Submitted:
27 April 2025
Posted:
28 April 2025
You are already at the latest version
Abstract
Background: The ethnobotanical use of medicinal plants is a widespread practice worldwide, serving various therapeutic and practical purposes due to their phytocompounds. This study investigates the chemical composition, antioxidant capacity, and in vitro ruminal fermentation properties of four plant species: Polypodium vulgare, Chamaemelum nobile, Ocimum forsskaolii Benth, and Lavandula stoechas. Methods: The chemical composition, including insoluble dietary fiber and phenolic compounds, was determined. Antioxidant activity was assessed using DPPH, and HPLC identified key bioactive compounds. In vitro fermentation trials evaluated gas production, metabolizable energy (ME), organic matter digestibility (OMd), and volatile fatty acid (VFA) production in sheep and goats. Results: All species contained high levels of insoluble dietary fiber, with L. stoechas exhibiting the highest neutral detergent fiber (62.39%) and hemicellulose (56.74%). C. nobile had the highest polyphenol and flavonoid levels (73.88 ± 0.79 mg GAE/g DW and 27.85 ± 0.54 mg QE/g DW, respectively), along with strong antioxidant activity (IC₅₀ = 0.38 mg/mL). HPLC identified catechol in P. vulgare, ferulic acid in C. nobile, chlorogenic acid in O. forsskaolii, and apigenin in L. stoechas as major compounds. In vitro fermentation showed that C. nobile and O. forsskaolii at D20 produced the highest gas volume. ME, OMd, and VFA production increased, with a greater effect in sheep than in goats. Conclusion: These findings highlight the potential of these plant extracts, rich in bioactive compounds, to offer a sustainable alternative to conventional feed additives by enhancing ruminal fermentation, energy efficiency, and livestock performance.
Keywords:
medicinal plant extracts
; biochemical composition
; phenolic compounds
; fermentation
; ruminants
1. Introduction
The ethnobotanical use of medicinal plant species is an ancient practice worldwide, particularly in economically disadvantaged communities and remote regions. In recent decades, research on natural compounds has significantly accelerated, focusing on the extraction, analysis, and integration of these phytocompounds for potential applications in the cosmetic, pharmaceutical, and agri-food industries. However, their use in animal nutrition remains limited, despite their potential as natural antioxidants, which could improve the quality of animal products and contribute to human health. Beyond their nutritional and health benefits, certain medicinal plants may also play a role in environmental sustainability. Livestock is responsible for about 12% of global greenhouse gas (GHG) emissions, primarily due to methane production during ruminal fermentation [1]. Addressing this issue has become a key focus in climate change mitigation strategies, with increasing attention on managing ruminal methane emissions [2]. In small ruminants, effective microbial digestion is essential to meet the nutritional needs of high-production animals. However, this digestion remains partially incomplete, leading to nitrogen losses (mainly in the form of urine and milk) and energy losses in the form of methane, with significant environmental and economic consequences. These losses highlight the urgency of optimizing digestive processes to improve animal performance while reducing emissions [3]. However, current strategies such as the use of chemical additives or dietary modifications have shown limited results in terms of long-term effectiveness and environmental impact [4]. In this context, the integration of medicinal plants into animal feed is emerging as a promising alternative, addressing multiple challenges, including food security, greenhouse gas (GHG) reduction, and the adaptation of livestock systems to climate change [5]. Research has shown that these plants can regulate ruminal function, lower methane emissions, modulate ruminal fermentation, and improve digestive efficiency in ruminants [6].
The Tunisian flora, comprising 2103 species across 115 families and 742 genera, represents an underexploited resource with the potential to offer innovative applications. This potential is further emphasized by its traditional ethnobotanical uses, particularly in veterinary care and herbal medicine, which highlight its broad utility [7]. The growing use of these plants can be attributed to their richness in bioactive compounds with antimicrobial, antiparasitic, and anti-inflammatory properties, which can play a vital role in improving ruminant health and boosting livestock performance.
Among these, P. vulgare (Polypodiaceae), known for its anti-inflammatory properties, has been traditionally used to treat respiratory ailments, digestive disorders, and skin diseases in human medicine [8]. C. nobile (Asteraceae), valued for its digestive and calming effects, has been widely consumed as an infusion to relieve gastrointestinal discomfort, anxiety, and skin inflammations [9]. O. forsskaolii Benth (Lamiaceae), recognized for its antimicrobial and analgesic properties, has been employed in folk medicine to combat infections, support digestion, and alleviate pain [10]. L. stoechas (Lamiaceae), appreciated for its antiseptic and expectorant effects, has been extensively used in treating respiratory conditions, nervous disorders, and skin infections, while also being incorporated into cosmetic applications [11]. Given their well-documented medicinal properties in humans, these plants have also attracted interest for their potential applications in animal health and nutrition. In this context, the present study aims to characterize these four medicinal species (P. vulgare, C.nobile, O. forsskaolii, and L. stoechas) by analyzing their biochemical properties and assessing their digestibility through kinetic indicators of gas production during in vitro ruminal fermentation. This study aims to investigate the potential of these plant extracts to enhance ruminal fermentation, providing a natural and sustainable alternative to conventional livestock feeding practices.
2. Resultats and Discussion
2.1. Dry Matter, Ash and Dietary Fiber Content
The results in Table 1 show that all parameters varied significantly (p ≤ 0.05) among the plant species. Notably, L. stoechas exhibited the highest amounts of dry matter and ash relative to the other species. Furthermore, the analysis of insoluble dietary fibers assessed the levels of NDF, ADF, ADL, HC, and CB. The results indicated that the studied plants were relatively rich in dietary fiber. Specifically, insoluble fiber levels across all plants ranged as follows: NDF (41.04–62.39%), ADF (23.94–40.22%), ADL (5.65–14.58%), HC (33.65–56.74%), and CC (12.55–34.5%). Except the NDL fraction, L. stoechas has the highest level of all insoluble fibers parameters, and the NDF and HC were the dominant fractions with values of 62.39± 0.71 and 56.74± 0.87% DW, respectively. However, the obtained results were consistent with those reported for similar species, other Asteraceae, and Mediterranean shrubs [12,13]. The observed variation among results may be attributed to diverse geographical location, seasonal conditions, and the maturity stage of the sampled plants.
On the other hand, these results highlight that the studied species can be considered a potential source of insoluble fibers. These fibers play a crucial role in digestive function, rumen health, and metabolism regulation. They contribute to improved digestion, support energy production through fermentation, and influence the quality of animal-derived products [14]. In livestock farming, ensuring an appropriate fiber intake is essential, taking into account the animal type (ruminant or monogastric), its developmental or production stage, and its specific nutritional requirements [15].

Table 1.
Dry matter, ash and insoluble dietary fibers content (%) in aerial part of P. vulgare; C.nobile, O. forsskaolii and L. stoechas.
Table 1.
Dry matter, ash and insoluble dietary fibers content (%) in aerial part of P. vulgare; C.nobile, O. forsskaolii and L. stoechas.
| Insoluble fiber fraction | Content (%) | |||
|---|---|---|---|---|
| P. vulgare | C. nobile | O. forsskaolii | L. stoechas | |
| DM Ash NDF ADF ADL HC CC |
24.25 ± 0.04b 4.55 ± 0.01ab 48.23 ±0.70b 27.13± 0.11 b 14.58 ± 0.06a 33.65 ± 0.63b 12.55 ± 0.18c |
21.46 ± 0.03c 3.99 ± 0.01b 41.04± 0.99d 23.94 ±0.24c 6.72± 0.05bc 34.32 ±0.94b 17.22 ±0.19b |
19.87 ± 0.17c 3.67± 0.03b 43.48 ±0.41cd 26.46± 1.01b 7.92 ±0.50 b 35.56 ±0.09b 18.54 ± 0.50b |
27.49 ± 0.12a 6.11 ± 0.02a 62.39 ±0.71a 40.22±1.12a 5.65 ± 0.15c 56.74± 0.87a 34.5 ± 0.97a |
In the same line, means followed by different letters are significantly different using Duncan's multiple range test at p ≤ 0.05), data are means ± SD of three replicates. DM: Dry matter, NDF: neutral detergent fibre, ADF: acid detergent fibre; ADL: acid detergent lignin; HC: hemicellulose; CC : crude cellulose.
2.2. Extraction Yield, Phenolic Compounds' Content, and DPPH Activity
As shown in Table 2, all parameters varied significantly among the species (p ≤ 0.05). The highest yields were recorded for C. nobile (16.22±0.41), O. forsskaolii (17.04±0.85%), and L stoechas (16.44±0.62), while P. vulgare exhibited the lowest yield (12.42±0.37%). Among the studied plants, C. nobile and L. stoechas exhibited the highest levels of polyphenols (73.88 ± 0.79 and 72.69 ± 0.05 mg GAE/g DW, respectively). Whereas, P. vulgare showed the lowest polyphenol content (30.45 ± 0.26 mg GAE/g DW). Similarly, flavonoid levels varied significantly, with C. nobile showing the highest content (27.85 ± 0.54 mg QE/g DW), followed by L. stoechas and O. forsskaolii (20.11 ± 0.12 and 19.25 ± 0.20 mg QE/g DW, respectively). In contrast, P. vulgare presented the lowest flavonoid level (9.03 ± 0.24 mg QE/g DW).
Previous studies have demonstrated the variability in total phenolic content among these plant species. Indeed, L stoechas exhibited a notably high phenolic content of 130.15 mg GAE/g of plant extract [16]. In contrast, C. nobile contained 1.72 mg/g DW in green extracts [17]. Similarly, O. forsskaolii was reported to have a phenolic content of 0.126 mgGAE/gDW [18]. The variability in phenolic compound levels observed in comparison to our results can be attributed to ecological and environmental factors, as well as differences in cultivation conditions, including the plant's phenological stage and extraction methodologies.
The antioxidant test of plant extracts showed a significant difference (p≤0.01) among the species, and the strongest antioxidant activities were recorded for O. forsskaolii (IC₅₀ = 0.36 mg/mL), C. nobile (IC₅₀ = 0.38 mg/mL), and L. stoechas (IC₅₀ = 0.40 mg/mL) (Table 2). In contrast, P. vulgare showed weaker effect with an IC₅₀ value of 1.79 mg/mL.The present findings demonstrated that the potent IC50 values observed can be explained by the significant concentrations of the phenolic compounds in the studied plants. Previous studies have reported a strong correlation between antioxidant activity and phenolic content, identifying these compounds being recognized as key indicators of antioxidant potential [19,20].
Table 2.
Yield, phenolic compounds and antioxydant activity of P. vulgare, C.nobile, O.forsskaolii and L. stoechas.
Table 2.
Yield, phenolic compounds and antioxydant activity of P. vulgare, C.nobile, O.forsskaolii and L. stoechas.
| Assay | P. vulgare | C. nobile | O. forsskaolii | L. stoechas |
|---|---|---|---|---|
| Yield (%) Polyphenols (mg GAE/g DW Flavonoids (mgQE/g DW DPPH IC50 (mg/mL) |
12.42±0.37b 30.45±0.26c 9.03±0.24c 1.79±0.08a |
16.22±0.41a 73.88±0.79a 27.85±0.54a 0.38±0.002b |
17.04±0.85a 60.70±0.62b 19.25±0.20b 0.36± 0.006b |
16.44±0.62a 72.69±0.05a 20.11±0.12b 0.40±0.003b |
In the same line, means followed by different letters are significantly different using Duncan's multiple range test at p ≤ 0.05), data are means ± SD of three replicates.
2.3. Compounds Profile Identified in the Aqueous Extract of P. vulgare; C. nobile, O. forsskaolii and L. stoechas
The HPLC analysis of the aqueous extracts from the studied plant species (Figure 1) revealed the presence of 11 compounds in P. vulgare, 13 in C. nobile, 9 in O. forsskaolii, and 12 in L. stoechas, classified into different phenolic acids and flavonoids (Table 3). Furthermore, in P. vulgare, catechol and ellagic acid were identified as the major compounds, accounting for 22.12% and 20.59%, respectively. In C. nobile, ferulic acid (10.42%) and apigenin-7-O-glucoside (9.39%) were the predominant compounds. Similarly, chlorogenic acid (23.53%) and ascorbic acid (14.22%) were the major constituents in O. forsskaolii. For L. stoechas, apigenin (21.40%) and chlorogenic acid (10.30%) were the dominant compounds. However, previous analyses have reported that 3-O-caffeoylquinic acid [8], quercetin and rutin [21], caffeic acid and quercetin [10], and vanillic acid [22] were the most abundant compounds in P. vulgare, C. nobile, O. forsskaolii, and L. stoechas, respectively.These compounds have been identified as the primary contributors to the biological activities of these species.
2.4. Effects of Plant Extracts on the In Vitro Kinetics of Gas Production
The kinetic factors of in vitro fermentation, determined using the exponential model of Orskov and Macdonald (1979) [23], were presented in Figure 2 and Table 4. This model describes the fermentation process in three distinct phases: the lag phase (a), during which gas production remains minimal as rumen microbes adapt to the substrate; the exponential growth phase (b), characterized by rapid fermentation of the fermentable substrate fraction, leading to a consistent increase in gas production; and the plateau phase (c), where gas production slows and stabilizes, reflecting the rate of substrate degradation during this phase. All these parameters are crucial for assessing substrate digestibility and fermentation efficiency.
As shown in Table 4, gas production was mainly influenced by the degradation rate and the characteristics of the carbohydrates in the feed, as well as by variation in plants species and extract doses.
The soluble fraction “a” showed significant variation between animals. In sheep, this parameter ranged from 1.49 ± 0.01 to 2.73 ± 0.01 mL/0.03 DW at D20, whereas in goats, the average values were generally lower, ranging from 1.01 ± 0.05 to 1.58 ± 0.06 mL/0.03 DW at the same dose (D20). These results are in agreement with those reported by Freitas et al., (2024) [24], who observed increases in the soluble fraction parameter in response to plant extract doses in other ruminant species. Additionally, the fraction of potentially available fermentable substrate, “b” varied significantly depending on plant species and doses. The highest values were recorded for the C. nobile extract at D20, with 261.63 ± 0.45 mL/0.03 DW in sheep and 177.66 ± 0.11 mL/0.03 DW in goats. The influence of dose has been documented in several studies, including Gonzalez Ronquillo et al. (2023) [25], who observed that increasing the dose of plant extract resulted in a greater fraction of available fermentable substrate. On the other hand, the rate of substrate degradation “c” varied among plant species and doses, the heighest values were observed for O. forsskaolii extract with 0.055 h-¹ at D20 in sheep.
Furthermore, the highest gas production values were recorded with C. nobile (42 ± 2.26 – 37 ± 1.46 mL/µL extract) and O. forsskaolii (39.5 ± 1.12 – 33 ± 1.15 mL/µL extract) at D20 in sheep and goats, respectively.
Previous studies found that plant extracts can influence the rate of substrate degradation in some ruminants depending on their bioactive compound content [24]. Particularly, flavonoid-rich extracts can stimulate specific microbial populations responsible for rapid substrate degradation and gas production in the rumen [26,27]. Messaoudi et al. (2018) [28] also confirmed that increasing the dose of plant extract improves fermentation speed due to the activation of microbial processes in the rumen. These results reinforce the idea that bioactive plant compounds playa key role in optimizing ruminal fermentation processes and improving substrate digestibility in ruminants.
Table 4.
Effect of Aqueous Extracts from P. vulgare, C. nobile, O. forsskaolii, and L. stoechas on in Vitro digestibility parameters in Sheep and Goats.
Table 4.
Effect of Aqueous Extracts from P. vulgare, C. nobile, O. forsskaolii, and L. stoechas on in Vitro digestibility parameters in Sheep and Goats.
| Animals | Plants | Doses (µL) | a (mL/DW) |
b (mL/DW) |
c (mL/h) |
gas production (mL at 24 h) | gas production (mL at 48 h) | ||
|---|---|---|---|---|---|---|---|---|---|
|
Sheep |
P. vulgare C. nobile O. forsskaolii L. stoechas |
D0 D10 D20 D0 D10 D20 D0 D10 D20 D0 D10 D20 |
1.98±0.05a 1.15±0.05b 1.49± 0.01b 1.49±0.01b 2.27±0.02a 2.73±0.01a 1.49±0.05b 1.65±0.07b 2.33±0.02a 1.98±0.05a 1.17±0.01b 1.83±0.05a |
104.1 ±0.08b 198.5±1.15a 105 ±0.95b 140.3 ±1.05c 256.43 ±0.46b 261.63 ±0.45a 140.3 ±1.05b 220.23 ±0.20a 219.7 ±0.17a 104.1±0.22b 85.06 ±0.31c 121.53±0.57a |
0.022b 0.032a 0.019b 0.016b 0.022a 0.018ab 0.028c 0.048b 0.055a 0.016 bc 0.022a 0.014 c |
23±1 ±1.08a 18.5 ±2.24b 15.5 ±2.16c 13.5 ±2.64b 24.5±2.06a 23.5 ±1.86a 13.5 ±2.12c 23.5 ±2.26b 26 ±1.18a 23 ±1.46a 21.5 ±1.76b 22 ±1.23ab |
34 ±0.56a 32 ±0.98ab 30 ±1.46b 31 ±2.51b 41 ±1.48a 42 ±2.26a 27 ±2.7c 35.5 ±0.98b 39.5 ±1.12a 33 ±1.8ab 34 ±2.7ab 35 ±2.3a |
||
| Plant effect Dose effect |
0.0038 0.0064 |
0.0006 0.0081 |
<0.0001 0.0067 |
<0.0001 <0.0001 |
<0.0001 <0.0001 |
||||
|
Goats |
P. vulgare C. nobile O. forsskaolii L. stoechas |
D0 D10 D20 D0 D10 D20 D0 D10 D20 D0 D10 D20 |
0.67±0.02b 0.83±0.03b 1.50±0.01a 1.36±0.03a 1.23±0.02a 1.01±0.05a 1.36±0.03a 1.06±0.21a 1.01±0.07a 1.07±0.09b 1.58±0.08a 1.58±0.06a |
93.36±0.08b 78.31±0.04c 165.83±0.98a 67.71±0.01c 172.56±0.23b 177.66±0.11a 67.71 ±0.22b 100.9±0.17a 33.63 ±0.41c 93.36 ±0.89bc 91.6 ±1.71c 96.69±0.64a |
0.072a 0.049b 0.022c 0.012b 0.004ab 0.005a 0.032a 0.028b 0.012c 0.028a 0.020b 0.022b |
14.5 ±0.89a 14.5 ±1.48a 12.5 b±1.16b 16.5 ±1.56c 18.5±1.76b 20±2.16a 16.5±1.34b 19±1.56a 17.5±1.84ab 14.5±2.06a 12.5±2.46b 12 ±1.42b |
25±1.68a 26±1.86a 22.5±2.13b 28 ±1.86c 35 ±2.49b 37±1.46a 27 ±1.56b 32 ±1.27a 33 ±1.15a 27±0.86a 26.5±1.96ab 25 ±1.92b |
||
| Plant effect Dose effect |
<0.0001 0.0006 |
0.002 0.0037 |
0.0002 <0.001 |
<0.0001 <0.0001 |
<0.0001 <0.0001 |
||||
Data are means ± SD, for each parameter and each plant species, the means followed by different letters in the same column are significantly different at p≤0.05. (a) and (b) represent the volume of gas produced from the soluble and insoluble fractions of the feed, respectively,(c) represents the fermentation rate. D0=0µL; D10=10µL and D20=20 µL of extract.
2.5. In Vitro Impact of Plant Extracts on Ruminal Fermentation Parameters
As shown in Table 5, a significant effect of plant species and extract doses (p≤ 0.05) was observed in sheep for metabolic energy (ME) production, organic matter digestibility (OMd), and volatile fatty acid (VFA) production. However, no significant effect (p>0.05) was obtained for the same parameters in goats. Also, for all parameters, the results indicated no significant difference between the D10 and D20 from all plant species in sheep, which showed the highest values compared to D0. However, in goats, no significant differences were observed across all doses for all parameters. In contrast, sheep showed the highest values across all fermentation parameters following the addition of the extract, with ranges of 5.26±0.4 to 5.88±0.13 MJ/kg DW for ME, 35.60±3.11 to 42.93 ±1.33% for OMd, and 0.41±0.03 to 0.56±0.2 mmol for VFA. In comparison, goats displayed lower values, ranging from 4.04±1.79 to 5.04 ±0.06 MJ/kg DW for ME, 27.15±1.77 to 34.27±5.33% for OMd, and 0.22±0.05 to 0.40±0.16 mmol for VFA.
The higher values of metabolizable energy (ME), organic matter digestibility (OMd), and volatile fatty acids (VFA) observed in sheep can be attributed to their superior digestive efficiency, likely due to their more homogeneous ruminal microbiota, as suggested by Moghaieb et al. (2020) and Ferchichi et al. (2019) [29,26]. These studies observed that sheep respond more positively to plant extracts in terms of ruminal fermentation and VFA production. However, goats appear to be less influenced by these extracts, as indicated by the work of Khelil et al. (2020) [30] and Moghaieb et al. (2020) [29], who reported less pronounced effects in goats, suggesting a species-specific difference in digestive responses.
On the other hand, research carried out by Khouja and Boussaid (2013) [31] have showed that the integration of medicinal plants in the diet of small ruminants in arid environments, such as mugwort and rosemary, not only improves the health of the animals in general, but also reduces the parasite load. However, these effects are more significant in sheep than in goats, probably due to differences in their intestinal microbiota and digestive system.
Furthermore, the improvement in ruminal fermentation can be attributed from the synergistic effects of insoluble fibers and bioactive compounds, such as phenolic acids and flavonoids, present in the studied plants. Insoluble fibers, including cellulose and hemicellulose, serve as essential substrates for ruminal microorganisms, promoting the production of volatile fatty acids (VFAs), a key energy source for ruminants [32]. However, fermentation efficiency is also influenced by the microbial composition of the rumen, which bioactive compounds can modulate by stimulating beneficial microorganisms.
Among these bioactive compounds, caffeic, ferulic, and chlorogenic acids, as well as flavonoids like luteolin and quercetin, enhance fiber degradation by promoting cellulolytic bacteria, leading to increased VFA production, particularly propionate, a highly energy-efficient metabolite for ruminants [33] [34]. Besides, ellagic acid and myricetin contribute to ruminal microbial balance by stimulating cellulolytic bacteria while inhibiting methanogenic microorganisms. This dual effect not only enhances fiber breakdown but also reduces methane emissions, an undesirable byproduct of ruminal fermentation, thereby improving overall energy efficiency [33] [35]. In addition to the previously mentioned bioactive compounds, certain flavonoids, such as apigenin and apigenin-7-O-glucoside, play a crucial role in modulating ruminal fermentation dynamics. These molecules not only influence microbial composition but also enhance fermentation efficiency by promoting beneficial microorganisms and regulating gas production [36].
In summary, the observed effects from each of the studied plant tests likely resulted from a synergy between bioactive compounds, such as phenolic acids and flavonoids, and insoluble fibers. This interaction enhances cellulolytic bacterial activity, modulates the ruminal microbial population, and promotes the degradation of complex fibers. Consequently, this process leads to increased VFA production and improved energy efficiency of ruminal fermentation, ultimately supporting better nutritional performance in ruminants.
Table 5.
Effect of aqueous extracts from P. vulgare, C. nobile, O. forsskaolii, and L. stoechas on metabolizable energy, organic matter digestibility, and volatile fatty acid production in Sheep and Goats.
Table 5.
Effect of aqueous extracts from P. vulgare, C. nobile, O. forsskaolii, and L. stoechas on metabolizable energy, organic matter digestibility, and volatile fatty acid production in Sheep and Goats.
| Plants | Doses | ME (MJ /kg DW) | OMd (%) | VFA (mmol) | |||
|---|---|---|---|---|---|---|---|
| Sheep | Goats | Sheep | Goats | Sheep | Goats | ||
|
P. vulgare C. nobile O. forsskaolii L. stoechas |
D0 D10 D20 D0 D10 D20 D0 D10 D20 D0 D10 D20 |
3.86 ±0.41b 5.47 ±0.2a 5.45 ±0.2a 4.18±0.76b 5.54 ±0.34a 5.54 ±0.34a 4.18 ±0.06b 5.54 ±0.87a 5.88 ±0.13a 4.22 ±0.4b 5.26 ±0.47ab 5.33±0.4a |
3.31±0.06b 4.31±0.06a 4.04±0.06ab 5.08±0.74a 4.67±1.94a 4.12 ±1.79a 4.58 ±0.34a 4.92 ±0.95a 4.77 ±0.88a 4.31±0.06a 5.04 ±0.06a 4.97 ±0.27a |
36.93 ±2.66b 42.93 ±1.33a 40.27 ±1.33ab 28.49 ±0.44b 37.38 ±2.22a 37.38±2.22a 28.49 ±0.44b 37.38±3.11a 39.60±0.88a 30.93±2.66b 35.60±3.11a 36.04±2.66a |
29.38±0.44a 29.38±0.44a 27.6±0.44a 31.15±2.22a 32.93±5.77a 34.27±5.33a 31.15±2.22a 33.38±6.22a 32.04±7.55a 29.38±0.44a 27.6±0.44a 27.15 ±1.77a |
0.38 ±0.03b 0.48±0.07a 0.41±0.03b 0.26±0.01b 0.50±0.06a 0.50±0.06a 0.26 ±0.01b 0.50±0.08a 0.56±0.2a 0.42±0.07b 0.57±0.14a 0.56 ±0.07a |
0.28±0.01a 0.28±0.01a 0.23±0.01a 0.33±0.05b 0.38 ±0.15ab 0.40±0.16a 0.33±0.06a 0.39±0.17a 0.35±0.20a 0.28±0.011a 0.23±0.14a 0.22±0.05a |
| Plant effect Dose effect |
0.028 0.0476 |
0.1602 0.6423 |
0.0155 0.0307 |
0.4968 0.101 |
0.0192 0.04 |
0.4968 0.101 |
|
Data are means ± SD, for each parameter and each plant species, the means followed by different letters in the same column are significantly different at p≤0.05. VFA: volatile fatty acid, OMd :Organic matter digestibility, ME : Metabolizable energy, D0=0µL ; D10=10µL and D20=20 µL of extract.
3. Materiel and Methods
3.1. Plant Material
Four spontaneous medicinal plants (P. vulgare, C. nobile, O. forsskaolii and L. stoechas) from scrubland were collected from the Aindrahim area in northwestern Tunisia in the springtime, during the flowering phase. The aerial parts of each plant were harvested and transported to the laboratory. Species identification was performed by Professor Marouani Ahmed from the Higher School of Agriculture of Kef (ESAK, Tunisia), with voucher specimens (PAr1, CAr2, OAr3, and LAr4) stored for future reference within the Herbarium of the Laboratory of Functional Physiology and Bio-Resource Valorization at the Higher Institute of Biotechnology of Béja. Before conducting the analyses, the samples were air-dried at room temperature for twenty days.
3.2. Animal Material
Fresh continuous rumens from sheep and goats were collected at the Tabarka municipal slaughterhouse.
3.3. Phytochemical Analysis
3.3.1. Dry Matter, Ash and Dietary Fiber Determination
Dry matter content was quantified by drying 1 g of fresh sample at 105°C until a constant weight was obtained.The ash content was determind after calcination of 1g from each sample at 550 °C in a muffle furnace for 4 hours, until complete combustion. Dietary fiber was analyzed following the method outlined by Van Soest et al. (1991) [37] using a FIBRESTEST RAYPA apparatus. Neutral detergent fiber (NDF) was assessed by dissolving cellular contents with sodium lauryl sulfate detergent and pectic substances were solubilized with a chelating agent in a buffered solution. This fraction predominantly includes hemicelluloses, cellulose and lignin. Acid Detergent Lignin (ADL) was measured by treating the acid detergent fiber (ADF) fraction, which consists of lignin and true cellulose, with sulfuric acid. Hemicellulose content was determined by subtracting the ADF from the NDF. Each sample was tested in triplicate.
3.3.2. Preparation and Yield of Plant Extracts
The aerial parts of each plant were extracted in distilled water with a 1:10 (w/v) ratio for 48 hours in the dark. After filtration through Whatman No. 4 filter paper, the extracts were freeze-dried using a LABCONCO freeze-dryer (USA). To ensure maximum yield, the extraction was repeated three times on the same plant residue. The extracts were stored in glass tubes at 4°C, away from light. The yield (%) of dried extracts was calculated as DWextr/DWsamp x 100, where DWextr is the weight of extract after freeze-drying, and DWsamp is the dry weight of sample.
3.3.3. Total Polyphenols Détermination
The polyphenol content was estimated according to Olsen and Sommer’s (1982) [38] method, with the Folin-Ciocalteu reagent. Absorbance was read at 760 nm using a UV-Vis spectrophotometer (Jenway 7205, UK), using a blank for reference and gallic acid as a standard (DO=0.003C+0.102; R2=0.971). The assay was conducted in triplicate, and the total polyphenol content was represented as mg of gallic acid equivalent/g of dry weight (mg GAE/g DW) of the plant material.
3.3.4. Total Flavonoids Determination
The flavonoid content was assessed using the method outlined by Tajini et al. (2020)[39]. Quercetin was used as a positive control (DO = 0.372C- 0.011; R² = 0.984), and the absorbance was read at 510 nm with a UV-visible spectrophotometer (Jenway 7205, UK), using a blank for calibration. The assay was performed in triplicate, and the flavonoid content was determine as mg of quercetin equivalent /g of dry weight (mg QE/g DW) of the plant material.
3.3.5. Antioxidant Activity
The evaluation of DPPH radical-scavenging assay was assessed using a method adapted from Sarr et al. (2015) [40], and the activity was measured accordingly to the following equation: % DPPH radical-scavenging effect = [(C0-Cs)/C0] x 100, where C0 and Cs are the concentration values of the (DPPH) solution before and after adding the extract sample, respectively. C0 and Cs were determined according to a calibration curve, and the absorbance at 517 nm was measured against a control, carried out with methanolic DPPH solution. The extract concentration required to inhibit 50% of the DPPH radicals (IC50) was determined using linear regression of the inhibition percentages. The IC50 assay was conducted in triplicate, and gallic acid (GA) was applied as a standard.
3.3.6. HPLC Analysis of Plant Aqueous Extracts
Phenolic compounds were identified using high-performance liquid chromatography (Agilent 1100 series HPLC system, Germany) equipped with a UV-vis multi-wavelength detector, following the protocol described in the study by Falleh et al. (2008) [41]. The identification process involved comparing the retention times and the mass spectra with those of authentic standards.
3.4. Evaluation of Gas Production In Vitro
The fermentation of substrates in the rumen was evaluated using the in vitro gas production method, outlined by Jedidi et al. (2019) [42]. Briefly, rumen fluid was obtained from sheep and goats at the municipal slaughterhouse in Tabarka. The collected material was blended, filtered using four layers of surgical gauze to eliminate any solid residues, and kept in an insulated container to maintain rumen-like conditions. The filtered fluid was subsequently transferred in an industrial mixer and continuously exposed to CO₂ to ensure the maintenance of anaerobic conditions.
To prepare the fermentation medium, each 100 mL syringe was filled with a mixture of 10 mL of filtered rumen fluid, 20 mL of artificial saliva, and 0.3 g of ground oat hay as the base substrate. Additional syringes were prepared using the same components, with varying doses of plant extracts added (D0 = 0 µL, D10 = 10 µL, and D20 = 20 µL). Afte incubation at 39 °C in a water bath, the syringes were purged with CO₂ to maintain anaerobic conditions. To ensure uniformity, the syringes were gently agitated 30 minutes before measurements. Gas volumes were recorded by holding the syringes horizontally without moving the piston for precise readings. Incubation ended when gas production stabilized. Gas generation was modeled using the exponential equation: G(t)=a+b×(1−e−ct), where “a” represents the gas production of the soluble part of the substrate (ml), “b” represents the gas production of the insoluble part of the substrate (ml), “a + b” represents the potential total gas production (ml), that is, the asymptote of the curve, “c” represents the gas production rate (in ml/hour), reflecting the fermentation speed, and “t” represents the culture time (in hours).
3.5. In Vitro Fermentation Parameters
The metabolizable energy (ME), digestibility of organic matter (OMd), and total volatile fatty acids (VFA) production were estimated following the method described by Menke and Steingass (1988) [43]
OMd (%) = 14.88 + 0.889 GP + 0.45 MAT + 0.0651 MM
EM (MJ/kg MS) = 2.20 + 0.136GP + 0.057PB
VFA (mmol/syringe) = 0.0239.Gp -0.0601
GP: Volume of gas produced (ml/300 mg dry matter) after 24 hours incubation; MAT:Total nitrogenous matter (g/100 g dry matter) ; MM:Mineral matter (g/100 g dry matter).
3.6. Statistical Analysis
Data were statistically analyzed using univariate analysis of varaiance (ANOVA), and mean comparaisons were determined using Duncan's multiple range test at a significance level of p≤0.05, with SAS (1997) software.
4. Conclusion
The obtained results showed that all plant species were relatively rich in dietary fiber, with L. stoechas exhibiting the highest levels. Regarding phenolic compounds, C. nobile and L. stoechas exhibited the highest polyphenol levels, with C. nobile also showing the highest flavonoid content compared to other species. Furthermore, strong antioxidant activities were recorded, and several phenolic acids and flavonoids were identified by HPLC analysis in all plant species. In vitro substrate digestibility showed significant effects from plant species and extract doses, and the highest values of gas production were observed with C. nobile and O. forsskaolii at D20. Additionally, all parameters of fermentation (ME, OMd and VFA), showed significant improvement following the addition of plant extracts, particularly in sheep, demonstrating considerable potential for improving animal digestibility. This suggests that the extracts may promote faster and more efficient degradation of substrates in the rumen, potentially improving energy production and the animals' overall performance.
Looking ahead, these plants present promising solutions for sustainable livestock production systems. By offering natural alternatives to conventional feed additives, they not only contribute to improving the health and productivity of small ruminants but also promote a more environmentally friendly approach to animal nutrition. Future in vivo studies will be essential to confirm these findings and assess their long-term effects on animal health, oxidative stress, and overall performance. Additionally, evaluating their broader ecological benefits in sustainable farming practices and their potential role in zootechnical applications will be crucial for advancing the adoption of these plants in the livestock sector.
Author Contributions
Conceptualization, Monia Bouzazi and Hichem Sebai; Data curation, Monia Bouzazi, Houcine Selmi, Fatma Tajini and Abid Ouerghui; Formal analysis, Monia Bouzazi and Hichem Sebai; Investigation, Monia Bouzazi and Fatma Tajini; Methodology, Monia Bouzazi, Houcine Selmi, Abid Ouerghui and Hichem Sebai; Writing – original draft, Monia Bouzazi, Fatma Tajini, Abid Ouerghui and Hichem Sebai; Writing – review & editing, Monia Bouzazi, Fatma Tajini and Hichem Sebai.
Acknowledgments
The Ministry of Higher Education and Scientific Research, the Higher Institute of Biotechnology of Beja, and the Sylvo-Pastoral Institute of Tabarka are deeply acknowledged for their support.
Conflicts of Interest
The authors declare that there are no conflicts of interest concerning the research, authorship, or publication of this article.
References
- FAO. Estimations des émissions de gaz à effet de serre en agriculture : Un manuel pour répondre aux exigences de données des pays en développement; Organisation des Nations Unies pour l'alimentation et l'agriculture. 2015. https://openknowledge.fao.org/items/49ce77d7-499f-49f0-a254-e2210235eaa8.
- Agriculture Victoria. Livestock methane and nitrogen emissions; Agriculture Victoria. 2024. https://agriculture.vic.gov.au.
- Climate and Clean Air Coalition (CCAC). Enteric fermentation and methane reduction; CCAC Coalition. 2024. https://www.ccacoalition.org.
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substances. J. Anim. Sci. Biotechnol. 2017, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Tan, S.; Chen, S.; Wu, Y.; Yang, Y.; Li, H.; et al. Effects of fermented Yupingfeng on intramuscular fatty acids and ruminal microbiota in Qingyuan black goats. Anim. Sci. J. 2021, 92, 13554. [Google Scholar] [CrossRef]
- Aziz, S.; Ölmez, M.; Nazir, R.; Mobashar, M.; Iqbal, Z.; Naeem, M.I.; Muzaffar, H.A.; Saif, D.; Faizan, M.; Kamran, M. Herbal feed additives for ruminant nutrition. In Abbas, R.Z.; Akhtar, T., Asrar, R., Khan, A.M.A., Saeed, Z., Eds.; Complementary and Alternative Medicine: Feed Additives; Unique Scientific Publishers: Faisalabad, Pakistan, 2024; pp. 15–21. [Google Scholar] [CrossRef]
- Bouzazi, M.; Tajini, F.; Selmi, H.; Ouerghui, A.; Sebai, H. Ethno-Veterinary Survey on the Traditional Use of Medicinal Plants with Therapeutic Properties in the Northwestern Regions of Tunisia. IOSR J. Agric. Vet. Sci. 2025, 18, 1–9. [Google Scholar] [CrossRef]
- Farràs, A.; Mitjans, M.; Maggi, F.; Caprioli, G.; Vinardell, M. P.; López, V. Polypodium vulgare L. (Polypodiaceae) as a source of bioactive compounds: Polyphenolic profile, cytotoxicity, and cytoprotective properties in different cell lines. Front. Pharmacol. 2021, 12, 727528. [Google Scholar] [CrossRef] [PubMed]
- Sah, A.; Naseef, P.P.; Kuruniyan, M.S.; Jain, G.K.; Zakir, F.; Aggarwal, G. A Comprehensive Study of Therapeutic Applications of Chamomile. Pharmaceuticals 2022, 15, 1284. [Google Scholar] [CrossRef]
- Alharbi, A.A. In vitro and in vivo anticancer, anti-inflammatory, and antioxidant activity of Dhimran (Ocimum forsskaolii Benth) extract and essential oil on carbon tetrachloride-induced hepatotoxicity in mice. Kafkas Univ Vet Fak Derg 2024, 30(5), 729–740. [Google Scholar] [CrossRef]
- Domingues, P.; Silva, A.; Mendes, F.; Rodrigues, A. Vanillic acid in L. stoechas and its biological activity. J. Agric. Food Chem. 2023, 71(9), 2035–2045. [Google Scholar] [CrossRef]
- Dupont, J.; Martin, P. Study of insoluble fibers in Mediterranean plants: Comparison between Asteraceae. J. Medit. Bot. 2017, 24(3), 145–157. [Google Scholar]
- George, C.F.; Lawrence, N.; Brian, L.; David, R.M. Critical Factors in Determining Fiber Content of Feeds and Foods and Their Ingredients. J. AOAC Int. 2019, 102, 52–62. [Google Scholar] [CrossRef]
- Sebastian, D.M.; Kim, L.J.; Walter, G.; Sonja, de V. Insoluble fibers affect digesta transit behavior in the upper gastrointestinal tract of growing pigs, regardless of particle size. J. Anim. Sci. 2023, 101, 299. [Google Scholar] [CrossRef]
- Chuang, W.-Y.; Lin, L.-J.; Shih, H.-D.; Shy, Y.-M.; Chang, S.-C.; Lee, T.-T. The potential utilization of high-fiber agricultural by-products as monogastric animal feed and feed additives: A review. Animals 2021, 11, 2098. [Google Scholar] [CrossRef] [PubMed]
- Ez Zoubi, Y.; Bousta, D.; Farah, A. A phytopharmacological review of a Mediterranean plant: Lavandula stoechas L. Clin. Phytosci. 2020, 6, 9. [Google Scholar] [CrossRef]
- Polcaro, L.M.; Cerulli, A.; Montella, F.; Ciaglia, E.; Masullo, M.; Piacente, S. Chemical Profile and Antioxidant and Tyrosinase Inhibitory Activity of Chamaemelum nobile L. Green Extracts. Cosmetics 2024, 11, 94. [Google Scholar] [CrossRef]
- Patel, F.; Nainesh, R.M. Estimation of total phenolic content in selected varieties of Ocimum species grown in different environmental conditions. J. Pharmacogn. Phytochem. 2018, 7(5), 144–148. [Google Scholar]
- Zongo, E.; Busuioc, A.; Meda, R.N.T.; Botezatu, A.V.; Mihaila, M.D.; Mocanu, A.M.; Avramescu, S.M.; Koama, B.K.; Kam, S.E.; Belem, H.; et al. Exploration of the antioxidant and anti-inflammatory potential of Cassia sieberiana DC and Piliostigma thonningii (Schumach.) Milne-Redh, traditionally used in the treatment of hepatitis in the Hauts-Bassins region of Burkina Faso. Pharmaceuticals 2023, 16, 133. [Google Scholar] [CrossRef]
- Tajini, F.; Jelassi, A.; Hamdani, A.; Salem, A.; Abdelhedi, O.; Ouerghui, A.; Sebai, H. Antioxidant Activities and Laxative Effect of Bioactive Compounds from Cynara cardunculus var. sylvestris. Sains Malays. 2024, 53(7), 1617–1630. [Google Scholar] [CrossRef]
- Dai, X.; Zhang, Y.; Chen, W.; Liu, J. Antioxidant and anti-inflammatory properties of quercetin and rutin in C. nobile extracts. nobile extracts. Phytochem. Rev. 2023, 22(2), 223–235. [Google Scholar] [CrossRef]
- Ndhlala, A.R.; Işık, M.; Kavaz Yüksel, A.; Dikici, E. Phenolic Content Analysis of Two Species Belonging to the Lamiaceae Family: Antioxidant, Anticholinergic, and Antibacterial Activities. Molecules 2024, 29(2), 480. [CrossRef]
- Orskov, E.R.; Macdonald, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. Camb. 1979, 92, 499–502. [Google Scholar] [CrossRef]
- Freitas, R.S.X.d.; da Silva, J.S.; Furtado, A.J.; Perna Junior, F.; de Oliveira, A.L.; Bueno, I.C.d.S. The production of Marandu grass (Urochloa brizantha) extracts as a natural modifier of rumen fermentation kinetics using an in vitro technique. Fermentation 2024, 10, 447. [Google Scholar] [CrossRef]
- Gonzalez Ronquillo, M.; Ghavipanje, N.; Sainz-Ramírez, A.; Danaee Celis-Alvarez, M.; Andrea Plata-Reyes, D.; Robles Jimenez, L.E.; Vargas-Bello Perez, E. Effects of plant extracts on in vitro gas production kinetics and ruminal fermentation of four fibrous feeds: towards sustainable animal diets. Chil. J. Agric. Anim. Sci. 2023, 39(3), 319–326. [Google Scholar] [CrossRef]
- Ferchichi, A.; Ben Salem, H.; Makkar, H.P.S.; Fodelianakis, S. Influence of phenolic-rich plants on in vitro ruminal fermentation and methane production. Anim. Feed Sci. Technol. 2019, 248, 138–146. [Google Scholar] [CrossRef]
- Huang, Q.; et al. Effects of plant polyphenols on rumen fermentation and microbial composition: A review. J. Anim. Sci. 2020, 6(4), 481–493. [Google Scholar] [CrossRef]
- Messaoudi, Z.; El Gharbi, S.; Khammassi, S. Antioxidant and antimicrobial properties of Lavandula stoechas extracts and their effect on ruminant digestion. Environ. Toxicol. Pharmacol. 2018, 63, 21–28. [Google Scholar] [CrossRef]
- Moghaieb, R.; El-Hack, M.E.A.; Abdelnour, S.A.; Rady, M.O. Incorporation of plant extracts into ruminant diets: Effects on fermentation and digestibility. J. Appl. Anim. Res. 2020, 48(1), 45–55. [Google Scholar] [CrossRef]
- Khelil, H.; Amrouche, N.; Ben Salem, H. Impact of plant secondary metabolites on ruminal fermentation and productivity in small ruminants under hot climatic conditions. Afr. J. Agric. Res. 2020, 15(8), 1184-1191. [CrossRef]
- Khouja, M.L.; Boussaid, M. Use of medicinal plants for small ruminants in arid regions. J. Arid Environ. 2013, 96, 1–10. [Google Scholar]
- Wang, L.; Zhang, G.; Li, Y.; Zhang, Y. Effects of high forage/concentrate diet on volatile fatty acid production and the microorganisms involved in VFA production in cow rumen. Animals 2020, 10, 223. [Google Scholar] [CrossRef]
- Antonius, A.; Pazla, R.; Putri, E.M.; Alma'I, M.I.; Laconi, E.B.; Diapari, D.; Jayanegara, A.; Ardani, L.R.; Marlina, L.; Purba, R.D.; Gopar, R.A.; Negara, W.; Asmairicen, S.; Negoro, P.S. Effects of herbal plant supplementation on rumen fermentation profiles and protozoan population in vitro. Vet. World 2024, 17(5), 1139–1148. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Y.; Wang, X.; Dai, C.; Tang, W.; Li, J.; Huang, P.; Li, Y.; Ding, X.; Huang, J.; Hussain, T.; Yang, H.; Zhu, M. Effects of dietary energy levels on rumen fermentation, microbiota, and gastrointestinal morphology in growing ewes. Food Sci. Nutr. 2020, 8(12), 6621–6632. [Google Scholar] [CrossRef]
- Li, M.; Hassan, F.; Peng, L.; Xie, H.; Liang, X.; Huang, J.; Huang, F.; Guo, Y.; Yang, C. Mulberry flavonoids modulate rumen bacteria to alter fermentation kinetics in water buffalo. PeerJ 2022, 10, e14309. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: metabolism of nutrients and other food components. Eur. J. Nutr. 2017, 57(1), 1–24. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Method for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Olsen, S.R.; Sommer, L.E. Phosphorus. Chem. Microbiol. Prop. 1982, 9, 403p. Am. Soc. Agron., Soil Sci. Soc. Am., Madison.
- Tajini, F.; Boualy, Y.; Ouerghui, A. Study of the Nutritional Quality of Phoenix dactylifera L. fruit: Measurement of Biochemical Parameters. Rev. Nat. Technol. 2020, 12(2), 39-49.
- Sarr, S.O.; Fall, A.D.; Gueye, R.; Diop, A.; Sene, B.; Diatta, K.; Ndiaye, B.; Diop, Y.M. Evaluation de l’activité antioxydante des extraits des feuilles de Aphania senegalensis (Sapindaceae) et de Saba senegalensis (Apocynaceae). Int. J. Biol. Chem. Sci. 2015, 9, 2676–2684. [Google Scholar] [CrossRef]
- Falleh, H.; Ksouri, R.; Chaieb, K.; Karray-Bouraoui, N.; Trabelsi, N.; Boulaaba, M.; Abdelly, C. Phenolic composition of Cynara cardunculus L. organs, and their biological activities. C. R. Biol. 2008, 331, 372–379. [CrossRef]
- Jedidi, S.; Selmi, H.; Aloui, F.; Rtibi, K.; Jridi, M.; Chaâbane, A.; Sebai, H. Comparative studies of phytochemical screening, HPLC-PDA-ESI-MS/MS-LC-HRESIMS analysis, antioxidant capacity, and in vitro fermentation of officinal sage (Salvia officinalis L.) cultivated in different biotopes of Northwestern Tunisia. Chem. Biodivers. 2019, 10, 1002. [Google Scholar] [CrossRef]
- Menke, K.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
Figure 1.
HPLC chromatogram of extracts from P. vulgare (A), C. nobile (B), O.forsskaolii (C) and L. stoechas (D). Peaks numbers corresponding as compounds in Table 3.
Figure 1.
HPLC chromatogram of extracts from P. vulgare (A), C. nobile (B), O.forsskaolii (C) and L. stoechas (D). Peaks numbers corresponding as compounds in Table 3.
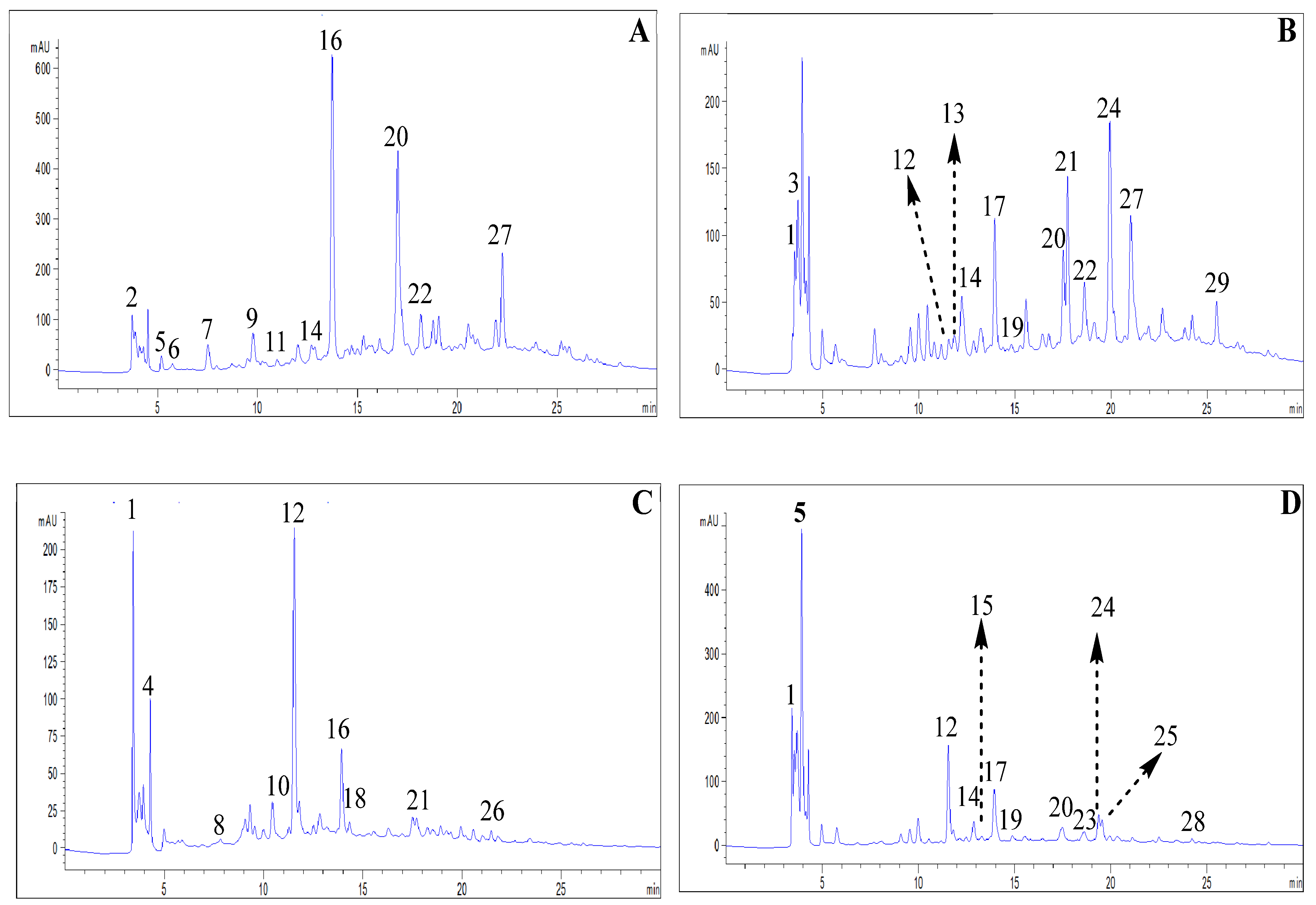
Figure 2.
Kinetic curves for gas production by P.vulgare (I), C. nobile (II), O. forsskaolii (III) and L. stoechas (IV), in sheep (A) and goat (B).
Figure 2.
Kinetic curves for gas production by P.vulgare (I), C. nobile (II), O. forsskaolii (III) and L. stoechas (IV), in sheep (A) and goat (B).
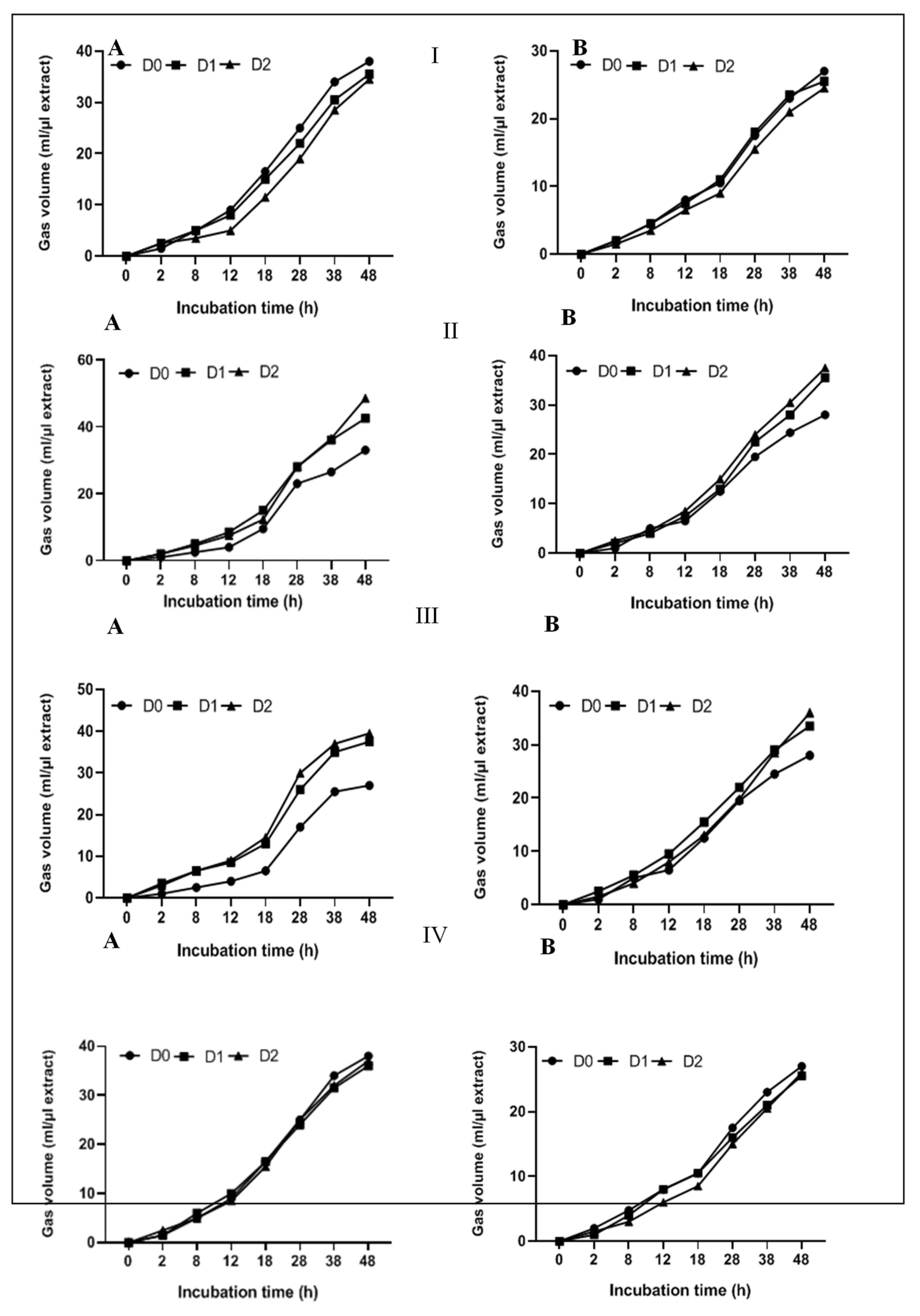
Table 3.
HPLC identification of phenolic compounds (%) in aqueous extracts from P. vulgare, C. nobile, O. forsskaolii, and L. stoechas.
Table 3.
HPLC identification of phenolic compounds (%) in aqueous extracts from P. vulgare, C. nobile, O. forsskaolii, and L. stoechas.
| Peaks numbers | Compoundsa | RTb | P. vulgare | C. nobile | O. forsskaolii | L. stoechas |
|---|---|---|---|---|---|---|
| 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 |
Ascorbic acid Shikimic acid Apigenine-7-O-glucoside Apigenin Gallic acid 5-O-caffeoylquinic acid 3-O-caffeoylquinic acid Rosmarinic acid Hyperoside Kaempferol 3,5-Dicaffeoylquinic acid Chlorogenic acid Résorcinol Catechin Vanillic acid Catechol Epicatechin Syringic acid Cafeic acid Ellagic acid Luteolin Sinapic acid p-Coumaric acid Ferulic acid m-Coumaric acid Quercetin Myricetin Cinnamic acid Ferulic-1-O-glucoside acid |
3.69 3.71 3.93 4.29 5.18 5.74 7.51 7.83 9.77 10.44 10.98 11.50 11.84 12.70 13.29 13.73 13.94 14.32 14.88 17.02 17.74 18.17 18.61 19.36 19.55 21.80 22.26 24.22 25.49 |
- 2.52 - - 0.88 0.50 2.29 - 2.75 - 0.41 - - 1.37 - 22.12 - - - 20.59 - 3.68 - - - - 6.89 - - |
5.81 - 9.39 - - - - - - - - 0.99 1.31 3.92 - - 5.80 - 0.34 3.92 6.83 3.13 - 10.42 - - 7.56 - 1.79 |
14.22 - - 6.89 - - - 1.06 - 3.76 - 23.53 - - - 7.63 - 1.17 - - 1.97 - - - - 0.51 - - - |
9.44 - - 21.40 - - - - - - - 10.30 - 2.56 0.66 - 7.63 - 0.73 2.65 - - 1.94 3.02 2.45 - - 0.41 - |
aIdentification was confirmed using authentic commercial standards, bretention time.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



