Submitted:
07 May 2025
Posted:
08 May 2025
You are already at the latest version
Abstract
Arundo donax L. (giant reed) is a perennial rhizomatous grass with high drought and salinity tolerance, making it a promising low-input bioenergy crop. However, the combined effects of irrigation and nitrogen application in salinized soil on physiological adaptations and biomass allocation are still limited. In this study, we conducted a three-factor orthogonal pot experiment with four levels per factor in 2023 and 2024: salinity (S0: non-saline, S1: low salinity, S2: moderate salinity, S3: high salinity); irrigation amount (W0: 605, W1: 770, W2: 935, W3: 1100 mm); and nitrogen application (N0: 0, N1: 60, N2: 120, N3: 180 kg/ha). This resulted in 14 irrigation-nitrogen-salinity combined treatments. The results showed the following: (1) Irrigation, nitrogen and salinity significantly affected leaf dimensions (length, width, area and leaf elongation rate, specific leaf weight), photosynthetic rate, plant height, biomass allocation and dry matter of total plant (p < 0.05). (2) Significant coupling interactions were observed between salinity and irrigation, as well as between nitrogen and irrigation, affecting leaf width, leaf elongation rate,plant height,leaf dry matter and total biomass accumulation; coupling interaction of salinity and nitrogen were found to have effect on leaf width, leaf area, leaf fresh weight, root, stem and leaf dry weight. (3) The S0N2W2 treatment produced the highest dry biomass (44.76 ± 0.68 g/plant), which was 2.2 times higher than the S3N2W2 treatment (20.13 ± 0.50 g/plant). (4) Under moderate salinity conditions (S2), biomass allocation favoured stems (42.70%, mean values of 2023 and 2024) and leaves (39.53%, mean values of 2023 and 2024), whereas under high salinity conditions (S3) biomass allocation shifted towards leaves, followed by stems and roots. A combination of 935 mm irrigation amount and 120 kg/ha nitrogen (N2W2) in S1 and S2 is recommended to optimise biomass production. Our study provides practical irrigation and nitrogen management strategies to enhance A. donax cultivation on marginal saline lands, supporting climate-resilient bio-economy initiatives.
Keywords:
Arundo donax L.
; biomass accumulation and allocation
; saline soil
; water consumption
; nitrogen application
1. Introduction
Arundo donax L. (giant reed, family Poaceae) is a perennial rhizomatous grass that is recognized as a high-yield biomass feedstock. Its biomass can be used for bioenergy, biofuels, paper pulp, wood-based building materials and other biobased products within a biorefinery scheme [1,2]. A. donax follows the C3 pathway for carbon assimilation, but with high photosynthetic capacity similar to C4 species [3,4], demonstrates broad adaptability to different climates, and maintains high yields even on marginal land [5]. The robust growth and high productivity of A. donax make it particularly valuable for biogas and bioenergy production [6]. A. donax is highly resistant to extreme soil and water conditions. It thrives in all types of soils—from loose sand to heavy clay and gravelly substrates—and tolerates high salinity. Although it prefers abundant water, it can withstand extended periods of severe drought [7].
A. donax is of Asian origin that can remain productive for over 15 years, yielding more than 30 t/ha dry matter annually[8,9]. As a perennial species, it reduces the energy costs associated with ploughing and sowing. When properly managed, A. donax can significantly contribute to the bioenergy supply while increasing soil carbon stocks and belowground biomass[10]. In Azul, Argentina, A. donax produced twice as much biomass in the second year when it was watered and fertilized compared to without these treatments [11]. Its extended soil coverage and reduced tillage can improve soil organic matter and decrease soil erodibility [12]. After 7 years of cultivation, A. donax increased organic carbon by 46% in the upper 0–0.2 m soil layer [7]. Under 9 years of nitrogen and phosphate fertilization application, A. donax accumulated 540 kg/ha carbon per year in the upper 0–0.4 m soil layer, maintaining the total nitrogen content of the soil from the start of the study [13].
In addition, A. donax possesses several advantages as an energy crop, reducing irrigation demand and the economic and environmental costs of cultivation. As a perennial species, it eliminates the need for annual tillage, can be cultivated without irrigation and efficiently recycles nitrogen, seasonally translocating it between above- and below-ground organs [14]. Interestingly, several adaptive mechanisms of A. donax have been observed, including an extensive root system, morphological modifications and physiological regulation of stomatal behaviour, which collectively enhance water-use efficiency and sustain high productivity under stress conditions [4,15].
Although A. donax is generally considered a low-maintenance crop with modest fertilizer and water requirements, recent studies indicate that moderate irrigation combined with increased fertilization during early growth, particularly after transplantation, can substantially enhance yield while ensuring both economic viability and environmental sustainability [16]. A. donax has been reported to tolerate water stress at the adult stage[17] and respond positively to reduced irrigation, although peak yield was reached 1 year earlier with increased irrigation [18]. Its energy production efficiency and net energy yield are influenced by fertilization and plant density [19]. Non-optimal nitrogen availability inhibits growth and biomass accumulation, whereas excessive nitrogen application severely suppresses both [20].
Salinity stress is a major threat to global agricultural production, with more than half of the world’s irrigated farmland severely affected by soil salinisation. Rising water stress has also led to increased salt crust formation on agricultural land. In arid and semi-arid regions with high evapotranspiration (ET), water and salinity stress are both the most significant environmental limitations on crop growth and yields. A. donax is classified as moderate salt-tolerant, which can maintain over 50% relative growth at salinity levels below 12 ds/m [21]. It copes with increasing salinity through a sensitive stomatal response, enhanced antioxidant biosynthesis and higher carbohydrate accumulation [22]. Furthermore, its antioxidative defence, mediated by multiple symplastic enzymes, contributes to its high physiological tolerance to alkaline salt stress [23]. The remarkable plasticity, adaptability and productivity of A. donax under challenging conditions make it a promising candidate for sustainable bioenergy production [6].
However, the interaction between irrigation amount and nitrogen fertilization in saline-alkali soils remains poorly understood, despite its importance in enhancing plant adaptability and productivity. Specifically, how the combination of water and nitrogen influences leaf traits, photosynthetic efficiency and biomass allocation in A. donax under different soil salinity levels remains unclear. The objectives of this study were as follows: (1) to evaluate the effects of irrigation amount, nitrogen application and their interaction with different salinity levels on leaf shape and function; (2) to determine how irrigation, nitrogen and salinity influence biomass accumulation and its allocation between above- and below-ground components; (3) to develop optimal irrigation and nitrogen management practices for different soil salinity conditions.
2. Materials and Methods
2.1. Experimental Design and Plant Material
The pot experiments were conducted at a research site of Taiyuan University of Technology in Taiyuan city, Shanxi province, China (37.85°N, 112.52°E) from April 20 to September of 2023 and 2024, which had typical saline loam soil and a temperate continental climate. The region received an average annual precipitation of 400–600 mm, whereas annual pan evaporation ranges from 1000 to 1200 mm. The soil had a field capacity of 33.4% (dry weight) and bulk density of 1.53 g/cm3.
The study examined three factors at four levels, each with three replicates: Soil salinity was classified according to Ayub et al. [24] and included S0 (non-saline, 0.31 g/kg), S1 (low salinity, 4.25 g/kg), S2 (moderate salinity, 8.18 g/kg) and S3 (high salinity, 12.12 g/kg) in 2023, S0 (non-saline, 0.44 g/kg), S1 (low salinity, 4.52 g/kg), S2 (moderate salinity, 8.75 g/kg) and S3 (high salinity, 12.00 g/kg) in 2024. The nitrogen application levels were N0 (0 kg/ha), N1 (60 kg/ha), N2 (120 kg/ha) and N3 (180 kg/ha). Irrigation amount levels were W0 (605 mm, 55% of ET), W1 (770 mm, 70% of ET), W2 (935 mm, 85% of ET) and W3 (1100 mm, 100% of ET), referenced ET values was determined by Federico et al. [25]. The orthogonal test design generated 14 treatment combinations as follows: S0N0W0, S0N2W2, S2N1W1, S1N1W2, S1N2W1, SIN2W2, S2N0W2, S2N1W2, S2N2W0, S2N2W1, S2N2W2, S2N2W3, S2N3W2 and S3N2W2.
To prevent the influence of precipitation, experimental pots (height: 55 cm, diameter: 35 cm) were placed under a canopy. The A. donax plantlets used in the pot experiment were commercially cloned seedlings of uniform height, each with a single main stem and three to five expanded leaves. Soils with different salinity levels were prepared in April and allowed to stabilise until their chemical properties remained stable. Pot soil beds were prepared and five seedlings were transplanted into each pot. Full irrigation was applied for 10 days after transplantation to ensure successful crop establishment. Irrigation treatments began at May 1st, nitrogen fertilization treatment was applied on June 1st and July 1st separately by 40% and 60% of total nitrogen application. Irrigation and nitrogen treatment both continued for 120 days to harvest until September 1st.
2.2. Characteristics of Leaf Morphology
Throughout the irrigation treatment, the newly emerged leaves at the top of the plants were systematically monitored. The leaf length was measured at consistent intervals each day until full expansion, which was defined as the point at which the leaf dimensions stabilised. The leaf elongation rate (LER) was calculated as the final leaf length divided by the number of days in its developmental period. After 120 days of treatment, the morphometric parameters were recorded. Plant height was measured using a flexible tape measure, and the total number of leaves per plant was counted. Three fully expanded leaves were selected for dimensional analysis, with length and width measured using a vernier calliper (precision: ± 0.01 mm). These leaves were then scanned with a calibration scale and digitised for precise leaf area quantification using Image J software (IJ 1.46r, revised edition) with standardised thresholding protocols.
2.3. Characteristics of Leaf Water Content
For each treatment, three fully expanded leaves were sampled to determine the relative water content (RWC, %) using the standardised gravimetric method. RWC values were recorded after 120 days and calculated as (FW − DW) / (TW − DW) × 100, where FW is fresh weight, DW is dry weight after oven-drying at 60°C until constant weight and TW is turgid weight after equilibration in distilled water for 24 h at 4 °C. The water saturation deficit (WSD) was calculated as 100 % − RWC. The specific leaf weight (SLW, g/cm2) was determined as the ratio of dry weight to leaf area.
2.4. Characteristics of Leaf Relative Chlorophyll Content and Photosynthesis
The relative chlorophyll content (SPAD value) was measured using a SPAD-502 chlorophyll metre (Konica Minolta, Japan). After 120 days of irrigation–nitrogen treatment, the second fully expanded leaf from the top of the plant was selected for SPAD measurements. Five readings were taken at different points on each leaf and averaged over three replicate plants per treatment.
Photosynthetic parameters were measured using a portable photosynthesis system (Li-6400, USA). Measurements were taken between 09:00 and 11:00 on clear, cloudless days, 120 days after starting the irrigation–nitrogen treatment. The recorded parameters included net photosynthetic rate (Pn, μmol/(m²·s)), transpiration rate (Tr, mmol/(m²·s)), intercellular CO₂ concentration (Ci, μmol/mol) and stomatal conductance (Gs, mmol/(m²·s)). Measurements were performed on the second fully expanded leaf, avoiding major veins, with three replicate readings per treatment. Leaf transpiration efficiency (TE) was calculated as the ratio of Pn to Tr.
2.5. Characteristics of Biomass Accumulation and Allocation
At the end of the experiment, the fresh and dry biomasses of roots, stems and leaves were recorded separately. Whole A. donax plants were brushed to remove any attached solid particles before separation into leaves, stems and roots. Fresh weight was measured immediately, whereas dry weight was recorded after drying at a constant temperature of 80°C for 24 h until a constant weight was achieved. Total biomass was calculated as the sum of root, stem and leaf biomass. Biomass allocation to roots, stems and leaves was determined based on the proportion of each component relative to total dry biomass.
2.6. Statistical Analysis
All data were recorded using Excel 2016. Statistical analysis were performed using SPSS 25, and figures were generated using Origin 2024. A three-way analysis of variance (ANOVA) was conducted to assess the effects of irrigation amount, nitrogen application and soil salinity on leaf morphology, photosynthetic characteristics, biomass accumulation and allocation. Significance was evaluated at probability levels of 0.05 and 0.01.
3. Results
3.1. Effects of Irrigation–Nitrogen–Salinity Coupling on Leaf Morphology in A. donax
Our results demonstrated that irrigation–nitrogen–salinity coupling treatments had distinct effects on leaf morphology in A. donax in 2023 (Table 1) and 2024 (Table 2). Among the 14 treatments, S0N2W2, S1N2W2, S2N2W2 produced higher leaf length, length–to –width ratio and area, S2N1W2 yielded the largest leaf width and S0N2W2 generated the largest leaf area. In non-saline soil (S0), the leaf length, width and area of the second fully expanded leaf at the top of A. donax plant were significantly greater in N2W2 treatment than those in N0W0 (p < 0.05). In low salinity soil (S1), no significant differences were observed in leaf length, width or area between the N1W2 and N2W1 treatments. However, the aspect ratio varied significantly, with the N2W2 treatment producing longer leaves and a larger leaf area than N1W2 and N2W1 (p < 0.05). The leaf area under N1W2 and N2W1 treatments showed no significant differences. In moderate saline soil (S2), leaf morphology indices in N0W2 and N2W0 did not differ significantly. However, significant differences were observed in leaf width, aspect ratio and leaf area between the N1W2 and N2W1 treatments, with N1W2 producing greater values (p < 0.05). No significant differences were found between N2W3 and N3W2, but leaf length, width and area varied significantly between N1W1 and N3W2 (p > 0.05). Our findings indicate that overall leaf morphology did not differ. However, leaf length, aspect ratio and area were significantly higher in the N2W2 treatment than in N1W1, whereas leaf length and aspect ratio were significantly lower in the N2W3 and N3W2 treatments than in N2W2. Leaf length, width, area and aspect ratio were significantly reduced in S3N2W2 compared with S2N2W2 (p < 0.05). At the S0, S1 and S2 salinity levels, the combination of nitrogen application and irrigation amount influenced leaf morphology.
The three-factor, four-level interaction analysis (Table 3) demonstrated that the combined effects of soil salinity, nitrogen application and irrigation amount significantly influenced leaf length, width and area (p < 0.05), but had no significant effect on leaf aspect ratio in 2024 (p > 0.05). The combined effect of soil salinity and irrigation amount, salinity and nitrogen application, nitrogenapplication and irrigation amount had significant effects on leaf width and length to width ration(p < 0.05), showed significant effect on leaf length in 2023. The influence of leaf length and area was determined as Salinity > Irrigation > Nitrogen. The interaction between soil salinity and nitrogen application also significantly affected leaf morphology (p < 0.05).
3.2. Effects of Irrigation–Nitrogen–Salinity Coupling on the Photosynthetic Characteristics of A. donax
3.2.1. SPAD Values
The effects of irrigation-nitrogen-salinity treatments on the SPAD values of A. donax leaves are depicted in Figure 1 (A (2023) and B (2024)). The highest SPAD value was recorded under S1N2W2 (2023)and S2N2W2 (2024), whereas the lowest was observed under S2N2W0 (2023) and S0N0W0 (2024). In the non-saline soil (S0), the SPAD value of the second fully expanded leaf at the top of A. donax was slightly higher under N2W2 treatment than under N0W0 treatment. However, this difference was not statistically significant (p > 0.05). In low salinity soil (S1), the highest SPAD value was observed in N2W2 treatment, but the difference from other treatments was not significant (p > 0.05). In moderate saline soil (S2), the SPAD value was significantly higher in the N2W2 treatment than in the N2W0 treatment in 2023 (p < 0.05), whereas no significant differences were found between the N0W2 and N2W0, N1W2 and N2W1 or N2W3 and N3W2 (p > 0.05). Additionally, SPAD values did not differ significantly between the S3N2W2 and S2N2W2 treatments. In non-saline soils, increased irrigation amount and nitrogen application resulted in significant increase in SPAD values. In low and moderate salinity soils, increasing water and nitrogen levels also resulted in higher SPAD values, although these differences were not significant statistically (p > 0.05).
The three-factor, four-level interaction significance analysis (Figure 1 A (2023) and B (2024)) revealed that SPAD values were significantly influenced by salinity (p < 0.05). The individual and combined effects of nitrogen application, irrigation amount and soil salinity (such as Salinity × Nitrogen, Salinity × Irrigation and Nitrogen × Irrigation) were not statistically significant in 2023 (Figure 1 A) (p > 0.05), whereas the combined of nitrogen and salinity, nitrogen and irrigation showed significant effection in 2024 (Figure 1 B) (p < 0.05). The relative impact of the three factors on SPAD values followed the order Salinity > Irrigation > Nitrogen.
3.2.2. Photosynthetic Characteristics
The effects of irrigation-nitrogen-salinity coupling on the photosynthetic parameters of A. donax leaves are presented in Table 4 (2023) and Table 5 (2024). Among the 14 treatments, S1N2W2 resulted in the highest Pn and Tr values, although with no significant differences observed compared with S0N2W2 (p > 0.05). However, Tr values were significantly higher in S1N2W2 than in S2N2W2 (p < 0.05), whereas Pn, Tr and Ci were significantly greater than in S3N2W2 in 2023 (p < 0.05). In the non-saline soil (S0), the N2W2 treatment significantly increased Pn and Tr compared with the N0W0 treatment in 2023 (p < 0.05), whereas Pn, Tr, Gs and Ci values were no significant difference beteen N0W0 and N2W2 in 2024. In low salinity soil (S1), there were no significant differences in photosynthetic parameters between N1W2 and N2W1 treatments in 2023 and 2024, the N2W2 treatment yielded the highest Pn, Tr and Gs values, with only Tr exhibiting significant differences compared with N2W1 in 2023 (p < 0.05), but Tr, Gs and Ci showed significant difference compared with N2W1 in 2024 (p < 0.05). In moderate saline soil (S2), no significant differences were observed between N0W2 and N2W0 or between N1W2 and N2W1 in 2023 and 2024, compared with N1W1, N1W2 and N2W1 improved Pn, Tr and Gs, but these differences were not statistically significant (p > 0.05). Although N2W1 exhibited non-significant increases in Pn, Tr and Gs over N2W0, N2W2 significantly improved Pn (p < 0.05) and reduced Ci. N2W3 treatment resulted in non-significant improvements in photosynthetic parameters. Notably, S3N2W2 exhibited lower Pn, Tr and Gs values than S2N2W2, although these differences were not statistically significant (p > 0.05).
The three-factor, four-level interaction significance analysis of A. donax photosynthetic parameters (Table 6) revealed that soil salinity had a significant effect on Pn, Tr and Ci (p < 0.05). Conversely, nitrogen application and irrigation amount significantly affected only Pn. Furthermore, the interactions between Salinity × Nitrogen, Salinity × Irrigation and Nitrogen × Irrigation did not have significant effect on any of the four photosynthetic parameters in 2023 (p > 0.05).
TE was assessed at the leaf level as the ratio of Pn to Gs in fully expanded leaves. The effects of irrigation-nitrogen-salinity coupling treatments on TE are shown in Figure 2 (A (2023) and B (2024)). Among the 14 treatments, the highest TE value was recorded under S1N2W1 in 2023 and S2N0W2 in 2024, whereas the lowest were under S2N2W0 in 2023 and S2N3W2 in 2024. Significant differences in TE were observed only between S1N2W2 and S2N0W2, as well as between S0N2W2 and S2N2W0 in 2023, between S2N0W2 and S0N0W0, between S2N0W2 and S2N1W1, as well as between S2N0W2 and S2N3W2 in 2024. In non-saline soil (S0), TE was higher in N2W2 treatment than in N0W0 treatment. In low salinity soil (S1), there were no significant difference among 3 treatments both in 2023 and 2024. In moderate saline soil (S2), the N2W2 treatment resulted in the highest TE value in 2023.
The three-factor, four-level interaction significance analysis (Figure 2) indicated that individual factor, such as salinity and irrigation did not have significant effect on TE both in 2023 and 2024, among three factors, such as salinity and nitrogen, salinity and irrigation, nitrogen and irrigation, all of them did not have significant coupling effect on TE either in 2023 and 2024 (p > 0.05).
3.3. Effects of Irrigation–Nitrogen–Salinity Coupling on Water-Related Physiological Parameters of Fully Expanded A. donax leaves
The effects of irrigation-nitrogen-salinity coupling treatments on the moisture content characteristics of fully expanded A. donax leaves were presented in 2023 (Table 7) and 2024 (Table 8). Compared with S0N2W2, S1N2W2 had no significant effect on LER, leaf dry weight (LDW), SLW and RWC (p > 0.05) both in 2023 and 2024. Similarly, S2N2W2 showed no significant effect on any parameters compared with S1N2W2 (p > 0.05). Compared with S2N2W2, S3N2W2 resulted in an increase in leaf fresh weight (LFW)—although this difference was not significant—a significant increase in LDW, significant decreases in LER and SLW and a reduction in RWC that was not statistically significant in 2023 (p > 0.05). In 2024, there were no significant difference among S0N2W2, S1N2W2 and S2N2W2 on LER, LFW and LDW(p > 0.05), the LER of S2N2W2 was significant higher than S3N2W2.
In Table 7, under non-saline conditions (S0), N2W2 increased LER, LFW, LDS, SLW and RWC compared with N0W0, with significant effects on LER, LFW and SLW (p < 0.05). In low-salinity soil (S1), N1W2 and N2W1 treatments significantly affected only LER and LDW (p < 0.05), whereas LFW, SLW and RWC remained unaffected (p > 0.05). Compared with N2W1, N2W2 treatment significantly reduced LFW and LDW while significantly increasing SLW but had no significant effects on LER or RWC. Under moderate salinity (S2), N1W2 significantly increased LER, LFW, LDW and SLW compared with N0W2, whereas RWC also increased but not significantly (p > 0.05). Compared with N1W1, N1W2 significantly improved LER but had no significant effects on LFW, LDW, SLW or RWC (p > 0.05). Compared with N2W0, N2W1 significantly decreased LDW and increased RWC (p > 0.05), but with no significant effects on LER, LFW or SLW. N2W2 significantly reduced LFW and LDW while significantly increasing LER, SLW and RWC compared with N2W0 (p < 0.05). Compared weith N2W0, N2W3 treatment significantly increased LER, LFW and LDW, whereas SLW and RWC showed non-significant increases (p < 0.05). Compared with N2W2, N3W2 significantly decreased LER while increasing LFW and SLW significantly (p < 0.05). Compared with S2N2W2, S3N2W2 showed significantly lower values of LER and SLW (p < 0.05).
In Table 8, under non-saline conditions (S0), N2W2 treatment significantly increased the values of LER and SLW compared with N0W0. In low salinity soil (S1), compared with N1W1, N2W2 treatment did not significantly increased the values of LER, whereas incresed the values of SLW significantly. Under moderate salinity (S2), there were no significant difference in LER, LFW, LDW, SLW and RWC between N0W2 and N1W2, there were no significant difference in LER, LFW, LDW, SLW and RWC between N1W1 and N2W1. Compared with S2N2W2, S3N2W2 showed significantly lower values of LER (p < 0.05).
The three-factor, four-level interaction significance analysis (Table 9) of water-related physiological parameters showed that salinity had significant effects on LER and SLW (p < 0.05), nitrogen and irriagtion had significant effects on LER, LFW, LDW and SLW individually in 2023 and 2024. Coupling effects, such as salinity and nitrogen, had significant effects on LFW, whereas no significant effects on LER and RWC, salinity and nitrogen, as well as salinity and irrigation had significant effects on LFW, nitrogen and irrigation also had no coupling effect on RWC.
3.4. Effects of Irrigation-Nitrogen-Salinity Coupling on Biomass Accumulation in A. donax
After 120 days of irrigation-nitrogen-salinity coupling treatments, plant height, leaf number and biomass accumulation in the roots, stems and leaves of A. donax were recorded in 2023 (Table 10) and 2024 (Table 11). Among the 14 treatments, the S0N2W2 treatment resulted in the greatest plant height, leaf number and stem biomass in 2023 and 2024. The optimal water–nitrogen combination for achieving the greatest plant height in both low- and moderate-salinity soils was identified as N2W2 in 2023 and 2024.
In Table 10, under the non-saline soil (S0), N2W2 treatment significantly increased plant height, leaf number, root, stem and leaf dry weight compared with N0W0 (p < 0.05). In low salinity soil (S1), N2W2 treatment significantly enhanced plant height, as well as root, stem and leaf biomass, compared with N1W2 (p < 0.05). Compared with N2W1, N2W2 treatment significantly increased plant height, leaf number, root, stem and leaf dry weight (p < 0.05). In moderate saline soil (S2), no significant differences were observed in plant height or leaf biomass accumulation between the N0W2 and N2W0 treatments. However, leaf number, root biomass and stem biomass showed significant differences. N1W2 and N2W1 treatments significantly increased plant height, stem biomass and leaf biomass compared with N1W1 (p < 0.05). The N2W1 treatment significantly improved plant height, leaf number, root, stem and leaf dry weight compared with N2W0 (p < 0.05), while the N2W2 treatment significantly enhanced plant height and total biomass accumulation compared with N2W1 (p < 0.05). Compared with N2W2, the N2W3 and N3W2 treatments significantly reduced plant height and total biomass (p < 0.05). Similarly, compared with S2N2W2, the S3N2W2 treatment resulted in significant reductions in plant height, leaf number and total biomass accumulation (p < 0.05).
In Table 11, under the non-saline soil (S0), N2W2 treatment significantly increased the plant height, leaf number, root, stem and leaf dry weight compared with N0W0 treatment (p < 0.05). In low salinity soil (S1), there were significant differences among the N1W2, N2W1 and N2W2 treatments in plant height, root and leaf dry weight (p < 0.05). Under moderate saline soil (S2), there were no significant differences in height, leaf number and stem dry weight between N0W2 and N2W0 treatments, N1W2 treatment showed significantly higher plant height and leaf dry weight values than N0W2(p < 0.05), and showed significantly lower plant height and leaf dry weight than N2W2 (p < 0.05). Compared with N2W2, the N2W3 and N3W2 treatment significantly changed the plant height, root, stem and leaf dry weight but with opposite direction (p < 0.05). Similarly, compared with S2N2W2, the S3N2W2 treatment resulted in significant reductions in plant height, leaf number, root and stem dry weight (p < 0.05) showing the same trend as 2023.
The three-factor, four-level interaction significance analysis (Table 12) showed that soil salinity, nitrogen application and irrigation amount as individual factor significantly influenced plant height and biomass accumulation in the roots, stems and leaves of A. donax (p < 0.05). The relative influence on plant height followed the order Salinity > Nitrogen > Irrigation. Regarding coupling interaction, Salinity × Nitrogen significantly influenced biomass accumulation in roots, stems and leaves (p < 0.05). Salinity × Irrigation significantly impacted plant height, leaf number, root biomass and leaf biomass accumulation, while Nitrogen × Irrigation significantly influenced plant height, stem biomass and leaf biomass accumulation (p < 0.05).
3.5. Effects of Irrigation-Nitrogen-Salinity Coupling on Biomass Accumulation and Allocation in A. donax
The effects of irrigation-nitrogen-salinity coupling treatment on biomass accumulation in A. donax were shown in Figure 3 (A (2023) and B (2024)). The biomass accumulation varied considerably across 14 treatments. The highest biomass was recorded in the S0N2W2 treatment, with a mean of 45.16 g/plant (2023) and 44.37 g/plant (2024), whereas the lowest biomass was observed in the S3N2W2 treatment in 2023, at 19.68 g/plant. There were significant difference between S0N0W0 and S0N2W2 in 2023 and 2024 (p < 0.05). In lower salinity soil (S1), all of nitrogen and irrigation coupling treatment could have significant effect on the biomass accumulation in 2023 and 2024 (p < 0.05). Under the moderate salinity soil (S2), no significant differences were found between N1W2 and N2W3, N2W1 and N3W2, or N0W2 and N1W1 in 2023 (p > 0.05), significant differences were found between N0W2 and N2W2, N1W1 and N2W1, N1W1 and N1W2, N2W0 and N2W1, N2W1 and N2W2, N2W2 and N2W3, N2W3 and N3W2 in 2024 (p < 0.05). There were significant reduction of S3N2W2 compared with S2N2W2 in 2023 and 2024 (p < 0.05). In non-saline and low saline soils, higher nitrogen and water inputs significantly (p < 0.05) promoted biomass accumulation.
The three-factor, four-level interaction analysis of biomass accumulation (Figure 3) indicated that salinity, nitrogen and irrigation as single factors, as well as the Salinity × Irrigation and Nitrogen × Irrigation interactions, had significant effects on biomass accumulation in A. donax (p < 0.05).
The dry matter allocation to the roots, stems and leaves of A. donax was determined using the ratio of root (AR), stem (AS) and leaf (AL) dry weight to the total dry weight of the plant. Changes in dry matter allocation under different nitrogen application rates (a), irrigation amount levels (b) and soil salinity levels (c) were shown in Figure 4 (A (2023) and B (2024)).
In moderate saline soil (S2), when irrigation amount was fixed at W2 and nitrogen application increased from N0 to N2 (Figure 4 A (a) and B (a)), root biomass allocation gradually decreased from 23.77% to 12.09% in 2023, aboveground biomass allocation increased and leaves accounted for a greater proportion than stems. Under moderate saline soil (S2), when nitrogen application was fixed at N2 and irrigation amount increased from W0 to W2 (Figure 4 A (b) and B (b)), root biomass allocation again decreased, aboveground biomass allocation increased from 82.59% to 87.90% in 2023, leaf biomass was more affected by irrigation amount than stem biomass. Under N2W2 treatment, as soil salinity increased from S0 to S1 (Figure 4 A (c) and B (c)), root biomass allocation was highest (24.86% in 2023 and 25.14% in 2024) in low-saline soil, whereas aboveground biomass allocation was lowest (75.14% in 2023 and 74.86% in 2024). In moderate saline soil, aboveground biomass allocation reached its maximum, with stems and leaves accounting for 30.37% and 40.95% in 2023 respectively. In highly saline soils, biomass allocation followed the order leaves > stems > roots. There were the same biomass allocation trend of root, stem and leaf under the nitrogen, irrigation and salinity changes in 2024.
3.6. Regression and Path Analysis of Irrigation-Nitrogen-Salinity Coupling Effects on Biomass Accumulation in A. donax
Soil salinity, irrigation amount and nitrogen application rate had complex interactive effects on leaf morphology, photosynthetic parameters and overall growth of A. donax. To assess their impact on biomass accumulation, regression and path analysis were conducted using data of 2023 and 2024. Standardised values of soil salinity, nitrogen application and irrigation amount were used in the regression analysis along with the dry biomass weight per plant. The regression equation was as follows:
Y = 18.464 − 1.660X1 + 0.061X2 + 0.003X3 (R = 0.831, R2 = 0.691, p < 0.05)
Here, Y represents the dry biomass weight per plant (including above- and belowground components), X1 represents soil salinity, X2 represents nitrogen application rate and X3 represents irrigation amount. The significance test showed that the partial regression coefficients of X₁, X₂ and X₃ were significant at p < 0.05, confirming a significant relationship between the independent variables and biomass accumulation.
An F-test of the regression equation indicated that at least one of the three factors—irrigation, nitrogen or salinity—had a linear relationship with biomass accumulation. The coefficient of determination (R²) was 0.691, and the residual factor (e) was 0.556, suggesting that factors other than soil salinity, nitrogen and irrigation amount influenced biomass accumulation. The Durbin–Watson value of 0.429, within the acceptable range of 0–4, confirmed that the residuals were independent. Collinearity diagnostics showed a tolerance of 0.309 and a VIF of 3.236, indicating no collinearity among the three variables. Further research is required to identify other significant factors affecting biomass accumulation in saline soils.
The relationships between the simple correlation coefficients, path coefficients and indirect path coefficients of irrigation, nitrogen and salinity on biomass accumulation are shown in Table 13. The direct effects of the independent variables (X₁, X₂ and X₃) on the dependent variable (Y) are as follows: P₁y = −0.707, P₂y = 0.391 and P₃y = 0.511. The indirect effects of soil salinity along with nitrogen application and irrigation amount on biomass accumulation were relatively large, with indirect path coefficients of 0.230. The indirect effects of nitrogen application along with soil salinity and irrigation amount were negative, with indirect path coefficients of -0.048. Similarly, the indirect effects of irrigation amount along with soil salinity and nitrogen application were similar, with indirect path coefficients of -0.079. The simple correlation coefficient of soil salinity on biomass accumulation was −0.477, indicating that soil salinity had a significant negative impact on biomass accumulation. Irrigation amount and nitrogen application positively impacted biomass accumulation, and the influence of irrigation amount was larger than that of nitrogen application under salinity stress.
4. Discussion
4.1. Leaf Plasticity of A. donax Under Irrigation-Nitrogen-Salinity Coupling
This study demonstrated the complex interactions among irrigation, nitrogen application and soil salinity on leaf morphology and function, highlighting the adaptive advantages of A. donax as a salt-tolerant bioenergy crop. In non-saline soil (S0), moderate irrigation and nitrogen application (N2W2) significantly increased leaf length, width and area (p < 0.05), consistent with the efficient nitrogen use mechanism proposed by Ceotto et al.[14]. Under saline conditions (S1 and S2), the effects of irrigation–nitrogen coupling varied. In low-salinity soil (S1), N2W2 maintained a larger leaf area, but the leaf morphology under N1W2 and N2W1 did not differ significantly. This suggested that salinity may weaken the dominant role of nitrogen, whereas water availability and nitrogen fertilization remained as key factors influencing leaf development, consistent with the findings of Cosentino et al. [26]. In moderate saline soil (S2), N1W2 resulted in superior leaf morphology compared with the other treatments, indicating that an appropriate irrigation–nitrogen ratio (e.g., N1W2) could mitigate salt stress effects on leaf growth. This finding is consistent with the conclusions of Romero-Munar et al. [17], who reported that A. donax exhibits leaf plasticity, stomatal regulation and osmotic adjustment to cope with stress. However, excessive nitrogen application (e.g., N3W2) led to deteriorated leaf morphology, reinforcing the concept of ‘non-optimal nitrogen inhibiting growth’ proposed by Zhang et al. [20]. This finding highlights the need for precise fertilizer management to optimise A. donax growth in saline environments.
The three-way analysis of variance further clarified the combined effects of environmental variables. Soil salinity had the greatest impact on leaf morphology and photosynthetic parameters, including length, width, area, SPAD, Pn, Tr and Ci (Salinity > Irrigation > Nitrogen), confirming that salinity was the primary limiting factor for leaf expansion in A. donax (Table 2. Morphological parameters of fully expanded leaves of A. donax under each treatment in 2024.
| Treatment | Leaf length (cm) | Leaf width (cm) | Leaf length–to –width ratio | Leaf area(cm2) |
| S0N0W0 | 15.24±1.41 de | 0.61±0.02 f | 25.02±2.76 ab | 6.13±0.52 f |
| S0N2W2 | 25.73±1.77 a | 1.04±0.07 b | 24.80±3.23 ab | 18.74±0.60 a |
| S1N1W2 | 20.55±2.94 bc | 0.87±0.06 cde | 23.94±4.90 b | 12.39±1.04 cd |
| S1N2W1 | 19.65±1.64 bcd | 0.94±0.04 bc | 20.81±1.09 b | 13.00±1.55 cd |
| S1N2W2 | 22.42±1.99 ab | 1.07±0.01 b | 21.00±1.63 b | 16.75±1.68 ab |
| S2N0W2 | 18.31±1.81 cde | 0.83±0.04 cde | 22.07±3.22 b | 10.56±0.80 de |
| S2N1W1 | 16.73±0.36 cde | 0.83±0.05 cde | 20.13±1.34 bc | 9.48±0.47 def |
| S2N1W2 | 17.14±3.27 cde | 1.28±0.19 a | 13.45±2.22 c | 15.49±4.64 bc |
| S2N2W0 | 14.68±0.71 e | 0.73±0.03 def | 20.11±0.63 bc | 7.29±0.58 ef |
| S2N2W1 | 16.88±1.31 cde | 0.91±0.04 bcd | 18.54±2.12 bc | 10.68±0.60 de |
| S2N2W2 | 23.01±1.87 ab | 0.77±0.07 cde | 30.11±4.33 a | 12.37±0.86 cd |
| S2N2W3 | 17.59±1.73 cde | 0.95±0.08 bc | 18.61±3.17 bc | 11.70±1.10 d |
| S2N3W2 | 17.16±1.39 cde | 0.81±0.08 cde | 21.26±1.95 b | 9.59±1.68 def |
| S3N2W2 | 13.58±0.32 e | 0.70±0.07 ef | 19.53±2.07 bc | 6.34±0.67 f |
Table 3). Although the independent effects of irrigation amount and nitrogen were relatively weak, their interactions with salinity were significant (p < 0.05), indicating that water management could partially mitigate the negative effects of salt stress. For instance, under S2 conditions, the leaf area in the N2W2 treatment was significantly larger than in N2W0 (Table 1 and Table 2). This finding supports Romero-Munar et al.[17], who found that leaf length and elongation rate in A. donax were influenced by soil water content. The leaf aspect ratio was not significantly affected by the three-factor interaction (p > 0.05), suggesting that A. donax maintains leaf shape stability to adapt to environmental fluctuations, a trait linked to its physiological stress tolerance mechanisms, such as stomatal regulation[4]. Similarly, Pompeiano et al.[27] reported that A. donax exhibits an integrated response mechanism and environmental plasticity under controlled drought conditions. Overall, A. donax sustains leaf functionality in saline soils by dynamically adjusting water and nitrogen use efficiency. However, its production potential requires further optimisation through phased and precise resource management strategies.
The findings on nitrogen, water and salinity coupling conditions highlighted the complex relationship between leaf structural adaptation and photosynthetic function in A. donax. The highest SPAD values, reflecting chlorophyll content and photosynthetic potential, were observed under S1N2W2 treatment, suggesting that mild salinity combined with optimal water and nitrogen inputs enhances chlorophyll synthesis and leaf structural integrity. This aligns with Sánchez et al.[4], who reported that A. donax employs morphological and physiological adaptations, such as stomatal regulation and nitrogen translocation to optimise photosynthetic efficiency under stress. However, the absence of significant differences in SPAD values among most treatments (p > 0.05) suggests that A. donax maintains relatively stable chlorophyll content across varying environmental conditions, likely due to its extensive root system and efficient resource allocation mechanisms [14]. The dominant influence of soil salinity on SPAD values (Salinity > Irrigation > Nitrogen) further highlights the critical role of salinity in shaping leaf structural properties, which in turn determine photosynthetic capacity.
The photosynthetic parameters (Pn, Tr, Gs and Ci) further illustrate the functional consequences of structural adaptation. The S1N2W2 treatment, which produced the highest Pn, Tr and Gs values, shows that mild salinity combined with sufficient water and nitrogen can enhance Gs and Tr, thus maximising carbon assimilation. This finding supports by Romero-Munar et al.[17], who found that A. donax optimises stomatal behaviour to balance water loss and CO₂ uptake under stress. The significant decline in Pn and Tr under severe salinity (S3N2W2) compared to moderate salinity (S2N2W2) highlights the threshold beyond which structural damage impairs photosynthetic function. The absence of significant interactions among nitrogen, irrigation and salinity (p > 0.05) suggests that A. donax relies on integrated structural adaptations rather than isolated responses to individual stressors. Furthermore, the TE results, with the highest values observed under S1N2W1 treatment reinforce the idea that optimal water and nitrogen management can improve water-use efficiency without compromising photosynthetic performance—a key adaptation for survival in resource-limited environments [18]. These findings highlight how the structural resilience of A. donax leaves underpins its functional plasticity, enabling it to maintain photosynthetic efficiency under diverse environmental conditions.
4.2. Water Physiology of A. donax Under Irrigation-Nitrogen-Salinity Coupling
Our results on water physiology of expanded leaves in A. donax demonstrated that LER, LFW and LDW were dynamically influenced by interactions between salinity, nitrogen and irrigation. Under non-saline conditions (S0), N2W2 significantly increased LER and LFW, indicating that optimal nitrogen and irrigation supply synergistically promoted leaf expansion and biomass accumulation. However, under salinity stress, these responses became more complex. For example, at moderate salinity (S2), N1W2 significantly increased LER, LFW and LDW compared to N0W2, suggesting that moderate nitrogen and adequate water consumpation (W2) could alleviate salinity-induced growth limitations. In contrast, higher salinity (S3) combined with N2W2 reduced LER but increased SLW, suggesting a shift towards leaf thickening under stress. The N2W3 treatment under S2 further increased LER, LFW and LDW, highlighting the importance of increased water input (W3) in sustaining growth under salinity. Notably, the significant Nitrogen×Irrigation interaction highlighted that water availability modulates nitrogen efficacy: while N2W1 under S1 improved LER and LDW, N2W2 reduced LFW and LDW, likely due to altered resource partitioning under different water consumption. These results highlighted the role of nitrogen and water management in balancing leaf expansion, biomass allocation and stress adaptation in saline environments.
In this study, the RWC values varied from 94.84 % to 98.33 % under all irrigation-nitrogen-salinity coupling treatments. These findings indicated no significant plasticity in the RWC of A. donax leaves, which would provide insights into its adaptive mechanisms under stress. Stable and high RWC performance was consistent with Claudia et al., they detected that A. donax presented the highest RWC with the value of 85-90 % under watered or non-watered conditions [15]. Müller et al.[23] similarly reported no significant differences in RWC between 0 mM and 80 mM NaHCO3 and Na2CO3 salinity stress, with A. donax exhibiting higher RWC and transpiration intensity. The stability of these parameters suggested that salinity did not affect stomatal movement. However, A. donax responded to elevated salinity with increased WSD, particularly under extreme salt stress (S3). This was consistent with Pompeiano et al.[16], who found that A. donax experienced physiological adjustments such as osmotic regulation and cellular water retention to mitigate salinity stress, and water availability could enhance biomass strongly. The significant reduction in RWC under severe salinity (S3N2W2) compared with moderate salinity (S2N2W2) suggested that while A. donax could maintain water balance under mild to moderate stress, extreme salinity disruptd its water retention capacity, leading to higher WSD. Additionally, the significant effects of nitrogen and irrigation interactions on LFW, LDW, and SLW highlighted the importance of balanced resource management in maintaining leaf structural integrity and water status. These results suggested that although A. donax exhibited a high degree of plasticity in leaf water content, enabling adaptation to diverse environmental conditions, its resilience was limited under extreme salinity.
4.3. Biomass Accumulation and Allocation of A. donax Under Irrigation-Nitrogen-Salinity Coupling
Our results demonstrated that A. donax employed dynamic biomass accumulation and allocation strategies under irrigation-nitrogen-salinity coupling conditions, reflecting its adaptability to environmental stress. The highest biomass accumulation observed in the S0N2W2 treatment in 2023 (45.16 g/plant) highlighted the synergistic benefits of optimal water and nitrogen inputs in non-saline soils, where resource availability aligned with the growth potential of plant [28]. However, even under mild or moderate salinity (S1 and S2), N2W2 treatment consistently maximised plant height, leaf number and total biomass, indicating that A. donax prioritised resource acquisition and photosynthetic efficiency under stress. This supported the findings of Sánchez et al.[4], who reported that A. donax optimised nitrogen translocation and stomatal regulation to sustain growth under suboptimal conditions. The significant biomass reduction under severe salinity (S3N2W2, 19.68 g/plant) highlighted a critical threshold beyond which salt toxicity irreversibly impaired physiological processes, aligning with Ullah et al.[29], who emphasised the coordination between structural and physiological traits in leaves as key to their response to environmental variation. This coordination enabled plant to adjust leaf structure and physiological functions to optimise photosynthetic efficiency and biomass allocation patterns [6,30]. The dominant influence of soil salinity on biomass accumulation underscored salinity as the primary limiting factor. At the same time, interactions such as Salinity × Irrigation and Nitrogen × Irrigation suggested that water and nitrogen management could partially mitigate salt-induced growth suppression. In moderate saline soils (S2), increasing nitrogen (from N0 to N2) or irrigation (from W0 to W2) significantly enhanced aboveground biomass allocation, demonstrating the morphological plasticity of A. donax in response to habitat heterogeneity [31]. Adequate water and nitrogen inputs were essential for high biomass yields in mild and moderate saline soils [12].
Plants compete for nutrients, water and light, which are often scarce or unevenly distributed. Biomass allocation provides insights into how species respond to environmental limitations [32]. The biomass allocation patterns of A. donax under irrigation-nitrogen-salinity treatments further illustrated its adaptive plasticity performance. Under low salinity (S1), the highest root biomass allocation (24.86%) in N2W2 treatment reflected a ‘root-foraging’ strategy to enhance water and nutrient uptake. In moderate salinity (S2), the shift towards aboveground dominance (75.14% of total biomass) indicated greater investment in leaves and stems to maximise light capture and carbon assimilation (Figure 4). In high saline soils (S3), preferential allocation to leaves ( > 40%) over stems and roots suggested a strategy to maintain photosynthetic capacity and offset salt-induced metabolic costs despite reduced total biomass. The decline in root allocation with increasing nitrogen or irrigation in moderate saline soils (S2) suggested that greater resource availability reduced the need for extensive root systems, allowing more investment in aboveground growth. These findings aligned with that of Zhang et al.[20], who reported that non-optimal nitrogen disrupted biomass partitioning, highlighting the importance of balanced fertilization. The significant interaction effects of Salinity × Nitrogen and Nitrogen × Irrigation on stem and leaf biomass further emphasised the need for tailored water–nitrogen management to optimise resource use efficiency. Collectively, A. donax employed a dual strategy: under low stress, it maximises total biomass through synergistic resource inputs, whereas under salinity, it reallocates biomass to sustain functional organs, ensuring resilience at the cost of reduced productivity—a trade-off essential for its ecological success in marginal environments [18]. In this study, the aboveground biomass allocation percentage (including stem and leaf biomass) of A. donax ranged from 75.14% to 87.91%, exceeding the 20%–60% reported by Di Nasso et al.[33]. The variation in dry matter percentage suggests differences in composition and specific fibre types, which can be used in different conversion technologies (e.g., thermochemical processes) for downstream applications. It is necessary to investigate the fibre pattern of A. donax under irrigation-nitrogen-salinity coupling conditions, as Ceotto et al.[14] reported that biomass composition is linked to growth duration and harvest time, with the fraction of leaves in total biomass decreasing progressively.
In the 3.6 section, we conducted regression analysis and revealed significant but complex interactions among soil salinity, nitrogen application, and irrigation amount in influencing biomass accumulation in A. donax. Soil salinity (X₁) exhibited a substantial negative direct effect (P₁y = −0.707), underscoring its detrimental impact on growth. Conversely, irrigation (X₃) and nitrogen (X₂) demonstrated positive direct effects (P₃y = 0.511, P₂y = 0.391), with irrigation exerting a stronger influence under different salinity conditions (Table 12). The high coefficient of determination (R² = 0.691) confirmed a robust linear relationship among these factors, though 30.9% of variation in biomass remained unexplained, suggesting additional unaccounted variables. Acturally, A. donax depends on its rhizome, acts alternatively as a source and sink of nutrient, furtherly optimizes nutrient recycing between under-ground and above-ground [16]. Our results clearly demostrate that irrigation and nitrogen have a significant impact on the leaf photosynthesis and yield during the experiment period. However, as a modified ARMIDA model of A. donax reported, the base temperature, leaf area index and radiation intercepted during the growth period were also the main factors conductive to final yields [11]. Moreover, the negative indirect effects of nitrogen and irrigation, coupled with salinity-mediated pathways, highlight potential trade-offs or antagonistic interactions in saline conditions. These findings emphasize the need for integrated management strategies in saline soils, considering the nuanced interplay among these factors. Further research should explore additional contributors, such as soil properties or microbial activity, to enhance biomass productivity.
5. Conclusions
In this study, we examined the adaptive strategies and resource utilisation efficiency of A. donax under salt-stressed conditions using a coupled irrigation-nitrogen experiment.
In low -to - moderate saline soils, moderate water-to-nitrogen ratios maintained leaf morphological stability and photosynthetic efficiency by optimising stomatal behaviour and nitrogen transport. Conversely, severe salinity significantly reduced leaf area and Pn, indicating a physiological damage threshold for salt stress. Soil salinity was the primary factor limiting growth and biomass accumulation in A. donax, indirectly affecting aboveground resource distribution by directly inhibiting root development and photosynthetic function. Nitrogen and irrigation interactions significantly increased stem and leaf biomass under low to moderate salinity, although this effect diminished under severe salinity. Optimising biomass production in marginal saline environments requires precise management of high water–nitrogen coupling in low –to – moderate saline soils, whereas extreme salinity necessitates salt-tolerant cultivar selection or soil improvement. Our study provides a theoretical foundation and technical parameters for the large-scale cultivation and sustainable biomass production of A. donax on marginal lands.
Author Contributions
Conceptualization, Y.J. and X.G.; methodology, Y.J. and Y.F.; formal analysis, Y.J. and S.L.; investigation, Y.J., T. Ch. and Z. D.; writing—original draft preparation, Y.J.; writing—review and editing, X.G.
Funding
This research was funded by the Natural Science Foudation of the Shanxi Province (grant number 202203021221075).
Conflicts of Interest
We all authors declare that there is no conflict on interest or personal relationships that could have appeared to influence the work reported in this paper.
References
- Jámbor, A.; Török, Á. The Economics of Arundo donax—A Systematic Literature Review. Sustainability 2019, 11, 4225. [Google Scholar] [CrossRef]
- Scordia, D.; Cosentino, S.L. Perennial Energy Grasses: Resilient Crops in a Changing European Agriculture. Agriculture (Basel) 2019, 9, 19. [Google Scholar] [CrossRef]
- Ngernsaengsaruay, C.; Puangsin, B.; Leksungnoen, N.; Khantayanuwong, S.; Chanton, P.; Thaepthup, T.; Wessapak, P.; Meeboonya, R.; Yimlamai, P.; Wanitpinyo, K.; et al. Morphology, Taxonomy, Culm Internode and Leaf Anatomy, and Palynology of the Giant Reed (Arundo donax L.), Poaceae, Growing in Thailand. Plants (Basel) 2023, 12, 1850. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Rivera-Vargas, P.; Serrat, X.; Nogués, S. Arundo donax L.: How High Photosynthetic Capacity Is Maintained under Water Scarcity Conditions. Agronomy (Basel) 2021, 11, 1089. [Google Scholar] [CrossRef]
- Goolsby, J.A.; Moran, P.J.; Martínez Jiménez, M.; Yang, C.; Canavan, K.; Paynter, Q.; Ota, N.; Kriticos, D.J. Biology of Invasive Plants 4. Arundo donax L. Invasive Plant Sci Manag 2023, 16, 81–109. [Google Scholar] [CrossRef]
- Lino, G.; Espigul, P.; Nogués, S.; Serrat, X. Arundo donax L. growth potential under different abiotic stress. Heliyon 2023, 9, e15521. [Google Scholar] [CrossRef]
- Ceotto, E.; Di Candilo, M. Shoot cuttings propagation of giant reed (Arundo donax L.) in water and moist soil: The path forward? Biomass and Bioenergy 2010, 34, 1614–1623. [Google Scholar] [CrossRef]
- Amaducci, S.; Perego, A. Field evaluation of Arundo donax clones for bioenergy production. Ind Crops Prod 2015, 75, 122–128. [Google Scholar] [CrossRef]
- Scordia, D.; Testa, G.; Cosentino, S.L. Perennial grasses as lignocellulosic feedstock for second-generation bioethanol production in Mediterranean environment. Ital J Agron 2014, 9, 84–92. [Google Scholar] [CrossRef]
- Martani, E.; Ferrarini, A.; Serra, P.; Pilla, M.; Marcone, A.; Amaducci, S. Belowground biomass C outweighs soil organic C of perennial energy crops: Insights from a long-term multispecies trial. Glob Change Biol Bioenergy 2021, 13, 459–472. [Google Scholar] [CrossRef]
- Rodriguez, L.D.; Confalone, A.E.; Lazaro, L.; Pimentel, R.M.; Lyra, G.B.; de Oliveira, J.F.; Singh, S.K.; Pereira, C.R. Growth of the energy crop giant reed (Arundo donax L.) and optimization of the ARMIDA model in the south-central region of Buenos Aires, Argentina. Ind Crops Prod 11812024, 211, 118190. [Google Scholar] [CrossRef]
- Visconti, D.; Fiorentino, N.; Cozzolino, E.; di Mola, I.; Ottaiano, L.; Mori, M.; Cenvinzo, V.; Fagnano, M. Use of giant reed (Arundo donax L.) to control soil erosion and improve soil quality in a marginal degraded area. Ital J Agron 2020, 15, 1764. [Google Scholar] [CrossRef]
- Siri Prieto, G.; Bustamante, M.; Battaglia, M.; Ernst, O.; Seleiman, M.F.; Sadeghpour, A. Effects of perennial biomass yield energy grasses and fertilization on soil characteristics and nutrient balances. Agron J 2021, 113, 4292–4305. [Google Scholar] [CrossRef]
- Ceotto, E.; Vasmara, C.; Marchetti, R.; Cianchetta, S.; Galletti, S. Biomass and methane yield of giant reed (Arundo donax L.) as affected by single and double annual harvest. Glob Change Biol Bioenergy 2021, 13, 393–407. [Google Scholar] [CrossRef]
- Arias, C.; Lino, G.; Sánchez, E.; Nogués, S.; Serrat, X. Drought Impact on the Morpho-Physiological Parameters of Perennial Rhizomatous Grasses in the Mediterranean Environment. Agriculture 2023, 13, 1233. [Google Scholar] [CrossRef]
- Pompeiano, A.; Guglielminetti, L.; Bargiacchi, E.; Miele, S. Responses in chemical traits and biomass allocation of Arundo donax L. to deficit resources in the establishment year. Chil J Agric Res 2013, 73, 377–384. [Google Scholar] [CrossRef]
- Romero-Munar, A.; Baraza, E.; Cifre, J.; Achir, C.; Gulías, J. Leaf plasticity and stomatal regulation determines the ability of Arundo donax plantlets to cope with water stress. Photosynthetica 2018, 56, 698–706. [Google Scholar] [CrossRef]
- Scordia, D.; D Agosta, G.M.; Mantineo, M.; Testa, G.; Cosentino, S.L. Life Cycle Assessment of Biomass Production from Lignocellulosic Perennial Grasses under Changing Soil Nitrogen and Water Content in the Mediterranean Area. Agronomy 2021, 11, 988. [Google Scholar] [CrossRef]
- Angelini, L.; Ceccarini, L.; Bonari, E. Biomass yield and energy balance of giant reed (Arundo donax L.) cropped in central Italy as related to different management practices. Eur J Agron 2005, 22, 375–389. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Y.; Wang, A.; Wu, T.; Guo, J.; Shi, G.; Tian, B.; Wei, F.; Cao, G. Integrated physiological and transcriptomic analysis reveals the involvement of photosynthesis and redox homeostasis in response of Arundo donax to low and high nitrogen supply. Ind Crops Prod 2024, 221, 119377. [Google Scholar] [CrossRef]
- Lloyd, L. Nackley, S.K. A salt on the bioenergy and biological invasions debate: salinity tolerance of the invasive biomass feedstock Arundo donax. Glob Change Biol Bioenergy 2015, 7, 752–762. [Google Scholar]
- Brilli, F.; Pignattelli, S.; Baraldi, R.; Neri, L.; Pollastri, S.; Gonnelli, C.; Giovannelli, A.; Loreto, F.; Cocozza, C. Root Exposure to 5-Aminolevulinic Acid (ALA) Affects Leaf Element Accumulation, Isoprene Emission, Phytohormonal Balance, and Photosynthesis of Salt-Stressed Arundo donax. International Journal of Molecular Sciences 2022, 23, 4311. [Google Scholar] [CrossRef] [PubMed]
- Müller, B.; Arcoverde Cerveira Sterner, V.; Papp, L.; May, Z.; Orlóci, L.; Gyuricza, C.; Sági, L.; Solti, Á.; Fodor, F. Alkaline Salt Tolerance of the Biomass Plant Arundo donax. Agronomy 2022, 12, 1589. [Google Scholar] [CrossRef]
- Ayub, M.A.; Ahmad, H.R.; Ali, M.; Rizwan, M.; Ali, S.; Zia Ur Rehman, M.; Waris, A.A. Chapter 3 - Salinity and its tolerance strategies in plants. In Plant Life Under Changing Environment; Tripathi, D.K., Pratap Singh, V., Chauhan, D.K., Sharma, S., Prasad, S.M., Dubey, N.K., Ramawat, N., Eds.; Academic Press, 2020; pp. 47–76. [Google Scholar]
- Triana, F.; Di Nasso, N.N.O.; Ragaglini, G.; Roncucci, N.; Bonari, E. Evapotranspiration, crop coefficient and water use efficiency of giant reed ( Arundo donax L.) and miscanthus ( Miscanthusxgiganteus Greef et Deu.) in a Mediterranean environment. Glob Change Biol Bioenergy 2015, 7, 811–819. [Google Scholar] [CrossRef]
- Cosentino, S.C.S.L.; Patanè, C.P.C.; Sanzone, E.S.E.; Testa, G.T.G.; Scordia, D.S.D. Leaf gas exchange, water status and radiation use efficiency of giant reed (Arundo donax L.) in a changing soil nitrogen fertilization and soil water availability in a semi-arid Mediterranean area. Eur J Agron 2016, 72, 56–69. [Google Scholar] [CrossRef]
- Pompeiano, A.; Landi, M.; Meloni, G.; Vita, F.; Guglielminetti, L. Allocation pattern, ion partitioning, and chlorophyll a fluorescence in Arundo donax L. in responses to salinity stress. Plant Biosyst 2017, 151, 613–622. [Google Scholar] [CrossRef]
- Cano-Ruiz, J.; Sanz, M.; Curt, M.D.; Plaza, A.; Lobo, M.C.; Mauri, P.V. Fertigation of Arundo donax L. with different nitrogen rates for biomass production. Biomass and Bioenergy 2020, 133, 105451. [Google Scholar] [CrossRef]
- Ullah, H.; Santiago-Arenas, R.; Ferdous, Z.; Attia, A.; Datta, A. Chapter Two - Improving water use efficiency, nitrogen use efficiency, and radiation use efficiency in field crops under drought stress: A review. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Pathum Thani, 2019; Volume 156, pp. 109–157. [Google Scholar]
- Pompeiano, A.; Remorini, D.; Vita, F.; Guglielminetti, L.; Miele, S.; Morini, S. Growth and physiological response of Arundo donax L. to controlled drought stress and recovery. Plant Biosyst 2017, 151, 906–914. [Google Scholar] [CrossRef]
- Galal, T.M.; Shehata, H.S. Growth and nutrients accumulation potentials of giant reed ( Arundo donax L.) in different habitats in Egypt. Int J Phytoremediation 2016, 18, 1221–1230. [Google Scholar] [CrossRef]
- De Battisti, D. Plant biomass allocation advances our understanding of plant adaptation to environmental gradients: a commentary on ‘Contrasting biomass allocations explain adaptations to cold and drought in the world’s highest-growing angiosperms’. Ann Bot 2024, 134, i–iii. [Google Scholar] [CrossRef]
- Di Nasso, N.N.O.; Roncucci, N.; Bonari, E. Giant reed (Arundo donax L.) as energy crop in Central Italy: a review. Ital J Agron 2013, 8, 10–17. [Google Scholar] [CrossRef]
Figure 1.
SPAD values of A. donax under different irrigation-nitrogen-salinity coupling treatments in 2023 (A) and 2024 (B).The values represent the means of each treatment, and the error bars indicate the standard deviation. Different letters indicate significant differences at p < 0.05. ANOVA p-value < 0.05, indicates the interaction is significant at the level of 0.05.
Figure 1.
SPAD values of A. donax under different irrigation-nitrogen-salinity coupling treatments in 2023 (A) and 2024 (B).The values represent the means of each treatment, and the error bars indicate the standard deviation. Different letters indicate significant differences at p < 0.05. ANOVA p-value < 0.05, indicates the interaction is significant at the level of 0.05.
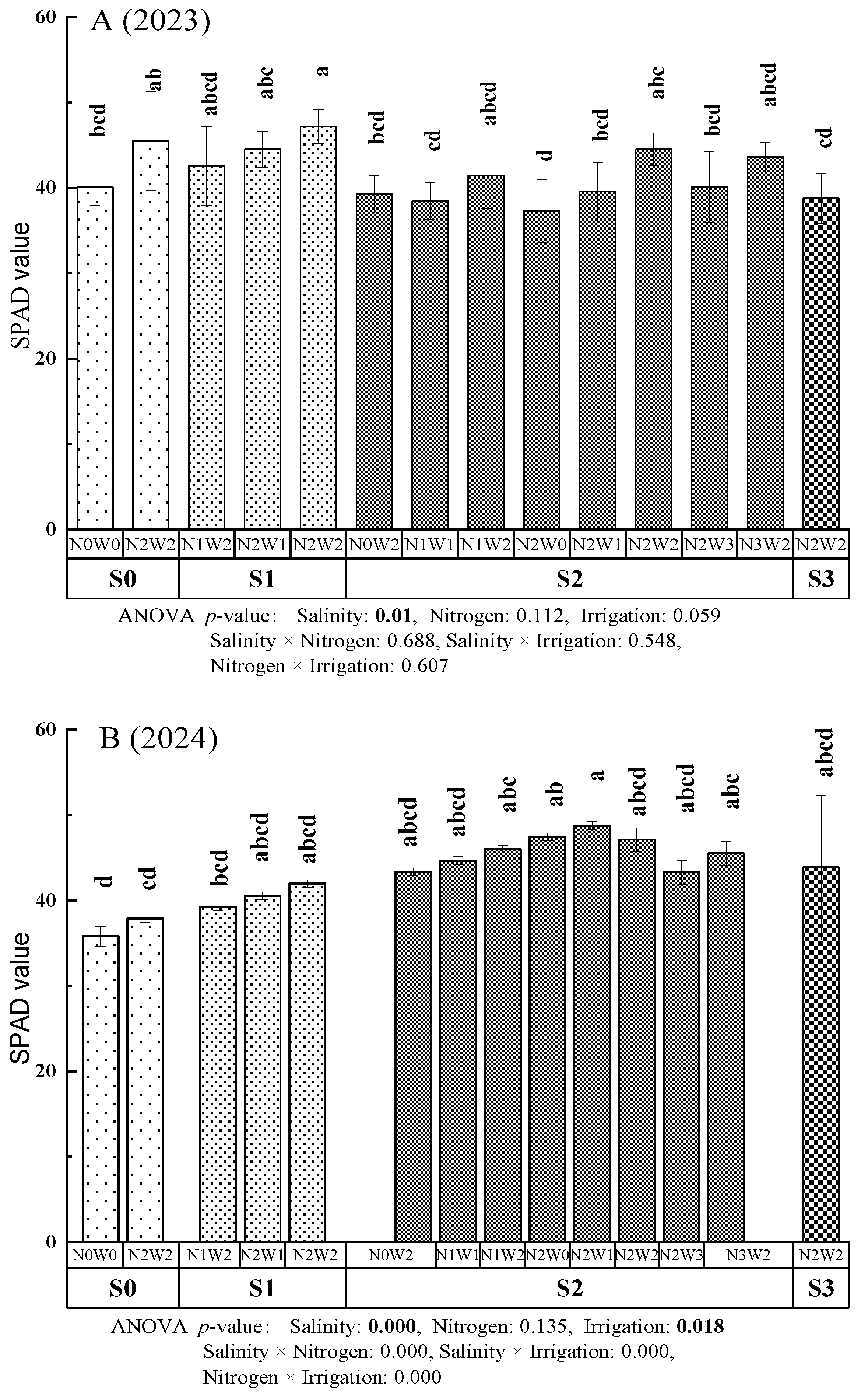
Figure 2.
Transpiration efficiency (TE) of A. donax under different irrigation-nitrogen-salinity coupling treatments in 2023 (A) and 2024 (B). The values represent the mean of each treatment, and the error bars indicate the standard deviation. Different letters indicate significant differences at p < 0.05. ANOVA p-value < 0.05, indicates the interaction is significant at the level of 0.05.
Figure 2.
Transpiration efficiency (TE) of A. donax under different irrigation-nitrogen-salinity coupling treatments in 2023 (A) and 2024 (B). The values represent the mean of each treatment, and the error bars indicate the standard deviation. Different letters indicate significant differences at p < 0.05. ANOVA p-value < 0.05, indicates the interaction is significant at the level of 0.05.
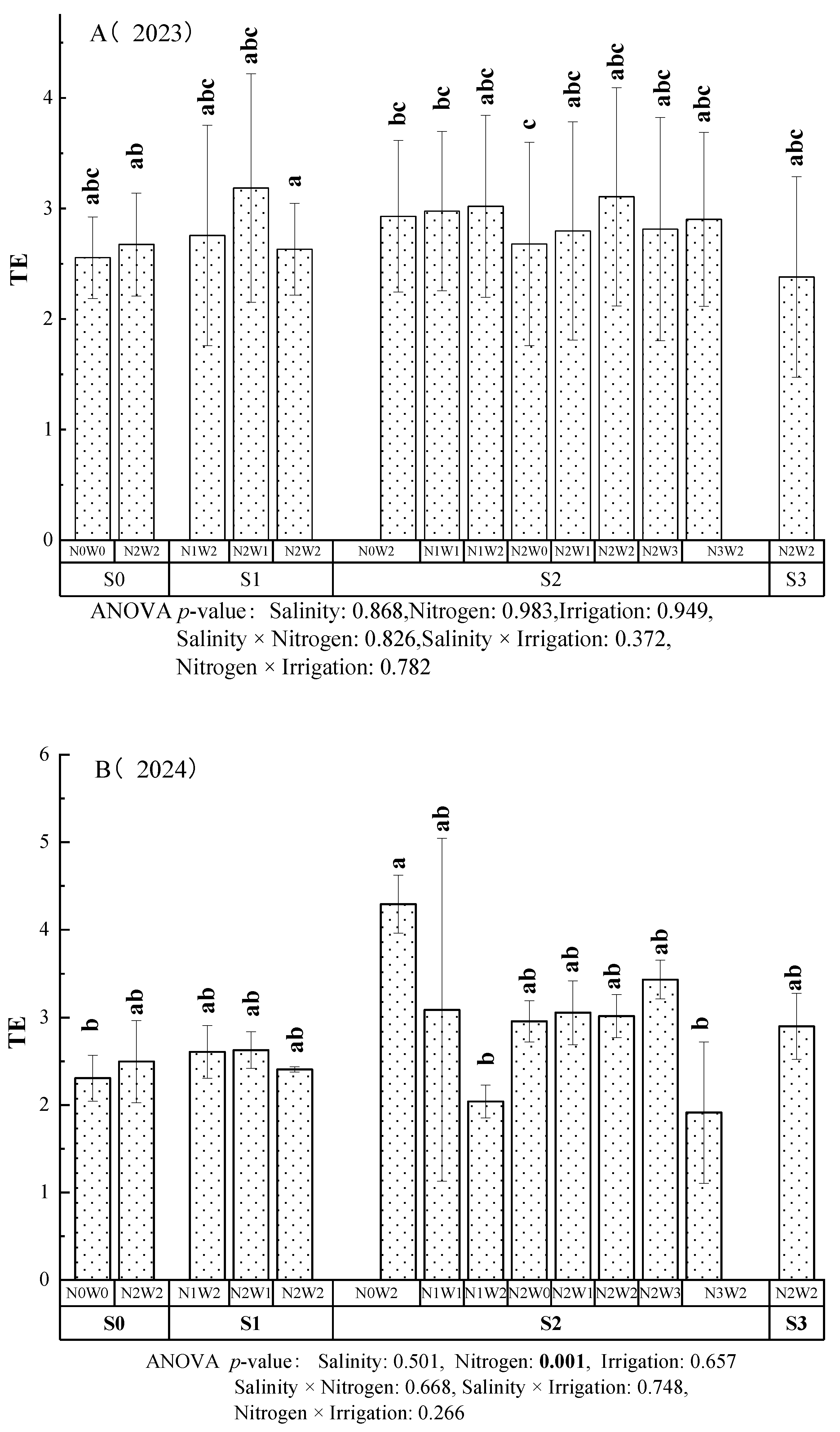
Figure 3.
Biomass accumulation of A. donax under different irrigation-nitrogen-salinity-coupling treatments in 2023 (A) and 2024 (B). The values represent the mean of each treatment, and the error bars indicated the standard deviation. Different letters indicate significance diffeences at p < 0.05. ANOVA p-value < 0.05, indicates the interaction is significant at the level of 0.05.
Figure 3.
Biomass accumulation of A. donax under different irrigation-nitrogen-salinity-coupling treatments in 2023 (A) and 2024 (B). The values represent the mean of each treatment, and the error bars indicated the standard deviation. Different letters indicate significance diffeences at p < 0.05. ANOVA p-value < 0.05, indicates the interaction is significant at the level of 0.05.
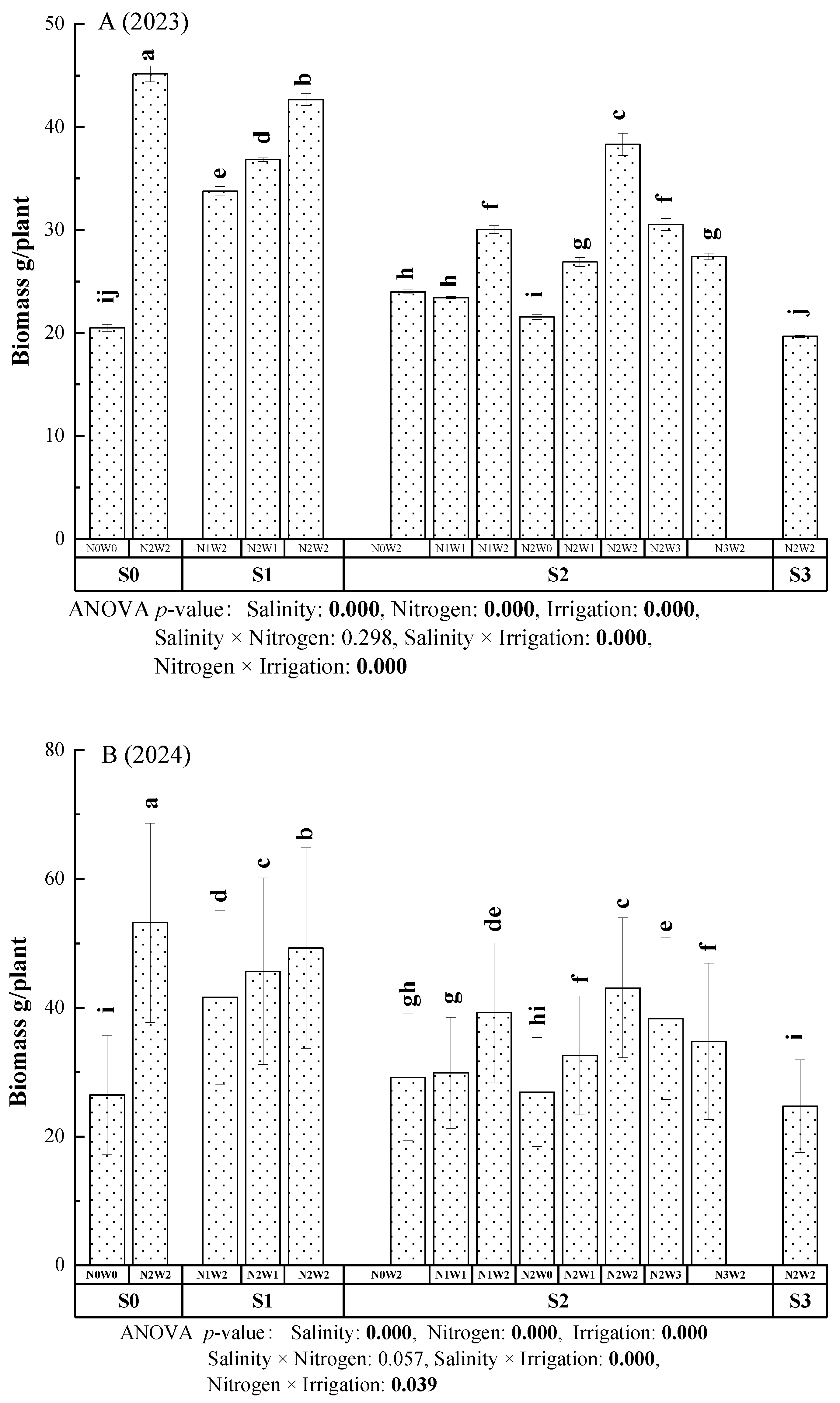
Figure 4.
Biomass allocation of A. donax under different treatments in 2023 (A) and 2024 (B). (a) Nitrogen application increasing under S2 and W2; (b) Irrigation amount increasing under S2 and N2; (c) Soil salinity increasing under N2 and W2.
Figure 4.
Biomass allocation of A. donax under different treatments in 2023 (A) and 2024 (B). (a) Nitrogen application increasing under S2 and W2; (b) Irrigation amount increasing under S2 and N2; (c) Soil salinity increasing under N2 and W2.
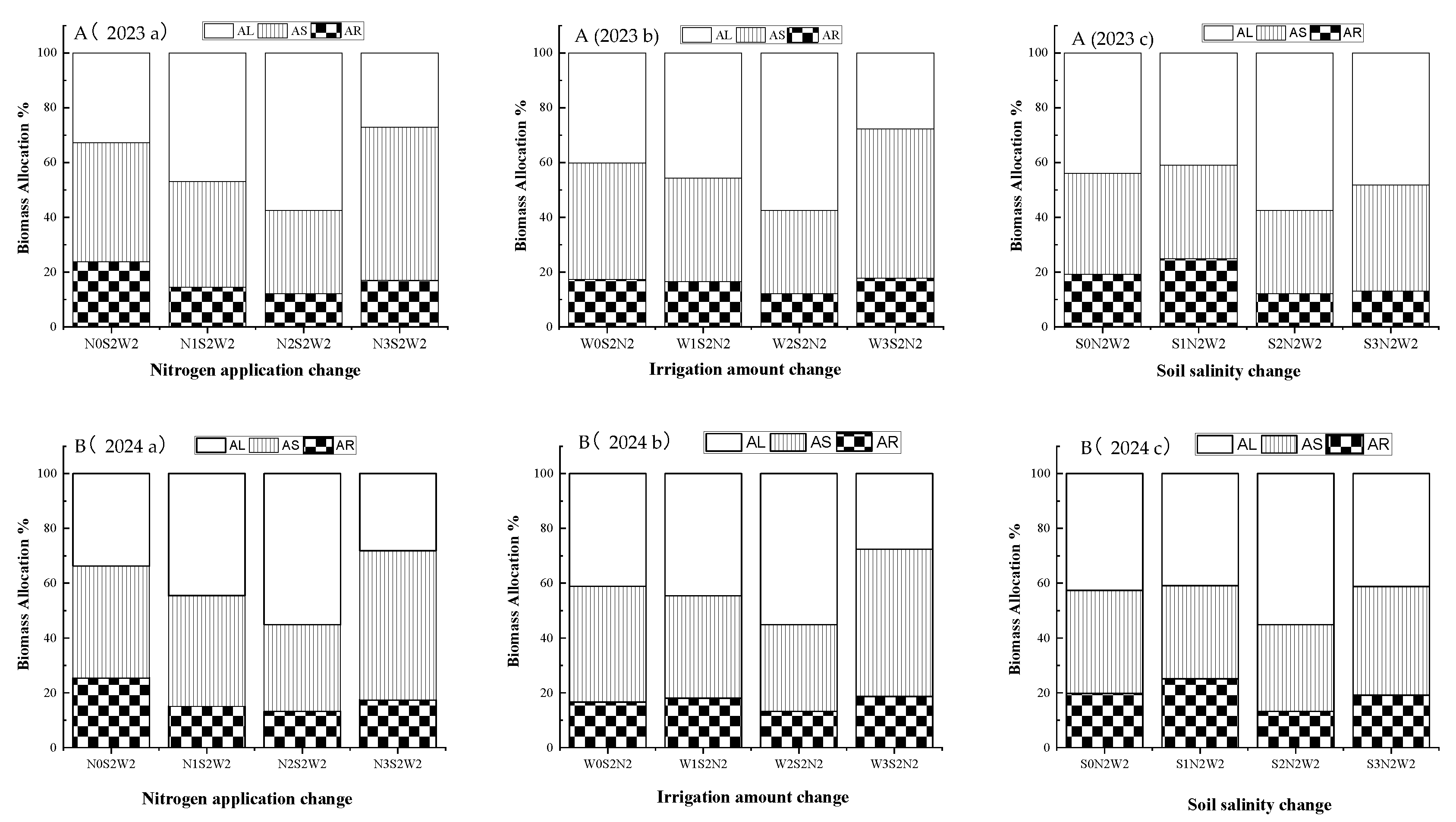

Table 1.
Morphological parameters of fully expanded leaves of A. donax under each treatment in 2023.
Table 1.
Morphological parameters of fully expanded leaves of A. donax under each treatment in 2023.
| Treatment | Leaf length (cm) | Leaf width (cm) | Leaf length–to –width ratio | Leaf area(cm2) |
| S0N0W0 | 15.95±0.53 gh | 0.64±0.01 f | 24.92±0.87 b | 8.36±0.32 fg |
| S0N2W2 | 23.75±0.25 a | 1.03±0.05 b | 23.09±1.10 bc | 20.06±1.11 a |
| S1N1W2 | 19.33±0.58 c | 0.84±0.04 cde | 22.97±1.79 bc | 13.35±0.25 cd |
| S1N2W1 | 18.43±1.12 cd | 0.94±0.04 bc | 19.66±1.87 d | 13.40±1.28 cd |
| S1N2W2 | 20.74±1.87 b | 1.06±0.04 b | 19.57±1.60 d | 17.08±2.88 b |
| S2N0W2 | 16.65±0.18 efgh | 0.84±0.05 cde | 19.77±0.93 d | 10.84±0.23 def |
| S2N1W1 | 16.30±0.51 fgh | 0.83±0.03 cde | 19.48±0.25 d | 10.23±0.68 ef |
| S2N1W2 | 17.04±0.81 defg | 1.32±0.30 a | 13.22±2.30 e | 15.64±4.16 bc |
| S2N2W0 | 15.17±0.55 h | 0.75±0.03 def | 20.23±0.21 cd | 9.33±0.66 efg |
| S2N2W1 | 16.94±0.68 defg | 0.91±0.04 bcd | 18.73±1.56 d | 11.85±0.46 de |
| S2N2W2 | 24.27±0.49 a | 0.78±0.06 cdef | 31.22±2.27 a | 15.52±1.16 bc |
| S2N2W3 | 17.83±1.01 de | 0.95±0.07 bc | 18.81±2.38 d | 13.11±0.65 cd |
| S2N3W2 | 17.55±0.27 def | 0.82±0.04 cde | 21.36±1.35 cd | 11.49±0.17 de |
| S3N2W2 | 13.44±0.92 i | 0.69±0.05 ef | 19.45±2.70 d | 7.16±0.39 g |
Table 2.
Morphological parameters of fully expanded leaves of A. donax under each treatment in 2024.
Table 2.
Morphological parameters of fully expanded leaves of A. donax under each treatment in 2024.
| Treatment | Leaf length (cm) | Leaf width (cm) | Leaf length–to –width ratio | Leaf area(cm2) |
| S0N0W0 | 15.24±1.41 de | 0.61±0.02 f | 25.02±2.76 ab | 6.13±0.52 f |
| S0N2W2 | 25.73±1.77 a | 1.04±0.07 b | 24.80±3.23 ab | 18.74±0.60 a |
| S1N1W2 | 20.55±2.94 bc | 0.87±0.06 cde | 23.94±4.90 b | 12.39±1.04 cd |
| S1N2W1 | 19.65±1.64 bcd | 0.94±0.04 bc | 20.81±1.09 b | 13.00±1.55 cd |
| S1N2W2 | 22.42±1.99 ab | 1.07±0.01 b | 21.00±1.63 b | 16.75±1.68 ab |
| S2N0W2 | 18.31±1.81 cde | 0.83±0.04 cde | 22.07±3.22 b | 10.56±0.80 de |
| S2N1W1 | 16.73±0.36 cde | 0.83±0.05 cde | 20.13±1.34 bc | 9.48±0.47 def |
| S2N1W2 | 17.14±3.27 cde | 1.28±0.19 a | 13.45±2.22 c | 15.49±4.64 bc |
| S2N2W0 | 14.68±0.71 e | 0.73±0.03 def | 20.11±0.63 bc | 7.29±0.58 ef |
| S2N2W1 | 16.88±1.31 cde | 0.91±0.04 bcd | 18.54±2.12 bc | 10.68±0.60 de |
| S2N2W2 | 23.01±1.87 ab | 0.77±0.07 cde | 30.11±4.33 a | 12.37±0.86 cd |
| S2N2W3 | 17.59±1.73 cde | 0.95±0.08 bc | 18.61±3.17 bc | 11.70±1.10 d |
| S2N3W2 | 17.16±1.39 cde | 0.81±0.08 cde | 21.26±1.95 b | 9.59±1.68 def |
| S3N2W2 | 13.58±0.32 e | 0.70±0.07 ef | 19.53±2.07 bc | 6.34±0.67 f |
Table 3.
Significance analysis of interaction among leaf morphology parameters in A. donax under different treatments in 2023 and 2024.
Table 3.
Significance analysis of interaction among leaf morphology parameters in A. donax under different treatments in 2023 and 2024.
| Source of variation | 2023 | 2024 | ||||||
| Length | Width | L:W | Area | Length | Width | L:W | Area | |
| S | ** | ** | ** | ** | ** | ** | ** | ** |
| N | ** | ** | * | ** | * | ** | ns | ** |
| I | ** | ** | ** | ** | ** | ** | * | ** |
| S×N | ** | ** | ** | * | ns | ** | ** | ** |
| S×I | ** | ** | ** | ns | ns | ** | ** | ns |
| N×I | ** | ** | ** | ns | ** | ** | ** | * |
Note: ‘*’ indicates p < 0.05, ‘**’ indicates p < 0.01 and ‘ns’ indicates no significance. L:W means leaf length–to –width ratio. S is salinity level, N is nitrogen application, I is irrigation amount.
Table 4.
Photosynthetic parameters of fully expanded leaves of A. donax under each treatment in 2023.
Table 4.
Photosynthetic parameters of fully expanded leaves of A. donax under each treatment in 2023.
| Treatment | Pn(μmol(CO2) m-2·s-1) | Tr(mmol·m-2·s-1) | Gs(mmol·m-2·s-1) | Ci(μmol·mol-1) |
| S0N0W0 | 8.86±0.59 bcd | 3.53±0.64 c | 10.68±5.49 ab | 414.80±17.26 ab |
| S0N2W2 | 14.62±1.74 a | 5.53±0.64 ab | 20.68±10.54 a | 352.80±8.80 c |
| S1N1W2 | 10.05±2.12 bcd | 3.90±1.14 c | 9.81±5.60 ab | 414.2±24.46 ab |
| S1N2W1 | 12.22±2.54 ab | 4.04±1.08 bc | 13.96±6.64 ab | 374.67±24.97 bc |
| S1N2W2 | 14.72±1.58 a | 5.64±0.60 a | 20.61±10.55 a | 352.67±6.11 c |
| S2N0W2 | 8.68±1.49 bcd | 3.06±0.78 c | 9.86±5.46 ab | 404.37±16.74 ab |
| S2N1W1 | 8.59±1.55 bcd | 2.98±0.75 c | 9.61±5.21 ab | 394.73±17.55 ab |
| S2N1W2 | 10.24±2.16 bcd | 3.53±0.96 c | 11.82±6.25 ab | 389.2±21.81 abc |
| S2N2W0 | 7.17±2.44 d | 2.72±0.43 c | 6.40±3.73 b | 425.63±27.77 a |
| S2N2W1 | 8.93±2.56 bcd | 3.29±0.59 c | 9.62±5.47 ab | 402.5±27.84 ab |
| S2N2W2 | 12.12±2.51 abc | 4.10±1.05 bc | 13.95±7.01 ab | 374.17±20.73 bc |
| S2N2W3 | 9.73±2.09 bcd | 3.70±1.10 c | 9.42±5.29 ab | 394.63±23.97 ab |
| S2N3W2 | 10.52±1.67 bcd | 3.79±1.03 c | 11.46±6.27 ab | 386.87±18.23 abc |
| S3N2W2 | 8.25±2.22 cd | 3.70±1.04 c | 9.28±4.37 ab | 409.30±23.83 ab |
Table 5.
Photosynthetic parameters of fully expanded leaves of A. donax under each treatment in 2024.
Table 5.
Photosynthetic parameters of fully expanded leaves of A. donax under each treatment in 2024.
| Treatment | Pn(μmol(CO2) m-2·s-1) | Tr(mmol·m-2·s-1) | Gs(mmol·m-2·s-1) | Ci(μmol·mol-1) |
| S0N0W0 | 7.33±1.06 d | 3.17±0.12 c | 13.34±1.76 cd | 401.00±17.62 abc |
| S0N2W2 | 8.97±0.58 cd | 3.65±0.52 bc | 23.03±1.67 a | 348.83±12.07 c |
| S1N1W2 | 10.23±0.36 bcd | 3.96±0.47 bc | 13.17±1.69 cd | 419.30±16.04 a |
| S1N2W1 | 11.55±0.53 abcd | 4.42±0.48 b | 17.28±2.09 bc | 384.97±27.73 abc |
| S1N2W2 | 13.29±0.58 abc | 5.53±0.31 a | 20.58±3.41 ab | 360.53±6.61 bc |
| S2N0W2 | 15.16±0.65 a | 3.55±0.37 bc | 13.03±1.95 cd | 413.03±18.34 ab |
| S2N1W1 | 9.75±5.78 bcd | 3.24±0.38 c | 12.96±2.07 cd | 396.03±23.11 abc |
| S2N1W2 | 7.64±0.54 d | 3.76±0.35 bc | 15.17±2.04 bcd | 395.80±17.01 abc |
| S2N2W0 | 9.30±0.55 cd | 3.15±0.21 c | 10.79±1.65 d | 428.23±25.84 a |
| S2N2W1 | 10.96±0.55 abcd | 3.61±0.37 bc | 12.98±2.06 cd | 405.53±23.69 abc |
| S2N2W2 | 12.65±0.55 abc | 4.21±0.36 bc | 17.25±2.28 bc | 377.80±15.65 abc |
| S2N2W3 | 14.30±0.55 ab | 4.18±0.35 bc | 13.10±2.24 cd | 396.10±36.03 abc |
| S2N3W2 | 7.13±1.71 d | 3.91±0.76 bc | 14.81±2.36 cd | 394.70±19.80 abc |
| S3N2W2 | 10.61±0.96 abcd | 3.67±0.16 bc | 11.30±1.16 cd | 396.40±13.15 abc |
Table 6.
Significance analysis of interaction among leaf photosynthetic paraments in A. donax under different treatments in 2023 and 2024.
Table 6.
Significance analysis of interaction among leaf photosynthetic paraments in A. donax under different treatments in 2023 and 2024.
| Source of variation | Leaf phothsynthtic paraments (2023) | Leaf phothsynthtic paraments (2024) | ||||||
| Pn | Tr | Gs | Ci | Pn | Tr | Gs | Ci | |
| S | ** | * | ns | * | ** | ** | ** | ** |
| N | * | ns | ns | ns | ** | ** | ** | ns |
| I | * | ns | ns | ns | ** | ** | ** | * |
| S×N | ns | ns | ns | ns | ns | * | ns | ns |
| S×I | ns | ns | ns | ns | ns | ns | ns | ns |
| N×I | ns | ns | ns | ns | ns | ns | ns | ns |
Note: ‘*’ indicates p < 0.05, ‘**’ indicates p < 0.01 and ‘n.s.’ indicates no significance. S is salinity level, N is nitrogen application, I is irrigation amount.
Table 7.
Water-related physiological parameters of fully expanded A. donax leaves under different treatments in 2023.
Table 7.
Water-related physiological parameters of fully expanded A. donax leaves under different treatments in 2023.
| Treatment | LER (mm/d) | LFW (g) | LDW (g) | SLW (g cm-2) | RWC (%) |
| S0N0W0 | 1.507±0.057 cd | 0.091±0.010 ef | 0.019±0.003 efg | 445.39±54.30 cd | 95.99 ab |
| S0N2W2 | 1.827±0.085 a | 0.118±0.007 cd | 0.020±0.002 efg | 986.24±103.78 a | 97.20 ab |
| S1N1W2 | 1.643±0.025 b | 0.160±0.024 b | 0.026±0.002 cd | 509.83±51.48 cd | 97.76 a |
| S1N2W1 | 1.837±0.116 a | 0.171±0.020 b | 0.031±0.004 b | 441.07±74.72 cd | 98.33 a |
| S1N2W2 | 1.837±0.025 a | 0.080±0.018 fg | 0.019±0.005 efg | 937.69±374.06 a | 96.91 ab |
| S2N0W2 | 1.437±0.021 d | 0.063±0.009 g | 0.014±0.002 h | 783.33±98.47 ab | 95.86 ab |
| S2N1W1 | 1.433±0.025 d | 0.134±0.007 c | 0.022±0.002 def | 460.44±53.77 cd | 97.66 a |
| S2N1W2 | 1.553±0.021 c | 0.114±0.009 cd | 0.025±0.003 d | 618.89±96.30 bc | 95.96 ab |
| S2N2W0 | 1.443±0.015 d | 0.133±0.005 c | 0.030±0.001 bc | 307.70±14.88 d | 94.84 b |
| S2N2W1 | 1.507±0.021 cd | 0.123±0.002 cd | 0.026±0.002 d | 463.31±36.57 cd | 97.76 a |
| S2N2W2 | 1.820±0.036 a | 0.081±0.004 fg | 0.016±0.003 gh | 982.07±255.23 a | 96.77 ab |
| S2N2W3 | 1.640±0.026 b | 0.216±0.008 a | 0.038±0.001 a | 342.76±9.67 d | 97.43 ab |
| S2N3W2 | 1.570±0.010 bc | 0.106±0.004 de | 0.019±0.002 fg | 619.00±59.82 bc | 96.64 ab |
| S3N2W2 | 0.983±0.006 e | 0.085±0.002 f | 0.023±0.001 de | 305.28±13.41 d | 95.01 b |
Table 8.
Water-related physiological parameters of fully expanded A. donax leaves under different treatments in 2024.
Table 8.
Water-related physiological parameters of fully expanded A. donax leaves under different treatments in 2024.
| Treatment | LER (mm/d) | LFW (g) | LDW (g) | SLW (g cm-2) | RWC (%) |
| S0N0W0 | 1.323±0.220 e | 0.094±0.010 bcd | 0.045±0.005 bcd | 136.49±6.08 d | 96.27 ab |
| S0N2W2 | 1.810±0.114 ab | 0.121±0.006 abcd | 0.058±0.003 abcd | 322.83±23.65 abc | 97.31 ab |
| S1N1W2 | 1.607±0.032 bcd | 0.162±0.024 abc | 0.078±0.012 abc | 159.93±12.39 cd | 97.75 ab |
| S1N2W1 | 1.817±0.064 ab | 0.174±0.020 ab | 0.083±0.010 ab | 158.16±32.61 cd | 97.91 a |
| S1N2W2 | 1.760±0.098 abc | 0.082±0.018 cd | 0.039±0.009 cd | 445.53±144.32 a | 96.48 ab |
| S2N0W2 | 1.437±0.015 de | 0.064±0.010 d | 0.031±0.005 d | 349.45±71.66 ab | 95.60 ab |
| S2N1W1 | 1.457±0.047 de | 0.136±0.007 abcd | 0.065±0.003 abcd | 145.15±10.20 d | 95.95 ab |
| S2N1W2 | 1.557±0.025 cde | 0.116±0.009 abcd | 0.056±0.004 abcd | 275.62±64.31 bcd | 95.41 ab |
| S2N2W0 | 1.553±0.125 cde | 0.133±0.009 abcd | 0.064±0.004 abcd | 115.20±17.45 d | 94.17 b |
| S2N2W1 | 0.510±0.036 de | 0.114±0.022 abcd | 0.055±0.011 abcd | 202.00±55.73 bcd | 96.77 ab |
| S2N2W2 | 1.853±0.031 a | 0.123±0.074 abcd | 0.059±0.036 abcd | 251.46±104.38 bcd | 96.37 ab |
| S2N2W3 | 1.627±0.040 abcd | 0.185±0.066 a | 0.089±0.031 a | 146.34±62.91 d | 97.08 ab |
| S2N3W2 | 1.550±0.030 cde | 0.100±0.012 abcd | 0.048±0.006 bcd | 200.91±25.75 bcd | 96.84 ab |
| S3N2W2 | 0.977±0.015 f | 0.087±0.001 cd | 0.042±0.001 cd | 152.63±14.72 cd | 94.51 ab |
Table 9.
Significance analysis of interaction among water-related physiological parameters of A. donax leaves under different treatments in 2023 and 2024.
Table 9.
Significance analysis of interaction among water-related physiological parameters of A. donax leaves under different treatments in 2023 and 2024.
| Source of variation | Leaf wate-related parameters (2023) | Leaf wate-related parameters (2024) | ||||||||||
| LER | LFW | LDW | SLW | RWC | LER | LFW | LDW | SLW | RWC | |||
| S | ** | ** | ** | ** | ns | ** | ns | ns | ** | * | ||
| N | ** | ** | ** | ** | ns | ** | * | ** | ** | ns | ||
| I | ** | ** | ** | ** | * | * | * | ** | ** | * | ||
| S×N | ns | ** | ns | ns | ns | ns | * | * | ** | ns | ||
| S×I | ** | ** | ns | ns | ns | ** | * | ** | ** | ns | ||
| N×I | ** | ns | ** | * | ns | * | ns | ns | ns | ns | ||
Note: ‘*’ indicates p < 0.05, ‘**’ indicates p < 0.01 and ‘ns’ indicates no significance.
Table 10.
Plant height, leaf number and biomass accumulation in A. donax under each treatment in 2023.
Table 10.
Plant height, leaf number and biomass accumulation in A. donax under each treatment in 2023.
| Treatment | Height(cm) | Number of Leaves | Root dry weight(g/plant) | Stem dry weight(g/plant) | Leaf dry weight(g/plant) |
| S0N0W0 | 83.12±5.08 e | 9.33±0.58 fg | 2.43±0.11 f | 11.50±0.51 h | 9.23±0.41 j |
| S0N2W2 | 141.23±5.25 a | 15.33±1.53 a | 5.00±0.08 d | 23.65±0.37 a | 18.97±0.30 b |
| S1N1W2 | 108.48±4.44 c | 13.00±1.00 bcd | 3.85±0.11 d | 18.20±0.51 e | 14.60±0.41 d |
| S1N2W1 | 122.12±3.14 b | 11.67±0.58 cde | 3.00±0.07 b | 14.20±0.33 d | 11.39±0.27 e |
| S1N2W2 | 137.35±4.00 a | 13.67±1.53 ab | 4.70±0.09 a | 22.23±0.42 c | 17.83±0.33 c |
| S2N0W2 | 78.24±5.47 ef | 11.00±1.00 ef | 2.92±0.15 e | 13.82±0.71 g | 11.09±0.57 hi |
| S2N1W1 | 76.65±5.49 ef | 11.33±1.15 df | 2.81±0.06 gh | 13.29±0.3 h | 10.66±0.24 g |
| S2N1W2 | 92.84±6.35 d | 11.00±1.00 ef | 3.52±0.09 g | 16.63±0.42 f | 13.34±0.33 de |
| S2N2W0 | 75.22±3.05 f | 9.00±1.00 g | 2.46±0.18 i | 11.64±0.83 i | 9.34±0.67 gh |
| S2N2W1 | 83.64±3.28 e | 11.67±0.58 cde | 3.03±0.11 gh | 14.33±0.52 g | 11.50±0.42 f |
| S2N2W2 | 122.12±2.13 b | 11.67±0.58 cde | 4.39±0.18 g | 20.75±0.86 f | 16.64±0.69 a |
| S2N2W3 | 102.91±4.24 c | 12.33±0.58 bcde | 3.42±0.06 f | 16.15±0.29 a | 12.96±0.23 h |
| S2N3W2 | 94.64±11.03 d | 13.33±1.15 bc | 3.07±0.05 g | 14.52±0.25 b | 11.65±0.20 i |
| S3N2W2 | 62.66±4.17 g | 8.00±1.00 g | 2.26±0.07 j | 10.67±0.32 j | 8.56±0.26 g |
Table 11.
Plant height, leaf number and biomass accumulation in A. donax under each treatment in 2024.
Table 11.
Plant height, leaf number and biomass accumulation in A. donax under each treatment in 2024.
| Treatment | Height(cm) | Number of Leaves | Root dry weight(g/plant) | Stem dry weight(g/plant) | Leaf dry weight(g/plant) |
| S0N0W0 | 97.91±9.82 de | 10.00±1.00 cd | 5.57±0.04 ef | 10.08±0.23 f | 5.50±0.16 j |
| S0N2W2 | 140.54±2.20 a | 14.67±1.53 ab | 8.79±0.10 c | 16.67±0.89 a | 18.91±0.90 b |
| S1N1W2 | 111.30±1.50 c | 12.00±1.00 abc | 7.49±0.38 d | 12.36±0.76 de | 14.80±0.52 d |
| S1N2W1 | 127.80±7.89 b | 11.00±1.00 bcd | 9.64±0.58 b | 14.20±0.10 bc | 13.65±0.32 e |
| S1N2W2 | 140.18±3.94 a | 15.00±2.00 a | 10.26±0.08 a | 13.89±1.04 bcd | 16.69±0.33 c |
| S2N0W2 | 79.10±0.37 gh | 12.00±2.65 abc | 5.99±0.36 e | 9.64±0.83 fg | 7.95±0.13i |
| S2N1W1 | 77.37±1.17 gh | 11.33±0.58 abcd | 5.35±0.26 ef | 9.58±0.27 fg | 9.82±0.18 fg |
| S2N1W2 | 93.76±1.15 ef | 12.00±1.00 abc | 4.95±0.19 f | 13.25±1.03 cde | 14.60±0.58 de |
| S2N2W0 | 76.84±0.58 gh | 8.67±0.58 cd | 3.68±0.36 g | 9.41±0.55 fg | 9.15±0.32 gh |
| S2N2W1 | 84.51±0.80 fg | 11.33±0.58 abcd | 4.94±0.10 f | 10.27±0.06 f | 12.26±0.25 f |
| S2N2W2 | 125.30±3.04 b | 11.67±1.15 abc | 4.93±0.12 f | 11.81±0.38 e | 20.59±0.58 a |
| S2N2W3 | 104.71±2.02 cd | 12.33±1.53 abc | 5.71±0.05 e | 16.49±0.72 a | 8.46±0.48 hi |
| S2N3W2 | 93.73±0.87 ef | 12.00±1.00 abc | 4.88±0.13 f | 15.26±0.07 ab | 7.87±0.44 i |
| S3N2W2 | 68.64±5.52 h | 8.00±1.00 d | 3.93±0.15 g | 8.17±0.16 g | 8.48±0.08 hi |
Table 12.
Significance analysis of interaction among plant height, leaves number and biomass accumulation in A. donax under different treatments in 2023 and 2024.
Table 12.
Significance analysis of interaction among plant height, leaves number and biomass accumulation in A. donax under different treatments in 2023 and 2024.
| Source of variation | 2023 | 2024 | ||||||||
| Height | Leaves Number | Root dry weight | Stem dry weight | Leaf dry weight | Height | Leaves Number | Root dry weight | Stem dry weight | Leaf dry weight | |
| S | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| N | ** | * | ** | ** | ** | ** | ns | ** | ** | ** |
| I | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| S×N | ns | ns | ** | ** | ** | ns | * | ** | ** | ** |
| S×I | ** | ns | ** | ns | ** | ** | * | ** | * | ** |
| N×I | ** | ns | ns | ** | ** | ** | ns | ns | ** | ** |
Note: ‘*’ indicates p < 0.05, ‘**’ indicates p < 0.01 and ‘ns’ indicates no significance.
Table 13.
Breakdown of correlation coefficients for key factors influencing biomass accumulation in A. donax.
Table 13.
Breakdown of correlation coefficients for key factors influencing biomass accumulation in A. donax.
| Variation | Simple correlation coefficient with biomass Y | path coefficient(direct function) | Correlation coefficient between independent variables | Indirect path coefficient | |||||
| X1 | X2 | X3 | X1-Y | X2-Y | X3-Y | SUM | |||
| X1 | ﹣0.477 | 0.707** | --- | 0.255 | 0.255 | --- | 0.100 | 0.130 | 0.230 |
| X2 | 0.342 | 0.391** | 0.255 | --- | 0.258 | -0.180 | ---- | 0.132 | -0.048 |
| X3 | 0.431 | 0.511** | 0.255 | 0.258 | --- | -0.180 | 0.101 | ---- | -0.079 |
Note: ‘**’ indicates a highly significant difference at p < 0.01.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



