
Submitted:
29 April 2025
Posted:
30 April 2025
You are already at the latest version
Abstract
Trichomes in Cannabis sativa are specialized structures responsible for cannabinoid and terpene biosynthesis, making their density a critical factor for both research and industrial applications. Despite their importance, trichome density analysis is hindered by variability across plant structures and the lack of standardized protocols. This study evaluates different plant structures—bracts, sugar leaves, calyxes, and the main cola—to determine the most reliable site for trichome counting. Among these, bracts emerge as the most consistent due to their homogeneous trichome distribution and high cannabinoid concentration. While sugar leaves and calyxes also contribute to trichome yield assessments, their variability necessitates careful sampling. Moreover, trichome shape and size must be taken into consideration when correlating trichome density with secondary metabolite levels. The integration of microscopic imaging and software-assisted counting enhances accuracy and reproducibility in trichome density analysis. Establishing a standardized protocol for trichome assessment will improve cannabinoid yield optimization, quality control, and overall Cannabis research methodologies. Incorporating morphological data (trichome density, distribution, shape, and size) with chemical assays (cannabinoid and terpene identification and quantification) thus provides a more robust assessment of Cannabis potency and value. Future work should refine imaging techniques and sampling strategies to further enhance trichome analysis reliability.
Keywords:
Cannabinoids
; Terpenes
; Calyxes
; Anatomic Analysis
; Plant structures
; Protocols
1. Introduction
Virtually all terrestrial plant species possess epidermal outgrowths that resemble hair-like projections. These structures, generally referred to as trichomes when located on aerial parts, are multifunctional adaptations that play significant roles in plant survival [1]. When similar outgrowths occur on roots, they are typically called root hairs, emphasizing their distinct physiological context. Etymologically, the term “trichomes” is derived from the Greek word “trichos,” meaning hair, highlighting their morphological resemblance. Despite their superficial resemblance to vascular or ground tissues, trichomes are in fact specialized epidermal extensions rather than integral components of the plant’s internal vascular system [1].
1.1. Diversity and Functional Roles of Trichomes
Trichomes exhibit considerable diversity in both form and function. They may be non-glandular (often acting as simple hairs) or glandular, wherein specialized secretory cells produce and store bioactive compounds [2]. Non-glandular trichomes can deter herbivores by creating physical barriers or by reducing palatability, while also regulating leaf temperature and water loss through light reflection—an adaptation particularly important in arid environments [3,4]. Meanwhile, glandular trichomes can synthesize an array of secondary metabolites, including essential oils, alkaloids, and terpenes, which serve defensive, allelopathic, or communicative functions [3]. Consequently, trichomes act as a protective barrier against biotic threats (herbivores, pests, and pathogens, e.g.) and abiotic stressors, such as UV radiation and excessive transpiration. Interestingly, in certain species, they also facilitate seed dispersal or seed protection [4,5].
Structurally, trichomes are typically classified according to their cellularity (single-celled vs. multicellular), branching pattern (branched vs. unbranched), and secretory activity (glandular vs. non-glandular) [2]. They also take on diverse shapes such as star-like (stellate), hooked, peltate (scale-like), or capitate (headed). Larger, more conspicuous trichomes are often found on leaf undersides (abaxial surfaces) or along leaf edges, while smaller trichomes can appear in stomatal regions or in association with vascular bundles [6]. Notably, the presence of dense trichomes in some species increases epidermal thickness and may enhance the production of long-chain fatty acids, thus reducing water loss and stabilizing leaf temperature [7]. Recent transcriptomic studies suggest that the formation and specialization of trichomes are controlled by intricate genetic networks—highlighting a balance between the plant’s developmental signaling and environmental cues [8,5].
1.2. Trichomes in Cannabis sativa L
Among the myriad plant species exhibiting trichomes, Cannabis sativa L. stands out for the medicinal and commercial significance of the metabolites produced in its glandular trichomes (Figure 1). Cannabis flowers, particularly those of female plants, have garnered attention due to their production of cannabinoids (e.g., Δ9-tetrahydrocannabinol [THC], cannabidiol [CBD]) and terpenes, compounds highly valued in both therapeutic and recreational contexts. Trichome biosynthesis of these specialized metabolites is influenced by the interplay of genetic factors, developmental stage, and environmental conditions, including light spectrum and intensity, temperature, and physical stress [9,10].
On a morphological level, female Cannabis flowers possess at least three distinct glandular trichome types based on their surface features and stalk length: bulbous, sessile capitate, and stalked (pedicellate) capitate [11].
Bulbous trichomes are the smallest and are dispersed across various aerial surfaces of the plant. Despite their ubiquity, they remain the least characterized, partly due to their diminutive size.
Sessile capitate trichomes have a more prominent glandular head and are typically abundant on sugar leaves and bracts.
Stalked (pedicellate) capitate trichomes are generally larger and often regarded as the principal source of cannabinoids and terpenes within female inflorescences. These structures are most visible to the naked eye during the later flowering stages, creating the characteristic “frosty” appearance on colas and sugar leaves [10].
Notably, bracts in female Cannabis flowers frequently harbor higher densities of these glandular trichomes and thus represent crucial sites for cannabinoid and terpene biosynthesis [10,12]. As a result, the distribution, density, and maturity of trichomes in specific floral regions—ranging from bracts and calyxes to sugar leaves—have become a focal point of breeding and cultivation strategies aimed at optimizing resin yield and the biochemical composition in terms of the bioactive compounds of interest. Importantly, beyond their biochemical significance, these trichomes also offer protective functions, forming a defensive barrier against pests and environmental stresses during critical reproductive phases. Furthermore, higher trichome density creates a desirable aesthetic attribute for consumers.
Considering this dual role—both functional defense and bioactive compound biosynthesis and storage—trichomes in C. sativa L. are of paramount interest to researchers and cultivators. Understanding the morphological, genetic, biochemical, and environmental factors that govern trichome development and metabolism enables more precise cultivation practices and efficient breeding programs, ultimately influencing the quality and consistency of Cannabis-derived products. The following sections delve deeper into the methods for assessing trichome density in various Cannabis structures, with the aim of establishing reproducible and standardized approaches in both research and industrial applications.
C. sativa L. flowers are consumed for their specific medicinal and recreational purposes according to the properties of their specialized metabolites (i.e., cannabinoids and terpenes). Based on their surface morphology, three different forms of glandular trichomes have been identified in female Cannabis flowers: bulbous, sessile, and stalked [11]. The bulbous trichomes are the smallest and least studied, distributed across the surface of the plant. The sessile capitate trichomes have a larger glandular head and are commonly found on sugar leaves and bracts. In their turn, pedicellate capitate trichomes, more abundant in tissues rich in cannabinoids, are typically found on the bracts of female flowers and are the primary sites of cannabinoid and terpene biosynthesis. These trichomes are visible to the naked eye and are most abundant during the flowering phase, covering the colas and sugar leaves [10].
The glandular trichomes of C. sativa play a central role in the biosynthesis and storage of cannabinoids and terpenes, compounds with recognized therapeutic, psychoactive, and commercial relevance. Trichome density is often considered an indicator of flower quality, particularly in relation to resin production and extraction yield. However, the lack of standardized protocols for measuring trichome density hinders comparison across studies and limits its reliability as a quality criterion in breeding and industry practices. Additionally, trichome distribution and morphology vary among floral structures—such as bracts, sugar leaves, calyces, and the main cola—raising the question of which plant region best represents overall trichome density in analytical assessments.
The density and morphology of glandular trichomes vary markedly among different floral structures of C. sativa, including bracts, sugar leaves, calyxes, and the main cola. At the anatomical level, glandular trichomes house specialized secretory cells in their head region, which synthesize and store secondary metabolites such as cannabinoids and terpenes within a subcuticular cavity [5,11]. The accumulation of these metabolites is a dynamic process influenced by a confluence of factors—environmental conditions, developmental stage, and genetic background.
Recent genotype × environment (G×E) studies have shown that variations in light spectrum, temperature, humidity, and even UV-B light exposure can significantly modulate trichome density and secondary metabolite profiles [8,9]. Meanwhile, the age of the plant and the onset of the flowering stage can shift the trichome’s developmental trajectory, impacting both the density and chemical composition over time [10]. For instance, Punja et al. (2023) [10] noted that the number of capitate trichomes and pedicel length differed significantly across genotypes and between the upper and lower surfaces of bracts, underscoring the intricate interplay between genetics and plant morphology. These findings illustrate how structural variations in trichomes can affect the quality and consistency of the final product by influencing the content of cannabinoids and terpenes.
Beyond their biochemical relevance, trichomes also play a pivotal role in the visual and commercial appeal of C. sativa flowers, particularly in the recreational market. A dense covering of glandular trichomes imparts a characteristic “frosty” appearance to flowers, as shown in Figure 2, which is often perceived by consumers as an indicator of higher potency and quality. Consequently, breeders and cultivators target enhanced trichome coverage through selective breeding, aiming to maximize both resin yield and market value [11]. For medicinal applications, consistent trichome density across harvests helps ensure reproducible therapeutic efficacy, while for recreational consumers, the visual allure can significantly influence purchasing decisions.
Despite its critical importance, accurately assessing trichome density remains challenging. Variability across plant structures—bracts, sugar leaves, calyxes, and main cola tissues—can be substantial, and lack of standardized protocols further complicates reliable comparisons across different studies and commercial operations [5]. Moreover, trichome shape and size must be taken into consideration when correlating trichome density with secondary metabolite levels. A robust, reproducible methodology for trichome counting would therefore be of immense value, facilitating consistent cannabinoid yield assessments and quality control in both research and industry contexts. Recent efforts have begun to explore more refined imaging techniques and computational tools for trichome quantification, but these methods require broader validation and consensus [8].
This review explores the methods used to count trichomes on different Cannabis structures, examines structural variation in trichome density, assesses available quantification techniques, and proposes future directions for standardization in both research and industrial applications.
2. Methods for Trichome Counting
Accurate trichome quantification in C. sativa is often challenging due to uneven distribution across different plant structures. Consequently, researchers and cultivators use various methods to determine where and how to count trichomes. Manual counts under a dissecting or compound microscope remain a foundational approach but can be labor-intensive and prone to observer bias [10]. To mitigate these limitations, digital imaging and software-assisted analysis commonly employing platforms like ImageJ software (ImageJ, U. S. National Institutes of Health, Bethesda, Maryland, USA, https://imagej.net/ij/), for instance, are increasingly popular, allowing for standardized area selection and semi-automated trichome detection [5]. On the other hand, advanced methods such as confocal or scanning electron microscopy offer detailed morphological insights, but require specialized equipment, whereas field-based techniques (e.g., portable microscopes) provide more rapid, if less precise, assessments. Regardless of the chosen method, consistent sampling protocols and clearly defined reference areas are vital for reproducibility. In the following sections, we examine how trichome density varies across key Cannabis structures depicted in Figure 3, as e.g., main cola, bracts, calyxes, and sugar leaves, and discuss the merits and limitations of focusing on each site for reliable trichome analysis.
2.1. Main Cola
As illustrated in Figure 4, the main cola, or apical bud, stands out as the plant’s primary flowering site and is frequently cited as having one of the highest trichome densities [13]. This dense coverage can be attributed to favorable light exposure at the canopy’s apex, along with preferential nutrient allocation during flowering. However, intra-cola variability still poses a challenge, as subtle differences in trichome distribution within the apical region can affect the reliability of quantitative assessments. Researchers often recommend consistent sampling protocols, such as selecting a defined portion of the upper cola to improve reproducibility when evaluating trichome density in this visually prominent structure.
The main cola, or apical bud, is widely regarded as a focal point of C. sativa inflorescence due to its high trichome density and substantial contribution to overall resin yield [13]. This prominence stems from favorable light exposure at the canopy’s apex and concentrated nutrient flow during the flowering stage, factors that collectively promote robust trichome development. Despite these advantages, trichome distribution within the main cola can be highly heterogeneous, reflecting micro-environmental variations (e.g., light penetration, airflow) and internal plant processes [9].
Such variability introduces significant challenges for researchers seeking standardized, repeatable trichome analysis in this key region. When sampling is not carefully controlled, measurements of trichome density may fluctuate widely, complicating cross-comparisons among different studies or cultivars. To mitigate these issues, many researchers advocate a targeted sampling strategy that focuses on well-defined substructures within the cola, such as a specified zone of upper bracts or the distal end of sugar leaves. For instance, selecting a consistent, clearly demarcated 1–2 cm section on the upper portion of the cola can help ensure that repeated measurements capture a comparable physiological zone, reducing both intra- and inter-plant variability [10]. By employing this localized approach, researchers can enhance reproducibility in trichome density measurements while still accounting for the main cola’s significant role in cannabinoid and terpene biosynthesis. Moreover, combining a structured sampling protocol with digital imaging tools (e.g., ImageJ or machine-learning platforms) can further standardize the evaluation process, minimizing observer bias and facilitating reliable quantitative data [5]. Ultimately, although the main cola remains a highly relevant site for resin production and overall flower quality, adherence to rigorous sampling protocols is essential to harness its full potential as a credible metric for trichome-related studies.
2.2. Bracts
Bracts in Cannabis are small, leaf-like structures that surround and protect the reproductive organs of the plant (Figure 5). They are often described as the most reliable site for trichome analysis due to their dense and homogeneous resin coverage. Unlike other floral structures, bracts exhibit a relatively consistent glandular trichome distribution, making them ideal for microscopic imaging and cannabinoid quantification [12]. Methods for analyzing trichomes on bracts typically involve imaging the surface under a dissecting microscope, followed by manual or software-assisted counting. Image-processing software can help standardize area selection, ensuring consistency across samples. Since bracts are larger and more exposed compared to inner floral structures, they provide an accessible and efficient target for cannabinoid research.
Although the term bract is sometimes used interchangeably with bractlet or calyx, botanical classifications distinguish bracts as distinct structures that envelop the developing flowers and seeds [14]. Their primary function is to protect the plant's reproductive components while simultaneously serving as a major site for cannabinoid and terpene production. Bracts in Cannabis are small, leaf-like structures that surround and protect the reproductive organs of the plant (Figure 5). They are often described as the most reliable site for trichome analysis due to their dense and homogeneous resin coverage. Unlike other floral structures, bracts exhibit a relatively consistent glandular trichome distribution, making them ideal for microscopic imaging and cannabinoid quantification [12]. Methods for analyzing trichomes on bracts typically involve imaging the surface under a dissecting microscope, followed by manual or software-assisted counting. Image-processing software can help standardize area selection, ensuring consistency across samples. Since bracts are larger and more exposed compared to inner floral structures, they provide an accessible and efficient target for cannabinoid research.
Although the term bract is sometimes used interchangeably with bractlet or calyx, botanical classifications distinguish bracts as distinct structures that envelop the developing flowers and seeds [14]. Their primary function is to protect the plant's reproductive components while simultaneously serving as a major site for cannabinoid and terpene production.
Researchers typically employ light or dissecting microscopy to capture high-resolution images of bract surfaces, followed by manual or software-assisted counting of trichomes. Image-processing tools such as ImageJ can standardize the counting area (e.g., 1 mm²), ensuring consistency across multiple samples and experimental replicates. Given that bracts are readily accessible and not as tightly packed as inner floral structures, they are conducive to rapid sampling and imaging workflows. This accessibility streamlines the quantification of cannabinoids and terpenes, which in turn facilitates both breeding research and commercial quality control efforts [5].
Significance of Bracts for Cannabinoid and Terpene Biosynthesis
The protective function of bracts aligns with their high density of glandular trichomes; secondary metabolites produced in these trichomes not only deter herbivores and pathogens but also serve as the foundation for the plant’s medicinal and recreational value [10]. As a result, focusing on bracts offers researchers and cultivators insight into the plant’s primary sites of cannabinoid and terpene biosynthesis. This dual role, i.e., physical protection and metabolite production, makes bracts a particularly appealing target for those aiming to optimize resin output or explore the genetic determinants of trichome development.
By coupling consistent sampling of bracts with rigorous imaging protocols, studies can achieve highly reproducible trichome density measurements, an essential factor for breeding programs and product standardization. Consequently, bracts represent one of the most reliable benchmarks for comparing both genotypic variation and environmental influences on trichome production in C. sativa.
2.3. Calyxes
The term calyx in C. sativa remains a point of contention, as some sources equate it with the bract, while others view it as a distinct inner structure that encases the ovule [14,15]. This inconsistency stems from overlapping morphological features and the protective roles that such structures perform, leading to varied usage across botanical literature and Cannabis research. Nevertheless, the generally accepted definition positions the calyx as an inward protective layer situated within the bracts, playing a critical role in reproductive organ development. A core challenge in studying calyxes arises from their small size and close integration with the plant’s reproductive tissues. Unlike the more accessible bracts, calyxes typically require meticulous dissection to isolate for microscopic examination. Although they often harbor abundant glandular trichomes, their compact architecture can exhibit spatial variability in trichome distribution, particularly if different segments of the calyx envelop the ovule with varying degrees of tightness or exposure. This heterogeneity not only complicates sampling, but also demands greater precision in quantification methods, whether manual or software assisted.
Despite these logistical hurdles, calyxes remain highly relevant for cannabinoid research. Their intimate involvement in enclosing the ovule and rich trichome content mean they contribute substantially to resin production, including key cannabinoids and terpenes. By refining the techniques used to identify and measure calyx-specific trichomes, researchers can gain deeper insights into the timing and mechanisms of resin accumulation. This knowledge can inform both cultivar selection for higher potency and cultivation practices tailored to optimize yield. Consequently, while calyxes may be more challenging to analyze than bracts, their significance in cannabinoid biosynthesis makes them a crucial focus in ongoing studies of C. sativa flower development and resin production.
2.4. Sugar Leaves
Sugar leaves, as evidenced in Figure 6, the small “frosty” leaves that surround Cannabis buds, represent another key site for trichome density analysis. These leaves can contribute significantly to overall trichome yield [15], with higher densities observed closer to their tips, an attribute that makes them particularly valuable for cannabinoid extraction. In addition to their role as trichome-bearing structures, sugar leaves retain some photosynthetic capacity, theoretically aiding energy supply for the developing bud [11]. Although their photosynthetic contribution may be modest compared to the larger fan leaves, they potentially assist in local nutrient support for resin production. Given these dual functions, i.e., limited photosynthesis and high trichome content, sugar leaves offer an intriguing focal point for both morphological and biochemical analyses in Cannabis research.
Although sugar leaves significantly contribute to total resin yield, they demonstrate a less uniform distribution of trichomes in comparison to bracts, with elevated densities being found at the margins and tips. This variability requires meticulous sample techniques, frequently concentrating on areas abundant in trichomes, to guarantee precise density assessments. Although sugar leaves may lack the uniformity of bracts, they provide a helpful supplementary site for trichome analysis, especially in research focused on evaluating whole-plant cannabinoid concentration. Their inclusion aids in capturing the comprehensive resin profile of the plant and provides more insights into the variability of trichome production across various leaf areas and developmental phases.
3. Discussion
Establishing reliable methods for trichome density analysis remains central to Cannabis sativa research, given that secondary metabolites, primarily cannabinoids and terpenes, accumulate in these glandular structures. Among the plant tissues evaluated, bracts consistently demonstrate the highest reproducibility due to their relatively uniform trichome distribution and significant cannabinoid content [10]. This consistency also translates into lower plant-to-plant variance, a critical attribute for industrial-scale applications where maintaining batch consistency directly impacts market value and therapeutic efficacy [11].
Still, sugar leaves and calyxes offer valuable perspectives depending on experimental objectives. Sugar leaves, although less uniform in trichome dispersion, can illuminate extraction efficiency since their edges and tips often host abundant glandular heads [15]. Calyxes, despite posing sampling challenges due to their smaller size and proximity to reproductive tissues, represent strategic focal points for investigating cannabinoid biosynthesis pathways [14]. In contrast, although visually prominent, the main cola can exhibit substantial variability in trichome coverage; hence, standardization for this region typically involves precisely defined substructures (e.g., uppermost bracts) rather than the entire apical bud [13].
Despite trichomes’ pivotal role, higher density alone does not guarantee greater cannabinoid potency [12]. Recent studies underscore that while trichome density is a valuable morphological marker, it does not consistently predict cannabinoid content across genotypes. For instance, research has shown that higher trichome densities may not correlate directly with increased cannabinoid levels, suggesting that factors such as biosynthetic enzyme activity and resin storage capacity play significant roles in determining final cannabinoid amounts [10]. Multiple studies, including recent metabolomic and transcriptomic work, confirm that biosynthetic enzyme activity, genotypic expression profiles, and resin storage capacity are equally, if not more, influential in determining final cannabinoid amounts [8,10]. A cultivar with moderate trichome density but upregulated synthase genes may surpass a denser but genetically less active cultivar in total cannabinoid contents. Consequently, relying solely on trichome counting could obscure the true biochemical potential of a given plant genotype. Integrating morphological data (trichome density, distribution, shape, and size) with chemical assays (cannabinoid and terpene identification and quantification) thus provides a more robust assessment of Cannabis potency and value.
From a breeding standpoint, uniform and precise trichome density metrics enable phenotypic selection for cultivars that exhibit stable, high-resin traits under diverse environmental conditions [9]. Furthermore, environmental factors such as plant density and architecture significantly influence cannabinoid uniformity. Studies indicate that higher planting densities can reduce cannabinoid uniformity across different parts of the plant, although they may increase overall yield per area. Architectural modifications, like defoliation or pruning, have been shown to mitigate some of these effects, enhancing chemical standardization [16].
Moreover, anatomical and morphological differences among Cannabis trichomes have been linked to variations in metabolite profiles. Microcapillary sampling of individual glandular trichomes revealed that stalked trichomes on floral tissues predominantly accumulate monoterpenes, with monoterpene to sesquiterpene ratios exceeding 12:In contrast, sessile trichomes on vegetative leaves exhibited lower monoterpene content and higher sesquiterpene proportions. Interestingly, despite these differences in terpene profiles, cannabinoid compositions between stalked and sessile trichomes were found to be similar. These findings underscore the importance of considering trichome type and sampling location when analyzing secondary metabolite profiles, as morphological variations can significantly influence terpene composition, while cannabinoid content may remain relatively consistent across trichome types. The study by Livingston et al. (2020) [11] explores the dynamic changes in Cannabis trichome morphology and its relationship with metabolite content throughout the plant's maturation, providing insights into the underlying mechanisms that drive metabolite synthesis.
Adding to this, Ferri et al. (2024) [17] demonstrated that even individual glandular trichomes isolated from the same Cannabis flower can display detectable variations in their phytocannabinoid profiles. In their study, single trichomes from two Cannabis sativa L. varieties—a medicinal strain (FM2) and a CBD-rich hemp cultivar—were analyzed using an untargeted metabolomics approach. Around 70 phytocannabinoids, including carboxylated and decarboxylated forms of CBD, THC, and CBG, were identified. Although the profiles of isolated trichomes and whole inflorescences were broadly similar, absolute cannabinoid levels varied among trichomes, while the CBD-to-THC ratio remained consistent, reflecting genetic chemotypes. These results highlight that chemical variability exists even within a single anatomical region, suggesting that differences across broader plant structures, such as leaves, bracts, and flowers, could be even more significant. Despite the overall consistency observed between isolated trichomes and whole flower extracts, subtle differences reinforce the hypothesis that cannabinoid biosynthesis is influenced not only by genetics but also by tissue-specific factors.
However, despite ongoing advancements in microscopic imaging and software-assisted quantification, standardizing sampling protocols still poses significant hurdles. Variability introduced by genotype, developmental stage, and environmental factors often leads to inconsistent data across research labs and commercial operations [5,9]. Moreover, cultivars can manifest unique developmental timelines for trichome initiation, and environmental stressors (e.g., UV-B light, nutrient shifts) can differentially modulate secondary metabolite biosynthesis and accumulation [8]. Such variability underlines the need for harmonized guidelines, including standardized sampling areas (e.g., 1 mm² or 2 mm² sectors), consistent plant growth stages (e.g., mid-flowering), and documented environmental parameters (e.g., light intensity, humidity ranges).
Microscopic imaging (optical, confocal, electron) combined with macro-photographic methods improves the resolution and accuracy of trichome assessments. Platforms like ImageJ or custom machine-learning algorithms help reduce observer bias, permitting faster and more reproducible data collection [5]. However, uniform calibration of imaging equipment and consistent magnification levels are crucial for cross-study comparability [10]. Tanney et al. (2021) [5] demonstrated that portable Raman spectroscopy can non-destructively and rapidly profile cannabinoids and terpenes in trichomes without extensive sample preparation, offering a powerful tool for real-time, in situ analysis. In parallel, machine learning approaches have been used to automate trichome identification and quantification from microscopic images, as shown by Ebersbach et al. (2018) [18] in Arabidopsis, improving throughput and reducing observer bias—methods that could be adapted for Cannabis research. Comparative studies employing multiple quantification techniques—such as manual counting, ImageJ analysis, and AI-based methods—across various Cannabis cultivars under controlled conditions are warranted to validate the reliability and reproducibility of these approaches.
As regulatory frameworks evolve worldwide, robust methods for trichome evaluation will be increasingly important not only for academic research but also for quality control in commercial settings. Validated trichome density assays may become integral to legal compliance, product labeling, and ensuring batch-to-batch consistency, particularly in medicinal markets where patients rely on reproducible dosage [11]. Also, it creates a metric for a trait that is desirable to consumers. The development and adoption of standardized best practices will be essential for advancing Cannabis sativa as a scientifically validated crop.
4. Conclusions
Standardizing trichome density assessment remains a critical priority for advancing Cannabis research. This review highlights that among various plant tissues, bracts stand out as the most reliable sampling site, offering both consistent glandular coverage and high cannabinoid concentrations—key attributes for robust and reproducible analyses. Nevertheless, morphological evaluations alone are insufficient to fully characterize Cannabis potency. Future methodologies must integrate trichome density, distribution, shape, and size metrics with biochemical profiling, capturing not only structural traits but also the metabolic potential of different tissues. Advanced imaging techniques, automated quantification tools, and standardized sampling protocols will be essential to improve reproducibility across research and industrial applications. As the Cannabis industry matures and regulatory frameworks evolve, reliable trichome analysis will play an increasingly central role in quality control, breeding programs, and therapeutic product validation. Building a foundation of standardized, multidimensional assessment practices will be key to unlocking the full scientific and commercial potential of C. sativa.
Author Contributions
Original draft preparation: T.A.; Literature review: T.A., F.D., S.S., R.R; Editing: F.D., S.S., R.R., A.A.D., M.M., J.F.D.C.L.F.; Figures preparation: R.R., S.S.; Supervision: M.M., J.F.D.C.L.F.; Conceptualization: J.F.D.C.L.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding authors.
Acknowledgments
The authors would like to thank Sanchit Kumar for capturing one of the images used in this study (Figure 6).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Glas, J.J.; Schimmel, B.C.; Alba, J.M.; Escobar-Bravo, R.; Schuurink, R.C.; Kant, M.R. Plant glandular trichomes as targets for breeding or engineering of resistance to herbivores. Int. J. Mol. Sci. 2012, 13, 17077. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, C.; Meng, P.; Tan, G.; Lv, L. Analysis and review of trichomes in plants. BMC Plant Biol. 2021, 21. [Google Scholar] [CrossRef] [PubMed]
- Wagner, G.J. Secreting glandular trichomes: more than just hairs. Plant Physiol. 1991, 96, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Stratmann, J.W.; Bequette, C.J. Hairless but no longer clueless: understanding glandular trichome development. J. Exp. Bot. 2016, 67, 5285. [Google Scholar] [CrossRef] [PubMed]
- Tanney, C.A.S.; Backer, R.; Geitmann, A.; Smith, D.L. Cannabis Glandular Trichomes: A Cellular Metabolite Factory. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Mao, X.; Lin, Y. Trichome, a Functional Diversity Phenotype in Plant. Mol. Biol. 2016, s. [Google Scholar] [CrossRef]
- Hegebarth, D.; Buschhaus, C.; Wu, M.; Bird, D.; Jetter, R. The composition of surface wax on trichomes of Arabidopsis thaliana differs from wax on other epidermal cells. Plant J. 2016, 88, 762. [Google Scholar] [CrossRef] [PubMed]
- Backer, R.; Schwinghamer, T.; Rosenbaum, P.; McCarty, V.; Eichhorn Bilodeau, S.; Lyu, D.; Ahmed, M.B.; Robinson, G.; Lefsrud, M.; Wilkins, O.; et al. Closing the Yield Gap for Cannabis: A Meta-Analysis of Factors Determining Cannabis Yield. Front. Plant Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Pauli, C.S.; Gostin, E.L.; Staples, S.K.; Seifried, D.; Kinney, C.; Vanden Heuvel, B.D. Effects of short-term environmental stresses on the onset of cannabinoid production in young immature flowers of industrial hemp (Cannabis sativa L.). J. Cannabis Res. 2022, 4. [Google Scholar] [CrossRef] [PubMed]
- Punja, Z.K.; Sutton, D.B.; Kim, T. Glandular trichome development, morphology, and maturation are influenced by plant age and genotype in high THC-containing cannabis (Cannabis sativa L.) inflorescences. J. Cannabis Res. 2023, 5. [Google Scholar] [CrossRef] [PubMed]
- Livingston, S.J.; Quilichini, T.D.; Booth, J.K.; Wong, D.C.J.; Rensing, K.H.; Laflamme-Yonkman, J.; Castellarin, S.D.; Bohlmann, J.; Page, J.E.; Samuels, A.L. Cannabis glandular trichomes alter morphology and metabolite content during flower maturation. Plant J. 2020, 101, 37. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.C. , Hemphill, J.K., & Mahlberg, P.G. Trichomes and Cannabinoid Content of Developing Leaves and Bracts of Cannabis sativa L. (Cannabaceae). Am. J. Botany 1980, 67, 1397. [Google Scholar] [CrossRef]
- Laverty, K.U.; Stout, J.M.; Sullivan, M.J.; Shah, H.; Gill, N.; Holbrook, L.; Deikus, G.; Sebra, R.; Hughes, T.R.; Page, J.E.; et al. A physical and genetic map of Cannabis sativa identifies extensive rearrangements at the THC/CBD acid synthase loci. Genome Res. 2019, 29, 146. [Google Scholar] [CrossRef] [PubMed]
- Riboulet-Zemouli, K. ‘Cannabis’ ontologies I: Conceptual issues with Cannabis and cannabinoids terminology. Drug Sci. Policy Law. 2020, 6. [Google Scholar] [CrossRef]
- Raman, V.; Lata, H.; Chandra, S.; Khan, I.A.; ElSohly, M.A. Morpho-Anatomy of Marijuana (Cannabis sativa L.). In Cannabis sativa L. - Botany and Biotechnology, Chandra, S., Lata, H., ElSohly, M.A. Eds.; Springer International Publishing, 2017; pp 123-136.
- Danziger, N.; Bernstein, N. Too Dense or Not Too Dense: Higher Planting Density Reduces Cannabinoid Uniformity but Increases Yield/Area in Drug-Type Medical Cannabis. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Ferri, E.; Cerrato, A.; Caprari, C.; Russo, F.; Laganà, A.; Capriotti, A.L.; Faggiana, G.; Moro, A.; Citti, C.; Cannazza, G. Single trichome phytocannabinomics of two different cannabis varieties. J. Pharm. Biomed. Anal. 2025, 261. [Google Scholar] [CrossRef] [PubMed]
- Ebersbach, P.; Stehle, F.; Kayser, O.; Freier, E. Chemical fingerprinting of single glandular trichomes of Cannabis sativa by Coherent anti-Stokes Raman scattering (CARS) microscopy. BMC Plant Biol. 2018, 18. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Detailed visualization of the resinous biomass produced by trichomes on the adaxial surface of Cannabis sativa sugar leaves, highlighting capitate-stalked trichomes, cystolithic hairs, and sessile capitate trichomes. The figure was created using Biorender (https://BioRender.com, accessed 10 April 2025).
Figure 1.
Detailed visualization of the resinous biomass produced by trichomes on the adaxial surface of Cannabis sativa sugar leaves, highlighting capitate-stalked trichomes, cystolithic hairs, and sessile capitate trichomes. The figure was created using Biorender (https://BioRender.com, accessed 10 April 2025).

Figure 2.
Details of the dense covering of glandular trichomes that imparts a frosty appearance to Cannabis sativa female flowers.
Figure 2.
Details of the dense covering of glandular trichomes that imparts a frosty appearance to Cannabis sativa female flowers.

Figure 3.
Illustration of Cannabis sativa’s anatomy highlighting anatomical regions for trichome identification, including main cola, sugar leaves, fan leaves, bracts, stigma, ovary, calyx, and trichomes. The figure was created using Biorender (https://BioRender.com, accessed 12 April 2025).
Figure 3.
Illustration of Cannabis sativa’s anatomy highlighting anatomical regions for trichome identification, including main cola, sugar leaves, fan leaves, bracts, stigma, ovary, calyx, and trichomes. The figure was created using Biorender (https://BioRender.com, accessed 12 April 2025).
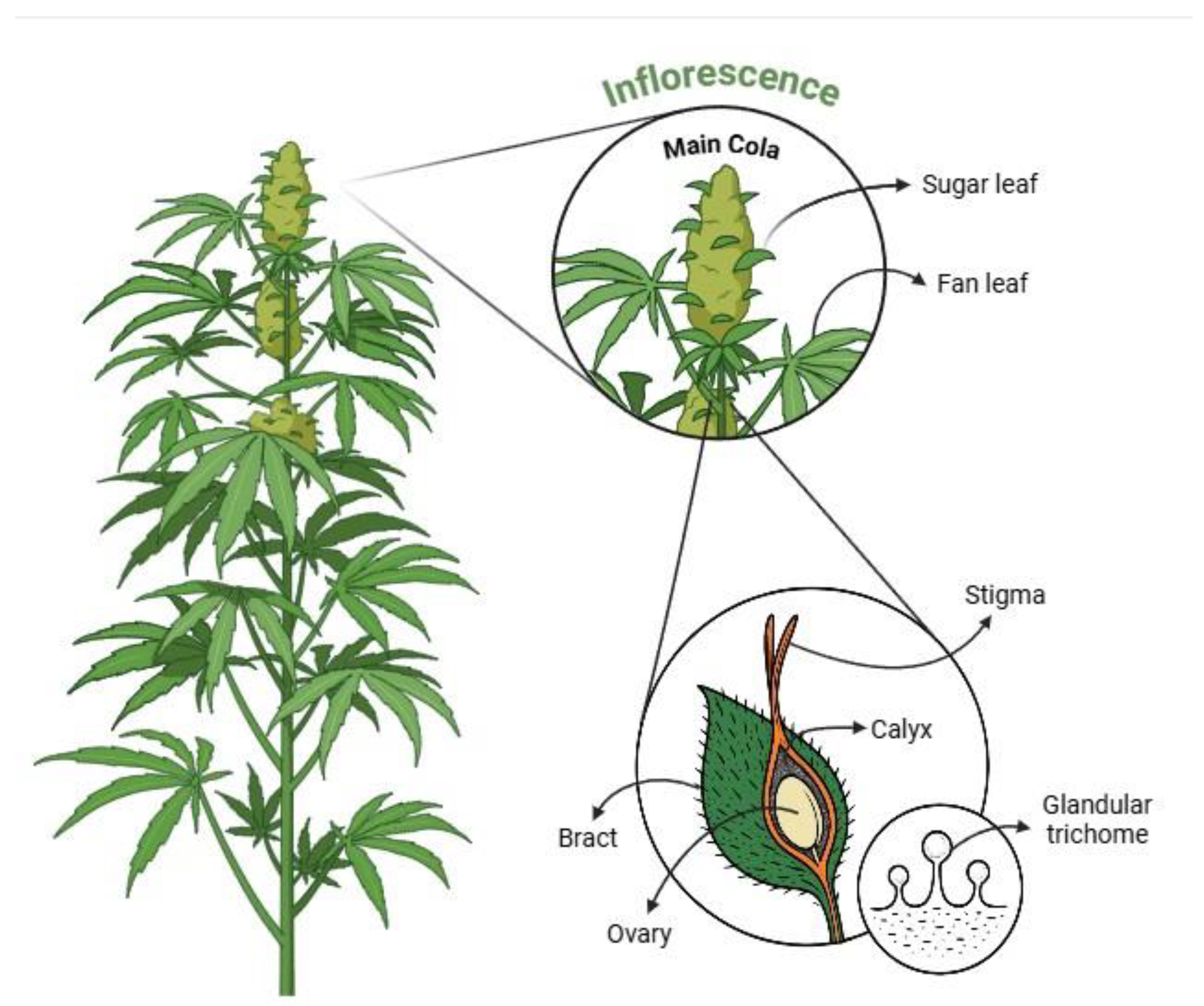
Figure 4.
Representative image of the main cola of Cannabis sativa L., illustrating the organization of the inflorescence, the morphology of sugar leaves, and sections of fan leaves extending from the floral cluster.
Figure 4.
Representative image of the main cola of Cannabis sativa L., illustrating the organization of the inflorescence, the morphology of sugar leaves, and sections of fan leaves extending from the floral cluster.

Figure 5.
Magnified detail of the resin accumulation on trichomes covering a dried female flower of Cannabis sativa L.
Figure 5.
Magnified detail of the resin accumulation on trichomes covering a dried female flower of Cannabis sativa L.

Figure 6.
Details of the resinous biomass produced by trichomes on Cannabis sativa’s sugar leaf adaxial surface.
Figure 6.
Details of the resinous biomass produced by trichomes on Cannabis sativa’s sugar leaf adaxial surface.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



