
Submitted:
25 April 2025
Posted:
25 April 2025
You are already at the latest version
Abstract
Background/Objectives: Acute kidney injury (AKI) remains an unsolved medical problem due to the lack of effective treatments, high mortality and increased susceptibility to progression to chronic kidney disease (CKD), especially in the elderly. Cellular senescence has been described in AKI, CKD and ageing, and has been proposed as a promising therapeutic target. The senolytic drugs combination dasatinib plus quercetin (D&Q) is beneficial in some pathological conditions, including experimental CKD, but there is no data in AKI. Methods: The effect of D&Q treatment was tested in folic acid-induced nephrotoxicity (AKI-FAN), a murine AKI model. Results: D&Q pretreatment did not prevent renal dysfunction in the acute phase of AKI-FAN, as determined by serum creatinine and BUN levels at 48 hours. Moreover, gene expression of the kidney damage biomarkers Lcn2 and Havcr1, the Cdkn1a gene, which encodes p21, and some genes encoding components of the senescent cell secretome were significantly increased in response to D&Q treatment. The number of senescent p21-positive cells in injured kidneys was similar in untreated or D&Q-treated FAN mice. In addition, D&Q did not prevent the downregulation of the anti-aging factor Klotho in damaged kidneys. Conclusions: D&Q treatment was not protective in AKI-FAN, exacerbating some deleterious responses. These results suggest caution when exploring the clinical translation of D&Q senolytic activity.
Keywords:
acute kidney injury
; senolytics
; senescence
; dasatinib
; quercetin
; Klotho
; p21
1. Introduction
Acute kidney injury (AKI) is the most common cause of death in hospitalized and clinically ill patients [1,2,3]. It is characterized by a sudden but often reversible reduction in renal function, resulting in changes in serum creatinine and urine levels, that may progress to chronic kidney disease (CKD) [4,5]. Although in 2013 the International Society of Nephrology launched an initiative to eradicate or, at least, reduce preventable AKI-related deaths worldwide by 2025 [6], this goal is far from being achieved. To date, no pharmacological strategies have been implemented to prevent nor to cure AKI episodes and renal replacement therapies remain the standard of care [7]. Population ageing is contributing to a higher burden of both AKI and CKD. CKD is estimated to become the 5th global leading cause of death by 2040 [8]. Both CKD and older age increase the risk of AKI and of the AKI-to-CKD progression [9]. This highlights the importance of further research to find interventions that prevent or alleviate AKI.
Activation of cellular senescence has been described both in AKI and CKD [10]. Cellular senescence occurs in both physiological (e.g., embryonic development, tissue regeneration and repair) and pathological (e.g., metabolic disorders, aging and age-related diseases) processes [11,12] in response to multiple stressors (e.g., DNA damage, epigenetic changes, metabolic shocks, oxidative stress). It is characterized by increased expression of cell-cycle-related molecules, such as the cyclin kinase inhibitors (CDK) p16ink4a (p16; encoded by CDKN2A) and p21WAF-1/CAP1 (p21, encoded by CDKN1A) that initiate and maintain a stable cell cycle arrest in the G1 or possibly G2 phase, promoting resistance to apoptosis [13]. Phenotype changes in senescent cells include an aberrant secretome, the “senescence-associated secretory phenotype” (SASP) containing proinflammatory cytokines, growth factors and chemokines [14]. Renal accumulation of senescent cells has been described in CKD patients and can contribute to kidney damage progression through SASP proinflammatory and profibrotic factors [15].
However, the role of cellular senescence in the early phase of AKI is not fully understood [16,17]. The acute phase of injury is followed by regeneration consisting of tubular cells proliferation, and restoration of epithelial phenotype and function [18]. Some cells do not recover and are thought to acquire a senescent phenotype, leading to maladaptive repair promoting the AKI-to-CKD transition [19,20]. In this sense, targeting senescence in maladaptive repair, renal fibrosis, and transplant failure was beneficial in preclinical models [21,22].
Senotherapeutic strategies to target senescence are divided into senolytics that eliminate senescent cells, and senomorphics that suppress the pathological SASP [23]. Senolytics may target prosurvival pathways, including the BCL-2/BCL-xL, p53/p21, and PI3K/AKT pathways, and antiapoptotic pathways, including serpins [24]. The combination of Dasatinib (D) and Quercetin (Q) was discovered by a hypothesis-driven bioinformatics approach and demonstrated senolysis in vivo [25]. Dasatinib is a tyrosine inhibitor of BCR-ABL, SRC family, c-KIT, and ephrin receptors, used to treat leukemias, that enhances apoptosis and inhibits proliferation and migration [26]. Quercetin is a flavonoid inhibitor of PI3Ks and serpins, with anti-inflammatory, antioxidant and anti-proliferative properties [27]. Compared to each drug alone, the combination magnifies senescent cell clearance through apoptosis in vitro [26]. In experimental diabetic nephropathy, D&Q administration during 10 weeks improved renal function [28], as described in other models of CKD by the same authors [22]. However, preclinical data on the safety and efficacy of D&Q are still insufficient and its effect in the acute phase of AKI is unknown. Therefore, our aim was to investigate the effect of the senolytic drug cocktail D&Q in experimental AKI caused by folic acid nephrotoxicity (AKI-FAN).
2. Results
2.1. Treatment with Dasatinib Plus Quercetin Did Not Prevent Renal Dysfunction in AKI-FAN
To investigate the effect of D&Q in AKI, nephrotoxicity was induced in C57BL/6 mice by an overdose of folic acid and mice were studied at 48 hours. FAN is an established model to investigate the molecular mechanism involved in the acute phase of AKI [29]. Serum BUN and creatinine were increased in AKI-FAN at 48 hours compared to controls (Figure 1A,B), indicating decreased kidney function in the acute phase of AKI. However, D&Q treatment did not improve serum renal functionality markers compared to untreated AKI-FAN mice (Figure 1A,B), suggesting that this senolytic treatment does not protect from the acute phase of AKI.
2.2. Dasatinib Plus Quercetin Increased the Gene Expression of Kidney Damage Biomarkers in AKI-FAN
To further evaluate the effect of D&Q on AKI-FAN, the gene expression of well-known kidney damage biomarkers Havcr1 and Lipocalin-2 (Lcn2), encoding the proteins KIM-1 and NGAL respectively [30] was studied. In AKI-FAN, qPCR disclosed increased kidney Lcn2 and Havcr1 gene expression (Figure 2A,B) compared to healthy controls, as previously described in the acute phase of AKI [31]. In response to D&Q treatment, the gene expression of both biomarkers significantly increased in injured AKI-FAN kidneys compared to untreated AKI-FAN (Figure 2A,B), suggesting a deleterious effect and an increased severity of kidney injury resulting from senolytic treatment.
2.3. Dasatinib and Quercetin Did Not Modify the Tubular Damage Marker KIM-1 in AKI-FAN
2.4. Dasatinib Plus Quercetin Did Not Modify the Number of Senescent Cells in AKI-FAN
To test the potential senolytic effect of D&Q in AKI-FAN, we evaluated its impact on the number of tubular senescent cells. To this aim, cells stained for p21, a universal marker of cell senescence, were evaluated by immunohistochemistry. In AKI-FAN, p21 positive cells were observed mainly among tubular cells, whereas no staining was found in healthy controls (Figure 4A). D&Q did not modify the number of p21 positive cells compared to untreated AKI-FAN mice (Figure 4B). These results suggest the lack of senolytic effect of D&Q in the acute phase of AKI-FAN.
2.5. Dasatinib Plus Quercetin and Senescence-Associated Biomarkers
To test whether D&Q modulated other senescence-associated processes in AKI-FAN, we analyzed the gene expression of well-established markers of cell senescence in mouse kidneys. First, the expression of the cell cycle arrest genes Cdkn1a and Cdkn2a, that encode p21 and p16 respectively, was evaluated. In AKI-FAN, Cdkn1a and Cdkn2a expression increased compared with healthy control (Figure 5A,B), as previously described [32]. However, in D&Q-treated mice renal Cdkn1a gene expression was significantly increased compared to untreated AKI-FAN mice (Figure 5A,B).
Senescent cells are characterized by gene reprograming, leading to increased SAPS gene expression [33]. The gene expression for several SASP components, such as the proinflammatory cytokine IL1β and the chemokines Ccl2 and Ccl5 was significantly higher in AKI-FAN compared to controls, as described [32,34]. However, D&Q did not diminish SASP proinflammatory gene expression, whereas significantly increasing Ccl2 expression (Figure 5D,E). These data indicate an absence of anti-inflammatory effect of D&Q in AKI-FAN.
2.6. Dasatinib Plus Quercetin Did Not Increase Apoptosis during AKI-FAN
D&Q targets prosurvival and anti-apoptotic pathways [35]. To assesses the proapoptotic activity of D&Q in AKI-FAN, the expression of apoptotic and anti-apoptotic genes (Bax and Bcl2l1 respectively) was evaluated. In AKI-FAN, Bax expression increased compared to untreated kidneys, and was not influenced by D&Q (Figure 6A). Moreover, the anti-apoptotic factor Bcl2l1 (which encodes BCLxL) was evaluated at both gene and protein levels. No significant changes were observed for gene expression (Figure 6B) whereas at BclxL protein increased in AKI-FAN when compared to control, but this was not modified by D&Q.
2.7. Dasatinib Plus Quercetin Did Not Modify Necroptosis Pathway Activation in AKI-FAN
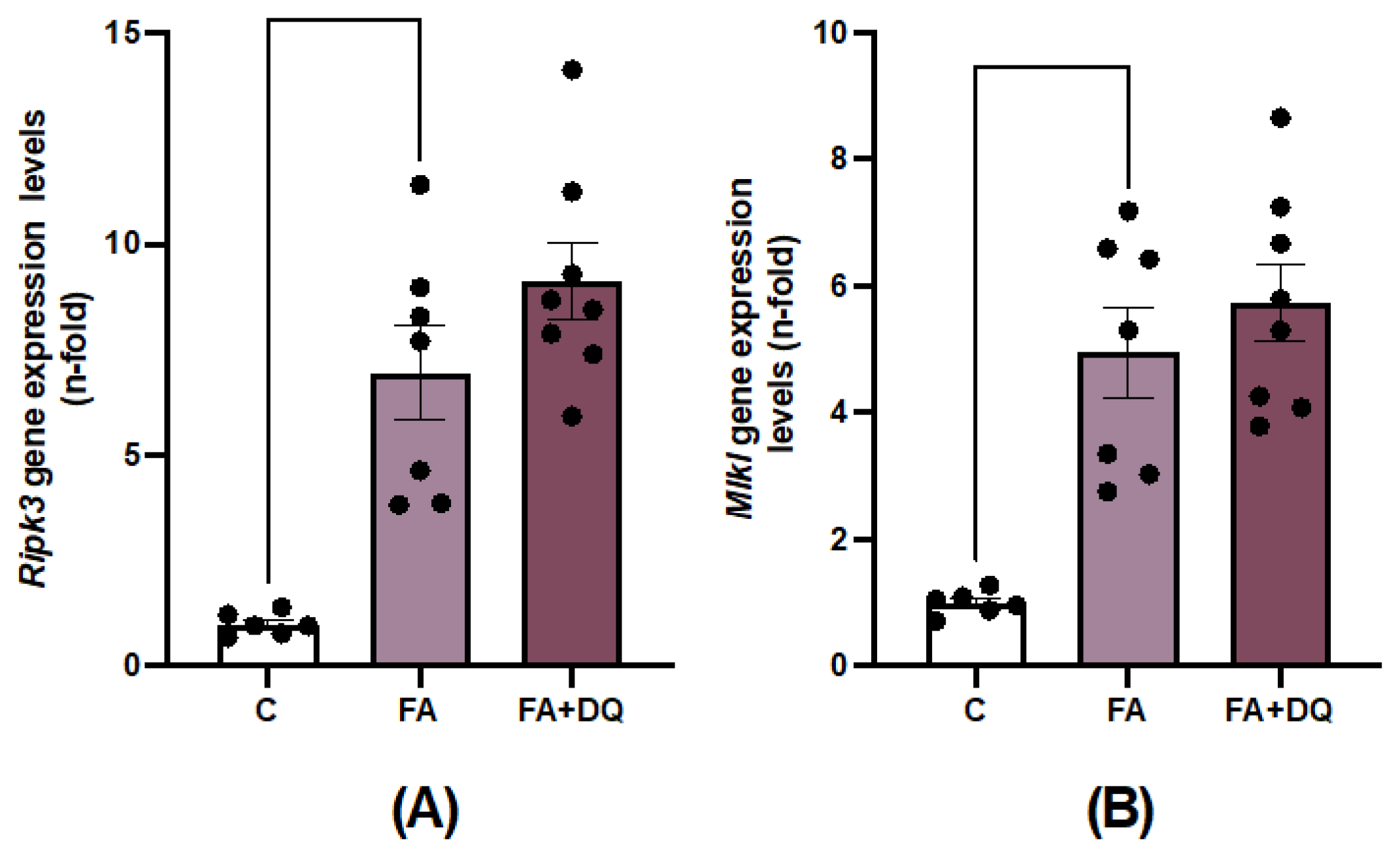
Previous studies have described the cell death mechanisms involved in experimental FAN-AKI [29,34], emphasizing the role of necroptosis, including the upregulation of key components in this cell death pathway, such as receptor-interacting protein kinase-3 (RIPK3) and mixed-lineage kinase domain-like protein (MLKL) [34]. Treatment with D&Q had no effect on the Ripk3 and Mlkl gene overexpression observed in AKI-FAN (Figure 7A,B).
3. Discussion
The main finding is that a senolytic drug combination, D&Q, can increase renal damage in the acute phase of an experimental nephrotoxic nephropathy. Clinical trials are evaluating the senolytic potential of D&Q in multiple conditions, including Alzheimer (NCT05422885), skeleton health (NCT04313634), pulmonary fibrosis (NCT02874989) and diabetic CKD (NCT02848131), but none has reported on kidney function. To our knowledge there are no reports evaluating the effect of D&Q in the acute phase of experimental models of AKI, neither evaluating their long-term kidney safety, emphasizing the need to report negative results of drugs already in clinical development in humans, especially in drug repurposing.
Genetic or pharmacological elimination of senescent cells was beneficial in many murine preclinical models of disease [24]. Therefore, targeting senescent cells in aging and age-related diseases has been proposed as a potential therapeutic option for humans. The beneficial effects of D&Q were initially described in old mice [36,37,38] observing a longer lifespan [39]. In an open label phase 1 clinical trial (NCT02848131), treatment with D&Q for 3 days decreased adipose tissue senescent cell burden in diabetic patients [40]. In diabetic db/db mice treatment with D&Q for 20 weeks reduced glycemia and improved functional and histopathological changes [41]. D&Q also attenuated adipose tissue inflammation and improved systemic metabolic function in old mice, without changing senescence and inflammatory SASP markers in the liver and skeletal muscle [36]. In line with these data, D&Q treatment did not diminish senescence and inflammatory SASP markers in injured AKI-FAN kidneys. These data suggest that the beneficial D&Q effects described in diabetes can be restricted to senolysis on adipose tissue or other specific tissues. In this sense, in a murine model of 5/6 nephrectomy that resembles human nephron reduction in CKD, D&Q treatment reduced muscle wasting, suggesting that skeletal muscle can also be a target tissue [42]. However, they did not report kidney function, damage or senescence parameters. In muscle wasting, idiopathic pulmonary fibrosis and intestinal senescence, D&Q decreased senescence markers (p21, p16) and SASP components [37,42,43]. By contrast, D&Q reduced the senescent-like myeloid cells in experimental autoimmune encephalomyelitis, but did not influence inflammation or mice recovery [44]. Other senolytics, such as Navitoclax (ABT-263) and Fisetin were beneficial in cardiovascular disease [45]. However, the role of senolytics in renal regeneration has not been fully elucidated.
Proximal tubule cells are the kidney cells most sensitive to nephrotoxins [46]. Nephrotoxic insults elicit different stress responses in injured tubular cells [47], including activation of CDK inhibitors, mainly p21 [48]. In different experimental AKI models, including ischemia-reperfusion and cisplatin nephrotoxicity, rapid activation of cell senescence was observed, mainly induction of p21 gene and protein expression in tubular cells [49,50,51,52,53], as observed for AKI-FAN. Early p21 activation has been interpreted as a protective mechanism, preventing uncontrolled progression to cell death, allowing damaged cells to remain in cell cycle arrest, and thus providing more time for DNA damage repair [51]. In addition, p21 activation modulates apoptosis and necrosis [54]. Indeed, AKI was more severe in p21 knockout mice [54]. D&Q significantly increased p21 gene expression in AKI-FAN and it did not diminish the number of p21 positive tubular senescent cells. A possible explanation for the loss of the senolytic effect could be the overall net balance between apoptosis and senescence in this acute phase of AKI characterized by cell death (necroptosis). Accordingly, D&Q did not modify AKI-induced changes on the anti-apoptotic protein BCLxL or on necroptosis-related components. Some data suggest that activation of cellular senescence can be protective in the early stages of AKI [55], indicating that D&Q could be deleterious in the acute phase of AKI, characterized by ongoing cell death. Supporting this finding, biomarkers of renal damage Havcr1 and Lcn2, and the proinflammatory mediator ccl2 genes were significantly overexpressed in response to D&Q treatment in mice with AKI-FAN.
Contrary to our findings, in preclinical septic AKI induced by LPS administration, the senolytic compound Fisetin [56] inhibited kidney dysfunction, inflammation and apoptosis [57]. Fisetin is a flavonoid that diminished renal inflammation in many preclinical models [58,59,60,61,62], therefore exerting senomorphic activities, by inhibiting SAPS overexpression. Moreover, anti-inflammatory treatments ameliorate LPS-induced AKI [63]. In this sense, fisetin inhibition of the TLR4-NF-κB p65 and MAPKs pathways in LPS-AKI [64] may explain or contribute to its beneficial effects, independent of potential senolytic actions. There is ample evidence of beneficial effects of SASP inhibitors in preclinical studies, such as observed for the IL-1β receptor inhibitor Anakinra used in rheumatoid arthritis, or Metformin, a treatment for type 2 diabetes which inhibits the transcription factor NF- κB [33]. Reports also support beneficial effect of SASP-targeting senomorphic therapies in the acute phase of AKI. Blockade of the SASP component CCN2 improved the acute phase of experimental IRI-AKI and AKI-FAN, by targeting oxidative stress [65] and the NLRP3/RIPK3/NRF2 pathway [66]. IKK/NF-κB and JAK inhibitors also suppress the SASP [67], however no protective effect has been described in acute AKI.
In contrast to AKI findings, there are reports of protective effects of senolytics in experimental CKD. Suppression of tubular senescence by Navitoclax and Fisetin attenuated renal fibrosis and improved tubular repair, as indicated by restoration of tubular regeneration and renal function [19,55]. Navitoclax inhibits Bcl-2 family members thereby activating apoptosis mechanisms in senescent cells. In murine cisplatin AKI, Navitoclax inhibited tubular senescence, improving fibrosis and renal function [19]. However, navitoclax did not exhibit anti-senescence properties in premature ageing in short telomere zebrafish [68]. Navitoclax may produce thrombocytopenia as a dose-limiting adverse effect [69].
Klotho is a protein mainly synthetized by the kidneys [70], that is downregulated in AKI and CKD [71]. The administration of recombinant Klotho protein in experimental CKD delayed fibrosis [72]. Therefore therapies restoring Klotho were proposed as nephroprotective and anti-aging agents [73]. Exposure to the senescent cell secretome reduces α-Klotho in different cultured human cells, and this was partially prevented by blocking some SASP factors, as IL1α [73]. In an idiopathic pulmonary fibrosis clinical trial (NCT02874989), D&Q improved physical function and increased urinary α-Klotho [73], suggesting that D&Q could modulate Klotho levels. However, the mechanisms involved are poorly understood. By contrast, in AKI-FAN, renal Klotho downregulation was not restored by D&Q treatment [74].
Aging is a complex process [12]. Senescent cell burden increases in multiple tissues with aging [75]. Cellular senescence may accelerate aging as the transplantation of a small numbers of senescent cells in young mice led to physical dysfunction [38]. Moreover, genetic or pharmacological elimination of senescent cells extended the health span and longevity of naturally aged mice [24,38], as well as in mice with accelerated aging [76]. In humans and in preclinical AKI, aging increased the severity of renal injury [9]. The acute phase of FAN-AKI was more severe in aged mice, as evidenced by higher tubular cell death, mainly regulated-necrosis, suppressed apoptosis and higher number of senescent cells, and increased levels of senescent markers (p16, p21, γH2AX, SASP components) [77]. These data suggest that overactivation of senescence mechanisms, including suppression of apoptosis, could contribute to increase the severity of AKI in the elderly. In this regard, FOXO4-DRI, a senolytic drug targeting the p53-FOX4 interaction that promotes apoptosis of senescent cells, improved kidney function in mice with spontaneous or accelerated aging [78]. However, strategies that eliminate senescent cells could be detrimental in certain disease conditions, as described here for AKI-FAN in the first study to evaluate D&Q in an AKI model. Our results indicate that targeting senescence in AKI may require a more tailored approach, as the benefits observed in other contexts, such as aging or chronic diseases, do not necessarily translate to AKI. Although this study does not support the use of D&Q in AKI, further research is essential to better understand the molecular mechanisms triggered by senolytic drugs in different contexts.
Certain limitations should be acknowledged. Although AKI-FAN shares some cellular and molecular mechanisms with other causes of AKI, future studies should address other causes of AKI, including IRI and sepsis-induced AKI. In addition, D&Q was tested at a single dose and timepoint, in a preventive manner. Whether this is the optimal therapeutic time window, and dose remains to be determined. Moreover, this study tested young male mice, so additional experiments should explore female and older mice. Finally, this preclinical study requires clinical confirmation that may be derived from the kidney safety evaluation of ongoing clinical trials.
In conclusion, in a preclinical nephrotoxic AKI, preventive D&Q treatment was not protective and even increased the severity of some features of kidney damage in the acute phase. This is a cautionary tale for the clinical translation of senolytic therapies.
4. Materials and Methods
4.1. Animals
Experiments were performed according to the European Community guidelines for animal experiments and the ARRIVE guidelines and with consent of the Experimental Animal Ethics Committee of the Health Research of the IIS-Fundación Jiménez Díaz and PROEX 242.2/21 and PROEX 065/18 of the Community of Madrid. Animals were sacrificed with an overdose of CO2 in a special chamber. Blood and urine were collected, and kidneys were perfused in situ with saline before removal. Half of each kidney (2/4) was fixed, embedded in paraffin, and used for immunohistochemistry, and the rest was snap-frozen in liquid nitrogen for renal cortex RNA studies.
4.2. Folic acid Model (AKI-FAN)
AKI was induced by a single intraperitoneal (i.p.) injection of 125 mg/kg body weight folic acid (Sigma-Aldrich) in 0.3 mol/L sodium bicarbonate. Animals were sacrificed 48 hours later. Some animals received 24 hours before folic acid, a single oral dose of Dasatinib (5 mg/kg body weight) and Quercetin (50 mg/kg body weight) (n=6 to 9 mice per group).
4.3. Gene Expression Studies
RNA from renal cortex was isolated with TRItidy GTM (PanReac; Barcelona, Spain). cDNA was synthesized by a High Capacity cDNA Archive kit (Applied Biosystems) using 2μg of total RNA and following the manufacturer’s instructions. Quantitative gene expression analysis was performed on a QuantStudio™ 3 fast real-time PCR system (Applied Biosystems) using fluorogenic TaqMan MGB probes and primers designed by Assay-on-DemandTM gene expression products or Predesigned qPCR. Mouse assays IDs were: Ccl2: Mm00441242_m1, Ccl5: Mm01302428_m1, Cdkn1a: Mm00432448_m, Cdkn2a: Mm00494449_m1, Ripk3: Mm00444947_m1 Havcr1: Mm00506686_m1, Il1β: Mm00434228_m1, Mlkl: Mm01244219_m1, Klotho: Mm00502002_m1 and Lcn2: Mm01324470_m1. Data were normalized to Gapdh: Mm99999915_g1. The mRNA copy numbers were calculated for each sample by the instrument software using Ct value (“arithmetic fit point analysis for the lightcycler”). Results were expressed in n-fold, calculated relative to control group after normalization against Gapdh.
4.4. Immunohistochemistry
Paraffin-embedded kidney sections were stained using standard histology procedures, as described elsewhere [79]. Immunohistochemistry (IH) were performed in 3 μm thick tissue sections. Antigens were retrieved using the PTlink system (DAKO) with sodium citrate buffer (10 mM) adjusted to pH 6–9, depending on the immunohistochemical marker. Endogenous peroxidase was blocked. Sections were incubated for 1 h at room temperature with Casein Solution (Vector Laboratories) to remove non-specific protein binding sites. Then, primary antibodies were incubated overnight at 4°C. Specific biotinylated secondary antibodies (Amersham Biosciences) were used. The latter was followed by Avidin-Biotin Complex incubation (Vector Laboratories). Signal was developed with Substrate solution and 3,3-diaminobenzidine as a chromogen (Abcam). Finally, slides were counterstained with Carazzi’s haematoxylin (Richard Allan Scientific).
The primary antibodies used were [dilution]: P21cip1 ( [1:200, Ab188224, Abcam), BCL-xL ( [1:2000], ab178844, Abcam) and KIM-1 ( [1:200]; AF 1817, R&D).
Specificity was checked by omission of primary antibodies (not shown). Quantification was made by using the Image-Pro Plus software (Maryland, USA) to determine the positive staining area relative to the total area or counting positive staining manually (in the case of P21cip1 immunohistochemistry), in 5-10 randomly chosen fields (×200 magnification).
4.5. Statistical Analysis
Kidneys from all groups were compared to control and to AKI-FAN, expressing results as fold-change over control values of 1. Results are expressed as fold increase with respect to the control average as mean ± standard error of the mean (SEM) of 4 to 9 animals per group. The Shapiro-Wilk test was used to evaluate sample normality distribution. When samples followed the Gaussian distribution, a one-way ANOVA followed by the corresponding post-hoc analyses was used. To compare non-parametric samples, a Kruskal-Wallis and a subsequent post-hoc analysis was performed. Graphics and statistical analysis were conducted using GraphPad Prism 9.5.1 (GrahPad Software, San Diego California USA). Values of p < 0.05 were considered statistically significant.
Author Contributions
Conceptualization A.B-V, V.M., L.T-S, S.R-M, A.O and M.R-O. Funding acquisition M.P.R-T, S.R-M, A.O and M.R-O. Methodology A.B-V, L.T-S, C.G-C and E.F-S. Validation M.P-R-T, S.R-M, A.O and M.R-O. Writing—original draft A.B-V and M.R-O. Review & editing, V.M., L T-S, C G-C, E.F-S, M.P-R-T, S-R-M and A.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from the Instituto de Salud Carlos III (ISCIII) and Fondos FEDER European Union (PI23/00394, PI24/00007, RD21/0005/0002, RD21/0005/0005 and RD24/0004/0021 and RD24/0004/0028 RICORS2040-RENAL funded by European Union—NextGenerationEU). INNOREN “P2022/BMD-7221: Nuevas estrategias diagnósticas y terapéuticas en enfermedad renal crónica. Miguel Servet CP23/00025 to S. R-M. PIPF-2023/SAL-GL-29648 to A.B-V.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the IIS-FJD Animal Research Ethical Committee guidelines (PROEX 065/18; PROEX 242.2/21).
Acknowledgments
We thank Laura Almenara for secretarial support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| AKI | Acute Kidney Injury |
| CKD | Chronic Kidney disease |
| D&Q | Dasatinib (D) and Quercetin (Q) |
| SASP | Senescence-associate secretory phenotype |
References
- Rodrigues FB, Bruetto RG, Torres US, Otaviano AP, Zanetta DMT, Burdmann EA. Incidence and Mortality of Acute Kidney Injury after Myocardial Infarction: A Comparison between KDIGO and RIFLE Criteria. PLoS One 2013;8. [CrossRef]
- Gameiro J, Fonseca JA, Outerelo C, Lopes JA. Acute kidney injury: From diagnosis to prevention and treatment strategies. J Clin Med 2020;9. [CrossRef]
- Havaldar AA, Sushmitha EAC, Shrouf S Bin, H. S M, N M, Selvam S. Epidemiological study of hospital acquired acute kidney injury in critically ill and its effect on the survival. Sci Rep 2024;14. [CrossRef]
- Tamargo C, Hanouneh M, Cervantes CE. Treatment of Acute Kidney Injury: A Review of Current Approaches and Emerging Innovations. J Clin Med 2024;13. [CrossRef]
- Gameiro J, Fonseca JA, Outerelo C, Lopes JA. Acute kidney injury: From diagnosis to prevention and treatment strategies. J Clin Med 2020;9. [CrossRef]
- Mehta RL, Cerdá J, Burdmann EA, Tonelli M, García-García G, Jha V, Susantitaphong P, Rocco M, Vanholder R, Sever MS, Cruz D, Jaber B, Lameire NH, Lombardi R, Lewington A, Feehally J, Finkelstein F, Levin N, Pannu N, Thomas B, Aronoff-Spencer E, Remuzzi G. International Society of Nephrology’s 0by25 initiative for acute kidney injury (zero preventable deaths by 2025): A human rights case for nephrology. The Lancet 2015;385:2616–43. [CrossRef]
- Negi S, Wada T, Matsumoto N, Muratsu J, Shigematsu T. Current therapeutic strategies for acute kidney injury. Ren Replace Ther 2023;9. [CrossRef]
- Ortiz A, Roger M, Jiménez VM, Perez JCR, Furlano M, Montero AM, Pérez NM, et al. RICORS2040: The need for collaborative research in chronic kidney disease. Clin Kidney J 2022;15:372–87. [CrossRef]
- Chang-Panesso M. Acute kidney injury and aging. Pediatric Nephrology 2021;36:2997–3006. [CrossRef]
- Huang W, Hickson LTJ, Eirin A, Kirkland JL, Lerman LO. Cellular senescence: the good, the bad and the unknown. Nat Rev Nephrol 2022;18:611–27. [CrossRef]
- Sturmlechner I, Durik M, Sieben CJ, Baker DJ, Van Deursen JM. Cellular senescence in renal ageing and disease. Nat Rev Nephrol 2017;13:77–89. [CrossRef]
- López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. Hallmarks of aging: An expanding universe. Cell 2023;186:243–78. [CrossRef]
- O’Sullivan EA, Wallis R, Mossa F, Bishop CL. The paradox of senescent-marker positive cancer cells: challenges and opportunities. Npj Aging 2024;10:41. [CrossRef]
- Rayego-Mateos S, Marquez-Expósito L, Rodrigues-Diez R, Sanz AB, Guiteras R, Doladé N, Rubio-Soto I, Manonelles A, Codina S, Ortiz A, Cruzado JM, Ruiz-Ortega M, Sola A. Molecular Mechanisms of Kidney Injury and Repair. Int J Mol Sci 2022;23. [CrossRef]
- Li Y, Lerman LO. Cellular Senescence: A New Player in Kidney Injury. Hypertension 2020;76:1069–75. [CrossRef]
- Chen J, Zhang H, Yi X, Dou Q, Yang X, He Y, Chen J, Chen K. Cellular senescence of renal tubular epithelial cells in acute kidney injury. Cell Death Discov 2024;10. [CrossRef]
- Sanz AB, Sanchez-Niño MD, Ramos AM, Ortiz A. Regulated cell death pathways in kidney disease. Nat Rev Nephrol 2023;19:281–99. [CrossRef]
- Ruiz-Ortega M, Rayego-Mateos S, Lamas S, Ortiz A, Rodrigues-Diez RR. Targeting the progression of chronic kidney disease. Nat Rev Nephrol 2020;16:269–88. [CrossRef]
- Li S, Livingston MJ, Ma Z, Hu X, Wen L, Ding HF, Zhou D, Dong Z. Tubular cell senescence promotes maladaptive kidney repair and chronic kidney disease after cisplatin nephrotoxicity. JCI Insight 2023;8. [CrossRef]
- Chen J, Zhang H, Yi X, Dou Q, Yang X, He Y, Chen J, Chen K. Cellular senescence of renal tubular epithelial cells in acute kidney injury. Cell Death Discov 2024;10. [CrossRef]
- Kurella M, Chertow GM, Fried LF, Cummings SR, Harris T, Simonsick E, Satterfield S, Ayonayon H, Yaffe K. Chronic kidney disease and cognitive impairment in the elderly: The Health, Aging, and Body Composition study. Journal of the American Society of Nephrology 2005;16:2127–33. [CrossRef]
- Li C, Shen Y, Huang L, Liu C, Wang J. Senolytic therapy ameliorates renal fibrosis postacute kidney injury by alleviating renal senescence. FASEB Journal 2021;35. [CrossRef]
- Zhang L, Pitcher LE, Prahalad V, Niedernhofer LJ, Robbins PD. Targeting cellular senescence with senotherapeutics: senolytics and senomorphics. FEBS Journal 2023;290:1362–83. [CrossRef]
- Robbins PD, Jurk D, Khosla S, Kirkland JL, Lebrasseur NK, Miller JD, Passos JF, Pignolo RJ, Tchkonia T, Niedernhofer LJ. Senolytic Drugs: Reducing Senescent Cell Viability to Extend Health Span n.d.
- Zhu Y, Tchkonia T, Pirtskhalava T, Gower AC, Ding H, Giorgadze N, Palmer AK, Ikeno Y, Hubbard GB, Lenburg M, O’hara SP, Larusso NF, Miller JD, Roos CM, Verzosa GC, Lebrasseur NK, Wren JD, Farr JN, Khosla S, Stout MB, McGowan SJ, Fuhrmann-Stroissnigg H, Gurkar AU, Zhao J, Colangelo D, Dorronsoro A, Ling YY, Barghouthy AS, Navarro DC, Sano T, Robbins PD, Niedernhofer LJ, Kirkland JL. The achilles’ heel of senescent cells: From transcriptome to senolytic drugs. Aging Cell 2015;14:644–58. [CrossRef]
- Zhang L, Pitcher LE, Prahalad V, Niedernhofer LJ, Robbins PD. Targeting cellular senescence with senotherapeutics: senolytics and senomorphics. FEBS Journal 2023;290:1362–83. [CrossRef]
- Deepika, Maurya PK. Health Benefits of Quercetin in Age-Related Diseases. Molecules 2022;27. [CrossRef]
- Guo X, Wen S, Wang J, Zeng X, Yu H, Chen Y, Zhu X, Xu L. Senolytic combination of dasatinib and quercetin attenuates renal damage in diabetic kidney disease. Phytomedicine 2024;130. [CrossRef]
- Guerrero-Mauvecin J, Villar-Gómez N, Rayego-Mateos S, Ramos AM, Ruiz-Ortega M, Ortiz A, Sanz AB. Regulated necrosis role in inflammation and repair in acute kidney injury. Front Immunol 2023;14. [CrossRef]
- Lane BR. Molecular markers of kidney injury. Urologic Oncology: Seminars and Original Investigations 2013;31:682–5. [CrossRef]
- Ichimura T, Hung CC, Yang SA, Stevens JL, Bonventre J V. Kidney injury molecule-1: a tissue and urinary biomarker for nephrotoxicant-induced renal injury 2004.
- Marquez-Exposito L, Tejedor-Santamaria L, Santos-Sanchez L, Valentijn FA, Cantero-Navarro E, Rayego-Mateos S, Rodrigues-Diez RR, Tejera-Muñoz A, Marchant V, Sanz AB, Ortiz A, Goldschmeding R, Ruiz-Ortega M. Acute Kidney Injury is Aggravated in Aged Mice by the Exacerbation of Proinflammatory Processes. Front Pharmacol 2021;12. [CrossRef]
- Di Micco R, Krizhanovsky V, Baker D, d’Adda di Fagagna F. Cellular senescence in ageing: from mechanisms to therapeutic opportunities. Nat Rev Mol Cell Biol 2021;22:75–95. [CrossRef]
- Martin-Sanchez D, Ruiz-Andres O, Poveda J, Carrasco S, Cannata-Ortiz P, Sanchez-Niño MD, Ruiz Ortega M, Egido J, Linkermann A, Ortiz A, Sanz AB. Ferroptosis, but not necroptosis, is important in nephrotoxic folic acid-induced AKI. Journal of the American Society of Nephrology 2017;28:218–29. [CrossRef]
- Robbins PD, Jurk D, Khosla S, Kirkland JL, Lebrasseur NK, Miller JD, Passos JF, Pignolo RJ, Tchkonia T, Niedernhofer LJ. Senolytic Drugs: Reducing Senescent Cell Viability to Extend Health Span n.d.
- Islam MT, Tuday E, Allen S, Kim J, Trott DW, Holland WL, Donato AJ, Lesniewski LA. Senolytic drugs, dasatinib and quercetin, attenuate adipose tissue inflammation, and ameliorate metabolic function in old age. Aging Cell 2023;22. [CrossRef]
- Saccon TD, Nagpal R, Yadav H, Cavalcante MB, Nunes ADDC, Schneider A, Gesing A, Hughes B, Yousefzadeh M, Tchkonia T, Kirkland JL, Niedernhofer LJ, Robbins PD, Masternak MM. Senolytic Combination of Dasatinib and Quercetin Alleviates Intestinal Senescence and Inflammation and Modulates the Gut Microbiome in Aged Mice. Journals of Gerontology - Series A Biological Sciences and Medical Sciences 2021;76:1895–905. [CrossRef]
- Xu M, Pirtskhalava T, Farr JN, Weigand BM, Palmer AK, Weivoda MM, Inman CL, Ogrodnik MB, Hachfeld CM, Fraser DG, Onken JL, Johnson KO, Verzosa GC, Langhi LGP, Weigl M, Giorgadze N, LeBrasseur NK, Miller JD, Jurk D, Singh RJ, Allison DB, Ejima K, Hubbard GB, Ikeno Y, Cubro H, Garovic VD, Hou X, Weroha SJ, Robbins PD, Niedernhofer LJ, Khosla S, Tchkonia T, Kirkland JL. Senolytics improve physical function and increase lifespan in old age. Nat Med 2018;24:1246–56. [CrossRef]
- Xu M, Pirtskhalava T, Farr JN, Weigand BM, Palmer AK, Weivoda MM, Inman CL, Ogrodnik MB, Hachfeld CM, Fraser DG, Onken JL, Johnson KO, Verzosa GC, Langhi LGP, Weigl M, Giorgadze N, LeBrasseur NK, Miller JD, Jurk D, Singh RJ, Allison DB, Ejima K, Hubbard GB, Ikeno Y, Cubro H, Garovic VD, Hou X, Weroha SJ, Robbins PD, Niedernhofer LJ, Khosla S, Tchkonia T, Kirkland JL. Senolytics improve physical function and increase lifespan in old age. Nat Med 2018;24:1246–56. [CrossRef]
- Hickson LTJ, Langhi Prata LGP, Bobart SA, Evans TK, Giorgadze N, Hashmi SK, Herrmann SM, Jensen MD, Jia Q, Jordan KL, Kellogg TA, Khosla S, Koerber DM, Lagnado AB, Lawson DK, LeBrasseur NK, Lerman LO, McDonald KM, McKenzie TJ, Passos JF, Pignolo RJ, Pirtskhalava T, Saadiq IM, Schaefer KK, Textor SC, Victorelli SG, Volkman TL, Xue A, Wentworth MA, Wissler Gerdes EO, Zhu Y, Tchkonia T, Kirkland JL. Senolytics decrease senescent cells in humans: Preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease. EBioMedicine 2019;47:446–56. [CrossRef]
- Zhu X, Zhang C, Liu L, Xu L, Yao L. Senolytic combination of dasatinib and quercetin protects against diabetic kidney disease by activating autophagy to alleviate podocyte dedifferentiation via the Notch pathway. Int J Mol Med 2024;53. [CrossRef]
- Huang Y, Wang B, Hassounah F, Price SR, Klein J, Mohamed TMA, Wang Y, Park J, Cai H, Zhang X, Wang XH. The impact of senescence on muscle wasting in chronic kidney disease. J Cachexia Sarcopenia Muscle 2023;14:126–41. [CrossRef]
- Schafer MJ, White TA, Iijima K, Haak AJ, Ligresti G, Atkinson EJ, Oberg AL, Birch J, Salmonowicz H, Zhu Y, Mazula DL, Brooks RW, Fuhrmann-Stroissnigg H, Pirtskhalava T, Prakash YS, Tchkonia T, Robbins PD, Aubry MC, Passos JF, Kirkland JL, Tschumperlin DJ, Kita H, LeBrasseur NK. Cellular senescence mediates fibrotic pulmonary disease. Nat Commun 2017;8. [CrossRef]
- Manavi Z, Melchor GS, Bullard MR, Gross PS, Ray S, Gaur P, Baydyuk M, Huang JK. Senescent cell reduction does not improve recovery in mice under experimental autoimmune encephalomyelitis (EAE) induced demyelination. J Neuroinflammation 2025;22:101. [CrossRef]
- Nieto M, Könisgberg M, Silva-Palacios A. Quercetin and dasatinib, two powerful senolytics in age-related cardiovascular disease. Biogerontology 2024;25:71–82. [CrossRef]
- Molitoris BA. Therapeutic translation in acute kidney injury: The epithelial/endothelial axis. Journal of Clinical Investigation 2014;124:2355–63. [CrossRef]
- Ferenbach DA, Bonventre JV. Mechanisms of maladaptive repair after AKI leading to accelerated kidney ageing and CKD. Nat Rev Nephrol. 2015;11(5):264-76. [CrossRef]
- Andrade L, Rodrigues CE, Gomes SA, Noronha IL. Acute Kidney Injury as a Condition of Renal Senescence. Cell Transplant 2018;27:739–53. [CrossRef]
- Megyesi J, Udvarhelyi N, Safirstein RL, Price PM. The p534ndependent activation of transcription of p21WAF1/C1P1/SD*1 after acute renal failure. 1996.
- Megyesi J, Safirstein RL, Price PM. p21 Affects Cisplatin Renal Failure Induction of p21 WAF1/CIP1/SDI1 in Kidney Tubule Cells Affects the Course of Cisplatin-induced Acute Renal Failure. vol. 101. 1998.
- Nishioka S, Nakano D, Kitada K, Sofue T, Ohsaki H, Moriwaki K, Hara T, Ohmori K, Kohno M, Nishiyama A. The cyclin-dependent kinase inhibitor p21 is essential for the beneficial effects of renal ischemic preconditioning on renal ischemia/reperfusion injury in mice. Kidney Int 2014;85:871–9. [CrossRef]
- Megyesi J, Andrade L, Vieira JM, Safirstein RL, Price PM. Coordination of the cell cycle is an important determinant of the syndrome of acute renal failure n.d.
- Hochegger K, Koppelstaetter C, Tagwerker A, Huber JM, Heininger D, Mayer G, Rosenkranz AR. p21 and mTERT are novel markers for determining different ischemic time periods in renal ischemia-reperfusion injury. Am J Physiol Renal Physiol 2007;292:762–8.
- Price PM, Safirstein RL, Megyesi J. The cell cycle and acute kidney injury. Kidney Int 2009;76:604–13. [CrossRef]
- Chen J, Zhang H, Yi X, Dou Q, Yang X, He Y, Chen J, Chen K. Cellular senescence of renal tubular epithelial cells in acute kidney injury. Cell Death Discovery 2024 10:1 2024;10:1–13. [CrossRef]
- Della Vedova L, Baron G, Morazzoni P, Aldini G, Gado F. The Potential of Polyphenols in Modulating the Cellular Senescence Process: Implications and Mechanism of Action. Pharmaceuticals 2025;18. [CrossRef]
- Ren Q, Guo F, Tao S, Huang R, Ma L, Fu P. Flavonoid fisetin alleviates kidney inflammation and apoptosis via inhibiting Src-mediated NF-κB p65 and MAPK signaling pathways in septic AKI mice. Biomedicine and Pharmacotherapy 2020;122. [CrossRef]
- Ge C, Xu M, Qin Y, Gu T, Lou D, Li Q, Hu L, Nie X, Wang M, Tan J. Fisetin supplementation prevents high fat diet-induced diabetic nephropathy by repressing insulin resistance and RIP3-regulated inflammation. Food Funct 2019;10:2970–85. [CrossRef]
- Chenxu G, Xianling D, Qin K, Linfeng H, Yan S, Mingxin X, Jun T, Minxuan X. Fisetin protects against high fat diet-induced nephropathy by inhibiting inflammation and oxidative stress via the blockage of iRhom2/NF-κB signaling. Int Immunopharmacol 2021;92. [CrossRef]
- Ren Q, Tao S, Guo F, Wang B, Yang L, Ma L, Fu P. Natural flavonol fisetin attenuated hyperuricemic nephropathy via inhibiting IL-6/JAK2/STAT3 and TGF-β/SMAD3 signaling. Phytomedicine 2021;87. [CrossRef]
- Ju HY, Kim J, Han SJ. The flavonoid fisetin ameliorates renal fibrosis by inhibiting SMAD3 phosphorylation, oxidative damage, and inflammation in ureteral obstructed kidney in mice. Kidney Res Clin Pract 2023;42:325–39. [CrossRef]
- Prasath GS, Subramanian SP. Modulatory effects of fisetin, a bioflavonoid, on hyperglycemia by attenuating the key enzymes of carbohydrate metabolism in hepatic and renal tissues in streptozotocin-induced diabetic rats. Eur J Pharmacol 2011;668:492–6. [CrossRef]
- Liu AB, Tan B, Yang P, Tian N, Li JK, Wang SC, Yang LS, Ma L, Zhang JF. The role of inflammatory response and metabolic reprogramming in sepsis-associated acute kidney injury: mechanistic insights and therapeutic potential. Front Immunol 2024;15. [CrossRef]
- Ren Q, Guo F, Tao S, Huang R, Ma L, Fu P. Flavonoid fisetin alleviates kidney inflammation and apoptosis via inhibiting Src-mediated NF-κB p65 and MAPK signaling pathways in septic AKI mice. Biomedicine and Pharmacotherapy 2020;122. [CrossRef]
- Valentijn FA, Knoppert SN, Pissas G, Rodrigues-Diez RR, Marquez-Exposito L, Broekhuizen R, Mokry M, Kester LA, Falke LL, Goldschmeding R, Ruiz-Ortega M, Eleftheriadis T, Nguyen TQ. Ccn2 aggravates the immediate oxidative stress–dna damage response following renal ischemia–reperfusion injury. Antioxidants 2021;10. [CrossRef]
- Rayego-Mateos S, Marquez-Exposito L, Basantes P, Tejedor-Santamaria L, Sanz AB, Nguyen TQ, Goldschmeding R, Ortiz A, Ruiz-Ortega M. CCN2 Activates RIPK3, NLRP3 Inflammasome, and NRF2/Oxidative Pathways Linked to Kidney Inflammation. Antioxidants 2023;12. [CrossRef]
- Xu M, Tchkonia T, Ding H, Ogrodnik M, Lubbers ER, Pirtskhalava T, White TA, Johnson KO, Stout MB, Mezera V, Giorgadze N, Jensen MD, LeBrasseur NK, Kirkland JL. JAK inhibition alleviates the cellular senescence-associated secretory phenotype and frailty in old age. Proc Natl Acad Sci U S A 2015;112:E6301–10. [CrossRef]
- Hernández-Silva D, López-Abellán MD, Martínez-Navarro FJ, García-Castillo J, Cayuela ML, Alcaraz-Pérez F. Development of a Short Telomere Zebrafish Model for Accelerated Aging Research and Antiaging Drug Screening. Aging Cell 2025. [CrossRef]
- Zhang L, Pitcher LE, Prahalad V, Niedernhofer LJ, Robbins PD. Targeting cellular senescence with senotherapeutics: senolytics and senomorphics. FEBS Journal 2023;290:1362–83. [CrossRef]
- Kuro-O* M, Matsumura Y, Aizawa H, Kawaguchi H, Suga T, Utsugi T, Ohyama Y, Kurabayashi M, Kaname T, Kumek E, Iwasakik H, Iida A, Shiraki-Iida T, Nishikawa# S, Nagai R, Yo-Ichi Nabeshima &. Mutation of the mouse klotho gene leads to a syndrome resembling ageing. vol. 390. 1997.
- Izquierdo MC, Perez-Gomez M V., Sanchez-Niño MD, Sanz AB, Ruiz-Andres O, Poveda J, Moreno JA, Egido J, Ortiz A. Klotho, phosphate and inflammation/ageing in chronic kidney disease. Nephrology Dialysis Transplantation 2012;27. [CrossRef]
- Li S, Yu L, He A, Liu Q. Klotho inhibits unilateral ureteral obstruction-induced endothelial-to-mesenchymal transition via TGF-β1/Smad2/Snail1 signaling in mice. Front Pharmacol 2019;10. [CrossRef]
- Zhu Y, Langhi Prata LGP, Wissler Gerdes EO, Machado J, Netto E, Pirtskhalava T, Giorgadze N, Tripathi U, Inman CL, Johnson KO, Xue A, Palmer AK, Chen T, Schaefer K, Justice JN, Nambiar AM, Musi N, Kritchevsky SB, Chen J, Khosla S, Jurk D, Schafer MJ, Tchkonia T, Kirkland JL. Orally-active, clinically-translatable senolytics restore a-Klotho in mice and humans 2022.
- Castillo RF. Pathophysiologic Implications and Therapeutic Approach of Klotho in Chronic Kidney Disease: A Systematic Review. Laboratory Investigation 2023;103. [CrossRef]
- Wang C, Jurk D, Maddick M, Nelson G, Martin-ruiz C, Von Zglinicki T. DNA damage response and cellular senescence in tissues of aging mice. Aging Cell 2009;8:311–23. [CrossRef]
- Zhu Y, Tchkonia T, Pirtskhalava T, Gower AC, Ding H, Giorgadze N, Palmer AK, Ikeno Y, Hubbard GB, Lenburg M, O’hara SP, Larusso NF, Miller JD, Roos CM, Verzosa GC, Lebrasseur NK, Wren JD, Farr JN, Khosla S, Stout MB, McGowan SJ, Fuhrmann-Stroissnigg H, Gurkar AU, Zhao J, Colangelo D, Dorronsoro A, Ling YY, Barghouthy AS, Navarro DC, Sano T, Robbins PD, Niedernhofer LJ, Kirkland JL. The achilles’ heel of senescent cells: From transcriptome to senolytic drugs. Aging Cell 2015;14:644–58. [CrossRef]
- Marquez-Exposito L, Tejedor-Santamaria L, Santos-Sanchez L, Valentijn FA, Cantero-Navarro E, Rayego-Mateos S, Rodrigues-Diez RR, Tejera-Muñoz A, Marchant V, Sanz AB, Ortiz A, Goldschmeding R, Ruiz-Ortega M. Acute Kidney Injury is Aggravated in Aged Mice by the Exacerbation of Proinflammatory Processes. Front Pharmacol 2021;12. [CrossRef]
- Baar MP, Brandt RMC, Putavet DA, Klein JDD, Derks KWJ, Bourgeois BRM, Stryeck S, Rijksen Y, van Willigenburg H, Feijtel DA, van der Pluijm I, Essers J, van Cappellen WA, van IJcken WF, Houtsmuller AB, Pothof J, de Bruin RWF, Madl T, Hoeijmakers JHJ, Campisi J, de Keizer PLJ. Targeted Apoptosis of Senescent Cells Restores Tissue Homeostasis in Response to Chemotoxicity and Aging. Cell 2017;169:132-147.e16. [CrossRef]
- Marquez-Exposito L, Tejedor-Santamaria L, Valentijn FA, Tejera-Muñoz A, Rayego-Mateos S, Marchant V, Rodrigues-Diez RR, Rubio-Soto I, Knoppert SN, Ortiz A, Ramos AM, Goldschmeding R, Ruiz-Ortega M. Oxidative Stress and Cellular Senescence Are Involved in the Aging Kidney. Antioxidants 2022;11. [CrossRef]
Figure 1.
Dasatinib plus Quercetin (D&Q) did not prevent renal dysfunction in AKI-FAN. Mice were injected with folic acid (125 mg/kg intraperitoneal injection) and sacrificed 48 h later. Two groups were studied: one received a single oral dose of the combination of D&Q (Dasatinib; 5 mg/kg body mass and Quercetin; 50 mg/kg body mass, via oral gavage), another group remained untreated. A) BUN and B) Creatinine were evaluated. Data are expressed as the mean of 5-6 animals per group ± SEM. *p < 0.05. One-way ANOVA was followed by the Holm-Šídák multiple comparison test for BUN while for creatinine, the Kruskal-Wallis non-parametric statistical test was followed by Dunn’s test without correction.
Figure 1.
Dasatinib plus Quercetin (D&Q) did not prevent renal dysfunction in AKI-FAN. Mice were injected with folic acid (125 mg/kg intraperitoneal injection) and sacrificed 48 h later. Two groups were studied: one received a single oral dose of the combination of D&Q (Dasatinib; 5 mg/kg body mass and Quercetin; 50 mg/kg body mass, via oral gavage), another group remained untreated. A) BUN and B) Creatinine were evaluated. Data are expressed as the mean of 5-6 animals per group ± SEM. *p < 0.05. One-way ANOVA was followed by the Holm-Šídák multiple comparison test for BUN while for creatinine, the Kruskal-Wallis non-parametric statistical test was followed by Dunn’s test without correction.
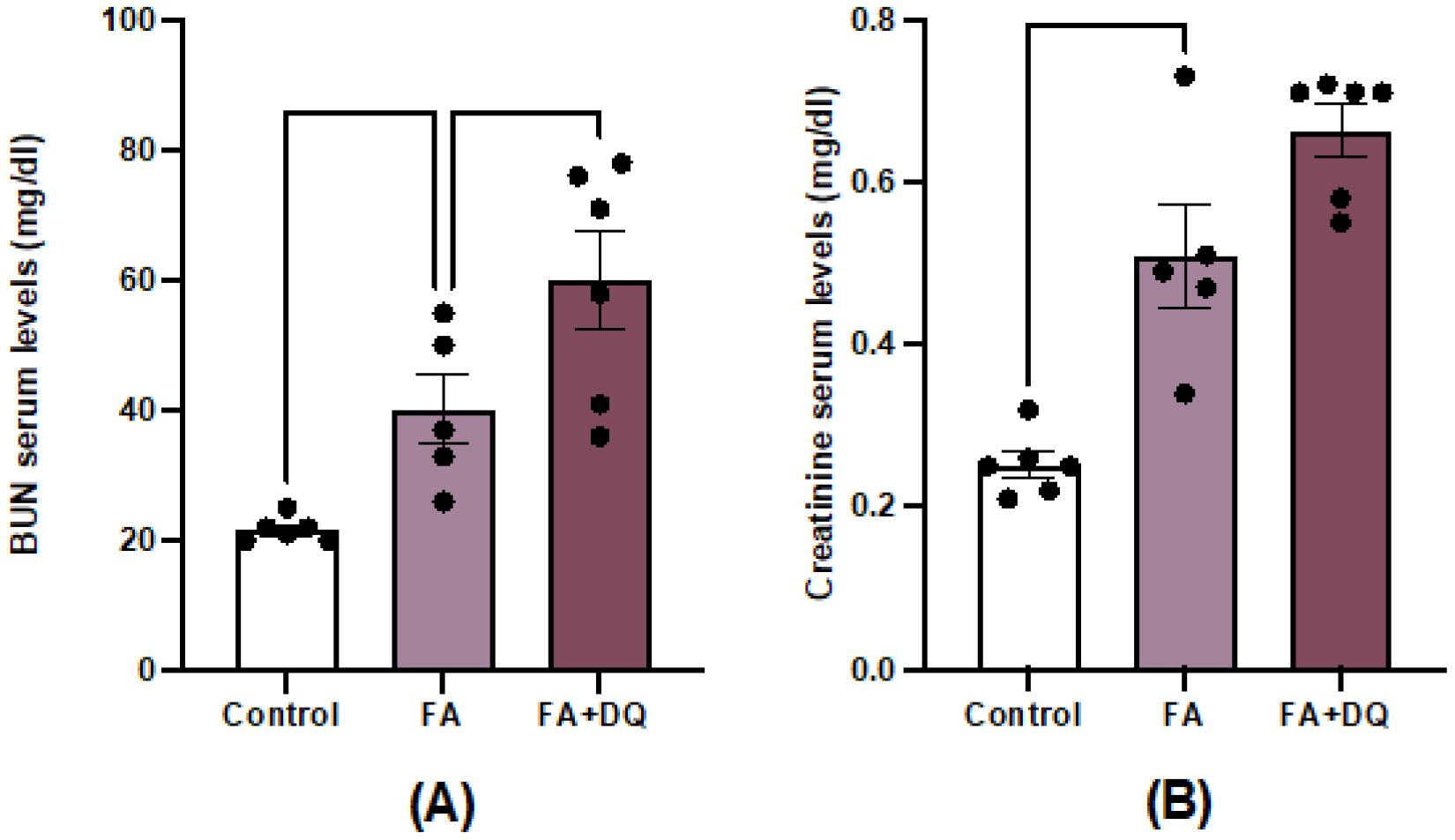
Figure 2.
Dasatinib plus Quercetin (D&Q) increased the gene expression of kidney injury biomarkers in AKI-FAN. RNA was extracted from total kidney cortex and Lcn2 and Havcr1 gene expression was evaluated by real time PCR. Data are expressed as the mean of 6-8 animals per group ± SEM. *p < 0.05, **p < 0.005, ***p < 0.0005. One-way ANOVA was followed by the Holm-Šídák multiple comparison test.
Figure 2.
Dasatinib plus Quercetin (D&Q) increased the gene expression of kidney injury biomarkers in AKI-FAN. RNA was extracted from total kidney cortex and Lcn2 and Havcr1 gene expression was evaluated by real time PCR. Data are expressed as the mean of 6-8 animals per group ± SEM. *p < 0.05, **p < 0.005, ***p < 0.0005. One-way ANOVA was followed by the Holm-Šídák multiple comparison test.
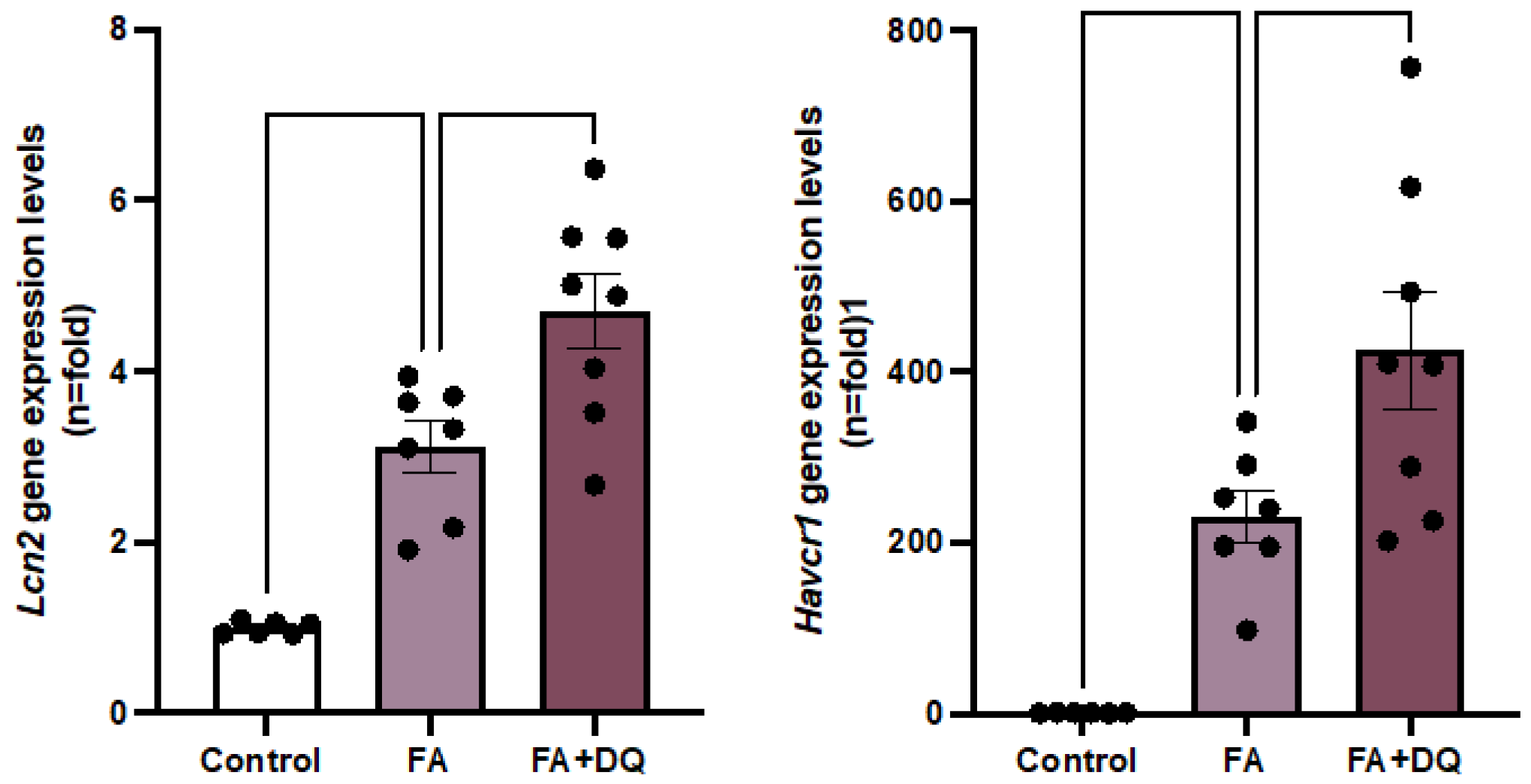
Figure 3.
Dasatinib + Quercetin (D&Q) did not modify the tubular damage marker KIM-1 in AKI-FAN. A) Representative microphotographs of KIM-1 immunohistochemistry at 20× magnification. B) Quantification of KIM-1 immunohistochemical staining expressed as mean stained area relative to the total area, represented as n-fold. Data are expressed as mean of 6 to 9 animals per group ± SEM. **p < 0.005. The Kruskal-Wallis non-parametric statistical test was followed by Dunn’s test without correction.
Figure 3.
Dasatinib + Quercetin (D&Q) did not modify the tubular damage marker KIM-1 in AKI-FAN. A) Representative microphotographs of KIM-1 immunohistochemistry at 20× magnification. B) Quantification of KIM-1 immunohistochemical staining expressed as mean stained area relative to the total area, represented as n-fold. Data are expressed as mean of 6 to 9 animals per group ± SEM. **p < 0.005. The Kruskal-Wallis non-parametric statistical test was followed by Dunn’s test without correction.
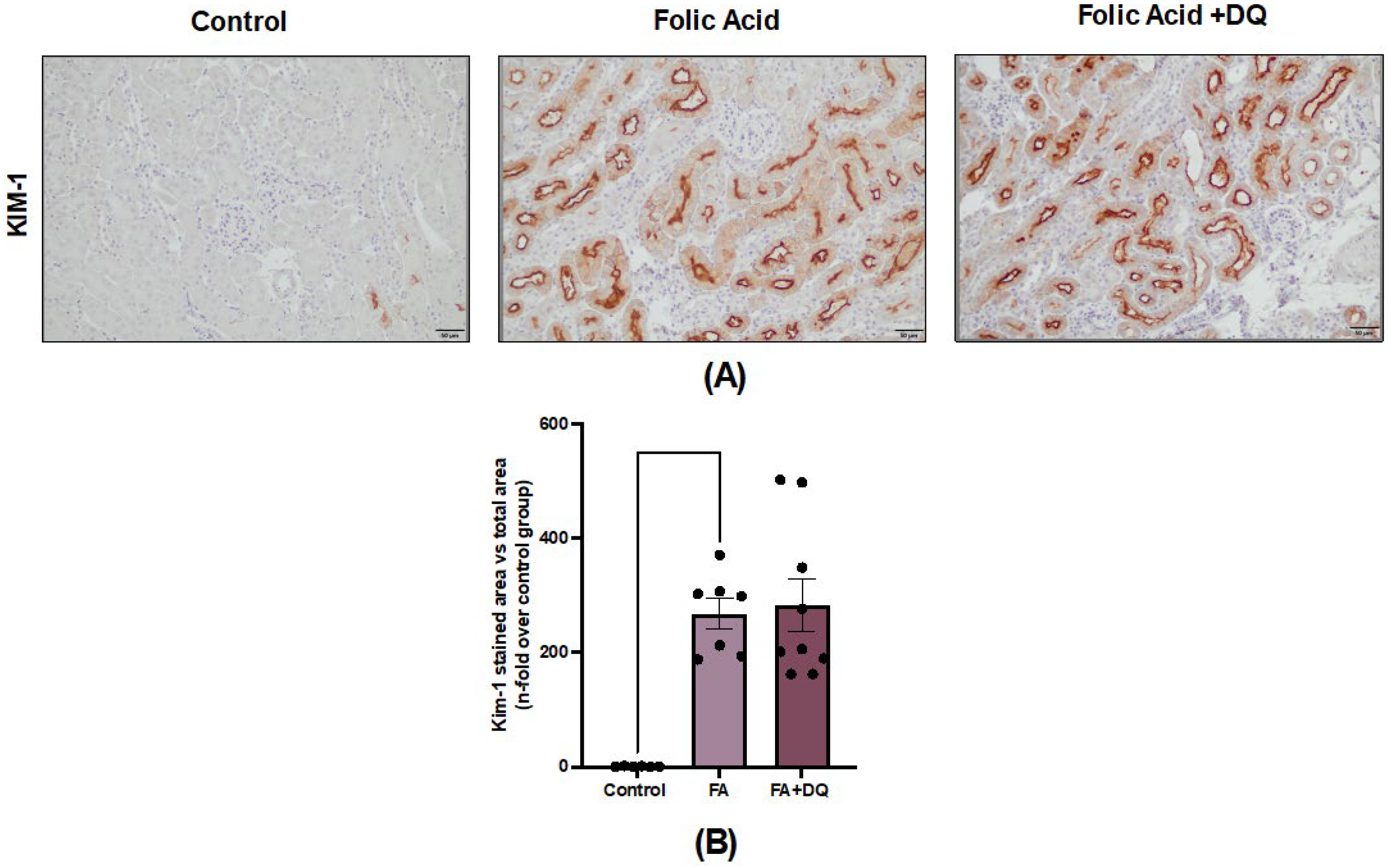
Figure 4.
Dasatinib + Quercetin (D&Q) did not modify the number of senescent cells in AKI-FAN. A) Representative microphotograph of p21 immunohistochemistry at 20× magnification. B) Representative microphotograph of p21 immunohistochemistry at 40× magnification. C) Quantification of p21 positive cells in eight fields for each sample, represented as n-fold. Data are expressed as the mean of 6 to 7 animals per group ± SEM. **p < 0.005. The Kruskal-Wallis non-parametric statistical test was followed by Dunn’s test without correction.
Figure 4.
Dasatinib + Quercetin (D&Q) did not modify the number of senescent cells in AKI-FAN. A) Representative microphotograph of p21 immunohistochemistry at 20× magnification. B) Representative microphotograph of p21 immunohistochemistry at 40× magnification. C) Quantification of p21 positive cells in eight fields for each sample, represented as n-fold. Data are expressed as the mean of 6 to 7 animals per group ± SEM. **p < 0.005. The Kruskal-Wallis non-parametric statistical test was followed by Dunn’s test without correction.
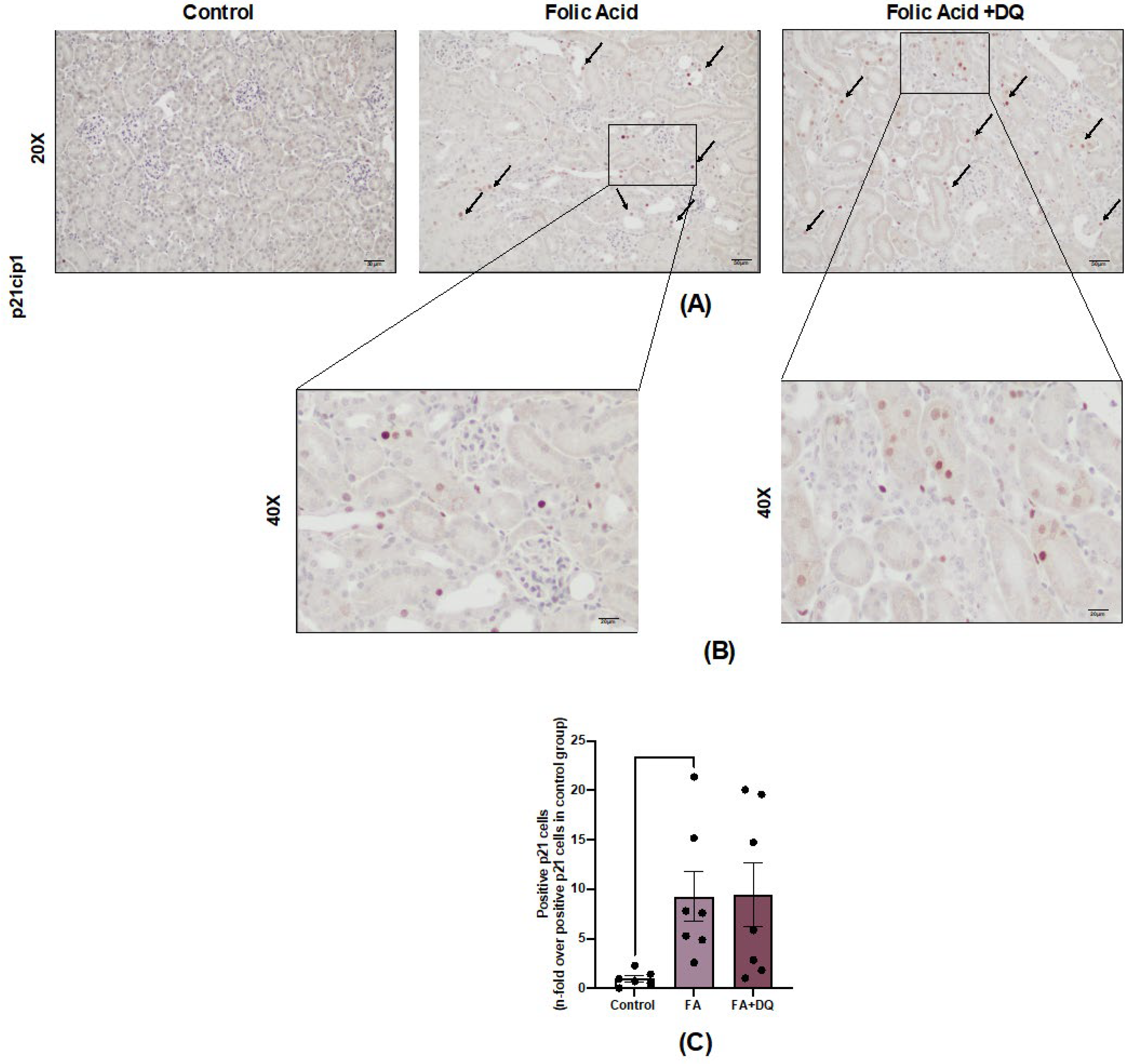
Figure 5.
Dasatinib + Quercetin (D&Q) did not decrease senescence-associated markers in AKI-FAN. The D&Q treatment effect on gene expression was evaluated by RT qPCR. A) Cdkn1a B) Cdkn2a, the SASP components C) Ccl2 D) Ccl5 E) IL1β and F) the anti-aging factor Klotho. Data are expressed as the mean of 6 to 7 animals per group ± SEM. *p < 0.05 **p < 0.005 ***p < 0.0005. One-way ANOVA was followed by the Holm-Šídák multiple comparison test, except for Ccl5 where the Kruskal-Wallis non-parametric statistical test was followed by Dunn’s test without correction.
Figure 5.
Dasatinib + Quercetin (D&Q) did not decrease senescence-associated markers in AKI-FAN. The D&Q treatment effect on gene expression was evaluated by RT qPCR. A) Cdkn1a B) Cdkn2a, the SASP components C) Ccl2 D) Ccl5 E) IL1β and F) the anti-aging factor Klotho. Data are expressed as the mean of 6 to 7 animals per group ± SEM. *p < 0.05 **p < 0.005 ***p < 0.0005. One-way ANOVA was followed by the Holm-Šídák multiple comparison test, except for Ccl5 where the Kruskal-Wallis non-parametric statistical test was followed by Dunn’s test without correction.
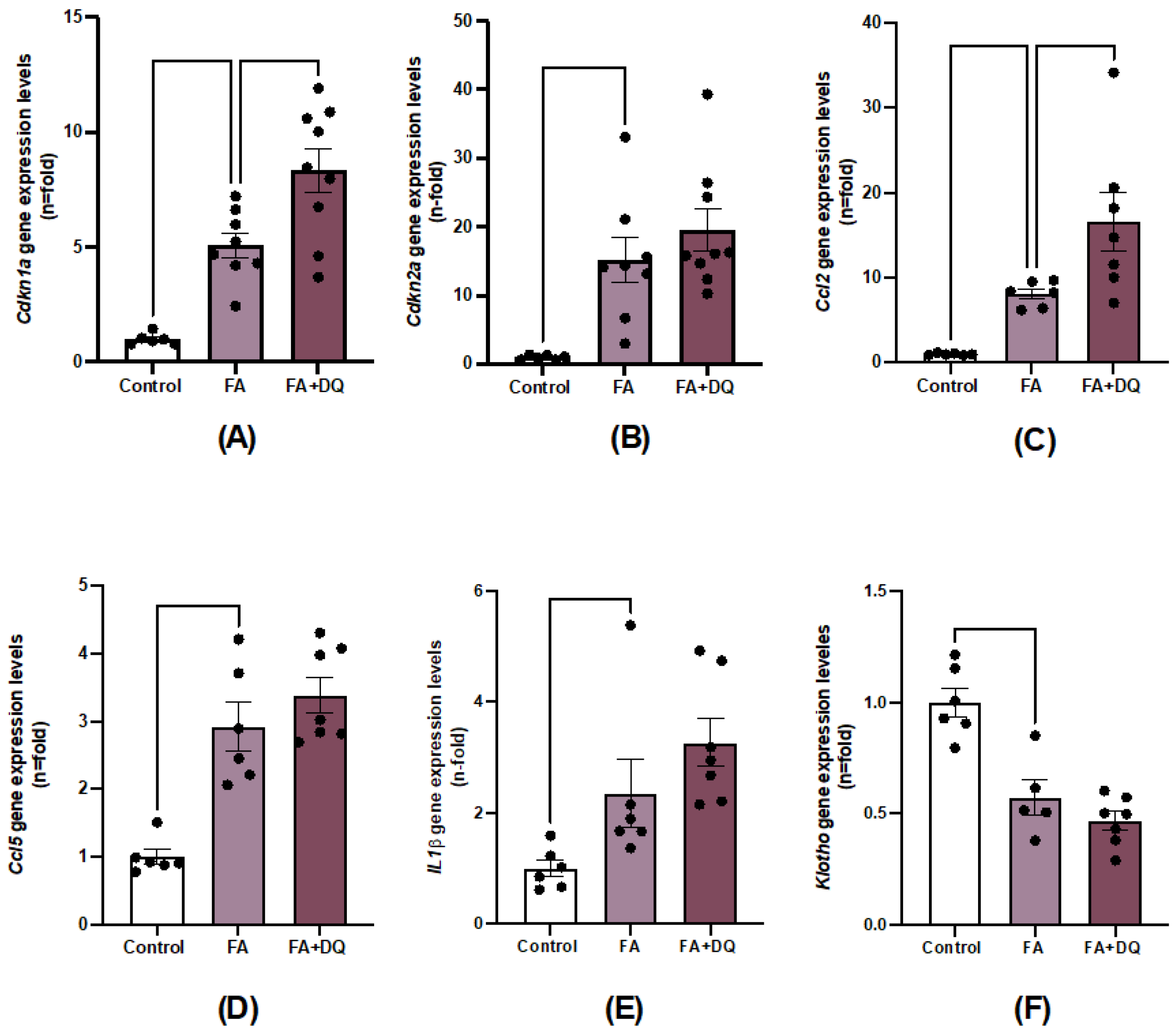
Figure 6.
Dasatinib + Quercetin (D&Q) did not increase apoptosis regulatory molecules in AKI-FAN. The impact of D&Q treatment on apoptosis was evaluated both at the gene (RT qPCR) and protein levels. A) BAX B) BCL2L1 and at protein levels C) Representative microphotographs of positive BCLxL per field at 20× magnification. D) Quantification of BCLxL immunohistochemical staining expressed as mean stained area relative to the total area, represented as n-fold. Data are expressed as mean of 4 to 9 animals per group ± SEM. *p < 0.05. The Kruskal-Wallis non-parametric statistical test followed by Dunn’s test without correction was performed.
Figure 6.
Dasatinib + Quercetin (D&Q) did not increase apoptosis regulatory molecules in AKI-FAN. The impact of D&Q treatment on apoptosis was evaluated both at the gene (RT qPCR) and protein levels. A) BAX B) BCL2L1 and at protein levels C) Representative microphotographs of positive BCLxL per field at 20× magnification. D) Quantification of BCLxL immunohistochemical staining expressed as mean stained area relative to the total area, represented as n-fold. Data are expressed as mean of 4 to 9 animals per group ± SEM. *p < 0.05. The Kruskal-Wallis non-parametric statistical test followed by Dunn’s test without correction was performed.

Figure 7.
Treatment with Dasatinib + Quercetin (D&Q) did not regulate the necroptosis pathway in AKI-FAN. Renal gene expression levels of the necroptosis pathways components A) Ripk3 B) Mlkl were analyzed by RT-qPCR. Data are expressed as the mean of 6 to 8 animals per group ± SEM. *p < 0.05 **p < 0.005 ***p < 0.0005 ****p<0.00005. One-way ANOVA was followed by the Holm-Šídák multiple comparison test.
Figure 7.
Treatment with Dasatinib + Quercetin (D&Q) did not regulate the necroptosis pathway in AKI-FAN. Renal gene expression levels of the necroptosis pathways components A) Ripk3 B) Mlkl were analyzed by RT-qPCR. Data are expressed as the mean of 6 to 8 animals per group ± SEM. *p < 0.05 **p < 0.005 ***p < 0.0005 ****p<0.00005. One-way ANOVA was followed by the Holm-Šídák multiple comparison test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



