
Submitted:
21 April 2025
Posted:
22 April 2025
You are already at the latest version
Abstract
It is a critical strategy for controlling agricultural non-point source pollution in the Yellow River Basin to utilize emergent plants for effectively absorbing nitrogen from water and converting it into organic nitrogen through nitrogen assimilation enzyme activity, thereby reducing nitrogen concentrations in water. However, the uptake rate and mechanism of different forms of nitrogen by emergent plants are still unclear. This study determined the nitrogen uptake rates, nitrogen assimilase activities, root properties, and photosynthetic parameters of four emergent plants, including Phragmites australis, Typha orientalis, Scirpus validus, and Lythrum salicaria, under five NH4+/NO3- ratios (9:1, 7:3, 5:5, 3:7 and 1:9) using 15N hydroponic simulations. The results demonstrated that both nitrogen form and plant species significantly influenced the nitrogen uptake rates of emergent plants. In water bodies with varying NH4+/NO3- ratios, P. australis and T. orientalis exhibited significantly higher inorganic nitrogen uptake rates compared to S. validus and L. salicaria, increasing by 11.83%–114.69% and 14.07%–130.46%, respectively. When the ratio of NH4+/NO3- in the water body is 9:1, the uptake rate of inorganic nitrogen by P. australis reaches its peak, which is 729.20 μg·N·g-1·h-1DW. When the ratio of NH4+/NO3- was 5:5, the uptake rate of T. orientalis was the highest, reaching 763.71 μg·N·g-1·h-1DW. Plant preferences for nitrogen forms exhibited significant environmental plasticity. At an NH4+/NO3- ratio of 5:5, P. australis and T. orientalis preferred NO3--N, whereas S. validus and L. salicaria favored NH4+-N. The uptake rate of NH4+-N by the four plants was significantly positively correlated with GS and GOGAT activities, while the uptake rate of NO3--N was significantly positively correlated with NR activity. These findings indicate that the nitrogen uptake and assimilation processes of these four plant species involve synergistic mechanisms of environmental adaptation and physiological regulation, enabling more effective utilization of different nitrogen forms in water. Additionally, the uptake rate of NH4+-N by P. australis and T. orientalis was significantly positively correlated with GDH, suggesting that they are better adapted to eutrophication via the GDH pathway. Specific root surface area plays a crucial role in regulating nitrogen uptake rates in plants. Nitrogen uptake amount exerted the greatest total impact on the nitrogen uptake rate, followed by root traits and nitrogen assimilation enzymes. Therefore, there were significant interspecific differences in the uptake rates and physiological response mechanisms of emergent plants to various nitrogen forms. It is recommended to prioritize the use of highly adaptable emergent plants such as P. australis and T. orientalis in the Yellow River irrigation area.
Keywords:
Nitrogen Uptake Rate
; NH4+/NO3- Ratio
; Nitrogen Assimilation Enzyme Activity
; Phragmites australis
; Typha orientalis
1. Introduction
Nitrogen plays a crucial role in plant physiological metabolism, growth and development, as well as in the nutrient cycling of ecosystems [1]. As a key component of aquatic ecosystems, aquatic plants are capable of efficiently absorbing nitrogen from water, and assimilating it into organic nitrogen such as amino acids and proteins via nitrogen assimilation enzyme activity. This process not only supports plant growth and metabolism but also contributes to the reduction of nitrogen concentrations in water [2]. Consequently, aquatic plants are essential for nitrogen cycling and water quality improvement in aquatic systems, making them a significant focus of contemporary research [3]. Different forms of nitrogen significantly influence the efficiency of nitrogen metabolism and the ecological functions of plants by modulating the activities of key enzymes involved in nitrogen assimilation [4]. Plants convert inorganic nitrogen into organic nitrogen via enzyme systems, including Nitrate Reductase (NR), Nitrite Reductase (NiR), Glutamine Synthetase (GS), Glutamate Synthase (GOGAT), and Glutamate Dehydrogenase (GDH). This process is fundamental to nitrogen uptake and assimilation in plants [5]. As illustrated in Figure 1-1, the nitrogen uptake and assimilation processes in plants proceed as follows: Aquatic plants are proficient in directly absorbing dissolved nitrogenous compounds by roots from the substrate, specifically ammonium nitrogen (NH4+-N) and nitrate nitrogen (NO3--N). The absorbed nitrogen is subsequently transported to various plant organs, such as stems and leaves. Through enzymatic activities associated with nitrogen assimilation, these inorganic nitrogen forms are converted into organic molecules, such as amino acids and proteins [6]. This process not only facilitates nutrient storage within the plant but also significantly contributes to plant growth and metabolism, thereby reducing nitrogenous pollutants in aquatic environments [2,7]. The primary biochemical pathways involved include: (1) Nitrate Reduction: NR in the cytoplasm reduces nitrate ions (NO3-) to nitrite ions (NO2-), which are further reduced to ammonium ions (NH4+) by NiR located in chloroplasts and plastids of root cells. (2) Ammonium Assimilation: GS in the cytoplasm and plastids catalyzes the reaction between NH4+ and glutamic acid (Glu) to form glutamine (Gln). Subsequently, GOGAT in chloroplasts and root cells catalyzes the reaction between Gln and α-ketoglutaric acid (α-KG) to produce two molecules of Glu. (3) Regulation under Ammonium Availability: In conditions of high ammonium concentration, GDH in mitochondria and the cytoplasm facilitates the binding of α-KG with NH4+ to generate Glu. Conversely, during ammonium starvation, GDH can catalyze the reverse reaction, decomposing Glu into α-KG and NH4+ [8,9].
The activities of key enzymes involved in nitrogen assimilation in aquatic plants are influenced by both water nitrogen forms and environmental factors. NO3--N predominantly induces NR and NiR activity [10], while NH4+-N predominantly induces GS GOGAT, andGDH activity [11]. NR serves as a rate-limiting enzyme for the assimilation of NO3--N, catalyzing the reduction of NO3- to NO2-, with its activity significantly affected by nitrogen form. Under NO3--N supply conditions, NR activity is high, whereas it is relatively low under NH4+-N supply conditions. NiR is a key enzyme that reduces NO2- to NH4+, and its activity is positively correlated with NR activity. However, in high NH4 environments, the activities of both NR and NiR may be inhibited [12]. GS and GOGAT together constitute the GS/GOGAT cycle, which is a critical metabolic pathway for plants to assimilate NH4+ into organic nitrogen [13]. Due to synergistic effects within this cycle, GOGAT activity is usually positively correlated with GS activity, and both GS and GOGAT activities are generally higher in plants when NH4-N is the primary nitrogen source [14]. The GDH pathway is one of the key pathways involved in nitrogen metabolism and energy metabolism [15]. GDH activity in aquatic plants is generally higher when NH4-N is the primary nitrogen source [11]. Additionally, GDH activity may significantly increase under stress conditions, serving as an important mechanism for plant ammonium detoxification [15,16].
Different forms of nitrogen (e.g., NH4+-N, NO3--N, etc.) in water may have different migration and transformation rules and bioavailability, which may lead to different absorption and utilization efficiency of different forms of nitrogen by aquatic plants [17]. Plant roots adopt different nitrogen uptake and assimilation strategies to sustain normal growth and development in response to changes in the availability and morphology of nitrogen in water [18], and these pathways and strategies are influenced by a combination of factors such as plant species, nitrogen morphology in the environment, and Ammonium to nitrate ratio (NH4+/NO3-) [19,20]. Different plant species demonstrate distinct nutrient preferences and utilization strategies across varying habitats [18]. Research has indicated that the absorption of amino acids and nitrogen utilization in plants are influenced by the form of the nitrogen source and exhibit a certain degree of adaptability to environmental changes in nitrogen forms [21]. NH4+-N treatment significantly enhanced rice biomass and nitrogen accumulation, demonstrating greater efficacy compared to NO3--N treatment in promoting rice growth. Furthermore, mixed nitrogen (NH4+-N and NO3--N) exhibited superior performance in increasing plant biomass and nitrogen use efficiency compared to single nitrogen forms [22]. In sandy constructed wetlands, varying NH4+/NO3- ratios directly influence plant diversity, nitrogen removal efficiency, and ecosystem stability [23]. Increasing the proportion of NH4+ can enhance the GOGAT and GS activities in plants involved in NH4+ assimilation, thereby accelerating nitrogen metabolism [24].
Significant differences were observed in nitrogen uptake preferences among different plant species, primarily characterized by a preference for either NH4+-N or NO3--N. In temperate desert ecosystems, ephemeral plants such as Centaurea pulchella and Lactuca undulata predominantly favor NO3--N absorption, whereas annual plants like Ceratocarpus arenarius and Suaeda glauca exhibit a stronger affinity for NH4+-N absorption [25]. While plants can efficiently utilize the available nitrogen forms in their growth medium, their preferences for specific nitrogen forms and rates of uptake may vary depending on geographic location, soil properties, climatic conditions, and other environmental factors [26]. Soil NH4+-N was the most preferred N form by plants in (sub)tropical regions, whereas NO3--N preference was significantly higher in high-latitude than low-latitude regions [27]. In a semi-arid grassland in Inner Mongolia, the three species of plants (Leymus chinensis, Stipa grandis and Cleistogenes squarrosa) altered their N-uptake preference in response to grazing intensity differently in the early growth and vigorous growth stages [28].
Different nitrogen forms exerted significant regulatory effects on root morphology and function. NH4+ generally enhances the accumulation of root biomass, leading to a significant increase in total root length, root surface area, and lateral root density [29]. The strategy of sacrificing the taproot and enhancing lateral root development may be linked to the higher energy demands for NO3- assimilation [30], which compels plants to enhance nitrogen capture efficiency by increasing the root absorption area. Modifying specific root length (SRL) and specific root surface area (SRA) to adjust nutrient uptake capacity constitutes a key adaptive mechanism for plants in response to environmental changes. [31] In addition, nitrogen is a crucial constituent of chlorophyll and photosynthetic enzymes, and varying nitrogen forms significantly impact plant photosynthetic efficiency [32].
Although the uptake rates and physiological characteristics of different nitrogen forms in aquatic plants have been studied, there is still a paucity of reports regarding the regulatory mechanisms of nitrogen metabolism under mixed nitrogen (NH4+-N and NO3--N) conditions. Specifically, the coupling mechanism among activity of key enzymes involved in nitrogen assimilation, root traits, photosynthetic characteristics, and nitrogen uptake rates remains to be fully elucidated. Consequently, it is crucial to investigate the uptake rates and physiological synergistic mechanisms of NH4+-N and NO3--N uptake by aquatic plants in aquatic environments. The Ningxia Yellow River Irrigation Area serves as a crucial irrigation zone and commercial grain production base in Northwest China, contributing over two-thirds of the output value of grain crops and agriculture in Ningxia. It plays a significant role in driving regional economic development [33]. The long-term extensive agricultural management practices, such as the overuse of chemical fertilizers and excessive irrigation, have led to water bodies in the Yellow River Basin confronting an increasingly severe issue of agricultural non-point source pollution. [34]. The utilization of aquatic phytoremediation technology for the removal of excess nitrogen from agricultural water bodies exhibits remarkable purification efficiency and resource recycling potential [35]. This constitutes a pivotal strategy for controlling agricultural non-point source pollution in the Yellow River irrigation area and for preventing and alleviating potential pollution risks to the Yellow River [36]. Furthermore, it signifies an emerging priority in the advancement of ecological environment research and pollution prevention and control technologies within the basin [37]. Emergent plants are a category of aquatic plants whose roots are embedded in mud, with the lower part of the stem or base submerged in water, and photosynthetic structures such as stems and leaves protruding above the water surface [38]. These plants display a wide ecological range and fulfill multiple ecological functions, including wind resistance, pollution tolerance, decontamination capacity, and the alleviation of eutrophication in aquatic ecosystems. Furthermore, they exhibit superior resilience compared to floating and submerged plants [39], and are widely distributed across the wetlands in Northwest China's arid region. The synergistic effect of pollutant removal achieved through the absorption and assimilation capabilities of water-lifting plants, in conjunction with the degradation and transformation activities of rhizosphere microorganisms [40,41], constitutes an environmentally friendly technology marked by low resource input, minimal energy consumption, and the prevention of secondary pollution. This approach has emerged as a pivotal strategy for ensuring sustained water quality in irrigation regions.
Therefore, in this study, four water-raising plants: P. australis, T. orientalis, S. validus, and L. salicaria were used as the research objects. In the experimental water maintained at a constant nitrogen concentration, five NH4+/NO3- ratios (9:1, 7:3, 5:5, 3:7, and 1:9) labeled with 15N isotopes were established. The nitrogen uptake rate, nitrogen assimilation enzyme activity, root properties and photosynthetic parameters of plants were measured to verify the following two hypotheses: (1) different forms of nitrogen significantly affected the nitrogen uptake rate and nitrogen assimilation-related enzyme activities of emergent plants; (2) The influencing factors of different forms of nitrogen uptake rate of emergent plants were mainly plant root attributes, nitrogen assimilation enzyme activity and photosynthetic parameters. This paper analyzes the responses of nitrogen uptake rate and physiological characteristics of emergent plants to different forms of nitrogen (NH4+-N, NO3--N), clarifies the adaptation strategies of plants to nitrogen forms, reveals the removal mechanism of nitrogen pollution by emergent plants, provides a scientific basis for optimizing the efficient purification of emergent plants and using emergent plants to remediate water nitrogen pollution, and deepens the correlation mechanism of "morphology-plant-efficiency" in the process of nitrogen removal from water.
2. Results
2.1. Nitrogen Uptake Rates of Emergent Plants in Water with Different NH4+/NO3- Ratios
The nitrogen form, plant species, NH4+/NO3- ratio, and their interactions significantly affected the nitrogen uptake rates of the four emergent plants (p < 0.01). Among these factors, the nitrogen form was the most influential, followed by plant species (Table 1).
As illustrated in Figure 2, in the experimental water with different NH4+/NO3- ratios, the uptake rates of inorganic nitrogen for P. australis and T. orientalis were significantly higher than those of the other two plant species (p < 0.05), with increases of 11.83–114.69% and 14.07–130.46%, respectively. When the NH4+/NO3- ratio was 9:1, the uptake rate of inorganic nitrogen peaked at 729.20 μg N·g-1·h-1DW for P. australis and 477.36 μg N·g-1·h-1DW for S. validus. By contrast, when the NH4+/NO3- ratio was 5:5, the highest uptake rates of inorganic nitrogen were observed in T. orientalis (763.71 μg N·g-1·h-1DW) and L. salicaria (609.92 μg N·g-1·h-1DW).
When NH4+-N was dominant (NH4+/NO3- ratios of 9:1 and 7:3), P. australis, T. orientalis, S. validus, and L. salicaria exhibited a preference for absorbing NH4+-N. When the NH4+/NO3- ratio was 9:1, the ratios of NH4+-N to NO3--N uptake rates were 2.66, 2.03, 4.32, and 3.86, respectively. When the NH4+/NO3- ratio was 7:3, the ratios of NH4+-N to NO3--N uptake rates were 2.05, 2.08, 2.29, and 1.48, respectively. When the NH4+/NO3- ratio was 1:9, the four emergent plants preferred to absorb NO3--N, with the ratios of NH4+-N to NO3--N uptake rates being 0.56, 0.53, 0.71, and 0.58, respectively. This indicates that the inorganic nitrogen uptake rates of these four species exhibit a certain degree of environmental plasticity. When the NH4+/NO3- ratio was 5:5, the four plants showed varying preferences for nitrogen forms. P. australis and T. orientalis preferred NO3--N absorption, while S. validus and L. salicaria favored NH4+-N absorption, with the respective uptake rate ratios being 0.77, 0.63, 4.50, and 1.64.
2.2. Nitrogen Assimilation Enzyme Activities of Emergent Plants in Water with Different NH4+/NO3- Ratios
As illustrated in Figure 3, with increasing NH4+/NO3- ratios, the contents of Soluble protein (SP) and NR activity decreased in the four emergent plants. In contrast,GS activity increased, while GDH activity initially increased and then decreased. There was no obvious regularity in the changes of NiR and GOGAT activities. When the NH4+/NO₃⁻ ratio was 1:9, the SP content in T. orientalis stems, leaves, and roots reached its highest levels, at 16.96 mg·g-1FW for stems and leaves, and 14.83 mg·g-1FW for roots. Similarly, NR activity in T. orientalis stems, leaves, and roots was also highest, at 43.08 μmol·h-1·g-1 FW for stems and leaves, and 46.62 μmol·h-1·g-1 FW for roots. At an NH4+/NO₃⁻ ratio of 9:1, GS activity in P. australis stems and leaves, as well as T. orientalis roots, was highest, reaching 39.52 μmol·h-1·g-1 FW for stems and leaves and 27.34 μmol·h-1·g-1 FW for roots. When the NH4+/NO₃⁻ ratio was 7:3, GDH activity in S. validus stems and leaves and L. salicaria roots was highest, at 66.15 μmol·h-1·g-1 FW for stems and leaves and 78.62 μmol·h-1·g-1 FW for roots. Furthermore, when theNH4+/NO₃⁻ ratio was 5:5, NiR activity in T. orientalis stems and leaves was relatively high, at 44.53 μmol·h-1·g-1 FW. At an NH4+/NO₃⁻ ratio of 7:3, NiR activity in T. orientalis roots was higher, at 36.83 μmol·h-1·g-1 FW. Lastly, when the NH4+/NO₃⁻ ratio was 7:3, GOGAT activity in P. australis stems and leaves, as well as T. orientalis roots was highest, at 37.80 μmol·h-1·g-1 FW for stems and leaves and 22.80 μmol·h-1·g-1 FW for roots.
In varying NH4+/NO3- ratios, the contents of SP in the stems and leaves of the four emergent plants, as well as the activities of NiR, GS and GOGAT, were all higher than those in the roots. Conversely, the GDH activity in the stems and leaves was lower than that in the roots for all four species. The NR activity in the stems and leaves of P. australis exceeded that in the roots, whereas in T. orientalis, NR activity was lower in the stems and leaves than in the roots. The NR activity of S. validus and L. salicaria is sometimes higher in the stems and leaves and sometimes in the roots (Figure 3).
2.3. Influencing Factors on Nitrogen Uptake Rates of Different Emergent Plants
As shown in Figure 4a, the NH4+-N uptake rate of P. australis exhibited a significantly positive correlation with NH4+/NO3- ratio, NH4+-N uptake amount, GS activity, GOGAT activity, and GDH activity (p < 0.05), while it was significantly negatively correlated with NO3--N uptake amount, SP content, NR activity, and biomass (p < 0.05). The NO3--N uptake rate demonstrated a significantly positive correlation with NO3--N uptake amount, SP content, and NR activity (p < 0.01), and it was significantly negatively correlated with NH4+-N uptake amount, NH4+/NO3- ratio, GS activity, GOGAT activity, and GDH activity (p < 0.05). Additionally, the inorganic nitrogen uptake rate was significantly positively correlated with specific root surface area (SRA) and total nitrogen (TN) content (p < 0.05), whereas it was negatively correlated with biomass and root tissue density (RTD) (p < 0.05).
As shown in Figure 4b, the NH4+-N uptake rate of T. orientalis exhibited a significantly positive correlation with the NH4+/NO3- ratio, NH4+-N uptake amount, GS activity, GOGAT activity, and GDH activity (p < 0.05). Conversely, it was significantly negatively correlated with NO3--N uptake amount, SP content, and NR activity (p < 0.01). The NO3--N uptake rate demonstrated a significant positive correlation with NO3--N uptake amount, SP content, and NR activity (p < 0.01), while it was significantly negatively correlated with NH4+-N uptake amount, NH4+/NO3- ratio, GS activity, GOGAT activity, and GDH activity (p < 0.05). Additionally, the uptake rate of inorganic nitrogen was significantly positively correlated with SRA and specific root length (SRL) (p < 0.05), whereas it was negatively correlated with RTD, biomass, and root diameter (AD) (p < 0.05).
As illustrated in Figure 4c, the NH4+-N uptake rate of S. validus showed a significantly positive correlation with the NH4+/NO3- ratio, GS activity, GOGAT activity, GDH activity, NH4+-N uptake amount, SRL, and SRA (p < 0.05). In contrast, it exhibited a significantly negative correlation with NO3--N uptake amount, SP, NR, RTD, AD (p < 0.05). Furthermore, the NO3--N uptake rate was significantly positively correlated with NO3--N uptake amount and NR activity (p < 0.05), whereas it was significantly negatively correlated with GS activity, GOGAT activity, and GDH activity (p < 0.05). Lastly, the inorganic nitrogen uptake rate demonstrated a significantly positive correlation with the NH4+/NO3- ratio, GS activity, SRA, and SRL (p < 0.05), while it was significantly negatively correlated with SP content, RTD, NR activity, NiR activity, and AD (p < 0.05).
As illustrated in Figure 4d, the NH4+-N uptake rate of L. salicaria showed a significantly positive correlation with the NH4+/NO3- ratio, NH4+-N uptake amount, GS activity, and GOGAT activity (p < 0.05), whereas it exhibited a significantly negative correlation with NO3--N uptake amount, SP content, and NR activity (p < 0.01). The NO3--N uptake rate was significantly positively correlated with the NO3--N uptake amount, SP content, and NR activity (p < 0.01), while it was significantly negatively correlated with the NH4+/NO3- ratio, NH4+-N uptake amount, GS activity, and GOGAT activity (p < 0.01). Furthermore, the inorganic nitrogen uptake rate demonstrated a significantly positive correlation with NH4+-N uptake amount and GS activity (p < 0.05), but it was significantly negatively correlated with SP content and NR activity (p < 0.05).
2.4. The Main Driving Factors of Nitrogen Uptake Rate of Emergent Plants
The redundancy analysis (RDA) analysis results (Figure 5) revealed that the interpretation rates of the first and second axes were 57.81% and 34.30%, respectively, with a cumulative interpretation rate of 92.11%. The main indicators influencing the nitrogen uptake rate of emergent plants, ranked from largest to smallest, were NO3--N uptake amount in stems and leaves, TN content in stems and leaves, NH4+-N uptake amount in stems and leaves, SRA, NH4+-N uptake amount in roots, NR activity in stems and leaves, NiR activity in roots, SP content in stems and leaves, SP content in roots, and NR activity in roots.
Among these factors, the NH4+-N uptake rate exhibited a significantly positive correlation with the TN content in stems and leaves, NH4+-N uptake amount, and SRA (p < 0.05). The NO3--N uptake rate demonstrated a significantly positive correlation with NO3--N uptake amount in stems and leaves, TN content in stems and leaves, SRA, NR activity in stems and leaves, NiR activity in roots, and SP content (p < 0.05). Furthermore, the inorganic nitrogen uptake rate was significantly positively correlated with NO3--N uptake amount in stems and leaves, TN content in stems and leaves, NH4+-N uptake amount, SRA, NR activity, NiR activity in roots, and SP content (p < 0.05).
The partial least squares path model (PLS-PM) was utilized to investigate the potential direct and indirect effects of nitrogen uptake amount, root traits, nitrogen assimilation enzymes, and photosynthetic parameters on the nitrogen uptake rate of emergent plants (Figure 6). The PLS-PM analysis results demonstrated that nitrogen uptake amount and root traits significantly positively influenced the nitrogen uptake rate directly (p < 0.001), with path coefficients of 0.786 and 0.460, respectively. Plant TN content contributed more than 0.8 to nitrogen uptake amount, while SRL and SRA accounted for over 0.8 to root traits. Furthermore, nitrogen assimilation enzymes had a significant negative direct effect on the nitrogen uptake rate (p < 0.01), with a path coefficient of -0.194. NiR, SP, NR to nitrogen assimilation enzymes were all greater than 0.8. In addition, root traits showed a significant positive direct effect on nitrogen assimilation enzymes, and nitrogen assimilation enzymes also had a significant positive direct effect on nitrogen uptake (p < 0.001). The main effects analysis of PLS-PM indicated that nitrogen uptake amount exerted the greatest total impact on the nitrogen uptake rate, followed by root traits and nitrogen assimilation enzymes (Figure 6).
3. Discussion
3.1. Effects of Different Forms of Nitrogen on Nitrogen Uptake Rate of Emergent Plants
Plant growth is optimized under a balanced supply of NH4+-N and NO3--N, and the efficiency of nitrogen acquisition is determined by the ionic availability in the habitat [18,42]. In this study, we demonstrated that both the nitrogen form and plant species significantly affected the nitrogen uptake rate of four emergent plants in the Yellow River irrigation area (p < 0.01) (Table 1). When NH4+-N was predominant, plants showed a clear preference for NH4+-N absorption. When NO3--N was predominant, plants favored the uptake of NO3--N (Figure 2). These results suggest that different nitrogen forms significantly affected the nitrogen uptake rate of emergent plants, and the nitrogen uptake rate exhibited a certain degree of environmental adaptability. These findings align with those reported by Daryanto et al. (2019) [43], thus confirming the part of our first hypothesis. This phenomenon can be explained from the following three aspects.
(1) Plants require different levels of energy for the absorption and assimilation of various nitrogen forms, and the associated metabolic pathways differ accordingly. NH4+ is directly involved in amino acid synthesis and requires less energy, thus eliminating the additional energy expenditure associated with NO3- reduction [44]. While the absorption and assimilation of NO3- demand more energy, its low toxicity reduces the metabolic burden on plants. [45,46]. Plants primarily rely on ammonium transporters for NH4+ uptake, whereas nitrate transporters are essential for NO3- uptake. Consequently, the predominant nitrogen form in the water may enhance the uptake of that specific nitrogen form by modulating the expression levels of the corresponding transporters [47].
(2) Changes in environmental factors directly or indirectly influence plant physiological activities, thereby modulating the nitrogen uptake rate of plants. For example, high temperatures significantly reduce the relative growth rate, nitrogen uptake rate, NR activity, and photosynthetic parameters of aquatic plants. [48]. Factors such as varying light intensities, nutrient concentrations, and hydraulic loads also influenced the absorption efficiency of plants. [49].
(3) To adapt to varying environmental conditions, aquatic plants modulate nitrogen uptake, assimilation, and transformation via their growth traits (e.g., root structure and photosynthetic capacity) and interactions with microorganisms. This not only meets their growth demands but also significantly decreases nitrogen concentrations in water bodies, thereby reflecting a certain degree of environmental adaptability [50,51]. Studies have shown that aquatic plants can increase their direct nitrogen uptake and induce changes in nitrogen cycling and the microenvironment by altering root morphology, growth characteristics, leaf biomass, and rhizosphere conditions, thus reducing nitrogen pollution in riparian waters [52]. The nitrogen assimilation efficiency of aquatic plants is closely linked to the root-to-shoot ratio. Aquatic plants indirectly affect nitrogen removal by microorganisms through modifying tailwater quality parameters and can also mediate nitrogen transformation by modulating bacterial community structures. [53]. In addition, as the nitrogen load in sewage increases, plant nitrogen preference shifts from NH4+-N to NO3--N. The elevated NR activity detected in downstream river plants provides evidence for their enhanced nitrate assimilation capacity and preference [18].
3.2. Effects of Different Forms of Nitrogen on Nitrogen Assimilation Enzyme Activity in Emergent Plants
Different forms of nitrogen substantially influence the activities of nitrogen assimilation-related enzymes in plants. Through these complex adaptive adjustments, plants sustain their growth and development under varying nitrogen supply conditions [12]. In this study, different forms of nitrogen significantly affected the activity of nitrogen assimilation-related enzymes in plants (Figure 3), which is largely in agreement with the findings reported by Chen et al. and further supports the other part of our first hypothesis. We found that as the NH4+/NO3- ratio in the water increased, the NR and SP content in emergent plants decreased, whereas GS activity increased. Moreover, GDH activity exhibited an initial increase followed by a subsequent decrease. This phenomenon may be explained by three potential factors:
(1) NR is a key enzyme involved in the assimilation of NO3- by plants, and its activity is influenced by nitrogen form. Under varying nitrogen supply conditions, plants from different ecological groups exhibit significantly different NR activities [55]. When NH4+-N is supplied, the NR activity in plant roots remains low. In contrast, when NO3--N is supplied, the NR activity in plant roots is significantly higher. This may be attributed to NH4+ competing with NO3- for absorption sites, thereby influencing the uptake of NO3- and the associated NR activity of plants [56]. In addition, NO3- can be assimilated in the roots and subsequently transported to the aerial parts for further assimilation. This process facilitates the rational and efficient utilization of the carbon skeleton across the entire system. Notably, NR acts as a rate-limiting enzyme, and its activity plays a critical role in regulating nitrogen metabolism and protein synthesis [57]. In this study, an increase in the NH4+/NO3- ratio was found to be associated with decreased NR activity and reduced SP content in the four emergent plants, thereby providing further support for the findings of this study.
(2) GS plays a crucial role in plant nitrogen metabolism by primarily mediating the assimilation of NH4+ [58]. Elevated NH4+ levels stimulate GS activity, leading to more efficient conversion of NH4+ into organic nitrogen compounds [24]. Studies have demonstrated that plants exhibit significant growth advantages and competitiveness under conditions with a high NH4+/NO3- ratio. This can be attributed to their efficient uptake of NH4+ and the increased activity of enzymes such as GS [4]. In addition, the genes most significantly influenced by NO3- and NH4+ treatment were those involved in glutamine metabolism, further supporting the link between changes in GS activity and nitrogen assimilation in plants [59]. In this study, an increase in the NH4+/NO3- ratio was correlated with enhanced GS activity in the four emergent plants.
(3) GDH plays a crucial role in NH4+ assimilation and glutamate synthesis in plants [15], thereby modulating the balance of nitrogen metabolism and, in turn, significantly contributing to plant growth, development, and stress adaptation [60]. As the NH4+/NO3- ratio increases, plants may mitigate the adverse effects of NH4+ by enhancing GDH activity; however, when this ratio becomes excessively high, GDH activity may decline due to impaired energy supply and disrupted metabolic balance [11,16].
3.3. Factors Influencing Nitrogen Uptake Rate of Different Emergent Plants
The nitrogen uptake rate in different plants is closely linked to root traits and the activity of nitrogen assimilation enzymes. These factors interact synergistically, thus affecting the efficiency of nitrogen uptake and utilization in plants. [61]. In this study, the NH4+-N uptake rate of the four emergent plants was significantly positively correlated with the NH4+/NO3- ratio, NH4+-N uptake amount, GS activity, and GOGAT activity (p < 0.05), whereas it was significantly negatively correlated with NO3--N uptake amount, SP content, and NR activity (p < 0.05). Moreover, the NO3--N uptake rate exhibited a significant positive correlation with NO3--N uptake amount and NR activity (p < 0.05), while showing a significant negative correlation with GS activity and GOGAT activity (p < 0.05) (Figure 4). The results demonstrated that the four emergent plants were influenced by the synergistic effects of environmental adaptation and physiological regulation during nitrogen uptake and assimilation. Specifically, a higher NH4+-N concentration in the water enhanced the activities of GS and GOGAT, promoted NH4+-N uptake amount, and thereby improved the efficiency of NH4+-N uptake and assimilation. In contrast, in NO3--N-enriched water, plants upregulated NR activity to enhance NO3--N uptake capacity, and this synergistic mechanism facilitated more efficient utilization of different nitrogen forms in the water [12,62].
The rate of NH4+-N uptake by plants was significantly and positively correlated with GS activity and GOGAT activity. This relationship can be explained from the following perspectives:(1) The GS/GOGAT cycle represents the primary pathway for NH4+ assimilation in plants, with GS catalyzing the synthesis of glutamine. Subsequently, GOGAT converts glutamine and α-ketoglutarate into glutamic acid, which serves as a critical precursor for the biosynthesis of nitrogenous compounds, including proteins and nucleic acids [13]. Elevated activities of GS and GOGAT not only enhance NH4+ uptake and assimilation but also provide additional nitrogen for plant growth, facilitating the synthesis of biological macromolecules such as proteins and nucleic acids, and thus promoting plant growth and development [63]. (2) GS is a pivotal enzyme in plant nitrogen metabolism [64], and variations in its activity can serve as a signaling cue for nitrogen uptake, thereby regulating nitrogen acquisition and utilization in plants [13]. When plant roots sense an increase in NH4+ concentration, they modulate the expression of GS-related genes, enhance GS activity, and consequently promote NH4+ uptake and assimilation via signaling pathways [65]. (3) By regulating the expression of key enzymes involved in nitrogen metabolism, such as GS and GOGAT, plants can effectively enhance their adaptability to environmental stress, thereby promoting growth and productivity [66]. Elevated activities of GS and GOGAT facilitate efficient NH4+ absorption and utilization, reduce intracellular NH4+ accumulation, and thus prevent NH4+ toxicity [45]. Furthermore, aquatic plants preferentially absorb NH4+, which enhances GS and GOGAT activities while suppressing NR and NiR activities. This mechanism not only ensures efficient utilization of nitrogen sources in water bodies but also minimizes energy consumption [67].
There are several plausible reasons for the significant positive correlation between NO3--N uptake rate and NR activity: (1) NR acts as the initial enzyme in the nitrate assimilation process and serves as the rate-limiting step [68]. Higher NR activity increases the plant's ability to reduce NO3- to NH4+, thereby enhancing NO3- uptake efficiency [69]. (2) Under NO3- induction, the expression of NR-related genes is markedly upregulated, resulting in elevated NR activity and consequently improving NO3- absorption and assimilation [70]. (3) The uptake of NO3- by plants demands a greater energy investment compared to NH4+uptake [44]. When NR activity is high, it suggests that plants prioritize energy allocation toward NO3- uptake to meet their nitrogen metabolism demands [18].
In this study, the inorganic nitrogen uptake rates of P. australis, T. orientalis and S. validus were significantly correlated with SRA and RTD. The main reasons for this correlation may be summarized as follows: (1) Most leaves of P. australis, T. orientalis and S. validus are positioned above the water layer, precluding direct nitrogen absorption. To adapt to their aquatic environment, these plants have evolved well-developed root systems, allowing them to predominantly absorb inorganic nitrogen via their roots [71]. (2) Larger SRAs increase the surface area of the root system, thereby enhancing diffusion and mass flow processes and improving the uptake rate of inorganic nitrogen [72]. (3) A larger SRA may reflect a greater density of root hairs on the root surface. These root hairs secrete organic acids that desorb adsorbed inorganic nitrogen, thus facilitating its absorption by the root system [73]. (4) Plants with lower RTDs tend to exhibit higher SRAs, which enlarge the contact area between roots and the environment, thereby promoting nitrogen diffusion and uptake [74].
In addition, the NH4+-N uptake rates of P. australis and T. orientalis were significantly positively correlated with GDH activity, which may be linked to their ammonium toxicity resistance. This implies that P. australis and T. orientalis could be better adapted to eutrophic water bodies through the GDH pathway [16]. The inorganic nitrogen uptake rates of P. australis and T. orientalis exhibited a negative correlation with biomass and RTD, suggesting that higher nitrogen uptake rates might lead to energy reallocation toward root development, consequently inhibiting aboveground growth. Alternatively, alterations in root structure could potentially affect nitrogen uptake efficiency [75].
3.4. The Main Drivers of Nitrogen Uptake Rate in Emergent Plants
In this study, plant nitrogen uptake amount was identified as the most influential factor affecting the overall nitrogen uptake rate, with plant nitrogen content being particularly significant. Root attributes, specifically SRA and SRL, were the second most important factors. Nitrogen assimilation enzymes, including NiR, SP, NR, played a subsequent role in influencing nitrogen uptake rate. However, photosynthetic parameters exhibited no significant impact on nitrogen uptake rate (Figure. 6). These findings confirm our second hypothesis. Furthermore, this study revealed the following key indicators influencing plant nitrogen uptake rates: NO3--N uptake amount in stems and leaves, TN content in stems and leaves, NH4+-N uptake amount in stems and leaves, SRA, NH4+-N uptake amount in roots, NR activity in stems and leaves, NiR activity in roots, SP content in stems and leaves, SP content in roots, and NR activity in roots (Figure 5).
Nitrogen uptake amount exerted the greatest total impact on the nitrogen uptake rate, especially the plant TN content and NO3--N uptake amount in stem and leaf, which were mainly due to the following reasons. (1) Plants with higher nitrogen uptake amount generally have more complete nitrogen metabolism pathways and more efficient nitrogen transport systems, and can more flexibly adjust the nitrogen uptake rate to adapt to changes in external nitrogen supply when faced with different concentrations of nitrogen [76]. (2) The roots of aquatic plants absorb different forms of nitrogen from water or sediment, and then transport and distribute them to various tissues and organs [77]; When the N content of stems and leaves is high, it indicates that the plant has abundant nitrogen stores, which can provide sufficient biological macromolecules such as enzymes and carrier proteins for related physiological activities, thereby maintaining a high nitrogen uptake capacity [78]. (3) Stem and leaf cells have high metabolic activity, can quickly absorb and transport nitrogen, especially stem and leaf NR activity is high, and can reduce NO3- to NH4+ and then be used by plants [79]. Moreover, the photosynthesis process of stems and leaves produces a large amount of energy and carbon backbone, which provides a material basis and dynamic support for nitrogen absorption and assimilation [80]. (4) NO3- in water has high solubility and mobility, can spread rapidly in water, and is in full contact with the stems and leaves of aquatic plants, so that it is more easily absorbed by the stems and leaves of aquatic plants [81]. Compared with NH4+, NO3- is relatively less toxic to aquatic plants, and can continuously absorb and accumulate NO3- within a certain range [82].
Root traits, especially SRA and SRL, have a greater impact on nitrogen uptake rates, which may be due to the following reasons: (1) Aquatic plants with larger SRAs and SRLs have better exposure to nitrogen in water, thereby increasing nitrogen uptake rates [71,72]. At the same time, plant roots adjust morphology and physiological functions with different forms of inorganic nitrogen, such as increasing SRA and SRL, to improve nitrogen uptake and utilization [29]. (2) Roots are the main part of nitrogen absorption in plants, and root cells are rich in nitrogen assimilation enzymes and nitrogen transporters, and adjusting root morphology can provide more substrates for root nitrogen assimilation enzymes, and by adjusting nitrogen assimilation enzyme activity, the absorbed nitrogen is converted into transportable and available organic nitrogen compounds, and nitrogen is transported to the aerial parts through redistribution strategies to meet the growth needs of stems and leaves, so it has a greater impact on the nitrogen uptake rate [83]. (3) P. australis, an emergent plant with well-developed aeration tissue, can promote microbial nitrogen conversion through root oxygen secretion, thereby increasing nitrogen uptake [84]. In this study, this was also confirmed by a significant positive correlation between SRA and nitrogen uptake rate in emergent plant (Figure 5).
Nitrogen assimilation enzymes, particularly NiR, SP, NR, play a crucial role in influencing nitrogen uptake rates. This can primarily be explained by the following factors: (1) The activity of nitrogen assimilation enzymes (e.g., NR and GS) serves as a key indicator of the metabolic efficiency of NO3- and NH4+ in plants. Under varying nitrogen sources, the activity of nitrogen assimilation enzymes in aquatic plants adjusts dynamically to adapt to environmental conditions. Specifically, under high NO3- conditions, NR activity is significantly induced; in contrast, under high NH4+ conditions, NR activity is markedly inhibited while GS expression is upregulated [85]. Moreover, soluble proteins, which include a variety of nitrogen assimilating enzymes (e.g., NR and GS), play a critical role in influencing nitrogen uptake rates by modulating the activities of key enzymes involved in nitrogen metabolism. Soluble proteins also function as a storage form of nitrogen and synergistically interact with photosynthesis [86]. These findings are consistent with the significant positive correlation observed between nitrogen assimilation enzyme activity and nitrogen uptake rates in emergent plants (Figure 5). (2) The nitrate assimilation process constitutes a critical pathway for plants to achieve sustainable growth and enhance productivity. In this process, NR and NiR function as pivotal enzymes. NR serves as the initial enzyme in the nitrate assimilation process and acts as the rate-limiting enzyme that catalyzes the reduction of NO3- to NO2-. Subsequently, NiR reduces NO2- to NH4+, thereby enabling further involvement in the synthesis of amino acids and proteins [57,68]. However, when NH4+ accumulates to high levels in plants, it may inhibit the activity of NR and NiR by modulating the cellular redox state or signal transduction pathways, thus reducing the demand for NO3- reduction and conserving energy [44]. This absorption mechanism not only supports the rapid growth of aquatic plants in high NH4+ environments but also enhances their competitive advantage. [87,88].
Since the samples in this study were collected at a single time point, they could only reflect the uptake rates of different nitrogen forms by emergent plants during a specific period and did not consider the dynamic regulation of enzyme activity throughout the plant growth cycle. Consequently, this study has certain limitations. Furthermore, the test water consisted of a 15N isotopically labeled NH4+/NO3- proportional gradient solution, which is relatively simplified compared to natural water bodies and neglects the influence of other environmental factors (e.g., light, temperature, pH, flow rate, etc.). In the future, we aim to conduct more comprehensive investigations into the nitrogen uptake rates and influencing factors of aquatic plants across different seasons using in-situ field experiments.
4. Materials and Methods
4.1. Experimental Desig
The 15N isotope labeling experiment was carried out at the Key Laboratory of Ningxia Academy of Agriculture and Forestry Sciences, Ministry of Agriculture and Rural Affairs. Considering the survival rate, ecotype, and purification efficiency, seedlings of four common emergent plants in the Yellow River Irrigation Area of Ningxia—P. australis, T. orientalis, S. validus and L. salicaria—were transplanted into an agricultural drainage ditch and grown for two months as experimental materials.
Based on previous studies [12,89] and preliminary monitoring conducted by our research group, the NH4+/NO3- ratio was observed to range from 0.41 to 3.67. A nitrogen-free Hoagland nutrient solution was used as the base solution, and five NH4+/NO3- gradients with identical total nitrogen concentrations were established for conducting 15N isotope labeling experiments (Table 2). Each emergent plant species was divided into two groups: one group was prepared using 15NH4Cl (99.12 atom%) and NaNO3 according to the five NH4+/NO3- gradients, while the other group was prepared using Na15NO3 (99.21 atom%) and NH4Cl. Each treatment was replicated three times, resulting in a total of 30 experimental beakers. Each beaker contained 1 L of test solution with a uniform total nitrogen concentration of 15 mg N L-1. Plant transplantation was initiated at 10:00 on July 2, 2023, and the experiment continued for 24 hours.
At 10:00 on July 3, the net photosynthetic rate of plant leaves was measured using a plant photosynthetic analyzer (CIRAS-3, PP Systems, USA), with three replicates per treatment and three plants measured per replicate. After the completion of the 15N isotope labeling experiment, the chlorophyll content of leaves was determined, roots were scanned for root attribute analysis, and the dry weight, N content, SP content, nitrogen assimilase activity, and 15N atomic percentage of stems and leaves, and roots samples were analyzed.
4.2. Sampling and Measurements
Following the completion of the 15N isotope labeling experiment, the plants were carefully rinsed with distilled water and subsequently separated into two components: the aboveground biomass (stems and leaves) and the belowground biomass (roots) for further analysis.
Chlorophyll a and chlorophyll b contents were determined using 95% ethanol extraction. Specifically, 0.5 g of fresh leaf samples were accurately weighed and transferred into a 10 mL centrifuge tube, followed by the addition of 10 mL of 95% ethanol. The mixture was incubated for 48 h at room temperature in the dark. Subsequently, the supernatant was carefully collected, and the absorbance values at wavelengths of 665 nm and 649 nm were measured with a multifunctional microplate reader (Thermo Varioskan LUX, Shanghai). Finally, the concentrations of chlorophyll a, chlorophyll b, and total chlorophyll were calculated based on the measured absorbance values.
Determination of Root Functional Traits: Roots were dissected following their branching order, carefully arranged on clear glass without overlapping, scanned with an Epson desktop scanner(Epson Expression 10 000 XL,Epson,Japan), and analyzed using the root analysis software WinRHIZO (Regent Instruments, Quebec, Canada). Subsequently, AD, total root length, root surface area, root volume, and number of root segments for each plant were determined.
Determination of Enzyme Activity in Plant Stems, Leaves, and Roots: Fresh samples of plant stems and leaves, and roots were carefully cleaned, finely minced, thoroughly homogenized, and accurately weighed at 0.1 g of fresh weight. The samples were subsequently ground into a fine powder under liquid nitrogen conditions. The SP content was quantified using the bicinchoninic acid (BCA) protein assay kit. The activities of NR, NiR, GS, GOGAT, and GDH were quantified using a double-antibody one-step sandwich enzyme-linked immunosorbent assay (ELISA).
Determination of Nitrogen Content in Different Forms of Plant Stems and Leaves, and Roots: Fresh samples of plant stems and leaves, and roots were dried at 65 °C for 48 h until a constant weight was achieved, then accurately weighed. Subsequently, the samples were ground into a fine powder using a ball mill (MM2, Fa. Retsch, Haan, Germany), passed through a 100-mesh sieve, and the 15N atomic percentage was analyzed using an isotope ratio mass spectrometer–elemental analyzer (IRMS-EA, Elementar, Manchester, UK).
The TN content of plant samples was determined using the Kjeldahl nitrogen determination method.
4.3. Calculations of Various Indicators
The contents of chlorophyll a, chlorophyll b and total chlorophyll can be calculated by equations (1)-(3).
where and represent the absorbance values of the chlorophyll extract at wavelengths of 665 nm and 649 nm, respectively. denotes the concentration of chlorophyll a (mg L-1), represents the concentration of chlorophyll b (mg L-1), and indicates the total chlorophyll concentration (mg L-1).
Formulas (4)-(6) for SRL, SRA, RTD are as follows:
where denotes the specific root length (m g-1); represents the specific root surface area (cm2 g-1); indicates the root tissue density (g cm-3); and , , , and correspond to the total length (m), dry weight (g), surface area (cm2), and volume (cm3) of each root segment, respectively.
The formula (7)-(8) for calculating the plant nitrogen uptake rate and the nitrogen uptake amount of each organ of plant roots, stems and leaves is as follows [90]:
where denotes the rate of plant nitrogen uptake (μg N g-1·h-1DW); , represent 15N atom percent excess in roots, stems and leaves, respectively. , (µg N g-1) indicate the nitrogen contents in roots, stems and leaves, respectively. , (g) refer to the dry weights of roots, stems and leaves, respectively. T represents the labeling duration (h). signifies the nitrogen uptake by each organ (root, stem and leaf) of the plant (μg·N), with the subscript X denoting the specific organ (root, stem and leaf). refers to the nitrogen concentration of NH4+ or NO3- in each beaker (mmol L-1), and indicates the 15N concentration (mmol L-1) of the added labeling solution.
4.4. Statistical Analysis
The effects of N form, plant species and NH4+/NO3- on plant nitrogen uptake rate were analyzed by multivariate ANOVA. The difference was tested by least square method (LSD), and the significance level was p < 0.05. The differences in nitrogen uptake rate and enzyme activities of nitrogen assimilation between different treatments were determined to be statistically significant at a level of p < 0.05 using one-way ANOVA and LSD tests. Pearson correlation analysis was employed to examine the associations among the nitrogen uptake rate, and influencing factors. The RDA was employed to examine the relationships among the rate of nitrogen uptake, and influencing factors. The PLS-PM showing the effects of nitrogen uptake amount, traits of roots n assimilation enzymes and photosynthetic parameter on nitrogen uptake rate. All the statistical analyses were carried out using R software (version 3.1.2).
5. Conclusions
The nitrogen form, plant species, NH4+/NO3- ratio, and their interactions significantly influenced the nitrogen uptake rates of four emergent plant species in the Yellow River irrigation area. Specifically, the nitrogen form exerted the strongest influence, followed by plant species. In water bodies with varying NH4+/NO3- ratios, the nitrogen uptake rates of P. australis and T. orientalis were significantly higher than those of S. validus and L. salicaria, with increases of 11.83%–114.69% and 14.07%–130.46%, respectively. The nitrogen form preference of four emergent plant species exhibited significant environmental plasticity, varying with the NH4+/NO3- ratio. However, at an NH4+/NO3- ratio of 5:5, P. australis and T. orientalis showed a preference for NO3--N absorption, whereas S. validus and L. salicaria favored NH4+-N uptake.
The nitrogen uptake rate of emergent plants was closely associated with their nitrogen assimilation enzyme activity. The NH4+-N uptake rate in the four emergent plants increased with the enhancement of GS and GOGAT activities, while the NO3--N uptake rate increased with the enhancement of NR activity. The nitrogen uptake and assimilation processes in the four plants involved a synergistic mechanism of environmental adaptation and physiological regulation, enabling them to absorb different forms of nitrogen from water more effectively. P. australis and T. orientalis exhibited enhanced adaptation to eutrophication through the GDH pathway. Root traits, particularly SRA, played a critical role in modulating the nitrogen uptake rate of the plants.
Nitrogen uptake amount exerted the greatest total impact on the nitrogen uptake rate, particularly on plant nitrogen content. The second most influential factor was root traits, specifically SRA and SRL. Nitrogen assimilation enzymes, especially NiR, SP and NR, also played a significant role in regulating nitrogen uptake. However, photosynthetic parameters showed no significant effect on the nitrogen uptake rate. Plants primarily adapt to different forms of nitrogen in water by modulating nitrogen assimilation enzyme activities and optimizing root traits.
It is evident that significant interspecific variations exist in the nitrogen uptake rates and physiological response mechanisms of emergent plants. In the remediation of nitrogen pollution in the Yellow River irrigation area, priority should be given to highly adaptable species such as P. australis and T. orientalis. Future research should focus on examining the seasonal dynamics of nitrogen uptake rates and physiological responses of aquatic plants in natural water bodies, thereby providing insights into the regulatory mechanisms underlying nitrogen uptake in these ecosystems.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Nitrogen uptake amounts in different aquatic plants. Table S2: Root traits and photosynthetic parameters in different aquatic plants.
Author Contributions
Conceptualization, Y.H.; Data curation, Y.H.; Formal analysis, Y.H.; Investigation, Y.H.; Methodology, Y.H.; Project administration, R.L.; Supervision, X.F.; Writing – original draft, Y.H.; Writing – review & editing, R.L., W.X., P.L. and X.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ningxia Key R&D Program (grant no. 2022BEG02007), the National Natural Science Foundation of China (32460324), and the Ningxia Agricultural High-quality Development and Ecological Protection Technology Innovation Demonstration Project (grant no. NGSB-2021-11).
Data Availability Statement
The data are not publicly available due to privacy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Khanna, K.; Kohli, S.K.; Sharma, N.; Kour, J.; Devi, K.; Bhardwaj, T.; Dhiman, S.; Singh, A.D.; Sharma, N.; Sharma, A.; et al. Phytomicrobiome communications: novel implications for stress resistance in plants. Front. Microbiol. 2022, 13, 912701. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, H.; Zang, S. Purification mechanism of emergent aquatic plants on polluted water: a review. J. Environ. Manage. 2025, 375, 124198. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Yang, R.; Cao, Q.; Dong, J.; Li, C.; Quan, Q.; Huang, M.; Liu, J. Differences of the microbial community structures and predicted metabolic potentials in the lake, river, and wetland sediments in dongping lake basin. Environ. Sci. Pollut. Res. 2020, 27, 19661–19677. [Google Scholar] [CrossRef]
- Chen, H.; Huang, X.; Shi, W.; Kronzucker, H.J.; Hou, L.; Yang, H.; Song, Q.; Liu, J.; Shi, J.; Yang, Q.; et al. Coordination of nitrogen uptake and assimilation favours the growth and competitiveness of moso bamboo over native tree species in high-NH4+ environments. J. Plant Physiol. 2021, 266, 153508. [Google Scholar] [CrossRef]
- Li, L.; Cheng, X.; Kong, X.; Jia, P.; Wang, X.; Zhang, L.; Zhang, X.; Zhang, Y.; Zhang, Z.; Zhang, B. Comparative transcriptomic analysis reveals the negative response mechanism of peanut root morphology and nitrate assimilation to nitrogen deficiency. Plants 2023, 12, 732. [Google Scholar] [CrossRef]
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: current status and future prospects. J. Genet. Genomics 2022, 49, 394–404. [Google Scholar] [CrossRef]
- Huangfu, S.; Zhou, F.; Zheng, X.; Zhang, X.; Hu, L. Removal of ammonia nitrogen from black and odorous water by macrophytes based on laboratory microcosm experiments. RSC Adv. 2023, 13, 3173–3180. [Google Scholar] [CrossRef]
- Sahay, S.; Robledo-Arratia, L.; Glowacka, K.; Gupta, M. Root NRT, NiR, AMT, GS, GOGAT and GDH expression levels reveal NO and ABA mediated drought tolerance in brassica juncea l. Sci. Rep. 2021, 11, 7992. [Google Scholar] [CrossRef]
- Jiao-jiao, G.; Hong-wei, Z.; Yan, J.; Bo-wen, H.; Zhuo-qian, W.; Zhao-jun, Q.; Feng-li, Y. Effect of salt stress on nitrogen assimilation of functional leaves and root system of rice in cold region. Journal of Northeast Agricultural University (English Edition). 2020, 27, 9–16. [Google Scholar]
- Lv, X.; Wang, X.; Pan, J.; Deng, W.; Li, Y. Role of nitrate reductase and nitrite reductase in NaCl tolerance in eelgrass (zostera marina l.). Ecological Chemistry and Engineering S 2022, 29, 111–125. [Google Scholar] [CrossRef]
- Du, C.; Lin, J.; Dong, L.; Liu, C.; Tang, D.; Yan, L.; Chen, M.; Liu, S.; Liu, X. Overexpression of an NADP(h)-dependent glutamate dehydrogenase gene, TrGDH, from trichurus improves nitrogen assimilation, growth status and grain weight per plant in rice. Breed. Sci. 2019, 69, 429–438. [Google Scholar] [CrossRef]
- Chen, M.; Zhu, K.; Xie, J.; Liu, J.; Qiao, Z.; Tan, P.; Peng, F. Ammonium-nitrate mixtures dominated by NH4+-N promote the growth of pecan (carya illinoinensis) through enhanced N uptake and assimilation. Front. Plant Sci. 2023, 14, 1186818. [Google Scholar] [CrossRef] [PubMed]
- Singh, S. Role of glutamine synthetase, glutamine and NH4+ in the regulation of NO2- uptake in the cyanobiont nostoc ANTH. J. Plant Physiol. 1993, 142, 403–406. [Google Scholar] [CrossRef]
- Song, J.; Yang, J.; Jeong, B.R. Growth, quality, and nitrogen assimilation in response to high ammonium or nitrate supply in cabbage (brassica campestris l.) And lettuce (lactuca sativa l.). Agronomy 2021, 11, 2556. [Google Scholar] [CrossRef]
- Robinson, S.A.; Slade, A.P.; Fox, G.G.; Phillips, R.; Ratcliffe, R.G.; Stewart, G.R. The role of glutamate dehydrogenase in plant nitrogen metabolism. Plant Physiol. 1991, 95, 509–516. [Google Scholar] [CrossRef]
- Xian, L.; Zhang, Y.; Cao, Y.; Wan, T.; Gong, Y.; Dai, C.; Ochieng, W.A.; Nasimiyu, A.T.; Li, W.; Liu, F. Glutamate dehydrogenase plays an important role in ammonium detoxification by submerged macrophytes. Sci. Total Environ. 2020, 722, 137859. [Google Scholar] [CrossRef]
- Knight, R.L.; Gosselink, J.G. Global treatment wetlands. Wetlands 2001, 21, 648–649. [Google Scholar] [CrossRef]
- Guo, H.R.; Wu, Y.; Hu, C.C.; Liu, X.Y. Elevated nitrate preference over ammonium in aquatic plants by nitrogen loadings in a city river. Journal of Geophysical Research: Biogeosciences 2022, 127, e2021JG006614. [Google Scholar] [CrossRef]
- Hong, J.; Qin, X.; Ma, X.; Xu, X.; Wang, X. Seasonal shifting in the absorption pattern of alpine species for NO3- and NH4+ on the tibetan plateau. Biol. Fertil. Soils 2019, 55, 801–811. [Google Scholar] [CrossRef]
- Zhuang, W.; Wang, M.; Xiao, Y.; Zhou, X.; Wu, N. Differential uptake of nitrogen forms by two herbs in the gurbantunggut desert, central asia. Plant Biol. 2022, 24, 758–765. [Google Scholar] [CrossRef]
- Czaban, W.; Rasmussen, J. Co-occurrence of organic and inorganic n influences asparagine uptake and amino acid profiles in white clover. J. Plant Nutr. Soil Sci. 2021, 184, 162–172. [Google Scholar] [CrossRef]
- Wang, P.; Wang, Z.; Sun, X.; Mu, X.; Chen, H.; Chen, F.; Lixing, Y.; Mi, G. Interaction effect of nitrogen form and planting density on plant growth and nutrient uptake in maize seedlings. J. Integr. Agric. 2019, 18, 1120–1129. [Google Scholar] [CrossRef]
- Du, Y.; Niu, S.; Jiang, H.; Luo, B.; Wang, Q.; Han, W.; Liu, Y.; Chang, J.; Ge, Y. Comparing the effects of plant diversity on the nitrogen removal and stability in floating and sand-based constructed wetlands under ammonium/nitrate ratio disturbance. Environ. Sci. Pollut. Res. 2021, 28, 69354–69366. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lv, J.; Xie, J.; Gan, Y.; Coulter, J.A.; Yu, J.; Li, J.; Wang, J.; Zhang, X. Nitrogen source affects the composition of metabolites in pepper (capsicum annuum l.) And regulates the synthesis of capsaicinoids through the GOGAT–GS pathway. Foods 2020, 9, 150. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Hou, B.; Song, B.; Zhang, X.; Zhuang, W. Preferences of annual and ephemeral plants for inorganic n versus organic n at different growth stages in a temperate desert ecosystem. J. Arid Environ. 2024, 225, 105264. [Google Scholar] [CrossRef]
- Zhuang, W.; Li, J.; Yu, F.; Dong, Z.; Guo, H. Seasonal nitrogen uptake strategies in a temperate desert ecosystem depends on n form and plant species. Plant Biol. 2020, 22, 386–393. [Google Scholar] [CrossRef]
- Mao, J.; Wang, J.; Liao, J.; Xu, X.; Tian, D.; Zhang, R.; Peng, J.; Niu, S. Plant nitrogen uptake preference and drivers in natural ecosystems at the global scale. New Phytol. 2025. [Google Scholar] [CrossRef]
- Yang, Z.; Li, Y.; Wang, Y.; Cheng, J.; Li, F.Y. Preferences for different nitrogen forms in three dominant plants in a semi-arid grassland under different grazing intensities. Agriculture, Ecosystems & Environment 2022, 333, 107959. [Google Scholar] [CrossRef]
- Meier, M.; Liu, Y.; Lay-Pruitt, K.S.; Takahashi, H.; von Wirén, N. Auxin-mediated root branching is determined by the form of available nitrogen. Nat. Plants 2020, 6, 1136–1145. [Google Scholar] [CrossRef]
- Tang, D.; Liu, M.; Zhang, Q.; Ma, L.; Shi, Y.; Ruan, J. Preferential assimilation of NH4+ over NO3- in tea plant associated with genes involved in nitrogen transportation, utilization and catechins biosynthesis. Plant Sci. 2020, 291, 110369. [Google Scholar] [CrossRef]
- Xiao, H.; Sheng, M.; Wang, L.; Guo, C.; Zhang, S. Effects of short-term n addition on fine root morphological features and nutrient stoichiometric characteristics of zanthoxylum bungeanum and medicago sativa seedlings in southwest china karst area. J. Soil Sci. Plant Nutr. 2022, 22, 1805–1817. [Google Scholar] [CrossRef]
- Sun, J.; Jin, L.; Li, R.; Meng, X.; Jin, N.; Wang, S.; Xu, Z.; Liu, Z.; Lyu, J.; Yu, J. Effects of different forms and proportions of nitrogen on the growth, photosynthetic characteristics, and carbon and nitrogen metabolism in tomato. Plants 2023, 12, 4175. [Google Scholar] [CrossRef] [PubMed]
- Aryal, K.; Maraseni, T.; Apan, A. How much do we know about trade-offs in ecosystem services? A systematic review of empirical research observations. Sci. Total Environ. 2022, 806, 151229. [Google Scholar] [CrossRef]
- Zhang, T.; Lei, Q.; Liang, X.; Lindsey, S.; Luo, J.; Pei, W.; Du, X.; Wu, S.; An, M.; Qiu, W.; et al. Optimization of the n footprint model and analysis of nitrogen pollution in irrigation areas: a case study of ningxia hui autonomous region, china. J. Environ. Manage. 2023, 340, 118002. [Google Scholar] [CrossRef]
- Kumwimba, M.N.; Zhu, B.; Muyembe, D.K. Assessing the influence of different plant species in drainage ditches on mitigation of non-point source pollutants (n, p, and sediments) in the purple sichuan basin. Environ. Monit. Assess. 2017, 189, 267. [Google Scholar] [CrossRef]
- Pei, W.; Yan, T.; Lei, Q.; Zhang, T.; Fan, B.; Du, X.; Luo, J.; Lindsey, S.; Liu, H. Spatio-temporal variation of net anthropogenic nitrogen inputs (NANI) from 1991 to 2019 and its impacts analysis from parameters in northwest china. J. Environ. Manage. 2022, 321, 115996. [Google Scholar] [CrossRef]
- Yu, W.; Zhang, J.; Liu, L.; Li, Y.; Li, X. A source-sink landscape approach to mitigation of agricultural non-point source pollution: validation and application. Environ. Pollut. 2022, 314, 120287. [Google Scholar] [CrossRef]
- Barko, J.W.; Michael Smart, R. The nutritional ecology of cyperus esculentus, an emergent aquatic plant, grown on different sediments. Aquat. Bot. 1979, 6, 13–28. [Google Scholar] [CrossRef]
- Qin, J.; Sun, Y.; Qiu, X.; Liu, H.; Zhang, E.; Mao, X. Distribution pattern simulation of multiple emergent plants in river riparian zones. River Res. Appl. 2021, 37, 1180–1190. [Google Scholar] [CrossRef]
- Lünsmann, V.; Kappelmeyer, U.; Taubert, A.; Nijenhuis, I.; von Bergen, M.; Heipieper, H.J.; Müller, J.A.; Jehmlich, N. Aerobic toluene degraders in the rhizosphere of a constructed wetland model show diurnal polyhydroxyalkanoate metabolism. Appl. Environ. Microbiol. 2016, 82, 4126–4132. [Google Scholar] [CrossRef]
- Rehman, F.; Pervez, A.; Khattak, B.N.; Ahmad, R. Constructed wetlands: perspectives of the oxygen released in the rhizosphere of macrophytes. CLEAN – Soil, Air, Water 2017, 45. [Google Scholar] [CrossRef]
- Liu, M.; Xu, X.; Wanek, W.; Sun, J.; Bardgett, R.D.; Tian, Y.; Cui, X.; Jiang, L.; Ma, Z.; Kuzyakov, Y.; et al. Nitrogen availability in soil controls uptake of different nitrogen forms by plants. New Phytol. 2025, 245, 1450–1467. [Google Scholar] [CrossRef] [PubMed]
- Daryanto, S.; Wang, L.; Gilhooly, W.P.; Jacinthe, P. Nitrogen preference across generations under changing ammonium nitrate ratios. J. Plant Ecol. 2019, 12, 235–244. [Google Scholar] [CrossRef]
- Krouk, G.; Crawford, N.M.; Coruzzi, G.M.; Tsay, Y. Nitrate signaling: adaptation to fluctuating environments. Curr. Opin. Plant Biol. 2010, 13, 265–272. [Google Scholar] [CrossRef]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: a critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, Z.; Sun, H.; Liu, X.; Xie, J.; Qiu, Y.; Chai, T.; Chu, C.; Hu, B. Nitrate confers rice adaptation to high ammonium by suppressing its uptake but promoting its assimilation. Mol. Plant. 2023, 16, 1871–1874. [Google Scholar] [CrossRef]
- Qian, J.; Jin, W.; Hu, J.; Wang, P.; Wang, C.; Lu, B.; Li, K.; He, X.; Tang, S. Stable isotope analyses of nitrogen source and preference for ammonium versus nitrate of riparian plants during the plant growing season in taihu lake basin. Sci. Total Environ. 2021, 763, 143029. [Google Scholar] [CrossRef]
- Jiang, H.; Huang, B.; Qian, Z.; Xu, Y.; Liao, X.; Song, P.; Li, X. Effects of increased carbon supply on the growth, nitrogen metabolism and photosynthesis of vallisneria natans grown at different temperatures. Wetlands 2020, 40, 1459–1467. [Google Scholar] [CrossRef]
- Wang, K.; Hu, Q.; Wei, Y.; Yin, H.; Sun, C.; Liu, G. Uptake kinetics of NH4+, NO3- and H2PO4- by Typha orientalis, Acorus calamus L., Lythrum salicaria L., Sagittaria trifolia L. and Alisma plantago-aquatica Linn. Sustainability 2021, 13, 434. [Google Scholar] [CrossRef]
- Guo, C.; Zhang, Q.; Hu, Y.; Zhao, Q.; Li, Q.; Wu, J. Influence of sediment, plants, and microorganisms on nitrogen removal in farmland drainage ditches. Agronomy 2023, 13, 2211. [Google Scholar] [CrossRef]
- Manolaki, P.; Mouridsen, M.B.; Nielsen, E.; Olesen, A.; Jensen, S.M.; Lauridsen, T.L.; Baattrup-Pedersen, A.; Sorrell, B.K.; Riis, T. A comparison of nutrient uptake efficiency and growth rate between different macrophyte growth forms. J. Environ. Manage. 2020, 274, 111181. [Google Scholar] [CrossRef]
- Franklin, H.M.; Robinson, B.H.; Dickinson, N.M. Plants for nitrogen management in riparian zones: a proposed trait-based framework to select effective species. Ecol. Manag. Restor. 2019, 20, 202–213. [Google Scholar] [CrossRef]
- Wu, H.; Hao, B.; You, Y.; Zou, C.; Cai, X.; Li, J.; Qin, H. Aquatic macrophytes mitigate the conflict between nitrogen removal and nitrous oxide emissions during tailwater treatments. J. Environ. Manage. 2024, 370, 122671. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xiang, Y.; Yao, Z.; Zhang, Q.; Li, H.; Cheng, M. Stability of c:n:p stoichiometry in the plant–soil continuum along age classes in natural pinus tabuliformis carr. Forests of the eastern loess plateau, china. Forests 2023, 14, 44. [Google Scholar] [CrossRef]
- Koyama, L.A.; Terai, M.; Tokuchi, N. Nitrate reductase activities in plants from different ecological and taxonomic groups grown in japan. Ecol. Res. 2020, 35, 708–712. [Google Scholar] [CrossRef]
- CHEN, H.; HUANG, L. Effect of nitrogen fertilizer application rate on nitrate reductase activity in maize. Appl. Ecol. Environ. Res. 2020, 18, 2879–2894. [Google Scholar] [CrossRef]
- Ali, A. Nitrate assimilation pathway in higher plants: critical role in nitrogen signalling and utilization. Plant Sci. Today 2020, 7, 182–192. [Google Scholar] [CrossRef]
- Du, J.; Shen, T.; Xiong, Q.; Zhu, C.; Peng, X.; He, X.; Fu, J.; Ouyang, L.; Bian, J.; Hu, L.; et al. Combined proteomics, metabolomics and physiological analyses of rice growth and grain yield with heavy nitrogen application before and after drought. BMC Plant Biol. 2020, 20, 556. [Google Scholar] [CrossRef]
- Zhou, T.; Wu, P.; Yue, C.; Huang, J.; Zhang, Z.; Hua, Y. Transcriptomic dissection of allotetraploid rapeseed (brassica napus l.) In responses to nitrate and ammonium regimes and functional analysis ofBnaA2.gln1;4in arabidopsis. Plant Cell Physiol. 2022, 63, 755–769. [Google Scholar] [CrossRef]
- Vega-Mas, I.; Rossi, M.T.; Gupta, K.J.; González-Murua, C.; Ratcliffe, R.G.; Estavillo, J.M.; González-Moro, M.B. Tomato roots exhibit in vivo glutamate dehydrogenase aminating capacity in response to excess ammonium supply. J. Plant Physiol. 2019, 239, 83–91. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, B.; Chen, Z.; Xu, K. Grafting enhances the photosynthesis and nitrogen absorption of tomato plants under low-nitrogen stress. J. Plant Growth Regul. 2022, 41, 1714–1725. [Google Scholar] [CrossRef]
- Ma, K.; Zhou, Y.; Ma, Y.; Zhang, T. Increased rice yield by improving the stay-green traits and related physiological metabolism under long-term warming in cool regions. Int. J. Plant Prod. 2024, 18, 175–186. [Google Scholar] [CrossRef]
- Harrison, J.; Pou De Crescenzo, M.; Sen; Olivier; Hirel, B. Does lowering glutamine synthetase activity in nodules modify nitrogen metabolism and growth of lotus japonicus? Plant Physiol. 2003, 133, 253–262. [CrossRef] [PubMed]
- Li, H.; Hu, B.; Chu, C. Nitrogen use efficiency in crops: lessons from arabidopsis and rice. J. Exp. Bot. 2017, 68, 2477–2488. [Google Scholar] [CrossRef]
- Cai, H.; Zhou, Y.; Xiao, J.; Li, X.; Zhang, Q.; Lian, X. Overexpressed glutamine synthetase gene modifies nitrogen metabolism and abiotic stress responses in rice. Plant Cell Rep. 2009, 28, 527–537. [Google Scholar] [CrossRef]
- Hasan, A.K.; Carrasco-G, F.E.; Carolina Lizana, X.; Calderini, D.F. Low seed rate in square planting arrangement has neutral or positive effect on grain yield and improves grain nitrogen and phosphorus uptake in wheat. Field Crops Res. 2022, 288, 108699. [Google Scholar] [CrossRef]
- Wang, H.; Tang, X.; Chen, J.; Shang, S.; Zhu, M.; Liang, S.; Zang, Y. Comparative studies on the response of zostera marina leaves and roots to ammonium stress and effects on nitrogen metabolism. Aquat. Toxicol. 2021, 240, 105965. [Google Scholar] [CrossRef]
- Lartigue, J.; Sherman, T.D. Response of enteromorpha sp. (Chlorophyceae) to a nitrate pulse: nitrate uptake, inorganic nitrogen storage and nitrate reductase activity. Mar. Ecol. Prog. Ser. 2005, 292, 147–157. [Google Scholar] [CrossRef]
- Qi, D.; Wu, Q. Physiological characteristics, lint yield, and nitrogen use efficiency of cotton under different nitrogen application rates and waterlogging durations. J. Soil Sci. Plant Nutr. 2023, 23, 5594–5607. [Google Scholar] [CrossRef]
- Wang, M.; Hasegawa, T.; Beier, M.; Hayashi, M.; Ohmori, Y.; Yano, K.; Teramoto, S.; Kamiya, T.; Fujiwara, T. Growth and nitrate reductase activity are impaired in RiceOsnlp4 mutants supplied with nitrate. Plant Cell Physiol. 2021, 62, 1156–1167. [Google Scholar] [CrossRef]
- Fang, Y,; Xie, P.; Tan, L.; Sun, Y.; Zhu, S.; Qiao, F. A review of the influence of habitat on emergent macrophyte growth and its feedback mechanism. Chinese Journal of Ecology 2021, 40(8): 2610-2619. [CrossRef]
- Wang, L.; Liao, D.; Rengel, Z.; Shen, J. Soil–plant–microbe interactions in the rhizosphere: incremental amplification induced by localized fertilization. Front. Agric. Sci. Eng. 2024, 12, 57–68. [Google Scholar] [CrossRef]
- Jia, Z.; Giehl, R.F.H.; Hartmann, A.; Estevez, J.M.; Bennett, M.J.; von Wirén, N. A spatially concerted epidermal auxin signaling framework steers the root hair foraging response under low nitrogen. Curr. Biol. 2023, 33, 3926–3941. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Tanaka-Oda, A.; Masumoto, T.; Akatsuki, M.; Makita, N. Different relationships of fine root traits with root ammonium and nitrate uptake rates in conifer forests. J. For. Res. 2023, 28, 25–32. [Google Scholar] [CrossRef]
- Lin, Q.; Gu, B.; Hong, J. Tracking uptake of submerged macrophytes (ceratophyllum demersum)—derived nitrogen by cattail (typha angustifolia) using nitrogen stable isotope enrichments. Ecol. Eng. 2017, 99, 114–118. [Google Scholar] [CrossRef]
- Choi, S.J.; Lee, Z.; Jeong, E.; Kim, S.; Seo, J.S.; Um, T.; Shim, J.S. Signaling pathways underlying nitrogen transport and metabolism in plants. BMB Rep. 2023, 56, 56–64. [Google Scholar] [CrossRef]
- Cai, Z.; Li, Q.; Bai, H.; Zhu, C.; Tang, G.; Zhou, H.; Huang, J.; Song, X.; Wang, J. Interactive effects of aquatic nitrogen and plant biomass on nitrous oxide emission from constructed wetlands. Environ. Res. 2022, 213, 113716. [Google Scholar] [CrossRef]
- OSONE, Y.; TATENO, M. Nitrogen absorption by roots as a cause of interspecific variations in leaf nitrogen concentration and photosynthetic capacity. Funct. Ecol. 2005, 19, 460–470. [Google Scholar] [CrossRef]
- Zhao, Y.; Xie, Z.; Hu, B.; Li, Y.; Teng, A.; Zhong, F. The effects of polypropylene microplastics on the removal of nitrogen and phosphorus from water by acorus calamus, iris tectorum and functional microorganisms. Chemosphere 2024, 364, 143153. [Google Scholar] [CrossRef]
- Xiong, Q.; Zhong, L.; Shen, T.; Cao, C.; He, H.; Chen, X. ITRAQ-based quantitative proteomic and physiological analysis of the response to n deficiency and the compensation effect in rice. BMC Genomics 2019, 20, 681. [Google Scholar] [CrossRef]
- Wu, H.; Ding, J. Abiotic and biotic determinants of plant diversity in aquatic communities invaded by water hyacinth [eichhorniacrassipes (mart.) Solms]. Front. Plant Sci. 2020, 11, 555774. [Google Scholar] [CrossRef]
- Hu, S.; He, R.; He, X.; Zeng, J.; Zhao, D. Niche-specific restructuring of bacterial communities associated with submerged macrophyte under ammonium stress. Appl. Environ. Microbiol. 2023, 89, e0071723. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Dijkstra, F.A.; Han, X.; Jiang, Y. Root nitrogen reallocation: what makes it matter? Trends Plant Sci. 2024, 29, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Zhao, R.; Cao, Q.; Quan, Q.; Sun, R.; Liu, J. Effects of emergent aquatic plants on nitrogen transformation processes and related microorganisms in a constructed wetland in northern china. Plant Soil 2019, 443, 473–492. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, T.; Song, S.; Li, C. Effects of nitrogenous nutrition on growth and nitrogen assimilation enzymes of dinoflagellate akashiwo sanguinea. Harmful Algae 2015, 50, 99–106. [Google Scholar] [CrossRef]
- Liu, G.; Du, Q.; Li, J. Interactive effects of nitrate-ammonium ratios and temperatures on growth, photosynthesis, and nitrogen metabolism of tomato seedlings. Sci. Hortic. 2017, 214, 41–50. [Google Scholar] [CrossRef]
- Guo, X.; Sheng, Y.; Yang, S.; Han, L.; Gao, Y.; Zhang, K.; Cheng, J.; Zhang, H.; Song, Z.; Su, Y. Isolation and characterization of a high-affinity ammonium transporter ApAMT1;1 in alligatorweed. Plant Growth Regul. 2019, 89, 321–330. [Google Scholar] [CrossRef]
- Zhou, Q.; Gao, J.; Zhang, R.; Zhang, R. Ammonia stress on nitrogen metabolism in tolerant aquatic plant— myriophyllum aquaticum. Ecotoxicol. Environ. Saf. 2017, 143, 102–110. [Google Scholar] [CrossRef]
- Jia, X.; Huangfu, C.; Hui, D. Nitrogen uptake by two plants in response to plant competition as regulated by neighbor density. Front. Plant Sci. 2020, 11, 584370. [Google Scholar] [CrossRef]
- Liu, Q.; Xu, M.; Yuan, Y.; Wang, H. Interspecific competition for inorganic nitrogen between canopy trees and underlayer-planted young trees in subtropical pine plantations. For. Ecol. Manage. 2021, 494, 119331. [Google Scholar] [CrossRef]
Figure 1.
The process model of nitrogen uptake and assimilation by plants in aquatic ecosystems. Arrows represent the key nitrogen assimilation processes.
Figure 1.
The process model of nitrogen uptake and assimilation by plants in aquatic ecosystems. Arrows represent the key nitrogen assimilation processes.
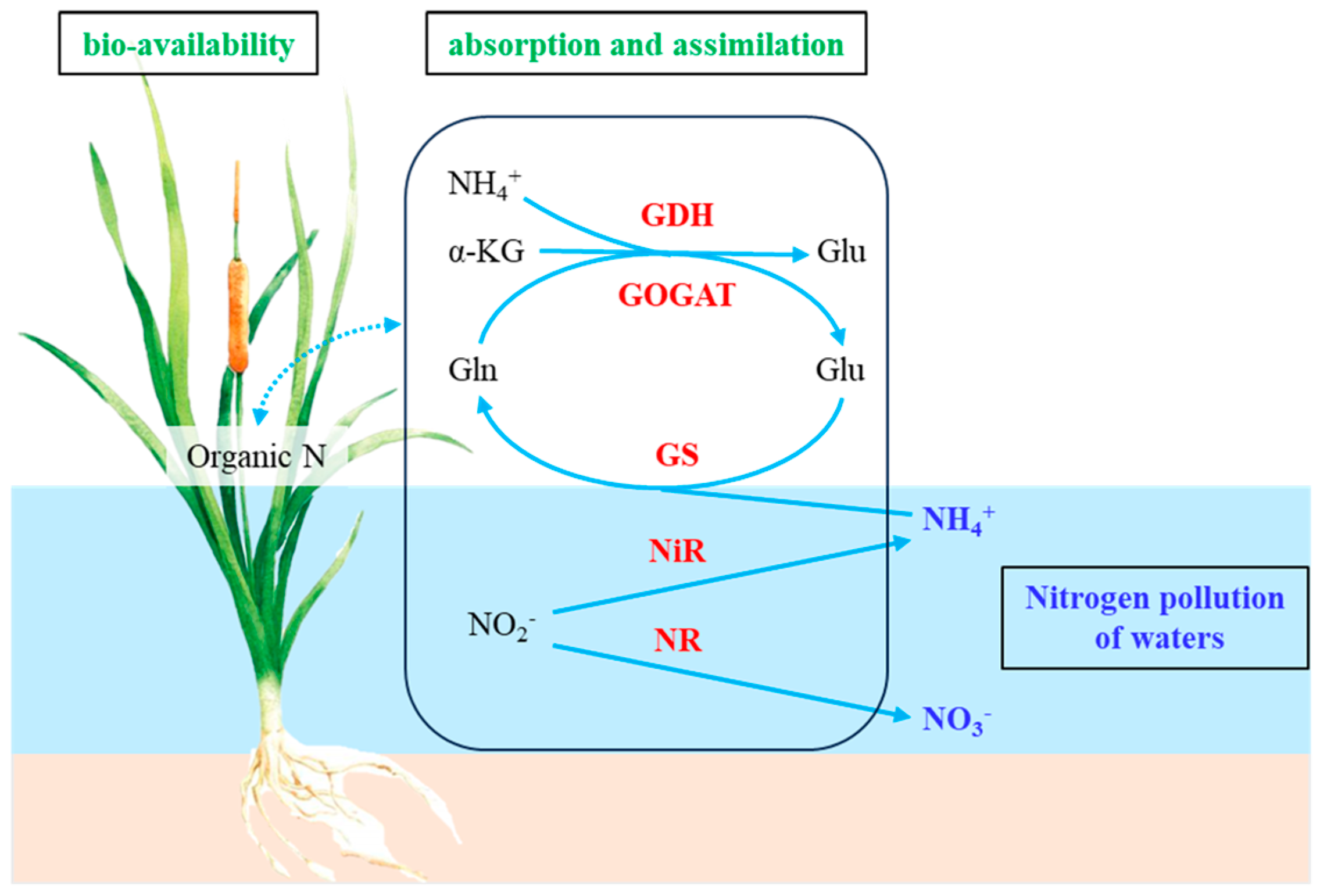
Figure 2.
Uptake rates and ratios of NH4+-N and NO3--N by different plants in water with different NH4+/NO3- ratios. PH, P. australis; TY, T. orientalis; SC, S. validus; LY, L. salicaria. Different lowercase letters indicate significant differences in the uptake rates/ratios of the same nitrogen form among different plants (p<0.05). Different uppercase letters indicate significant differences in the uptake rates of the same plant among different NH4+/NO3- ratios (p<0.05).
Figure 2.
Uptake rates and ratios of NH4+-N and NO3--N by different plants in water with different NH4+/NO3- ratios. PH, P. australis; TY, T. orientalis; SC, S. validus; LY, L. salicaria. Different lowercase letters indicate significant differences in the uptake rates/ratios of the same nitrogen form among different plants (p<0.05). Different uppercase letters indicate significant differences in the uptake rates of the same plant among different NH4+/NO3- ratios (p<0.05).
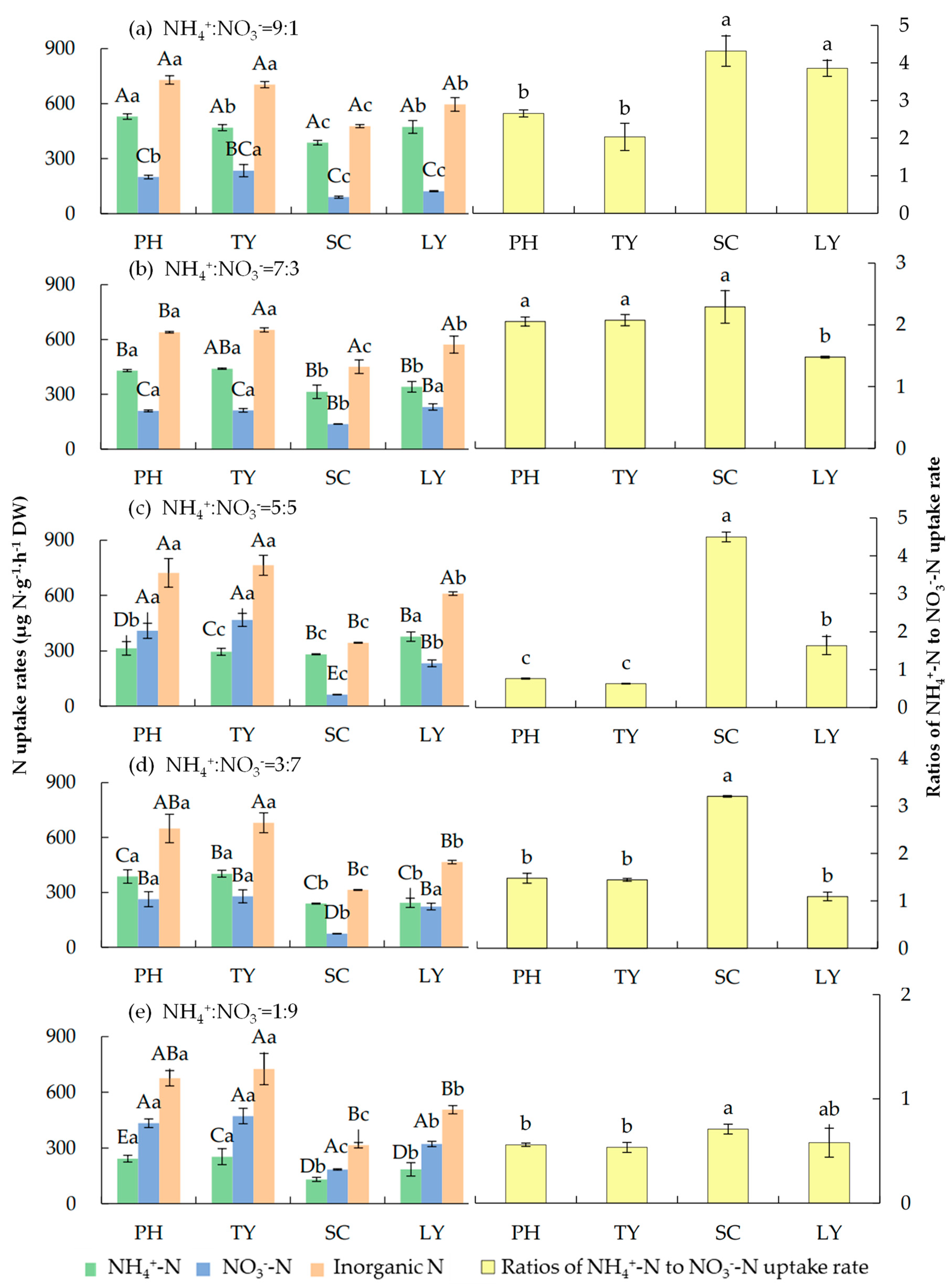
Figure 3.
Nitrogen assimilation enzyme activities in stems, leaves and roots of different plants in water with different NH4+/NO3- ratios. (a) SP content, (b) NR activity, (c) NiR activity, (d) GS activity, (e) GOGAT activity, (f) GDH activity. PH, P. australis; TY, T. orientalis; SC, S. validus; LY, L. salicaria. Different lowercase letters indicate significant differences in nitrogen assimilation enzyme activities among different plants (p<0.05).
Figure 3.
Nitrogen assimilation enzyme activities in stems, leaves and roots of different plants in water with different NH4+/NO3- ratios. (a) SP content, (b) NR activity, (c) NiR activity, (d) GS activity, (e) GOGAT activity, (f) GDH activity. PH, P. australis; TY, T. orientalis; SC, S. validus; LY, L. salicaria. Different lowercase letters indicate significant differences in nitrogen assimilation enzyme activities among different plants (p<0.05).
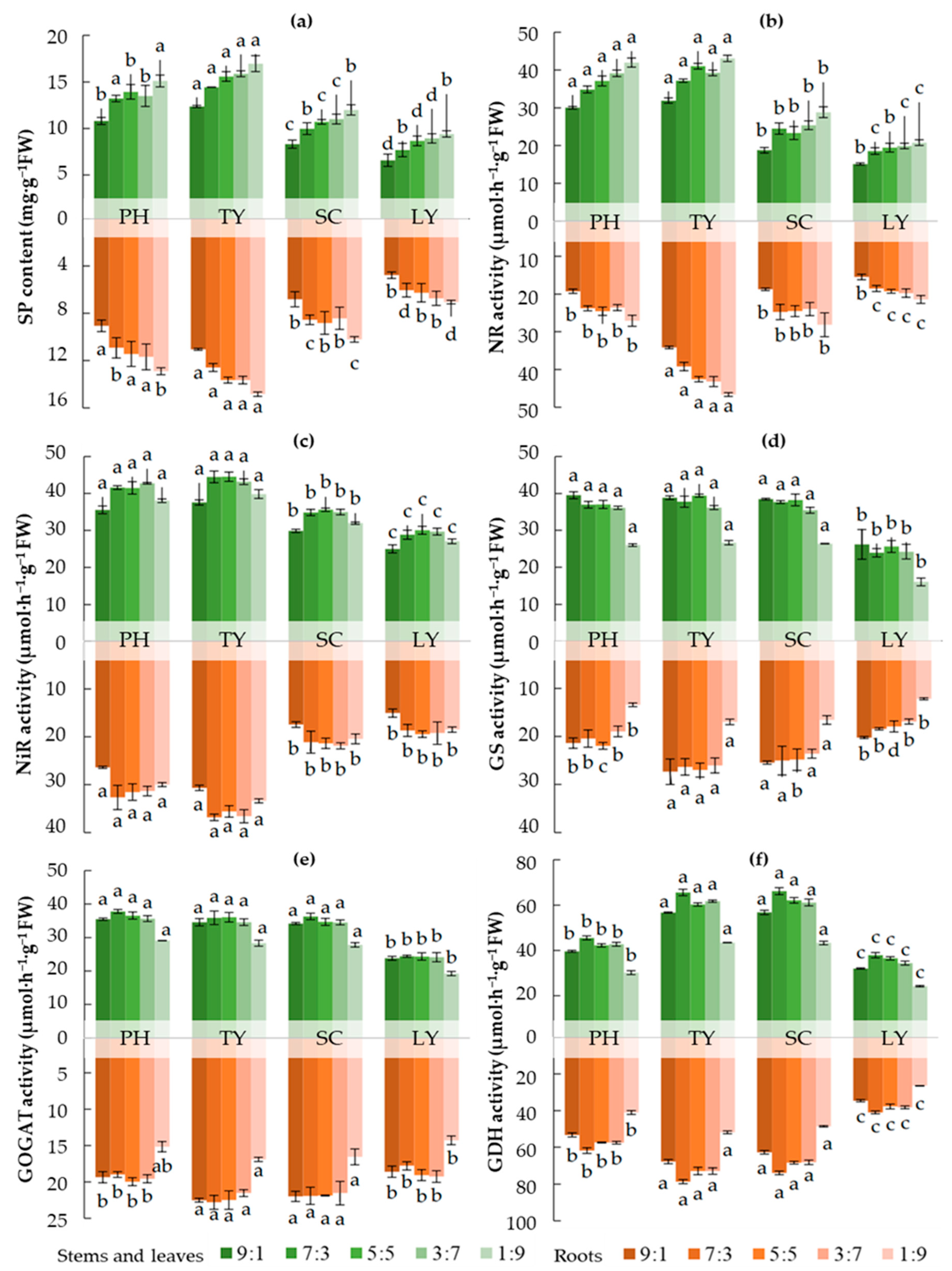
Figure 4.
Figure 4. Correlations of factors affecting nitrogen uptake rate of emergent plants. (a) PH, P. australis; (b) TY, T. orientalis; (c) SC, S. validus; (d) LY, L. salicaria. AMU, the uptake rate of NH4+-N; NTU, the uptake rate of NO3--N; INU, the uptake rate of inorganic nitrogen; RAN, NH4+/NO3- ratio; AMA, the uptake amount of NH4+-N; NTA, the uptake amount of NO3--N; TN, the total nitrogen content of plant; TM, the plant biomass; CHA, the content of chlorophyll a; CHB, the content of chlorophyll b; CHR, the ratio of Chlorophyll a to Chlorophyll b; CHT, the total chlorophyll content; Pn, the net photosynthetic rate; SP, the content of soluble protein; NR, the activity of nitrate reductase; NiR, the activity of nitrite reductase; GS, the activity of glutamine synthetase; GOGAT, the activity of glutamate synthase; GDH, the activity of glutamate dehydrogenase; AD, the diameter of fine roots; SRL, the specific root length of fine roots; SRA, the specific root surface area of fine roots; RTD, the tissue density of fine roots.
Figure 4.
Figure 4. Correlations of factors affecting nitrogen uptake rate of emergent plants. (a) PH, P. australis; (b) TY, T. orientalis; (c) SC, S. validus; (d) LY, L. salicaria. AMU, the uptake rate of NH4+-N; NTU, the uptake rate of NO3--N; INU, the uptake rate of inorganic nitrogen; RAN, NH4+/NO3- ratio; AMA, the uptake amount of NH4+-N; NTA, the uptake amount of NO3--N; TN, the total nitrogen content of plant; TM, the plant biomass; CHA, the content of chlorophyll a; CHB, the content of chlorophyll b; CHR, the ratio of Chlorophyll a to Chlorophyll b; CHT, the total chlorophyll content; Pn, the net photosynthetic rate; SP, the content of soluble protein; NR, the activity of nitrate reductase; NiR, the activity of nitrite reductase; GS, the activity of glutamine synthetase; GOGAT, the activity of glutamate synthase; GDH, the activity of glutamate dehydrogenase; AD, the diameter of fine roots; SRL, the specific root length of fine roots; SRA, the specific root surface area of fine roots; RTD, the tissue density of fine roots.
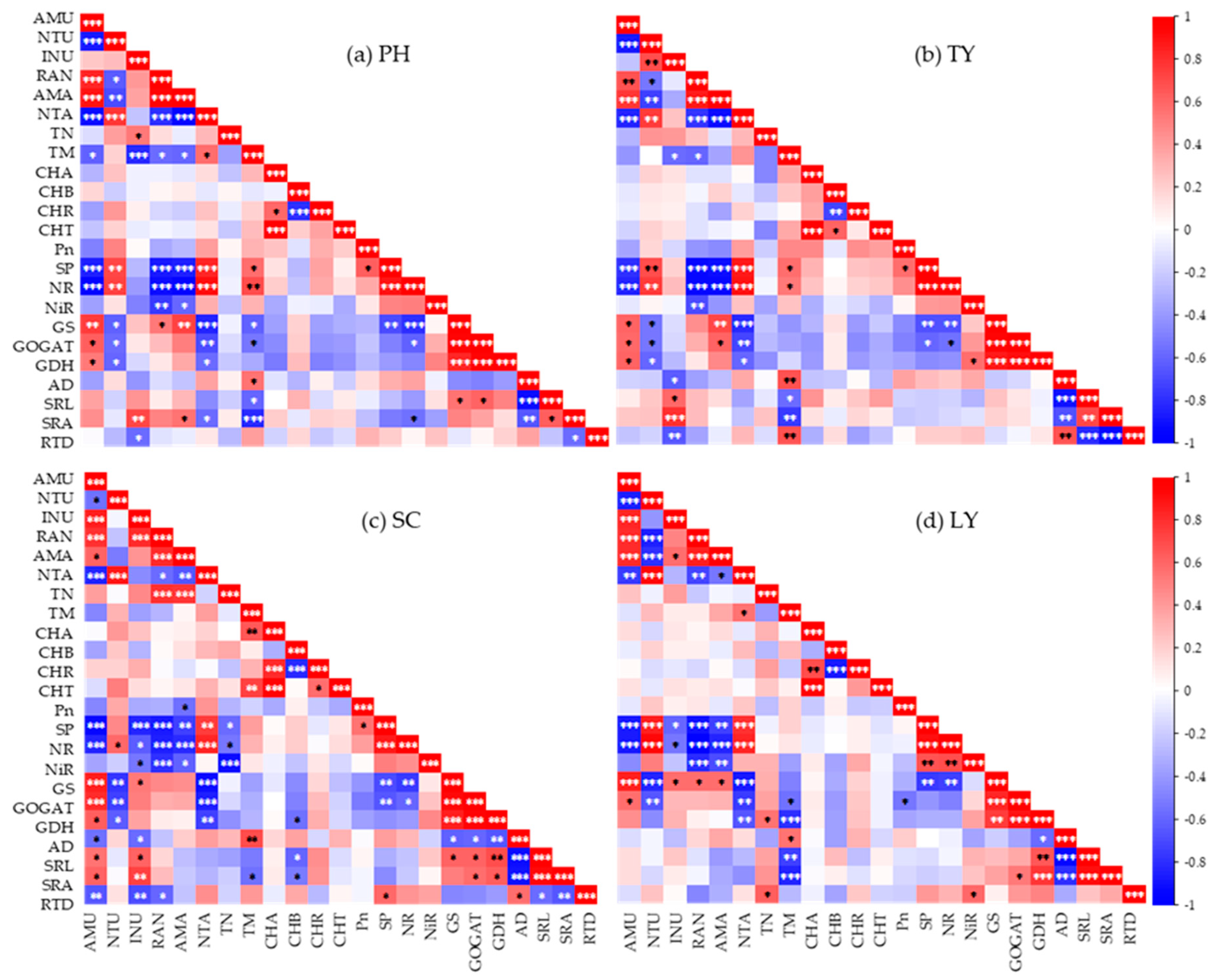
Figure 5.
Figure 5. RDA of factors affecting nitrogen uptake rate of emergent plants. AMA-L, the uptake amount of NH4+-N in stems and leaves; AMA-R, the uptake amount of NH4+-N in roots; NTA-L, the uptake amount of NO3--N in stems and leaves; NTA-R, the uptake amount of NO3--N in roots; TN-L, the total nitrogen content in stems and leaves; TN-R, the total nitrogen content in roots; TM-L, the biomass in stems and leaves; TM-R, the biomass in roots; SP-L, the content of soluble protein in stems and leaves; SP-R, the content of soluble protein in roots; NR-L, the activity of nitrate reductase in stems and leaves; NR-R, the activity of nitrate reductase in roots; NiR-L, the activity of nitrite reductase in stems and leaves; NiR-R, the activity of nitrite reductase in roots; GS-L, the activity of glutamine synthetase in stems and leaves; GS-R, the activity of glutamine synthetase in roots; GOGAT-L, the activity of glutamate synthase in stems and leaves; GOGAT-R, the activity of glutamate synthase in roots; GDH-L, the activity of glutamate dehydrogenase in stems and leaves; GDH-R, the activity of glutamate dehydrogenase in roots.
Figure 5.
Figure 5. RDA of factors affecting nitrogen uptake rate of emergent plants. AMA-L, the uptake amount of NH4+-N in stems and leaves; AMA-R, the uptake amount of NH4+-N in roots; NTA-L, the uptake amount of NO3--N in stems and leaves; NTA-R, the uptake amount of NO3--N in roots; TN-L, the total nitrogen content in stems and leaves; TN-R, the total nitrogen content in roots; TM-L, the biomass in stems and leaves; TM-R, the biomass in roots; SP-L, the content of soluble protein in stems and leaves; SP-R, the content of soluble protein in roots; NR-L, the activity of nitrate reductase in stems and leaves; NR-R, the activity of nitrate reductase in roots; NiR-L, the activity of nitrite reductase in stems and leaves; NiR-R, the activity of nitrite reductase in roots; GS-L, the activity of glutamine synthetase in stems and leaves; GS-R, the activity of glutamine synthetase in roots; GOGAT-L, the activity of glutamate synthase in stems and leaves; GOGAT-R, the activity of glutamate synthase in roots; GDH-L, the activity of glutamate dehydrogenase in stems and leaves; GDH-R, the activity of glutamate dehydrogenase in roots.
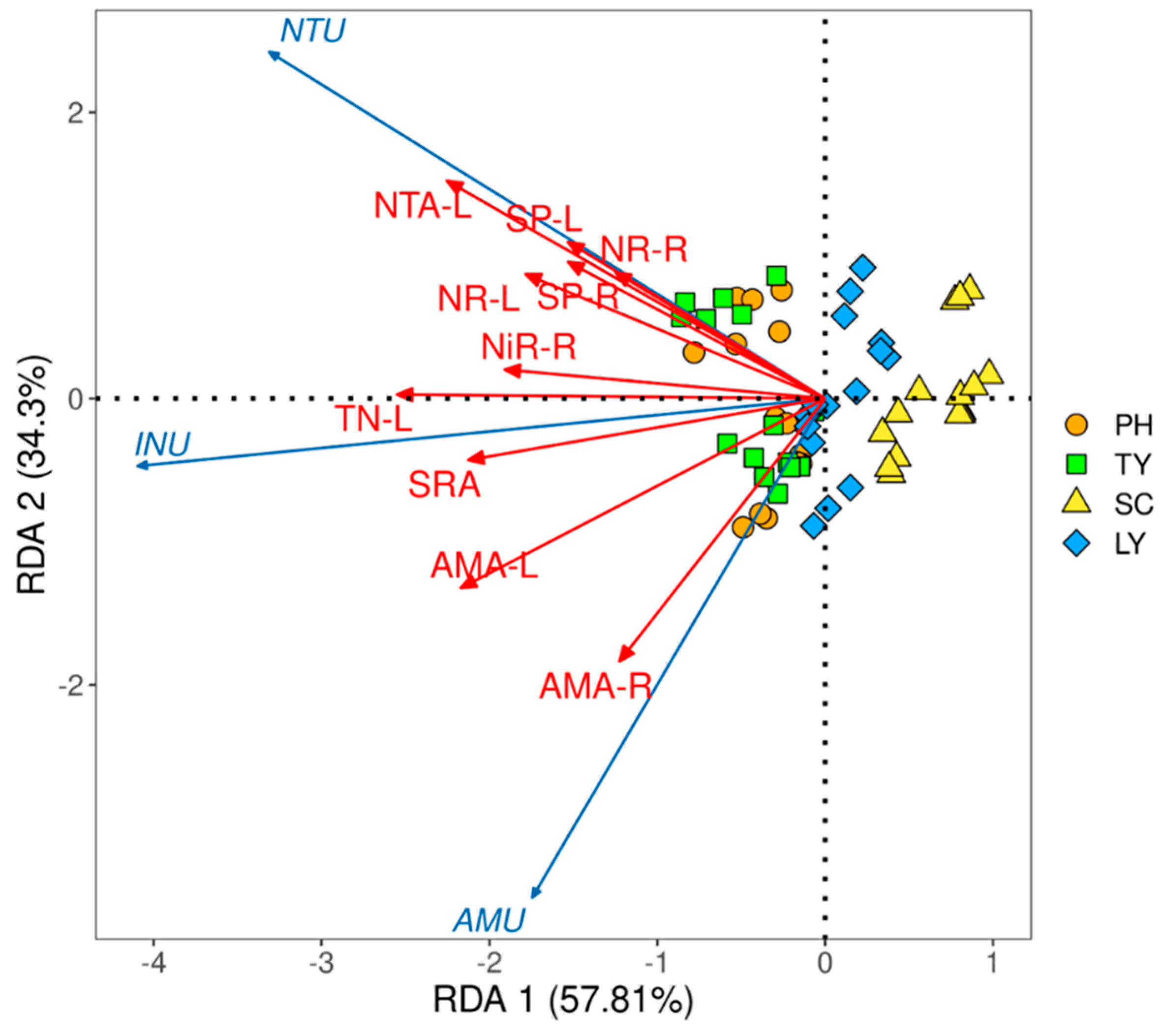
Figure 6.
PLS-PM showing the effects of nitrogen uptake amounts, root traits, nitrogen assimilation enzymes, and photosynthetic parameters on nitrogen uptake rates. The blue and red lines indicate positive and negative effects, respectively. The solid and dashed arrows indicate significant and insignificant of the path coefficients, respectively. The numbers adjacent to each arrow denote the path coefficients (significance codes: ***p< 0.001, **p< 0.01, *p< 0.05). The number next to each indicator represents the contribution of each predictor variable to the latent variable. R2 values display the proportion of variance in nitrogen uptake rate explained by each factor.
Figure 6.
PLS-PM showing the effects of nitrogen uptake amounts, root traits, nitrogen assimilation enzymes, and photosynthetic parameters on nitrogen uptake rates. The blue and red lines indicate positive and negative effects, respectively. The solid and dashed arrows indicate significant and insignificant of the path coefficients, respectively. The numbers adjacent to each arrow denote the path coefficients (significance codes: ***p< 0.001, **p< 0.01, *p< 0.05). The number next to each indicator represents the contribution of each predictor variable to the latent variable. R2 values display the proportion of variance in nitrogen uptake rate explained by each factor.
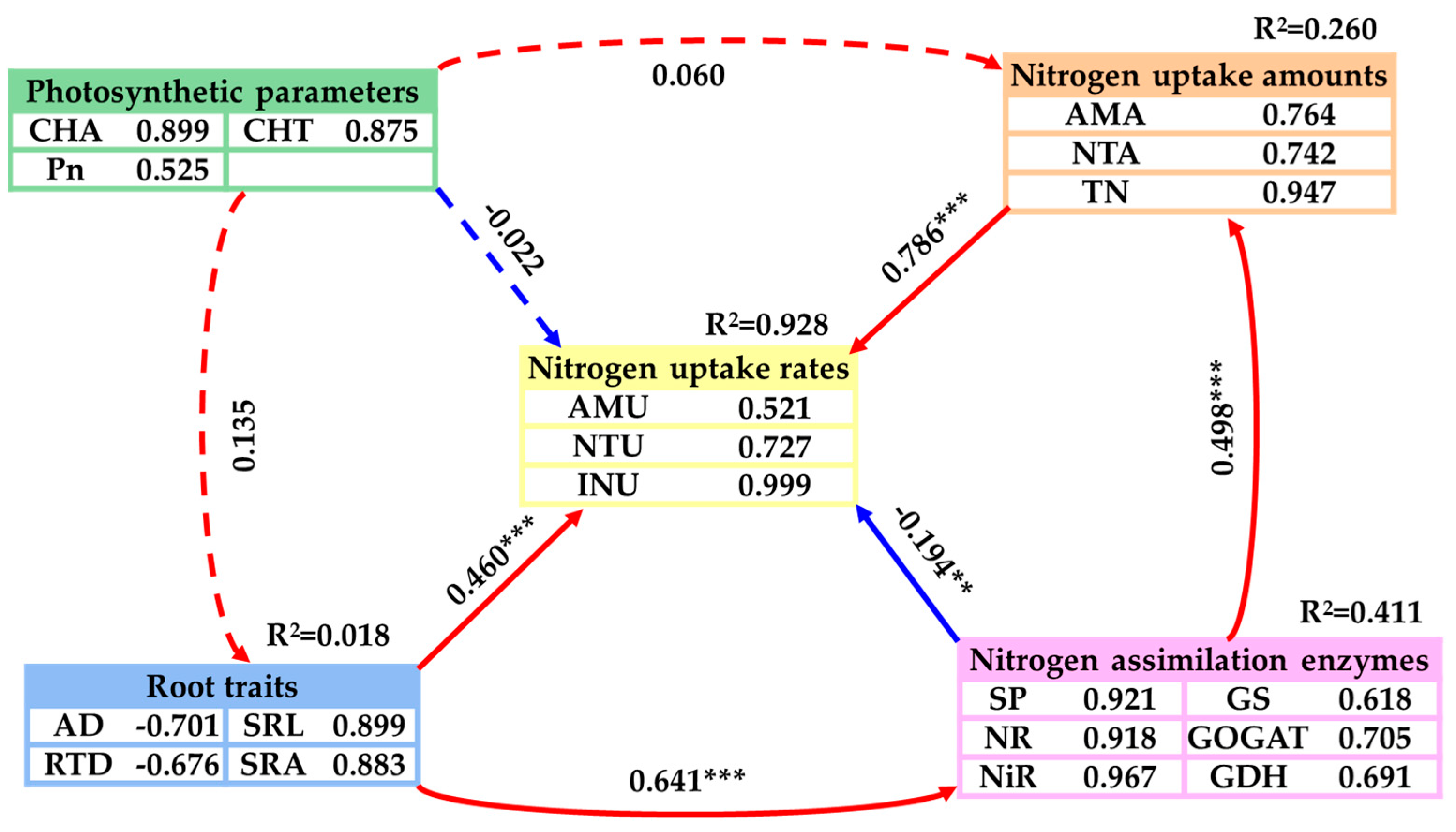

Table 1.
Multi-way analysis of variance (ANOVA) of the effects of species, NH4+/NO3- ratio, and nitrogen form on plant nitrogen uptake rate.
Table 1.
Multi-way analysis of variance (ANOVA) of the effects of species, NH4+/NO3- ratio, and nitrogen form on plant nitrogen uptake rate.
| Factors | Mean square | Df | F value | P value |
|---|---|---|---|---|
| Species | 167945.969 | 3 | 305.340 | <0.001 |
| NH4+/NO3- | 9714.229 | 4 | 17.661 | <0.001 |
| Nitrogen form | 263812.486 | 1 | 479.633 | <0.001 |
| Species × NH4+/NO3- | 3482.750 | 12 | 6.332 | <0.001 |
| Species × Nitrogen form | 19500.487 | 3 | 35.453 | <0.001 |
| NH4+/NO3-×Nitrogen form | 174690.090 | 4 | 317.601 | <0.001 |
| Species×NH4+/NO3-×Nitrogen form | 13835.820 | 12 | 25.155 | <0.001 |
| Error | 550.030 | 80 |
* Bold P values indicate significance.
Table 2.
Experimental design for 15N isotope labeling.
| Number | NH4+/NO3- | NH4Cl (mg L-1) | NaNO3 (mg L-1) |
|---|---|---|---|
| 1 | 9:1 | 13.5 | 1.5 |
| 2 | 7:3 | 10.5 | 4.5 |
| 3 | 5:5 | 7.5 | 7.5 |
| 4 | 3:7 | 4.5 | 10.5 |
| 5 | 1:9 | 1.5 | 13.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



