Submitted:
18 April 2025
Posted:
18 April 2025
You are already at the latest version
Abstract
Betalains are natural bioactive pigments characterized by their nitrogen-containing structure, water solubility, and non-toxic properties, occurring naturally in various plant species across different families. Among these plants, Celosia argentea, a member of the Amaranthaceae family, has been identified as a particularly rich source of betalains, synthesizing and accumulating significant quantities of both red-purple betacyanins and yellow-orange betaxanthins. The value of betalains extends far beyond their role as brilliant natural colorants; these bioactive substances possess an impressive array of health-promoting properties that have attracted significant scientific interest, with research demonstrating that betalains exhibit potent antioxidant and anti-inflammatory activities, alongside notable antimicrobial properties, while studies have revealed their anticancer, antidiabetic, and antilipidemic effects, as well as their ability to provide hepatoprotective and neuroprotective benefits. This diverse profile of biological activities has positioned betalains as valuable ingredients across multiple industries, with applications spanning food and beverages, where they serve as natural colorants and functional ingredients; textiles, where they provide sustainable dyeing alternatives; and the cosmetic and pharmaceutical sectors, where their therapeutic properties can be harnessed for various health-promoting products. This review presents a comprehensive examination of the current research status regarding betalain production in C. argentea, exploring the biosynthetic pathways responsible for betalain formation, analyzing their diverse biological properties, discussing their wide-ranging applications across different industries, and offering perspectives on future research directions in this promising field.
Keywords:
betalain
; biological property
; Celosia argentea
; natural pigment
; ornamental plant
; plant secondary metabolites
1. Introduction
Celosia belongs to the Amaranthaceae family, which comprises approximately 60 species. This genus is native to subtropical and temperate zones of Africa, South America, and Southeast Asia. The name “Celosia” derives from the Greek word “kelos,” meaning “burned” or “burning,” referring to the vibrant colors of its inflorescences (yellow, red, and orange). Among the various species, Celosia argentea var. cristata (L.) Kuntze (also known as C. cristata L.) and C. argentea var. plumosa (Burvenich) Voss (C. plumosa Burvenich) are widely recognized ornamental plants cultivated globally. C. argentea var. cristata is commonly known as cockscomb, while C. argentea var. plumosa is referred to as feathered amaranth [1,2,3]. According to Miguel et al. [4], C. argentea is considered the wild form, whereas C. cristata and C. plumosa are regarded as cultivar types.
Beyond their ornamental value, Celosia species serve multiple purposes. The seedlings, young leaves, and inflorescences are consumed as vegetables in Asia, Africa, and South America. Additionally, they play a significant role in traditional medicine, where they are used as disinfectants and remedies for eye and liver ailments, as well as treatments for dysentery, dysuria, blood disorders, gynecological conditions, hypertension, and sarcoptidosis, among other health issues [5,6,7,8]. C. argentea contains a diverse array of bioactive compounds, including saponins [8,9], phenols [6,10,11], flavonoids [12], bicyclic peptides [13], and betalains [1,14,15]. These compounds contribute to various biological properties, such as antioxidant [16,17], antimicrobial [12,18], anti-inflammatory [12], antidiabetic [19], antimetastatic [20], and anticancer activities [6,11]. Further benefits include immunomodulatory and hepatoprotective effects [4,9,21].
Among the various bioactive compounds found in C. argentea, betalains have garnered significant scientific interest. This review aims to comprehensively describe current research on betalain production in C. argentea, including its biosynthesis pathway, biological properties, and applications. The review also discusses future prospects for betalain production from this valuable plant resource.
2. Celosia argentea: Taxonomy, Morphology, and Ethnobotanical Applications
Celosia argentea, commonly known as plumed cockscomb or silver cock’s comb, is an annual plant belonging to the Amaranthaceae family. Native to subtropical and temperate zones of Africa, South America, and Southeast Asia, the genus Celosia comprises approximately 60 species worldwide [1]. This plant is known by various regional names, including Qingxiang in China [7], Mawal in India [22], and soi kai or ngon kai in Thailand. Among the diverse species within this genus, two primary varieties are widely cultivated globally: C. cristata (characterized by cylindrical pink or rose flower heads) and C. plumosa (distinguished by feathery plume-like flower heads) (Figure 1).
Morphologically, C. argentea is classified as an herbaceous plant due to its lack of woody tissue and the presence of a hairy covering. It is a short-lived, predominantly annual plant with stems reaching approximately 15–100 cm in height. The plant features simple leaves arranged in opposite pairs, with shapes varying from lance-like to linear. These leaves exhibit pointed tips, tapered bases, and wavy edges, and are relatively thin with light green coloration. The midrib of the leaves ranges in color from creamy white to dark green or reddish. The inflorescence of C. argentea consists of compact flower clusters emerging from leaf axils and branch tips. The flower stalks typically measure 10–20 cm in length, with upright-standing flowers. These flowers are characterized by hairy bracts that overlap densely in multiple layers. They are bisexual and cylindraceous or trochiform, positioned at the apex of stems and branches. The flowers display a vibrant palette of colors, including red, orange, pink, and yellow, with some clusters exhibiting multiple colors simultaneously. Owing to their striking coloration, these plants are widely cultivated as ornamental specimens, cut flowers, and are frequently used in dried flower arrangements [7,23,24].
Beyond their ornamental value, C. argentea plants serve significant nutritional and medicinal purposes. The seedlings, young leaves, and inflorescences are consumed as vegetables across various regions, including Asia, Africa, and South America. From an ethnomedicinal perspective, different parts of the plant have been traditionally utilized to address various health conditions. The dried ripe seeds, for instance, have been employed in the treatment of hepatitis, hypertension, and sarcoptidosis, as well as to enhance vision [7,25]. Similarly, dried leaves and inflorescences have served as disinfectants and remedies for ocular and hepatic ailments, and have been used to treat dysentery, dysuria, blood disorders, and gynecological conditions [5,6,7,8]. In addition, the radix extract of this plant has been used to treat stomachache [26].
3. Bioactive Compounds in C. argentea
C. argentea contains a diverse array of bioactive compounds isolated from various parts of the plant. The major constituents include saponins, polyphenols, fatty acids, peptides, amino acids, betalains, and minerals, as summarized in Table 1. Each of these compounds contributes to the plant’s medicinal properties.
Saponins in C. argentea exist as oleanane-type triterpenoids, with various forms identified, including celosin A to J, celosin I and II, and cristatain. These compounds demonstrate significant biological properties such as anti-inflammatory, antitumor, and hepatoprotective effects [7].
Polyphenols represent another important class of bioactive compounds with similar biological properties to saponins. Research has shown that polyphenol content varies across Celosia species. For example, C. laxa contains significantly higher levels of polyphenols and flavonoids than red and green variants of C. argentea [17]. The primary polyphenols in C. laxa are rosmarinic acid and phenol glycoside, while rutin and epigallocatechin predominate in the green variant, and epigallocatechin is the main constituent in the red variant of C. argentea [7].
Japanese researchers first discovered cyclic peptides in C. argentea [31]. The most notable peptides include moroidin, celogentins, and celogenamide A, which have demonstrated anti-mitotic effects. Beyond these cyclic peptides, various amino acids are present in this plant. Lin et al. [37] reported that approximately 42.85% of total amino acids in C. argentea are essential, with glutamic and aspartic acid being the predominant forms. Regarding fatty acid composition, C. argentea contains primarily unsaturated fatty acids (approximately 79.28%), with oleinic and linoleic acids being the most abundant [37].
The mineral content of C. argentea varies depending on the specific variety. For instance, C. argentea var. plumosa contains higher levels of Fe, Al, Mn, Cu, K, Ni, Ti, and Se compared to C. argentea var. cristata [40]. According to Lin et al. [37], the content of Fe, Zn, Mn, and Cu is 197, 160, 56, and 30 mg/g, respectively. Additional bioactive compounds found in C. argentea include lutein, β-carotene, stigmasterol, and ascorbic acid [38,39].
Recent studies have highlighted that C. argentea inflorescences contain significant amounts of natural pigments called betalains. These compounds confer numerous biological and pharmaceutical properties, including anti-inflammatory, anticancer, antimicrobial, antioxidant, and antidiabetic effects, as well as immunological activity, cytoprotective, and hepatoprotective benefits [15,41,42]. Schliemann et al. [1] found that C. argentea contained 0.157 g/g fresh weight (FW) of total betalains in yellow inflorescence and 0.293 g/g FW in orange-red inflorescence. More recently, Mueangnak et al. [15] reported that inflorescences of 3-month-old field-cultivated C. argentea contained approximately 2.95 mg/g dry weight (DW) of total betalains.
4. Betalains in C. argentea: Biosynthesis Pathway, Biological Properties, and Applications
Betalains are nitrogen-containing, water-soluble, non-toxic major plant secondary metabolites derived from the metabolism of amino acid L-tyrosine. They comprise two major groups: red-purple betacyanins, with a maximum absorbance of 530 nm, and yellow-orange betaxanthins, with a maximum absorbance of 480 nm. Both compounds are based on betalamic acid [4-(2-oxoethylindene)-1,2,3,4-tetrahydropyridine-2,6-dicarboxylic acid], which forms through condensation with either cyclo-3,4-dihydroxyphenylalanine (cyclo-DOPA) derivatives or various amino acids/amines, respectively (Figure 2) [43,44,45]. The composition and concentration of betacyanins and betaxanthins vary across plant sources, resulting in diverse colorations in the plant organs where they accumulate [46,47,48].
Over the past decade, scientists have isolated and characterized more than a hundred betalains, organizing them into different structural categories [43,50]. Within the betacyanin group, compounds such as betanin, gomphrenin, amaranthin, and bougainvillein have been identified, while the betaxanthin group includes amino acid conjugates and amine-derived conjugates [49,50]. Compared to anthocyanins (another class of plant pigments), betalains demonstrate superior water solubility, greater dyeing capacity, and enhanced stability across a pH range of 3 to 7 [43,51], making them exceptionally versatile for various applications.
The distinctive properties of betalains make them valuable candidates for use as natural colorants in foods and beverages, dyeing agents in textiles and cosmetics, and film materials for coating metal surfaces. Beyond their colorant applications, betalains exhibit remarkable biological activities, including anti-inflammatory, antioxidant, anticancer, and antimicrobial properties, highlighting their significant potential for pharmaceutical and medical applications [45]. Recent research has begun exploring the molecular mechanisms behind these health-promoting effects, further expanding the potential therapeutic uses of these natural compounds in treating various disorders and diseases.
4.1. Biosynthesis Pathway of Betalains
The biosynthesis pathway of betalains in C. argentea has not yet been fully elucidated. Most studies have been carried out in Beta vulgaris, Portulaca grandiflora, Mirabilis jalapa, Hylocereus undatus, H. monacanthus, and Amaranthus tricolor [44,45,52,53]. However, based on comprehensive literature reviews, betalains are synthesized from the aromatic amino acid tyrosine, which is a product of the shikimic acid pathway [54]. These pigments are produced in the cytosol and endoplasmic reticulum before being transported and accumulated in vacuoles, particularly in the epidermal and subepidermal tissues [55,56].
As illustrated in Figure 3, the biosynthesis begins with the conversion of tyrosine to 3,4-dihydroxy-L-phenylalanine (L-DOPA). Initially, this conversion was reported to be catalyzed by tyrosinase in the presence of molecular oxygen [47,50]; however, more recent studies have demonstrated that cytochrome P450 enzymes belonging to the CYP76AD family are responsible for this tyrosine hydroxylase activity [57]. Following this conversion, the aromatic ring of L-DOPA undergoes oxidation and cleavage in the presence of the enzyme 4,5-DOPA-estradiol-dioxygenase (4,5-DODA), resulting in the formation of 4,5-seco-DOPA [58]. The resulting 4,5-seco-DOPA then spontaneously converts through a cyclization reaction to form betalamic acid [59], which serves as a key intermediate for the formation of both betacyanins and betaxanthins. Betaxanthins are formed through a relatively simple process involving the condensation of betalamic acid with amino acids or other amines, which occurs without enzymatic catalysis [43].
In contrast, the formation of betacyanins follows a more complex pathway. First, L-DOPA is converted into cyclo-DOPA through an oxidation and cyclization reaction catalyzed by CYP76AD enzymes. Cyclo-DOPA is subsequently glycosylated by cyclo-DOPA-5-O-glucosyltransferase to yield cyclo-DOPA-glucoside. The condensation of this glycosylated compound with betalamic acid results in the formation of betanin, a member of the betacyanin group, which is characterized by a hydroxy group (-OH) at the C6 position and a glycosyl residue linked to -OH at the C5 position [60,61]. The formation of other betacyanins involves additional structural modifications catalyzed by various glucosyltransferase enzymes. For example, gomphrenin features a glucosyl residue linked to the OH at the C6 position, while amaranthine contains a glucuronyl glucosyl residue at the C6 position. Bougainvillein is distinct in having glucosyl residues at both the C5 and C6 positions of cyclo-DOPA [4,62].
4.2. Biological Properties of Betalains
Betalains exhibit a wide spectrum of biological activities as demonstrated in both in vitro and cellular assays, including antioxidant, anti-inflammatory, antimicrobial, anticancer, antidiabetic, antilipidemic, and hepatoprotective effects. B. vulgaris (beetroot) and Opuntia ficus-indica (prickly pear) serve as the primary sources for betalains in biological property investigations, with betanin and indicaxanthin being the most extensively studied compounds among this pigment family [45]. The mechanisms through which betalains exert these diverse biological effects are property-specific and comprehensively summarized in Table 2.
4.3. Applications of Betalains
Betalains exhibit diverse biological properties, making them valuable across multiple industries, including food and beverages, packaging, textiles, cosmetics, and pharmaceuticals, as well as other aspects. The Food and Drug Administration (FDA) has classified betalains as generally recognized as safe (GRAS) under the color additive petition, permitting their use as natural products in foods, beverages, cosmetics, and pharmaceuticals (Simon et al., 2017; Sadowska-Bartosz and Bartoz, 2021; FDA U.S. Food and Drug Administration, 2023). Among betalains, betanin derived from beetroot (E162) is the most widely utilized compound. While commercial betalains are predominantly sourced from beetroot, limited information exists regarding betalains from C. argentea. Table 3 summarizes the applications of betalains derived from various plant sources across different industries.
5. Production of Betalains from C. argentea
Betalains can be extracted from both wild and cultivated Celosia argentea, with the plant’s inflorescences serving as particularly rich sources of these compounds. However, obtaining betalains from natural plants presents several significant challenges. The process is notably labor-intensive, requires extensive land area for cultivation, and remains highly vulnerable to seasonal, climatic, and geographical variations. Furthermore, the susceptibility of these plants to pathogens significantly reduces betalain yield. A recent study by Mueangnak et al. [15] highlighted these limitations, reporting that betalain content from a 3-month-old field-cultivated plant reached only 2.95 mg/g dry weight (DW), with productivity at a mere 0.03 mg/g·day. These constraints have prompted researchers to develop alternative production techniques, with plant cell and tissue cultures emerging as particularly promising platforms for efficiently producing these valuable bioactive compounds.
Compared to other plant sources, research on betalain production from Celosia argentea remains relatively limited. Warhade and Badere [138] conducted pioneering work in this field, successfully investigating betalain production from callus cultures of C. argentea var. cristata. Their research documented significant betalain content, calculated by measuring amaranthin, betanin, betalamic acid, and betaxanthin levels, ranging from 9.62 to 29.90 mg/g fresh weight (FW). Building on this foundation, in 2018, the same researchers expanded their investigations to explore the effects of various elicitors on betalain production in cell suspension cultures derived from the same plant variety. Their findings revealed that after fungal elicitation, cultures achieved a total betalain content of approximately 1.44 mg/g FW [139].
More recent advancements have shown promising results. Sang A Roon et al. [14] established a betalain-producing cell line from C. argentea var. plumosa and achieved a maximum betalain concentration of 2.16 mg/g DW. This was accomplished by cultivating the cells in MS medium supplemented with 43.88 g/L sucrose, 0.15 mg/L tyrosine, and 0.77 mg/L BAP. Building on this research, Mueangnak et al. [15] investigated the effects of both biotic and abiotic elicitation on betalain production in cell suspension cultures of C. argentea var. plumosa. Their findings revealed that chitosan at 5.0 mg/L and copper sulfate (CuSO₄) at 6.4 mM were the most effective elicitors, enhancing betalain production to achieve impressive concentrations of 4.65 and 4.99 mg/g DW, respectively, demonstrating both the potential and challenges of enhanced betalain production through biotechnological approaches.
6. Perspectives
Betalains represent a class of bioactive natural pigments found in several plant species, including C. argentea. These valuable compounds exhibit a diverse range of biological properties that make them increasingly attractive for applications across multiple industries. While beetroot (Beta vulgaris) has traditionally served as the main commercial source of betalains, several limitations restrict its widespread utilization. These constraints include a limited color spectrum of pigments, the presence of undesirable earthy-musty odorants (primarily geosmin and pyrazines), and high nitrate content—compounds that can serve as precursors to potentially carcinogenic nitrosamines [140]. In contrast, C. argentea offers significant advantages as an alternative source, providing a broader variety of colorants while notably lacking the problematic earthy-musty odorants found in beetroot.
Despite these promising attributes, research focusing on betalain production from C. argentea has been relatively sparse over the past decade, especially when compared to extensively studied plants such as B. vulgaris (beetroot), O. ficus-indica (prickly pear), H. polyrhizus (dragon fruit), and A. tricolor (amaranth). This research gap extends to extraction methodologies, processing techniques, and characterization of betalains’ physicochemical properties under various environmental and experimental conditions. Comprehensive investigations into these aspects are urgently needed to fully realize the potential of C. argentea as a commercial betalain source.
The complexity of betalain chemistry presents both challenges and opportunities for researchers. The betalain family comprises more than one hundred distinct compounds, each potentially exhibiting unique biological properties and activities. Isolation, identification, and thorough characterization of individual betalain compounds from C. argentea represent crucial steps toward understanding their specific beneficial effects. Moreover, elucidating the precise mechanisms through which these compounds exert their biological activities is essential for their targeted application. This knowledge gap is particularly significant in the pharmaceutical and cosmetic industries, where betalains show tremendous promise but currently lack sufficient scientific documentation to support their commercial development and regulatory approval.
7. Conclusion
C. argentea stands as one of the most promising sources of bioactive compounds in the plant kingdom. This remarkable plant not only accumulates various beneficial compounds, including saponins, polyphenols, peptides, and amino acids, but also synthesizes and stores relatively high concentrations of natural betalain pigments. These vibrant compounds extend far beyond their aesthetic value as brilliant colorants, offering an impressive array of health benefits that have captured significant scientific interest. Extensive research has demonstrated that betalains possess potent antioxidant, anti-inflammatory, and antimicrobial properties. Furthermore, they exhibit anticancer, antidiabetic, and antilipidemic activities, while also providing hepatoprotective and neuroprotective effects—a comprehensive profile that positions them as valuable ingredients across diverse industries, including food and beverages, textiles, cosmetics, and pharmaceuticals.
The future development of C. argentea as a commercial source of betalains necessitates focused research efforts in several key areas. Scientists should prioritize the optimization of cultivation methods and extraction techniques specifically tailored to maximize betalain yield and quality from this plant species. Equally important is the identification and characterization of novel betalain compounds unique to C. argentea, as these may possess distinct biological activities not found in more commonly studied sources. Comprehensive investigations into the bioavailability, potential toxicity, and therapeutic efficacy of these compounds will be essential for establishing their safety and effectiveness in various applications. Recent advancements in analytical technologies and biotechnological approaches, particularly in areas such as metabolomics, genomics, and cell culture systems, could substantially accelerate progress in this promising field, ultimately enabling more sustainable, efficient, and economically viable production of these valuable natural compounds for global markets.
Author Contributions
Conceptualization, P.K. (Preekamol Klanrit), S.T., P.K. (Poramaporn Klanrit), P.K. (Poramate Klanrit) and P.T.; resources, P.K. (Preekamol Klanrit), K.M. and P.T.; writing—original draft preparation, P.K. (Preekamol Klanrit), S.T., P.K. (Poramaporn Klanrit), P.K. (Poramate Klanrit), K.M. and P.T.; writing—review and editing, P.K. (Preekamol Klanrit), S.T., P.K. (Poramaporn Klanrit), P.K. (Poramate Klanrit) and P.T.; visualization, P.T.; project administration, P.K. (Preekamol Klanrit) and P.T.; funding acquisition, P.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fundamental Fund of Khon Kaen University in fiscal year 2024 and the National Science, Research, and Innovation Fund (NSRF).
Acknowledgments
The authors thank the Department of Biotechnology, Faculty of Technology, and the Fermentation Research Center for Value Added Agricultural Products (FerVAAPs), Khon Kaen University, for all facilities during the writing of this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schliemann, W.; Cai, Y.; Degenkolb, T.; Schmidt, J.; Corke, H. Betalains of Celosia argentea. Phytochem. 2001, 58, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Surse, S.N.; Shrivastava, B.; Sharma, P.; Gide, P.S.; Attar, S. Celosia cristata: Potent pharmacotherapeutic herb—A review. Int. J. Pharm. Phytopharm. Res. 2014, 3, 444–446. [Google Scholar]
- Nidavani, R.B.; Mahalakshmi, A.M.; Shalawadi, M. Towards a better understanding of an updated of ethnopharmacology of Celosia argentea L. Int. J. Pharm. Pharm. Sci. 2013, 5 (Suppl. 3), 54–59. [Google Scholar]
- Miguel, M.G. Betalains in some species of the Amaranthaceae family: A review. Antioxidants. 2018, 7, 53. [Google Scholar] [CrossRef]
- Cai, Y.; Sun, M.; Schliemann, W.; Corke, H. Chemical stability and colorant properties of betaxanthin pigments from Celosia argentea. J. Agric. Food Chem. 2001, 49, 4429–4435. [Google Scholar] [CrossRef]
- Rub, R.A.; Pati, M.J.; Siddiqui, A.A.; Moghe, A.S.; Shaikh, N.N. Characterization of anticancer principles of Celosia argentea (Amaranthaceae). Pharmacogn. Res. 2016, 8, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Xin, H.L.; Guo, M.L. Review on research of the phytochemistry and pharmacological activities of Celosia argentea. Braz. J. Pharmacogn. 2016, 26, 787–796. [Google Scholar] [CrossRef]
- Wu, Q.B.; Wang, Y.; Liang, L.; Jiang, Q.; Guo, M.L.; Zhang, J.J. Novel triterpenoid saponins from the seeds of Celosia argentea L. Nat. Prod. Res. 2013, 27, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Sun, Z.L.; Guo, M.L.; Wang, Y.; Zhang, G. Two new compounds from Semen celosiae and their protective effects against CCl4-induced hepatotoxicity. Nat. Prod. Res. 2011, 25, 772–780. [Google Scholar] [CrossRef]
- Rub, R.A.; Patil, M.J.; Shaikh, N.N.; Haikh, T.; Ahmed, J.; Siddiqui, A.A. Immunomodulatory profile of Celosia argentea–Activity of isolated compounds I and II. Int. J. Adv. Biotechnol. Res. 2015, 6, 270–277. [Google Scholar]
- Shen, S.; Ding, X.; Ouyang, M.A.; Wu, Z.J.; Xie, L.H. A new phenolic glycoside and cytotoxic constituents from Celosia argentea. J. Asian Nat. Prod. Res. 2010, 12, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Bhujbal, S.S.; Chitlange, S.S.; Suralkar, A.A.; Shinde, D.B.; Patil, M.J. Anti-inflammatory activity of an isolated flavonoid fraction from Celosia argentea Linn. J. Med. Plant Res. 2008, 2, 52–54. [Google Scholar]
- Suzuki, H.; Morita, H.; Iwasaki, S.; Kobayashi, J. New antimitotic bicyclic peptides, celogentins D-H and J, from the seeds of Celosia argentea. Tetrahedron. 2003, 59, 5307–5315. [Google Scholar] [CrossRef]
- Sang, A.; Roon, T.; Klanrit, P.; Klanrit, P.; Thanonkeo, P.; Apiraksakorn, J.; Thanonkeo, S.; Klanrit, P. Establishment of betalain-producing cell line and optimization of pigment production in cell suspension cultures of Celosia argentea var. plumosa. Plants. 2024, 13, 3225. [Google Scholar] [CrossRef] [PubMed]
- Mueangnak, K.; Kitwetcharoen, H.; Thanonkeo, S.; Klanrit, P.; Apiraksakorn, J.; Klanrit, P.; Klanrit, P.; Thanonkeo, P. Enhancing betalains production and antioxidant activity in Celosia argentea cell suspension cultures using biotic and abiotic elicitors. Sci. Rep-UK. 2025, 15, 376. [Google Scholar] [CrossRef]
- Odukoya, O.A.; Inya-Agha, S.I.; Segun, F.I.; Sodifiya, M.O.; Ilori, O.O. Antioxidant activities of selected Nigerian green leafy vegetables. Am. J. Food Technol. 2007, 2, 169–175. [Google Scholar] [CrossRef]
- Molehin, O.R.; Adefegha, S.A.; Oboh, G.; Saliu, J.A.; Athayde, M.L.; Boligon, A.A. Comparative study on the phenolic content, antioxidant properties and HPLC fingerprinting of three varieties of Celosia species. J. Food Biochem. 2014, 38, 575–583. [Google Scholar] [CrossRef]
- Wiart, C.; Mogana, S.; Khalifah, S.; Mahan, M.; Ismail, S.; Buckle, M.; Narayana, A.K.; Sulaiman, M. Antimicrobial screening of plants used for traditional medicine in the state of Perak, Peninsular Malaysia. Fitoterapia. 2004, 75, 68–73. [Google Scholar] [CrossRef]
- Vetrichelvan, T.; Jegadeesan, M.; Devi, B.A.U. Anti-diabetic activity of alcoholic extract of Celosia argentea Linn. seeds in rats. Biol. Pharm. Bull. 2002, 25, 526–528. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Fujii, H.; Hase, K.; Ohnishi, Y.; Sakukawa, R.; Kadota, S.; Namba, T.; Saiki, I. Anti-metastatic and immunomodulating properties of the water extract from Celosia argentea seeds. Biol. Pharm. Bull. 1998, 21, 1154–1159. [Google Scholar] [CrossRef]
- Hase, K.; Kadota, S.; Basnet, P.; Takahashi, T.; Namba, T. Hepatoprotective effects of traditional medicines. Isolation of the active constituent from seeds of Celosia argentea. Phytother. Res. 1996, 10, 387–392. [Google Scholar] [CrossRef]
- Showkat, S.; Rafiq, A.; Richa, R.; Sidique, Q.; Hussain, A.; Lohani, U.C.; Bhat, O.; Kumar, S. Stability enhancement of betalain pigment extracted from Celosia cristata L. flower through copigmentation and degradation kinetics during storage. Food Chem. X. 2025, 26, 102312. [Google Scholar] [CrossRef]
- Palada, M.C.; Crossman, S.M.A. Evaluation of tropical leaf vegetables in the Virgin Islands. In Perspectives on New Crops and New Uses; Janick, J., Ed.; ASHS press: Alexandria, VA, 1999; pp. 388–393. [Google Scholar]
- Lock, M.; Grubben, G.J.H.; Denton, O.A. Plant resources of tropical Africa 2. Vegetables. Kew Bull. 2004, 59, 650. [Google Scholar] [CrossRef]
- Zhang, H.C.; Zhang, T.T.; Du, B.; Cheng, D.Y.; Li, Z.G. Chemical constituents of Celosia cristata L. Chinese Trad. Patent Med. 2014, 36, 122–125. [Google Scholar]
- Zheng, X.L.; Wei, J.H.; Sun, W.; Li, R.T.; Liu, S.B.; Dai, H.F. Ethnobotanical study on medicinal plants around Limu Mountains of Hainan Island, China. J. Ethnopharmacol. 2013, 148, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.L.; Wang, Y.; Guo, M.L.; Li, Y.X. Two new hepaprotective saponins from Semen celosiae. Fitoterapia. 2010, 81, 375–380. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, Y.; Guo, M. Triterpenoid saponins from the seeds of Celosia argentea and their anti-inflammatory and antitumor activities. Chem. Pharm. Bull. 2011, 59, 666–671. [Google Scholar] [CrossRef]
- Wu, Q.B.; Wang, Y.; Liang, L.; Jiang, Q.; Guo, M.L.; Zhang, J.J. Novel triterpenoid saponins from the seeds of Celosia argentea L. Nat. Prod. Res. 2013, 27, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Yan, H.X.; Wang, Z.F.; Fan, M.X.; Zhao, Y.; Fu, X.T.; Xiong, C.Q.; Zhang, J.; Ma, B.P.; Guo, H.Z. New oleanane-type triterpenoid saponins isolated from the seeds of Celosia argentea. J. Asian Nat. Prod. Res. 2014, 16, 240–247. [Google Scholar] [CrossRef]
- Morita, H.; Shimbo, K.; Shigemori, H.; Kobayashi, J. Antimitotic activity of moroidin, a bicyclic peptide from the seeds of Celosia argentea. Bioorg. Med. Chem. Lett. 2000, 10, 469–471. [Google Scholar] [CrossRef]
- Kobayashi, J.; Suzuki, H.; Shimbo, K.; Takeya, K.; Morita, H. Celogentins A–C, new antimitotic bicyclic peptides from the seeds of Celosia argentea. J. Org. Chem. 2001, 66, 6626–6633. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Morita, H.; Iwasaki, S.; Kobayashi, J. New antimitotic bicyclic peptides, celogentins D–H, and J, from the seeds of Celosia argentea. Tetrahedron. 2003, 59, 5307–5315. [Google Scholar] [CrossRef]
- Suzuki, H.; Morita, H.; Shiro, M.; Kobayashi, J.I. Celogentin K, a new cyclic peptide from the seeds of Celosia argentea and X-ray structure of moroidin. Tetrahedron. 2004, 60, 2489–2495. [Google Scholar] [CrossRef]
- Morita, H.; Suzuki, H.; Kobayashi, J. Celogenamide A, a new cyclic peptide from the seeds of Celosia argentea. J. Nat. Prod. 2004, 67, 1628–1630. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.H.; Cui, X.; Zhou, P.; Li, S.L. A comparative study of fatty acids and inorganic elements in Semen celosiae and cockscomb. J. Chinese Med. Mat. 1995, 18, 466–467. [Google Scholar]
- Lin, W.Q.; Chen, Z.; Liu, J.Q. The chemical constituents of Perilla frutescens (L.) Britt. var. acute (Thunb.) and Celosia argentea L. seeds grown in Fujian province. Chinese Acad. Med. Magazing Organisms 2002, 57–59. [Google Scholar]
- Fu, H.Z.; Meng, X.Y.; Li, S.S.; Wu, L.J. Study on the chemical constituents of Semen celosiae. Chinese Trad. Herbal Drugs. 1992, 23, 344–345. [Google Scholar]
- Xue, Q.; Guo, M.; Zhang, G. Study of chemical constituents of Semen celosiae. Pharm. Care Res. 2006, 6, 345–347. [Google Scholar]
- Markandeya, A.G.; Firke, N.P.; Pingale, S.S.; Salunke-Gawali, S. Quantitative elemental analysis of Celosia argentea leaves by ICP-OES technique using various digestion methods. Int. J. Chem. Anal. Sci. 2013, 4, 175–181. [Google Scholar] [CrossRef]
- Thorat, B.R. Review on Celosia argentea L. plant. Res. J. Pharmacognosy Phytochem. 2018, 10, 109–119. [Google Scholar] [CrossRef]
- Divya, B.J.; Jyothi Sravani, M.; Hari Chandana, J.; Sumana, T.; Thyagaraju, K. Phytochemical and phytotherapeutic activities of Celosia argentea: a review. World J. Pharm. Pharmaceu. Sci. 2019, 8, 488–505. [Google Scholar]
- Khan, M.I.; Harsha, P.S.C.S.; Chauhan, A.S.; Vijayendra, S.V.N.; Asha, M.R.; Giridhar, P. Betalains rich Rivina humilis L. berry extract as natural colorant in product (fruit spread and RTS beverage) development. J. Food Sci. Technol. 2015, 52, 1808–1813. [Google Scholar] [CrossRef]
- Polturak, G.; Aharoni, A. “La Vie en Rose”: biosynthesis, sources, and applications of betalain pigments. Mol Plant 2018, 11, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Carreón-Hidalgo, J.P.; Franco-Vásquez, D.C.; Gómez-Linton, D.R.; Pérez-Flores, L.J. Betalain plant sources, biosynthesis, extraction, stability enhancement methods, bioactivity, and applications. Food Res. Int. 2022, 151, 110821. [Google Scholar] [CrossRef]
- Azeredo, H.M.C. Betalains: properties, sources, applications, and stability—A review. Int. J. Food Sci. Technol. 2009, 44, 2365–2376. [Google Scholar] [CrossRef]
- Gandía-Herrero, F.; García-Carmona, F. Biosynthesis of betalains: yellow and violet plant pigments. Trends Plant Sci. 2013, 18, 334–343. [Google Scholar] [CrossRef]
- Slimen, I.B.; Najar, T.; Abderrabba, M. Chemical and antioxidant properties of betalains. J. Agric. Food Chem. 2017, 65, 675–689. [Google Scholar] [CrossRef]
- Murthy, H.N.; Joseph, K.S.; Paek, K.Y.; Park, S.Y. Production of betalains in plant cell and organ cultures: A review. Plant Cell Tiss Org 2024, 158, 28. [Google Scholar] [CrossRef]
- Strack, D.; Vogt, T.; Schliemann, W. Recent advances in betalain research. Phytochemistry 2003, 62, 247–269. [Google Scholar] [CrossRef]
- Khan, M.I. Stabilization of betalains: A review. Food Chem. 2016, 197, 1280–1285. [Google Scholar] [CrossRef]
- Xie, F.; Hua, Q.; Chen, C.; Zhang, L.; Zhang, Z.; Chen, J.; Zhang, R.; Zhao, J.; Hu, G.; Zhao, J.; Qin, Y. Transcriptomics-based identification and characterization of glucosyltransferases involved in betalain biosynthesis in Hylocereus megalanthus. Plant Physiol. Biochem. 2020, 152, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Chiu, Y.C.; Tsao, N.W.; Chou, Y.L.; Tan, C.M.; Chiang, Y.H.; Liao, P.C.; Lee, Y.C.; Hsieh, L.C.; Wang, S.Y.; Yang, J.Y. Elucidation of the core betalain biosynthesis pathway in Amaranthus tricolor. Sci. Rep.-UK. 2021, 11, 6086. [Google Scholar] [CrossRef]
- Tzin, V.; Galili, G. The biosynthetic pathways for shikimate and aromatic amino acids in Arabidopsis thaliana. Arabidopsis Book. 2010, 8, e0132. [Google Scholar] [CrossRef] [PubMed]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef]
- Chen, N.; Yu, Z.H.; Xiao, X.G. Cytosolic and nuclear colocalization of betalain biosynthetic enzymes in tobacco suggests that betalains are synthesized in the cytoplasm and/or nucleus of betalainic plant cells. Front Plant Sci 2017, 8, 831. [Google Scholar] [CrossRef]
- Sunnadeniya, R.; Bean, A.; Brwon, M.; Akhavan, N.; Hatlestad, G.; Gonzalez, A.; Symonds, V.V.; Lloyd, A. Tyrosine hydroxylation in betalain pigment biosynthesis is performed by cytochrome P450 enzymes in beets (Beta vulgaris). PLoS ONE. 2016, 11, e0149417. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, N.; Aabe, Y.; Goda, Y.; Adachi, T.; Kasahara, K.; Ozeki, Y. Detection of DOPA 4,5-dioxygenase (DOD) activity using recombinant protein prepared from Escherichia coli cells harboring cDNA encoding DOD from Mirabilis jalapa. Plant Cell Physiol. 2009, 50, 1012–1016. [Google Scholar] [CrossRef]
- Fischer, N.; Dreiding, A.S. Biosynthesis of betalains. On the cleavage of aromatic ring during enzymatic transformation of DOPA into betalamic acid. Helv. Chim. Acta. 1972, 55, 649–658. [Google Scholar] [CrossRef]
- Sasaki, N.; Adachi, T.; Koda, T.; Ozeki, Y. Detection of UDP glucose:cyclo-DOPA 5-O-glucosyltransferase activity in four o’clocks (Mirabilis jalapa L). FEBS Lett. 2004, 568, 159–162. [Google Scholar] [CrossRef]
- Sasaki, N.; Wada, K.; Koda, T.; Kasahara, K.; Adachi, T.; Ozeki, Y. Isolation and characterization of cDNAs encoding an enzyme with glucosyltransferase activity for cyclo-DOPA from four o’clocks and feather cockscombs. Plant Cell Physiol. 2005, 46, 666–670. [Google Scholar] [CrossRef]
- Belhadj Slimen, I.; Najar, T.; Abderrabba, M. Chemical and antioxidant properties of betalains. J. Agric. Food Chem. 2017, 65, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Esatbeyoglu, T.; Wagner, A.E.; Schini-Kerth, V.B.; Rimbach, G. Betanin– a food colorant with biological activity. Mol. Nutr. Food Res. 2015, 59, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Vieira Teixeira da Silva, D.; dos Santos Baião, D.; de Oliveira Silva, F.; Alves, G.; Perrone, D.; Mere Del Aguila, E.; Flosi Paschoalin, V.M. Betanin, a natural food additive: stability, bioavailability, antioxidant and preservative ability assessments. Molecules. 2019, 24, 458. [Google Scholar] [CrossRef]
- Gonçalves, L.C.P.; Lopes, N.B.; Augusto, F.A.; Pioli, R.M.; Machado, C.O.; Freitas-Dörr, B.C.; Suffredini, H.B.; Bastos, E.L. Phenolic betalain as antioxidants: Meta means more. Pure Appl. Chem. 2020, 92, 243–253. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Venkidasamy, B.; Subramanian, U.; Samynathan, R.; Ali Shariati, M.; Rebezov, M.; Girish, S.; Thangavel, S.; Dhanapal, A.R.; Fedoseeva, N.; et al. Bioactive compounds in oxidative stress-mediated diseases: targeting the NRF2/ARE signaling pathway and epigenetic regulation. Antioxidants. 2021, 10, 1859. [Google Scholar] [CrossRef]
- Attanzio, A.; Frazzitta, A.; Busa, R.; Tesoriere, L.; Livrea, M.A.; Allegra, M. Indicaxanthin from Opuntia ficus-indica (L. Mill) inhibits oxidized LDL-mediated human endothelial cell dysfunction through inhibition of NF-κB activation. Oxid. Med. Cell. Longe. 2019, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, H.; Nayeri, Z.; Minuchehr, Z.; Sabouni, F.; Mohammadi, M. Betanin purification from red beetroots and evaluation of its antioxidant and anti-inflammatory activity on LPS-activated microglial cells. PLoS One. 2020, 15, 1–18. [Google Scholar] [CrossRef]
- Shunan, D.; Yu, M.; Guan, H.; Zhou, Y. Neuroprotective effect of betalain against AlCl3-induced Alzheimer’s disease in Sprague Dawley rats via putative modulation of oxidative stress and nuclear factor kappa B (NF-ΚB) signaling pathway. Biomed. Pharmaco. 2021, 137, 111369. [Google Scholar] [CrossRef]
- Wang, Y.; Fernando, G.S.N.; Sergeeva, N.N.; Vagkidis, N.; Chechik, V.; Do, T.; Marshall, L.J.; Boesch, C. Uptake and immunomodulatory properties of betanin, vulgaxanthin I and indicaxanthin towards Caco-2 intestinal cells. Antioxidants. 2022, 11, 1627. [Google Scholar] [CrossRef]
- Fernando, G.S.N.; Sergeeva, N.N.; Vagkidis, N.; Chechik, V.; Marshall, L.J.; Boesch, C. Differential effects of betacyanin and betaxanthin pigments on oxidative stress and inflammatory response in murine macrophages. Mol. Nutri. Food Res. 2023, 67, 2200583. [Google Scholar] [CrossRef]
- Moreno-Ley, C.M.; Osorio-Revilla, G.; Hernández-Martínez, D.M.; Ramos-Monroy, O.A.; Gallardo-Velázquez, T. Anti-inflammatory activity of betalains: A comprehensive review. Human Nutri. Metab. 2021, 25, 200126. [Google Scholar] [CrossRef]
- Canadanović-Brunet, J.M.; Savatović, S.S.; Ćetković, G.S.; Vulić, J.J.; Djilas, S.M.; Markov, S.L.; Cvetković, D.D. Antioxidant and antimicrobial activities of beet root pomace extracts. Czech J. Food Sci. 2011, 29, 575–585. [Google Scholar] [CrossRef]
- Yong, Y.Y.; Dykes, G.; Lee, S.M.; Choo, W.S. Comparative study of betacyanin profile and antimicrobial activity of red pitahaya (Hylocereus polyrhizus) and red spinach (Amaranthus dubius). Plant Foods Human Nutri. 2017, 72, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Melgar, B.; Dias, M.I.; Ciric, A.; Sokovic, M.; Garcia-Castello, E.M.; Rodriguez-Lopez, A.D.; Barros, L.; Ferreira, I. By-product recovery of Opuntia spp. peels: Betalainic and phenolic profiles and bioactive properties. Ind. Crops Prod. 2017, 107, 353–359. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Bartosz, G. Biological properties and applications of betalains. Molecules. 2021, 26, 2520. [Google Scholar] [CrossRef]
- Sandate-Flores, L.; Romero-Esquivel, E.; Rodríguez-Rodríguez, J.; Rostro-Alanis, M.; Melchor-Martínez, E.M.; Castillo-Zacarías, C.; Ontiveros, P.R.; Celaya, M.F.M.; Chen, W.-N.; Iqbal, H.M.N.; et al. Functional attributes and anticancer potentialities of Chico (Pachycereus Weberi) and Jiotilla (Escontria Chiotilla) fruits extract. Plants. 2020, 9, 1623. [Google Scholar] [CrossRef]
- Henarejos-Escudero, P.; Hernández-García, S.; Guerrero-Rubio, M.A.; García-Carmona, F.; Gandía-Herrero, F. Antitumoral drug potential of tryptophan-betaxanthin and related plant betalains in the Caenorhabditis elegans tumoral model. Antioxidants. 2020, 9, 646. [Google Scholar] [CrossRef] [PubMed]
- Zielińska-Przyjemska, M.; Olejnik, A.; Dobrowolska-Zachwieja, A.; Łuczak, M.; Baer-Dubowska, W. DNA damage and apoptosis in blood neutrophils of inflammatory bowel disease patients and in Caco-2 cells in vitro exposed to betanin. Postepy Hig Med Dosw (Online). 2016, 70, 265–271. [Google Scholar] [CrossRef]
- Nowacki, L.; Vigneron, P.; Rotellini, L.; Cazzola, H.; Merlier, F.; Prost, E.; Ralanairina, R.; Gadonna, J.P.; Rossi, C.; Vayssade, M. Betanin-enriched red beetroot (Beta vulgaris L.) extract induces apoptosis and autophagic cell death in MCF-7 cells. Phytochem. Rev. 2015, 29, 1964–1973. [Google Scholar]
- Allegra, M.; De Cicco, P.; Ercolano, G.; Attanzio, A.; Busá, R.; Cirino, G.; Tesoriere, L.; Livrea, M.A.; Ianaro, A. Indicaxanthin from Opuntia ficus-indica (L. Mill) impairs melanoma cell proliferation, invasiveness, and tumor progression. Phytomedicine. 2018, 50, 19–24. [Google Scholar] [CrossRef]
- Yin, Z.; Yang, Y.; Guo, T.; Veeraraghavan, V.P.; Wang, X. Potential chemotherapeutic effect of betalain against human non-small cell lung cancer through PI3K/Akt/mTOR signaling pathway. Environ. Toxicol. 2021, 36, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Yu, K.; Chu, X.; Yang, L. Betanin alleviates inflammation and ameliorates apoptosis on human oral squamous cancer cells SCC131 and SCC4 through the NF-κB/PI3K/Akt signaling pathway. J. Biochem. Mol. Toxicol. 2022, 36, e23094. [Google Scholar] [CrossRef]
- Tsiailanis, A.D.; Chatzigiannis, C.M.; Papaemmanouil, C.D.; Chatziathanasiadou, M.V.; Chaloulos, P.; Riba, I.; Mullard, G.; Wiczkowski, W.; Koutinas, A.; Mandala, J.; Tzakos, A.G. Exploration of betalains and determination of the antioxidant and cytotoxicity profile of orange and purple Opuntia spp. cultivars in Greece. Plant Foods Human Nutri. 2022, 77, 198–205. [Google Scholar] [CrossRef]
- Coimbra, P.P.S.; Silva-e-Silva, A.C.A.G.d.; Antonio, A.d.S.; Pereira, H.M.G.; Veiga-Junior, V.F.d.; Felzenszwalb, I.; Araujo-Lima, C.F.; Teodoro, A.J. Antioxidant capacity, antitumor activity and metabolomic profile of a beetroot peel flour. Metabolites. 2023, 13, 277. [Google Scholar] [CrossRef]
- Madadi, E.; Mazloum-Ravasan, S.; Yu, J.S.; Ha, J.W.; Hamishehkar, H.; Kim, K.H. Therapeutic application of betalains: A review. Plants. 2020, 9, 1219. [Google Scholar] [CrossRef]
- Ragusa, M.A.; Naselli, F.; Cruciata, I.; Volpes, S.; Schimmenti, C.; Serio, G.; Mauro, M.; Librizzi, M.; Luparello, C.; Chiarelli, R.; et al. Indicaxanthin induces autophagy in intestinal epithelial cancer cells by epigenetic mechanisms involving DNA methylation. Nutrients. 2023, 15, 3495. [Google Scholar] [CrossRef]
- Alian, D.M.E.; Helmy, M.W.; Haroun, M.; Moussa, N. Modulation of autophagy and apoptosis can contribute to the anticancer effect of Abemaciclib/celecoxib combination in colon cancer cells. Med. Oncol. 2024, 41, 43. [Google Scholar] [CrossRef] [PubMed]
- Montazersaheb, S.; Jafari, S.; Aytemir, M.D.; Ahmadian, E.; Ardalan, M.; Zor, M.; Nasibova, A.; Monirifar, A.; Aghdasi, S. The synergistic effects of betanin and radiotherapy in a prostate cancer cell line: An in vitro study. Mol. Biol. Rep. 2023, 50, 9307–9314. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Tan, C.; Wang, Y.; Yang, S.; Tan, D. Betanin reduces the accumulation and cross-links of collagen in high-fructose-fed rat heart through inhibiting non-enzymatic glycation. Chem-Biol. Interact. 2015, 227, 37–44. [Google Scholar] [CrossRef]
- Khan, M.I. Plant betalains: Safety, antioxidant activity, clinical efficacy, and bioavailability. Comp. Rev. Food Sci. Food Safety. 2016, 15, 316–330. [Google Scholar] [CrossRef]
- Rivera, A.; Becerra-Martínez, E.; Pacheco-Hernández, Y.; Landeta-Cortés, G.; Villa-Ruano, N. Synergistic hypolipidemic and hypoglycemic effects of mixtures of Lactobacillus nagelii/betanin in a mouse model. Trop. J. Pharm. Res. 2020, 19, 1269–1276. [Google Scholar] [CrossRef]
- Gómez-Maqueo, A.; García-Cayuela, T.; Fernández-López, R.; Welti-Chanes, J.; Cano, M.P. Inhibitory potential of prickly pears and their isolated bioactives against digestive enzymes linked to type 2 diabetes and inflammatory response. J. Sci. Food Agric. 2019, 99, 6380–6391. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, P.; Mesbah-Namin, S.A.; Ostadrahimi, A.; Abedimanesh, S.; Separham, A.; Jafarabadi, M.A. Effects of betalains on atherogenic risk factors in patients with atherosclerotic cardiovascular disease. Food Func. 2019, 10, 8286–8297. [Google Scholar] [CrossRef]
- Ydjedd, S.; Chaalal, M.; Bahri, S.; Mokadem, S.; Radji, H. Antioxidant and α-amylase inhibition activities of prickly pears (Opuntia ficus indica L.) betalains extracts and application in yogurt as natural colorants. J. Agro. Proc. Technol. 2021, 27, 140–150. [Google Scholar]
- da Silva, D.V.T.; Pereira, A.D.; Boaventura, G.T.; Ribeiro, R.S.d.A.; Verícimo, M.A.; Carvalho-Pinto, C.E.d.; Baião, D.d.S.; Del Aguila, E.M.; Paschoalin, V.M.F. Short-term betanin intake reduces oxidative stress in Wistar Rats. Nutrients. 2019, 11, 1978. [Google Scholar] [CrossRef]
- Martínez-Rodríguez, P.; Henarejos-Escudero, P.; Hernández-García, S.; Sánchez-Ferrer, Á.; Gandía-Herrero, F. In vitro, in vivo, and in silico evidence for the use of plant pigments betalains as potential nutraceuticals against Alzheimer’s disease. Food Front. 2024, 5, 2137–2154. [Google Scholar] [CrossRef]
- Abedimanesh, N.; Asghari, S.; Mohammadnejad, K.; Daneshvar, Z.; Rahmani, S.; Shokoohi, S.; Motlagh, B.; et al. The anti-diabetic effects of betanin in streptozotocin-induced diabetic rats through modulating AMPK/SIRT1/NF-κB signaling pathway. Nutri. Meta. 2021, 18, 92. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Wang, Y.; Zhang, Z. Radioprotective activity of betalains from red beets in mice exposed to gamma irradiation. Eur. J. Pharm. 2009, 615, 223–227. [Google Scholar] [CrossRef]
- Krajka-Kuźniak, V.; Szaefer, H.; Ignatowicz, E.; Adamska, T.; Baer-Dubowska, W. Beetroot juice protects against N-nitrosodiethylamine-induced liver injury in rats. Food Chem. Toxic. 2012, 50, 2027–2033. [Google Scholar] [CrossRef]
- Motawi, T.K.; Ahmed, S.A.; El-Boghdady, N.A.; Metwally, N.S.; Nasr, N.N. Impact of betanin against paracetamol and diclofenac induced hepato-renal damage in rats. Biomarkers. 2020, 25, 86–93. [Google Scholar] [CrossRef]
- Liu, Y.; Miao, L.; Guo, Y.; Tian, H.; Dos Santos, J.M. Preclinical evaluation of safety, pharmacokinetics, efficacy, and mechanism of radioprotective agent HL-003. Oxid. Med. Cell. Longev. 2021, 2021, 6683836. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rodríguez, P.M.; Guerrero-Rubio, A.; Henarejos-Escudero, P.; García-Carmona, F.; Gandía-Herrero, F. Health-promoting potential of betalains in vivo and their relevance as functional ingredients: A review. Trends Food Sci. Technol. 2022, 122, 66–82. [Google Scholar] [CrossRef]
- Attia, G.Y.; Moussa, M.E.M.; Sheashea, E.R. Characterization of red pigments extracted from red beet (Beta vulgaris L.) and its potential uses as antioxidant and natural food colorants. Egypt. J. Agric. Res. 2013, 91, 1095–1110. [Google Scholar] [CrossRef]
- Kumar, S.S.; Manoj, P.; Shetty, N.P.; Prakash, M.; Giridhar, P. Characterization of major betalain pigments-gomphrenin, betanin and isobetanin from Basella rubra L. fruit and evaluation of efficacy as a natural colourant in product (ice cream) development. J. Food Sci. Technol. 2015, 52, 4994–5002. [Google Scholar] [CrossRef]
- Roriz, C.L.; Barreira, J.C.M.; Morales, P.; Barros, L.; Ferreira, I.C.F.R. Gomphrena globosa L. as a novel source of food-grade betacyanins: Incorporation in ice-cream and comparison with beet-root extracts and commercial betalains. LWT—Food Sci. Technol. 2018, 92, 101–107. [Google Scholar] [CrossRef]
- Khan, M.I.; Harsha, P.S.C.S.; Chauhan, A.S.; Vijayendra, S.V.N.; Asha, M.R.; Giridhar, P. Betalains rich Rivina humilis L. berry extract as natural colorant in product (fruit spread and RTS beverage) development. J. Food Sci. Technol. 2015, 52, 1808–1813. [Google Scholar] [CrossRef]
- Güneşer, O. Pigment and color stability of beetroot betalains in cow milk during thermal treatment. Food Chem. 2016, 196, 220–227. [Google Scholar] [CrossRef]
- Gengatharan, A.; Dykes, G.A.; Choo, W.S. Stability of betacyanin from red pitahaya (Hylocereus polyrhizus) and its potential application as a natural colourant in milk. Int. J. Food Sci. Technol. 2016, 51, 427–434. [Google Scholar] [CrossRef]
- Gengatharan, A.; Dykes, G.A.; Choo, W.S. The effect of pH treatment and refrigerated storage on natural colourant preparations (betacyanins) from red pitahaya and their potential application in yoghurt. LWT—Food Sci. Technol. 2017, 80, 437–445. [Google Scholar] [CrossRef]
- Coria-Cayupán, Y.; Nazareno, M.A. Cactus betalains can be used as antioxidant colorants protecting food constituents from oxidative damage. Acta Hort. 2015, 1067, 319–325. [Google Scholar] [CrossRef]
- Rodríguez-Sánchez, J.A.; María, T.C.V.; Barragán-Huerta, B.E. Betaxanthins and antioxidant capacity in Stenocereus pruinosus: Stability and use in food. Food Res. Int. 2017, 91, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Leal-Alcazar, M.C.; Bautista-Palestina, F.; Rocha-Pizana, M.R.; Mojica, L.; Hernández-Álvarez, A.J.; Luna-Vital, D.A. Extraction, stabilization, and health application of betalains: An update. Food Chem. 2025, 481, 144011. [Google Scholar] [CrossRef] [PubMed]
- Amnah, M.A. Nutritional, sensory and biological study of biscuits fortified with red beetroots. Life Sci. J. 2013, 10, 1579–1584. [Google Scholar]
- Kumar, V.; Kushwaha, R.; Goyal, A.; Tanwar, B.; Kaur, J. Process optimization for the preparation of antioxidant-rich ginger candy using beetroot pomace extract. Food Chem. 2018, 245, 168–177. [Google Scholar] [CrossRef]
- Kumar, S.S.; Singh Chauhan, A.; Giridhar, P. Nanoliposomal encapsulation mediated enhancement of betalain stability: Characterisation, storage stability and antioxidant activity of Basella rubra L. fruits for its applications in vegan gummy candies. Food Chem. 2020, 333, 127442. [Google Scholar] [CrossRef] [PubMed]
- Iahtisham-Ul-Haq; Butt, M.S.; Randhawa, M.A.; Shahid, M. Nephroprotective effects of red beetroot-based beverages against gentamicin-induced renal stress. J. Food Biochem. 2019, 43, 12873. [Google Scholar] [CrossRef]
- Zhu, F.; Cai, Y.Z.; Corke, H. Evaluation of Asian salted noodles in the presence of Amaranthus betacyanin pigments. Food Chem. 2010, 118, 663–669. [Google Scholar] [CrossRef]
- Yavuzer, E.; Ozogul, F.; Ozogul, Y. Impact of icing with potato, sweet potato, sugar beet, and red beet peel extract on the sensory, chemical, and microbiological changes of rainbow trout (Oncorhynchus mykiss) fillets stored at (3 ± 1 °C). Aquacult. Int. 2020, 28, 187–197. [Google Scholar] [CrossRef]
- Jamróz, E.; Kulawik, P.; Guzik, P.; Duda, I. The verification of intelligent properties of furcellaran films with plant extracts on the stored fresh Atlantic mackerel during storage at 2 °C. Food Hydrocolloids 2019, 97, 105211. [Google Scholar] [CrossRef]
- Qin, Y.; Liu, Y.; Zhang, X.; Liu, J. Development of active and intelligent packaging by incorporating betalains from red pitaya (Hylocereus polyrhizus) peel into starch/polyvinyl alcohol films. Food Hydrocolloids. 2020, 100, 105410. [Google Scholar] [CrossRef]
- Yao, X.; Hu, H.; Qin, Y.; Liu, J. Development of antioxidant, antimicrobial and ammonia-sensitive films based on quaternary ammonium chitosan, polyvinyl alcohol and betalains-rich cactus pears (Opuntia ficus-indica) extract. Food Hydrocolloids. 2020, 106, 105896. [Google Scholar] [CrossRef]
- Hu, H.; Yao, X.; Qin, Y.; Yong, H.; Liu, J. Development of multifunctional food packaging by incorporating betalains from vegetable amaranth (Amaranthus tricolor L.) into quaternary ammonium chitosan/fish gelatin blend films. Int. J. Biol. Macromolecules. 2020, 159, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Guesmi, A.; Ben Hamadi, N.; Ladhari, N.; Sakli, F. Dyeing properties and colour fastness of wool dyed with indicaxanthin natural dye. Ind. Crops Prod. 2012, 37, 493–499. [Google Scholar] [CrossRef]
- Popescu, V.; Blaga, A.C.; Caşcaval, D.; Popescu, A. Beta vulgaris L.—A source with a great potential in the extraction of natural dyes intended for the sustainable dyeing of wool. Plants. 2023, 12, 1933. [Google Scholar] [CrossRef]
- El-Zawahry, M.M.; Kamel, M.M.; Hassabo, A.G. Development of bio-active cotton fabric coated with betalain extract as encapsulating agent for active packaging textiles. Ind. Crops Prod. 2024, 222, 119583. [Google Scholar] [CrossRef]
- Khalaf, M.M.; Gouda, M.; Abou Taleb, M.F.; Abdelaziz, M.A.; Abd El-Lateef, H.M. Development of betalain-finished plasma-treated nonwoven cotton textiles from beetroot (Beta vulgaris L.) and recycled fabrics for identification of ammonia. J. Environ. Chem. Eng. 2025, 13, 116145. [Google Scholar] [CrossRef]
- Azwanida, N.N.; Normasarah, A.A. Utilization and evaluation of betalain pigment from red dragon fruit (Hylocereus Polyrhizus) as a natural colorant for lipstick. J. Teknol. 2014, 69, 139–142. [Google Scholar] [CrossRef]
- Scoles, G.E.; Pattacini, S.H.; Covas, G.F. Separation of the pigment of an amaranth. Molecules. 2000, 5, 566–567. [Google Scholar] [CrossRef]
- Zhang, D.; Lanier, S.M.; Downing, J.A.; Avent, J.L.; Lumc, J.; McHalea, J.L. Betalain pigments for dye-sensitized solar cells. J. Photochem. Photobiol. Chem. 2008, 195, 72–80. [Google Scholar] [CrossRef]
- Calogero, G.; Yumb, J.H.; Sinopoli, A.; Di Marco, G.; Grätzel, M.; Nazeeruddin, M.K. Anthocyanins and betalains as light-harvesting pigments for dye-sensitized solar cells. Sol. Energy. 2012, 86, 1563–1575. [Google Scholar] [CrossRef]
- Knorr, F.J.; Malamen, D.J.; McHale, J.L.; Marchioro, A.; Moser, J.E. Two-electron photo-oxidation of betanin on titanium dioxide and potential for improved dye-sensitized solar energy conversion. Physic. Chem. Interfaces Nanomaterials XIII. 2014, 9165, 48–56. [Google Scholar]
- Knorr, F.J.; McHale, J.L.; Clark, A.E.; Marchioro, A.; Moser, J.E. Dynamics of interfacial electron transfer from betanin to nanocrystalline TiO2: the pursuit of two-electron injection. J. Physic. Chem. 2015, 119, 1903019041. [Google Scholar] [CrossRef]
- Guerrero-Rubio, M.A.; Escribano, J.; García-Carmona, F.; Gandía-Herrero, F. Light emission in betalains: From fluorescent flowers to biotechnological applications. Trends Plant Sci. 2020, 25, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Rubio, M.A.; Martínez-Zapata, J.; Henarejos-Escudero, P.; García-Carmona, F.; Gandía-Herrero, F. Reversible bleaching of betalains induced by metals and application to the fluorescent determination of anthrax biomarker. Dyes Pigment. 2020, 180, 108493. [Google Scholar] [CrossRef]
- Fares, M.M.; Bani-Domi, A. Sustainable betalain pigments as eco-friendly film coating over aluminium surface. J. Materials Sci. 2021, 56, 13556–13567. [Google Scholar] [CrossRef]
- Fernandes, D.L.A.; Paun, C.; Pavliuk, M.V.; Fernandes, A.B.; Bastos, E.L.; Sá, J. Green microfluidic synthesis of monodisperse silver nanoparticles via genetic algorithm optimization. RSC Adv. 2016, 6, 95693–95697. [Google Scholar] [CrossRef]
- Warhade, M.I.; Badere, R.S. Isolation of callus lines of Celosia cristata L. with variation in betalain content. J Indian Bot Soc 2015, 94, 89–96. [Google Scholar]
- Warhade, M.I.; Badere, R.S. Fusarium oxysporum cell elicitor enhances betalain content in the cell suspension culture of Celosia cristata. Physiol Mol Biol Plants. 2018, 24, 285–293. [Google Scholar] [CrossRef]
- Pavokovic, D.; Krsnik-Rasol, M. Complex biochemistry and biotechnological production of betalains. Food Technol. Biotechnol. 2011, 49, 145–155. [Google Scholar]
Figure 1.
Celosia argentea var. plumosa (A) and C. argentea var. cristata (B).

Figure 2.
Betalamic acid (A), betacyanin (B), and betaxanthin (C) structures [49].
Figure 2.
Betalamic acid (A), betacyanin (B), and betaxanthin (C) structures [49].
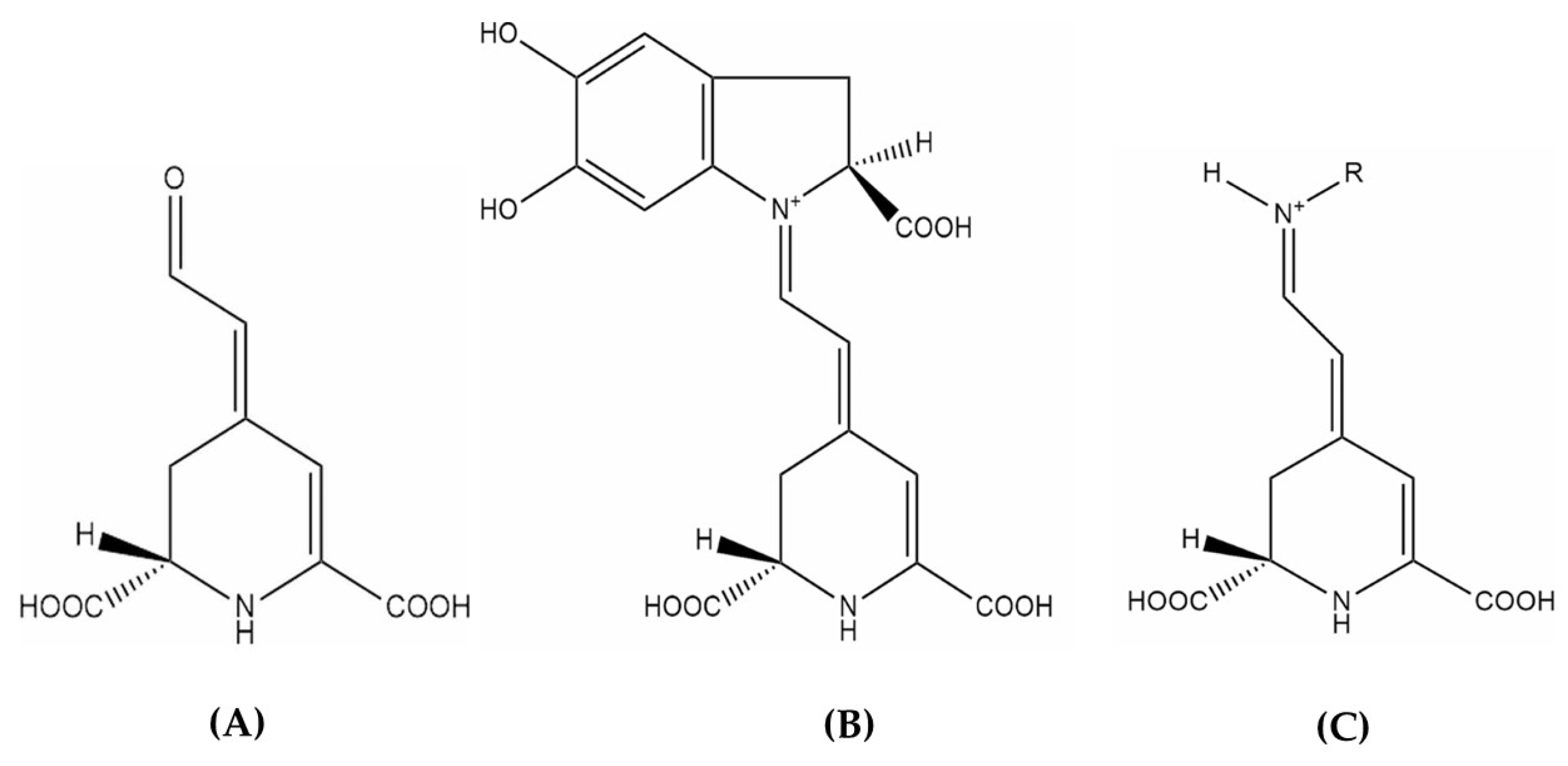
Figure 3.
Biosynthesis pathway of betalains [49].
Figure 3.
Biosynthesis pathway of betalains [49].
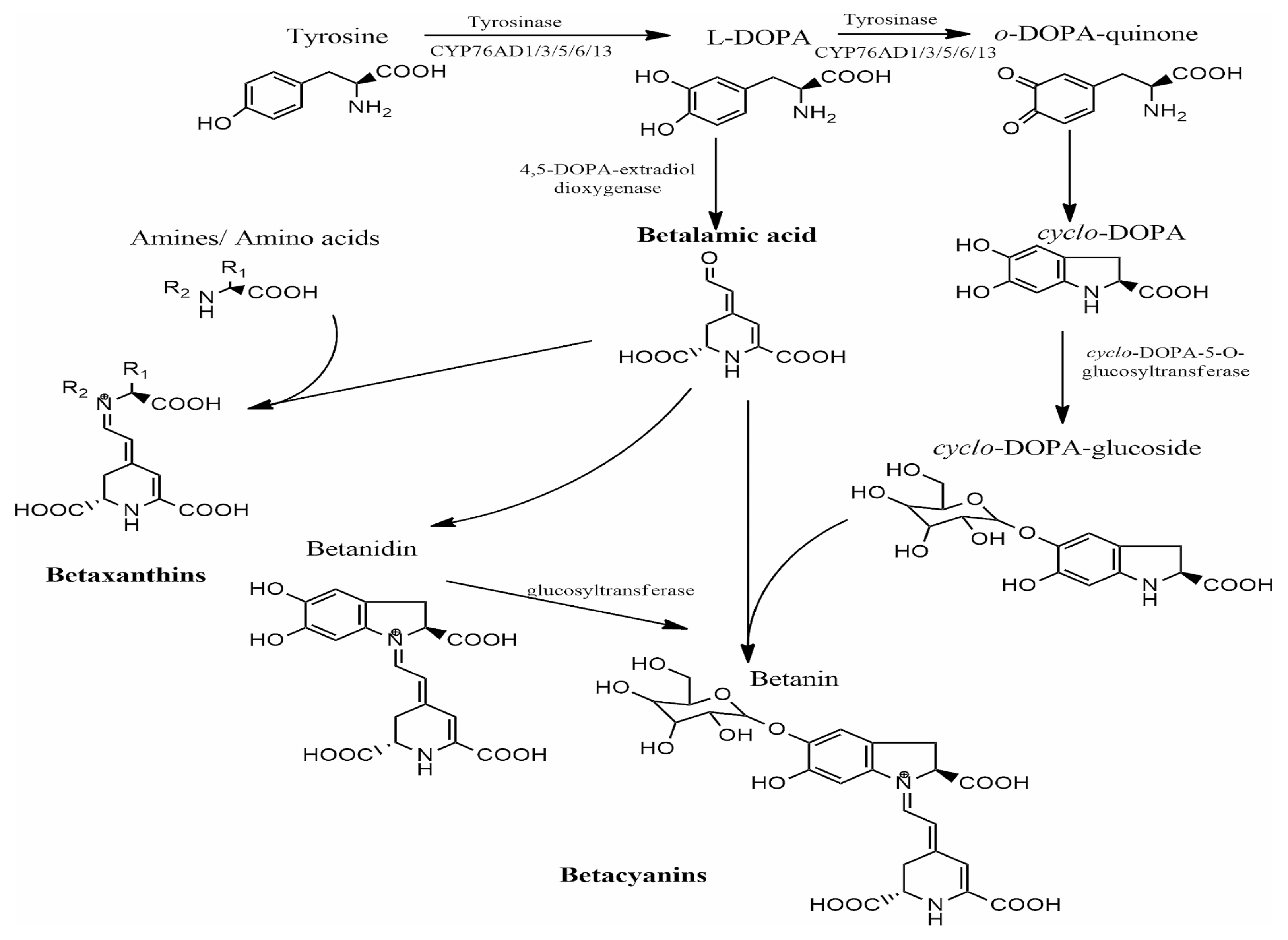

Table 1.
The major bioactive compounds found in C. argentea (modified from Tang et al. [7].
Table 1.
The major bioactive compounds found in C. argentea (modified from Tang et al. [7].
| Bioactive compound | Chemical | Analytical technique | Plant part | Reference |
| Saponins | Celosin A, Celosin B, Celosin C, Celosin D, Celosin E, Celosin F, Celosin G, Celosin I, Celosin II, Celosin H, Celosin I, Celosin J, Cristatain | NMR, HPLC-ELSD | Seed | [9,27,28,29,30] |
| Polyphenols | Lutin, Epigallocatechin, Gallic acid, Caffeic acid, Rosmarinic acid, Quercetin, 4-O-β-d-apifuranosyl-(1→2)-β-d-glucopyranosyl-2-hydroxy-6-methoxyacetophenone | HPLC | Leaf | [11,17] |
| Peptides | Moroidin, Celogentins A, Celogentins B, Celogentins C, Celogentins D, Celogentins E, Celogentins F, Celogentins G, Celogentins H, Celogentins J, Celogentins K, Celogenamide A | NMR, MS/MS, CD spectra | Seed | [31,32,33,34,35] |
| Amino acids | Glycine, Alanine, Arginine, Lysine, Glutamic acid, Valine, Methionine, Isoleucine, Phenylalanine, Serine, Tyrosine, Proline, Leucine, Histidine, Aspartic acid, Cysteine, Cytine, Threonine, Ornithine | Amino acid analyzer | Seed, Leaf | [36,37] |
| Fatty acids | Arachic acid, Arachidonic acid, Linolenic acid, Hexadecanoic acid, Palmitoleic acid, Octadecanoic acid, Octadecanoic monoenoic acid, Oleinic acid, Linoleic acid | GC | Seed | [36,37] |
| Betalains | Betaxanthins (Indicaxanthin, Dopaxanthin), Betacyanins (Betanin, Gomphrenin, Amaranthine, and Bougainvillein) | Spectrophotometry | Leaf, Inflorescence | [1,14,15] |
| Minerals | K, Ca, Mg, Na, Fe, Mn, Cu, Zn, S, Si, Ti, Cd, Hg, Cr, Mo, Pb | AA | Seed, Leaf | [36,37] |
| Others | Β-Sitosterol, Stigmasterol, β-Carotene, Ascorbic acid | - | Seed, Leaf | [38,39] |
Table 2.
Biological properties of betalains (modified from Carreón-Hidalgo et al. [45].
Table 2.
Biological properties of betalains (modified from Carreón-Hidalgo et al. [45].
| Biological activity | Mode of action | Reference |
| Antioxidant |
|
[45,63,64,65,66] |
| Anti-inflammatory |
|
[45,67,68,69,70,71,72] |
| Antimicrobial | Betalains exert antimicrobial activity against both Gram-positive and Gram-negative bacteria by targeting microbial cell membranes, similar to phenolic compounds. They alter membrane function and structure while increasing membrane permeability, ultimately leading to microbial cell death | [45,73,74,75,76] |
| Anticancer |
|
[77,78,79,80,81,82,83,84,85,86,87,88,89] |
| Antidiabetic and antilipidemic |
|
[86,90,91,92,93,94,95,96,97,98] |
| Hepatoprotective, neuroprotective, and other effects |
|
[99,100,101,102,103] |
Table 3.
The applications of betalains from different plant species.
| Industry | Product | Plant source | Reference |
| Foods and beverages | Jelly, ice cream, and ice sherbets | B. vulgaris | [104,105,106] |
| Banana juice, fruit spread | Rivina humilis | [107] | |
| Dairy (cow milk) | B. vulgaris, Hylocereus polyrhizus | [108,109] | |
| Yogurt | H. polyrhizus | [110] | |
| Yogurt and cream | Opuntia ficus-indica, O. megacantha | [111] | |
| Jelly gummy and drink | Salicornia fruticosa | [112] | |
| Juice | B. vulgaris | [113] | |
| Biscuits | B. vulgaris | [114] | |
| Candies | B. vulgaris | [115] | |
| Banana spread | Basella rubra | [116] | |
| Beverages, smoothie-like beverages | B. vulgaris | [113,117] | |
| Noodle | Amaranthus tricolor | [118] | |
| Pork meat | - | [64] | |
| Rainbow trout fillets | B. vulgaris | [119] | |
| Food packaging | Furcellaran films | B. vulgaris | [120] |
| Starch/polyvinyl alcohol films | Stenocereus stellatus | [121] | |
| Ammonium chitosan/polyvinyl alcohol films | O. ficus-indica | [122] | |
| Ammonium chitosan films | A. tricolor | [123] | |
| Textiles | Colored wool | O. ficus-indica | [124] |
| Colored wool | B. vugaris | [125] | |
| Bioactive cotton fabrics | B. vugaris | [126] | |
| Betalain-dyed nonwoven cotton fibers | B. vugaris | [127] | |
| Cosmetics and pharmaceuticals | Lipsticks | H. polyrhizus | [128] |
| Facial cosmetic | Amaranthus sp. | [129] | |
| Other applications | Dye-sensitized solar cells | Phytolacca americana, B. vulgaris; Bougainvillea sp. | [130,131,132,133] |
| Betalain-based biosensors | B. vulgaris | [134,135] | |
| Metal coating | B. vulgaris | [136] | |
| Organometallic reductants, stabilizing agents | B. vulgaris | [137] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



