Submitted:
06 May 2025
Posted:
07 May 2025
You are already at the latest version
Abstract
Kidney fibrosis and inflammation are significant contributors to the decline in renal function associated with aging. These processes are characterized by structural changes such as glomerular sclerosis and interstitial fibrosis, which exacerbate kidney injury and inflammation in aged individuals. Probiotics have gained increasing attention for their potential health-promoting effects. However, further investigation is required to fully understand the mechanisms underlying these benefits. We hypothesize probiotics could ameliorate fibrosis by immunomodulatory effects of probiotics and improving kidney tissue inflammation. Sixteen-month-old aging mice were administered Lacticaseibacillus paracasei MSMC39-1 for four months compared to young mice (6-month-old) and aged mice (20-month-old). The research found that following the administration of probiotic MSMC39-1, there were significant improvements in kidney inflammation, as evidenced by reductions in pro-inflammatory cytokines, fibrosis and inflammatory cells within the tissue. Moreover, the findings demonstrated that probiotic MSMC39-1 significantly normalized levels of MDA, and rescued antioxidant SOD and GPx in kidney tissue which was consistent with a low mitochondria biogenesis. More investigations revealed that conditioned medium from MSMC39-1 rescued epithelial kidney cell being damaged induced by high glucose. This research provides information and insights into the mechanisms underlying the beneficial health effects of probiotics, offering a deeper understanding of how these probiotics contribute to anti-aging processes in kidney.
Keywords:
Aging
; Chronic inflammation
; Anti-inflammation
; Antioxidant
; Probiotics
1. Introduction
Aging is a natural and complex process that involves a series of biological changes over time. It involves a variety of molecular and cellular changes, such as DNA damage, telomere shortening, and the accumulation of cellular waste products [1]. These are associated with oxidative stress by cell hoarding of oxygen reactive species (ROS), leading closely to inflammation [2]. Inflammation is characterized by an increase in pro-inflammatory cytokines; interleukin 1 beta (IL-1β), interleukin 6 (IL-6) and tumor necrosis factor-α (TNF-α), even in the absence of overt infection or injury [3]. This sterile inflammation is believed to play a role in the development of various age-related diseases and affect multiple systems including kidneys. Senescent cells in the renal tubular epithelium contribute to renal aging and are associated with increased levels of inflammatory cytokines and fibrosis markers [4,5,6]. The accumulation of senescent cells leads to a pro-inflammatory environment, often referred to as "inflammaging," which exacerbates kidney damage and fibrosis in aged individuals [5,7]. For example, aberrantly activated lysophosphatidic acid receptor 1 (LPAR1) is associated with chronic inflammation and renal fibrosis in aged kidneys and blocking it may reduce excessive inflammation in aged mice, suggesting sterile inflammation leading to fibrosis occurs in kidney during aging process [4].
Inflammatory response is a key driven factor in aging process in kidneys. Aged mice experienced more severe acute kidney injury (AKI) compared to young mice with increased pro-inflammatory pathways and cellular senescence [5]. AKI is associated with increased inflammation and cellular senescence, leading to higher mortality and morbidity in older patients [7]. Kidney where is resident and recruited macrophages, increased macrophage infiltration in aged mice contributes to chronic low-grade inflammation and fibrosis in aged kidneys, with ferroptosis signalling playing a crucial role [6]. Research evidence intestinal microbiota rejuvenation reduces renal interstitial fibrosis and delays renal senescence in aged male mice by inhibiting the NF-κB signaling pathway [8]. Additionally, probiotic supplementation can improve renal function and fibrosis in elderly mice by reducing intestinal inflammation and promoting M1-dominant renal inflammation [9]. Therefore, there has been a crosstalk of gut-kidney axis in maintenance homeostasis, microbiota or their metabolites may have a promising effect on kidney inflammation in aged mice either direct or indirect way. The gut microbiota experiences substantial shifts as one ages, which affects both its diversity and functional capacity. These shifts may lead to greater gut permeability and an increase in systemic inflammation [10,11,12].
Probiotics are live microorganisms that, when taken in sufficient quantities, provide health benefits to the host [13]. Most used probiotics include bacteria belonging to the Lactobacillus and Bifidobacterium genera [14]. Lactobacillus, notably, have proven to be an important genus with numerous beneficial biological characteristics. Potentially, L. reuteri LR12 and L. lactis LL10 illustrate positively scavenging activity of 2,2-diphenyl-1-picryl-hydrazyl-hydrate (DPPH) free radical, hydroxyl radical, and superoxide radical [15]. More specifically to a mixture of L. acidophilus La-14, L. casei Lc-11, L. lactis Ll-23, B. lactis Bl-04 and B. bifidum Bb-0, has been shown to lower the concentrations of TNF-α and IL-6 [16]. In this context, L. paracasei HII01 demonstrates the most significant impact on promoting longevity and anti-aging effects in Caenorhabditis elegans [17]. Based on our evidence, Lacticaseibacillus paracasei MSMC39-1 (formerly name: Lactobacillus paracasei MSMC39-1), previously, isolated from neonatal feces manifested anti-inflammatory effect on alcohol-induced hepatitis in rats and dextran sulfate sodium-induced colitis in rats [18,19]. Gaining insight into the processes of sterile inflammation and fibrosis in aging kidneys is essential for creating effective treatment strategies. This study aimed to assess the anti-inflammatory and antioxidant effects of the probiotic L. paracasei MSMC39-1 in an aging mouse model. Our findings suggest that L. paracasei MSMC39-1 may serve as a beneficial anti-aging probiotic, potentially slowing down the aging processes in the kidneys and offering a foundation for supplements aimed at preventing age-related diseases and enhancing gut health.
2. Results
2.1. MSMC39-1-Derived Condition Medium Rescues Kidney Epithelia Cell Being Damaged
Aging naturally affects various organs, especially kidneys, leading to structural and functional decline. This decline is often exacerbated by inflammation and oxidative stress. Additionally, high glucose levels significantly contribute to kidney damage and fibrosis by inducing oxidative stress, inflammation, and activating fibrogenic pathways. We treated epithelial kidney cell line with high glucose (HG) 25.5 mM for 48 hours with and without conditioned media (CM) compared to De Man, Rogosa, and Sharpe (MRS) broth control. Fresh medium was added and kept cells in low glucose medium until 192 hours. as shown in schematic experimental (Figure 1A). We examined the morphology of the cells and noted that high glucose (HG) conditions led to a change in cell shape at 96 hours, resulting in an irregular appearance (Figure 2B). At 192 hours, significant cell shrinkage was evident in the HG treatment. In contrast, the MRS treatment showed only minor alterations in cell morphology at 96 hours, but these changes became more pronounced over time (Figure 2B). Importantly, after the introduction of CM for 48 hours alongside HG treatment, CM successfully preserved the normal cellular architecture from 96 to 192 hours, exhibiting a morphology comparable to that of the control group (Figure 2B). The findings revealed that high glucose treatment led to a decline in cell proliferation, indicating possible cellular damage (Figure 1C). Conversely, epithelial kidney cells exposed to CM demonstrated a slight improvement in proliferation compared to the HG and MRS control groups (Figure 1C). Consistently, high glucose conditions lead to cellular damage and reduced proliferation, while CM may help to alleviate some of this damage, though its impact on overall cell proliferation is minimal.
2.2. Attenuation of Kidney Fibrosis and Injury in Aged Mice by MSMC39-1
To further determine whether MSMC 39-1 could protect kidney damaged in aged mice, mice were divided into 3 group, including normal young control age (YC) at 6-month-old, aged control (AC) 20-month-old, and mice that were started fed at 16-month-old with MSM39-1 via oral gavage till 20-month-old, called AM (Figure 2A). We assessed kidney pathology using hematoxylin and eosin (H&E) staining, which highlighted the pathological changes in both aged mice and those treated with MSMC39-1. As illustrated in Figure 2B, both the AM group and the YC group displayed normal glomerular architecture, size, and shape, with no signs of hypertrophy, fibrosis, or sclerosis in the kidney tissue. In contrast, the AC group exhibited mild glomerulosclerosis, slight mesangial cell proliferation, and mild glomerular hyperplasia, as shown in Figure 2B. In Figure 2C, there was a marked increase in the glomerular fibrotic area, as evidenced by collagen deposition observed through Masson’s Trichrome staining, in the AC group relative to the YC group. Furthermore, an increase in glomerular size was noted in the AC group. The administration of MSMC39-1 was found to reverse these changes. Overall, the results indicate that MSMC39-1 supplementation during middle age could mitigate kidney fibrosis related to aging.
2.3. Reducing Blood Glucose Levels and Enhancement of Kidney Antioxidant Defense by MSMC39-1
Elevated glucose levels, particularly in the context of diabetes or chronic hyperglycemia, play a crucial role in the development of kidney fibrosis, especially among aged individual. We subsequently assessed blood glucose levels in YC, AC, and AM mice. Our findings indicated that blood glucose was significantly higher in the AC group compared to the YC group. Importantly, the AM exhibited a notable reduction in blood sugar levels when compared to AC (Figure 3A). Additionally, we evaluated kidney lipid peroxidation by measuring malondialdehyde (MDA), an indicator of oxidative stress. The results showed that MDA levels were highest in the AC group, returning to normal following MSMC39-1 treatment in middle-aged mice, while MDA levels were elevated in AC compared to YC (Figure 3B). Glutathione peroxidase (GPx) and superoxide dismutase (SOD) are vital in alleviating oxidative stress and protecting cells from DNA damage. Following this, we measured the activities of GPx and SOD in kidney tissue. Our analysis showed that GPx (Figure 3C) and SOD (Figure 3D) activities were significantly lower in the AC group compared to the YC group. However, in the AM group, these activities were restored, indicating that MSMC39-1 possesses robust antioxidant properties. The findings from the MSMC39-1 treatment suggest that it can effectively improve both blood glucose levels and oxidative stress when compared to the aging group, aligning more closely with the young group.
2.4. Improving of Kidney Mitochondria DNA (mtDNA) Content in Aged Mice by MSMC39-1
The levels and integrity of mitochondrial DNA (mtDNA) are frequently affected by the cumulative impacts of aging, oxidative stress, and various age-related factors [20]. In our study, we assessed the mtDNA content in kidney tissue, which encompasses the total mtDNA present. The results revealed that kidneys from the aged group displayed a significantly elevated copy number of both ND1 and D-loop mtDNA (Figure 4A and 4B), in contrast to the relatively lower levels observed in young control mice. Following the administration of MSMC39-1, mtDNA levels in the aged group were restored to those typical of the young control group. This suggests that MSMC39-1 may serve as a potential agent for normalizing mtDNA levels in aged kidney tissue.
2.5. Anti-Inflammation in Kidney by MSMC39-1
The probiotic MSMC39-1 demonstrated a notable capacity in this study to reduce pro-oxidative stress and fibrosis in the kidneys. The interplay between oxidative stress and inflammation may contribute to the development of tissue fibrosis. Consequently, we assessed the levels of the inflammatory cytokines TNF-α and IL-1β in kidney tissue. In the MSMC39-1 group, there was a significant reduction in TNF-α and IL-1β levels compared to the aged group, while the values were not significantly different when compared to the young group (Figure 5A and 5B).
2.6. Positive Effects of MSMC39-1 on Maintained Colon Integrity and Anti-Inflammation
The interplay between the kidneys and the gut is significant, as they collaboratively contribute to the maintenance of homeostasis and the prevention of inflammation and oxidative stress at both local and systemic levels [21]. We hypothesized that MSMC39-1 might safeguard against alterations in gut architecture induced by aging, which could lead to inflammation in the kidneys. Subsequently, we assessed the gut histology across three groups: AC, YC, and AM. The AM group demonstrated a reduction in inflammatory conditions; the colonic mucosa was preserved, exhibiting uniform colonic tissue and epithelial cells with normal villi and crypt structures, along with the lamina propria showing minimal chronic inflammatory cell infiltration (Figure 6A). These findings were comparable to those observed in the young group (Figure 6A). In contrast, the colonic tissue of the AC group displayed areas with depleted goblet cells, structural loss of villi, and crypts (Figure 6A). Furthermore, inflammatory cell infiltration was evident throughout the lamina propria, submucosa, and deeper layers (Figure 6A). Histological grading indicated that the probiotic MSMC39-1 significantly improved the colitis score compared to the AC group, as illustrated in Figure 6B. In summary, MSMC39-1 appears to be a promising target for preserving gut architecture and exerting anti-inflammatory effects in aged mice.
3. Discussion
Aging is associated with physiological alterations in the body that can result in diminished functionality and the emergence of diseases, such as dementia, cardiovascular ailments, diabetes mellitus, and several chronic conditions, including kidney diseases [22]. Probiotics is one of the saftest supplementations that has potential to protect or restore the aging process. Probiotics have been suggested that they relief systemic cytokine makers, improved antioxidant level resulting to anti-aging effects [23]. The kidney plays a vital role in the aging process and is susceptible to the effects of oxidative stress and inflammation. This study investigates the kidneys of aging mice, based on a model created by Flurkey et al. (2007). In this model, young mice are represented by those aged 3–6 months, while the aging group consists of mice aged 18–24 months [24]. It is consistent with the mice ages in our study. The probiotic MSMC39-1 was administered to mice starting in middle age and continued throughout their old age. The results indicated that treatment leads to significant improvements in oxidative stress, inflammation, and fibrosis in the kidneys of aging mice, highlighting the important roles of probiotics in antioxidant and anti-inflammation.
Kidney fibrosis is a common pathological condition linked to aging, characterized by the excessive buildup of extracellular matrix components, leading to tissue scarring and reduced renal function. Our research demonstrates that MSMC39-1 effectively prevents collagen accumulation associated with aging through various cellular and molecular mechanisms, including inflammation, and oxidative stress. MSMC39-1 inhibits the production of inflammatory cytokines, highlighting the connection between inflammation and fibrosis. In agreement, Lacticaseibacillus paracasei HY7207 has shown efficacy in reducing hepatic steatosis, inflammation, and fibrosis in a mouse model of non-alcoholic fatty liver disease (NAFLD) [25]. While the study focused on liver health, the mechanisms by which Lacticaseibacillus paracasei reduce inflammation and fibrosis in the liver could
potentially be applicable to the kidneys and other tissues, this suggests the systematic effect of probiotics supplementation in mouse model. Chronic kidney disease (CKD) is associated with systemic inflammation and oxidative stress in the kidneys, which may result in various complications and a decline in renal function [26]. As a result, the findings align with the work of Wagner et al. (2022), indicating that dysbiosis facilitates the colonization of bacteria that produce urease, indole, and p-cresol-forming enzymes, ultimately leading to the accumulation of uremic toxins [27]. We did not measure those toxic agents produced in aged kidney, but tissue inflammation was probably due to toxic agents’ accumulation in aged organ in our model. Instead, we found that blood glucose is being increased in aged mice, this is likely to be a damaged molecule in aged mice that leads to systematic effect. In addition, studies indicate that the use of Lactobacillus paracasei and Lactobacillus plantarum can lead to improved kidney function and a reduction in inflammation, potentially serving as a preventive measure against the progression of chronic kidney disease [28]. This is achieved through the enhancement of the downregulation of harmful metabolites, uremic toxin. Likewise, probiotics develop natural defenses that inhibit harmful bacteria, including those that generate uremic toxins. Recent research has demonstrated that the administration of probiotics can modify the intestinal microbiota in aging mice, resulting in improved outcomes for both acute and chronic kidney injuries [9]. MSMC39-1 supplementation exhibits an anti-inflammatory effect by reducing pro-inflammatory cytokines level; IL-1β and TNF-α in aged kidney mice. Similarly, colon histology score, immune cell infiltration, crypt length distortion, and villi damaged in aged mice is also rescued by MSMC39-1 supplementation. This effect underscores probiotic properties such as immune system modulation via communication between renal and the gut system, which are known as gut-kidney axis. Moreover, strains of L. paracasei have demonstrated the ability to enhance the integrity of the gut barrier by elevating the levels of tight junction proteins and mucin, thereby contributing to the reduction of inflammation and the preservation of gut health [29,30]. This is crucial in preventing conditions like colitis and other inflammatory bowel diseases [29,30]. L. paracasei X11 supplementation has been shown to lower serum uric acid levels and reduce renal inflammation, which is beneficial in preventing chronic kidney disease [31]. This suggests a protective role in the gut-kidney axis.
Aging in kidneys is characterized by increased inflammation and oxidative stress, which further promote fibrosis. This can be achieved through several mechanisms, including NF-κB, along with the production of reactive oxygen species (ROS) [32,33]. Our research indicates that the restoration of pro-inflammatory cytokine expression in the kidneys of aged mice may be attributed to the inhibitory influence of NF-κB activation, as both TNF-α and IL-1β are recognized as well-characterized target genes of NF-kB. We found both cytokines are significantly restored in aged kidney that is consistent with improved kidney histology. Nevertheless, we were unable to establish a clear link between oxidative stress and inflammation, as both factors may exacerbate each other in a feedback loop. An imbalance between the production and neutralization of ROS results in the onset of oxidative stress [34]. As a result, an overabundance of hydroxy radicals can damage cell membranes, resulting in lipid peroxidation. This process subsequently elevates the levels of MDA in aging kidneys. The study revealed that MSMC39-1 reduces MDA levels while enhancing the activities of SOD and GPx in the kidney. Based on these findings, an examination of the organ's morphology was conducted, which may correlate with the antioxidative levels indicated by H&E staining. In alignment with the findings of Oguntoye et al. (2023), it was observed that Wistar rats receiving provitamin A cassava hydrolysate in conjunction with Lactobacillus rhamnosus exhibited a notable enhancement in antioxidant biomarkers within their kidneys, heart, and liver [35]. In individuals with chronic kidney disease (CKD), the use of probiotics, prebiotics, and synbiotics has been associated with improvements in markers of oxidative stress, such as elevated glutathione (GSH) and total antioxidant capacity (TAC); however, the specific impacts on GPx and SOD remain unspecified [36]. This indicates potential for probiotics to enhance overall antioxidant capacity, which could indirectly benefit kidney function. The ability of probiotic MSMC39-1 to neutralize free radicals at the cellular and tissue levels is apparent. This aligns with the findings of Lin et al. (2022), which showed that the combination of Lactobacillus and Bifidobacterium probiotics elevates SOD, catalase, and GPx levels in the brain and liver, while decreasing MDA levels in mice undergoing natural aging [37].
Findings from a population-based study suggest that higher levels of fasting glucose are related to glomerular hyperfiltration and microalbuminuria in adults of middle age, in contrast to insulin resistance measures, which do not exhibit such a connection [38]. Our research indicated that aged mice exhibited an increased blood glucose level, which aligns with kidney damage characterized by fibrosis and inflammation. The heightened glucose levels in aging mice contribute to kidney fibrosis and inflammation via multiple mechanisms, such as cellular senescence and oxidative stress. How did MSMC30-1 has an effect in improving inflammation and oxidative stress in aged kidney remains elusive. We observed in aging kidneys, mtDNA levels are elevated, correlating positively with inflammation, oxidative stress, and histological changes. This suggests that increased glucose levels may lead to mitochondrial damage, either directly or indirectly. Additionally, aging kidneys seem to activate compensatory mechanisms to reduce mitochondrial damage by enhancing the production of new mitochondria, thereby maintaining cellular energy and function, as indicated by mtDNA levels. Research indicated that a reduction in cytochrome c levels within kidney mitochondria plays a significant role in the impairment of oxidative phosphorylation in older rats. This decline is essential for the process of oxidative phosphorylation (OXPHOS) and results in diminished mitochondrial function in the kidneys of aged rats [39]. Moreover, aging is associated with a reduction in mitochondrial mass and function, which exacerbates renal fibrosis and cellular senescence, primarily through the activation of Wnt/catenin/RAS signaling pathways that are implicated in age-related renal fibrosis [40]. Studies have found that high levels of glucose aggravate renal inflammation and fibrosis through multiple pathways, including cellular senescence and the production of inflammatory cytokines. One study highlights that elevated glucose induces senescence in renal tubular epithelial cells, resulting in increased fibrosis and inflammation in diabetic kidney disease. This mechanism is driven by the secretion of Sonic hedgehog (Shh), which mediates the activation and proliferation of fibroblasts in diabetic kidney disease (DKD) [41]. Future studies are required to evaluate how gut microbiome in aged gut and kidney is being altered by supplementation MSMC39-1. In addition, how dose MSMC39-derived metabolites involve mitigating the inflammatory and oxidative stress in aged kidney are also needed to address.
4. Materials and Methods
4.1. Vero Cells Culture
The African green monkey kidney cell line, known as Vero cells (CCL-81) was obtained by Dr. Sakdiphong Punpai, Inovative Learning Center and Center of Excellence in Medical and Environmental Innovative Research (CEMEIR), Srinakharinwirot University, Bangkok, Thailand. Cells were regularly maintained in a medium consisting of DMEM low glucose with 1000 mg/L D-glucose and sodium pyruvate (Hyclone, USA). This medium was enriched with 10% fetal bovine serum and penicillin-streptomycin (Hyclone, USA). The Vero cells were incubated at a temperature of 37 °C in a humidified incubator continuously flushed with a mixture of 5% CO2 and 95% air.
4.2. Probiotic Strain and Culture Conditions
A probiotic Lacticaseibacillus paracasei MSMC39-1 strain was sourced from Center of Excellence in Probiotics, Faculty of Medicine, Srinakharinwirot University, Bangkok, Thailand. The probiotics was cultivated in de Man-Rogosa-Sharpe (MRS) medium (HiMedia Lab., Mumbai, India) at 37°C under anaerobic conditions for 24 hours and 48 hours, respectively. Afterwards, the bacterial suspension density was adjusted to 109 colony forming units (CFU) in phosphate-buffered saline for oral administration in the mice. For in vitro experiment, supernatant culture was collected and centrifuged at 4000 rpm for 10 min at 4 °C. Then, the supernatant was filtered through 0.22 µm and stored at -80 °C until use.
4.3. In Vitro Cell Damaging Induced by Glucose Experiment
Vero cells were seeded in a 96-well plate at a density of 2,000 cells per 100 µl and incubated overnight. The next day, they were treated with high glucose (Sigma, USA) at 25.5 mM, with and without conditioned medium, for 48 hours. The medium was then replaced with fresh warm low glucose medium, and the cells were maintained until day 8. Throughout this period, the cells were monitored and photographed using a Matero TL Digital Transmitted Light Microscope (Leica Microsystems). Cell proliferation was assessed using the WST-8 (MedChem Express, USA) assay at 460 nm with a Multiskan SkyHigh (Thermo Scientific).
4.4. Mitochondria DNA Measurement
Total genomic DNA was extracted from kidney tissue using GF-1 Tissue DNA Extraction (Vivantis, Malasia) according to the manufacturer’s instructions. Ten nanograms of total cellular DNA was used as a template for mitochondria DNA amplification by real time PCR with the following sequences encoded for mtDNA ND1 (mtND1) 5’ CTAGCAGAAACAAACCGGGC 3’, 5’ CCGGCTGCGTATTCTACGTT 3’; mtDNA D-loop (mtD-loop) 5’ AATCTACCATCCTCCGTGAAACC 3’, 5’ TCAGTTTAGCTACCCCCAAGTTTAA 3’; Total cellular mtDNA was normalized using endogenous genomic HK2; 5’ GCCAGCCTCTCCTGATTTTAGTGT 3’, 5’ GGGAACACAAAAGACCTCTTCTGG 3’.
Total cellular mtDNA was quantified by ExcelTaq™ 2X Fast Q-PCR Master Mix (SYBR, ROX) (SMBIO, Taiwan) using the StepOnePlus™ Real-Time PCR System (Applied Biosystems). The relative abundance of total mtDNA level was calculated using = 2(-∆∆Cq) according to reference method [42].
4.5. Experimental Animals
Two-month-old male C56BL/6NJcl mice (Mus musculus) were purchased from Nomura Siam International Co, Ltd (Bangkok, Thailand). Animal experiments were carried out according to the guidelines of the Ethics and Research Standardization Section, Srinakharinwirot University (approval number: COA/AE-015-2564). In short, all mice were housed in a room under controlled temperature and humidity ranges of 22± 2°C and 55± 10%, respectively, on 12 h light–dark cycle and free access to food and water. After 1 week of acclimatization, mice were randomly divided by the design of the aim of this study following into three groups: (1) young group aged 6 months, (2) aging group aged 20 months, and (3) MSMC39-1 group aged 20 months. MSMC39-1 group was fed normal food and water freely for 14 months, then received 100 µL MSMC39-1 via oral gavage at 109 CFU/mL for 4 months then were euthanized.
4.6. Organ Collection and Preparation
The mice were euthanized using isoflurane anesthesia. Briefly, their abdomens were opened, and the diaphragm barrier between the lungs and liver was cut for reaching the heart and excision, then mice died peacefully without pain. After that, the kidney and colon were all removed. The organs were stored at -80 °C then homogenization. The 50-100 mg of kidney was weighed and homogenized in 500 µL 10% (w/v) RIPA buffer using an ultrasonic homogenizer (Sonoplus HD 2070; Bandelin, Germany) at 25-30% power for 3 min. All procedures were performed on ice. Thereafter, the homogenates were centrifuged at 15,000 x g for 10 min at 4 °C. Each supernatant was collected and stored at -80 °C for inflammatory cytokines measurement and oxidative stress study. Histopathological analysis: the samples were fixed in 4% (w/v) paraformaldehyde for 48 hours. The kidney was sliced in 4 µm thickness and colon was sliced in 5-7 µm thickness. All organs were prepared for further study.
4.7. Blood Glucose Evaluation
The blood sample was collected by heparinized blood and centrifuged at 1500 x g for 15 min at 4 °C. Total sugar was measured using plasma by the Professional Laboratory Management Corp, Co., Ltd., Bangkok, Thailand.
4.8. Inflammatory Cytokines Evaluation by ELISA
The samples were measured inflammatory cytokines; TNF-α and IL-1β, using mouse TNF-α DuoSet ELISA (#DY410-05, Minnepolis, MN), a Mouse IL-1 beta/IL-1F2 DuoSet ELISA (#DY401-05, Minnepolis, MN). All the operations were performed under the manufacturer’s protocols. Briefly, ELISA plate (#DY990, R&D systems) was coated with capture antibody overnight at 4 °C. After washing the plate, reagent diluent was added for blocking the wells and incubate for 2 hours at room temperature. Next, the wells were washed and added supernatant (100 µL/well) overnight at 4 °C. Each supernatant was washed off and incubated with detecting antibody for 2 h at room temperature. The streptavidin-HRP and substrate were added for starting the reaction. After that, the wells were added stop solution and read at 450 nm using Biochrom microplate reader (Anthos 2010, Austria). The concentration of TNF-α, IL-1β were calculated using a standard curve which obtained from TNF-α, IL-1β (mouse) ELISA standard following the manufacturer’s protocols.
4.9. Lipoperoxidation and Anti-Oxidative Stress Evaluation
The samples were measured oxidative markers, including Superoxide dismutase (SOD), Malondialdehyde (MDA) using TBARS Assay Kit (#10009055, Cayman Chemical Company, Ann Arbor, Michigan), and Superoxide Dismutase (SOD) using Superoxide Dismutase Kit (#706002, Cayman Chemical Company, Ann Arbor, Michigan), and Glutathione (GPx) using Glutathione Peroxidase Assay Kit (#703102, Cayman Chemical Company, Ann Arbor, Michigan). All the operations were performed under the manufacturer’s protocols. The specific absorbance was read using Biochrom microplate reader (Anthos 2010, Austria). Each activity of enzymes was measured following the manufacturer’s protocols and using a standard curve.
4.10. Histopathological Analysis
The kidney and colon tissues were labeled with hematoxylin and eosin (H&E). In addition, kidney tissue was labeled with Masson’s Trichrome for fibrosis. The stained sections were observed and examined under a light microscope (OlympusBX53, Olympus Corporation, Tokyo, Japan). Masson’s Trichrome staining positive area and glomerulus area were analyzed using ImageJ software. For colitis manifestation, the sections were graded by two independent investigators blinded to the different cohorts. Score ranged from 0 to 18 (total score), which represents the sum of the scores from grade of inflammation (0-3), extent within the intestine layers (0-3), regeneration (0-4), crypt damage (0-4) and percentage of involvement (0-4), as following Table 1.
4.11. Statistical Analysis
The data were analyzed using GraphPad Prism 9 software, the bars on the graph depict mean ± SD. The statistical analysis of the data was carried out using one-way or two-way analysis of variance (ANOVA). Values of p < 0.05 and 0.01 were considered statistically significant.
5. Conclusions
Inflammation and oxidative stress are closely associated with the aging process and key driver in kidney fibrosis. The research demonstrates that probiotic MSMC39-1 exerts a preventive effect in an aging mice model, likely due to its ability to improve gut inflammation in aging mice. The administration of MSMC39-1 demonstrated its efficacy through several mechanisms: a reduction in pro-inflammatory cytokines, downregulation of oxidative stress, improvement in antioxidant system in aged kidney and enhancement of the colon's tight junctions and histological structure. Aged kidney tissue showed significant change in morphology and fibrosis area. Our finding suggests that MSMC39-1 may play a crucial role in establishing new gut homeostasis in aging mice targeting kidney fibrosis.
Author Contributions
Conceptualization, M.T. and T.J.; methodology, P.S., J.N., K.W., C.C. and P.J.; software, P.S. and J.N.; validation, T.J. and J.N.; formal analysis, P.S., J.N. and K.W.; investigation, P.S., J.N., K.W., C.C., and P.J.; resources, M.T. and T.J..; data curation, P.S., J.N. and T.J.; writing—original draft preparation, P.S.; writing—review and editing, T.J., M.T. and J.N..; visualization, T.J., M.T. and J.N.; supervision, M.T. and T.J.; project administration, M.T.; funding acquisition, M.T. and T.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundamental Fund, Thailand Science Research and Innovation (grant number 028/2567), Faculty of Medicine and HRH Princess Maha Chakri Sirindhorn Medical Center, Srinakharinwirot University (grant number149/2566, grant number 505/2566), and Excellence Center in Probiotics Srinakharinwirot University, (grant number 055/2567) and This research was partly supported by Walailak Univeristiy grant for New Researcher Development Scheme (grant number WU67264).
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board of Srinakharinwirot University (No. SWU-A-015-2564). Institutional animal care and use committee statement: All protocols were carried out in accordance with relevant guidelines and regulations.
Informed Consent Statement
This study did not involve human subjects or human-derived materials.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lopez-Otin, C. , et al., The hallmarks of aging. Cell, 2013, 153, 1194-217.
- Aleksandrova, K., L. Koelman, and C.E. Rodrigues, Dietary patterns and biomarkers of oxidative stress and inflammation: A systematic review of observational and intervention studies. Redox Biol, 2021, 42, 101869,.
- Zhang, J.M. and J. An, Cytokines, inflammation, and pain. Int Anesthesiol Clin, 2007, 45, 27-37.
- Jin, Y. , et al., Role of Aberrantly Activated Lysophosphatidic Acid Receptor 1 Signaling Mediated Inflammation in Renal Aging. Cells, 2021, 10(10).
- Marquez-Exposito, L. , et al., Acute Kidney Injury is Aggravated in Aged Mice by the Exacerbation of Proinflammatory Processes. Front Pharmacol, 2021, 12, 662020.
- Wu, L. , et al., Macrophage iron dyshomeostasis promotes aging-related renal fibrosis. Aging Cell, 2024, 23, e14275.
- Marquez-Exposito, L. , et al., MO338INCREASED INFLAMMATORY RESPONSE IS A HALLMARK OF AGE-RELATED AGGRAVATION OF EXPERIMENTAL AKI. Nephrology Dialysis Transplantation, 2021. 36(Supplement_1).
- Cui, S. , et al., Reduction in Renal Interstitial Fibrosis in Aged Male Mice by Intestinal Microbiota Rejuvenation. Gerontology, 2024, 70, 1161-1170.
- Kim, M.G. , et al., Altered gut microbiome plays an important role in AKI to CKD transition in aged mice. Front Med (Lausanne), 2023, 10, 1238960.
- Bosco, N. and M. Noti, The aging gut microbiome and its impact on host immunity. Genes Immun, 2021, 22(5-6), 289-303.
- Duong, S., T. Patel, and F. Chang, Dementia: What pharmacists need to know. Can Pharm J (Ott), 2017, 150, 118-129.
- Ragonnaud, E. and A. Biragyn, Gut microbiota as the key controllers of "healthy" aging of elderly people. Immun Ageing, 2021, 18, 2.
- Hill, C. , et al., Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol, 2014, 11, 506-14.
- Miri, S.T. , et al., The impact of Lactobacillus and Bifidobacterium probiotic cocktail on modulation of gene expression of gap junctions dysregulated by intestinal pathogens. Arch Microbiol, 2022, 204, 417.
- Al-Dhabi, N.A. , et al., Probiotic and Antioxidant Potential of Lactobacillus reuteriLR12 and Lactobacillus lactisLL10 Isolated from Pineapple Puree and Quality Analysis of Pineapple-Flavored Goat Milk Yoghurt during Storage. Microorganisms, 2020, 8(10).
- Cannarella, L.A.T. , et al., Mixture of probiotics reduces inflammatory biomarkers and improves the oxidative/nitrosative profile in people with rheumatoid arthritis. Nutrition, 2021, 89, 111282.
- Kumaree, K.K. , et al., Lactobacillus paracasei HII01 enhances lifespan and promotes neuroprotection in Caenorhabditis elegans. Sci Rep, 2023, 13, 16707.
- Boonyarut Ladda, P.T. , Poolpol Padungchaichot, Wisuit Pradidarcheep, Anongnard Kasorn, Malai Taweechotipatr, Anti-inflammatory effect of probiotic Lactobacillus paracasei MSMC39-1 on alcohol-induced hepatitis in rats. Vol. Volume: 11. 2021: Issue: 4. 046-056.
- Ladda, B. , et al., Anti-Inflammatory and Gut Microbiota Modulating Effects of Probiotic Lactobacillus paracasei MSMC39-1 on Dextran Sulfate Sodium-Induced Colitis in Rats. Nutrients, 2023, 15(6).
- Kujoth, G.C. , et al., Mitochondrial DNA mutations, oxidative stress, and apoptosis in mammalian aging. Science, 2005, 309, 481-4.
- Liu, X. , et al., Intestinal homeostasis in the gut-lung-kidney axis: a prospective therapeutic target in immune-related chronic kidney diseases. Front Immunol, 2023, 14, 1266792.
- Jaul, E. and J. Barron, Age-Related Diseases and Clinical and Public Health Implications for the 85 Years Old and Over Population. Front Public Health, 2017, 5, 335.
- Kwok, K.O. , et al., Effects of Probiotic Intervention on Markers of Inflammation and Health Outcomes in Women of Reproductive Age and Their Children. Front Nutr, 2022, 9, 889040.
- Flurkey, K., J. M. Currer, and D.E. Harrison, Chapter 20 - Mouse Models in Aging Research, in The Mouse in Biomedical Research (Second Edition), J.G. Fox, et al., Editors. 2007, Academic Press: Burlington. p. 637-672.
- Kim, H.J. , et al., Lacticaseibacillus paracsei HY7207 Alleviates Hepatic Steatosis, Inflammation, and Liver Fibrosis in Mice with Non-Alcoholic Fatty Liver Disease. Int J Mol Sci, 2024, 25(18).
- Rapa, S.F. , et al., Inflammation and Oxidative Stress in Chronic Kidney Disease-Potential Therapeutic Role of Minerals, Vitamins and Plant-Derived Metabolites. Int J Mol Sci, 2019, 21(1).
- Wagner, S. , et al., Probiotic Intake and Inflammation in Patients With Chronic Kidney Disease: An Analysis of the CKD-REIN Cohort. Front Nutr, 2022, 9, 772596.
- Huang, H. , et al., Preventive Effects of Lactobacillus Mixture against Chronic Kidney Disease Progression through Enhancement of Beneficial Bacteria and Downregulation of Gut-Derived Uremic Toxins. J Agric Food Chem, 2021, 69, 7353-7366.
- Kim, W.-K. , et al., Administration of Lactobacillus paracasei strains improves immunomodulation and changes the composition of gut microbiota leading to improvement of colitis in mice. Journal of Functional Foods, 2019, 52, 565-575.
- Simeoli, R. , et al., Preventive and therapeutic effects of Lactobacillus paracasei B21060-based synbiotic treatment on gut inflammation and barrier integrity in colitic mice. J Nutr, 2015, 145, 1202-10.
- Cao, J. , et al., Lactobacillus paracasei X11 Ameliorates Hyperuricemia and Modulates Gut Microbiota in Mice. Front Immunol, 2022, 13, 940228.
- Wang, D. , et al., Loss of legumain induces premature senescence and mediates aging-related renal fibrosis. Aging Cell, 2022, 21, e13574.
- Ha, S. , et al., Diminished Tubule Epithelial Farnesoid X Receptor Expression Exacerbates Inflammation and Fibrosis Response in Aged Rat Kidney. J Gerontol A Biol Sci Med Sci, 2023, 78, 60-68.
- Hajam, Y.A. , et al., Oxidative Stress in Human Pathology and Aging: Molecular Mechanisms and Perspectives. Cells, 2022, 11(3).
- Oguntoye, M.A., O. A. Oridupa, and O.O. Ezekiel, Phenolic, flavonoid, and beta-carotene contents of provitamin A cassava hydrolysate with free and encapsulated Lactobacillus rhamnosus GG improves antioxidant biomarkers in kidney, heart and liver of Wistar rats. J Food Sci, 2023, 88, 491-502.
- Zheng, H.J. , et al., Probiotics, prebiotics, and synbiotics for the improvement of metabolic profiles in patients with chronic kidney disease: A systematic review and meta-analysis of randomized controlled trials. Crit Rev Food Sci Nutr, 2021, 61, 577-598.
- Lin, W.Y. , et al., Probiotics and their Metabolites Reduce Oxidative Stress in Middle-Aged Mice. Curr Microbiol, 2022, 79, 104.
- Schroijen, M.A. , et al., The association of glucose metabolism and kidney function in middle-aged adults. Clin Kidney J, 2021, 14, 2383-2390.
- O'Toole, J.F. , et al., Decreased cytochrome c mediates an age-related decline of oxidative phosphorylation in rat kidney mitochondria. Biochem J, 2010, 427, 105-12.
- Miao, J. , et al., Wnt/beta-catenin/RAS signaling mediates age-related renal fibrosis and is associated with mitochondrial dysfunction. Aging Cell, 2019, 18, e13004.
- Wang, D. , et al., Senescent renal tubular epithelial cells activate fibroblasts by secreting Shh to promote the progression of diabetic kidney disease. Front Med (Lausanne), 2022, 9, 1018298.
- Bryant, J.D. , et al., Assessing Mitochondrial DNA Release into the Cytosol and Subsequent Activation of Innate Immune-related Pathways in Mammalian Cells. Curr Protoc, 2022, 2, e372.
Figure 1.
A condition medium generated from L. paracasei MSMC39-1 protects damaged kidney epithelial cells. (A) Schematic representation of treatment strategy of L. paracasei MSMC39-1-derived condition medium in kidney epithelial cell. (B) Representative microscopic images and (C) relative cell proliferation of Vero cells determined by WST-8 assay. The cells were co-treated with high glucose 25.5 mM and L. paracasei MSMC39-1-derived condition medium (HG + MSMC39-1 CM) for 48 hours followed by low glucose media for another 144 hours. De Man, Rogosa, and Sharpe (MRS) broth were used as a vehicle control (HG + MRS). Data are means ± SD (n = 4). Statistical analysis was two-way ANOVA, **p < 0.01 compared to HG + MRS treated cells. Data are representative of three reproducible and independent experiments.
Figure 1.
A condition medium generated from L. paracasei MSMC39-1 protects damaged kidney epithelial cells. (A) Schematic representation of treatment strategy of L. paracasei MSMC39-1-derived condition medium in kidney epithelial cell. (B) Representative microscopic images and (C) relative cell proliferation of Vero cells determined by WST-8 assay. The cells were co-treated with high glucose 25.5 mM and L. paracasei MSMC39-1-derived condition medium (HG + MSMC39-1 CM) for 48 hours followed by low glucose media for another 144 hours. De Man, Rogosa, and Sharpe (MRS) broth were used as a vehicle control (HG + MRS). Data are means ± SD (n = 4). Statistical analysis was two-way ANOVA, **p < 0.01 compared to HG + MRS treated cells. Data are representative of three reproducible and independent experiments.
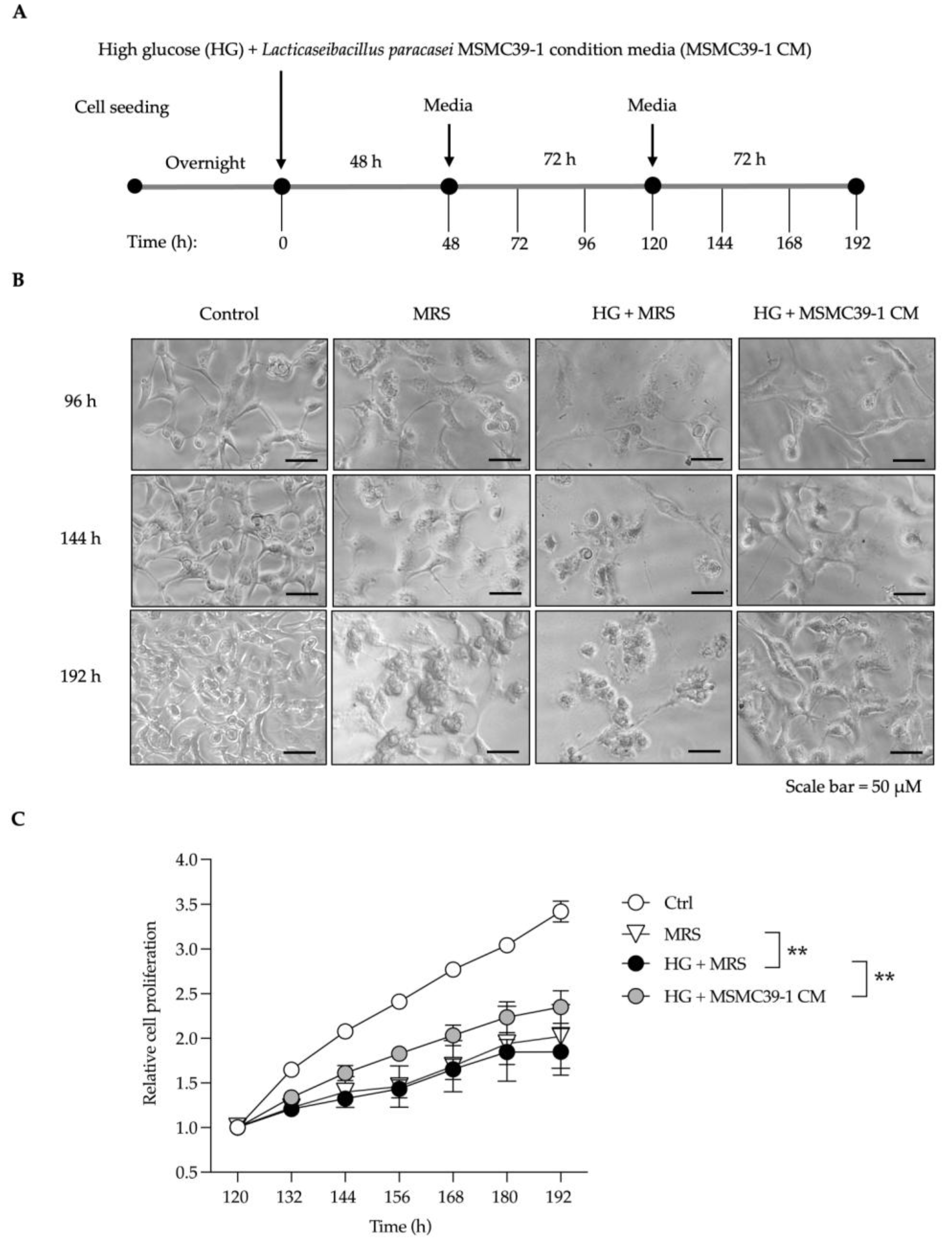
Figure 2.
Effects of L. paracasei MSMC39-1 on kidney fibrosis and injury in aged mice. (A) Schematic representation of in vivo treatment strategy of L. paracasei MSMC39-1 in aged mice. (B) Representative microscopic images of hematoxylin and eosin (H&E) and Masson’s trichrome staining of mice administered with L. paracasei MSMC39-1. (C) Quantification of masson's trichrome staining fibrotic area (n = 15) and glomerulus area (n = 20) of mice administered with L. paracasei MSMC39-1 for 4 months (AM). Young mice (6 months of age) were included as a control group (YC). Data are means ± SD (n = 5 mice, each containing 3 slices for masson's trichrome staining fibrotic area quantification and 4 glomeruli for glomerulus area quantification). Statistical analysis was one-way ANOVA followed by Dunnett's multiple comparisons test, *p < 0.05, **p < 0.01 compared to aged control (AC) group.
Figure 2.
Effects of L. paracasei MSMC39-1 on kidney fibrosis and injury in aged mice. (A) Schematic representation of in vivo treatment strategy of L. paracasei MSMC39-1 in aged mice. (B) Representative microscopic images of hematoxylin and eosin (H&E) and Masson’s trichrome staining of mice administered with L. paracasei MSMC39-1. (C) Quantification of masson's trichrome staining fibrotic area (n = 15) and glomerulus area (n = 20) of mice administered with L. paracasei MSMC39-1 for 4 months (AM). Young mice (6 months of age) were included as a control group (YC). Data are means ± SD (n = 5 mice, each containing 3 slices for masson's trichrome staining fibrotic area quantification and 4 glomeruli for glomerulus area quantification). Statistical analysis was one-way ANOVA followed by Dunnett's multiple comparisons test, *p < 0.05, **p < 0.01 compared to aged control (AC) group.
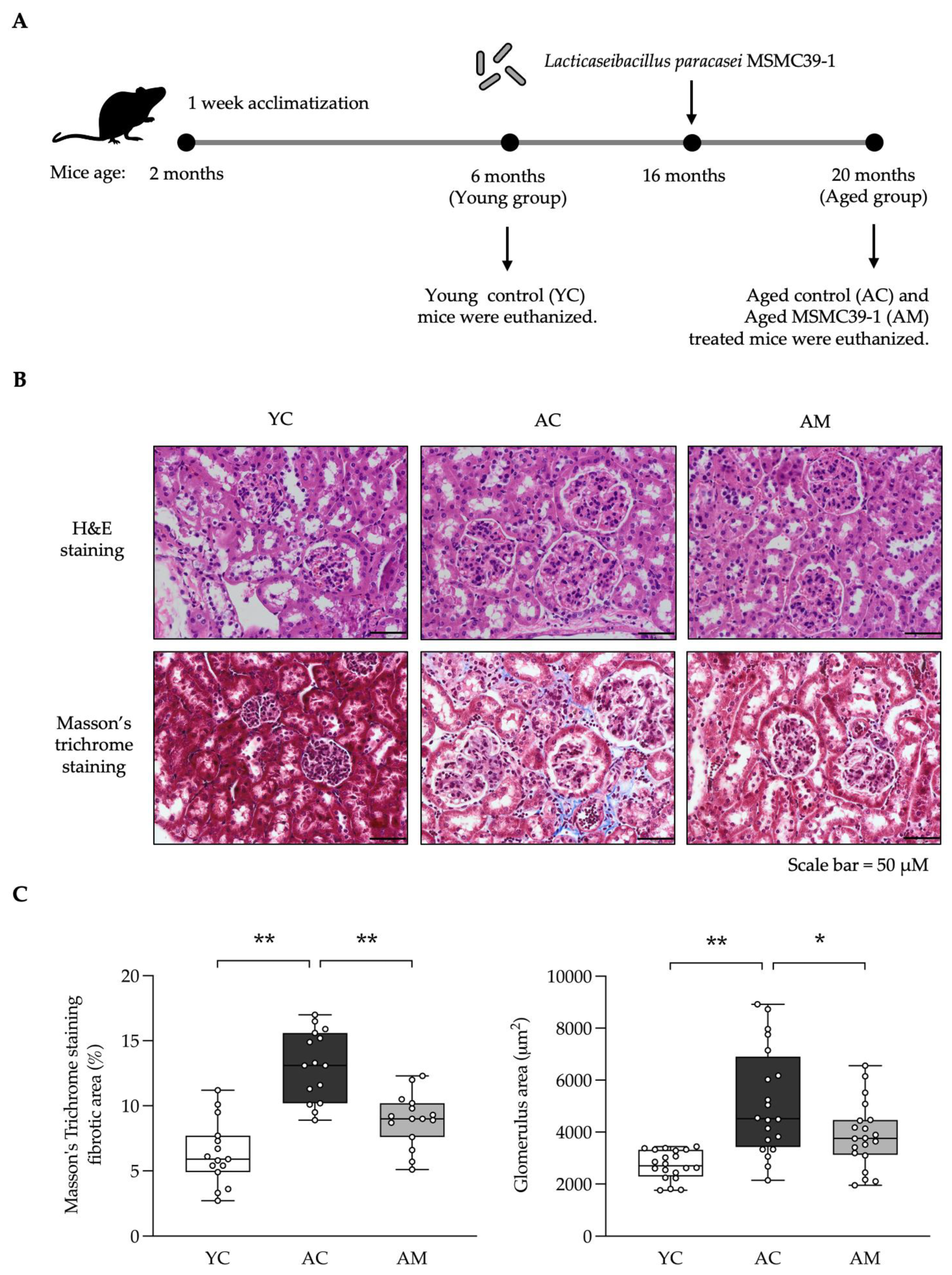
Figure 3.
Effects of L. paracasei MSMC39-1 on high glucose and oxidative stress in aging mice. (A) Serum blood glucose, (B) kidney malondialdehyde (MDA) levels, (C) kidney glutathione pe-roxidase (Gpx) levels and (D) kidney superoxide dismutase (SOD) levels in aged mice treated with L. paracasei MSMC39-1 for 4 months (AM). Young mice (6 months of age) were included as a control group (YC). Data are means ± SD (n = 5 mice). Statistical analysis was one-way ANOVA followed by Dunnett's multiple comparisons test, *p < 0.05, **p < 0.01 compared to aged control (AC) group.
Figure 3.
Effects of L. paracasei MSMC39-1 on high glucose and oxidative stress in aging mice. (A) Serum blood glucose, (B) kidney malondialdehyde (MDA) levels, (C) kidney glutathione pe-roxidase (Gpx) levels and (D) kidney superoxide dismutase (SOD) levels in aged mice treated with L. paracasei MSMC39-1 for 4 months (AM). Young mice (6 months of age) were included as a control group (YC). Data are means ± SD (n = 5 mice). Statistical analysis was one-way ANOVA followed by Dunnett's multiple comparisons test, *p < 0.05, **p < 0.01 compared to aged control (AC) group.
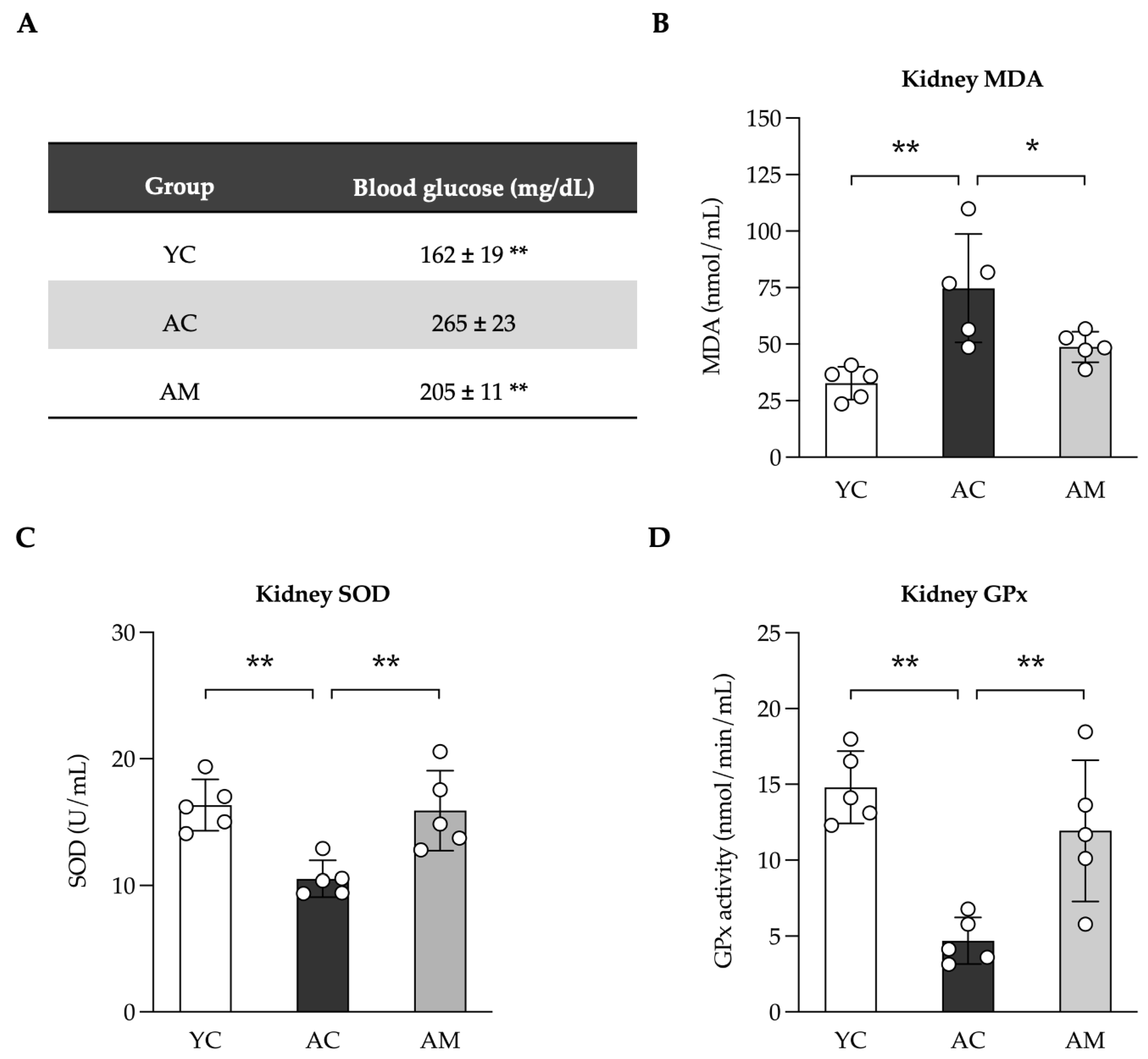
Figure 4.
Improvements of kidney mitochondria DNA (mtDNA) content in aged mice by L. paracasei MSMC39-1. (A) qPCR analysis of kidney mtND1 and (B) mtD-loop in aged mice treated with L. paracasei MSMC 39-1 for 4 months (AM). Young mice (6 months of age) were included as a control group (YC). Data are means ± SD (n = 5 mice). Statistical analysis was one-way ANOVA followed by Dunnett's multiple comparisons test, **p < 0.01 compared to aged control (AC) group.
Figure 4.
Improvements of kidney mitochondria DNA (mtDNA) content in aged mice by L. paracasei MSMC39-1. (A) qPCR analysis of kidney mtND1 and (B) mtD-loop in aged mice treated with L. paracasei MSMC 39-1 for 4 months (AM). Young mice (6 months of age) were included as a control group (YC). Data are means ± SD (n = 5 mice). Statistical analysis was one-way ANOVA followed by Dunnett's multiple comparisons test, **p < 0.01 compared to aged control (AC) group.
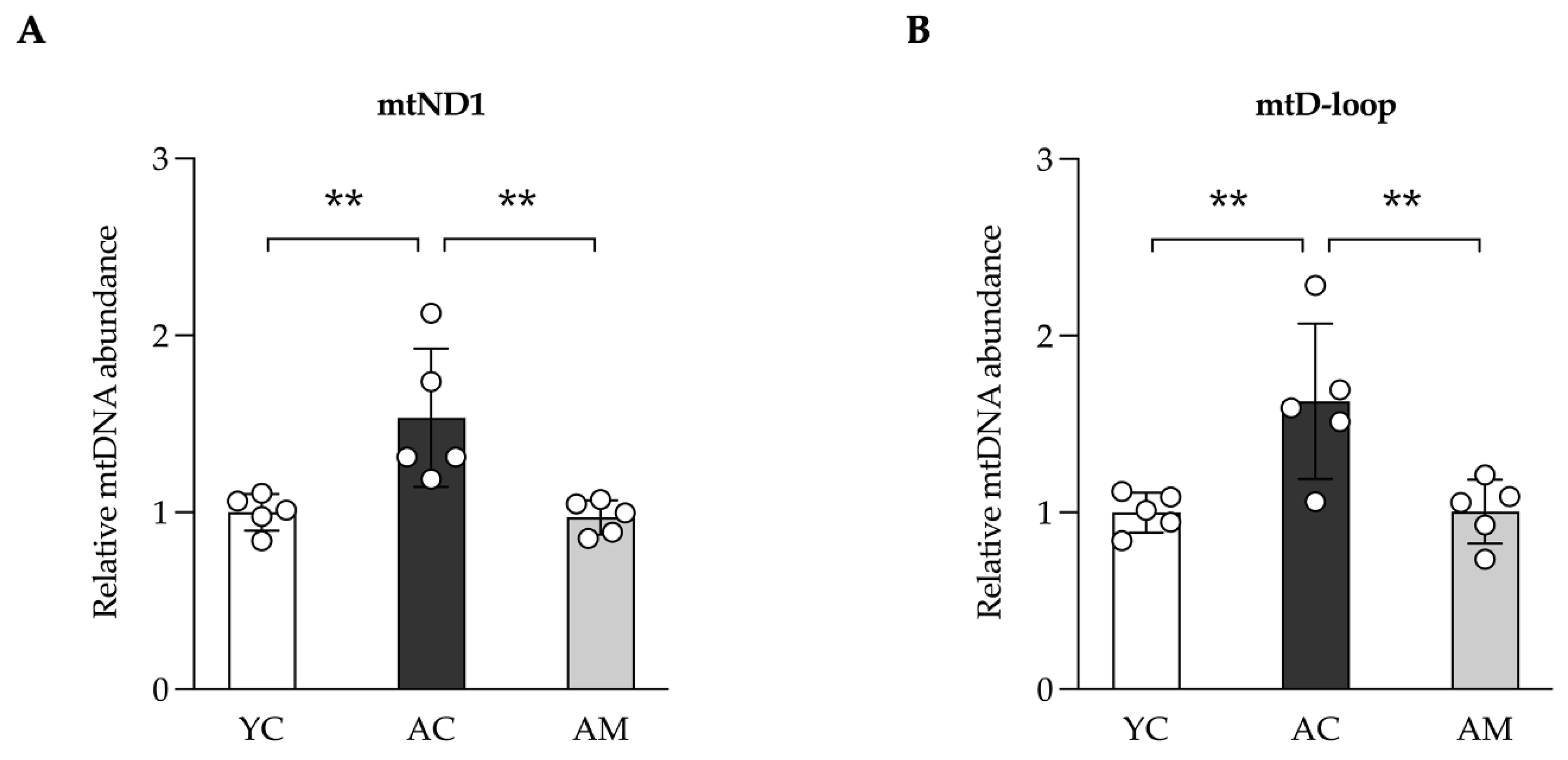
Figure 5.
Anti-inflammatory effects in kidney in aged mice administration of L. paracasei MSMC39-1. (A) Kidney levels of TNF-α and (B) IL-1β in aged mice treated with L. paracasei MSMC39-1 for 4 months (AM). Young mice (6 months of age) were included as a control group (YC). Data are means ± SD (n = 5 mice). Statistical analysis was one-way ANOVA followed by Dunnett's multiple comparisons test, **p < 0.01 compared to aged control (AC) group.
Figure 5.
Anti-inflammatory effects in kidney in aged mice administration of L. paracasei MSMC39-1. (A) Kidney levels of TNF-α and (B) IL-1β in aged mice treated with L. paracasei MSMC39-1 for 4 months (AM). Young mice (6 months of age) were included as a control group (YC). Data are means ± SD (n = 5 mice). Statistical analysis was one-way ANOVA followed by Dunnett's multiple comparisons test, **p < 0.01 compared to aged control (AC) group.
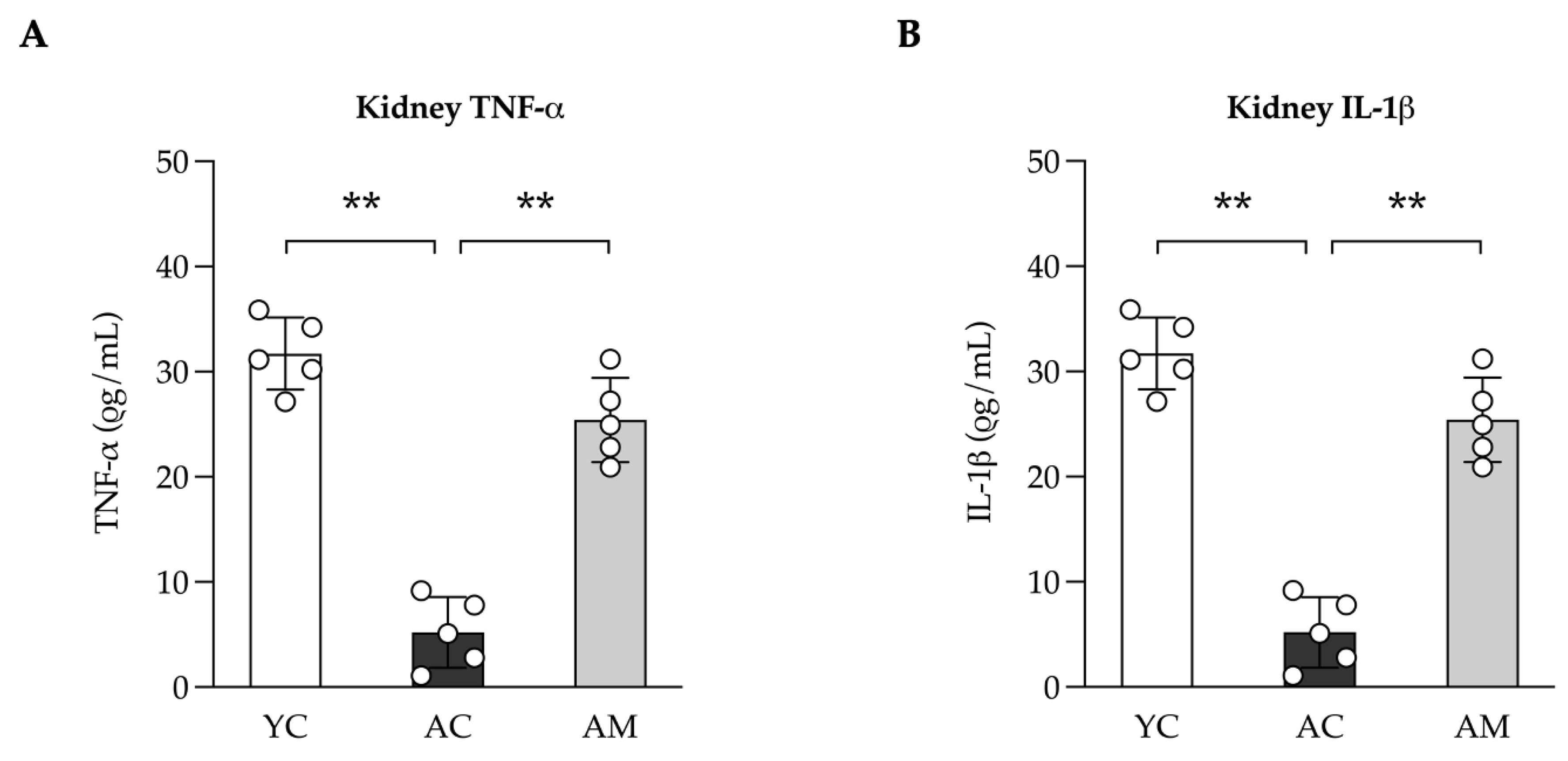
Figure 6.
Improvements of colitis symptoms in aged mice administered with L. paracasei MSMC39-1. (A) Representative microscopic images of hematoxylin and eosin (H&E) and (B) quantification of colitis manifestation in aged mice treated with L. paracasei MSMC39-1 for 4 months (AM). Young mice (6 months of age) were included as a control group (YC). Data are means ± SD (n = 5 mice, each containing 2 slices for the quantification). Statistical analysis was one-way ANOVA followed by Dunnett's multiple comparisons test, **p < 0.01 compared to aged control (AC) group.
Figure 6.
Improvements of colitis symptoms in aged mice administered with L. paracasei MSMC39-1. (A) Representative microscopic images of hematoxylin and eosin (H&E) and (B) quantification of colitis manifestation in aged mice treated with L. paracasei MSMC39-1 for 4 months (AM). Young mice (6 months of age) were included as a control group (YC). Data are means ± SD (n = 5 mice, each containing 2 slices for the quantification). Statistical analysis was one-way ANOVA followed by Dunnett's multiple comparisons test, **p < 0.01 compared to aged control (AC) group.
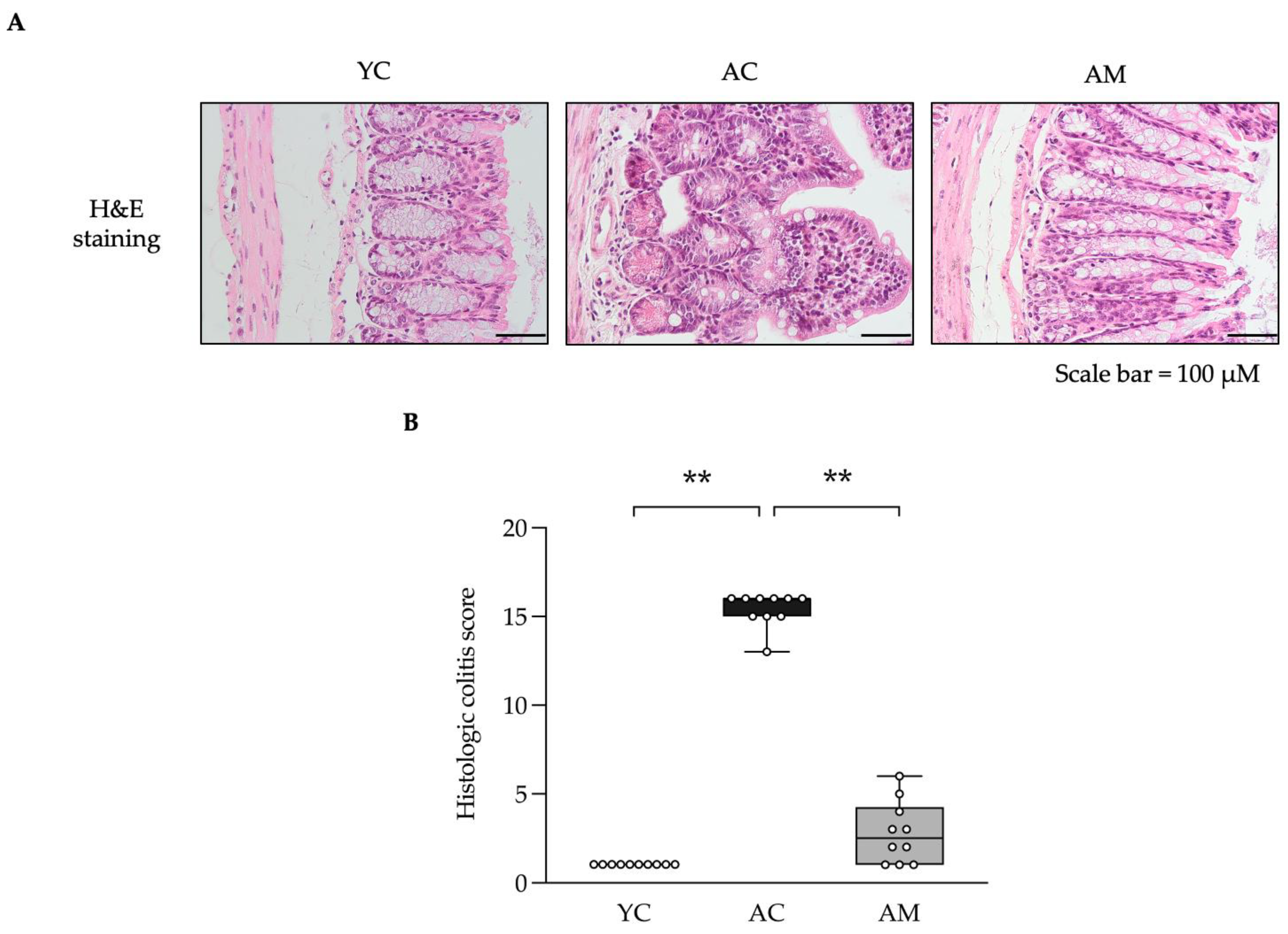

Table 1.
Histological grading of colitis.
| Feature graded | Grade | Description |
|---|---|---|
| Inflammation | 0 1 2 3 |
None Slight Moderate Severe |
| Extent | 0 1 2 3 |
None Mucosa Mucosa and submucosa Transmural |
| Regeneration | 4 3 2 1 0 |
No tissue repair Surface epithelium not intact Regeneration with crypt depletion Almost complete regeneration Complete regeneration or normal tissue |
| Crypt damage | 0 1 2 3 4 |
None Basal 1/3 damaged Basal 2/3 damaged Only surface epithelium intact Entire crypt and epithelium lost |
| Percent involvement | 1 2 3 4 |
1-25% 1-25% 26-50% 51-100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



