
Submitted:
09 May 2025
Posted:
13 May 2025
You are already at the latest version
Abstract
The aim of this research was to characterize the genetic structure of brown trout (Salmo trutta L.) complex populations in 10 river basins in the Aosta Valley (Italy) and neighbouring regions in northern Italy. The markers used were nuclear Lactate Dehydrogenase (LDH-C1*), mtDNA-CR (Control Region) and ddRAD-seq (SNPs). By this way, the degree of hybridization of wild populations with the Atlantic hatchery lineage was calculated. The results of the genetic analyses showed a composite situation in the studied area with different levels of introgression at the sampling sites. The mitochondrial lineages (Atlantic (AT), Mediterranean (ME), Adriatic (AD) and Marmoratus (MA) were present with varying percentages across study sites. For the Vertosan River, a distinctive native population and a level of hybridization close to zero were identified. Data allowed the identification in the Aosta Valley of four cluster units. In the future, within the study area, some interventions to support conservation actions can be envisaged.
Keywords:
ddRAD-seq
; mtDNA-CR
; LDH-C1*
; genomic analysis
; hybridization
; conservation
; Salmo trutta complex
1. Introduction
The brown trout (Salmo trutta L.) complex is one of most important salmonid species in Europe. This species shows a great phenotypic diversity related to its evolutionary history. In Italy, intensive restocking with brown trout of Atlantic origin, domesticated for decades, are carried out every year to increase the density of local populations, particularly for sport fishing. In recent years, numerous genetic analyses conducted in Italy on brown trout populations have shown a high level of introgression of the Atlantic lineage [1,2,3].
In brown trout, fragments of the mitochondrial non-coding Control Region (CR) [4], nuclear LDH-C1* gene and various microsatellite arrays have been commonly used to assess genetic diversity across populations [5,6,7,8,9,10]. According to [4] the CR mitochondrial marker, discriminates seven main clusters: Danubian (DA), Atlantic (AT), Marmoratus (MA), Adriatic (AD), Mediterranean (ME), Duero (DU) and Tigris (TI) lineages [4,11]. Of these, the ME, AD, AT and MA lineages are most relevant in the Alpine and Italian context. The ME lineage is typical of trout living in the Mediterranean coastal basins of Southwestern Europe. It is found in Spain, Southern France, Corsica, etc., and represents trout that likely survived glaciations in refugia around the Western Mediterranean. Notably, ME haplotypes have been documented in some North Italian waters connected to France. The AD lineage is associated with trout from the Adriatic Sea drainages (historically on the Italian and Balkan sides of the Adriatic). The AD lineage includes the genetic stock of Italy’s so-called Mediterranean trout (sometimes referred to as Salmo ghigii in recent literature) as well as the marble trout’s maternal lineage. In fact, the most common haplotype of this lineage (termed ADcs-1) has been found historically in trout from Lake Garda, Lake Maggiore, and the Adige River, suggesting a wide Adriatic lineage presence in pre-stocking times. The MA lineage refers to genetic variants characteristic of the marble trout (Salmo trutta marmoratus) and closely related forms. Marble trout, an endemic of Northern Italy and adjacent regions, has a distinctive appearance and historically dominated lower elevation rivers. Genetically, marble trout typically falls within or very close to the Adriatic lineage. For instance, the endemic lake trout of Lake Garda (Salmo carpio, or Garda “carpione”) carries unique haplotypes that belong to both AD and MA lineages, underlining that the marble trout lineage contributed to post-glacial trout diversity in the region. These seven lineages are considered evolutionarily significant units (ESUs), making it possible to implement ESU-based management programs.
In Italy, the original occurrence of the ME and AD lineages in the Alpine area has long been debated. Over the last 30 years, some authors have indicated brown trout ME and AD as the only native species in the Alpine area, while other authors have indicated S. trutta marmoratus as the only native species [12]. The original range of brown trout has been indicated as the Northern part of the Apennines and the Alpine area. Splendiani et al. [2] confirmed the autochthony of brown trout populations in the South-Western area of the Alps and indicated that the Cottian and Maritime Alps acted as a dispersal corridor from the Adriatic Sea to the French side.
A contact zone with marble trout has been established in that area. Polgar et al. [13] proposed several scenarios of distribution in the Po Valley before of the last glacial period. According to D’Agaro et al. [14], the entire Po Valley basin can be considered as a potential dispersal area for the ME and AD brown trout lineages. Splendiani et al. [1] conducted studies on DNA museum samples collected in Northern Italy prior to stocking activities with trout of the AT lineage. The results of the research showed the presence of ME and AD haplotypes (e.g., museum sample dated 1876 of ME lineage from Lake Moncenisio).
The aim of this study was to increase the knowledge on brown trout living in the North-West Alps, particularly in the Dora Baltea catchment and determine whether the dispersion of brown trout is limited to the Stura di Lanzo basin or whether it has also affected the Western-Northern area. The analysis of the mtDNA-CR genes (to identify the ME, AD, AT and MA lineages), nuclear LDH-C1* (to identify the 90 and 100 lalleles) and ddRAD-seq were used to improve the study of the genetic structure of brown trout populations. Thanks to ddRAD-seq, it is, now, possible to achieve very high marker densities (SNPs). Simply increasing the number of markers greatly increases the robustness and precision of estimates of various parameters traditionally used in conservation genetics. The amount of markers available also allows for the identification of outliers whose parameters deviate significantly from the rest of the genome. These outliers may reflect genomic regions involved in local adaptation. They are classically distinguished by high levels of genetic variation between populations living in different habitats [15,16].
2. Materials and Methods
2.1. Sample Collection
The objective of the present study was to sample a representative sample of the all the wild populations of the Aosta Valley (Italy) and nearby regions and assess their possible origin. In the period between October and December 2024, 10 locations in the main watercourses of the Aosta Valley region were sampled by electrofishing (Table 1).
At least twenty individuals per site were sampled across the 10 locations of Aosta Valley (Italy) (Figure 1).
Samples from the closed regions were added as reference groups: Piemonte (Chisone, Chiusola, Gesso, Loana, Ovarda, Stura di Lanzo, Ripa, Valcellina), Liguria (Mallero, Samolaco, Vara) and Emilia Romagna (Rio delle pozze), France (Petit Buëch) and Switzerland (Poschiavo). Figure 2 shows some brown trouts sampled in the study.
A small fin fragment was taken from the individuals sampled and preserved in ethyl alcohol at -20 °C. After removal of the fin fragment, each individual was immediately released at the catch site. Samples of n=192 individuals were selected for genetic analysis. Furthermore, samples collected (n=96) in a previous research [17] (Molise, Sardegna, Corsica (France), Lake Garda and Friuli Venezia Giulia) were used for the genetic analysis.
2.2. Genetic Analysis
2.2.1. mtDNA Control Region and Nuclear LDH-C1
The mtDNA gene (992 bp) was amplified with primers L19 and HN20 [4]. PCR conditions were as follows: 94°C for 5 min; 35 cycles at 94°C for 30 s; 50°C for 30 s and 72°C for 1 min; extension at 72°C for 10 min. Sequencing was performed with the ABI Prism 3130 instrument (Applied Biosystems, Carlsbad, CA, USA). The sequences obtained were aligned using Clustal W software (Mega 12). CR haplotypes were identified with DNASP 6 [18] software and assigned to the AD, MA, ME and AT lineages by phylogenetic analysis.
The LDH-C1* gene (440 bp) was digested with the enzyme BseLI (Fermentas Inc., Burlington, Canada) and the fragments obtained were separated using an agarose gel (*90/90, AT lineage and *100/100, ME lineage) [19].
2.2.2. Genomic Analysis
DNA extraction was performed with the MagAttract HMW DNA kit (Qiagen, Hilden, Germany). Sample sequencing was performed at IGA Technology Services S.r.l. (Udine, Italy). The ddRAD libraries were obtained according to the manufacturer’s instructions. Genomic DNA was digested with SbfI and SphI enzymes and an agarose gel was used to separate the DNA fragments. Paired-end sequences (2x150 bp) were obtained using the Novaseq6000 instrument (Illumina, San Diego, CA).
3. Results
3.1. Animal Genotyping
A total of 973,900,734 sequences were obtained. 19,222,079 was the maximum number of demultiplexed sequences per sample and the average number was 10,144,799. In total, 263,198 SNPs passed the filtering criteria and were identified on the 40 chromosomes of Salmo trutta. All samples achieved excellent qualitative values. After merging with the samples analysed by Magris et al. (2022), the new group of individuals consisted of 288 individuals with 93,678 SNPs.
3.2. Population Structure
Table 2 shows a summary of the results obtained in the Aosta Valley for the mtDNA-CR, LDH-C1* and ddRAD-seq (q quantifies the hybridisation to the AT lineage) analysis. Sequencing of the mtDNA-CR gene resulted in a 992 bp sequence. Eight different haplotypes were identified (MEcs1; ADrh1; ADporh1; ADcs1; MA-S1; MA-S2; MA-S3; MA-S4; Masl1. LDH-C1* analysis confirmed the presence of the 90 allele in the AT lineages.
To explore the brown trout genetic population structure and hybridisation level in the Aosta Valley and closed regions, a genetic analysis was performed using the Admixture software. Analysis was performed on two different datasets. The first dataset includes all the samples of Aosta Valley and nearby regions from the Alpine area (Piemonte, Liguria, France and Switzerland) and Sardinia (Figure 3 and Figure 4). Based on these values, the number of clusters that best support the populations studied in the Aosta Valley and closed regions is 5 (K=5) (AT, ME+AD, MA, Native (Vertosan) and Native (Sardinia)). The number of clusters (K) was chosen based on the Cross Validation Error (CVE), selecting the value of K matching the lowest CVE (K=1 -7). Comparing samples from the Aosta Valley, those from Vargno, Stuba, Graines, Giassit, Ayasse and Crest populations showed intermediate levels of hybridisation with the AT lineage while the Vertosan population showed a distinctive native cluster and a level of hybridisation close to zero. These results confirm the findings of other research conducted over the past thirty years [14]. The restocking of the main rivers in the Alpine regions with populations of AT lineage resulted in the genetic erosion of the native populations. The Vertosan River in the Aosta Valley was the only site with a zero introgression level (Q=0.04) and the Morgex site showed the highest value (Q=0.96). Genetic traces of the MA lineage were found in almost all samples from the Aosta Valley, with varying percentages. Specimens from Vargno and Stuba rivers showed more than 30% of MA lineage. Hybridization with the AT lineage confirms was confirmed also in the Piemonte, Liguria and Apennines area as already reported by Magris et al. [17]. The hybridisation of marbled populations with the AT lineage indicates that the restocking activity is taking place also in typical marbled areas.
In Figure 4, results of the genetic analysis were grouped according to the different Italian regions.
The second set of data included samples from Magris et al. [17] as the Garda Lake (Salmo carpio), Apennines area, Sardegna and the reference AT and MA lineages. In Figure 5 is shown a Neighboor-Net network graph of the brown trout complex populations living across the Alpine area.
The graph shows that populations from Vertosan, Ripa and Petit Buech Buëch (France) rivers are located at the opposite end of the the marbled trout from the Central and Eastern Alps. Specimens of Chisone, Crest and Vargno rivers are arranged along intermediate nodes between these two extremes. The intermediate position in the phylogenetic network of the S. Carpio from the Garda Lake, support the hypothesis of the hybrid origin of this differentiated and isolated form. In the graph, for each population, were reported by means of pie charts, the mitochondrial haplotypes (CR). Populations in intermediate position in the network, show a variable percentage of MA haplotypes.
Finally, in Figure 6 the phylogenetic trees is elaborated into a broader framework that also includes the Apennines area and Sardinia lineage group.
4. Discussion
MtDNA-CR has been used for many years to identify the lineage of trout populations [23]. Early studies by Giuffra et al. [11], first revealed that brown trout in the Po River basin of Northern Italy did not all belong to the Atlantic lineage as stocking records might suggest, but instead many carried ME lineage haplotypes. Giuffra identified distinct mtDNA haplotypes in North Italian trout corresponding to the AD and ME lineages. Mitochondrial haplotyping remains a cornerstone in detecting remnant native trout. Nuclear markers like the LDH-C1 locus* may provide an early genetic differentiation between Atlantic and Mediterranean trout. Splendiani et al. [2] recently combined mtDNA data with LDH-C1* genotypes to confirm that trout in the South-Western Alps carry the Mediterranean-specific allele. Allozyme studies in the 1990s [8] similarly noted distinct allele patterns in Alpine trout, reflecting limited gene flow between native and introduced stocks. New Next-Generation Sequencing (NGS) techniques such as ddRAD-seq allow to obtain thousands of markers to be analysed simultaneously. This high number of markers is a very effective and precise method for calculating the level of introgression at the individual level. Numerous studies have used this technique in conservation and management studies of different species, especially non-model species. By this way, it is possible to determine the level of introgression between different ecotypes or lineages [24]. Splendiani et al. [2] carried out a comprehensive genetic survey in the Southwest Alps (spanning Italian and French sides). By examining LDH-C1* and mtDNA, they confirmed that the ME lineage trout in the Italian South-Western Alps are native, retaining haplotypes distinct from e Atlantic stocks.
Results of the present study showed a considerable degree of introgression by the Atlantic lineage of domestic origin in almost all sampled streams, excluding the Vertosan river. However, the genetic structure of brown trout populations in the Aosta Valley and neighbouring regions is consistent with what has been found in other Western Alpine basins [2]. In the light of the possible dispersal dynamics of brown trout and recent findings acquired by several research groups in recent years [1,2,3], it is believed that there are still some nuclei of native trout in the Aosta Valley and Sardinia that require active safeguard efforts and future management practices.
In the light of recent studies [2,14,25] which confirm the autochthony of brown trout in the Alpine area (from Liguria to Lake Garda), it is reasonable to assume that originally the Po Valley, was set up of a dynamic and varied group of populations with different genetic and ecological characteristics, which, today, has largely disappeared. Throughout the 1990s and 2000s, researchers tried to determine if trout, bearing Mediterranean haplotypes in regions like the Po basin and Alpine lakes, were relics of post-glacial colonization or if they had been imported from elsewhere (such as France or Italy’s Apennine streams). The South-Western Alps (Cottian and Maritime Alps on the Italy-France border) emerge as a critical refugial zone for the Mediterranean trout. In this region, including rivers like the upper Dora Riparia, Chisone, Stura di Lanzo, upper Po, and others, researchers have discovered viable native populations of Mediterranean brown trout persisting alongside the marble trout [6]. These trout belong to the Italian Adriatic/Mediterranean lineages (historically termed Salmo ghigii by some authors) and show a distinctive altitudinal segregation from marble trout. Genetic surveys found that ME and AD haplotypes occur predominantly in higher-altitude streams (~1000–2000 m elevation) of the southwest Alps, whereas MA (marble) haplotypes/fish dominate lower elevations (0–1000 m). In other words, the native brown trout (Mediterranean lineages) in these headwaters appear to be a separate gene pool, largely confined to upland habitats where marble trout competition is lower.
On a broad geographical scale, several hypotheses have been proposed for the taxonomic classification and evolution over time of populations of Salmo trutta complex from the Southern European range. In Northern Italy, postglacial recolonization may have occurred from different refugia with different lineages. Based on the results obtained, there is no reason to rule out the possibility that, following the Last Glacial Maximum (LGM), brown trout populations recolonized suitable alpine streams in the North-Westem Alpine region, including the Dora Baltea basin. In many European countries, at a local level, a high genetic differentiation between populations has been highlighted. These differences are also due to the homing instinct of the brown trout and the possible presence of artificial barriers, which reduces the gene flow. It should be noted that little is known about the relationships between sedentary and anadromous populations. While admitting, therefore, that in such a dynamic context, at the contact areas between the Alpine are and the Apennines, river catches could have promoted the exchange of genetic material in both directions.
The MEcs1 mitochondrial haplotype was found in the Vertosan (Aosta Valley) and Chisone and Pipa (Piemonte) populations. A possible hypothesis is that the MEcs1 haplotype reached the Liguria basins from Piemonte. However, the hypothesis that the MEcs1 haplotype is originated from a marine colonization of Liguria appears equally valid and probable, given its distribution in Vara,Serchio and in Corsica rivers [1,2]. The area of the Western Alps is characterized by a considerable asymmetry, with the Po Valley slope being significantly steeper than the transalpine slope. It is not surprising, therefore, that the MEcsl haplotype may have reached the Po basins through a river capture phenomena, not uncommon during the Quaternary period. Furthermore, we cannot exclude the possibility that the ME haplotype m ay also arrived from the Liguria basins, given the presence of the MEcs1 haplotype in the populations of Baracca river, a tributary of Orba (Liguria-Piemonte Apennines, Tanaro Basin) and Sansobbia (Liguria-Piemonte Apennines, Liguria Sea Basin) [2]. A museum specimen from the Lerca, a tributary of the Leirone (Cogoleto, Genoa, Liguria Sea Basin) also shows a haplotype that can be ascribed to MEcs-1. According to Splendiani et al. [2], a corridor existed in remote times that would have transferred the ADporh1 haplotype (typically associated with the Cuneo Po Valley populations) to the French catchment areas. It is presumable, therefore, that the Western Alps have acted as a bidirectional corridor of genetic flow between the Mediterranean species of the Po basin and those of the Rhone basins. Splendiani et al. [1] reconstructed the post-glacial colonization history: during the last glaciation and the following deglaciation, this region served as a one-way corridor for trout dispersal from East to West. During the Last Glacial Maximum (LGM), much of the higher Alps was ice-covered and uninhabitable for fish. Native trout survived in refugia at the periphery, for example, in rivers South of the Alps (Padania plain), in the Liguria coastal streams, and in some large lake basins that remained ice-free (like parts of Lake Maggiore and Lake Garda). This means that as glaciers receded, trout that had survived in refugia on the Adriatic/Padanian side (perhaps in the Po basin or Northern Apennines) expanded westward through the alpine foothills into what is now the French Alpine region. This scenario directly refutes the older “French origin” hypothesis. For Polgar et al. [13], the main factors that would have limited the dispersion of Mediterranean trout after the last glacial m aximum (LGM) would be the total coverage of the main valleys by glaciers and the distance from the glacial refuges.
According to Antognazza et al. [3], plans aimed at preserving local genetic lines are urgently needed in order to avoid the complete impoverishment of the original biodiversity of the Alpine area. It is, therefore, advisable that active management of fish stocks should be directed towards the preservation of native local forms. It is also considered important that the marble trout should enjoy maximum protection, by means of appropriate fishing regulations and by increasing efforts to protect its habitats of choice. Worthy of mention are, for example, the marble trout conservation projects, one of which is underway in the Dora Baltea basin in the Aosta Valley. Logically, careful management of fish stocks must also pass through an assessment of the suitability of each river. New introductions, even of native species, should be accompanied by an assessment of the impacts on other native species. Furthermore, the implementation of ’buffer zones’ limiting the possibility of introgression between species, forced by human activities, is reasonable [14].
5. Conclusions
This section is not mandatory but can be added to the manuscript if the discussion is unusually long or complex. The combined use of the markers mtDNA-CR (allowing characterisation of the AT, AD, ME and MA lineages), LDH-C1* (allowing identification of the 90 and 100 alleles) and ddRAD-seq analysis allowed to obtainan an efficient and reliable analysis of the genetic structure of brown trout populations . In particular, the ddRAD technique made it possible to obtain a very high density of markers (SNPa) with an increase in the robustness and precision of estimates of various parameters traditionally used in conservation genetics (e.g., hydridation level). The quantity of markers available also makes it possible to identify outliers whose parameters deviate significantly from the rest of the genome.
The results of the present study highlight a composite genetic structure of the brown trout populations in the Aosta Valley, with the presence of different mitochondrial lineages (Mediterranean (ME), Adriatic (AD) Atlantic (AT) and Marmoratus (MA)) and different introgression rates among the sampling sites. For the Vertosan River, a distinctive new native population was identified with a hybridisation level close to zero.
Author Contributions
All authors have contributed equally. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the START-UP 2018 grant from the University of Udine (project GENSAL) to Michele Morgante.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Splendiani, A.; Fioravanti, T.; Giovannotti, M.; Olivieri, L.; Ruggeri, P.; Cerioni, P.N.; Vanni, S.; Enrichetti, F.; Barucchi, V.C. Museum samples could help to reconstruct the original distribution of Salmo trutta complex in Italy. J. Fish Biol. 2017, 90, 2443–2451. [Google Scholar] [CrossRef] [PubMed]
- Splendiani, A.; Berrebi, P.; Tougard, C.; Righi, T.; Reynaud, N.; Fioravanti, T.; Lo Conte, P.; Delmastro, G.B.; Baltieri, M.; Ciuffardi, L.; et al. The role of the south-western Alps as a unidirectional corridor for Mediterranean brown trout (Salmo trutta complex) lineages. Biol. J. Linn. Soc. 2020, 131, 909–926. [Google Scholar] [CrossRef]
- Antognazza, C. M.; Palandaćić, A.; Delmastro, G. B.; Crosa, G.; Zaccara, S. Current and Historical Genetic Variability of Native Brown Trout Populations in a Southern Alpine Ecosystem: Implications for Future Management. Fishes 2023, 8, 411. [Google Scholar] [CrossRef]
- Bernatchez, L. The evolutionary history of brown trout (Salmo trutta L.) inferred from phylogeographic, nested clade, and mismatch analyses of mitochondrial DNA variation. Evolution 2001, 55, 351–379. [Google Scholar]
- Berrebi, P. Three brown trout Salmo trutta lineages in Corsica described through allozyme variation. J. Fish Biol. 2015, 86, 60–73. [Google Scholar] [CrossRef]
- Berrebi, P.; Barucchi, V.C.; Splendiani, A.; Muracciole, S.; Sabatini, A.; Palmas, F.; Tougard, C.; Arculeo, M.; Maric, S. Brown trout (Salmo trutta L.) high genetic diversity around the Tyrrhenian Sea as revealed by nuclear and mitochondrial markers. Hydrobiologia 2019, 826, 209–231. [Google Scholar] [CrossRef]
- Berrebi, P.; Horvath, Á.; Splendiani, A.; Palm, S.; Berna’s, R. Genetic diversity of domestic brown trout stocks in Europe. Aquaculture 2021, 544, 737043. [Google Scholar] [CrossRef]
- Giuffra, E.; Bernatchez, L.; Guyomard, R. Mitochondrial control region and protein coding genes sequence variation among phenotypic forms of brown trout Salmo trutta from northern Italy. Mol. Ecol. 1994, 3, 161–171. [Google Scholar] [CrossRef]
- Gratton, P.; Allegrucci, G.; Sbordoni, V.; Gandolfi, A. The evolutionary jigsaw puzzle of the surviving trout (Salmo trutta L. complex) diversity in the Italian region. A multilocus Bayesian approach. Mol. Phylogenetics Evol. 2014, 79, 292–304.
- Snoj, A.; Mari’c, S.; Bajec, S.S.; Berrebi, P.; Janjani, S.; Schöffmann, J. Phylogeographic structure and demographic patterns of brown trout in North-West Africa. Mol. Phylogenetics Evol. 2011, 61, 203–211. [Google Scholar] [CrossRef]
- Giuffra, E.; Bernatchez, L.; Guyomard, R. Mitochondrial control region and protein coding genes sequence variation among phenotypic forms of brown trout Salmo trutta from northern Italy. Mol. Ecol. 1994, 3, 161–171. [Google Scholar] [CrossRef]
- Meraner, A.; Gandolfi, A. Genetics of the genus Salmo in Italy: Evolutionary history, population structure, molecular ecology and conservation. Brown trout: Biol. Ecol. Manag. 2018, 65–102. [Google Scholar]
- Polgar, G.; Iaia, M.; Righi, T.; Volta, P. The Italian Alpine and Subalpine trouts: taxonomy, evolution, and conservation. Biology 2022, 11, 576. [Google Scholar] [CrossRef] [PubMed]
- D’Agaro, E.; Gibertoni, P.; Marroni, F.; Messina, M.; Tibaldi, E.; Esposito, S. Genetic and phenotypic characteristics of the Salmo trutta complex in Italy. Appl. Sci. 2022, 12, 3219. [Google Scholar] [CrossRef]
- Lecaudey, L.A.; Schliewen, U.K.; Osinov, A.G.; Taylor, E.B.; Bernatchez, L.; Weiss, S.J. Inferring phylogenetic structure, hybridization and divergence times within Salmoninae (Teleostei: Salmonidae) using RAD-sequencing. Mol. Phylogenet. Evol. 2018, 124, 82–99. [Google Scholar] [CrossRef]
- Lemopoulos, A.; Prokkola, J.M.; Uusi-Heikkilä, S.; Vasemägi, A.; Huusko, A.; Hyvärinen, P.; Koljonen, M.; Koskiniemi, J.; Vainikka, A. Comparing RADseq and microsatellites for estimating genetic diversity and relatedness—Implications for brown trout conservation. Ecol. Evol. 2019, 9, 2106–2120. [Google Scholar] [CrossRef] [PubMed]
- Magris, G.; Marroni, F.; D’Agaro, E.; Vischi, M.; Chiabà, C.; Scaglione, D.; Kijas, J.; Messina, M.; Tibaldi, E.; Morgante, M. ddRAD-seq reveals the genetic structure and detects signals of selection in Italian brown trout. Genet. Sel. Evol. 2022, 54, 8. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- McMeel, O.M.; Hoey, E.M.; Ferguson, A. Partial nucleotide sequences, and routine typing by polymerase chain reactionrestriction fragment length polymorphism, of the brown trout (Salmo trutta) lactate dehydrogenase, LDH-C1*90 and *100 alleles. Mol. Ecol. 2001, 10, 29–34. [Google Scholar] [CrossRef]
- Rochette, N. C.; Rivera-Colon, A.G.; Catchen, J.M. Stacks 2: Analytical methods for paired-end sequencing improve RADseq-based population genomics. Mol. Ecol. 2019, 28, 4737–4754. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv, 2013; arXiv:arXiv:1303.3997v2. [Google Scholar]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef]
- Segherloo, I.H.; Freyhof, J.; Berrebi, P.; Ferchaud, A.L.; Geiger, M.; Laroche, J.; Levin, B.A.; Normandeau, E.; Bernatchez, L. A genomic perspective on an old question: Salmo trouts or Salmo trutta (Teleostei: Salmonidae)? Mol. Phylogenetics Evol. 2021, 162, 107204. [Google Scholar] [CrossRef] [PubMed]
- Keller, I.; Schuler, J.; Bezault, E.; Seehausen, O. Parallel divergent adaptation along replicated altitudinal gradients in Alpine trout. BMC Evol. Biol. 2012, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Segherloo, I.H. , Freyhof, J.; Berrebi, P.; Ferchaud, A.L.; Geiger, M.; Laroche, J.; Bernatchez, L. A genomic perspective on an old question: Salmo trouts or Salmo trutta (Teleostei: Salmonidae)? Mol. Phylogenetics Evol. 2021, 162, 107204. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map of the Aosta Valley and nearby regions where the fish of the study were sampled.
Figure 2.
Morphological features of brown trout sampled in Vertosan (Aosta Valley), Ripa and Chisone (Piemonte).
Figure 2.
Morphological features of brown trout sampled in Vertosan (Aosta Valley), Ripa and Chisone (Piemonte).

Figure 3.
Q values (% of ancestry) obtained with the Admixture software using the smallest cross validation error. The coloured bars (height) indicate the probability of that individual being assigned to a particular lineage: AT lineage (lilac), ME+AD lineage (red), Marmoratus (green) and native wild (blue and grey).
Figure 3.
Q values (% of ancestry) obtained with the Admixture software using the smallest cross validation error. The coloured bars (height) indicate the probability of that individual being assigned to a particular lineage: AT lineage (lilac), ME+AD lineage (red), Marmoratus (green) and native wild (blue and grey).
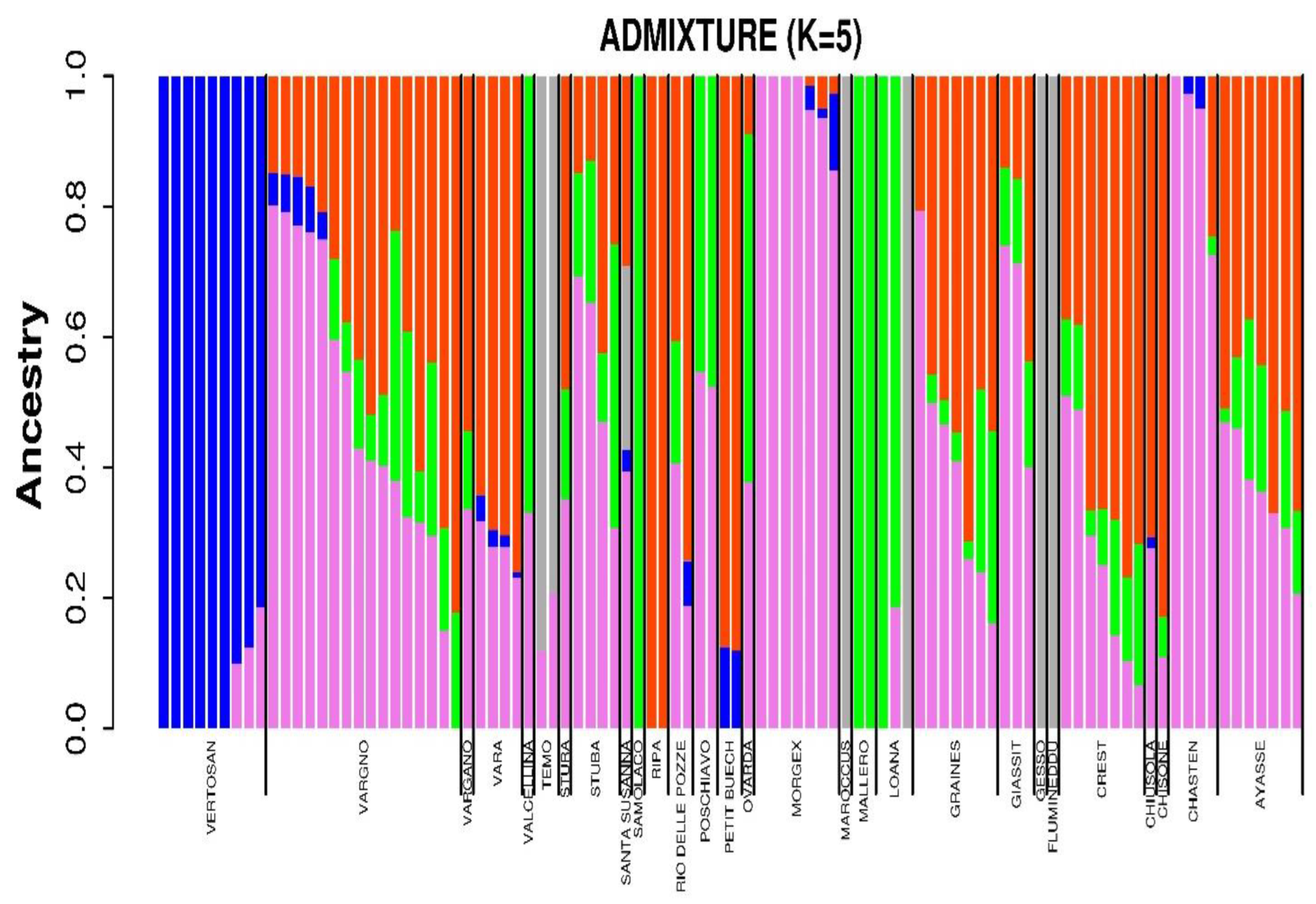
Figure 4.
Q values (% of ancestry) obtained with the Admixture software using the smallest cross validation error. The coloured bars (height) indicate the probability of that individual being assigned to a particular lineage. lineage: AT lineage (lilac), ME+AD lineage (red), Marmoratus (green) and native wild (blue and grey).
Figure 4.
Q values (% of ancestry) obtained with the Admixture software using the smallest cross validation error. The coloured bars (height) indicate the probability of that individual being assigned to a particular lineage. lineage: AT lineage (lilac), ME+AD lineage (red), Marmoratus (green) and native wild (blue and grey).
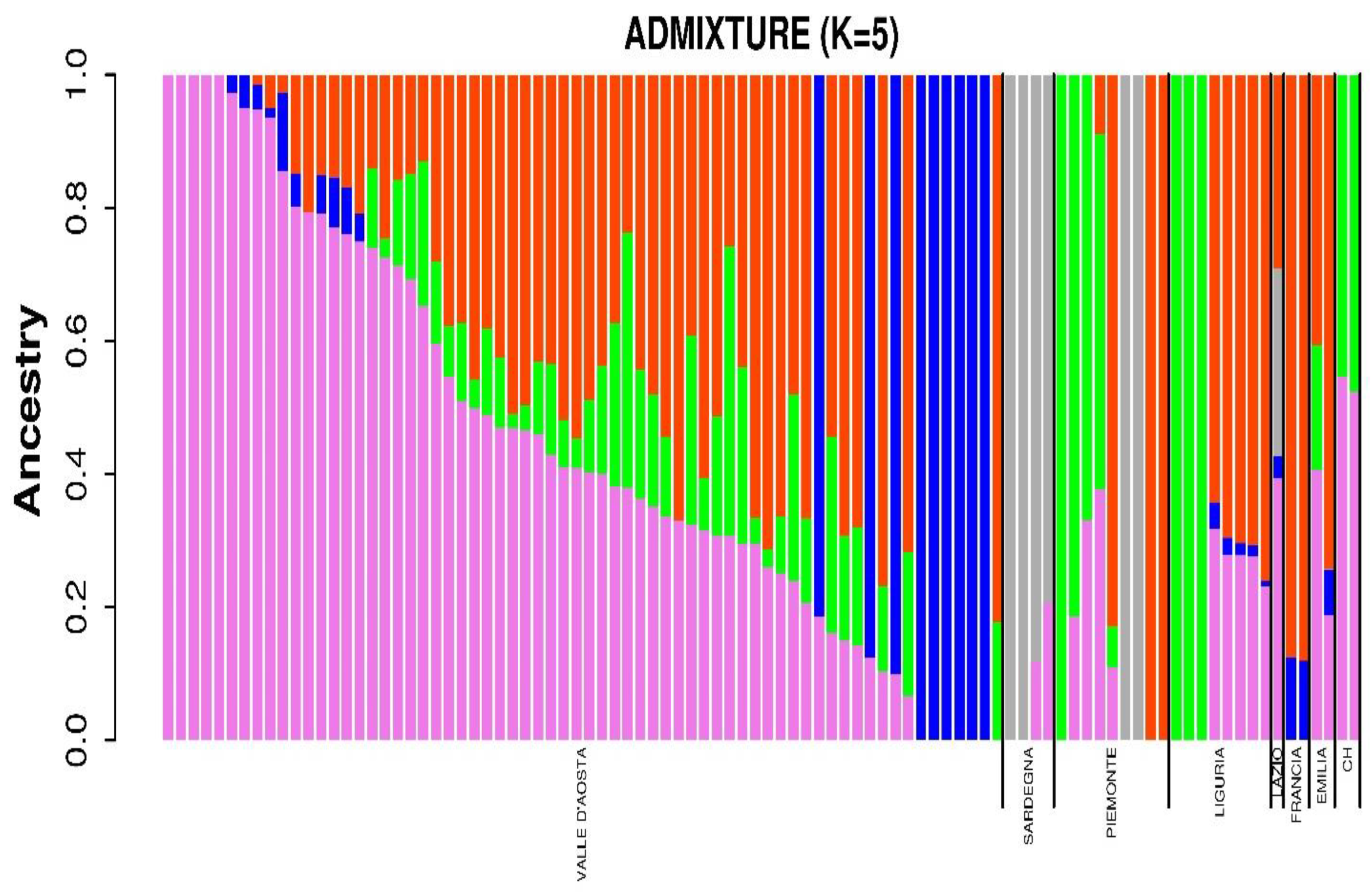
Figure 5.
Neighboor-Net network of the brown trout complex populations living across the Alpine area. Mitochondrial haplotype (CR) membership (ME, AD, MA lineages) and relative frequency in each population are shown (pie charts).
Figure 5.
Neighboor-Net network of the brown trout complex populations living across the Alpine area. Mitochondrial haplotype (CR) membership (ME, AD, MA lineages) and relative frequency in each population are shown (pie charts).
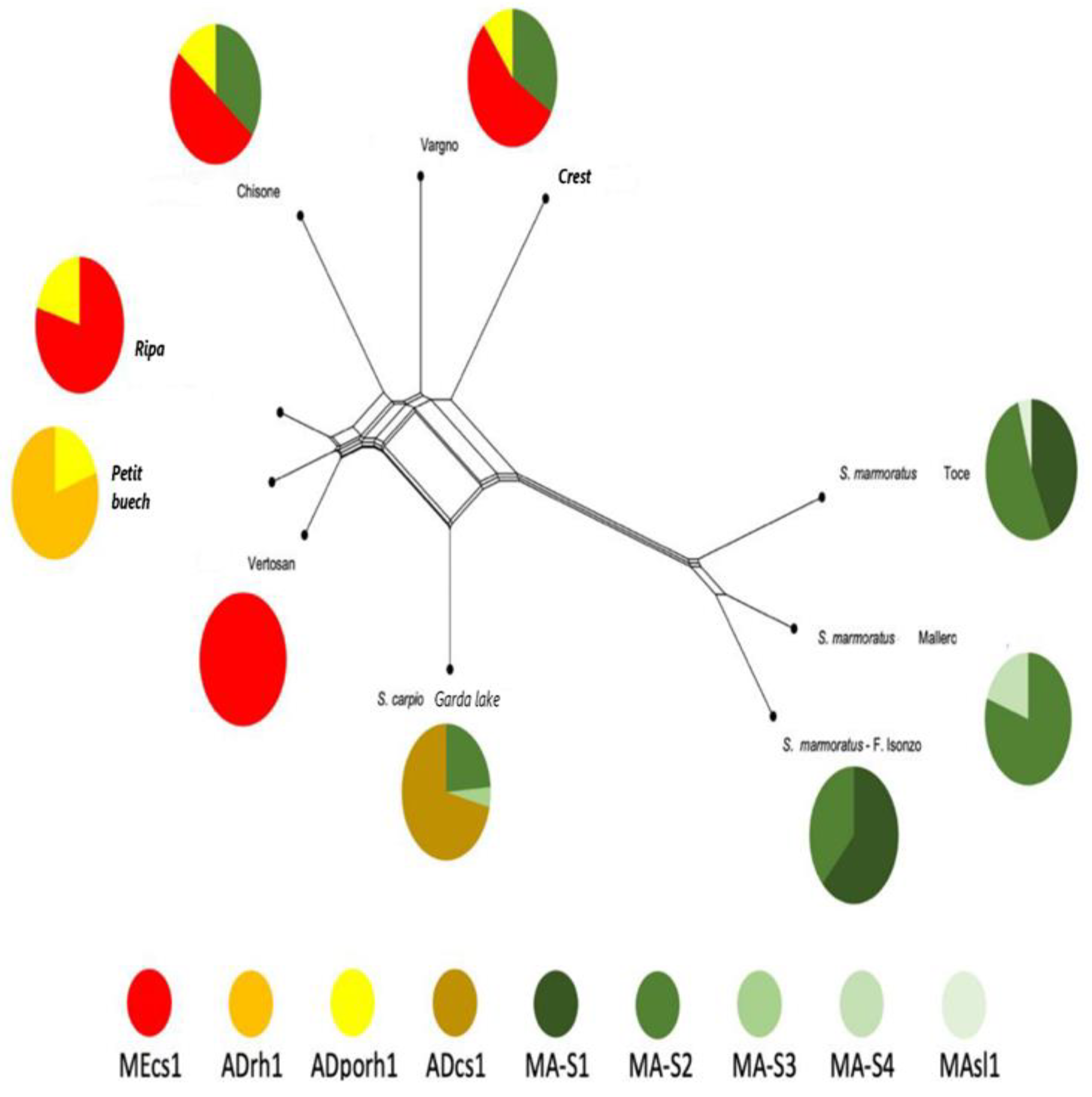
Figure 6.
Neghboor-net Network brown trout complex populations living across the Alpine, Apennines and Sardinia areas.
Figure 6.
Neghboor-net Network brown trout complex populations living across the Alpine, Apennines and Sardinia areas.
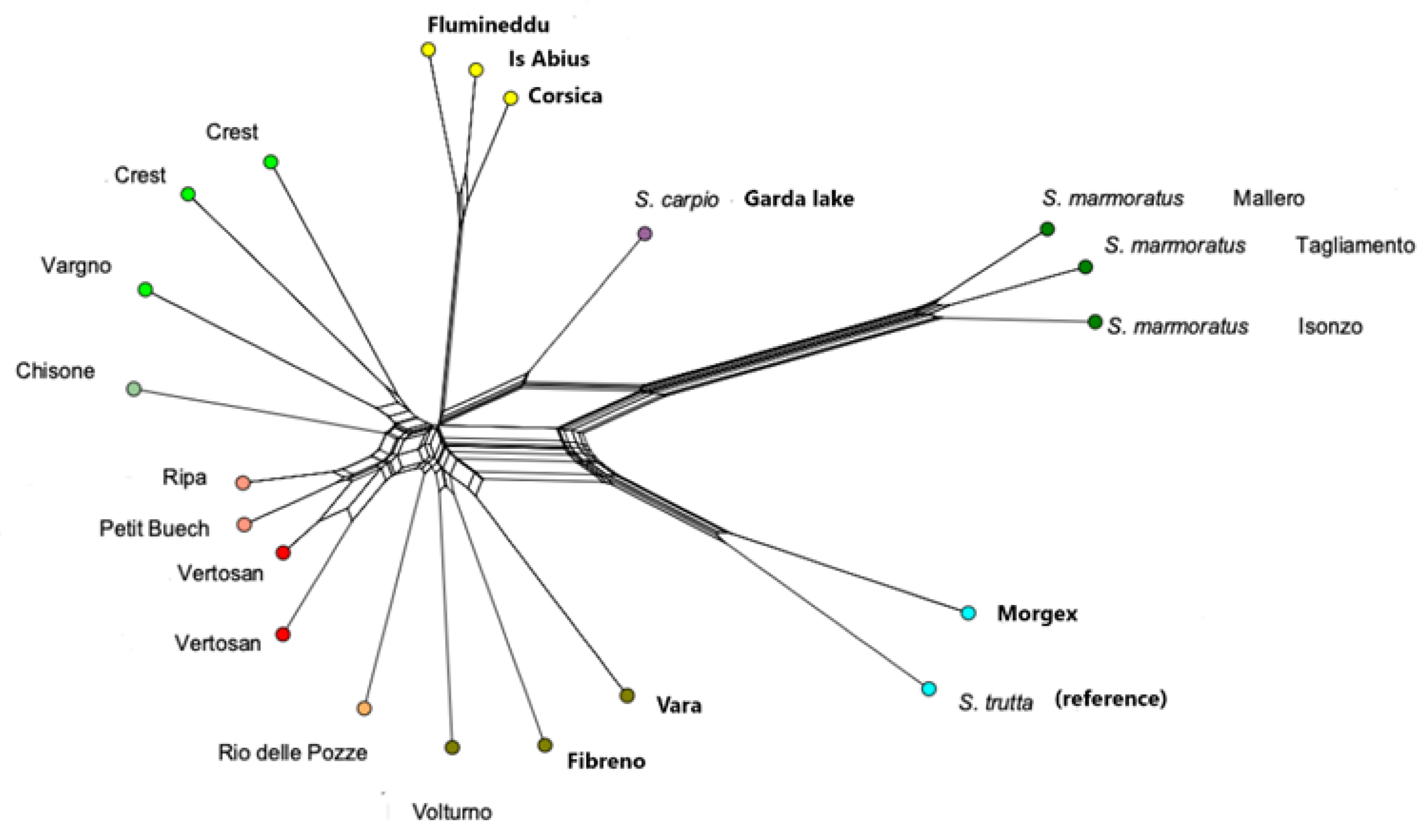

Table 1.
List of sampling sites of brown trout and geographic features.
| River catchment | Number of samples | Lon °(N) | Lat °(E) |
| Ayasse | 20 | 39°93′05.25” | 74°11′1.94 |
| Chasten | “ | 40°23′28.53” | 74°73′3.40 |
| Chisone | “ | 44°49′13.2” | 72°45′1.76 |
| Chiusola | “ | 44°17′37.44” | 94°03′1.54 |
| Crest | “ | 39°72′07.72” | 75°43′5.79 |
| Flumineddu | “ | 39°32′43.39” | 92°25′3.04 |
| Gesso | “ | 44°24′26.78” | 73°32′1.66 |
| Giassit | “ | 41°16′92.99” | 74°72′7.17 |
| Isonzo | “ | 46°24′42.56” | 134°32′6.79 |
| Loana | “ | |44°07′44.41” | 81°53′5.36 |
| Mallero | “ | 46°20′29.85” | 94°35′4.69 |
| Maroccu | “ | 39°13′04.65” | 84°91′5.45 |
| Morgex | “ | 34°85′80.76” | 70°24′5.45 |
| Ovarda | “ | 44°52′19.39” | 83°06′5.68 |
| Petit Buëch | “ | 44°11′51.28” | 55°63′5.16 |
| Poschiano | “ | 46°25′00.79” | 100°30′8.95 |
| Rio delle pozze | “ | 46°16′35.06” | 113°65′5.50 |
| Ripa | “ | 44°57′20.05” | 64°73′2.53 |
| Samolacco | “ | 46°16′46.16” | 92°34′6.23 |
| Santa susanna | “ | 42°30′09.82” | 125°19′6.56 |
| Stuba | “ | 40°47′70.10” | 77°34′3.39 |
| Stura | “ | 45°08′55.26” | 82°05′2.47 |
| Temo | “ | 40°17′26.57” | 82°83′3.47 |
| Toce | “ | 45°56′08.07” | 82°93′0.04 |
| Valcellina | “ | 46°10′00.30” | 124°00′0.21 |
| Vara | “ | 44°09′09.30” | 95°31′4.97 |
| Vargno 1 | “ | 41°08′94.27” | 75°44′0.97 |
| Vargno 2 | “ | 41°23′91.34” | 75°23′0.21 |
| Vertosan | “ | 35°42′22.44” | 71°54′0.29 |
| Volturno | “ | 41°38′14.60” | 140°41′6.81 |
Table 2.
Results of MtDNA CR, DDH-C1* and ddRAD analysis.
| River catchment | Mt DNA-CRaplotypes | LDH-C1*alleles | ddRAD-seq Q (Ancestry to AT) |
| Morgex | AT | 90/90 | 0.96 |
| Ayasse | AD AT MA | 90/100 | 0.36 |
| Chasten | AT | 90/90 | 0.91 |
| Crest | AD AT MA | 90/100 | 0.25 |
| Giassit | AT AD | 90/100 | 0.61 |
| Graines | ME AT MA | 90/100 | 0.40 |
| Stuba | AT AD MA | 90/100 | 0.53 |
| Vargno | ME AT MA | 90/100 | 0.46 |
| Vertosan | ME | 100/100 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



