
Submitted:
08 May 2025
Posted:
09 May 2025
You are already at the latest version
Abstract
Drought stress is a critical constraint limiting seed germination and seedling establish-ment in field crops such as jalapeño pepper (Capsicum annuum L.). Nanopriming, a seed enhancement technique using nanoparticle suspensions, has emerged as a sustainable approach to improve water-use efficiency during early development. This study evaluated the effects of zinc oxide (ZnO, 100 mg·L⁻¹), silicon dioxide (SiO₂, 10 mg·L⁻¹), and their com-bination (ZnO + SiO₂), stabilized with chitosan, on the germination performance and stress tolerance of four commercial jalapeño hybrids under mannitol-induced drought conditions (0%, 15%, and 30%). Seeds were primed for 12 hours, and physiological, mor-phological, and biochemical responses were measured. Nanopriming significantly im-proved germination percentage, speed, and seedling vigor under both moderate (15%) and severe (30%) osmotic stress. SiO₂ enhanced chlorophyll content and germination rate un-der 15% stress, while ZnO promoted proline and flavonoid accumulation under 30%. The combined ZnO + SiO₂ treatment delivered the most consistent improvements across all traits and stress levels. Multivariate analysis revealed clear treatment-specific and drought-dependent response patterns, emphasizing the role of osmolytes and antioxidant activity. These results highlight the potential of nanopriming as a low-cost, scalable strat-egy to enhance early-stage drought resilience and promote efficient water use in jalapeño cultivation under semi-arid conditions.
Keywords:
Drought-stress
; germination
; jalapeño pepper
; nanopriming
; seedling
; SiO2
; ZnO
1. Introduction
Drought is one of the most limiting abiotic factors affecting seed germination and early seedling establishment, significantly reducing agricultural productivity in arid and semi-arid regions [1-4]. Under water-deficit conditions, plants experience several physiological disruptions, including decreased water potential [5], excessive accumulation of reactive oxygen species (ROS) [6], and reduced germination rate, seedling vigor, and growth [7-8]. Water is essential for plant metabolism and structure, comprising approximately 85% of the fresh biomass of plants [9].
In experimental settings, mannitol is frequently used to simulate drought by lowering the osmotic potential of the germination medium, allowing the assessment of plant responses to water stress under controlled conditions [10]. In crops such as jalapeño pepper (Capsicum annuum L.), water stress at early stages reduces emergence rate, vigor, and photosynthetic capacity [11]. Jalapeño belongs to the Solanaceae family and is a nutritionally rich crop with significant antioxidant compounds including capsaicinoids, flavonoids, and vitamins A, C, and E [12-13]. Mexico is the second-largest producer worldwide, concentrating over 50% of its national production in the northern states—semi-arid zones where water scarcity is a critical constraint [14-15]).
Improving seedling establishment under drought conditions is crucial to increasing water productivity. Among emerging seed enhancement technologies, seed nanopriming has shown promise. This technique involves soaking seeds in aqueous suspensions of mineral nanoparticles (NPs) at specific concentrations and durations, improving hydration kinetics, antioxidant activity, and seedling vigor under stress conditions [16-17]). Studies with ZnO and SiO₂ nanopriming have reported enhanced germination and early growth in cereals such as wheat and maize under water stress [2, 18-19].
Physiological and biochemical indicators such as proline, phenolic compounds, flavonoids, and soluble sugars are commonly used to evaluate stress responses. These metabolites are involved in osmotic adjustment, membrane stabilization, and ROS scavenging, playing central roles in plant adaptation to drought [20-22].
However, there is limited research on the efficacy of ZnO and SiO₂ nanopriming in commercial jalapeño pepper varieties, especially under mannitol-induced water stress. Therefore, this study aimed to evaluate the effects of seed nanopriming with ZnO and SiO₂ on germination, early growth traits, and oxidative metabolism in Capsicum annuum L. under both optimal and drought-simulated conditions. The outcomes of this study may pave the way for the implementation of nanopriming as a viable pre-sowing strategy to improve seed performance under water-deficit conditions, contributing to more sustainable and climate-resilient jalapeño pepper cultivation in semi-arid agroecosystem.
2. Results
2.1. Germination Parameters
Seed germination was significantly influenced by nanopriming treatments, drought stress levels, and jalapeño pepper varieties (p ≤ 0.05), as shown in Table 1. Treatments were coded using a combination of the variety abbreviation, treatment number, and stress level (see Table 2). For example, MT215 corresponds to the Mixteco variety treated with ZnO (Treatment 2) under 15% drought stress.
Under optimal conditions (0%), treatments such as IMT10, FT10, IMT20, FT20, IMT30, and IMT40 achieved 100% germination, while IT10 (Ideal + hydropriming) showed the lowest germination (75%). Nanopriming with SiO₂ (IT30) resulted in the shortest average germination time (2.33 ± 0.08 days) and the highest germination rate (0.429 ± 0.015 day⁻¹), outperforming FT30 (4.13 ± 0.14 days and 0.242 ± 0.009 day⁻¹, respectively).
2.2. Morphological Parameters
Nanopriming significantly improved seedling morphology, especially under water stress (Figure 1 and Figure 2). With 0% drought, FT10 recorded the longest shoot length (27 ± 0.91 mm) and the highest fresh weight (68.67 ± 4.16 mg), while FT20 and MT30 showed the longest roots. MT30 also had the largest stem diameter (0.3 ± 0.1 mm), three times larger than IMT30.
With 15% stress, FT415 (Forajido + ZnO + SiO₂) produced the highest fresh weight of the seedlings, while IT415 had the longest shoots. FT315 showed the most extensive root growth (87.5 ± 2.88 mm). The highest shoot-to-root ratio was observed in IMT215 (0.54), while FT115 had the lowest (0.13), indicating a variation in resource allocation among the treatments.
Under 30% water stress, IT130 had the longest shoots (16.3 ± 3.1 mm), and MT330 and MT130 had the longest roots. IMT330 and MT130 also had the highest fresh weights (>34 mg). IT230 showed the highest shoot-to-root ratio (1.56), suggesting preferential shoot development under extreme drought conditions.
2.3. Biochemical Parameters
2.3.1. Photosynthetic Pigments
Pigment levels (chlorophyll a, chlorophyll b, total chlorophyll, and carotenoids) were analyzed using a two-way ANOVA (water stress level x priming treatment) performed separately for each jalapeño variety, followed by Tukey's HSD test (p ≤ 0.05).
Chlorophyll and carotenoid content varied significantly between treatments and stress levels (Figure 3A–D). In the absence of water stress (0%), the Mixteco variety with the ZnO + SiO₂ treatment (MT40) had the highest values of total chlorophyll (787.26 ± 31.79 µg·g⁻¹ PF) and carotenoids (103.49 ± 8.25 µg·g⁻¹ PF). Under moderate stress (15%), IT415 (Ideal + ZnO + SiO₂) achieved the highest total chlorophyll content (831.64 ± 173.63 µg·g⁻¹ PF). Under severe stress conditions (30%), IT130 (Ideal + hydropriming) showed the highest concentrations in all pigment variables, indicating remarkable stress tolerance in that specific treatment.
2.3.2. Soluble Sugars
Glucose, fructose, and sucrose levels were analyzed using two-way ANOVA (drought stress level x priming treatment) performed separately for each jalapeño variety, followed by Tukey’s HSD test (p ≤ 0.05). Sugar content significantly increased in response to both drought stress and nanopriming treatments (Figure 3E–G).
Under non-stress conditions (0% drought), the Forajido variety treated with ZnO + SiO₂ (FT40) accumulated the highest total sugar content (24.39 mg·g⁻¹ FW). At 15% drought stress, IMT415 (Imperial + ZnO + SiO₂) exhibited significantly elevated levels of glucose, fructose, and sucrose. Under 30% drought stress, IMT230 (Imperial + ZnO + Q) recorded the highest sugar accumulation.
2.3.3. Free Proline
Proline content showed significant variation between treatments and drought levels (p ≤ 0.05), as shown in Figure 4A. Nanopriming with ZnO, especially when combined with SiO₂, resulted in substantial proline accumulation.
Under stress-free conditions (0%), FT40 (Forajido + ZnO + SiO₂) recorded the highest proline level (790.29 ± 254.64 µg·g⁻¹ FW), more than 50 times higher than IT10 (Ideal + hydropriming), which had the lowest value. With 15% drought, FT415 showed the highest accumulation (410.36 ± 104.89 µg·g⁻¹ FW), while IT315 had the lowest (22.64 ± 18.88 µg·g⁻¹ FW). Under severe stress (30%), IT430 led all treatments with 448.50 ± 189.00 µg·g⁻¹ FW, compared to only 53.08 ± 8.61 µg·g⁻¹ FW in IT130. These findings confirm the role of proline in osmotic adjustment and indicate that ZnO and ZnO + SiO₂ priming effectively promote stress tolerance through osmoprotective mechanisms.
2.3.4. Total Phenolic Content (TPC)
Significant differences in TPC were detected between treatments and stress levels (Figure 4B). With 0% stress, FT40 (Forajido + ZnO + SiO₂) had the highest TPC (55.28 ± 5.89 mg GAE·g⁻¹ FW), almost 4.8 times higher than MT10 (Mixteco + Hydropriming), which had the lowest content (11.50 ± 0.41 mg GAE·g⁻¹ FW).
With 15% stress, IT115 (Ideal + hydropriming) showed the highest TPC (26.41 ± 3.68 mg GAE·g⁻¹ FW), while FT215 (Forajido + ZnO) had the lowest (10.81 ± 0.89 mg GAE·g⁻¹ FW). With 30% drought, IT430 maintained the highest phenolic content (51.40 ± 6.35 mg GAE·g⁻¹ FW), representing a 3.2-fold increase compared to IT130 (16.08 ± 10.45 mg GAE·g⁻¹ FW). These data suggest that the combination of ZnO and SiO₂ increases phenol accumulation, especially under severe drought conditions.
2.3.5. Total Flavonoid Content (TFC)
The TFC also showed differences depending on treatment and stress (Figure 4C). Under optimal conditions, FT40 recorded the highest TFC (21.97 ± 8.40 mg QE·g⁻¹ FW), almost 100 times higher than IT40 (0.22 ± 0.13 mg QE·g⁻¹ FW). Under 15% stress, FT115 had the highest TFC (9.00 ± 1.90 mg QE·g⁻¹ FW), while IT315 had the lowest (0.14 ± 0.67 mg QE·g⁻¹ FW). With 30% drought, FT430 retained the highest flavonoid levels (16.20 ± 5.03 mg QE·g⁻¹ FW), 70 times higher than those of FT230. These results highlight the improvement in flavonoid biosynthesis caused by ZnO and SiO₂, especially in combined treatments.
2.3.6. DPPH Antioxidant Activity
Antioxidant activity, evaluated by DPPH radical scavenging, varied significantly among treatments and stress levels (p ≤ 0.05) (Figure 4D). With 0% stress, FT40 showed the highest activity (15.72 ± 1.01 mg TE·g⁻¹ FW), indicating a strong baseline antioxidant potential. Under 15% stress, IT415 showed the highest DPPH activity (5.61 ± 0.01 mg TE·g⁻¹ FW), while under 30% drought, IT430 again led the way (11.53 ± 0.63 mg TE·g⁻¹ FW). These results suggest that nanopriming with ZnO + SiO₂ effectively stimulates antioxidant defense systems over a wide range of drought intensities.
2.4. PCA and Heat Map Analyses
2.4.1. PCA and Heatmap Under 0% Stress
Under stress-free conditions, principal component analysis (PCA) explained 55.23% of the total variance between treatments. PC1 accounted for 33.56% and was mainly associated with morphological traits such as shoot and root length, fresh weight, and chlorophyll content. PC2 explained 21.67% of the variance and was related to biochemical parameters such as proline, total flavonoid content (TFC), and DPPH activity (Figure 5A). This separation indicates that, even in the absence of drought, nanopriming influenced growth and antioxidant responses differently.
The heat map (Figure 5B) revealed strong positive correlations between total chlorophyll content and phenolic content (TPC), as well as DPPH activity (r > 0.87), indicating a functional relationship between photosynthetic pigments and antioxidant capacity under stress-free conditions.
2.4.2. PCA and Heatmap under 15% Stress
With a water stress of 15%, PCA explained 36.61% of the total variation. PC1 grouped morphological traits such as germination speed, shoot length, fresh weight, and vigor indices. PC2 was mainly driven by variables related to antioxidants, such as proline, TFC, and DPPH (Figure 5C). This suggests that nanopriming under moderate drought conditions triggers an integrated physiological and biochemical response.
The corresponding heat map (Figure 5D) highlighted a strong clustering of proline, DPPH, and TFC, with correlation coefficients above 0.85. This indicates a coordinated response of osmoprotective and antioxidant pathways under moderate stress.
2.4.3. PCA and Heatmap under 30% Stress
Under severe water stress conditions (30% mannitol), PCA accounted for 40.74% of the total variance. PC1 captured most of the variation and was dominated by osmoprotective and antioxidant variables—proline, soluble sugars, TPC, and DPPH activity—reflecting a metabolic shift toward biochemical protection. PC2 was largely associated with photosynthetic pigments (chlorophyll a, b, and carotenoids) (Figure 5E).
The heat map (Figure 5F) showed very strong correlations between proline, DPPH, and TPC (r > 0.90), supporting the hypothesis that responses to oxidative and osmotic stress are closely coordinated under high water stress. In addition, chlorophylls and carotenoids showed a high mutual correlation (r = 0.89), indicating synchronized photoprotective mechanisms.
These multivariate patterns demonstrate that the response to drought stress varies depending on the treatment and severity level, and that priming with ZnO and SiO₂, especially in combination, is consistently associated with stress resistance traits.
2.5. Radar Chart: Multivariate Comparison by Drought Level
A radial graph (Figure 6) was used to visualize the comparative performance of nanopriming treatments on 15 key variables and three stress levels (0%, 15%, and 30%). Each axis represents a trait, and each line corresponds to a treatment: ZnO (green), SiO₂ (red), and ZnO + SiO₂ (blue).
Under optimal conditions (0%), ZnO led in carotenoids, shoot length, and total chlorophyll, while SiO₂+ZnO outperformed the others in phenols and DPPH activity. SiO₂ alone improved fresh weight and sugar content, indicating osmotic benefits even in stress-free environments.
With 15% stress, SiO₂ treatment consistently outperformed the others in germination speed, DPPH activity, phenol accumulation, and fresh biomass. ZnO was favorable for vigor and shoot-root balance, while the combined treatment showed a balanced profile in all biochemical and morphological parameters.
With 30% stress, the combination of ZnO + SiO₂ showed superior performance in total chlorophyll, DPPH activity, and TPC, indicating a strong photoprotective and antioxidant response. ZnO improved root development and germination, while SiO₂ stood out in proline accumulation and TPC, reinforcing its role in osmotic adjustment and biochemical defense.
Overall, the radar chart illustrates that nanopriming combined with ZnO and SiO₂ consistently elicited synergistic responses under drought conditions, offering a robust strategy for improving drought resistance in jalapeño pepper seedlings.
3. Discussion
This study demonstrated that nanopriming with ZnO, SiO₂, and their combination effectively improved germination dynamics, early growth, and biochemical responses in jalapeño peppers (Capsicum annuum L.) under different levels of drought stress. The responses observed depended on the type of nanoparticle, the severity of water stress, and the plant variety, highlighting the relevance of interactions between variety, environment, and nanomaterials.
These results support previous findings indicating that nanopriming can improve crop performance under abiotic stress by triggering physiological and biochemical responses that enhance water use efficiency, osmotic regulation, and oxidative stress management [16, 21-22].
Under optimal conditions (0% water stress), priming with ZnO and SiO₂ significantly improved germination parameters. Treatments such as IMT20, FT20, and IMT40 promoted faster and more uniform germination compared to the hydroprimed control. This improved performance can be attributed to increased α-amylase activity and gibberellin signaling, which accelerate starch degradation and reserve mobilization, favoring early seedling development [22]. The significant reduction in average germination time and uncertainty in IT30, a SiO₂ priming treatment, suggests early metabolic activation potentially related to membrane stabilization and increased aquaporin activity [23-24]. Even without water stress, the activation of these physiological mechanisms through nanopriming likely creates a “priming state,” which may confer long-term resilience.
Under moderate water stress (15% mannitol), nanoprimed seeds in treatments such as FT415, IT315, and MT315 maintained high germination rates—up to 100% in FT415 and IT315—and showed vigorous root elongation, shoot development, and biomass accumulation. These responses suggest that nanopriming facilitates the maintenance of cell turgidity, osmotic balance, and early stress signal transduction. This is consistent with previous findings in chickpeas, tomatoes, and okra, where ZnO and SiO₂ NPs improved drought tolerance by maintaining root growth and physiological function [25-28]. Specifically, ZnO-treated seedlings showed improved root architecture, likely due to increased mitotic activity and root system plasticity, which allowed for greater water uptake under conditions of limited availability [29]. In addition, ZnO contributed to the balance between shoots and roots, while SiO₂ improved osmotic regulation and preserved tissue integrity by reinforcing cell wall stability and hydration dynamics [30].
Under severe water stress (30% mannitol), although biomass accumulation decreased, treatments such as FT430, IT430, and MT230 maintained high germination rates and showed strong accumulation of osmoprotectants and antioxidant compounds. This shift from growth promotion to biochemical defense enhancement indicates a physiological balance that prioritizes survival over expansion, a well-established adaptive response under abiotic stress [31]. It is noteworthy that proline levels increased dramatically, especially in ZnO+SiO₂ treatments, confirming its role as an osmoprotectant and ROS scavenger [20].
Proline also showed strong correlations with phenols and DPPH activity, consistent with stress responses in Mentha pulegium and corn [32-33]. Similar synergistic effects of silicon and zinc NPs in improving osmoprotectant accumulation and stress mitigation in wheat under salinity have been reported, where the joint application of proline with Zn and Si NPs significantly improved proline content, antioxidant activity, and plant growth (Abd-Elzaher et al., 2025). These results reinforce the concept that, under conditions of intense drought, nanopriming modulates seedling metabolism towards greater biochemical resilience rather than growth, particularly through the synergistic effects of ZnO and SiO₂.
Similarly, total phenolic content (TPC) and total flavonoid content (TFC) increased significantly in nanoparticle treatments, particularly in FT40 and IT430. These secondary metabolites play an essential role in non-enzymatic antioxidant defense, redox regulation, and cell protection under stress. Their accumulation under optimal and drought conditions suggests that nanopriming may induce a mild oxidative signal that preactivates antioxidant pathways, improving the plant's preparedness for stress [21]. This state of preparedness was supported by the high DPPH radical scavenging activity in the same treatments, indicating a robust antioxidant response. These findings are consistent with previous studies showing that ZnO and SiO₂ NPs improve phenolic biosynthesis and antioxidant potential in crops such as potato and wheat under water-limited conditions [34-36]. Furthermore, evidence obtained with NPs based on Mimusops elengi extracts also supports their role in stimulating phenol production and ROS detoxification [37]. In line with this, the antioxidant effects observed here are consistent with reports on plant systems treated with pink pepper and citrus extracts under oxidative conditions [38].
Multivariate analyses further confirmed the integrative role of nanopriming in modulating growth and defense responses under water stress. Principal component analysis (PCA) revealed that, under 0% stress, germination and morphological traits—such as root length, shoot length, fresh weight, and chlorophyll content—had a large weight in PC1, while antioxidant markers such as proline, total flavonoids (TFC), and DPPH activity contributed predominantly to PC2. This clear separation reflects the dual action of nanopriming in promoting early vigor and activating defense pathways. As drought intensified to 15% and 30% mannitol, the PCA structure changed markedly: osmoprotectants and antioxidants explained most of the variance, indicating a physiological transition toward biochemical prioritization. Similar patterns have been described in chickpea, where PCA-based multivariate selection was driven by biochemical markers under stress conditions [39]. At 30%, variables such as proline and glucose became the main contributors to PC1, highlighting their central role in osmotic adjustment and metabolic reprogramming.
Heat map correlation analysis complemented these findings, revealing strong associations (r > 0.90) between proline, DPPH, phenols, and TFC, especially under moderate and severe stress, indicating a closely coordinated antioxidant and osmotic defense network. These trends mirror observations made in Cicer arietinum and maize, where ROS scavenging, osmotic balance, and root system architecture (RSA) were crucial for drought adaptation [26, 40]. Under low stress, high correlations between chlorophylls, phenols, and DPPH reflected physiological homeostasis. At 15%, the synergy between TFC and proline with fresh weight suggested metabolic flexibility and maintenance of turgidity. At 30%, increased correlations between sugars and antioxidant compounds revealed a trade-off favoring stress survival over growth, a phenomenon described as an adaptation strategy in plants under drought stress [31].
To better visualize the multidimensional performance of each treatment, a radial graph (Figure 6) was constructed that summarizes 15 variables at all stress levels. This graph provided an intuitive and comprehensive view of the relative strengths of each nanopriming treatment. With 0% stress, ZnO treatments stood out in photosynthetic pigment content and shoot development, while SiO₂ + ZnO showed superior antioxidant capacity. SiO₂ alone improved fresh weight and sugar content, highlighting its role in osmotic stabilization even in unstressed seedlings. At 15% stress, SiO₂ consistently obtained the best results in germination speed, antioxidant activity, phenol accumulation, and biomass, suggesting strong ROS detoxification and greater water use efficiency. ZnO was most effective in maintaining the balance between shoots and roots, and the combined ZnO+SiO₂ treatment showed a balanced profile in morphological and biochemical dimensions.
Under severe drought conditions (30%), the superiority of the ZnO+SiO₂ treatment became even more evident. This combined treatment outperformed all others in total chlorophyll, DPPH activity, and phenols, reflecting integrated photoprotective and antioxidant defense. ZnO favored root development and germination under stress, while SiO₂ treatments achieved the highest levels of proline, further reinforcing its osmoprotective role. Thus, the radial graph confirmed the complementarity and potential synergism between these NPs, especially under difficult environmental conditions. It also illustrated the ability of nanopriming to activate different but interconnected response mechanisms depending on the level of drought.
Together, these results underscore the value of nanopriming as a specific, economical, and scalable technology for improving crop performance under drought conditions. In regions such as northern Mexico, where more than 50% of the national jalapeño production occurs and water scarcity is a major constraint [13, 41], the implementation of nanopriming protocols could significantly improve seedling establishment and productivity. Furthermore, the consistency of responses across varieties and stress levels suggests that these technologies could be extended to other crops and production systems.
Together, the evidence indicates that nanopriming with ZnO, SiO₂, and their combination plays a crucial role in improving the physiological and biochemical performance of jalapeño pepper seedlings under drought conditions. These treatments promoted more efficient germination, improved early structural development, and activated antioxidant defense pathways, especially under moderate and severe water deficit conditions. Integrative analysis using a radar chart highlighted the multidimensional advantages of the combined ZnO+SiO₂ treatment, reinforcing its value as a synergistic and promising approach for improving drought resilience during the most vulnerable stages of plant establishment.
4. Materials and Methods
4.1. Experimental Conditions and Plant Material
The experiment was conducted at the Food and Development Research Center (CIAD), Delicias Unit, Chihuahua, Mexico. Four commercial varieties of jalapeño pepper F1 (Capsicum annuum L.) were used: Mixteco, Ideal, Imperial, and Forajido. The seeds were visually inspected to ensure uniformity and then disinfected with 4% sodium hypochlorite for 2 minutes, followed by three rinses with sterile distilled water, as described by Pandya et al. [42].
4.2. Nanoparticle Preparation and Priming Treatments
Zinc oxide (ZnO) and silicon dioxide (SiO₂) NPs were synthesized using the coprecipitation method and characterized by transmission electron microscopy (TEM), which confirmed an average size of 50-80 nm (Figure 7). A solution of 100 mg L⁻¹ of each nanoparticle was prepared in triple-distilled water with 0.1% chitosan as a dispersant. The solutions were homogenized by magnetic stirring (60 min) and sonication (15 min), as described by Waqas et al. [22]. The priming treatments consisted of soaking the seeds in 30 ml of each solution for 12 hours at 25 °C in the dark, followed by drying at room temperature for 24 hours. The treatments were coded according to variety (M: Mixteco, I: Ideal, IM: Imperial, F: Forajido), nanoparticle type (T1: hydropriming, T2: ZnO, T3: SiO₂, T4: ZnO + SiO₂) and stress level (10 = 0%, 115 = 15%, 130 = 30%) (see Table 1). These codes were used consistently in data collection, figures, and analysis (see Figure 1, Figure 2, Figure 3, Figure 4, Figure 5 and Figure 6).
4.3. Experimental Design
A completely randomized design (CRD) was used, with a 4 x 4 x 3 factorial arrangement, where: factor A corresponded to four varieties of jalapeño pepper seeds; factor B consisted of four priming treatments; and factor C consisted of three levels of induced water stress. The effects of nanopriming and water stress levels on germination, initial growth, chlorophyll content, total soluble carbohydrates, free proline, total phenols, total flavonoids, and antioxidant capacity were evaluated using the DPPH method (Figure 8). Each analysis was performed in triplicate.
Each experimental unit corresponded to a Petri dish with 8 seeds, considering each seed as a replicate within its respective treatment.
4.4. Drought-Induced Stress
Water stress was induced using mannitol at concentrations equivalent to 0% (control), 15% (-0.24 MPa), and 30% (-0.48 MPa), following the method of Ertuş and Yazıcılar [10]. Germination was carried out in Petri dishes (9 cm in diameter) lined with Whatman No. 4 filter paper moistened with 5 ml of the corresponding mannitol solution. Eight seeds per dish were incubated under controlled conditions (28 ± 2 °C, 16/8 h light/dark cycle).
4.5. Germination Assay
Germination was monitored daily for 8 days. Seeds were considered germinated when the radicle exceeded 1 mm (Figure 9). Germination indices (percentage, mean germination time, mean germination rate, speed, and uncertainty) were calculated using the GerminaR package [43]. Germination evaluation followed the ISTA [44] guidelines. Results are shown in Table 1.
4.6. Morphological Measurements
On day 8, seedlings were evaluated for shoot length, root length, fresh weight, and stem diameter using a digital caliper. Radicle length was measured from the base of the hypocotyl to the apex of the radicle, while plumule (shoot) length was measured from the radicle–hypocotyl junction to the base of the cotyledons [41]. Stem diameter was measured at approximately 2 mm above the root–hypocotyl transition zone. Vigor indices I and II were calculated according to Abdul-Baki and Anderson [45]
4.7. Biochemical Analyses
Photosynthetic pigments (chlorophyll a, b, total chlorophyll, and carotenoids) were extracted with 99% methanol and quantified spectrophotometrically at 665, 652, and 470 nm, using the equations of Wellburn [47]. The proline content was quantified using the ninhydrin method, using benzene extraction and absorbance at 520 nm. Soluble sugars (glucose, fructose, sucrose) were extracted with 80% ethanol and analyzed using the anthrone method at 620 nm [48].
The total phenolic content (TPC) was determined using the Folin-Ciocalteu method with gallic acid as a standard [49], and the total flavonoid content (TFC) was measured using the AlCl₃ colorimetric method with quercetin as a standard [38]. Antioxidant activity was determined by DPPH radical scavenging [35]. The absorbance for TPC, TFC, and DPPH was measured at 765, 510, and 517 nm, respectively. The data for these biochemical traits are shown in Figure 3 and Figure 4.
4.8. Multivariate and Correlation Analysis
Principal component analysis (PCA) and Pearson correlation matrices were performed using OriginPro 2025 to evaluate relationships between morphological and biochemical traits. PCA biplots were generated separately for each drought level (0%, 15%, 30%) (Figure 5A,C, E), and heatmaps were constructed to visualize correlations between traits and treatments (Figure 5B,D,F) [39]. A radar chart (Figure 6) was created to integrate germination, growth, and biochemical performance of ZnO, SiO₂, and ZnO+SiO₂ treatments across the three stress levels, normalizing 15 key variables.
4.10. Statistical Analysis
Each variable was represented by three replicates. Germination parameters were analyzed using the nonparametric Kruskal-Wallis test followed by Dunn's post hoc test (p ≤ 0.05), due to the non-normal distribution of the data.
For early growth variables and biochemical parameters such as photosynthetic pigments and soluble sugars, the assumptions of normality, homogeneity of variances, and independence were tested. When these assumptions were met, a two-way ANOVA (drought stress level × priming treatment) was performed separately by variety. Tukey’s HSD test (p ≤ 0.05) was used to determine significant differences between means.
For antioxidant-related variables—total phenols, flavonoids, proline content, and DPPH radical scavenging activity—a three-way ANOVA was conducted to evaluate the effects of priming treatment, drought stress level, and variety, including their interactions. Post hoc comparisons were performed using Tukey’s HSD test (p ≤ 0.05).
All statistical analyses were carried out using SAS® software version 9.0 (SAS Institute Inc., Cary, NC, USA [50]). Figures, bar charts, radar charts, Pearson correlation matrices, and principal component analysis (PCA) plots were generated using OriginPro 2025 (v10.2.0.196).
5. Conclusions
Nanopriming with ZnO, SiO₂ and their combination significantly improved drought tolerance in jalapeño peppers by enhancing germination, seedling vigor, and antioxidant capacity under both moderate and severe water stress conditions. The combined treatment with ZnO + SiO₂ produced the most consistent improvements in physiological and biochemical parameters, indicating a synergistic effect in promoting stress resilience in the early stages. These findings support the integration of nanopriming as a pre-planting strategy to optimize seed performance under water-scarce conditions. By strengthening seedling establishment and improving metabolic responses under drought conditions, this approach contributes to improving water use efficiency and promoting more sustainable water resource management in crop production systems.
Author Contributions
The authors confirm contribution to the paper as follows: study conception and design: E.H.O.-C. and E.S.; data collection: L.U.C.-E., S.P.-Á. and J.J.P.-C..; analysis and interpretation of results: C.C.-M., J.C.A.-P. and E.M.-M.; draft manuscript preparation: E.H.O.-C. and E.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The authors declare that all data discussed in the study are available in the manuscript.
Acknowledgments
We would like to thank the Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI—Mexico) for the support provided by the scholarship granted to Erick Humberto Ochoa Chaparro through the “Becas Nacionales SECIHTI” program with CVU No. 843732.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Dietz, K.J.; Zörb, C.; Geilfus, C.M. Drought and crop yield. Plant Biol. 2021, 23, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Rai-Kalal, P.; Tomar, R.S.; Jajoo, A. Seed nanopriming by silicon oxide improves drought stress alleviation potential in wheat plants. Funct. Plant Biol. 2021, 48, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Figueroa, K.I.; Sánchez, E.; Ramírez-Estrada, C.A.; Anchondo-Páez, J.C.; Ojeda-Barrios, D.L.; Pérez-Álvarez, S. Efficacy and Differential Physiological–Biochemical Response of Biostimulants in Green Beans Subjected to Moderate and Severe Water Stress. Crops 2024, 4, 27–42. [Google Scholar] [CrossRef]
- Hossain, M.S.; Li, J.; Wang, C.; Monshi, F.I.; Tabassum, R.; Islam, M.A.; Faruquee, M.; Muktadir, M.A.; Mia, M.S.; Islam, A.K.M.M.; Hasan, A.K.; Sikdar, A.; Feng, B. Enhanced antioxidant activity and secondary metabolite production in tartary buckwheat under polyethylene glycol (PEG)-induced drought stress during germination. Agronomy 2024, 14, 619. [Google Scholar] [CrossRef]
- Takahashi, F.; Kuromori, T.; Urano, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Drought stress responses and resistance in plants: From cellular responses to long-distance intercellular communication. Front. Plant Sci. 2020, 11, 556972. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Saadaoui, W.; Tarchoun, N.; Msetra, I.; Pavli, O.; Falleh, H.; Ayed, C.; Amami, R.; Ksouri, R.; Petropoulos, S.A. Effects of drought stress induced by D-mannitol on the germination and early seedling growth traits, physiological parameters and phytochemicals content of Tunisian squash (Cucurbita maxima Duch.) landraces. Front. Plant Sci. 2023, 14, 1215394. [Google Scholar] [CrossRef] [PubMed]
- Qiao, M.; Hong, C.; Jiao, Y.; Hou, S.; Gao, H. Impacts of drought on photosynthesis in major food crops and the related mechanisms of plant responses to drought. Plants 2024, 13, 1808. [Google Scholar] [CrossRef]
- Ievinsh, G. Water content of plant tissues: So simple that almost forgotten? Plants 2023, 12, 1238. [Google Scholar] [CrossRef]
- Ertuş, M.M.; Yazıcılar, B. CaO nanoparticle enhances the seedling growth of Onobrychis viciifolia under drought stress via mannitol use. Biologia 2023, 78, 1119–1127. [Google Scholar] [CrossRef]
- Altaf, M.A.; Hao, Y.; He, C.; Mumtaz, M.A.; Shu, H.; Fu, H.; Wang, Z. Physiological and biochemical responses of pepper (Capsicum annuum L.) seedlings to nickel toxicity. Front. Plant Sci. 2022, 13, 950392. [Google Scholar] [CrossRef] [PubMed]
- García-Vásquez, R.; Vera-Guzmán, A.M.; Carrillo-Rodríguez, J.C.; Pérez-Ochoa, M.L.; Aquino-Bolaños, E.N.; Alba-Jiménez, J.E.; Chávez-Servia, J.L. Bioactive and nutritional compounds in fruits of pepper (Capsicum annuum L.) landraces conserved among indigenous communities from Mexico. AIMS Agric. Food 2023, 8, 832–850. [Google Scholar] [CrossRef]
- Lozada, D.N.; Pulicherla, S.R.; Holguin, F.O. Widely targeted metabolomics reveals metabolite diversity in jalapeño and serrano chile peppers (Capsicum annuum L.). Metabolites 2023, 13, 288. [Google Scholar] [CrossRef]
- Magaña, V.; Herrera, E.; Ábrego-Góngora, C.J.; Ávalos, J.A. Socioeconomic drought in a Mexican semi-arid city: Monterrey Metropolitan Area, a case study. Front. Water 2021, 3, 579564. [Google Scholar] [CrossRef]
- Pacheco-Treviño, S.; Manzano-Camarillo, M.G. The socioeconomic dimensions of water scarcity in urban and rural Mexico: A comprehensive assessment of sustainable development. Sustainability 2024, 16, 1011. [Google Scholar] [CrossRef]
- Singh, A.; Sengar, R.S.; Sharma, R.; Rajput, P.; Singh, A.K. Nano-priming technology for sustainable agriculture. Biogeosyst. Tech. 2021, 8, 79–92. [Google Scholar]
- Tamindžić, G.; Azizbekian, S.; Miljaković, D.; Ignjatov, M.; Nikolić, Z.; Budakov, D.; Vasiljević, S.; Grahovac, M. Assessment of various nanoprimings for boosting pea germination and early growth in both optimal and drought-stressed environments. Plants 2024, 13, 1547. [Google Scholar] [CrossRef]
- Abbas, S.F.; Bukhari, M.A.; Raza, M.A.S.; Abbasi, G.H.; Ahmad, Z.; Alqahtani, M.D.; Almutairi, K.F.; Abd_Allah, E.F.; Iqbal, M.A. Enhancing drought tolerance in wheat cultivars through nano-ZnO priming by improving leaf pigments and antioxidant activity. Sustainability 2023, 15, 5835. [Google Scholar] [CrossRef]
- Sharf-Eldin, A.A.; Alwutayd, K.M.; El-Yazied, A.A.; El-Beltagi, H.S.; Alharbi, B.M.; Eisa, M.A.M.; Alqurashi, M.; Sharaf, M.; Harbi, N.A.; Al-Qahtani, S.M.; Ibrahim, M.F.M. Response of maize seedlings to silicon dioxide nanoparticles (SiO₂NPs) under drought stress. Plants 2023, 12, 2592. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Pratyusha, S. Phenolic compounds in the plant development and defense. In Plant Stress Physiology: Perspectives in Agriculture; IntechOpen: London, UK, 2022; p. 125. [Google Scholar]
- Waqas Mazhar, M.; Ishtiaq, M.; Hussain, I.; Parveen, A.; Hayat Bhatti, K.; Azeem, M.; Thind, S.; Ajaib, M.; Maqbool, M.; Sardar, T. Seed nano-priming with zinc oxide nanoparticles in rice mitigates drought and enhances agronomic profile. PLoS ONE 2022, 17, e0264967. [Google Scholar] [CrossRef]
- Chávez Suárez, L.; Álvarez Fonseca, A.; Ramírez Fernández, R. Aspectos de interés sobre las acuaporinas en las plantas. Cultiv. Trop. 2014, 35, 45–54. [Google Scholar]
- Ashkavand, P.; Tabari, M.; Zarafshar, M.; Tomášková, I.; Struve, D. Effect of SiO₂ nanoparticles on drought resistance in hawthorn seedlings. Leśne Pr. Badawcze 2015, 76, 350–359. [Google Scholar]
- Haghighi, M.; Da Silva, J.T.; Mozafarian, M.; Afifipour, Z. Can Si and nano-Si alleviate the effect of drought stress induced by PEG in seed germination and seedling growth of tomato. Minerva Biotecnol. 2013, 25, 17–22. [Google Scholar]
- Shah, T.M.; Imran, M.; Atta, B.M.; Ashraf, M.Y.; Hameed, A.; Waqar, I.; Shafiq, M.; Hussain, K.; Naveed, M.; Aslam, M.; Maqbool, M.A. Selection and screening of drought tolerant high yielding chickpea genotypes based on physio-biochemical indices and multi-environmental yield trials. BMC Plant Biol. 2020, 20, 171. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, J.; Yang, M.; Miao, M. Influence of SiO₂, Al₂O₃ and TiO₂ nanoparticles on okra seed germination under PEG-6000 simulated water deficit stress. J. Seed Sci. 2023, 45, e202345029. [Google Scholar] [CrossRef]
- Hanif, S.; Farooq, S.; Kiani, M.Z.; Zia, M. Surface modified ZnO NPs by betaine and proline build up tomato plants against drought stress and increase fruit nutritional quality. Chemosphere 2024, 362, 142671. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Ilyas, N.; Mashwani, Z.-u.-R.; Hayat, R.; Yasmin, H.; Nouredeen, A.; Ahmad, P. Synergistic effects of plant growth promoting rhizobacteria and silicon dioxide nanoparticles for amelioration of drought stress in wheat. Plant Physiol. Biochem. 2021, 166, 160–176. [Google Scholar] [CrossRef]
- Behboudi, F.; Tahmasebi Sarvestani, Z.; Kassaee, M.Z.; Modares Sanavi, S.A.M.; Sorooshzadeh, A. Improving growth and yield of wheat under drought stress via application of SiO₂ nanoparticles. J. Agric. Sci. Technol. 2018, 20, 1479–1492. [Google Scholar]
- He, Z.; Webster, S.; He, S.Y. Growth–defense trade-offs in plants. Curr. Biol. 2022, 32, R634–R639. [Google Scholar] [CrossRef]
- El-Naggar, H.M.; Osman, A.R. Enhancing growth and bioactive metabolites characteristics in Mentha pulegium L. via silicon nanoparticles during in vitro drought stress. BMC Plant Biol. 2024, 24, 657. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Song, F.; Guo, J.; Zhu, X.; Liu, S.; Liu, F.; Li, X. Nano-ZnO-induced drought tolerance is associated with melatonin synthesis and metabolism in maize. Int. J. Mol. Sci. 2020, 21, 782. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elzaher, M.A.; El-Desoky, M.A.; Khalil, F.A.; Eissa, M.A.; Amin, A.E.-E.A. Exogenously applied proline with silicon and zinc nanoparticles to mitigate salt stress in wheat plants grown on saline soil. J. Plant Nutr. 2025, 48, 1559–1576. [Google Scholar] [CrossRef]
- Pagani, A.A.C.; de Souza, A.L.G.; Souza, D.S.; Batista, R.A.; Xavier, A.C.R.; Pagani, G.D. Quantification of bioactive compounds of pink pepper (Schinus terebinthifolius, Raddi). Int. J. Eng. Innov. Technol. 2014, 4, 37–41. [Google Scholar]
- Al-Selwey, W.A.; Alsadon, A.A.; Alenazi, M.M.; Tarroum, M.; Ibrahim, A.A.; Ahmad, A.; Osman, M.; Seleiman, M.F. Morphological and biochemical response of potatoes to exogenous application of ZnO and SiO₂ nanoparticles in a water deficit environment. Horticulturae 9, 883. [CrossRef]
- Rakhman, S.A.; Utaipan, T.; Pakhathirathien, C.; Khummueng, W. Metal and metal oxide nanoparticles from Mimusops elengi Linn. extract: Green synthesis, antioxidant activity, and cytotoxicity. Sains Malays. 2022, 51, 2857–2871. [Google Scholar] [CrossRef]
- Ghasemi, K.; Ghasemi, Y.; Ebrahimzadeh, M.A. Antioxidant activity, phenol and flavonoid contents of 13 citrus species peels and tissues. Pak. J. Pharm. Sci. 2009, 22, 277–281. [Google Scholar]
- Basavaraj, P.S.; Rane, J.; Jangid, K.K.; Babar, R.; Kumar, M.; Gangurde, A.; Shinde, S.; Boraiah, K.M.; Harisha, C.B.; Halli, H.M.; Reddy, K.S.; Prabhakar, M. Index-based selection of chickpea (Cicer arietinum L.) genotypes for enhanced drought tolerance. Sci. Rep. 2025, 15, 8282. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Sinha, R.; Singla-Pareek, S.L.; Pareek, A.; Singh, A.K. Shaping the root system architecture in plants for adaptation to drought stress. Physiol. Plant. 2022, 174, e13651. [Google Scholar] [CrossRef]
- Ochoa-Chaparro, E.H.; Ramírez-Estrada, C.A.; Anchondo-Páez, J.C.; Sánchez, E.; Pérez-Álvarez, S.; Castruita-Esparza, L.U.; Muñoz-Márquez, E.; Chávez-Mendoza, C.; Patiño-Cruz, J.J.; Franco-Lagos, C.L. Nanopriming with zinc–molybdenum in jalapeño pepper on imbibition, germination, and early growth. Agronomy 2024, 14, 1609. [Google Scholar] [CrossRef]
- Pandya, P.; Kumar, S.; Sakure, A.A.; Rafaliya, R.; Patil, G.B. Zinc oxide nanopriming elevates wheat drought tolerance by inducing stress-responsive genes and physio-biochemical changes. Curr. Plant Biol. 2023, 35, 100292. [Google Scholar] [CrossRef]
- Lozano-Isla, F.; Benites-Alfaro, O.E.; Pompelli, M.F. GerminaR: An R package for germination analysis with the interactive web application “GerminaQuant for R”. Ecol. Res. 2019, 34, 339–346. [Google Scholar] [CrossRef]
- International Seed Testing Association (ISTA). International Rules for Seed Testing 2020; ISTA: Bassersdorf, Switzerland, 2020. [Google Scholar]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor determination in soybean seed by multiple criteria. Crop Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Elizondo-Cabalceta, E.; Monge-Pérez, J.E. Caracterización morfológica de 15 genotipos de pimiento (Capsicum annuum L.) cultivados bajo invernadero en Costa Rica. InterSedes 2017, 18, 1–27. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Ornelas-Paz, J.; Martínez-Burrola, J.M.; Ruiz-Cruz, S.; Santana-Rodríguez, V.; Ibarra-Junquera, V.; Olivas, G.I.; Pérez-Martínez, J.D. Effect of cooking on the capsaicinoids and phenolics contents of Mexican peppers. Food Chem. 2010, 119, 1619–1625. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT® 15.2 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2021. [Google Scholar]
Figure 1.
Morphological response of four varieties under nanopriming and drought stress. Panels (A-L) show shoot length, root length, stem diameter, and fresh weight in four jalapeño varieties (Mixteco, Ideal Imperial and Forajido) under three drought levels (0%, 15% and 30%) and four treatments : T1 (Hydropriming), T2 (ZnO +Q), T3 (SiO2 + Q), and T4 (ZnO + SiO2 + Q). Bars represent mean ± SD. Different letters indicate significant differences (Tukey HSD, p ≤ 0.05).
Figure 1.
Morphological response of four varieties under nanopriming and drought stress. Panels (A-L) show shoot length, root length, stem diameter, and fresh weight in four jalapeño varieties (Mixteco, Ideal Imperial and Forajido) under three drought levels (0%, 15% and 30%) and four treatments : T1 (Hydropriming), T2 (ZnO +Q), T3 (SiO2 + Q), and T4 (ZnO + SiO2 + Q). Bars represent mean ± SD. Different letters indicate significant differences (Tukey HSD, p ≤ 0.05).
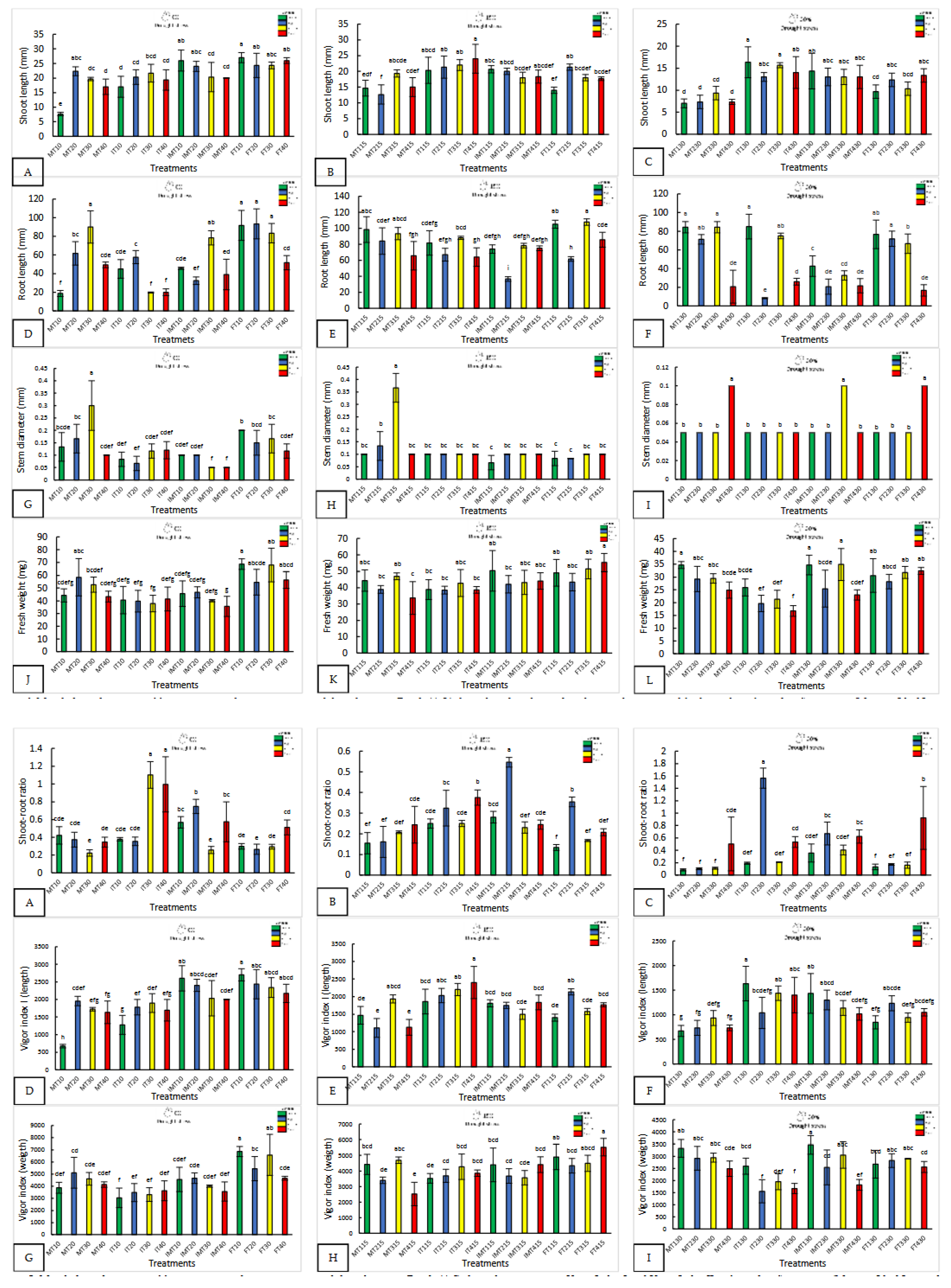
Figure 2.
Morphological response of four varieties under nanopriming and drought stress. Panels (A-I) show shoot-root ratio, Vigor Index I and Vigor Index II in four jalapeño varieties (Mixteco, Ideal Imperial and Forajido) under three drought levels (0%, 15% and 30%) and four treatments: T1 (Hydropriming), T2 (ZnO +Q), T3 (SiO2 + Q), and T4 (ZnO + SiO2 + Q). Bars represent mean ± SD. Different letters indicate significant differences (Tukey HSD, p ≤ 0.05).
Figure 2.
Morphological response of four varieties under nanopriming and drought stress. Panels (A-I) show shoot-root ratio, Vigor Index I and Vigor Index II in four jalapeño varieties (Mixteco, Ideal Imperial and Forajido) under three drought levels (0%, 15% and 30%) and four treatments: T1 (Hydropriming), T2 (ZnO +Q), T3 (SiO2 + Q), and T4 (ZnO + SiO2 + Q). Bars represent mean ± SD. Different letters indicate significant differences (Tukey HSD, p ≤ 0.05).
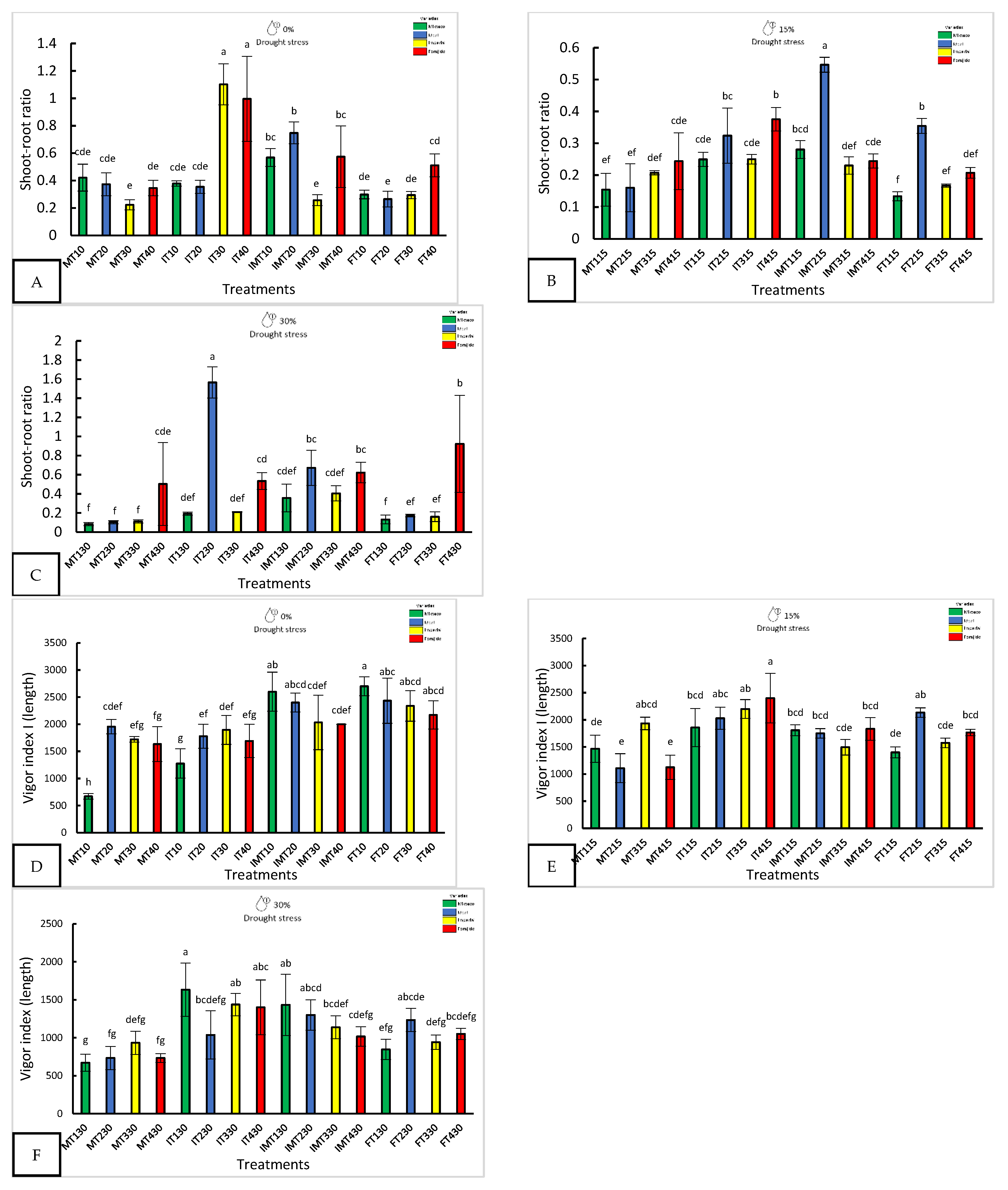
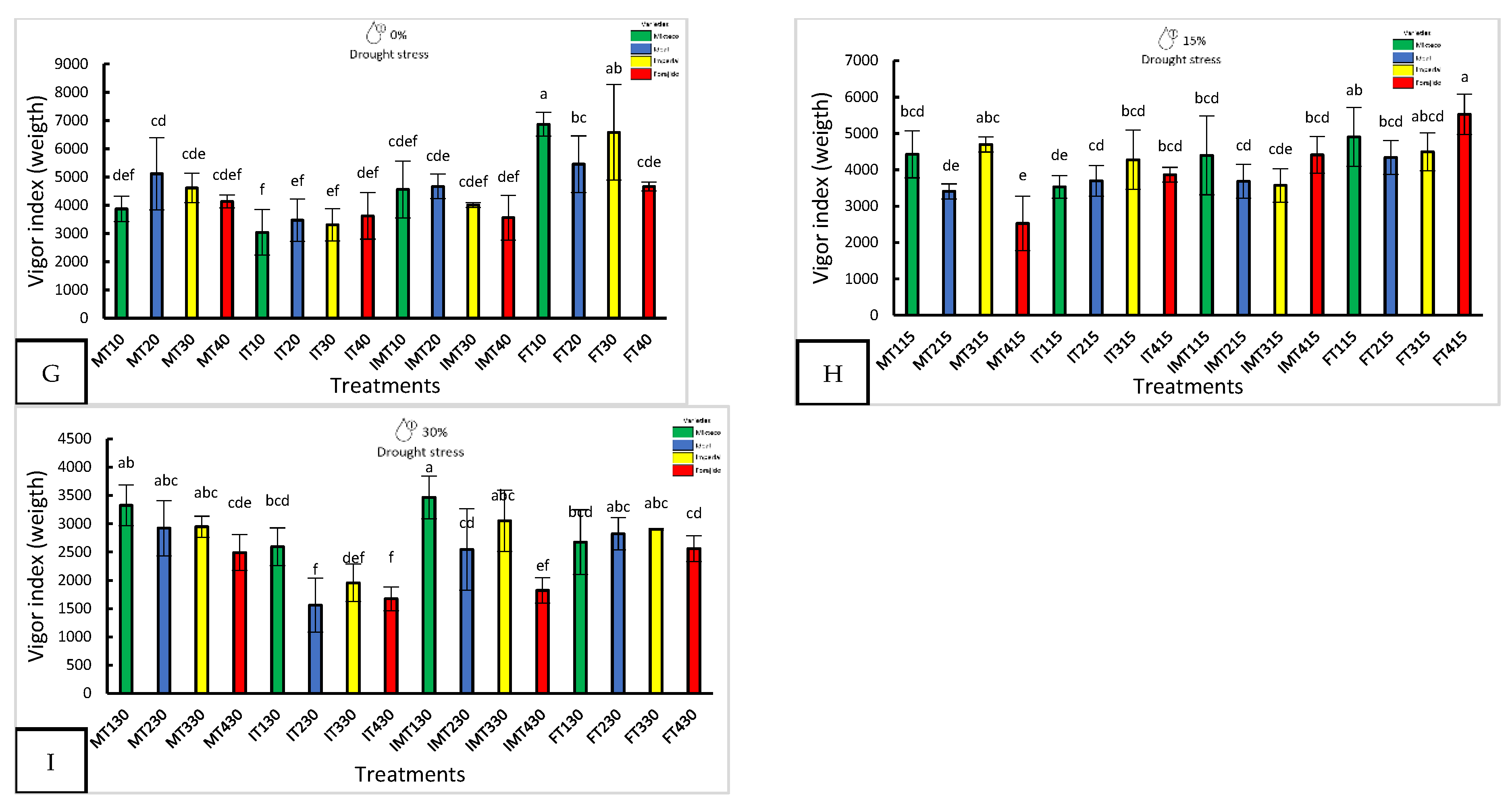
Figure 3.
Effect of nanopriming treatments and drought stress levels on the photosynthetic pigments and soluble sugar composition of four jalapeños pepper varieties. Panels (A–G) display the response in the varieties Mixteco, Ideal, Imperial and Forajido under three drought stress levels (0%, 15%, and 30%) and four nanopriming treatments: T1 (Hydropriming), T2 (ZnO + Q), T3 (SiO₂ + Q), and T4 (ZnO + SiO₂ + Q). The variables assessed were: (A) Chlorophyll (a); (B) Chlorophyll (b); (C) Chlorophyll total; (D) Glucose; (E) Fructose; (F) Sucrose. Each data point represents the mean ± standard deviation. Different letters indicate statistically significant differences (Tukey HSD, p ≤ 0.05Under severe drought conditions (30%), FT230 and MT330 achieved complete germination, while IMT430 and FT430 showed reduced performance (79.16%). Notably, FT230 also had the lowest uncertainty index (0.86 ± 0.08), indicating greater synchronization in germination events.
Figure 3.
Effect of nanopriming treatments and drought stress levels on the photosynthetic pigments and soluble sugar composition of four jalapeños pepper varieties. Panels (A–G) display the response in the varieties Mixteco, Ideal, Imperial and Forajido under three drought stress levels (0%, 15%, and 30%) and four nanopriming treatments: T1 (Hydropriming), T2 (ZnO + Q), T3 (SiO₂ + Q), and T4 (ZnO + SiO₂ + Q). The variables assessed were: (A) Chlorophyll (a); (B) Chlorophyll (b); (C) Chlorophyll total; (D) Glucose; (E) Fructose; (F) Sucrose. Each data point represents the mean ± standard deviation. Different letters indicate statistically significant differences (Tukey HSD, p ≤ 0.05Under severe drought conditions (30%), FT230 and MT330 achieved complete germination, while IMT430 and FT430 showed reduced performance (79.16%). Notably, FT230 also had the lowest uncertainty index (0.86 ± 0.08), indicating greater synchronization in germination events.
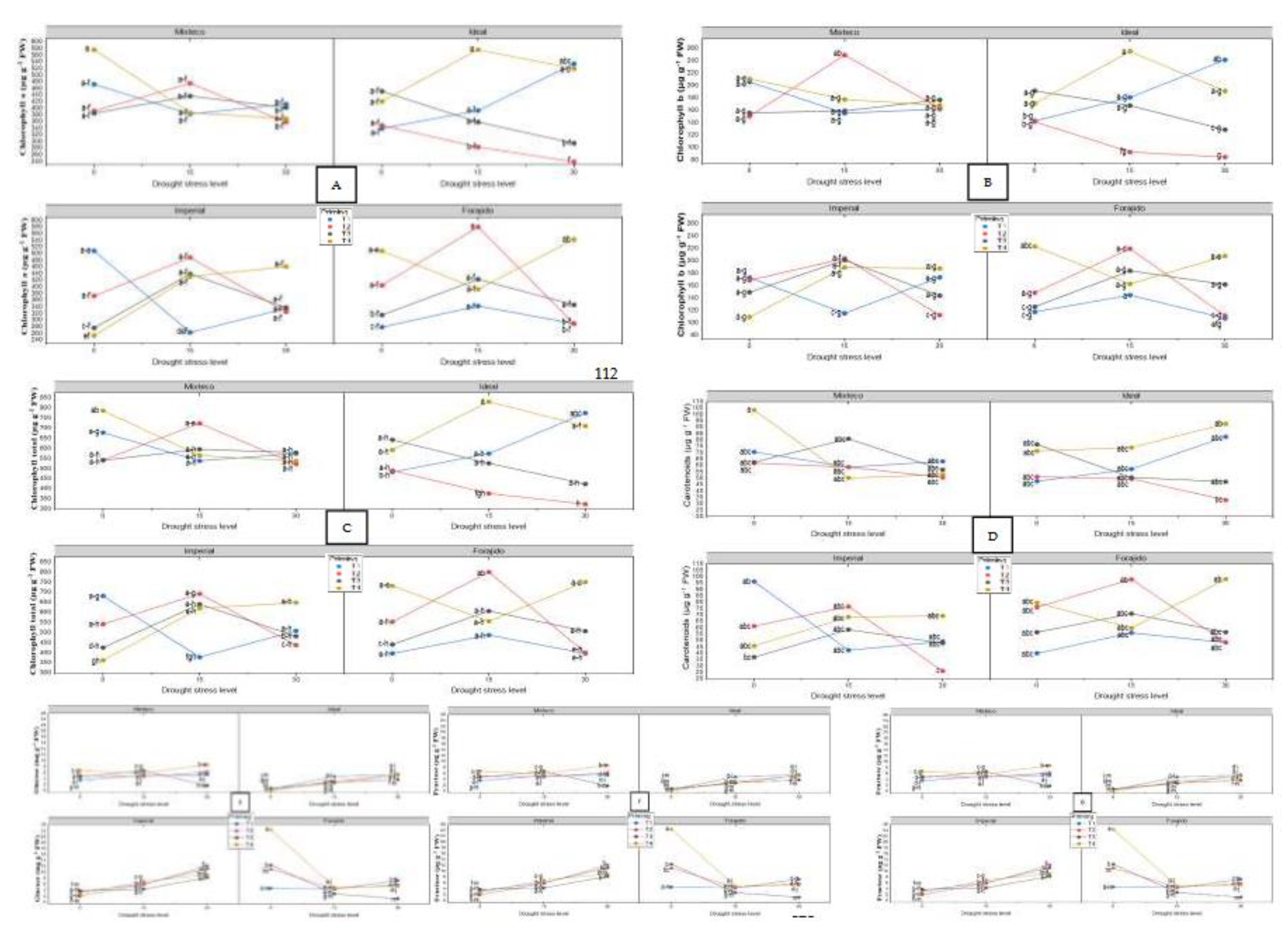
Figure 4.
Biochemical responses in four jalapeño pepper varieties under different nanopriming treatments and drought stress levels. The panels show: (A) Free proline content (μg g⁻¹ FW), (B) Total phenolic content (mg GAE g⁻¹ FW), (C) Total flavonoid content (mg QE g⁻¹ FW), and (D) DPPH radical scavenging activity (mg TE g⁻¹ FW). Bars represent the mean ± standard deviation for each variable. The treatments applied were T1 (Hydropriming), T2 (ZnO + Q), T3 (SiO₂ + Q), and T4 (ZnO + SiO₂ + Q), across three drought stress levels (0%, 15%, and 30%) in the varieties Mixteco, Ideal, Imperial, and Forajido. Letters above the bars indicate significant differences between treatments according to a Tukey HSD test (p ≤ 0.05).
Figure 4.
Biochemical responses in four jalapeño pepper varieties under different nanopriming treatments and drought stress levels. The panels show: (A) Free proline content (μg g⁻¹ FW), (B) Total phenolic content (mg GAE g⁻¹ FW), (C) Total flavonoid content (mg QE g⁻¹ FW), and (D) DPPH radical scavenging activity (mg TE g⁻¹ FW). Bars represent the mean ± standard deviation for each variable. The treatments applied were T1 (Hydropriming), T2 (ZnO + Q), T3 (SiO₂ + Q), and T4 (ZnO + SiO₂ + Q), across three drought stress levels (0%, 15%, and 30%) in the varieties Mixteco, Ideal, Imperial, and Forajido. Letters above the bars indicate significant differences between treatments according to a Tukey HSD test (p ≤ 0.05).
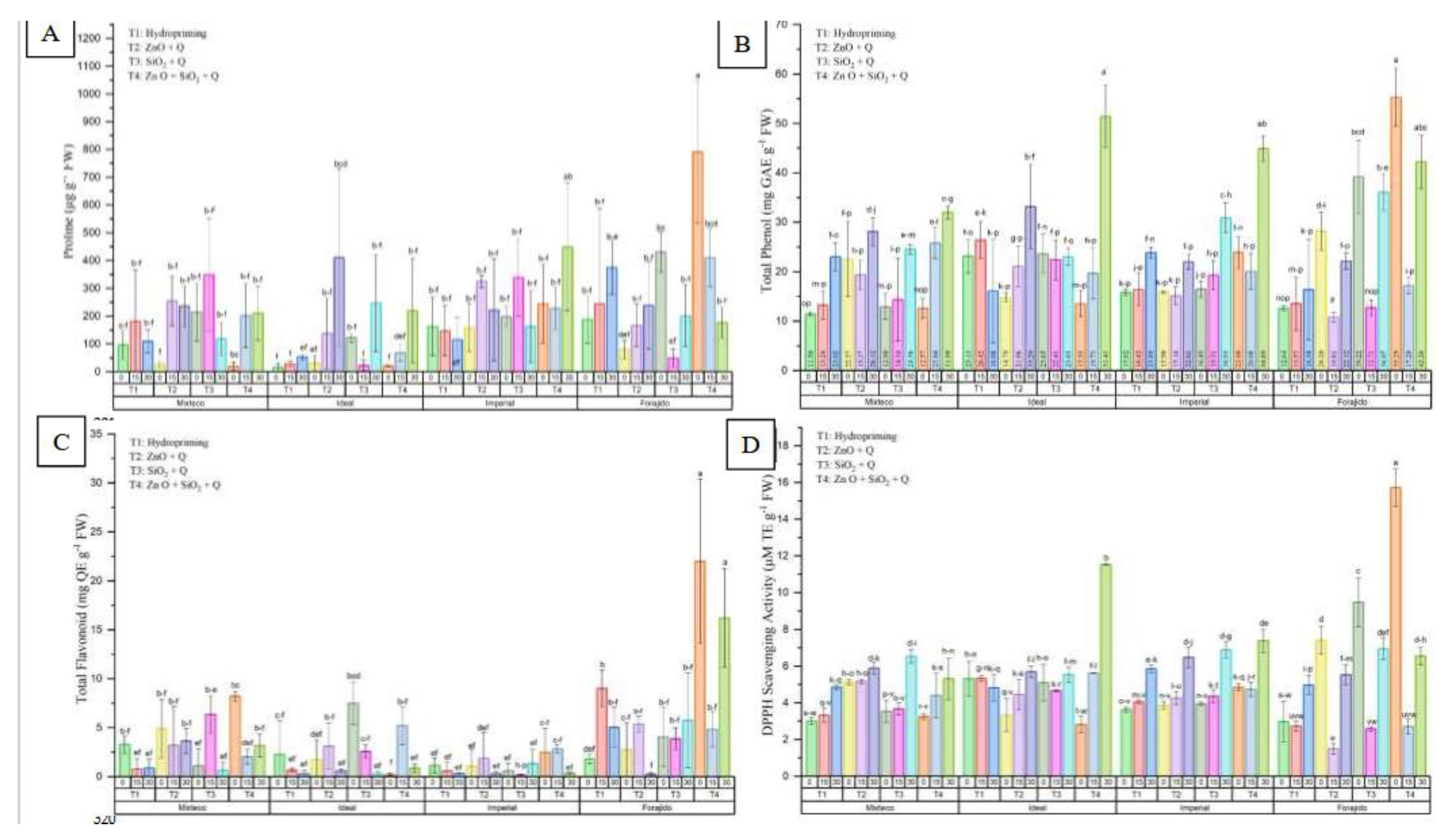
Figure 5.
Principal Component Analysis (PCA) and Pearson Correlation Heatmaps under Different Water Stress Levels. Panels (A), (C), and (E) represent the PCA biplots of morphophysiological and biochemical traits evaluated in four jalapeño pepper varieties (MIXTECO, IDEAL, IMPERIAL, and FORAJIDO) under 0%, 15%, and 30% water stress conditions, respectively. The ellipses represent the distribution and clustering of each variety based on the first two principal components. Panels (B), (D), and (F) show the corresponding Pearson correlation heatmaps for the same variables and stress levels. Positive correlations are shown in red and negative in blue, with the intensity indicating the strength of the correlation. These analyses highlight the multivariate relationships and changes in trait associations as water stress intensifies.
Figure 5.
Principal Component Analysis (PCA) and Pearson Correlation Heatmaps under Different Water Stress Levels. Panels (A), (C), and (E) represent the PCA biplots of morphophysiological and biochemical traits evaluated in four jalapeño pepper varieties (MIXTECO, IDEAL, IMPERIAL, and FORAJIDO) under 0%, 15%, and 30% water stress conditions, respectively. The ellipses represent the distribution and clustering of each variety based on the first two principal components. Panels (B), (D), and (F) show the corresponding Pearson correlation heatmaps for the same variables and stress levels. Positive correlations are shown in red and negative in blue, with the intensity indicating the strength of the correlation. These analyses highlight the multivariate relationships and changes in trait associations as water stress intensifies.
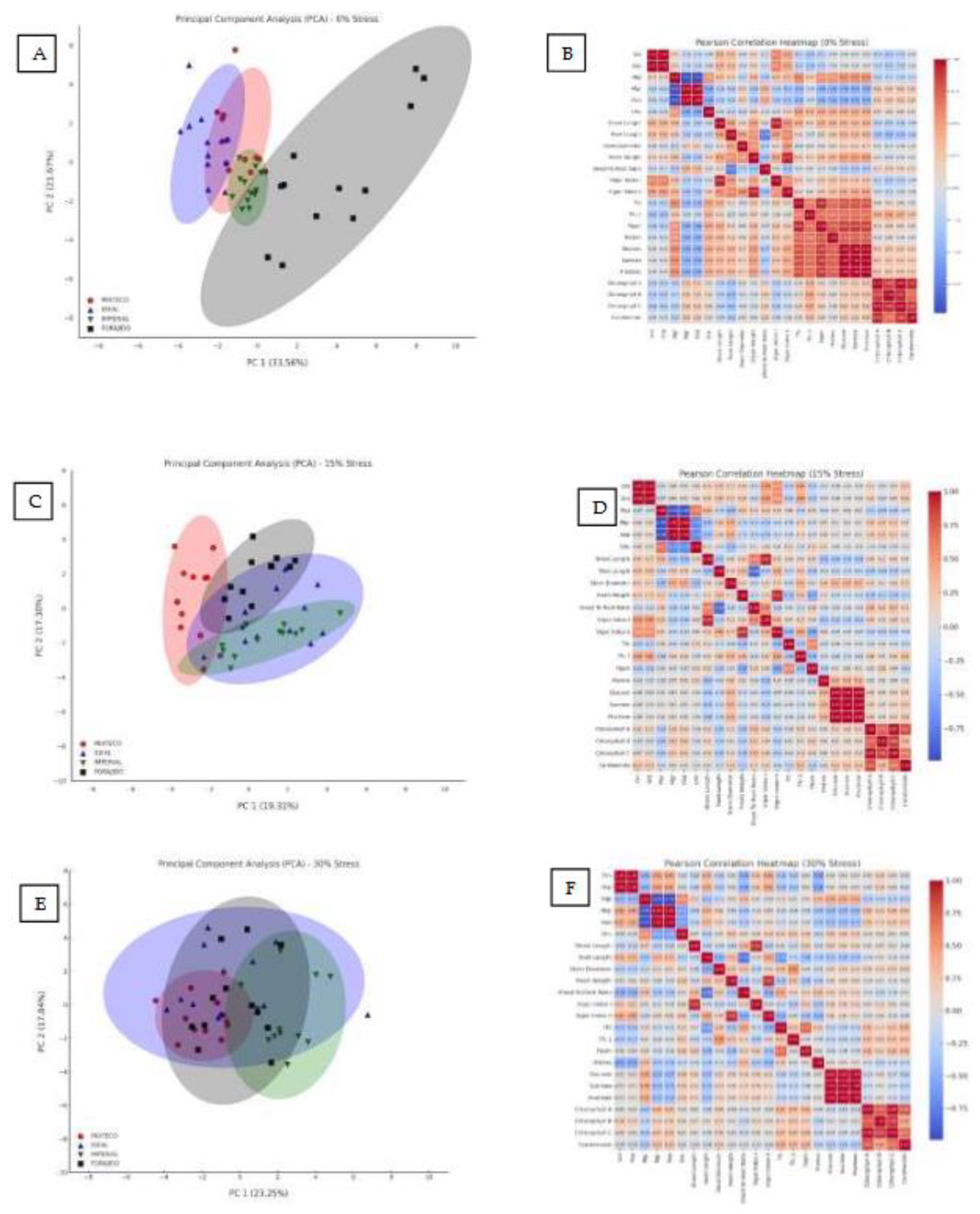
Figure 6.
Radar chart comparing ZnO (green), SiO₂ (red), and ZnO+SiO₂ (blue) nanopriming in jalapeño under 0%, 15%, and 30% drought stress. Fifteen variables were evaluated. ZnO+SiO₂ showed the best overall performance at 30% stress, while SiO₂ stood out at 15%. Combined treatments improved both growth and antioxidant responses under drought.
Figure 6.
Radar chart comparing ZnO (green), SiO₂ (red), and ZnO+SiO₂ (blue) nanopriming in jalapeño under 0%, 15%, and 30% drought stress. Fifteen variables were evaluated. ZnO+SiO₂ showed the best overall performance at 30% stress, while SiO₂ stood out at 15%. Combined treatments improved both growth and antioxidant responses under drought.
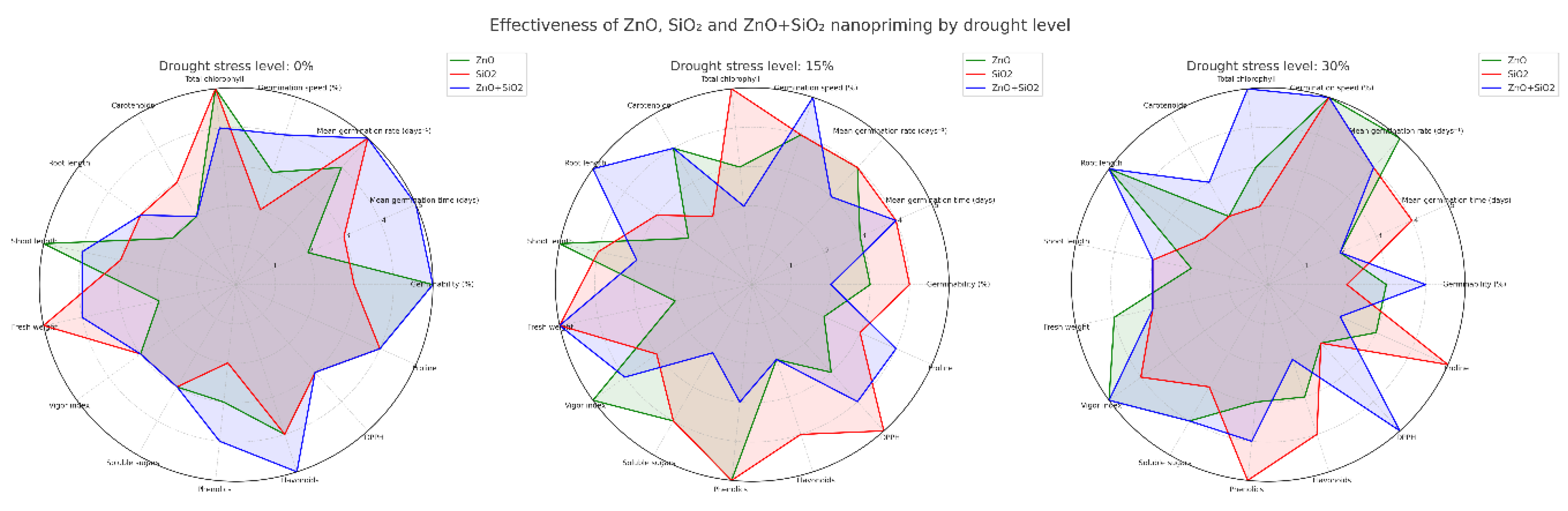
Figure 7.
Morphology of the sample by Transmission Electron Microscopy (TEM), (A) ZnO, (B) SiO2.

Figure 8.
Experimental design with a 4 × 4 × 3 factorial arrangement, evaluating four varieties of jalapeño pepper, four priming treatments and three levels of induced water stress.
Figure 8.
Experimental design with a 4 × 4 × 3 factorial arrangement, evaluating four varieties of jalapeño pepper, four priming treatments and three levels of induced water stress.
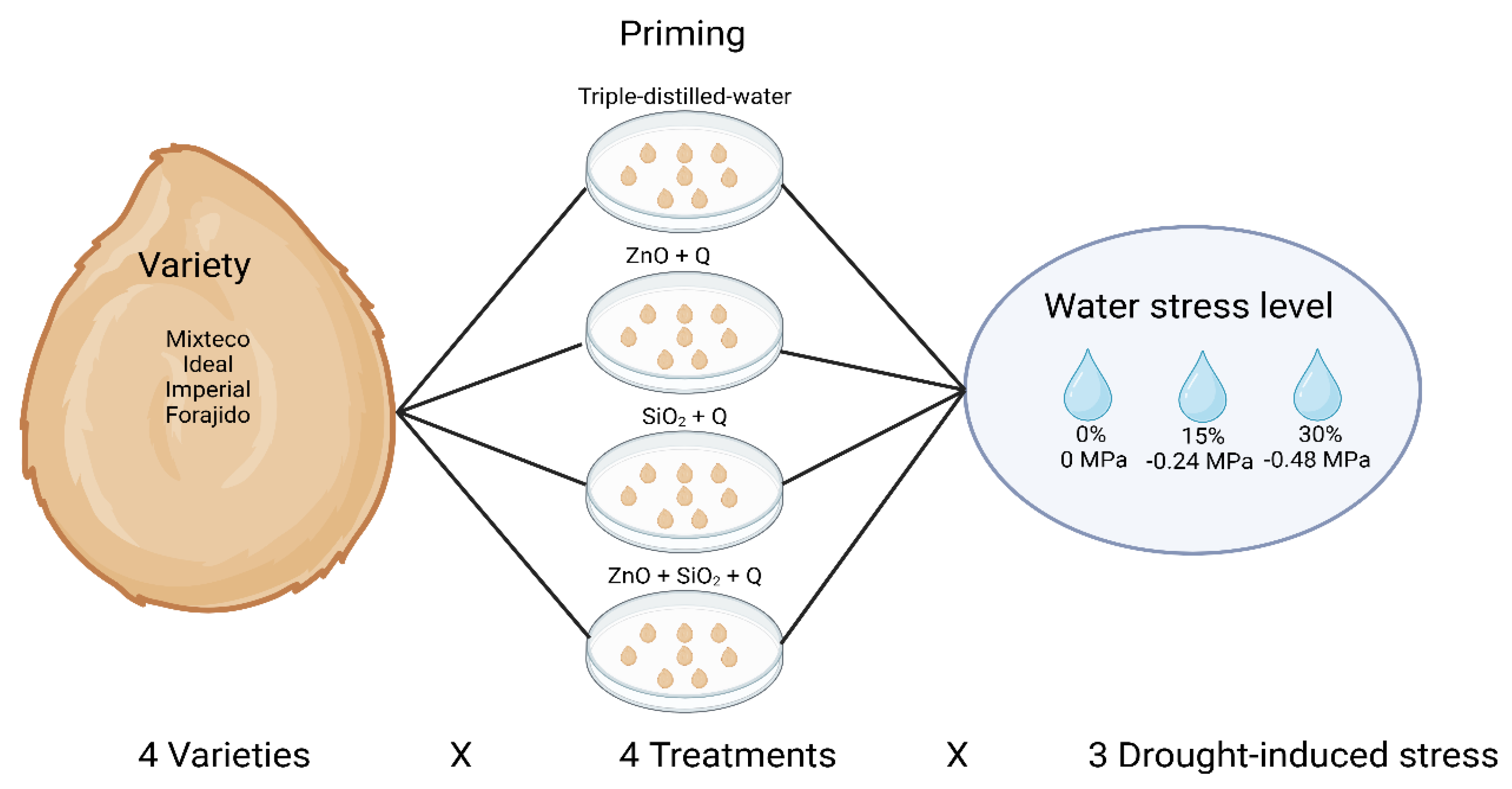
Figure 9.
Germination criteria radicle length 1 to 2 mm (Binocular stereo microscope with VE-S4 zoom system (Velab Co., Pharr, Texas, USA).
Figure 9.
Germination criteria radicle length 1 to 2 mm (Binocular stereo microscope with VE-S4 zoom system (Velab Co., Pharr, Texas, USA).

Table 1.
Germination parameters of four commercial varieties of jalapeño pepper under different priming treatments.
Table 1.
Germination parameters of four commercial varieties of jalapeño pepper under different priming treatments.
| Under 15% water stress, MT215 (Mixteco + ZnO) improved germination dynamics by reducing the average germination time by 26% and increasing the germination rate by 34% compared to IMT115. Several treatments, such as MT115, IT315, and IT415, maintained 100% germination even under water stress. Drought Stress | Treatment | Variety | Code | Germinated seeds (n) | Germinability (%) | Mean germinationtime (days) | Mean germination rate (days-1) | Germination speed (%) | Uncertainty Index (bit) |
|---|---|---|---|---|---|---|---|---|---|
| 0% |
T1 Hydropriming |
MT10 | 7.00 ± 0.00bc | 87.50 ± 0.00bc | 3.334 ± 0.08bcde | 0.300 ± 0.007cde | 30.0 ± 0.7cde | 1.43 ± 0.04a | MT10 |
| IT10 | 6.00 ± 0.00d | 75.00 ± 0.00d | 3.444 ± 0.10bcd | 0.291 ± 0.008de | 29.0 ± 0.8de | 0.97 ± 0.05ab | IT10 | ||
| IMT10 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.458 ± 0.07bcd | 0.289 ± 0.006de | 28.9 ± 0.6de | 0.98 ± 0.03ab | IMT10 | ||
| FT10 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.458 ± 0.07bcd | 0.289 ± 0.006de | 28.9 ± 0.6de | 0.98 ± 0.03ab | FT10 | ||
| T2 ZnO + Q |
MT20 | 7.00 ± 0.00bc | 87.50 ± 0.00bc | 3.191 ± 0.08cde | 0.313 ± 0.008bcd | 31.4 ± 0.8bc | 1.40 ± 0.04a | MT20 | |
| IT20 | 7.00 ± 0.00bc | 87.50 ± 0.00bc | 3.334 ± 0.08bcde | 0.300 ± 0.007cde | 30.0 ± 0.7bde | 0.90 ± 0.07ab | IT20 | ||
| IMT20 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.583 ± 0.26bc | 0.280 ± 0.020def | 28.0 ± 2.0def | 1.12 ± 0.25ab | IMT20 | ||
| FT20 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.458 ± 0.26bcd | 0.290 ± 0.021de | 29.0 ± 2.1de | 0.86 ± 0.08b | FT20 | ||
| T3 SiO2 + Q |
MT30 | 7.00 ± 0.00bc | 87.50 ± 0.00bc | 2.905 ± 0.08e | 0.344 ± 0.010b | 34.4 ± 1.0b | 0.78 ± 0.32b | MT30 | |
| IT30 | 7.00 ± 0.00bc | 87.50 ± 0.00bc | 2.334 ± 0.08f | 0.429 ± 0.015a | 42.9 ± 1.5a | 0.90 ± 0.07ab | IT30 | ||
| IMT30 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.292 ± 0.07bcde | 0.304 ± 0.007cde | 30.4 ± 0.7cde | 0.86 ± 0.08b | IMT30 | ||
| FT30 | 7.66 ± 0.57ab | 95.83 ± 7.21ab | 4.137 ± 0.14a | 0.242 ± 0.009g | 24.2 ± 0.8f | 1.42 ± 0.10a | FT30 | ||
| T4 ZnO + SiO2 + Q |
MT40 | 7.66 ± 0.57ab | 95.83 ± 7.21ab | 3.000 ± 0.13de | 0.334 ± 0.014bc | 33.4 ± 1.4bc | 1.00 ± 0.40ab | MT40 | |
| IT40 | 7.00 ± 0.00bc | 87.50 ± 0.00bc | 3.334 ± 0.08bcde | 0.300 ± 0.007cde | 30.0 ± 0.7cde | 0.90 ± 0.07ab | IT40 | ||
| IMT40 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.708 ± 0.26ab | 0.271 ± 0.019efg | 27.1 ± 1.8ef | 1.27 ± 0.23ab | IMT40 | ||
| FT40 | 6.33 ± 0.57cd | 83.33 ± 7.21cd | 4.095 ± 0.17a | 0.244 ± 0.010fg | 24.4 ± 1.0f | 1.42 ± 0.15a | FT40 | ||
| 15% | T1 Hydropriming |
MT115 | 8.00 ± 0.00a | 100.00 ± 0.00a | 2.917 ± 0.07fg | 0.343 ± 0.009a | 34.3 ± 0.8a | 0.72 ± 0.30ab | MT115 |
| IT115 | 7.33 ± 0.57abc | 91.66 ± 7.21abc | 3.696 ± 0.28abc | 0.271 ± 0.021de | 27.2 ± 2.1de | 1.20 ± 0.16ab | IT115 | ||
| IMT115 | 7.00 ± 0.00bc | 87.50 ± 0.00bc | 3.952 ± 0.41a | 0.255 ± 0.025e | 25.5 ± 2.5e | 1.25 ± 0.34ab | IMT115 | ||
| FT115 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.375 ± 0.13 bcdefg | 0.297 ± 0.011bcd | 29.7 ± 1.1bcd | 1.02 ± 0.25ab | FT115 | ||
| T2 ZnO + Q |
MT215 | 7.00 ± 0.00bc | 87.50 ± 0.00bc | 2.905 ± 0.08g | 0.344 ± 0.010a | 34.4 ± 1.0a | 0.78 ± 0.32ab | MT215 | |
| IT215 | 7.66 ± 0.57ab | 95.83 ± 7.21ab | 3.476 ± 0.04abcde | 0.288 ± 0.003cde | 28.8 ± 0.3cde | 1.09 ± 0.18ab | IT215 | ||
| IMT215 | 7.00 ± 0.00bc | 87.50 ± 0.00bc | 3.334 ± 0.08 bcdefg | 0.300 ± 0.007bcd | 30.0 ± 0.7bcd | 0.90 ± 0.07ab | IMT215 | ||
| FT215 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.375 ± 0.08 bcdefg | 0.297 ± 0.007bcd | 29.7 ± 0.7bcd | 0.92 ± 0.07ab | FT215 | ||
| T3 SiO2 + Q |
MT315 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.042 ± 0.07 efg | 0.329 ± 0.008ab | 32.9 ± 0.8ab | 0.54 ± 0.53ab | MT315 | |
| IT315 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.208 ± 0.07cdefg | 0.312 ± 0.007abc | 31.2 ± 0.7abc | 0.72 ± 0.15b | IT315 | ||
| IMT315 | 6.66 ± 0.57cd | 83.33 ± 7.21cd | 3.595 ± 0.11abcd | 0.278 ± 0.009cde | 27.8 ± 0.8cde | 1.36 ± 0.10a | IMT315 | ||
| FT315 | 7.00 ± 0.00bc | 87.50 ± 0.00bc | 3.429 ± 0.14 bcdef | 0.292 ± 0.012bcde | 29.2 ± 1.2bcde | 0.94 ± 0.07ab | FT315 | ||
| T4 ZnO + SiO2 + Q |
MT415 | 6.00 ± 0.00d | 75.00 ± 0.00d | 3.167 ± 0.17defg | 0.316 ± 0.017abc | 31.6 ± 1.7abc | 0.52 ± 0.47fb | MT415 | |
| IT415 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.417 ± 0.07 bcdefg | 0.293 ± 0.006bcde | 29.3 ± 0.6bcde | 0.97 ± 0.03ab | IT415 | ||
| IMT415 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.792 ± 0.14ab | 0.264 ± 0.010de | 26.4 ± 1.0de | 1.46 ± 0.09a | IMT415 | ||
| FT415 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.458 ± 0.14 abcde | 0.289 ± 0.012efg | 28.9 ± 1.2bcde | 1.14 ± 0.24abcde | FT415 | ||
| 30% | T1 Hydropriming |
MT130 | 7.66 ± 0.57ab | 95.83 ± 7.21ab | 3.268 ± 0.15d | 0.307 ± 0.014a | 30.6 ± 1.4a | 1.03 ± 0.25bcd | MT130 |
| IT130 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.375 ± 0.13 cd | 0.297 ± 0.011abc | 29.7 ± 1.1abc | 0.92 ± 0.10d | IT130 | ||
| IMT130 | 8.00 ± 0.00a | 100.00 ± 0.00a | 4.292 ± 0.19ab | 0.233 ± 0.010e | 23.3 ± 1.0e | 1.82 ± 0.23a | IMT130 | ||
| FT130 | 7.00 ± 0.00ab | 87.50 ± 0.00ab | 4.095 ± 0.08abc | 0.244 ± 0.005de | 24.4 ± 0.5de | 1.36 ± 0.20abcd | FT130 | ||
| T2 ZnO + Q |
MT230 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.708 ± 0.07 abcd | 0.270 ± 0.005abcde | 27.0 ± 0.5abcde | 1.44 ± 0.05abcd | MT230 | |
| IT230 | 6.33 ± 1.52b | 79.16 ± 19.04b | 3.992 ± 0.46 abcd | 0.253 ± 0.028cde | 25.3 ± 2.8cde | 1.17 ± 0.18bcd | IT230 | ||
| IMT230 | 8.00 ± 0.00a | 100.00 ± 0.00a | 4.000 ± 0.33abcd | 0.251 ± 0.020cde | 25.1 ± 2.0cde | 1.45 ± 0.14abc | IMT230 | ||
| FT230 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.292 ± 0.07d | 0.304 ± 0.007ab | 30.4 ± 0.7ab | 0.86 ± 0.08d | FT230 | ||
| T3 SiO2 + Q |
MT330 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.625 ± 0.13 bcd | 0.276 ± 0.010abcde | 27.6 ± 1.0abcde | 1.12 ± 0.25bcd | MT330 | |
| IT330 | 7.33 ± 0.57ab | 91.66 ± 7.21ab | 3.690 ± 0.27 abcd | 0.272 ± 0.019abcde | 27.2 ± 1.9abcde | 1.22 ± 0.21bcd | IT330 | ||
| IMT330 | 7.00 ± 0.00ab | 87.50 ± 0.00ab | 3.857 ± 0.38 abcd | 0.261 ± 0.025abcde | 26.1 ± 2.4abcde | 1.56 ± 0.25abc | IMT330 | ||
| FT330 | 7.33 ± 0.57ab | 91.66 ± 7.21ab | 3.958 ± 0.29 abcd | 0.253 ± 0.018bcde | 25.4 ± 1.8bcde | 1.62 ± 0.20ab | FT330 | ||
| T4 ZnO + SiO2 + Q |
MT430 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.417 ± 0.07 cd | 0.293 ± 0.006abcd | 29.3 ± 0.6abcd | 0.97 ± 0.03cd | MT430 | |
| IT430 | 8.00 ± 0.00a | 100.00 ± 0.00a | 3.500 ± 0.25 cd | 0.287 ± 0.021abcd | 28.7 ± 2.1abcd | 1.27 ± 0.23abcd | IT430 | ||
| IMT430 | 6.33 ± 0.57b | 79.16 ± 7.21b | 4.436 ± 0.36a | 0.226 ± 0.017e | 22.6 ± 1.8e | 1.18 ± 0.24bcd | IMT430 | ||
| FT430 | 6.33 ± 0.57b | 79.16 ± 7.21b | 3.690 ± 0.13 abcd | 0.271 ± 0.010abcde | 27.1 ± 1.0abcde | 1.43 ± 0.05abcd | FT430 |
T1: (Hydropriming); T2: (ZnO + Q); T3: (SiO₂ + Q); T4: (ZnO + SiO₂ + Q), and water stress levels (0%, 15%, and 30%). Statistical analysis was performed using the Kruskal-Wallis test, followed by Dunn's multiple comparisons (p ≤ 0.05). Superscript letters indicate significant differences.
Table 2.
Description of the distribution of the varieties of jalapeño pepper used in the study, the treatments, the level of induced stress, and the code used.
Table 2.
Description of the distribution of the varieties of jalapeño pepper used in the study, the treatments, the level of induced stress, and the code used.
| Jalapeño pepper varieties |
Treatment NPs + Chitosan |
Code |
||
|---|---|---|---|---|
| Level of osmotic stress | ||||
| 0% | 15% | 30% | ||
|
Mixteco (M) |
(T1) Triple-distilled water Control | MT10 | MT115 | MT130 |
| (T2) ZnO 100 mgL-1 + Q 100 mgL-1 | MT20 | MT215 | MT230 | |
| (T3) SiO2 10 mgL-1 + Q 100 mgL-1 | MT30 | MT315 | MT330 | |
| (T4) ZnO 100 mgL-1 + SiO2 10 mgL-1 + Q 100 mgL-1 | MT40 | MT415 | MT430 | |
|
Ideal (I) |
(T1) Triple-distilled water Control | IT10 | IT115 | IT130 |
| (T2) ZnO 100 mgL-1 + Q 100 mgL-1 | IT20 | IT215 | IT230 | |
| (T3) SiO2 10 mgL-1 + Q 100 mgL-1 | IT30 | IT315 | IT330 | |
| (T4) ZnO 100 mgL-1 + SiO2 10 mgL-1 + Q 100 mgL-1 | IT40 | IT415 | IT430 | |
|
Imperial (IM) |
(T1) Triple-distilled water Control | IMT10 | IMT115 | IMT130 |
| (T2) ZnO 100 mgL-1 + Q 100 mgL-1 | IMT20 | IMT215 | IMT230 | |
| (T3) SiO2 10 mgL-1 + Q 100 mgL-1 | IMT30 | IMT315 | IMT330 | |
| (T4) ZnO 100 mgL-1 + SiO2 10 mgL-1 + Q 100 mgL-1 | IMT40 | IMT415 | IMT430 | |
|
Forajido (F) |
(T1) Triple-distilled water Control | FT10 | FT115 | FT130 |
| (T2) ZnO 100 mgL-1 + Q 100 mgL-1 | FT20 | FT215 | FT230 | |
| (T3) SiO2 10 mgL-1 + Q 100 mgL-1 | FT30 | FT315 | FT330 | |
| (T4) ZnO 100 mgL-1 + SiO2 10 mgL-1 + Q 100 mgL-1 | FT40 | FT415 | FT430 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



