.79 | 36.60 | 2.33 | 5.59e-13 |
Table 2.
The top 10 down-regulated DEGs in Longissimus dorsi in CX and LAN pigs.
| Gene_name | Gene_id | CX | LAN | Log2FC | Adjusted p-value |
| VPS72 | ENSSSCG00045022741 | 471.19 | 1721.26 | -1.86 | 6.47e-24 |
| UBL5 | ENSSSCG00045039454 | 207.27 | 1137.93 | -2.45 | 6.84e-21 |
| GDE1 | ENSSSCG00045015707 | 616.04 | 1837.79 | -1.57 | 7.50e-21 |
| AQP7 | ENSSSCG00045001378 | 41.03 | 217.30 | -2.40 | 1.49e-19 |
| CUL1 | ENSSSCG00045021962 | 456.96 | 949.08 | -1.05 | 7.38e-19 |
| DUSP27 | ENSSSCG00045009424 | 740.94 | 2549.34 | -1.78 | 1.46e-18 |
| LYSMD2 | ENSSSCG00045031255 | 103.88 | 403.52 | -1.95 | 1.26e-17 |
| DDX24 | ENSSSCG00045022939 | 150.26 | 411.68 | -1.45 | 2.73e-17 |
| LRRC42 | ENSSSCG00045022986 | 321.70 | 889.54 | -1.46 | 3.18e-17 |
| PDZD9 | ENSSSCG00045028818 | 181.87 | 1576.16 | -3.11 | 9.77e-17 |
Table 3.
Relative expression levels of selected differentially expressed genes validated by qRT-PCR.
Table 3.
Relative expression levels of selected differentially expressed genes validated by qRT-PCR.
| Gene name | CX | LAN | Fold change (CX/LAN) | Log2FC | p-value | RNA-Seq Log2FC | Consistency |
| AP1S2 | 9.87±1.45 | 1.06±0.21 | 9.31 | 3.22 | <0.001 | 3.33 | 96.70% |
| FCGR3A | 5.64±0.82 | 1.03±0.18 | 5.48 | 2.45 | <0.001 | 2.39 | 97.50% |
| SLA-DQA | 7.42±1.16 | 1.05±0.22 | 7.07 | 2.82 | <0.001 | 2.83 | 99.60% |
| CD37 | 5.32±0.94 | 1.04±0.19 | 5.12 | 2.35 | <0.001 | 2.45 | 95.90% |
| EMP2 | 3.38±0.56 | 0.97±0.15 | 3.48 | 1.8 | <0.001 | 1.82 | 98.90% |
| VPS72 | 0.26±0.04 | 0.95±0.17 | 0.27 | -1.89 | <0.001 | -1.86 | 98.40% |
| UBL5 | 0.17±0.03 | 0.96±0.18 | 0.18 | -2.47 | <0.001 | -2.45 | 99.20% |
| GDE1 | 0.34±0.05 | 0.98±0.20 | 0.35 | -1.51 | <0.001 | -1.57 | 96.20% |
| AQP7 | 0.19±0.04 | 1.02±0.21 | 0.19 | -2.39 | <0.001 | -2.40 | 99.60% |
| CUL1 | 0.48±0.07 | 0.99±0.16 | 0.48 | -1.06 | <0.001 | -1.05 | 99.00% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



