
Submitted:
05 May 2025
Posted:
06 May 2025
You are already at the latest version
Abstract
Tetrodotoxin (TTX), earlier known as a tropical paralytic neurotoxin from the pufferfish poisonings, has increasingly been occurring in edible marine species, including filter-feeding bivalves, from relatively cold marine waters of some European countries. The defined conditions that promote the production of TTX, its origin or the processes of its accumulation in seafood are still not clarified. Recent studies in temperate waters show, however, that the accumulation of quantifiable levels of TTX in bivalves appear to be influenced by seawater temperature (>15 C) which indicates a seasonal occurrence. Uncertainties still remain regarding how sea water temperature interacts with other climate and environmental factors or organisms in the marine ecosystem to result in detectable levels of TTX in shellfish. Knowledge of the occurrence and distribution of TTX in the marine environment where the edible bivalves grow is important for maintaining seafood safety as the toxin is heat-stabile and remains potent even after cooking. Therefore, in this study 264 bivalve samples collected in 2019 and 2021 from 17 sites along the Swedish west coast were analyzed with LC-MS/MS to search for TTX. The study explores the hypothesis of TTX presence in Swedish marine waters, outlines the sample screening strategy and objectives, and reports no evidence of TTX presence in Swedish bivalve shellfish (≥ 7,8 µg/kg) based on the analyzed samples and the time periods in which the studied samples were collected.
Keywords:
Tetrodotoxin (TTX)
; bivalve shellfish
; screening
; Sweden
1. Introduction
Tetrodotoxin (TTX), is a heat-stable acute paralytic neurotoxin originally mostly known from poisonings caused by pufferfish (Tetraodontidae family) or local intoxications in Southeast Asia after consumption of seafood from areas associated with high-risk of TTX contamination [1,2,3]. TTX has been found not only in marine species but also in various terrestrial animals, as was earlier reviewed [4,5,6]. Yet, the consumption of edible marine species is the most common reason for human intoxication [1,7]. Following the ingestion of a TTX-contaminated meal a clinical picture of a poisoning in human can include a number of symptoms that appear from within 20 minutes up to 4 hours [8,9]. The symptoms are a consequence of a non-functioning neurotransmission between the nerve cells, which might result in numbness (in lips and mouth that can spread to face and extremities), gastrointestinal issues, slurred speech and muscle weakness that at higher doses progresses into paralysis, respiratory distress and sometimes fatality [7]. Although TTX became known as a pufferfish toxin, studies that later showed its absence in pufferfish specimens living in aquariums [10] or in other representative TTX-carrying organisms, like amphibians in captivity [11], raised the debate of the biogenic origin of TTX including its biosynthesis that is yet to be elucidated [6,9] The dynamics of TTX in ecosystems is complex and the toxins obvious presence in organisms has so far appeared to be based on an interplay between different factors, including surrounding habitat and environmental influences [9,12,13]. Also, the process of TTX accumulation has been complicated to clarify as the sources of TTX presented in studies so far are multifaceted. At least in the case of marine organisms, and unlike other marine biotoxins, TTX has been primarily associated with production by bacterial species [14,15], including Vibrio, Bacillus, Pseudomonas and Actinobacteria [6,14,15,16,17,18], but also phytoplankton [19,20], presence of nemertean species (ribbon worms) [12,21,22] and especially the links between the toxin occurrence, bacterial presence and climate change with emphasis on elevated seawater temperatures [23,24,25,26].
In Europe, the TTX has been a non-issue until almost two decades ago when the first case of human poisoning was reported in 2007 from Spain after consumption of a gastropod Charonia lampas lampas originating from Portugal [27,28,29]. This poisoning case, as evidence of emerging TTX in European marine waters, could likely correlate to the contemporary discovery on the presence of the pufferfish Lagocephalus sceleratus in the Mediterranean Sea reported in 2005, where its entrance (as a toxin-bearing species) was made possible through the Suez Canal [30,31,32]. The first detection of TTX in European bivalve molluscs was then reported in 2015 [16,19], which raised more concerns about seafood safety resulting in a scientific opinion on the risks for public health related to the presence of TTX and its analogues in marine bivalves and gastropods (above a concentration of 44 µg/kg) from the European Food Safety Authority (EFSA) [33]. Since that, there has been growing evidence of TTX presence in edible marine species sourced from European marine waters as well as in other marine organisms sought as seafood [7,34,35,36]. However, there are no European regulatory requirements for monitoring the presence of TTX in bivalves yet. The responsibility for developing analytical methods for a reliable detection of TTX, as well as regular monitoring in seafood, remains a matter for each of the EU-member states.
So far, the threat from TTX has been deemed negligible in Scandinavian marine waters. Anyhow, there is evidence that TTX-contamination in seafood is spreading northwards. Several studies on the occurrence of TTX and its analogues in bivalve molluscs along the west-European coasts have been published during the recent years, as in northern France [26], The Netherlands [34], in southern England [16,22,23,37] and Scotland (North Sea) being at the closest geographical distance to Sweden where the presence of TTX has been dated to 2014 [37]. Almost a decade later, it is reasonable to assume the presence of TTX in Swedish bivalve species, which rises the concerns about health risks for consumers prompting the research on its discovery and handling.
The aim of this study was to search for the presence of TTX and its analogues in Swedish bivalve shellfish as a preventive measure in shellfish safety and, upon the discovery of TTX, respond to the call of EFSA on the need of more occurrence data from European waters in different EU Member States.
2. Results and Discussion
Proximity to the countries where TTX has been detected in shellfish over a decade ago and the different environmental conditions in Swedish marine waters, including the risk of bacterial production of TTX that increases with warmer climate, led to the hypothesis that TTX is present in Swedish bivalves at a detectable level. Given the complex origin of TTX and the interplay of various ecological factors in its emergence, conducting the present screening study was considered relevant to thereby confirm or reject this hypothesis, at present.
Sampling sites for the study were selected, as far as possible, according to the risk profile proposed by Turner et al. [37]. The samples analyzed for TTX presence consisted of mussels (Mytilus edulis), Pacific oysters (Magallana gigas), native oysters (Ostrea edulis) and cockles (Cerastoderma edule). Most of these samples were official control (OC) samples from routine monitoring collected from classified production areas along the western Swedish archipelago (Bohuskusten), Figure 1, during warmer months April-September in 2019 and April-September in 2021. Sampling was limited to this part of the Swedish coastline, as it represents the only region in Sweden where production of live bivalve molluscs (LBM) occurs.
Additional five samples of Magallana gigas were collected from a selected area characterized by shallow warm water. When the presence of Prorocentrum cordatum was detected in the water column in September 2022 yet another group of mussel samples (Mytilus edulis) was collected, since earlier studies have assumed a linkage between the presence of Prorocentrum spp. and detectable levels of TTX [19,38]. Figure 2 shows the number of samples and proportion of representing species in each of the sampling seasons.
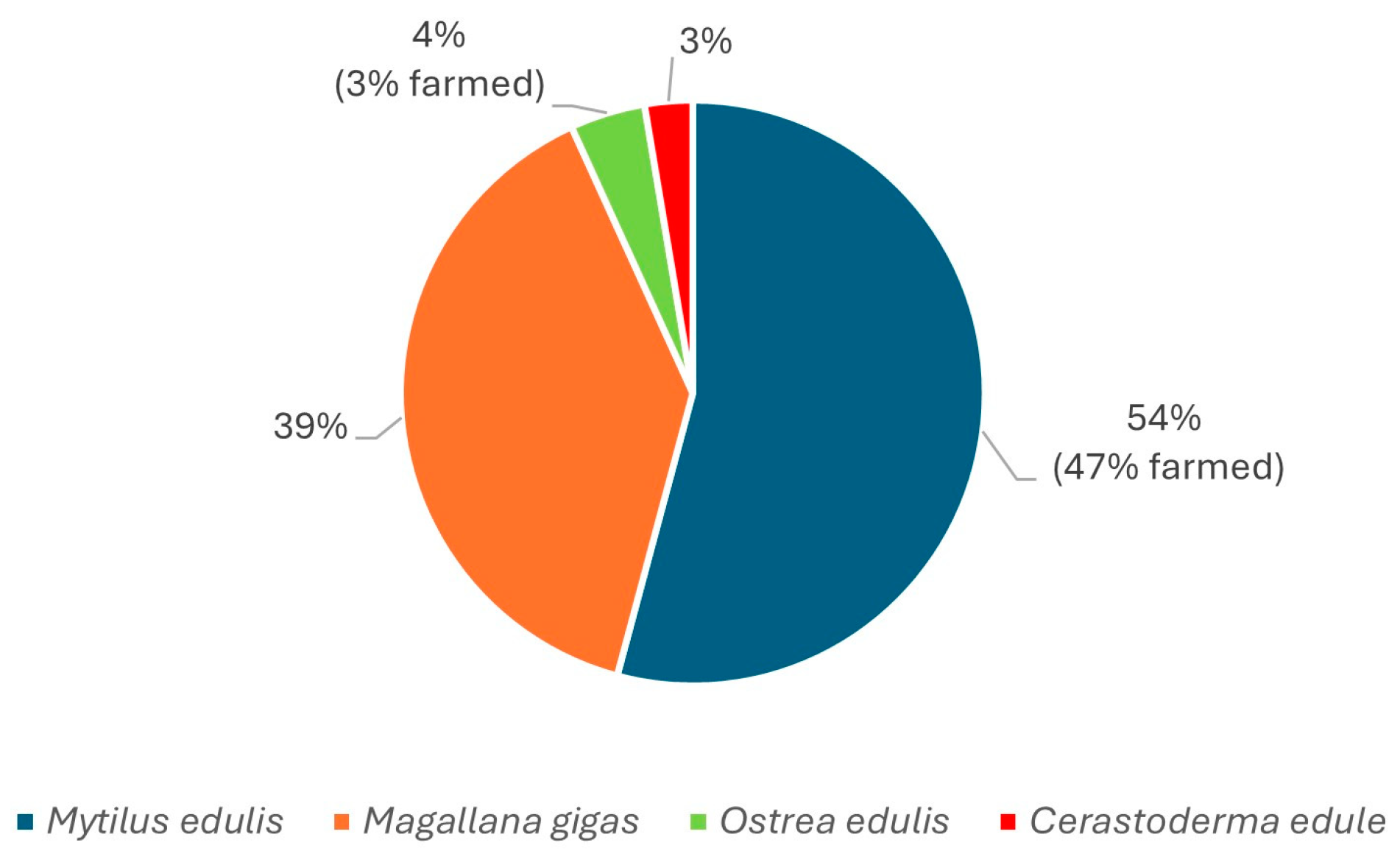
The sampling strategy did not intend the collection of representative species for each monitoring site, rather, the goal was to broadly search across all species during the two warm seasons. Mussels (Mytilus edulis) is the slightly overrepresented species among the analyzed samples Figure 3, which also reflects its dominance among the species harvested for human consumption in Sweden.
None of the tested samples were found to contain TTX, or the analogues for which the standards were available, above the limit of detection (LOD) for these toxins in the method, Table S1 [39]. Some samples that did not meet the specificity criteria for confirmation of TTX presence, yet exhibiting interesting signals (either through the presence of one of the two confirming fragment ions or a signal within the reasonable retention time window of a TTX-analogue that was included in the acquisition method although no standards were available) were sent to the national reference laboratory for marine biotoxins in The Netherlands for analysis using high-resolution mass spectrometry (HRMS). This approach could possibly clarify whether the signals could be from an isobaric toxin analogue to TTX with a similar elution time that could not be detected with the dictating triple quadrupole principle. These samples were also found to be TTX-negative.
As the purpose was primarily to conduct a first screening study of a potential presence of TTX in Swedish bivalves no specific hydrographic parameters (salinity and depth) or seawater temperatures were recorded during sampling. Whether there is a connection between the absence of TTX in samples tested in this study and unfavorable conditions for its production at the time point of sampling at their corresponding sampling sites is therefore challenging to discuss. Factors influencing the temporal variability of TTX accumulation in bivalve molluscs appear to be difficult to reach a definitive conclusion about, based on data from previous European findings [13].
However, several of the proposed sources of TTX, and the conditions in marine environments discussed in earlier studies as possible links or influencing factors that might interact in or trigger TTX production, are also present in Swedish marine waters:
- The relationship between TTX and Vibrio bacteria is particularly interesting given the strong correlation between bacterial prevalence, TTX occurrence and warmer climate, as highlighted in previous research [14,40,41,42,43,44,45]. Coastal waters and brackish areas with moderate salinity, typically dominating in Scandinavian marine environments, favors Vibrio populations [46,47,48]. However, high abundance of Vibrio bacteria has also been reported in more saline waters of the Skagerrak Sea, especially in warm summer days [49]. One notable Vibrio outbreak in the Baltic Sea occurred during the warm summer in 2014, when the sea surface temperatures became unusually high [50]. This Vibrio outbreak temporally corresponded with a TTX-positive mussel sample (26 µg/kg) in Scottland (North Sea) [37], assumptively indicating presence of TTX in Swedish marine waters (no samples were tested for TTX presence in Sweden during this period).
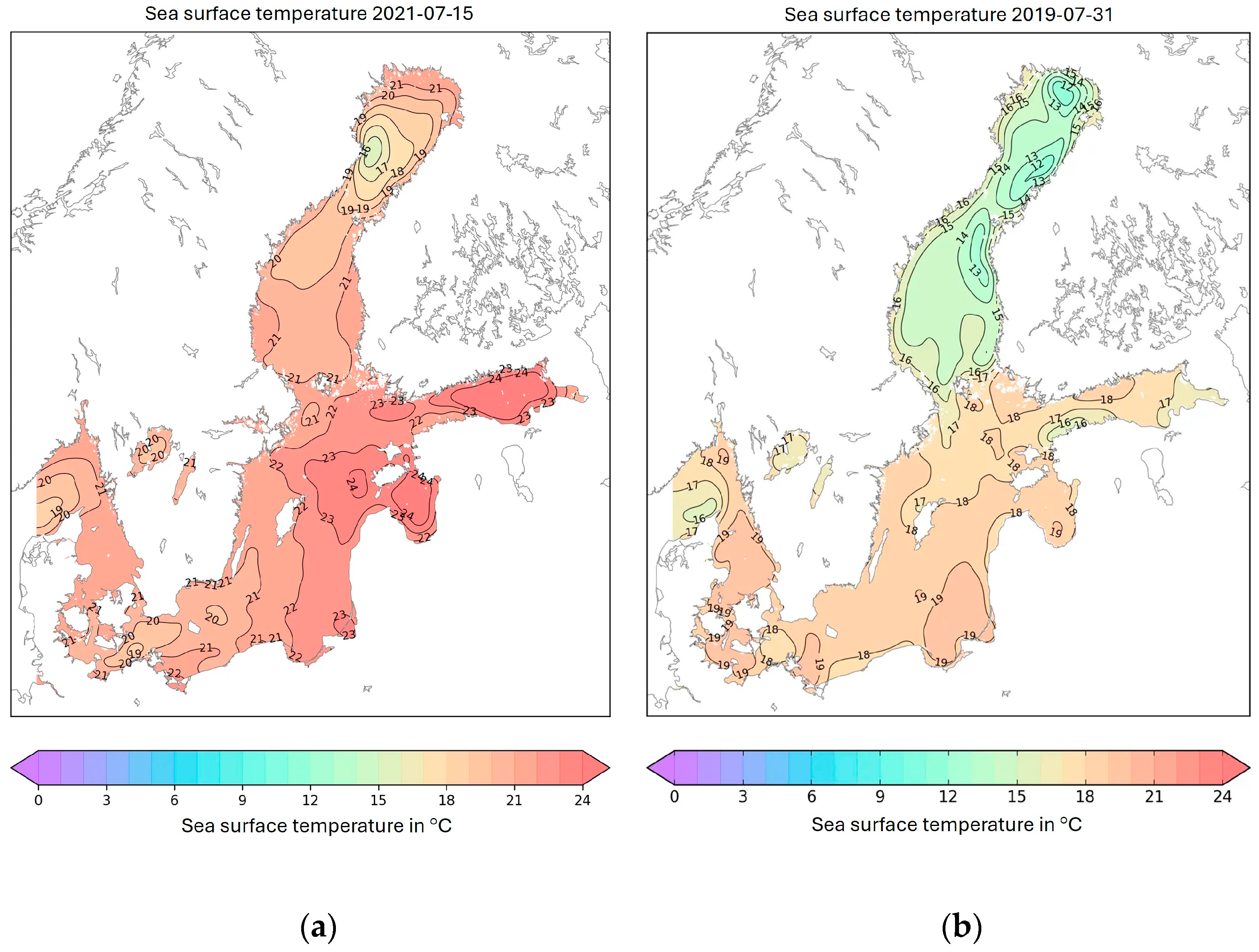
- Furthermore, the assumption of TTX presence in Swedish marine environment is supported with the knowledge that sea surface temperature seems to be one of the ultimate criteria earlier reported for the occurrence of TTX [23,34,37]. The threshold of 15 °C was suggested to indicate a limit above which TTX is more likely to occur in shellfish. Certainly, surface temperatures of the seawater around Sweden can reach well above that threshold during the summer season, as exemplified for the sampling seasons of this study in Figure 4, and the summer of 2014 described above.
This notion could be further strengthened by the climate benefits from the Gulf Stream that in combination with the climatic warming of coastal waters [53,54] interact and could create favorable conditions for the emergence of TTX even at these, normally cold areas when compared to similar latitudes.
- Introduction of invasive species, like the pufferfish or the spread of the invasive ribbon worm Cephalotrix simula to southern and northern Europe in the last decade, are strongly associated with the occurrence of TTX in European marine waters [21,55,56,57]. To the best of our knowledge, the latest biodiversity survey concerning nemerteans along the Swedish west coast was conducted in 2007 [58]. Various nemertean species have been studied in Sweden since then [59] but the occurrence of C. simula has not been reported yet. It is, however, reasonable to assume that the species could have become introduced into Swedish marine waters. One of the main vectors for the introduction of marine invasive species globally is ballast water of ships by which aquatic organisms are transported to new environments. Aspects on its constantly increasing impact on the existing ecosystems have been addressed elsewhere [60,61,62,63]. A majority of Sweden’s blue mussel production is located within 60 km north of the Port of Gothenburg, which is the largest port in Scandinavia.
- With the above outlined conditions 1) – 3) one could suggest that there is no major difference in the general conditions for TTX occurrence in the marine environment, compared to previous reports on TTX presence in northern Europe, which supports our hypothesis while the results of this screening study could not confirm it.
3. Materials and Methods
3.1. Standards, Reagents and Chemicals
All the used standards, reagents and chemicals were as previously described by Patria et. al. [39]. Briefly: certified reference material (CRM) of TTX and the analogues (in µmol/L concentrations 78.6, 9.9 of TTX and 4,9-anhydro TTX, and 2.50 of 11-deoxyTTX as non-certified, respectively) were obtained from Laboratorio CIFGA (Lugo, Spain). Stock standard solutions were prepared by diluting the CRM 10 times in Milli-Q® water, from which the LC-MS/MS calibration standards were prepared in 80% acetonitrile with 0.25% acetic acid. Amorphous graphitized polymer carbon Supelco ENVI-Carb 250 mg/3 mL cartridges (Sigma-Aldrich, St. Louis, MO, USA) were used for clean-up of the raw shellfish sample extracts. Instrument solvents were LC-MS-grade where possible, acetonitrile (ACN, Fisher Scientific, Loughborough, UK), methanol and formic acid 98–100% (Merck, Darmstadt, Germany). Sample preparation reagents were of HPLC grade.
3.2. Sampling and Sample Preparation
Shellfish samples collected in warmer months were only considered relevant for screening purposes as the risk profile described in previous studies from northern Europe showed that temperature plays a key role for TTX findings. 264 bivalve samples (Mytilus edulis, Magallana gigas, Ostrea edulis and Cerastoderma edule), which according to the sampling procedure for regulatory monitoring of marine biotoxins all included at least 15 individuals, were collected within the period April-September in 2019 and 2021. All the 17 selected sampling sites shown in Figure 1 were marine areas along the Swedish west coast (salinity data not shown). After passing the official control of marine biotoxins monitoring in Sweden, samples selected for analysis in this study were stored homogenized and frozen at −20 °C. Additional samples (Mytilus edulis) were collected in September 2022 from a site where Prorocentrum cordatum was present in the water column surrounding the bivalves. The number of samples tested per month of each season and the species represented in the sampling are shown in Figure 2. The number of samples per each of the 17 sampling sites, including the proportion (in %) of farmed or wild per species are shown in Figure 3.
3.3. Screening Analysis
All samples were prepared, extracted and analyzed for the presence of TTX following the method published by Patria et al. [39] without modifications, except that the multiple-reaction-monitoring (MRM) method comprised only the precursor and fragment ions of TTX and the analogues in the ion acquisition (Table S2). Samples resulting in non-specific although interesting signals within the expected retention time window were further investigated with HRMS to exclude or verify a possible presence of less known TTX analogues.
4. Conclusions
This study is the first to perform a large screening of a potential occurrence of TTX in bivalve molluscs collected along the coastal areas of the western Swedish archipelago.
The study was conducted using an in-house validated HILIC-MS/MS method [39], an analytical approach recommended by EFSA for the acquisition of occurrence data [33,36,64]. Based on analysis of 264 samples, mostly oysters and mussels from 17 harvesting sites during two warm seasons in 2019 and 2021, this study provides no evidence of TTX presence in Swedish bivalve shellfish higher than the LOD of the applied method (≥7,8 µg/kg). At present, as per these results, the risk to consumers of Swedish bivalves appears to be negligible. Nevertheless, the potential future presence of TTX in Swedish marine waters and bivalves should not be overlooked. Global warming and environmental changes may create favorable conditions for TTX production to prosper locally along a coast, thereby being represented in a low percentage of all bivalve shellfish, as previous studies have indicated [13,23,25]. This section is not mandatory but can be added to the manuscript if the discussion is unusually long or complex.
Further research on Swedish bivalves is needed to increase the chance of detecting any possible presence of TTX and thereby prevent human intoxications, whether it would occur occasionally in specific locations described as “hotspots” [42,65,66], or at low concentrations. This is particularly important to highlight considering the occurrence of recreational shellfish harvesting during summers, as well as informal advice and trends to collect the easily accessible invasive oyster species Magallana gigas from shallow waters, thereby contributing in reducing its spread. Furthermore, the niche aquaculture in Sweden does not afford the research on their product safety, while the routine analysis using non-LC-MS/MS methods lacks the detection capability for TTX within the regulated monitoring of paralytic shellfish toxins (PSTs). A more effective strategy to discover a potential TTX-contamination in Swedish shellfish could be a continuous screening plan that should focus on the sites with the environmental features known to promote TTX accumulation (shallow water) during the warm season and with a higher sampling frequency (once-twice per week) over a period of several years.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Supplementary Material, Table S1: LOD (limit of detection) and LOQ (limit of quantification) levels of toxins in the HILIC-MS/MS method applied in the screening study, Table S2: Settings of the LC-MS/MS parameters.
Author Contributions
Conceptualization, A.Z.M.; methodology, A.Z.M. and H.P.; sampling, M.P. and E.R.; formal analysis, S.D., M.K. and A.Z.M.; investigation, S.D., M.K. and A.Z.M.; resources, A.Z.M., J.B., and H.P.; writing—original draft preparation, A.Z.M.; writing—review and editing, S.D., M.P., E.R., M.K., J.B., H.P.; visualization, M.P., E.R. and A.Z.M.; funding acquisition, A.Z.M., J.B. and H.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the Swedish Foundation for Strategic Research (SSF), grant number SM19-0005.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Guardone, L.; Maneschi, A.; Meucci, V.; Gasperetti, L.; Nucera, D.; Armani, A. A Global Retrospective Study on Human Cases of Tetrodotoxin (TTX) Poisoning after Seafood Consumption. Food Reviews International 2019, 36, 1–23. [Google Scholar] [CrossRef]
- Chowdhury, F.R.; Nazmul Ahasan, H.A.; Mamunur Rashid, A.K.; Al Mamun, A.; Khaliduzzaman, S.M. Tetrodotoxin poisoning: a clinical analysis, role of neostigmine and short-term outcome of 53 cases. Singapore Med J 2007, 48, 830–833. [Google Scholar]
- Lee, K.J.; Ha, K.S.; Jung, Y.J.; Mok, J.S.; Son, K.T.; Lee, H.C.; Kim, J.H. Paralytic shellfish toxins (PSTs) and tetrodotoxin (TTX) of Korean pufferfish. Fisheries and Aquatic Sciences 2021, 24, 360–369. [Google Scholar] [CrossRef]
- Bane, V.; Lehane, M.; Dikshit, M.; O'Riordan, A.; Furey, A. Tetrodotoxin: chemistry, toxicity, source, distribution and detection. Toxins (Basel) 2014, 6, 693–755. [Google Scholar] [CrossRef]
- Melnikova, D.I.; Magarlamov, T.Y. An Overview of the Anatomical Distribution of Tetrodotoxin in Animals. Toxins 2022, 14, 576. [Google Scholar] [CrossRef] [PubMed]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the origins and biosynthesis of tetrodotoxin. Aquat Toxicol 2011, 104, 61–72. [Google Scholar] [CrossRef]
- Katikou, P.; Gokbulut, C.; Kosker, A.R.; Campàs, M.; Ozogul, F. An Updated Review of Tetrodotoxin and Its Peculiarities. Marine Drugs 2022, 20, 47. [Google Scholar] [CrossRef]
- Bucciarelli, G.M.; Lechner, M.; Fontes, A.; Kats, L.B.; Eisthen, H.L.; Shaffer, H.B. From Poison to Promise: The Evolution of Tetrodotoxin and Its Potential as a Therapeutic. Toxins 2021, 13, 517. [Google Scholar] [CrossRef]
- Zhang, X.; Qiao, K.; Cui, R.; Xu, M.; Cai, S.; Huang, Q.; Liu, Z. Tetrodotoxin: The State-of-the-Art Progress in Characterization, Detection, Biosynthesis, and Transport Enrichment. Marine Drugs 2024, 22, 531. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O. Tetrodotoxin--distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Gilhen, J.; Russell, R.W.; Krysko, K.L.; Melaun, C.; Kurz, A.; Kauferstein, S.; Kordis, D.; Mebs, D. Variability of tetrodotoxin and of its analogues in the red-spotted newt, Notophthalmus viridescens (Amphibia: Urodela: Salamandridae). Toxicon 2012, 59, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Malykin, G.V.; Velansky, P.V.; Magarlamov, T.Y. Levels and Profile of Tetrodotoxins in Spawning Cephalothrix mokievskii (Palaeonemertea, Nemertea): Assessing the Potential Toxic Pressure on Marine Ecosystems. Toxins (Basel) 2025, 17. [Google Scholar] [CrossRef]
- Varini, C.; Manganelli, M.; Scardala, S.; Antonelli, P.; Losasso, C.; Testai, E. An Update of Tetrodotoxins Toxicity and Risk Assessment Associated to Contaminated Seafood Consumption in Europe: A Systematic Review. Toxins 2025, 17, 76. [Google Scholar] [CrossRef]
- Pratheepa, V.; Vasconcelos, V. Microbial diversity associated with tetrodotoxin production in marine organisms. Environ Toxicol Pharmacol 2013, 36, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Magarlamov, T.Y.; Melnikova, D.I.; Chernyshev, A.V. Tetrodotoxin-Producing Bacteria: Detection, Distribution and Migration of the Toxin in Aquatic Systems. Toxins 2017, 9, 166. [Google Scholar] [CrossRef]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Euro Surveill 2015, 20. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Higgins, C.; Higman, W.; Hungerford, J. Potential Threats Posed by Tetrodotoxins in UK Waters: Examination of Detection Methodology Used in Their Control. Mar Drugs 2015, 13, 7357–7376. [Google Scholar] [CrossRef]
- Wu, Z.; Xie, L.; Xia, G.; Zhang, J.; Nie, Y.; Hu, J.; Wang, S.; Zhang, R. A new tetrodotoxin-producing actinomycete, Nocardiopsis dassonvillei, isolated from the ovaries of puffer fish Fugu rubripes. Toxicon 2005, 45, 851–859. [Google Scholar] [CrossRef]
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First Detection of Tetrodotoxin in Greek Shellfish by UPLC-MS/MS Potentially Linked to the Presence of the Dinoflagellate Prorocentrum minimum. Toxins (Basel) 2015, 7, 1779–1807. [Google Scholar] [CrossRef]
- Kodama, M.; Sato, S.; Sakamoto, S.; Ogata, T. Occurrence of tetrodotoxin in Alexandrium tamarense, a causative dinoflagellate of paralytic shellfish poisoning. Toxicon 1996, 34, 1101–1105. [Google Scholar] [CrossRef]
- Turner, A.D.; Fenwick, D.; Powell, A.; Dhanji-Rapkova, M.; Ford, C.; Hatfield, R.G.; Santos, A.; Martinez-Urtaza, J.; Bean, T.P.; Baker-Austin, C. , et al. New Invasive Nemertean Species (Cephalothrix Simula) in England with High Levels of Tetrodotoxin and a Microbiome Linked to Toxin Metabolism. Mar Drugs 2018, 16. [Google Scholar] [CrossRef]
- Dhanji-Rapkova, M.; Hatfield, R.G.; Walker, D.I.; Hooper, C.; Alewijnse, S.; Baker-Austin, C.; Turner, A.D.; Ritchie, J.M. Investigating Non-Native Ribbon Worm Cephalothrix simula as a Potential Source of Tetrodotoxin in British Bivalve Shellfish. Mar Drugs 2024, 22. [Google Scholar] [CrossRef]
- Dhanji-Rapkova, M.; Teixeira Alves, M.; Triñanes, J.A.; Martinez-Urtaza, J.; Haverson, D.; Bradley, K.; Baker-Austin, C.; Huggett, J.F.; Stewart, G.; Ritchie, J.M. , et al. Sea temperature influences accumulation of tetrodotoxin in British bivalve shellfish. Science of The Total Environment 2023, 885, 163905. [Google Scholar] [CrossRef] [PubMed]
- Gerssen, A.; Gago-Martínez, A. Emerging Marine Biotoxins. Toxins (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Hort, V.; Arnich, N.; Guérin, T.; Lavison-Bompard, G.; Nicolas, M. First Detection of Tetrodotoxin in Bivalves and Gastropods from the French Mainland Coasts. Toxins (Basel) 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Réveillon, D.; Savar, V.; Schaefer, E.; Chevé, J.; Halm-Lemeille, M.-P.; Hervio-Heath, D.; Travers, M.-A.; Abadie, E.; Rolland, J.-L.; Hess, P. Tetrodotoxins in French Bivalve Mollusks—Analytical Methodology, Environmental Dynamics and Screening of Bacterial Strain Collections. Toxins 2021, 13, 740. [Google Scholar] [CrossRef]
- Rodriguez, P.; Alfonso, A.; Vale, C.; Alfonso, C.; Vale, P.; Tellez, A.; Botana, L.M. First toxicity report of tetrodotoxin and 5,6,11-trideoxyTTX in the trumpet shell Charonia lampas lampas in Europe. Anal Chem 2008, 80, 5622–5629. [Google Scholar] [CrossRef]
- Fernández-Ortega, J.F.; Santos, J.M.M.-d.l.; Herrera-Gutiérrez, M.E.; Fernández-Sánchez, V.; Loureo, P.R.; Rancaño, A.A.; Téllez-Andrade, A. Seafood Intoxication by Tetrodotoxin: First Case in Europe. The Journal of Emergency Medicine 2010, 39, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.R.; Giráldez, J.; Rodrigues, S.M.; Leão, J.M.; Pinto, E.; Soliño, L.; Gago-Martínez, A. High Levels of Tetrodotoxin (TTX) in Trumpet Shell Charonia lampas from the Portuguese Coast. Toxins (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Akyol, O.; Ünal, V.; Ceyhan, T.; Bilecenoglu, M. First confirmed record of Lagocephalus sceleratus(Gmelin, 1789) in the Mediterranean Sea. Journal of Fish Biology 2005, 66, 1183–1186. [Google Scholar] [CrossRef]
- Ulman, A.; Abd Rabou, A.F.N.; Al Mabruk, S.; Bariche, M.; Bilecenoğlu, M.; Demirel, N.; Galil, B.S.; Hüseyinoğlu, M.F.; Jimenez, C.; Hadjioannou, L. , et al. Assessment of Human Health Impacts from Invasive Pufferfish (Attacks, Poisonings and Fatalities) across the Eastern Mediterranean. Biology 2024, 13, 208. [Google Scholar] [CrossRef] [PubMed]
- Ben Souissi, J.; Rifi, M.; Ghanem, R.; Ghozzi, L.; Boughedir, W.; Azzurro, E. Lagocephalus sceleratus (Gmelin, 1789) expands through the African coasts towards the Western Mediterranean Sea: a call for awareness. Management of Biological Invasions 2014, 5, 357–362. [Google Scholar] [CrossRef]
- EFSA. Panel on Contaminants in the Food Chain. Scientific opinion on the risks for public health related to the presence of tetrodotoxin (TTX) and TTX analogues in marine bivalves and gastropods. EFSA J. 2017, 15. [Google Scholar]
- Gerssen, A.; Bovee, T.H.F.; Klijnstra, M.D.; Poelman, M.; Portier, L.; Hoogenboom, R. First Report on the Occurrence of Tetrodotoxins in Bivalve Mollusks in The Netherlands. Toxins (Basel) 2018, 10. [Google Scholar] [CrossRef]
- Katikou, P. Public Health Risks Associated with Tetrodotoxin and Its Analogues in European Waters: Recent Advances after The EFSA Scientific Opinion. Toxins (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Biessy, L.; Boundy, M.J.; Smith, K.F.; Harwood, D.T.; Hawes, I.; Wood, S.A. Tetrodotoxin in marine bivalves and edible gastropods: A mini-review. Chemosphere 2019, 236, 124404. [Google Scholar] [CrossRef]
- Turner, A.D.; Dhanji-Rapkova, M.; Coates, L.; Bickerstaff, L.; Milligan, S.; O'Neill, A.; Faulkner, D.; McEneny, H.; Baker-Austin, C.; Lees, D.N. , et al. Detection of Tetrodotoxin Shellfish Poisoning (TSP) Toxins and Causative Factors in Bivalve Molluscs from the UK. Mar Drugs 2017, 15. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, I.; Alfonso, A.; Alonso, E.; Rubiolo, J.A.; Roel, M.; Vlamis, A.; Katikou, P.; Jackson, S.A.; Menon, M.L.; Dobson, A. , et al. The association of bacterial C9-based TTX-like compounds with Prorocentrum minimum opens new uncertainties about shellfish seafood safety. Scientific Reports 2017, 7, 40880. [Google Scholar] [CrossRef]
- Patria, F.P.; Pekar, H.; Zuberovic-Muratovic, A. Multi-Toxin Quantitative Analysis of Paralytic Shellfish Toxins and Tetrodotoxins in Bivalve Mollusks with Ultra-Performance Hydrophilic Interaction LC-MS/MS-An In-House Validation Study. Toxins (Basel) 2020, 12. [Google Scholar] [CrossRef]
- Noguchi, T.; Jeon, J.K.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Shida, Y.; Hashimoto, K. Occurrence of tetrodotoxin and anhydrotetrodotoxin in Vibrio sp. isolated from the intestines of a xanthid crab, Atergatis floridus. J Biochem 1986, 99, 311–314. [Google Scholar] [CrossRef]
- Leão, J.M.; Lozano-Leon, A.; Giráldez, J.; Vilariño, Ó.; Gago-Martínez, A. Preliminary Results on the Evaluation of the Occurrence of Tetrodotoxin Associated to Marine Vibrio spp. in Bivalves from the Galician Rias (Northwest of Spain). Mar Drugs 2018, 16. [Google Scholar] [CrossRef] [PubMed]
- Bacchiocchi, S.; Campacci, D.; Siracusa, M.; Dubbini, A.; Accoroni, S.; Romagnoli, T.; Campanelli, A.; Griffoni, F.; Tavoloni, T.; Gorbi, S. , et al. A Hotspot of TTX Contamination in the Adriatic Sea: Study on the Origin and Causative Factors. Mar Drugs 2022, 21. [Google Scholar] [CrossRef] [PubMed]
- Bacchiocchi, S.; Campacci, D.; Siracusa, M.; Dubbini, A.; Leoni, F.; Tavoloni, T.; Accoroni, S.; Gorbi, S.; Giuliani, M.E.; Stramenga, A. , et al. Tetrodotoxins (TTXs) and Vibrio alginolyticus in Mussels from Central Adriatic Sea (Italy): Are They Closely Related? Mar Drugs 2021, 19. [Google Scholar] [CrossRef] [PubMed]
- Hernroth, B.E.; Baden, S.P. Alteration of host-pathogen interactions in the wake of climate change - Increasing risk for shellfish associated infections? Environ Res 2018, 161, 425–438. [Google Scholar] [CrossRef]
- Vezzulli, L.; Brettar, I.; Pezzati, E.; Reid, P.C.; Colwell, R.R.; Höfle, M.G.; Pruzzo, C. Long-term effects of ocean warming on the prokaryotic community: evidence from the vibrios. The ISME Journal 2011, 6, 21–30. [Google Scholar] [CrossRef]
- Gyraitė, G.; Kataržytė, M.; Bučas, M.; Kalvaitienė, G.; Kube, S.; Herlemann, D.P.; Pansch, C.; Andersson, A.F.; Pitkanen, T.; Hokajärvi, A.M. , et al. Epidemiological and environmental investigation of the 'big four' Vibrio species, 1994 to 2021: a Baltic Sea retrospective study. Euro Surveill 2024, 29. [Google Scholar] [CrossRef]
- Larsen, J.L.; Farid, A.F.; Dalsgaard, I. Occurrence of Vibrio parahaemolyticus and Vibrio alginolyticus in marine and estuarine bathing areas in Danish coast. Zentralblatt fur Bakteriologie, Mikrobiologie und Hygiene. 1. Abt. Originale B, Hygiene 1981, 173, 338–345. [Google Scholar]
- Gjerde, J.; Böe, B. Isolation and characterization of Vibrio alginolyticus and Vibrio parahaemolyticus from the Norwegian coastal environment. Acta Vet Scand 1981, 22, 331–343. [Google Scholar] [CrossRef]
- Eiler, A.; Johansson, M.; Bertilsson, S. Environmental Influences on <i>Vibrio</i> Populations in Northern Temperate and Boreal Coastal Waters (Baltic and Skagerrak Seas). Applied and Environmental Microbiology 2006, 72, 6004–6011. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Trinanes, J.A.; Salmenlinna, S.; Löfdahl, M.; Siitonen, A.; Taylor, N.G.; Martinez-Urtaza, J. Heat Wave-Associated Vibriosis, Sweden and Finland, 2014. Emerg Infect Dis 2016, 22, 1216–1220. [Google Scholar] [CrossRef]
- Swedish Meteorological and Hydrological Institute (SMHI). Available online: https://www.smhi.se/klimat/klimatet-da-och-nu/manadens-vader-och-vatten-i-sverige/manadens-vader-och-vatten-i-sverige/2019-07-05-juni-2019---normala-observationer (accessed on 5th March 2025).
- Swedish Meteorological and Hydrological Institute (SMHI). Available online: https://www.smhi.se/klimat/klimatet-da-och-nu/manadens-vader-och-vatten-i-sverige/manadens-vader-och-vatten-i-sverige/2021-08-09-juli-2021---sma-variationer-under-hogtrycksbetonad-julimanad (accessed on 5th March 2025).
- Albretsen, J.; Aure, J.; Sætre, R.; Danielssen, D.S. Climatic variability in the Skagerrak and coastal waters of Norway. ICES Journal of Marine Science 2011, 69, 758–763. [Google Scholar] [CrossRef]
- Dutheil, C.; Meier, H.E.M.; Gröger, M.; Börgel, F. Warming of Baltic Sea water masses since 1850. Climate Dynamics 2023, 61, 1311–1331. [Google Scholar] [CrossRef]
- Kajihara, H.; Sun, S.C.; Chernyshev, A.V.; Chen, H.X.; Ito, K.; Asakawa, M.; Maslakova, S.A.; Norenburg, J.L.; Strand, M.; Sundberg, P.; et al. Taxonomic identity of a tetrodotoxin-accumulating ribbon-worm Cephalothrix simula (Nemertea: Palaeonemertea): a species artificially introduced from the Pacific to Europe. Zoolog Sci 2013, 30, 985–997. [Google Scholar] [CrossRef]
- Fernández-Álvarez, F.Á.; Machordom, A. DNA barcoding reveals a cryptic nemertean invasion in Atlantic and Mediterranean waters. Helgoland Marine Research 2013, 67, 599–605. [Google Scholar] [CrossRef]
- Faasse, M.; Turbeville, J.M. The first record of the north-west Pacific nemertean Cephalothrix simula in northern Europe. Marine Biodiversity Records 2015, 8. [Google Scholar] [CrossRef]
- Sundberg, P.; Gibson, R.; Strand, M. Swedish nemerteans (phylum Nemertea), with description of a new hoplonemertean genus and species. Journal of Natural History 2007, 41, 2287–2299. [Google Scholar] [CrossRef]
- Strand, M.; Hedström, M.; Seth, H.; McEvoy, E.G.; Jacobsson, E.; Göransson, U.; Andersson, H.S.; Sundberg, P. The Bacterial (Vibrio alginolyticus) Production of Tetrodotoxin in the Ribbon Worm Lineus longissimus-Just a False Positive? Mar Drugs 2016, 14. [Google Scholar] [CrossRef]
- Carney, K. Marine bioinvasion prevention : understanding ballast water transportation conditions and the development of effective treatment systems. PhD Thesis, Newcastle University, School of Marine Science and Technology, 2011. doi: http://theses.ncl.ac.uk/jspui/handle/10443/1246. [Google Scholar]
- Flagella, M.M.; Abdulla, A.A. Ship ballast water as a main vector of marine introductions in the Mediterranean Sea. WMU Journal of Maritime Affairs 2005, 4, 95–104. [Google Scholar] [CrossRef]
- Pierce, R.W.; Carlton, J.T.; Carlton, D.A.; Geller, J.B. Ballast water as a vector for tintinnid transport. Mar Ecol Prog Ser 1997, 149, 295–297. [Google Scholar] [CrossRef]
- Pagenkopp Lohan, K.M.; Darling, J.A.; Ruiz, G.M. International shipping as a potent vector for spreading marine parasites. Diversity and Distributions 2022, 28, 1922–1933. [Google Scholar] [CrossRef]
- Katikou, P.; Vlamis, A. Tetrodotoxins: recent advances in analysis methods and prevalence in European waters. Current Opinion in Food Science 2017, 18, 1–6. [Google Scholar] [CrossRef]
- Dhanji-Rapkova, M.; Turner, A.D.; Baker-Austin, C.; Huggett, J.F.; Ritchie, J.M. Distribution of Tetrodotoxin in Pacific Oysters (Crassostrea gigas). Mar Drugs 2021, 19. [Google Scholar] [CrossRef] [PubMed]
- Boundy, M.J.; Biessy, L.; Roughan, B.; Nicolas, J.; Harwood, D.T. Survey of Tetrodotoxin in New Zealand Bivalve Molluscan Shellfish over a 16-Month Period. Toxins (Basel) 2020, 12. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map of Swedish bivalve molluscs production areas (purple) and selected sampling sites (blue). n represents the number of samples per site.
Figure 1.
Map of Swedish bivalve molluscs production areas (purple) and selected sampling sites (blue). n represents the number of samples per site.
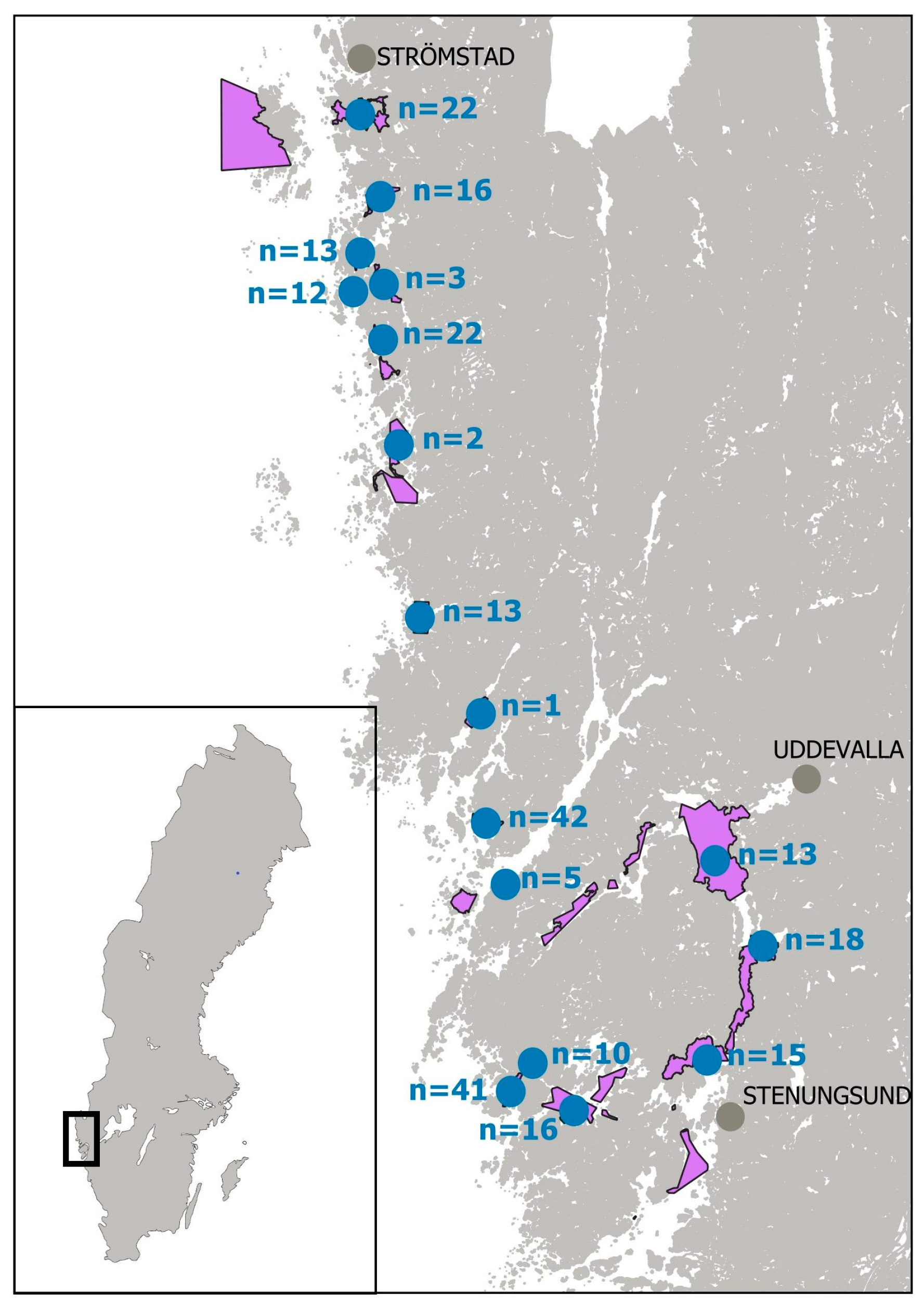
Figure 2.
The number of samples tested in the study and their distribution across four species.
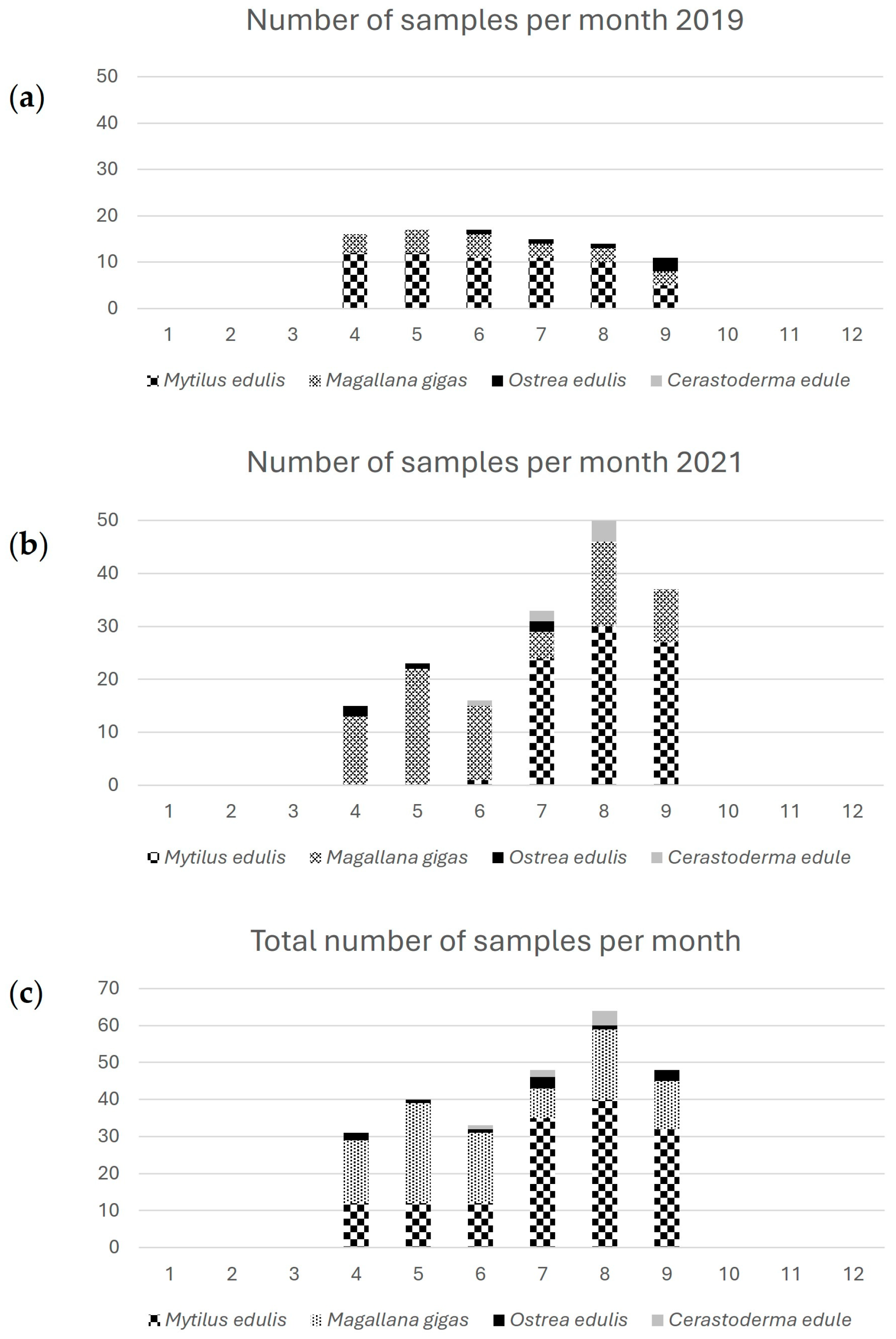
Figure 3.
Proportion of each species in the study, of which farmed (or wild) per species.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



