Submitted:
16 April 2025
Posted:
17 April 2025
You are already at the latest version
Abstract
Gut microbiota composition plays a crucial role in host health and may be influenced by age and disease conditions. This study investigated the gut microbiota diversity of 175 dogs across three age groups (Junior (20–46 months), Adult (47–92 months), and Senior (93–168 months), and examined the impact of osteoarthritis on microbial composition. Alpha diversity analysis using the Shannon and Chao1 indices showed no statistically significant differences (P > 0.05) among age groups or between healthy and osteoarthritic dogs, suggesting stable microbial diversity across life stages and health conditions. Beta diversity analysis based on Bray-Curtis dissimilarity indices demonstrated substantial overlap in gut microbiota composition across age groups, with no significant clustering observed (P > 0.05). This indicates that aging and osteoarthritis do not induce significant shifts in microbial beta diversity, although high inter-individual variability was noted. Linear Discriminant Analysis (LDA) Effect Size (LEfSe) analysis identified distinct bacterial taxa associated with different age groups. Linear Discriminant Analysis (LDA) Effect Size (LEfSe) analysis identified distinct bacterial taxa associated with different age groups. Junior dogs exhibited enrichment in Blautia, Erysipelotrichaceae, and Clostridium, while Adult dogs were characterized by higher abundances of Prevotella, Streptococcus, and Ruminococcaceae. Senior dogs had increased representation of Prevotella and Ruminococcus. In osteoarthritic dogs, Peptococcus, Peptostreptococcus, Clostridiaceae, and Coprobacillus were significantly enriched, suggesting potential microbiota shifts associated with osteoarthritis. Overall, these findings indicate that while gut microbiota diversity remains stable across different life stages, specific bacterial taxa were differentially enriched in relation to age and osteoarthritis. This study enhances our understanding of gut microbiota dynamics in dogs and provides insights into potential age- and disease-related microbial signatures.
Keywords:
gut microbiota
; dogs
; aging
; osteoarthritis
; 16S rRNA gene amplicon sequencing
1. Introduction
The gastrointestinal tract harbors a vast collection of microorganisms, collectively known as the gut microbiome. This diverse community is primarily composed of bacteria, alongside commensal fungi, viruses, archaea, and protists [1]. Compared to other colonized organs, the gut contains the highest concentration of microbiota. The intestinal microbiota is estimated to consist of approximately 10¹³ to 10¹⁴ cells [2], with a genome that is 150 times larger than that of humans.
A growing number of studies are exploring the various factors that influence the intestinal microbiota [3,4,5,6]. These factors can be broadly categorized into external and internal influences, such as diet and pharmaceutical treatments for external factors, and age and genetics for internal factors. Moreover, certain pathological conditions, including inflammation and type 2 diabetes, can lead to alterations in the gut microbiota, resulting in an imbalance known as dysbiosis. Among these factors, age is considered one of the most significant variables when examining the development and changes in the gut microbiota over time, which are believed to be linked to age-related diseases [7,8].
The relationship between aging, gut health, and gut microbiota is currently being investigated in mammals, with well-documented findings in humans. Badal et al. [9] reviewed existing knowledge on the human gut microbiota, examining its composition, function, and metabolic products in the context of aging and lifespan. Age-related changes in the human microbiota have been observed and appear to be linked to the host's overall health. In older individuals, microbial diversity tends to decrease, with an increased presence of Bacteroidetes and a reduced abundance of Bifidobacterium, Firmicutes, and Clostridium cluster IV when compared to younger individuals [7,10]. In contrast, Clostridium perfringens, Lactobacilli, Enterococci, and Enterobacteriaceae show an increase with aging [11].
Companion animals, especially dogs, serve as valuable models for studying microbiomes in relation to aging. They have relatively long lifespans and develop age-related health issues under living conditions similar to those of humans. Unlike in humans, Bifidobacterium may not have a significant role in canine gut health. Findings by Masuoka et al. [11] indicated that Bifidobacteria were present in only half of the youngest dogs examined and were absent in adult dogs. However, methodological differences should be taken into account when interpreting these results. Garrigues et al. [12] recently examined the development of gut microbiota in the early stages of canine life, highlighting changes in bacterial communities from day 2 after birth up to 52 weeks. By the second day, microbial richness begins to increase, and between days 2 and 21, the initial dominance of Bacillota shifts to a codominance of Bacteroidota, and Fusobacteriota [13]. During the first few weeks of life, puppies exhibit an immature microbiota, characterized by a higher dysbiosis index (DI), an increased presence of Clostridium difficile, and a lower abundance of Clostridium hiranonis compared to adult dogs [14]. Around 4 to 6 months of age, the microbiota transitions to a composition similar to that of adult dogs and remains relatively stable throughout adulthood [12].
The microbiome has emerged as a compelling factor that may integrate multiple influences, including aging, especially considering recent findings highlighting the presence of a low-grade chronic inflammatory state in osteoarthritis (OA) [15]. Furthermore, it is widely recognized that OA development is influenced by the interaction of three key factors: genetic predisposition, aging, and environmental influences [16]. In dogs, osteoarthritis often begins early in life due to developmental joint conditions such as hip dysplasia. Additionally, joint injuries are another significant cause of osteoarthritis in this species [17,18]. With approximately 9 million pet dogs in the UK [19] and 63.4 million households in the US owning a dog [20], the global burden of OA in dogs is substantial, posing a serious threat to canine welfare. In North America, OA is estimated to affect 20% of dogs over the age of one, based on data from 200 veterinarians [21]. Prevalence estimates for OA in the UK dog population vary significantly, likely due to differences in reporting methods. Estimates range from 2.5% to 6.6% for dogs of any age and breed attending primary-care practices [22,23], with figures rising to 20% for dogs over one year old [24]. Beyond its impact on canine welfare, OA presents significant challenges for veterinarians, owners, and breeders worldwide. Due to its chronic, progressive, and potentially debilitating nature, OA can negatively affect the quality of life of both dogs and their owners [25,26,27]. This condition can also affect owners' well-being, as managing OA in dogs often involves substantial financial costs [28]. Cachon et al. [29] published consensus guidelines for treating canine osteoarthritis (OA) using the COASTeR staging tool, excluding radiography. These recommendations, based on evidence and clinical experience, aim to provide veterinarians with a practical reference for stage-specific OA management. The goal is to support informed treatment decisions and personalized care.
The connection between intestinal microbiota, gut health, and the aging process in older dogs remains largely unexplored. Further research is essential to unravel this complex relationship and to identify potential strategies for enhancing the aging experience in dogs through gut health interventions. This study sets out to examine variations in intestinal microbiota composition and gut health across different age groups of dogs, with particular emphasis on senior dogs with osteoarthritis. Gut microbial markers were analyzed to gain deeper insights into how aging influences the gut environment, ultimately aiming to contribute to the development of targeted approaches for promoting healthier aging in canine populations.
2. Materials and Methods
2.1. Animal Ethics Statement
The Study Facility was committed to complying with all local regulations governing the care and use of laboratory animals. Procedures were designed to prevent or minimize discomfort, distress, and pain to the animals in accordance with the principles of the Animals for Research Act of Ontario and the guidelines of the Canadian Council on Animal Care (CCAC). The CCAC Guide for the Care and Use of Experimental Animals and related policies were regarded as guidelines to follow. To ensure compliance, this protocol was reviewed and approved by the Study Facility’s Institutional Animal Care and Use Committee (IACUC) before the start of the trial, as per IACUC standard operating procedures.
2.2. Animals and Diets
All Beagle dogs were from the Transpharmation Canada Ltd. colony. All dogs from the colony had fecal samples collected unless their study treatment may have interfered with the microbiome or if collection interfered with Sponsored Study objectives.
Dogs were fed using a standard commercial kibble (Purina® ProPlan® All Ages Sport Active 27/17 Chicken & Rice Formula) for the duration of the study. Dogs were individually fed once daily to maintain body condition according to standard operating procedures. Feed was offered in stainless-steel bowls at the end of each day. Water was provided ad libitum (animals could drink water at any time, without restriction) via automatic sippers or stainless-steel bowls. The water source was an on-site well. For the study, 175 Beagle dogs, 98 females and 77 males, were involved int the research and the inclusion criteria were without pharmacological treatments, specifically no NSAIDs, antibiotics, corticosteroids or deworming drugs during the 30 days before the sampling. The medical history and records were available for each dog, as well as body condition score (BCS) and age. None of the dogs presented watery, soft stools or clinical signs of gastrointestinal disease at the time of sampling. Animals were housed in kennels alone or in two individuals, depending on the size of the box. The area for each dog was at least 15 m2. Animals have free access to water and were allowed to have access to an outdoor area for 2 to 4 hours for socialization with other dogs. The diet was a Purina kibble, fed to the amount indicated by the manufacturing, to cover the nutrient requirements of NRC (NRC, 2008)
Table 1 provides an overview of a population of 175 dogs, categorized by age group, sex, and health conditions. The data distinguishes between spayed females (Fs) and castrated males (Mc) and categorizes health conditions into three groups: Healthy, Osteo-arthritis, and other diseases.
Table 2 presents data on a subset of 136 dogs categorized by age group and health condition (Healthy or Osteo-arthritic). The "Other" disease category from Table 1 were not included here, focusing solely on healthy vs. osteo-arthritic dogs. The new groups were Junior healthy (Junior_HE; n=26), Adult healthy (Adult_HE, n=26), Senior healthy (Seinor_HE, n=9), Adult osteo-arthritic (Adult_OA, n=23) and Senior osteo-arthritic (Senior_OA, n=52).
2.3. Data Recording and Samples Collections
Each dog was placed in a metabolic cage or individual pen to facilitate fecal collections. Animals were isolated for collections in the evening after being fed. Each dog was put into individual housing for up to 1 hour following feeding. After 1 hour, the metabolic cage was inspected for a naturally voided sample. If a sample had been produced, it was collected, and the animal was returned to its home pen. The metabolic cage was then cleaned, and another animal was placed into the same metabolic space for overnight collections. If a sample had not been produced, the animal remained in individual housing overnight.
For up to 4 days and 3 nights, the procedure was repeated, and each dog was placed in a metabolic cage or isolated pen until a fecal sample was obtained. Overnight samples were collected the following morning and documented accordingly. Animals returned to their home pens each morning. If fecal samples were not produced, dogs were walked post-feeding to encourage defecation or monitored within their home pens.
Using a gloved hand, the fecal sample was picked up and placed on clean weigh paper. The sample collected had the outer edges removed using a clean plastic knife to minimize bacterial growth. The center of the sample was divided into three aliquots of 250 mg each. Each aliquot was weighed using clean weigh paper that had been tared on the scale and was transferred using a clean plastic knife per sample. Each tube was inverted to ensure the sample was well mixed.
Aliquots were placed into three separate tubes: two ZYMO tubes (brown top) and one eNAT tube (blue top). Tubes were labeled with the animal ID, the date, and the time of fecal collection.
These tubes were stored in a refrigerator at 2–8°C until shipping. Aliquot 1 was shipped to the bioanalytical laboratory at the address below. Aliquots 2 and 3 remained in Transpharmation Canada Ltd. storage until sample analysis.
2.4. Microbiota Analysis
DNA extraction was performed within 6 weeks from the collection date. Total DNA extraction for microbiome analysis was performed on 150 mg of faeces using the Quick-DNA™ Fecal/Soil Microbe Miniprep Kit (Zymo Research, Irvine, CA, USA), following the manufacturer’s instructions. Quantification and quality check of the DNA were carried out using a QubitTM 3 Fluorometer (Thermo Scientific; Waltham, MA, USA). Following DNA extraction, libraries were prepared by amplifying the hypervariable regions V3 and V4 of the 16S rRNA (primers: 341F CCTAYGGGRBGCASCAG and 806R GGACTACNNGGGTATCTAAT) and incorporating sequencing indexes. This step utilized the NEBNextR Ultra™ IIDNA Library Prep Kit (Cat No. E7645), following the manufacturer’s guidelines. The resulting amplicons underwent sequencing on a Novaseq 6000 platform, SP flow cell (Illumina; San Diego, CA, USA) in 2 × 250 paired-end mode, following standard protocols for an intended depth of sequencing of 200,000 reads per sample.
The raw sequence data obtained was deposited in the NCBI Sequence Read Archive under the accession number (PRJNA1246572 for other dieses; PRJNA1247267 for H& OA).
2.5. Bioinformatic
The raw sequences (FASTQ) of the samples were processed using the bioinformatics tool Quantitative Insights into Microbial Ecology 2 (QIIME 2) [30]. The following steps were undertaken: demultiplexing was performed, sequenced reads meeting the quality threshold (Phred score ≥ 30) were identified, denoising with DADA2, and chimeras were filtered out. Overall, a total of 63003270 reads with an average count per sample of 360018 reads (lowest count of 188460 reads, highest 514750reads). These sequences of high-quality were clustered into amplicon sequence variants (ASVs) and annotated against the greengenes database 2022.10.backbone.full-length.nb.qza (https://ftp.microbio.me/greengenes_release/2022.10/) [31].
ASVs were also annotated to the latest 2024.09 backbone.full-length.nb (https://ftp.microbio.me/greengenes_release/current/) [32] classifier to take into account the recent new classification of the International Code of Nomenclature of Prokaryotes (ICNP, https://www.the-icsp.org/index.php/code-of-nomenclatur) [33].
2.6. Statistical Analysis
Dogs were grouped into Junior, Adult and Senior categories (factor Age), considering the range of age: from 20 to 46 (Junior, 43 dogs), from 47 to 92 (Adult, 58 dogs) and more than 92 (Senior, 74 dogs). A preliminary computation included all the 175 dogs (Table 1). Considering that some of the dogs suffered from murmur, diabetes and other non-communicable disease, a second study involved only healthy or arthritic animals. The number of dogs for Junior, Adult and Senior groups accounted for 30, 44 and 76 respectively (Table 2). Arthrosis is a common disorder during ageing and the Adult and Senior dogs were mildly affected and without severe pain, and no NSAIDs were administered during the 4 weeks before faecal sampling.
Reads were assigned to taxa until genus level and the data were uploaded to the Microbiome Analyst (https://www.microbiomeanalyst.ca/, accessed on January 2025) [34] for statistical and integrative analysis of microbiome (Lu et al., 2023) [35] and normalized as (RA) based on total sum. The reads RA were used to compute Shannon and Choa1 alpha diversity indexes and the comparisons for the factors age and sex were tested with Mann–Whitney test, with multi-testing adjustment based on Benjamini–Hochberg procedure (FDR). Bray–Curtis beta diversity differences between ages were also computed, and the results were visualized using Principal Coordinate Analysis (PCoA) plots. Permutational multivariate analysis of variance (PERMANOVA) was applied to evaluate differences in community composition. Linear Discriminant Analysis (LDA) Effect Size (LEfSe) was subsequently applied (Segata et al., 2011) [36] to compare RA between age.
3. Results
3.1. Rarefaction Curve and Relative Abundance of Whole Population
The rarefaction curves show that species richness increases with sequencing depth across all age groups, and Firmicutes were the most abundant taxa, followed by Bacteroidetes (Supplementary Figure S1). The relative abundance (RA) at the phylum level, annotated using the 2022 Greengenes version, indicated a prevalence of Firmicutes and Bacteroidetes, followed by Fusobacteria, Proteobacteria, Actinobacteria, Tenericutes, and Deferribacteres (Supplementary Figure S2-A). Using the latest version of the greengenes classifier from 2024, the ranking of phyla was Bacillota_A, Bacteroidota, Fusobacteriota, Bacillota_I, Pseudomonadota, Bacillota_C, Actinomycetota, and Deferribacteriota (Supplementary Figure S2-B). However, considering that most of the data still refers to the previous classification, for comparison purpose the annotations of 2022 greengene versions were used.
3.2. Alpha Diversity
Figure 1 presents the Shannon and Chao1 alpha diversity indices of the gut microbiota in the entire dog population (n = 175), categorized into Junior, Adult, and Senior groups. The boxplot illustrates the distribution of alpha diversity within each age group, highlighting median values, interquartile ranges, and individual data points. Significantly higher values (P < 0.05) were observed for the Shannon index in the Senior group compared to the Adult group.
The alpha diversity of the gut microbiota, assessed using the Shannon index and Chao1 index, was also analyzed to compare microbial diversity across different age groups and health conditions (Junior_HE, Adult_HE, Senior_HE, Adult_OA, and Senior_OA). As shown in Figure 2, variations in microbial diversity were observed among groups; however, no significant differences (P > 0.05) were detected for either the Shannon index or the Chao1 index. Despite this, the spread of values, indicated by the interquartile range and whiskers, showed high variability within each group. These results suggest that alpha diversity remains relatively stable across different age groups and health conditions in the studied dog population.
3.3. Beta Diversity
The beta diversity of the gut microbiota across different age groups of dogs was analysed using the Bray-Curtis dissimilarity index and visualized through a Principal Coordinate Analysis (PCoA) plot (Figure 3). The distribution of data points representing Junior, Adult, and Senior dogs does not indicate distinct clustering among the age groups (P > 0.05). Instead, there is considerable overlap, suggesting that the gut microbiota composition remains relatively similar across different life stages.
The Bray-Curtis beta diversity index was also used to assess differences in gut microbiota composition among Junior_HE, Adult_HE, Senior_HE, Adult_OA, and Senior_OA dogs, and no significant differences (P > 0.05) were detected (Figure 4). The distribution of points did not form distinct clusters, indicating substantial overlap in gut microbiota composition across all groups. Some outliers, represented by points positioned far from the main cluster, suggest that certain individuals have unique microbial compositions. The lack of significant differences implies that despite aging and osteoarthritis, gut microbiota remains relatively stable in terms of beta diversity.
3.4. LEfSe Analysis
Figure 5 presents a LEfSe analysis identifying bacterial taxa that were significantly different across the three age classes (Junior, Adult, and Senior) and in the Supplementary Table S1 the comparison of abundances between groups is reported. The LDA score (x-axis) quantifies the effect size of each significantly enriched bacterial taxon. The bacterial features (y-axis) represent taxa with an LDA score >2 and an FDR <0.10, indicating statistical significance. The results show that different bacterial taxa were associated with specific age groups. In the Adult group, the taxa S24_7, Ruminococcaceae, Mogibacteriaceae, Streptococcus, Peptococcus, Bacteroidales and Oscillospira significantly differed from the other groups.
In the Junior group, Blautia, Erysipelotrichaceae, Clostridium and Lachnospiraceae were significantly enriched. For the Senior group, Prevotella and Ruminococcus were identified as the most enriched taxa. The bar chart in Figure 6 represents the LDA of bacterial taxa that significantly differs (FDR < 0.10; LDA score >2) across Junior_HE, Adult_HE, Senior_HE, Adult_OA, and Senior_OA dogs. In the Supplementary Table S2 the comparison of abundances between groups is reported. Junior_HE (healthy young dogs, red) was notably associated with Erysipelotrichaceae and Mucispirillum. Adult_HE (healthy adult dogs, blue) had a higher RA of Prevotella, Bacteroidales, Phascolarctobacterium, Lachnospiraceae, Bacteroidales and Coprococcus. Adult_OA (osteoarthritic adult dogs, green) showed significant associations with Peptococcus and Peptostreptococcus, indicating a shift in microbiota linked to osteoarthritis in adult dogs. Senior_HE (healthy senior dogs, purple) was dominated by Fusobacteriaceae and Eubacterium, and Senior_OA (osteoarthritic senior dogs, orange) had a higher abundance of Clostridiaceae, Coprobacillus, and Bacillus.
4. Discussion
The composition of gut microbiota is essential for host health and is influenced by factors such as age and disease conditions. This study examined gut microbiota diversity in dogs across different life stages (Junior, Adult, and Senior) and health conditions (healthy vs. osteoarthritic).
4.1. Microbial Diversity
Shannon and Chao1 alpha diversity indices showed high individual variability across all age classes (Figure 1), with only the Adult dogs exhibiting a higher mean value for the Shannon index than the Senior dogs (P<0.05). Mizukami et al. [37] reported a moderate decline in the alpha diversity of the gut microbiome with age, with both the Chao1 and Shannon indices showing a decreasing trend, although the changes were not statistically significant. However, they found that the Faith-PD index significantly decreased, indicating reduced phylogenetic diversity in aging dogs. Similarly, another study on the canine gut microbiome by Omatsu et al. [38] also reported a decrease in alpha diversity in aged dogs. In contrast, Fernández-Pinteño et al. [39] did not find notable differences in alpha diversity measures or the fecal dysbiosis index across age groups. The significant variation of the Shannon index and the stability of the Chao1 index observed in this study (Figure 1) could be attributed to the standardized dietary and environmental conditions of the dog population. Also, the comparison of healthy and OA dogs across the three age classes (Figure 2), after the exclusion of dogs with other pathologies, revealed a substantial stability of the bacterial community. Stevens et al. [40] did not find significant differences in the Shannon diversity index between healthy and OA-pain dogs. Indeed, Cintio et al. [41] reported that the gut microbiota of OA dogs was significantly less diverse at the family taxonomic level compared to healthy dogs. The dogs in this latter study were client-owned and were fed the same diet for 45 days, conditions that differed from study of Stevens et al. [40], where privately owned dogs were fed different diets.
The lack of variation of beta diversity among Junior, Adult, and Senior dogs (Figure 3) suggested that gut microbiota composition remains relatively stable throughout life. However, Fernández-Pinteño et al. [39] and Mizukami et al. [37] found a significant difference in beta diversity with age when using the unweighted UniFrac metric, but not with the weighted UniFrac metric or Bray-Curtis distances. A limited number of studies reported the effect of OA on gut microbiomes in dogs. Stevens et al. [40] did not find variations of beta diversity between healthy and OA dogs, supporting the results obtained (Figure 4), and suggesting that the disease does not induce major shifts in gut microbial communities. Indeed, Cintio et al. [41] observed significant differences of beta diversity at the Family level between healthy and OA dogs. Studies investigating the effect of OA on the gut microbiome of dogs are still limited and warrant further investigation.
4.2. Microbial Signatures Associated with Age and OA
LEfSe analysis identified distinct microbial taxa enriched across the age groups, indicating age-related changes in gut microbiota composition (Figure 5). In Junior dogs, a higher abundance of Lachnospiraceae [39] and Blautia was reported compared to Adult and Senior dogs [42]. This agrees with the present results, suggesting a healthy gut microbiota [43,44].
An increase in Bacteroidales in Adult dogs could indicate immune homeostasis in the gut, as they are involved in carbohydrate degradation and the production of short-chain fatty acids [45]. The diverse family Erysipelotrichaceae, more abundant in Junior dogs, has been correlated with dietary carbohydrate and fiber digestion as well as short-chain fatty acid production [46]. This family has also been associated with healthier microbiota in dogs compared to those affected by enteropathies [44,47].
Fernández-Pinteño et al. [39] observed that S24-7 did not vary significantly with age but was found in higher abundance in Adult dogs. Senior dogs, on the other hand, showed higher levels of Prevotella and Ruminococcus, suggesting microbiota adaptations linked to the aging process. This is consistent with known microbial community shifts that occur as dogs age [39]. However, the variation in relative abundances (RAs) of other taxa did not align with the present study.
Mizukami et al. [37] found no significant correlation between aging and microbial changes at the genus or phylum levels. However, they identified a single ASV that showed a significant positive correlation with aging, suggesting the possibility of more subtle age-related shifts in the microbiome. This is in line with findings in humans, where certain microbes like Absiella dolichum have been found to be elevated in frail individuals [48] and lower in those on a high-fiber diet [49].
4.3. Microbiota and Osteoarthritis
When the data were analyzed separately by age class in healthy and OA dogs, higher relative abundances (RAs) of Erysipelotrichaceae and Bacteroidales were confirmed in Junior and Adult dogs, respectively (Figure 6). Interestingly, Fusobacteriaceae was more abundant in healthy Senior_HE dogs. You and Kim, [50] also reported that the Fusobacterium genus was more abundant in older dogs (6–10 years) compared to younger dogs (0.5–1 year).
For healthy dogs (Junior_HE, Adult_HE, Senior_HE), the taxa that significantly changed did not correspond with those presented in Figure 5, suggesting that OA may influence the gut microbiome, potentially through inflammation-mediated changes. Notably, none of the Junior dogs were affected by OA, and the differences in taxa that appeared significantly higher in the analysis of the entire dataset may be attributed to statistical factors.
Research by Cintio et al. [41] found that OA dogs had an elevated relative abundance (RA) of the Megamonas genus and reduced levels of the Paraprevotellaceae, Porphyromonadaceae, and Mogibacteriaceae families compared to healthy dogs, indicating that dysbiosis or a disruption of the normal gut microbial balance could occur. In the study by Stevens et al. [40], dogs with OA showed higher representations of specific species, including Bacteroides vulgatus, Eubacterium dolichum, Collinsella stercoris, Clostridium ramosum, and Ruminococcus torques. The study did not report notable differences in the Firmicutes to Bacteroidetes ratio or in factors such as pain severity, mobility, activity level, age, or body composition score.
In the present study, the gut microbiota of Adult_OA dogs showed increased levels of Peptococcus and Peptostreptococcus, while that of Senior_OA dogs showed increased levels of Clostridiaceae, Coprobacillus, and Bacillus (Figure 6).
The role of specific microbial taxa in OA has also been investigated in human studies. Boer et al. [51] identified a significant association between Streptococcus species abundance in stool and OA-related knee pain and inflammation. Similarly, Yu et al. [52] suggested that specific microbial families and orders, such as Methanobacteriaceae and Desulfovibrionales, might play a causal role in OA development.
Despite the lack of strong evidence linking gut dysbiosis to OA in dogs, Stevens et al. [40] suggest that subtle changes in the gut microbiota could influence microbial product translocation and intestinal permeability. Elevated levels of lipopolysaccharide (LPS) and LPS-binding protein (LBP) are associated with increased OA severity in humans, and preliminary data from the same group suggests a positive correlation between LBP levels and the number of joints affected by OA-related pain in dogs. This indicates that intestinal permeability may play a critical role in OA progression and warrants further investigation [53].
Limitations in sample size and the imbalance between healthy and OA-pain groups, along with the lack of measurements for markers of intestinal permeability (e.g., proinflammatory cytokines or LPS), suggest that further research is needed to better understand the role of microbiota in OA.
5. Conclusions
Our study confirmed that the gut microbiota undergoes distinct age-related changes in dogs, with different taxa becoming enriched or depleted across life stages, even though our results did not always align with those reported in the literature. While age influenced specific bacterial taxa, the overall microbial composition remained largely unchanged. OA appeared to induce microbial shifts in Adult and Senior dogs but did not significantly alter global diversity. These findings emphasize the need to explore the complex dynamics of microbial communities in aging dogs, particularly in relation to health conditions such as OA. Further studies with larger sample sizes and more diverse breeds are warranted to confirm these observations and gain a deeper understanding of the microbiome’s role in aging and disease.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Rarefaction curves at the phylum level of gut microbiota of the 175 dogs Junior, Adult and Senior based on age; Figure S2: Relative abundances at the phylum level of gut microbiota of the 175 dogs grouped in Junior, Adult and Senior based on age; Figure S3: Result of Linear discriminant analysis (LDA) effect Size of taxa which differ (FDR<0.01; LDA score >2) between age in the dog population (n=175); Figure S4: Result of Linear discriminant analysis (LDA) effect Size of taxa which differ (FDR<0.01; LDA score >2) between age in healthy and osteoarthritic dog population (n=136).Table S1: Result of Linear discriminant analysis (LDA) effect Size of taxa which differ (FDR <0.10; LDA score >2) between age in the dog population (n=175);Table S2: Result of Linear discriminant analysis (LDA) effect Size of taxa which differ (FDR <0.10; LDA score >2) between age between age in healthy and osteoarthritic dog population (n=136).
Author Contributions
F.B.: Data curation, Formal analysis, Writing—original draft, and Writing, review and editing; Ch. R.: Conceptualization, supervision and Writing—review& editing; A.P.: methodology, investigation, validation and resources ;B.S.: Funding acquisition, Conceptualization, supervision, project administration, Data curation, Formal analysis, Methodology, Writing—original draft and Writing—review& editing; S. K.: methodology, investigation, validation and resources; N.M.: methodology, investigation, validation and resources; P.M.: Conceptualization and Funding acquisition.
Funding
This work was supported by University of Udine, Special grant for Collaborative Research, ex DM 737/2021, Year 2023-2025.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The raw sequence data obtained was deposited in the NCBI Sequence Read Archive under the accession number PRJNA1246572 & PRJNA1247267.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Matijaši´c, M.; Meštrovi´c, T.; Paljetak, H.C.; Peri´c, M.; Bareši´c, A.; Verbanac, D. Gut microbiota beyond bacteria mycobiome, virome, archaeome, and eukaryotic parasites in IBD. Int J Mol Sci. 2020, 21, 2668. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.R.; Pop, M.; DeBoy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; et al. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–9. [Google Scholar] [CrossRef] [PubMed]
- Bajinka, O.; Darboe, A.; Tan, Y.; Abdelhalim, K.A.; Cham, L.B. Gut microbiota and the human gut physiological changes. Ann Microbiol. 2020, 70, 65. [Google Scholar] [CrossRef]
- Wen, L.; Duffy, A. Factors Influencing the gut microbiota, inflammation, and type 2 diabetes. J Nutrition. 2017, 147, 1468S–75S. [Google Scholar] [CrossRef]
- Hasan, N.; Yang, H. Factors affecting the composition of the gut microbiota, and its modulation. PeerJ. 2019, 7, e7502. [Google Scholar] [CrossRef] [PubMed]
- Pilla, R.; Suchodolski, J.S. The gut microbiome of dogs and cats, and the influence of diet. Vet Clin North Am Small Animal Pract. 2021, 51, 605–21. [Google Scholar] [CrossRef]
- Saraswati, S.; Sitaraman, R. Aging and the human gut microbiota from correlation to causality. Front Microbiol 2014, 5, 764. [Google Scholar]
- Biragyn, A.; Ferrucci, L. Gut dysbiosis: a potential link between increased cancer risk in ageing and inflammaging. Lancet Oncol 2018, 19, e295–304. [Google Scholar] [CrossRef]
- Badal, V.D.; Vaccariello, E.D.; Murray, E.R.; Yu, K.E.; Knight, R.; Jeste, D.V.; et al. The gut microbiome, aging, and longevity: a systematic review. Nutrients. 2020, 12, 3759. [Google Scholar] [CrossRef]
- Kumar, M.; Babaei, P.; Ji, B.; et al. Human gut microbiota and healthy aging: Recent developments and future prospective. Nutr Healthy Aging 2016, 4, 3–16. [Google Scholar] [CrossRef]
- Masuoka, H.; Shimada, K.; Kiyosue-Yasuda, T.; Kiyosue, M.; Oishi, Y.; Kimura, S.; et al. Transition of the intestinal microbiota of dogs with age. Biosci Microbiota Food Heal. 2016, 36, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Garrigues, Q.; Apper, E.; Chastant, S.; Mila, H. Gut microbiota development in the growing dog: a dynamic process influenced by maternal, environmental and host factors. Front Vet Sci. 2022, 9, 964649. [Google Scholar] [CrossRef]
- Guard, B.C.; Mila, H.; Steiner, J.M.; Mariani, C.; Suchodolski, J.S.; Chastant-Maillard, S. Characterization of the fecal microbiome during neonatal and early pediatric development in puppies. PLoS ONE. 2017, 12, e0175718. [Google Scholar] [CrossRef] [PubMed]
- Blake, A.B.; Cigarroa, A.; Klein, H.L.; Khattab, M.R.; Keating, T.; Coevering, P.V.D.; et al. Developmental stages in microbiota, bile acids, and clostridial species in healthy puppies. J Vet Intern Med. 2020, 34, 2345–56. [Google Scholar] [CrossRef] [PubMed]
- Scanzello, C.R.; Plaas, A.; Crow, M.K. Innate immune system activation in osteoarthritis: is osteoarthritis a chronic wound? Curr Opin Rheumatol. 2008, 20, 565–72. [Google Scholar] [CrossRef]
- Sandell, L.J. Etiology of osteoarthritis: genetics and synovial joint development. Nat Rev Rheumatol. 2012, 8, 77–89. [Google Scholar] [CrossRef]
- Fox, S.M. Pathophysiology of osteoarthritic pain. In:Northcott J, editor. Chronic Pain in Small Animal Medicine. London: Manson Publishing Ltd. 2011, 74–96.
- Pet Population PFMA. Avalable online at: https://www.pfma.org.uk/pet-population-2018 (accessed July 7, 2018).
- APPA 2019-2020 APPA National Pet Owners Survey. American Pet Products Association; 2019.
- Johnston, S.A. Osteoarthritis. Joint anatomy, physiology, and pathobiology. Vet Clin North Am Small Anim Pract. 1997, 27, 699–723. [Google Scholar] [CrossRef]
- O'Neill, D.G.; Church, D.B.; McGreevy, P.D.; Thomson, P.C.; Brodbelt, D.C. Prevalence of disorders recorded in dogs attending primary-care veterinary practices in England. PLoS ONE. 2014, 9, e90501–10.1371. [Google Scholar] [CrossRef]
- Anderson, K.L.; O'Neill, D.G.; Brodbelt, D.C.; Church, D.B.; Meeson, R.L.; Sargan, D.; et al. Prevalence, duration and risk factors for appendicular osteoarthritis in a UK dog population under primary veterinary care. Sci Rep. 2018, 8, 5641–10.1038. [Google Scholar] [CrossRef]
- Clements, D.N.; Carter, S.D.; Innes, J.F.; Ollier, W.E. Genetic basis of secondary osteoarthritis in dogs with joint dysplasia. Am J Vet Res. 2006, 67, 909–18. [Google Scholar] [CrossRef]
- Belshaw, Z.; Dean, R.; Asher, L. “You can be blind because of loving them so much”: The impact on owners in the United Kingdom of living with a dog with osteoarthritis. BMC Vet. Res. 2020, 16, 190. [Google Scholar] [CrossRef] [PubMed]
- Belshaw, Z.; Dean, R.; Asher, L. Slower, shorter, sadder: A qualitative study exploring how dog walks change when the canine participant develops osteoarthritis. BMC Vet. Res. 2020, 16, 85. [Google Scholar] [CrossRef] [PubMed]
- Gore, M.; Lana, S.E.; Bishop, G.A. Colorado State University, Pet Hospice Program. Vet. Clin. N. Am. Small Anim. Pract. 2019, 49, 339–349. [Google Scholar] [CrossRef]
- Wilke, V.; Robinson, D.; Evans, R.; Rothschild, M.; Conzemius, M. Estimate of the annual economic impact of treatment of cranial cruciate ligament injury in dogs in the United States. J Am Vet Med Assoc. 2005, 227, 1604–7. [Google Scholar] [CrossRef]
- Cachon, T.; Frykman, O.; Innes, J.F.; Lascelles, B.D.X.; Okumura, M.; Sousa, P.; Staffieri, F.; Steagall, P.V.; Van Ryssen, B. COAST Development Group's international consensus guidelines for the treatment of canine osteoarthritis. Front Vet Sci. 2023, 10, 1137888. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A. , et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol. 2019, 37, 852–7. [Google Scholar] [CrossRef]
- (https://ftp.microbio.me/greengenes_release/2022.10/).
- (https://ftp.microbio.
- (ICNP, https://www.the-icsp.org/index.
- (https://www.microbiomeanalyst.ca/, accessed on January 2025).
- Lu, Y.; Zhou, G.; Ewald, J.; Pang, Z.; Shiri, T.; Xia, J. MicrobiomeAnalyst 2.0: comprehensive statistical, functional and integrative analysis of microbiome data. Nucleic Acids Res 2023, 51, W310–W318. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol 2011, 12, (6)-R60. [Google Scholar] [CrossRef]
- Mizukami, K.; Uchiyama, J.; Igarashi, H.; Murakami, H.; Osumi, T.; Shima, A.; Ishiahra, G.; Nasukawa, T.; Une, Y.; Sakaguchi, M. Age-related analysis of the gut microbiome in a purebred dog colony. FEMS Microbiol Lett. 2019, 366, fnz095. [Google Scholar] [CrossRef] [PubMed]
- Omatsu, T.; Omura, M.; Katayama, Y.; et al. Molecular diversity of the faecal microbiota of Toy Poodles in Japan. J Vet Med Sci. 2018, 80, 749–54. [Google Scholar] [CrossRef]
- Fernández-Pinteño, A.; Pilla, R.; Manteca, X.; Suchodolski, J.; Torre, C.; Salas-Mani, A. Age-associated changes in intestinal health biomarkers in dogs. Front Vet Sci. 2023, 10, 1213287. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Stevens, C.; Norris, S.; Arbeeva, L.; Carter, S.; Enomoto, M.; Nelson, A.E.; Lascelles, B.D.X. Gut Microbiome and Osteoarthritis: Insights From the Naturally Occurring Canine Model of Osteoarthritis. Arthritis Rheumatol. 2024, 76, 1758–1763. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cintio, M.; Scarsella, E.; Sgorlon, S.; Sandri, M.; Stefanon, B. Gut Microbiome of Healthy and Arthritic Dogs. Vet Sci. 2020, 7, 92. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fernández-Pinteño, A.; Pilla, R.; Suchodolski, J.; Apper, E.; Torre, C.; Salas-Mani, A.; Manteca, X. Age-Related Changes in Gut Health and Behavioral Biomarkers in a Beagle Dog Population. Animals (Basel). 2025, 15, 234. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Honneffer, J.B.; Minamoto, Y.; Suchodolski, J.S. Microbiota alterations in acute and chronic gastrointestinal inflammation of cats and dogs. World J Gastroenterol. 2014, 20, 16489–97. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Suchodolski, J.S.; Dowd, S.E.; Wilke, V.; Steiner, J.M.; Jergens, A.E. 16S rRNA gene pyrosequencing reveals bacterial dysbiosis in the duodenum of dogs with idiopathic inflammatory bowel disease. PLoS ONE. 2012, 7, e39333. [Google Scholar] [CrossRef]
- Lapébie, P.; Lombard, V.; Drula, E.; Terrapon, N.; Henrissat, B. Bacteroidetes use thousands of enzyme combinations to break down glycans. Nat. Commun. 2019, 10, 2043. [Google Scholar] [CrossRef]
- Bermingham, E.N.; Maclean, P.; Thomas, D.G.; Cave, N.J.; Young, W. Key bacterial families (Clostridiaceae, Erysipelotrichaceae and Bacteroidaceae) are related to the digestion of protein and energy in dogs. PeerJ. 2017, 5, e3019. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Vázquez-Baeza, Y.; Hyde, E.; Suchodolski, J.; et al. Dog and human inflammatory bowel disease rely on overlapping yet distinct dysbiosis networks. Nat Microbiol 2016, 1, 16177. [Google Scholar] [CrossRef]
- Jackson, M.A.; Jeffery, I.B.; Beaumont, M.; et al. Signatures of early frailty in the gut microbiota. Genome Med 2016, 8, 8. [Google Scholar] [CrossRef]
- Lin, D.; Peters, B.A.; Friedlander, C.; et al. Association of dietary fibre intake and gut microbiota in adults. Br J Nutr 2018, 120, 1014–22. [Google Scholar] [CrossRef] [PubMed]
- You, I.; Kim, M.J. Comparison of Gut Microbiota of 96 Healthy Dogs by Individual Traits: Breed, Age, and Body Condition Score. Animals 2021, 11, 2432. [Google Scholar] [CrossRef] [PubMed]
- Boer, C.G.; Radjabzadeh, D.; Medina-Gomez, C.; et al. Intestinal microbiome composition and its relation to joint pain and inflammation. Nat Commun 2019, 10, 4881. [Google Scholar] [CrossRef]
- Yu, X.H.; Yang, Y.Q.; Cao, R.R.; et al. The causal role of gut microbiota in development of osteoarthritis. Osteoarthritis Cartilage 2021, 29, 1741–1750. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Chen, J.; Li, B.; et al. Faecal microbiota transplantation from metabolically compromised human donors accelerates osteoarthritis in mice. Ann Rheum Dis 2020, 79, 646–656. [Google Scholar] [CrossRef]
Figure 1.
Alpha diversity index of Shannon and Chao1 of gut microbiota of whole dog population (n=175) grouped in Junior, Adult and Senior based on age. Shannon index significantly differed between Adult and Senior for P<0.05. Junior: dog with age ranging 20–46 months; Adult dog with age ranging 47–92 months; Senior: dogs with age ranging 93–168 months.
Figure 1.
Alpha diversity index of Shannon and Chao1 of gut microbiota of whole dog population (n=175) grouped in Junior, Adult and Senior based on age. Shannon index significantly differed between Adult and Senior for P<0.05. Junior: dog with age ranging 20–46 months; Adult dog with age ranging 47–92 months; Senior: dogs with age ranging 93–168 months.
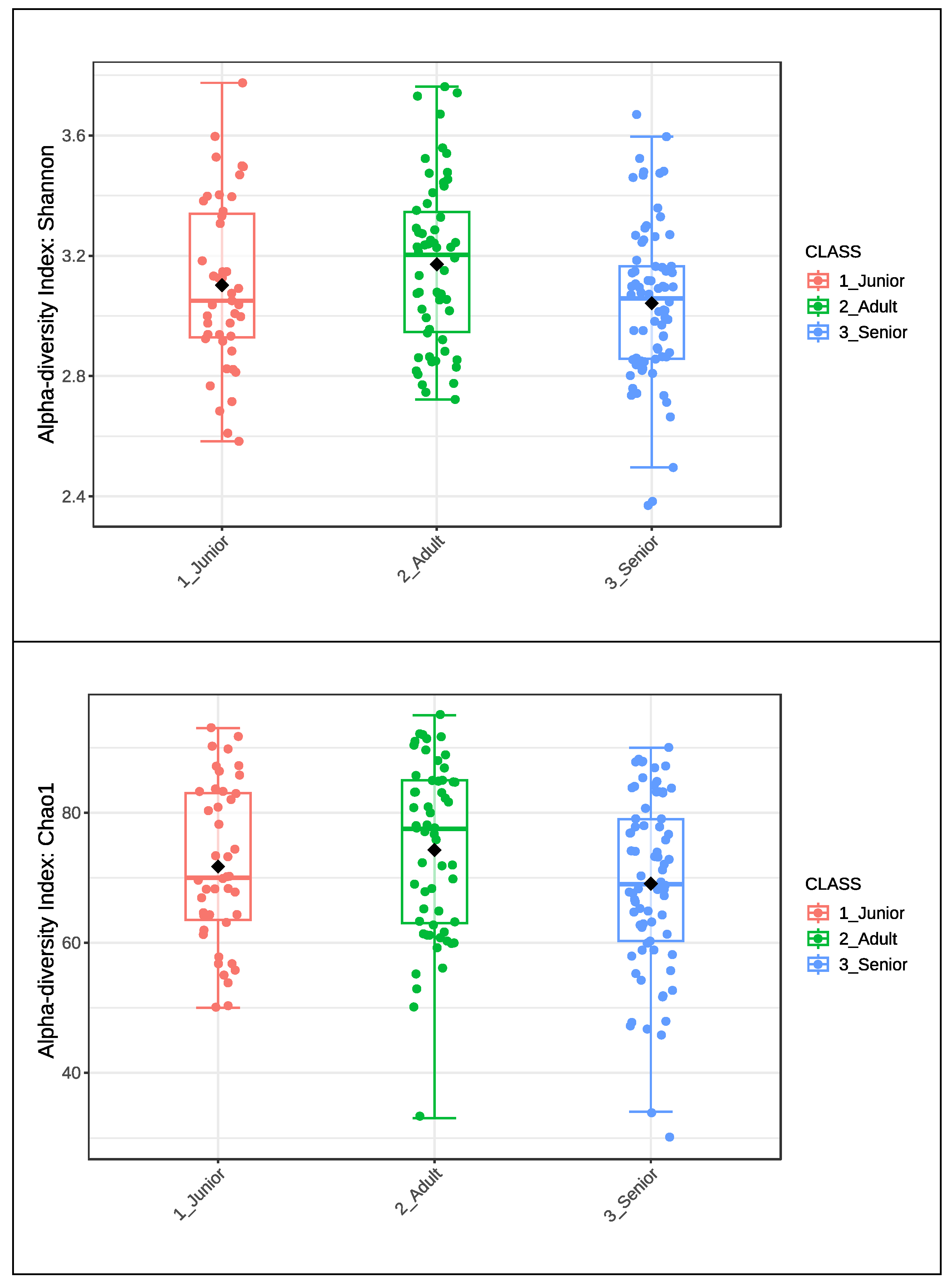
Figure 2.
Alpha diversity index of Shannon and Chao1 of gut microbiota of selected healthy and osteoarthritic dogs grouped in Junior, Adult and Senior based on age. Groups did not significantly differ for P<0.05. Junior_HE: healthy dog with age ranging 20–46 months; Adult_HE: healthy dog with age ranging 47–92 months; Adult_OA: osteoarthritic dogs with age ranging 47–92 months; Senior_HE: healthy dogs with age ranging 93–168 months; Senior_OA: osteoarthritic dogs with age ranging 93–168 months.
Figure 2.
Alpha diversity index of Shannon and Chao1 of gut microbiota of selected healthy and osteoarthritic dogs grouped in Junior, Adult and Senior based on age. Groups did not significantly differ for P<0.05. Junior_HE: healthy dog with age ranging 20–46 months; Adult_HE: healthy dog with age ranging 47–92 months; Adult_OA: osteoarthritic dogs with age ranging 47–92 months; Senior_HE: healthy dogs with age ranging 93–168 months; Senior_OA: osteoarthritic dogs with age ranging 93–168 months.
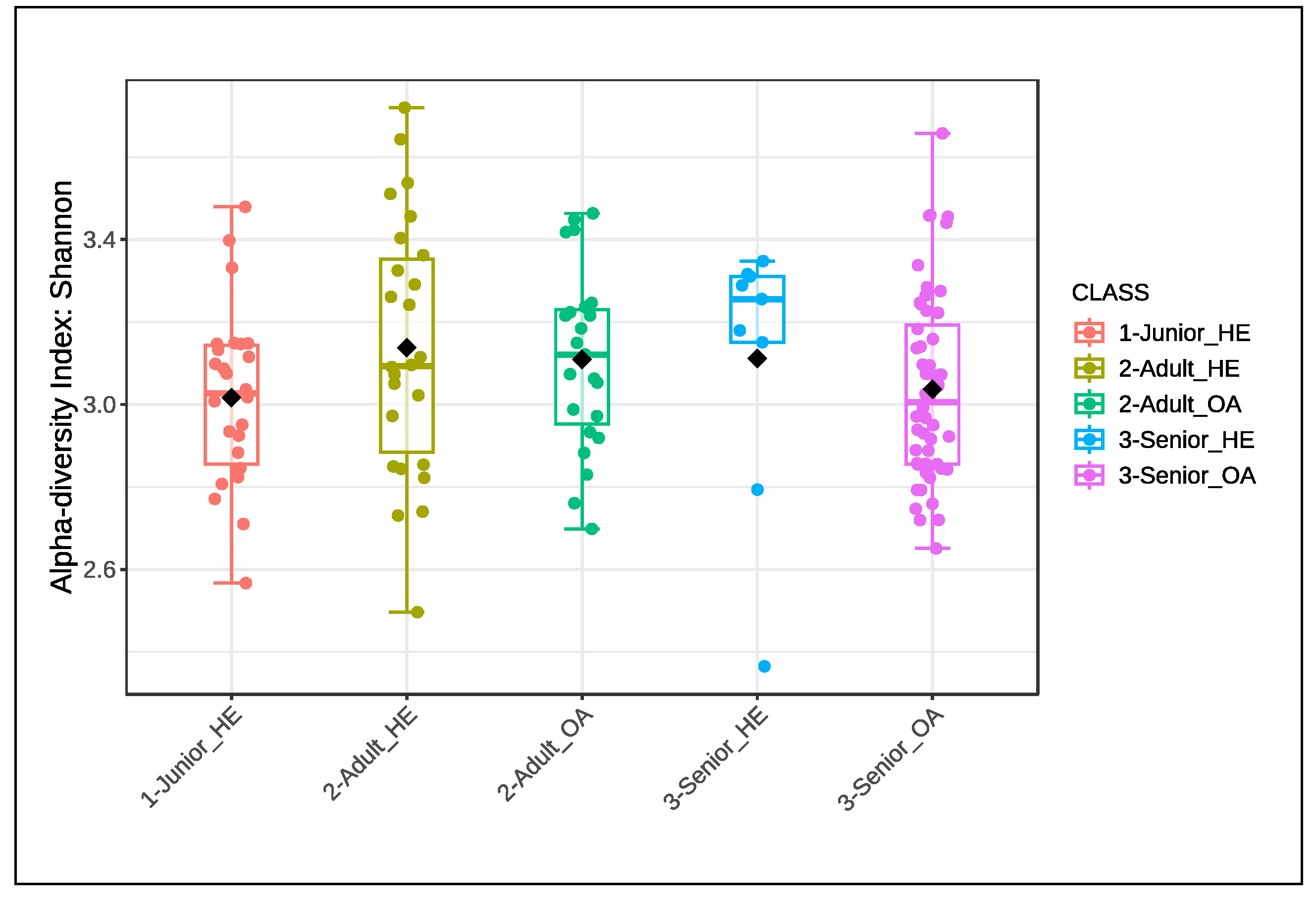
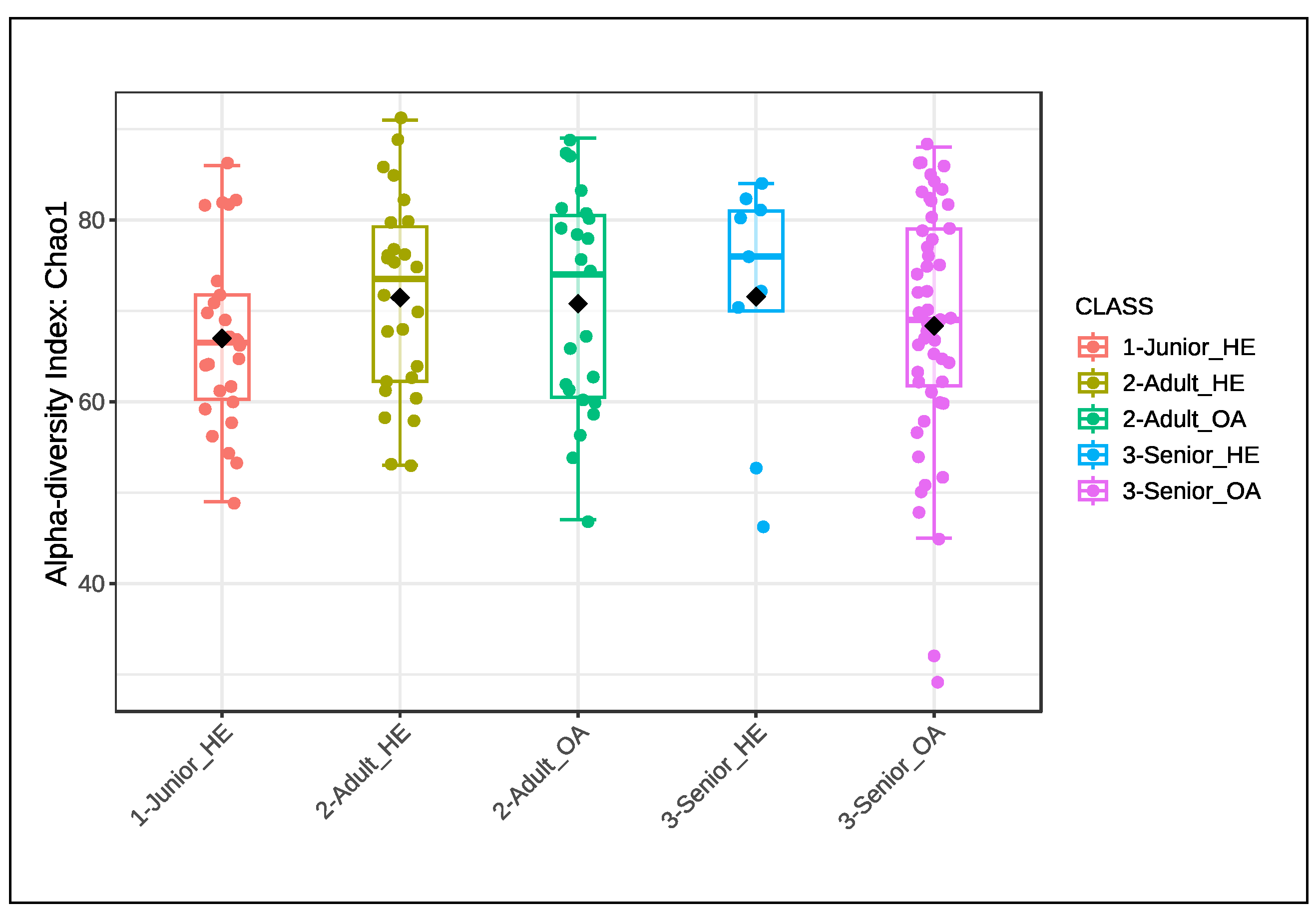
Figure 3.
Bray-Curtis beta diversity index of gut microbiota of dog population (n=175) grouped in Junior, Adult and Senior on the basis of age. Group of ages did not significantly differ for P<0.05 at the PERMANOVA analysis. Junior: dog with age ranging 20–46 months; Adult dog with age ranging 47–92 months; Senior: dogs with age ranging 93–168 months.
Figure 3.
Bray-Curtis beta diversity index of gut microbiota of dog population (n=175) grouped in Junior, Adult and Senior on the basis of age. Group of ages did not significantly differ for P<0.05 at the PERMANOVA analysis. Junior: dog with age ranging 20–46 months; Adult dog with age ranging 47–92 months; Senior: dogs with age ranging 93–168 months.
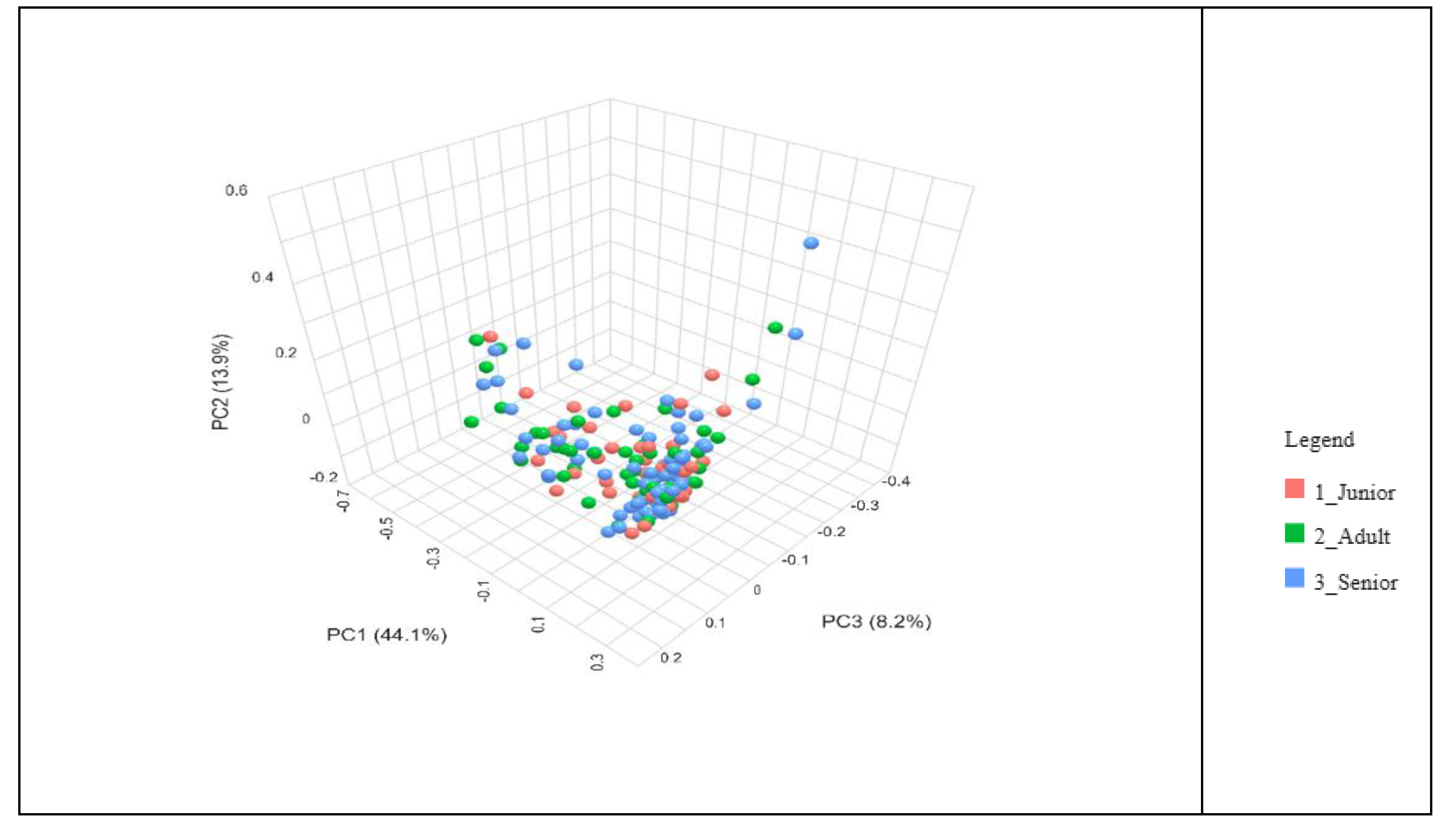
Figure 4.
Bray-Curtis beta diversity index of gut microbiota of health and osteoarthritic dogs (n=136) grouped in Junior, Adult and Senior on the basis of age and healthy conditions. Groups did not significantly differ for P<0.05 at the PERMANOVA analysis. Junior_HE: healthy dog with age ranging 20–46 months; Adult_HE: healthy dog with age ranging 47–92 months; Adult_OA: osteoarthritic dogs with age ranging 47–92 months; Senior_HE: healthy dogs with age ranging 93–168 months; Senior_OA: osteoarthritic dogs with age ranging 93–168 months.
Figure 4.
Bray-Curtis beta diversity index of gut microbiota of health and osteoarthritic dogs (n=136) grouped in Junior, Adult and Senior on the basis of age and healthy conditions. Groups did not significantly differ for P<0.05 at the PERMANOVA analysis. Junior_HE: healthy dog with age ranging 20–46 months; Adult_HE: healthy dog with age ranging 47–92 months; Adult_OA: osteoarthritic dogs with age ranging 47–92 months; Senior_HE: healthy dogs with age ranging 93–168 months; Senior_OA: osteoarthritic dogs with age ranging 93–168 months.
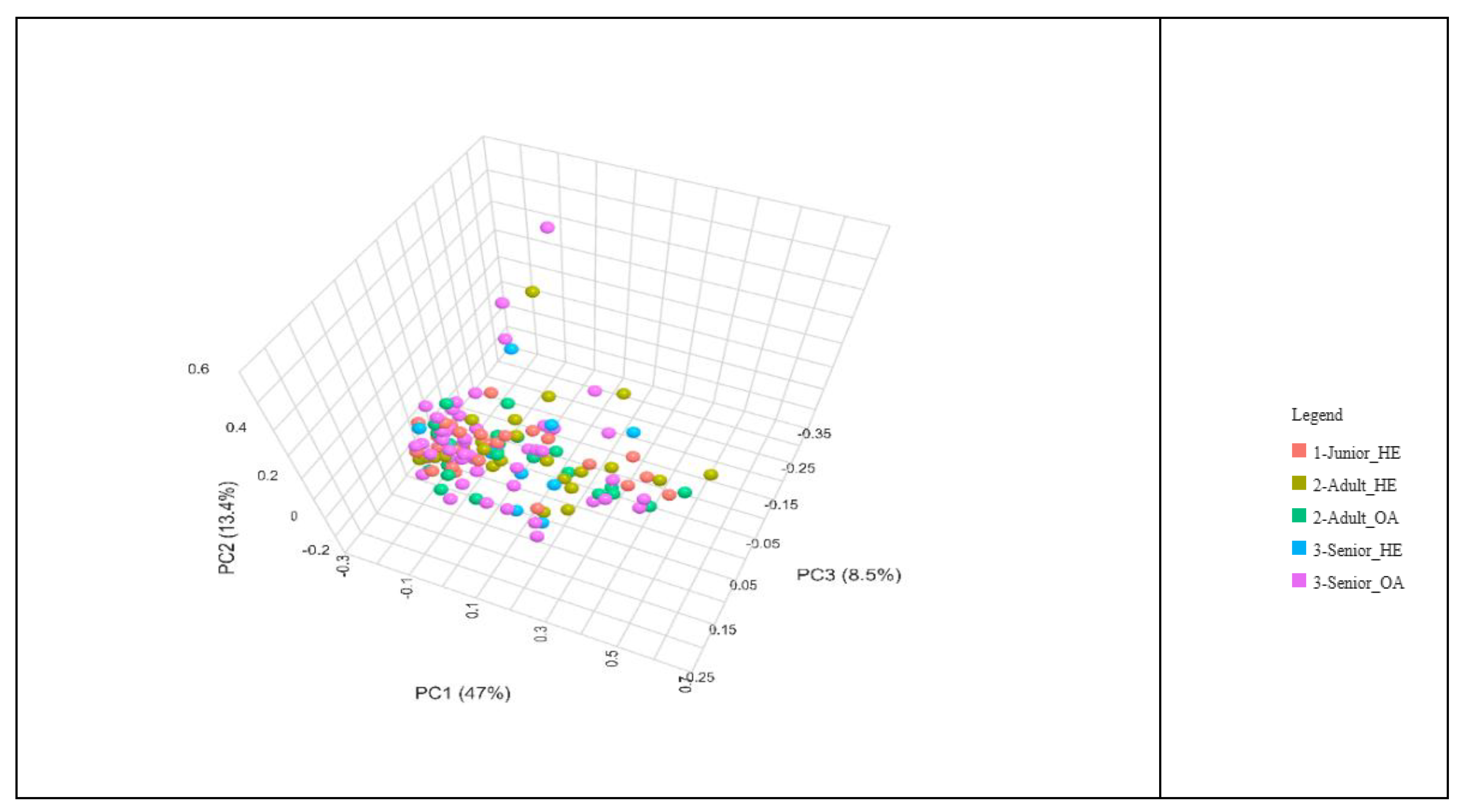
Figure 5.
Result of Linear discriminant analysis (LDA) effect Size of taxa which differ (FDR <0.10; LDA score >2) between age in the dog population (n=175). Junior: dog with age ranging 20–46 months; Adult dog with age ranging 47–92 months; Senior: dogs with age ranging 93–168 months.
Figure 5.
Result of Linear discriminant analysis (LDA) effect Size of taxa which differ (FDR <0.10; LDA score >2) between age in the dog population (n=175). Junior: dog with age ranging 20–46 months; Adult dog with age ranging 47–92 months; Senior: dogs with age ranging 93–168 months.
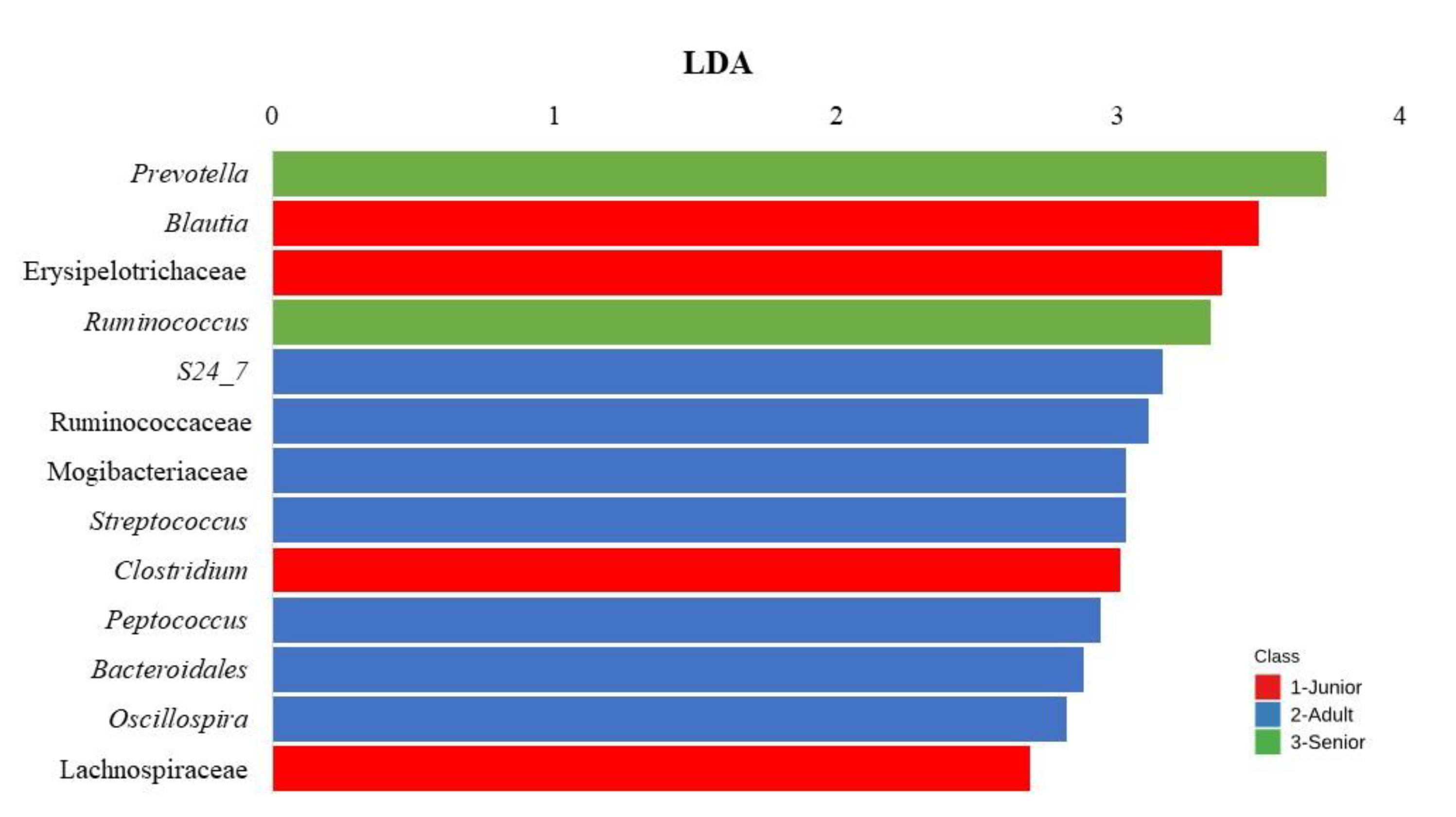
Figure 6.
Result of Linear discriminant analysis (LDA) effect Size of taxa which differs (FDR<0.10; LDA score >2) between age in healthy and osteoarthritic dog population (n=136). Junior_HE: healthy dog with age ranging 20–46 months; Adult_HE: healthy dog with age ranging 47–92 months; Adult_OA: osteoarthritic dogs with age ranging 47–92 months; Senior_HE: healthy dogs with age ranging 93–168 months; Senior_OA: osteoarthritic dogs with age ranging 93–168 months.
Figure 6.
Result of Linear discriminant analysis (LDA) effect Size of taxa which differs (FDR<0.10; LDA score >2) between age in healthy and osteoarthritic dog population (n=136). Junior_HE: healthy dog with age ranging 20–46 months; Adult_HE: healthy dog with age ranging 47–92 months; Adult_OA: osteoarthritic dogs with age ranging 47–92 months; Senior_HE: healthy dogs with age ranging 93–168 months; Senior_OA: osteoarthritic dogs with age ranging 93–168 months.
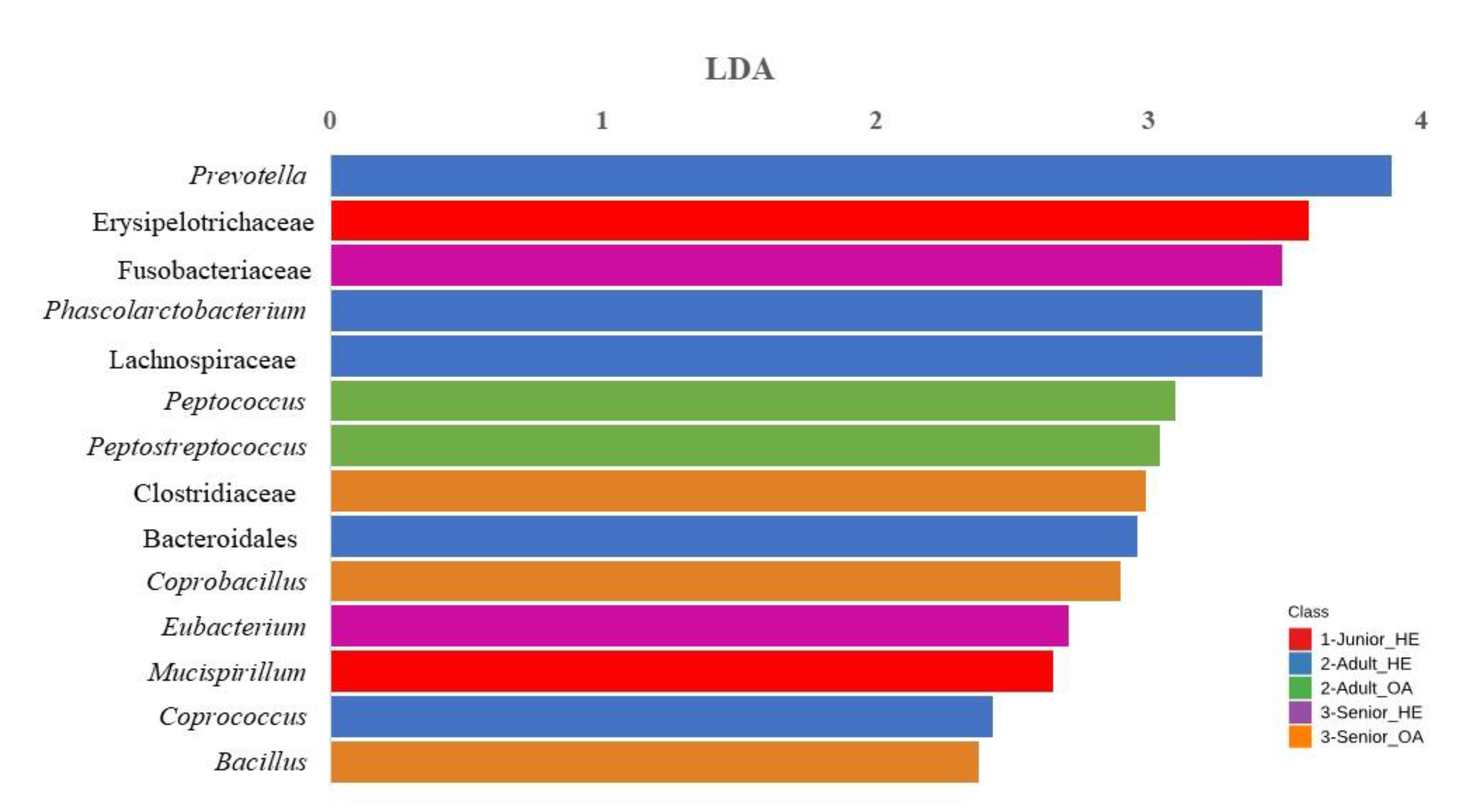

Table 1.
Contingency table of the whole dog population by age, sex and healthy conditions.
| Age | Sex | Healthy | Osteo-arthritis | Other | Total | ||
| Junior | Fs | 11 | 0 | 0 | 11 | ||
| Adult | 17 | 16 | 2 | 35 | |||
| Senior | 4 | 41 | 7 | 52 | |||
| Junior | Mc | 19 | 0 | 13 | 32 | ||
| Adult | 13 | 8 | 2 | 24 | |||
| Senior | 5 | 16 | 1 | 21 | |||
| Junior | Total | 30 | 0 | 13 | 43 | ||
| Adult | 30 | 24 | 4 | 58 | |||
| Senior | 9 | 57 | 8 | 74 | |||
| Total | 69 | 81 | 25 | 175 | |||
Fs: Spayed females; Mc: Castrated males; Junior: dog with age ranging 20–46 months; Adult dog with age ranging 47–92 months; Senior: dogs with age ranging 93–168 months; Other: 9 Junior dogs with diabetes and 4 Junior dogs with adverse drug reaction; 3 Adult dogs with seizure and 1 Adult dog with Cranial Cruciate Tear; 8 Senior dogs were affected by different diseases, as heart murmur (2), atopic dermatitis (1), congestive heart disease (1), seizure (4).
Table 2.
Contingency table by age, sex and healthy conditions of the selected dog population.
| Junior | Healthy | Osteo-arthritic | Total |
| Junior | 26 | 0 | 26 |
| Adult | 26 | 23 | 49 |
| Senior | 9 | 52 | 61 |
| Total | 61 | 75 | 136 |
Junior: dog with age ranging 20–46 months; Adult dog with age ranging 47–92 months; Senior: dogs with age ranging 93–168 months.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



