
Submitted:
11 April 2025
Posted:
14 April 2025
You are already at the latest version
Abstract
Leishmania infantum is widely recognized as the primary causative agent of visceral leishmaniasis (VL) in Turkey, while Leishmania tropica predominantly causes cutaneous leishmaniasis (CL). Although L. tropica is capable of causing VL, such cases remain exceedingly rare. This study aimed to identify genetic factors underlying the visceralization potential of L. tropica by comparing isolates from VL and CL patients.
Fourteen patients diagnosed with L. tropica infection, confirmed by parasite detection and genotyping between 2012 and 2022, were included: seven patients with VL and seven with CL. Clinical specimens were cultured for parasite isolation, and genotyping was performed via real-time PCR targeting the internal transcribed spacer 1 (ITS1) region. Differential gene expression was analyzed using quantitative real-time PCR (qRT-PCR), focusing on genes previously implicated in visceralization, including Cytochrome C Oxidase subunit IV, Metallo-peptidase (Clan MA(E), Family M32), Oligopeptidase B, Peroxidoxin 1, Peroxidoxin 2, Pyruvate kinase, and Succinyl-CoA:3-ketoacid-coenzyme A transferase. Next-generation sequencing (NGS) was conducted to detect potential mutations in these genes expressions was assessed by qRT-PCR.
Results revealed significantly elevated mRNA expression levels of Peroxidoxin 1 and 2 and Cytochrome C Oxidase subunit IV in VL isolates compared to CL isolates and reference strains, showing increases of approximately 17-fold and 21-fold, respectively. These findings suggest that enhanced expression of these genes contributes to parasite survival and proliferation in visceral organs. NGS analyses identified multiple mutations within key genes such as Oligopeptidase B and Metallo-peptidase (Clan MA(E), Family M32), potentially explaining distinct pathogenic traits between VL and CL isolates.
Collectively, this study identifies critical genetic factors involved in the pathogenesis of viscerotropic L. tropica infections. These findings offer significant insights into disease mechanisms, highlight potential therapeutic targets, and challenge the traditional perception of L. tropica as solely a causative agent of CL. The implications extend to developing improved diagnostic approaches, targeted therapies, and preventive strategies, thus providing a robust framework for future research.
Keywords:
Leishmania tropica
; Visceral Leishmaniasis
; Cutaneous Leishmaniasis
; Leishmania RNA Virus
; Virulenc
Highlights
- Increased Peroxidoxin expression linked to L. tropica visceralization in VL cases.
- No Leishmania RNA virus 1 detected in visceralized L. tropica isolates.
- L. tropica VL isolates show significant gene expression differences vs. CL strains.
- Novel mutations in Oligopeptidase B and Metallo-peptidase linked to VL pathogenesis.
- L. tropica identified as a potential cause of visceral leishmaniasis.
1. Introduction
There are approximately 30 distinct Leishmania species known to infect mammals; ten species are identified in the Old World, whereas the remaining 20 species are present in the New World. Leishmania parasites exhibit a digenetic life cycle comprising two distinct morphological stages: extracellular promastigotes within the invertebrate vector, and intracellular amastigotes residing in the macrophages of vertebrate hosts. Of these, 21 species have been identified as pathogenic to humans [1,2,3]. Leishmaniasis, a disease caused by intracellular parasites, is endemic primarily in tropical and subtropical regions. It is transmitted predominantly by the bite of infected female sand flies belonging to the genera Phlebotomus and Lutzomyia, prevalent across Europe, North Africa, the Middle East, Asia, and parts of South America. Recognized by the World Health Organization (WHO) as one of the seven most significant neglected tropical diseases, leishmaniasis constitutes a major public health concern due to its high morbidity, diverse clinical presentations, and considerable mortality. Endemic areas include Central and South America, parts of Southeast Mexico, Southern Europe, the Middle East, Africa, and Asia [4]. Leishmaniasis disproportionately affects impoverished populations, and its incidence is closely associated with socioeconomic factors such as malnutrition, population displacement, inadequate housing, compromised immunity, and limited healthcare resources. As of 2022, among the 200 countries and territories reporting to WHO, 99 were classified as endemic for leishmaniasis; of these, 71 countries reported both visceral (VL) and cutaneous leishmaniasis (CL), nine reported exclusively VL, and 19 reported exclusively CL. Approximately 85% of global VL cases originated from Brazil, Ethiopia, India, Kenya, Somalia, South Sudan, and Sudan, while eight countries—Afghanistan, Algeria, Brazil, among others—accounted for 85% of global CL cases [5].
Clinical manifestations of Leishmania infections include visceral leishmaniasis (VL), cutaneous leishmaniasis (CL), and mucocutaneous leishmaniasis (MCL), each correlated with distinct Leishmania species and characteristic clinical symptoms. VL, primarily caused by Leishmania donovani and Leishmania infantum, is characterized clinically by prolonged fever, hepatosplenomegaly, significant weight loss, anemia, leukopenia, and thrombocytopenia. CL, most commonly associated with Leishmania major and Leishmania braziliensis, typically presents with localized nodular lesions that may ulcerate. The severity and progression of CL lesions are heavily influenced by the host’s immune response. MCL, predominantly linked to L. braziliensis infection, involves extensive mucosal damage and severe morbidity. Although VL represents the most severe and potentially fatal form, CL remains more prevalent, particularly in endemic regions. Accurate diagnosis is complicated by clinical presentations overlapping with other diseases, underscoring the necessity for precise diagnostic methodologies [6,7,8,9,10].
Recent studies have indicated that certain Leishmania species previously believed to cause exclusively cutaneous or visceral disease may, in fact, be capable of inducing both clinical forms [11,12]. Nevertheless, significant evidence demonstrates clear species-level divergence between cutaneous and visceral strains, notably regarding their distinct adaptations for visceral tropism [1]. These adaptations include tolerance to higher temperatures found within internal organs [13], enhanced resistance to oxidative stress, a crucial aspect of host immune defense [14] and differential host cell tropism [15]. Additionally, parasite, host, and vector factors collectively influence visceralization, with the A2 gene family notably implicated as essential for establishing visceral infection [15,16,17].
The genus Leishmania is characterized by a meiosis-like reproductive mechanism, contributing significantly to intraspecific genetic variat [18]. Specifically, Leishmania tropica exhibits remarkable genomic plasticity, frequently undergoing genetic exchange events, including sexual reproduction, at rates higher than observed in other Leishmania species. Observed intra- and inter-chromosomal rearrangements in L. tropica suggest active regulatory processes involving mitotic, meiotic, and parasexual mechanisms [19]. The extensive genetic diversity observed in L. tropica is attributed to its digenetic life cycle, inherent chromosomal instability, frequent hybridization events, and high allelic variability. Such factors collectively enable rapid adaptation and increased pathogenic potential, complicating disease control and therapeutic intervention strategies [20,21,22]. Despite knowledge of tropism toward either cutaneous or visceral involvement, the underlying molecular mechanisms remain incompletely understood [23]. The primary objective of this study was to identify specific genetic determinants associated with visceralization of Leishmania tropica isolates from immunocompetent hosts, contributing to a deeper understanding of its pathogenicity and clinical outcomes.
2. Material and Methods
2.1. Patients
This study involved promastigotes isolated and cultured from 14 individuals diagnosed with Leishmania tropica infections, comprising seven patients presenting with visceral leishmaniasis (VL) and seven patients with cutaneous leishmaniasis (CL), to investigate genetic determinants associated with viscerotropism. Inclusion criteria were clearly defined as follows: patients diagnosed with VL exhibited clinical manifestations including hepatomegaly, splenomegaly, and pancytopenia, while CL patients presented with persistent cutaneous lesions lasting at least two months. All participants were diagnosed and treated at the Parasitology Laboratory of the Faculty of Medicine, Manisa Celal Bayar University, between 2012 and 2022. Parasite genotyping was systematically performed to confirm the identification of L. tropica. Patients diagnosed with VL received intravenous liposomal amphotericin B therapy at a dose of 3 mg/kg/day administered on days 1 through 5, followed by additional doses on days 14 and 21, totaling seven doses. Clinical improvement, characterized by resolution of fever, normalization of liver and spleen size, and restoration of hematological parameters, was observed by the end of the first month of treatment. Abdominal ultrasonography confirmed the resolution of hypoechoic nodules in the spleen. Patients were discharged upon full recovery and continued to be monitored during regular outpatient follow-up visits. Patients diagnosed with CL were treated using intralesional injections administered twice weekly. A minimum of eight injections were applied directly to lesion sites until complete blanching of lesions was observed [5]. Ethical approval for the study was granted by the Manisa Celal Bayar University Local Ethics Committee for Research Studies (Approval No: 20478486–050.04.04–20.478.486). Written informed consent was obtained from all participants prior to their inclusion in the study.
2.2. Clinical Sample Collection and Parasite Culture
Preliminary diagnoses of VL were confirmed by analyzing clinical specimens collected via fine needle aspiration from pelvic bone marrow by authorized physicians. For CL cases, the lesion area and surrounding healthy skin were first cleansed with 70% ethanol, followed by injection of 0.2–0.5 mL sterile saline solution directly into the lesion (Figure 1). Amastigotes were identified microscopically after staining the aspirate samples with Giemsa stain. All clinical samples were cultured initially on nutrient-enriched Novy-MacNeal-Nicolle (NNN) medium supplemented with cow milk and cow liver extract (EM medium), as previously described [24]. Promastigote cultures were incubated at 26°C and monitored daily for one month. Subsequently, established promastigote cultures were transferred to RPMI 1640 medium supplemented with 10% fetal calf serum (FCS), 200 U/mL penicillin, and 0.2 mg/mL streptomycin, and expanded in 25 mL flasks (5 mL culture volume) to yield sufficient quantities for genomic and gene expression analyses. Parasite density was adjusted to 1×10⁸ cells/mL, washed five times in sterile saline, and utilized in subsequent experiments.
2.3. Leishmania Genotyping
Leishmania genotyping was performed using real-time PCR assays targeting the ribosomal internal transcribed spacer 1 (ITS1) region, which separates the genes encoding small subunit (ssu) and 5.8S ribosomal RNAs in Leishmania species. Primers for amplification were ITS1 Forward (5'-CTGGATCATTTTCCGATG-3') and ITS1 Reverse (5'-GAAGCCAAGTCATCCATCGC-3'), utilizing the QuantiTect Probe PCR Master Mix. Species differentiation was achieved by melting curve analysis employing specific fluorescent probes: Probe 1 (5'-CCGTTTATACAAAAAATATACGGCGTTTCGGTTT-Fluo-3') and Probe 2 (5'-LCRed640-GCGGGGTGGGTGCGTGTGTG-Pho-3') [25]. Reference strains included in genotyping were L. tropica (MHOM/AZ/1974/SAF-K27), L. major (MHOM/SU/1973/5ASKH), L. infantum (MHOM/TN/1980/IPT1), and L. donovani (MHOM/IN/1980/DD8).
2.4. Real-Time qRT-PCR
Expression levels of seven previously identified genes associated with viscerotropism—Peroxidoxin 1 and 2, Oligopeptidase B, Metallo-peptidase (Clan MA(E), Family M32), Cytochrome C Oxidase subunit IV, Succinyl-CoA:3-ketoacid-coenzyme A transferase, and Pyruvate kinase—were quantified at the mRNA level [26]. Promastigotes were selected for gene expression analyses due to their ease of culture and suitability for standardized laboratory conditions. Although amastigotes represent the pathogenic intracellular stage in human hosts, technical limitations associated with their consistent in vitro cultivation necessitated using promastigotes as an experimental model to investigate genes potentially involved in viscerotropism. Promastigotes from primary NNN cultures exhibiting growth within 5–6 days were subcultured into RPMI-1640 medium and utilized at the logarithmic growth phase. Cultured parasites were cryopreserved without undergoing a second passage. Total RNA was extracted from promastigotes (3 × 10⁷ cells) of seven CL, seven VL, and reference L. tropica strains (MHOM/AZ/1974/SAF-K27) and L. infantum strains (MHOM/TR/2006/CBU20) using Trizol reagent according to the manufacturer’s instructions. Complementary DNA (cDNA) synthesis was conducted using SuperScript II Reverse Transcriptase, followed by purification with the QIAquick PCR purification kit, following the manufacturer’s guidelines. Real-time quantitative reverse transcriptase PCR (qRT-PCR) assays were performed utilizing QuantiTect Rotor-Gene SYBR Green PCR Kit with gene-specific primers (second primer pairs for each gene provided in Table S1) on the Rotor-Gene Q instrument. Gene expression normalization was performed using 18S rRNA as the internal control, and relative fold-changes were calculated using the 2−ΔΔCt method in comparison to reference L. tropica gene expression.
2.5. Evaluation of Genes by Comparing Gene Sequences
Next-generation sequencing (NGS) was employed to sequence gene regions hypothesized to be responsible for viscerotropism, using genomic DNA (gDNA) derived from Leishmania tropica isolates obtained from visceral leishmaniasis (VL) and cutaneous leishmaniasis (CL) patients. NGS provides deep sequencing coverage (≥100 reads per region), facilitating the detection of low-frequency single nucleotide polymorphisms (SNPs) and copy number variations (CNVs). Although the L. tropica genome assembly remains incomplete, relevant gene sequences are accessible via publicly available genome databases.
Genomic DNA was extracted from seven VL and seven CL isolates using a commercial genomic DNA extraction kit (Thermo Scientific) according to the manufacturer's instructions. For sequencing on the Ion Torrent PGM platform, approximately 100 ng of gDNA per isolate was utilized. Targeted gene regions were amplified using specific primers listed in Table S1. Following amplification, fragment libraries were prepared using the Ion Xpress Plus Fragment Library Kit, following the manufacturer’s protocol [27].
Barcoding of each Leishmania isolate library was achieved through ligation of Ion Xpress Barcode adapters to the fragmented DNA, according to the kit guidelines. Library normalization prior to template preparation was performed using the Library Equalizer Kit. Libraries were subsequently amplified using emulsion PCR on Ion Sphere Particles (ISPs) via the Ion PGM Hi-Q OT2 Kit, according to the manufacturer's recommendations [28]. Enrichment of templated ISPs was performed with the Ion PGM Enrichment Beads kit. Quality control assessments of enriched ISPs were conducted using the Ion Sphere Quality Control Kit before sequencing. Sequencing was performed on an Ion 316 Chip Kit, enabling ISPs to be sequenced across millions of wells by the Ion PGM sequencer.
Data analysis was performed using the Torrent Suite software, where base sequences were evaluated, and genetic variations between the L. tropica strains were identified with the Variant Caller software integrated into the CLC Genomics Workbench platform. The Variant Caller software identifies highly specific genetic variants using a fixed ploidy model (ploidy set to 2, variant probability threshold ≥50%), employing a Bayesian statistical approach combined with maximum likelihood analysis. Identified gene variants were compared against reference sequences from known VL-causing Leishmania species to evaluate their biological significance and relevance [29]. Reads were aligned to target gene regions, and variant analyses were performed to detect mutations.
Additionally, gene expression levels correlating with identified genetic mutations were quantified via quantitative real-time PCR (qRT-PCR) analysis using total RNA extracted from the same Leishmania isolates, as described in Section 2.4.
2.6. Statistical Analysis
Statistical comparisons among the three experimental groups were conducted using one-way analysis of variance (ANOVA) following confirmation of data normality through appropriate tests. All statistical analyses were performed using GraphPad Prism software version 9.5. Differences were considered statistically significant at a p-value of <0.05.
3. Results
3.1. Leishmaniasis Cases
Clinical samples collected from patients preliminarily diagnosed with cutaneous leishmaniasis (CL) were cultured, and promastigote growth was successfully achieved in enriched Novy-MacNeal-Nicolle (NNN) medium. The microscopic examination confirmed the presence of amastigotes in seven patient samples. Additionally, promastigotes were successfully isolated and cultured from bone marrow aspirates obtained from seven patients residing in the Aegean and Mediterranean regions, clinically diagnosed with visceral leishmaniasis (VL). Clinical assessments, including comprehensive medical history evaluations and routine laboratory examinations, confirmed that these VL patients had intact immune systems with no underlying systemic or immunological conditions other than VL itself. The clinical diagnosis of VL was based on the presentation of characteristic symptoms such as prolonged fever, pancytopenia, hepatosplenomegaly, significant weight loss, and microscopic confirmation of amastigotes in bone marrow aspirates. Quantitative PCR (qPCR) analyses of promastigotes isolated from these patients confirmed the species identification as Leishmania tropica in all 14 isolates through genotyping (Figure 2a). Detailed clinical information and genotyping data of the study participants are summarized in Table 1.
3.2. mRNA Expression Alterations of Genes Associated with Viscerotropism
The genes previously associated with viscerotropism in Leishmania tropica include Peroxidoxin 1, Peroxidoxin 2, Oligopeptidase B, Metallo-peptidase (Clan MA(E), M32 family protein), Pyruvate kinase, Succinyl-CoA:3-ketoacid-coenzyme A transferase, and Cytochrome C Oxidase subunit IV [26]. No significant differences were observed between isolates from patients with cutaneous leishmaniasis (CL) and reference L. tropica strains concerning mRNA expression levels of these viscerotropic genes (Figure 3). Additionally, Pyruvate kinase expression did not differ significantly in the visceral leishmaniasis (VL) group. Due to the high degree of homology between Peroxiredoxin 1 and Peroxiredoxin 2, it was not feasible to distinguish these two proteins at the mRNA level. However, combined Peroxiredoxin 1/2 mRNA expression in the VL group demonstrated a significant 17-fold increase compared to reference L. tropica expression levels (p < 0.0001). Among the genes examined, Cytochrome C oxidase subunit IV displayed the highest fold increase, with a 21-fold elevation in the VL group (p < 0.0001). In addition, Metallo-peptidase, Clan MA(E) M32 family protein, Oligopeptidase B, and Succinyl-CoA:3-ketoacid-coenzyme A transferase exhibited 6.1-, 3.9-, and 3.1-fold increases in mRNA expression, respectively, in the VL group (p < 0.05). When reference L. infantum expression levels were compared with those of reference L. tropica, Peroxiredoxin 1 and 2 mRNA expression showed a 9.16-fold increase, similar to that observed in the VL group. Another notable increase was detected in Oligopeptidase B mRNA expression, which increased by 9.64-fold. The remaining genes—Metallo-peptidase, Clan MA(E) M32 family protein, Pyruvate kinase, Succinyl-CoA:3-ketoacid-coenzyme A transferase, and Cytochrome C oxidase subunit IV—demonstrated fold changes of 1.84, 1.84, 1.40, and 1.37-fold, respectively.
Gene Ontology (GO) analyses were employed to categorize the molecular functions of these seven viscerotropism-associated genes. The identified molecular functions included "catalytic activity" (GO:0003824), which was further subclassified into five specific categories. "Catalytic activity, acting on a gene" (22.22%, GO:0140096) encompasses enzymes that modify the structure or function of proteins via enzymatic reactions, post-translational modifications, or protein-protein interactions. "Transferase activity" (22.22%, GO:0016740) describes enzymes that catalyze the transfer of functional groups (e.g., methyl, glycosyl, acyl, phosphate) from donor to acceptor molecules, typically enzymes within EC class 2. "Hydrolase activity" (22.22%, GO:0016787) involves enzymes that catalyze hydrolytic cleavage of chemical bonds such as C–O, C–N, C–C, and phosphoric anhydride bonds. "Oxidoreductase activity" (22.22%, GO:0016491) refers to enzymes that catalyze oxidation-reduction reactions, facilitating electron or hydrogen transfer and modifying the oxidation states of substrates. Lastly, "antioxidant activity" (11.11%, GO:0016209) includes compounds that inhibit oxidative reactions initiated by dioxygen or peroxide, thereby protecting molecules from oxidative damage by undergoing oxidation preferentially (Figure 4).
3.3. Gene-Level Analysis of Visceralized Leishmania Tropica
Peroxidoxins are thiol-specific peroxidases responsible for catalyzing the reduction of organic hydroperoxides and hydrogen peroxide into alcohol and water, respectively. Due to the high sequence homology between Peroxidoxin 1 and Peroxidoxin 2, a combined sequence representative of Peroxidoxin genes was submitted to GenBank (accession number: OQ689280). Targeted next-generation sequencing (NGS) revealed no distinct mutations within the Peroxidoxin 1 and 2 gene regions across the seven visceral leishmaniasis (VL) strains, seven cutaneous leishmaniasis (CL) strains, and the reference Leishmania tropica strain.
Oligopeptidase B, an enzyme that hydrolyzes oligopeptides at proline and alanine residues, was sequenced and submitted to GenBank (accession number: OQ689281). NGS analysis identified several mutations unique to VL isolates compared to CL and reference strains. These included silent mutations c.255C>T (p.Asp85Asp), c.1332G>A (p.Val444Val), c.1479G>T (p.Leu493Leu), c.1501G>T (p.Leu501Leu), and c.1950G>T (p.Glu650Glu), as well as missense mutations c.1031A>G (p.Asp344Gly) and c.1306C>G (p.Pro436Ala), all detected heterozygously in CL and reference strains.
Metallo-peptidase (Clan MA(E), Family M32), known for its metallocarboxypeptidase activity, was also sequenced and submitted to GenBank (accession number: OQ689282). Targeted NGS analysis revealed nine mutations in this gene across VL, CL, and reference strains. Notably, the c.169G>T (p.Ala57Ser) missense mutation within the peptidase M32 domain was heterozygous in CL and reference strains. The silent mutation c.228G>A (p.Ala76Ala) was homozygous exclusively in VL strains, while silent mutations c.525T>C (p.Asn175Asn), c.588G>C (p.Val196Val), c.594G>A (p.Ala198Ala), and c.1308G>A (p.Glu436Glu) were homozygous in VL strains but heterozygous in CL and reference strains.
Cytochrome C Oxidase subunit IV, exhibiting oxidoreductase activity, was sequenced and submitted to GenBank (accession number: OQ689285). Three mutations were detected within its gene region. The silent mutations c.21A>C (p.Val7Val) and c.23C>T (p.Ser8Ser), both located in the Signal Peptide Region C domain, as well as c.282C>T (p.Leu94Leu), were homozygous in VL strains and heterozygous in CL and reference strains.
Succinyl-CoA:3-ketoacid-coenzyme A transferase, an enzyme critical for ketone body metabolism, was sequenced and submitted to GenBank (accession number: OQ689284). Five mutations were detected, including the silent mutations c.48T>G (p.Gln16Gln) and c.321G>T (p.Phe107Phe), both homozygous in VL strains. Other mutations—c.378C>T (p.Ala126Ala), c.1152A>G (p.Ile384Ile), and c.1254T>C (p.Ser418Ser)—were heterozygous in CL and reference strains.
Pyruvate kinase, responsible for catalyzing the conversion of phosphoenolpyruvate to pyruvate, was also sequenced and submitted to GenBank (accession number: OQ689283). Four mutations were identified: silent mutations c.67C>A (p.Thr22Thr), c.588G>T (p.Gly196Gly), and c.786G>A (p.Arg262Arg), along with a missense mutation c.1362G>A (p.Glu454Lys). All four mutations were homozygous in VL strains, whereas CL and reference strains exhibited heterozygosity.
Table 3 summarizes the identified missense mutations from the targeted NGS analysis, highlighting significant genetic variations between VL and CL isolates of L. tropica and their potential implications in disease pathogenesis and therapeutic strategies.

Table 2.
Clinical and laboratory characteristics of visceral leishmaniasis (VL) patients.
| Clinical feature | V1 | V2 | V3 | V4 | V5 | V6 | V7 |
|---|---|---|---|---|---|---|---|
| Fever | + | + | + | + | + | + | + |
| Weight loss | + | + | + | – | + | + | – |
| Fatigue | + | + | – | + | + | + | + |
| GIS symptoms | – | + | + | + | – | + | + |
| Epistaxis/Gingival bleeding | + | – | – | – | – | – | + |
| Splenomegaly | + | + | + | + | + | + | + |
| Hepatomegaly | + | + | + | + | - | + | + |
| Pancytopenia | + | + | + | - | + | + | + |
| Leukopenia | – | – | – | + | – | – | – |
| Thrombocytopenia | – | – | – | + | – | – | – |
| Microscopy | Positive | Negative | Positive | Positive | Positive | Positive | Positive |
| Modified NNN | Positive | Positive | Positive | Positive | Positive | Positive | Positive |
| Seropositivity (IFAT) | 1/512 | 1/512 | 1/512 | 1/1024 | 1/512 | 1/1024 | 1/1024 |
| qPCR (Bone marrow) | Positive | Positive | Positive | Positive | Positive | Positive | Positive |
4. Discussion
Viscerotropic Leishmania tropica is characterized by its capacity to initiate visceral or systemic infections in humans. Numerous studies have investigated the pathogenesis and clinical features of viscerotropic leishmaniasis caused by L. tropica. Although Leishmania tropica is predominantly recognized as the causative agent of CL, its involvement in visceral leishmaniasis (VL) has been reported, challenging conventional understanding. Initial evidence of VL associated with L. tropica emerged in 1989 following isolation from two Kenyan VL patients [30]. Subsequently, among eight U.S. military personnel returning from the Gulf War diagnosed with VL, six isolates were identified as L. tropica using isoenzyme analysis [31]. A case study involving a patient initially presenting with skin lesions and subsequently developing VL nine months later confirmed L. tropica as the causative species via PCR analysis of skin, blood, and bone marrow samples [32]. Recent studies employing molecular techniques frequently confirm L. tropica as a VL-causing agent. For instance, sequencing of splenic puncture samples from VL patients in endemic regions of Iran identified L. tropica with 99.9% certainty [33]. Further, genetic analyses of the internal transcribed spacer-1 (ITS-1) region confirmed VL-associated L. tropica isolates [34]. Genetic investigations revealed that expression of the viscerotropic leishmaniasis antigen gene (VTL) was elevated threefold in VL-causing L. tropica compared to L. infantum [35].
This study identified seven genes in human-derived L. tropica isolates that may contribute to visceral tropism, as confirmed by quantitative qRT-PCR analyses demonstrating elevated mRNA expression levels. Among these genes, the most striking finding was the 21-fold increase in Cytochrome C oxidase subunit IV mRNA expression in L. tropica isolates obtained from VL patients. This observation supports previous reports suggesting that elevated Cytochrome C oxidase expression is associated with higher virulence and resistance to antimonial therapy in Leishmania species [36,37]. Cytochrome C oxidase, located in the inner mitochondrial membrane, serves as the terminal component of the electron transport chain and plays a critical role in ATP production. The pronounced overexpression of this subunit likely reflects the heightened energy demands of the visceral milieu, possibly driven by rapid intracellular proliferation and the requirement to overcome the host’s immune responses. Studies in Leishmania major have shown that Cytochrome C oxidase subunit IV (LmCOX4) expression is regulated by a protein called LACK, which governs the parasite’s thermotolerance and virulence; disruption of LACK leads to reduced LmCOX4 levels, impaired mitochondrial function, and decreased ATP production. Restoring LmCOX4 expression in these LACK-deficient parasites rescues their ability to tolerate mammalian temperatures and improves their infectivity in macrophages [36]. The 21-fold increase observed in visceralized L. tropica isolates is thus consistent with the broader mechanism utilized by various Leishmania species, including L. donovani, to adapt and replicate within visceral organs [38].
Another key finding of this study was the 17-fold increase in Peroxiredoxin 1 and 2 mRNA expression in VL isolates compared to reference L. tropica strains. Peroxiredoxins are a family of antioxidant enzymes that protect cells against oxidative stress by neutralizing reactive oxygen species (ROS), which are abundantly generated by the host immune system during infection [37,39]. In line with these results, Hajjaran et al. reported that increased Peroxiredoxin expression supports parasite survival in visceral organs by mitigating macrophage-derived oxidative stress [37]. Studies in Leishmania infantum and L. donovani further emphasize the importance of mitochondrial peroxiredoxin in infection, showing that its absence or reduction significantly compromises parasite survival in murine models and increases sensitivity to oxidative damage [39,40]. Notably, even a peroxidase-inactive version of Peroxiredoxin can restore infectivity, suggesting an additional role for this enzyme as a molecular chaperone [39]. In the present study, reference L. infantum displayed a 9.16-fold increase in Peroxiredoxin 1 and 2 expression compared to reference L. tropica, underscoring a conserved and robust antioxidant defense mechanism in visceralization across different Leishmania species. This finding also aligns with evidence that strains causing self-healing cutaneous leishmaniasis are more susceptible to oxidative stress, indicating that resistance to oxidative damage is pivotal in facilitating visceral disease [41]. Additional genes that showed significant increases in mRNA expression in VL L. tropica isolates included Metallo-peptidase (Clan MA(E), M32 family) (6.1-fold), Oligopeptidase B (3.9-fold), and Succinyl-CoA:3-ketoacid-coenzyme A transferase (SCOT) (3.1-fold). Metallo-peptidases participate in various processes critical to parasite survival and proliferation—such as nutrient acquisition, protein processing, and immune evasion [40,42]. Consequently, the upregulation of this M32 family protease may bolster L. tropica adaptation within visceral organs. Oligopeptidase B, a serine protease associated with macrophage infection and intracellular survival [43,44], was also more highly expressed in VL isolates. Notably, reference L. infantum exhibited a 9.64-fold increase in Oligopeptidase B gene expression compared to reference L. tropica, possibly highlighting a species-specific significance for this enzyme in visceralization. SCOT plays a key role in ketone body metabolism, thereby furnishing the parasite with an alternative energy source in glucose-limited environments such as host macrophages [45]. Taken together, the collective overexpression of these genes suggests a multifaceted suite of adaptive strategies that enables L. tropica to survive and proliferate in the visceral environment.
When VL L. tropica was compared to reference L. infantum, a comparable elevation was observed in Peroxiredoxin 1/2 and Oligopeptidase B expression, indicating shared mechanisms that mitigate oxidative stress and support intracellular proliferation. However, the fold increases of Metallo-peptidase, Pyruvate kinase, SCOT, and Cytochrome C oxidase subunit IV in reference L. infantum were generally lower than those in VL L. tropica. These differences may reflect distinct evolutionary trajectories and host adaptation strategies, whereby each species employs diverse metabolic and immune-evasive pathways to establish infection in visceral tissues. It is noteworthy that many of the enzymes found to be overexpressed in this study—particularly antioxidant and peptidase enzymes—can disrupt the chain reactions initiated by free radicals, thereby reducing the extent of cellular damage [46]. In considering the catalytic activities underlying L. tropica’s visceralization, we propose that oxidoreductase and antioxidant functions are critical for maintaining the redox balance necessary for parasite survival, while hydrolase activities may facilitate nutrient acquisition or modulate host immune responses. Overall, these results illuminate key molecular pathways that allow L. tropica to adapt, proliferate, and persist within visceral environments, thereby offering potential avenues for future therapeutic intervention against visceral leishmaniasis.
Genetic mutation analysis of the seven candidate genes revealed no missense or nonsense mutations in Peroxidoxin 1 and 2, Cytochrome C Oxidase subunit IV, or Succinyl-CoA:3-ketoacid-coenzyme A transferase. However, Oligopeptidase B exhibited two missense mutations, and Metallo-peptidase showed one missense mutation. Oligopeptidase B, belonging to the S9 prolyl oligopeptidase family, is recognized as a significant virulence factor in Leishmania [60]. Metallo-peptidases (M32 family) are exclusively expressed by trypanosomatids and not found in other eukaryotes [47]. Detected heterozygous mutations (p.Asp344Gly, p.Pro436Ala in Oligopeptidase B, and p.Ala57Ser in Metallo-peptidase) exclusive to CL isolates suggest potential contributions to cutaneous tropism, possibly influencing tissue lesion formation and remodeling during infection.
5. Conclusions
This study represents one of the few comprehensive analyses investigating the genetic determinants of visceral tropism in Leishmania tropica. Seven key genes associated with human visceral leishmaniasis were identified, including the significantly upregulated Cytochrome C oxidase subunit IV, Peroxiredoxin 1 and 2, Metallo-peptidase (Clan MA(E), M32 family), Oligopeptidase B, and Succinyl-CoA:3-ketoacid-coenzyme A transferase. These findings provide compelling evidence for the differential expression of specific genes in VL-derived L. tropica isolates compared to cutaneous and reference strains, underscoring a complex interplay of factors that facilitate the parasite’s capacity to infect and persist in visceral organs. Collectively, this research greatly advances our understanding of L. tropica pathogenesis and highlights the importance of further investigation into the roles of these identified genes in the progression and severity of visceral leishmaniasis. By elucidating the molecular mechanisms of visceralization, the present findings lay crucial groundwork for the development of novel therapeutic interventions, targeted vaccines, precise diagnostic methodologies, and optimized preventive strategies. In turn, these advancements hold the potential to significantly improve the clinical management and control of leishmaniasis.
Author Contributions
Statement Ahmet Ozbilgin conceptualized the study and conducted data curation. Ibrahim Cavus and Varol Tunali performed formal analysis. Melike Dinc, Merve Beyaz, and Talat Yalcin carried out PhD thesis. Sukran Kose evaluated the Leishmania RNA virus studies. Asli Tetik Vardarli and Cumhur Gunduz conducted genomic studies and compared the results with expression analyses, in addition to overseeing the writing and revision of the manuscript.
Funding
This study was supported by the Scientific and Technological Research Council of Turkey (TÜBİTAK) with Project No. 119S469.
Acknowledgments
The authors extend their sincere gratitude to the Manisa Celal Bayar University Parasite Bank for their valuable contributions and to TUBITAK for their support. .
Conflicts of Interest
The authors affirm that the investigation was carried out without any existing commercial or financial affiliations that could be interpreted as potential conflicts of interest.
References
- McCall, L.I.; Zhang, W.W.; Matlashewski, G. Determinants for the Development of Visceral Leishmaniasis Disease. PLoS Pathog 2013, 9, e1003053. [Google Scholar] [CrossRef]
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; de Boer, M. Leishmaniasis Worldwide and Global Estimates of Its Incidence. PLoS One 2012, 7, e35671. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, J. Leishmania. JAMA 2007, 298, 277–277. [Google Scholar] [CrossRef]
- Arenas, R.; Torres-Guerrero, E.; Quintanilla-Cedillo, M.R.; Ruiz-Esmenjaud, J. Leishmaniasis: A Review. F1000Res 2017, 6. [Google Scholar] [CrossRef]
- Clinical Care of Leishmaniasis | Leishmaniasis | CDC. Available online: https://www.cdc.gov/leishmaniasis/hcp/clinical-care/index.html (accessed on 26 October 2024).
- Bailey, M.S.; Lockwood, D.N.J. Cutaneous Leishmaniasis. Clin Dermatol 2007, 25, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Mathers, C.D.; Ezzati, M.; Lopez, A.D. Measuring the Burden of Neglected Tropical Diseases: The Global of Disease Framework. PLoS Negl. Trop. Dis. 2007, 1, e114. [Google Scholar] [CrossRef]
- Pedrosa, C.M.S.; Pedrosa, C.M.S. Clinical Manifestations of Visceral Leishmaniasis (American Visceral Leishmaniasis). The Epidemiology and Ecology of Leishmaniasis 2017. [Google Scholar] [CrossRef]
- Scorza, B.M.; Carvalho, E.M.; Wilson, M.E. Cutaneous Manifestations of Human and Murine Leishmaniasis. International Journal of Molecular Sciences 2017, 18, 1296. [Google Scholar] [CrossRef]
- Kumar, V.; Madhu, M.; Murti, K. An Overview on Leishmaniasis. Viral, Parasitic, Bacterial, and Fungal Infections: Antimicrobial, Host Defense, and Therapeutic Strategies 2023, 389–406. [Google Scholar] [CrossRef]
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. The Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Thakur, L.; Singh, K.K.; Shanker, V.; Negi, A.; Jain, A.; Matlashewski, G.; Jain, M. Atypical Leishmaniasis: A Global Perspective with Emphasis on the Indian Subcontinent. PLoS Negl Trop Dis 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- Callahan, H.L.; Portal, I.F.; Bensinger, S.J.; Grogl, M. Leishmaniaspp: Temperature Sensitivity of Promastigotesin Vitroas a Model for Tropismin Vivo. Exp Parasitol 1996, 84, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Ghosh, S.; Pakrashi, S.; Roy, D.; Sen, S.; Chatterjee, M. Leishmania Strains Causing Self-Healing Cutaneous Leishmaniasis Have Greater Susceptibility towards Oxidative Stress. Free Radical Research 2012, 46, 665–673. [Google Scholar] [CrossRef]
- Zhang, W.W.; Matlashewski, G. Loss of Virulence in Leishmania Donovani Deficient in an Amastigote-Specific Protein, A2. Proc Natl Acad Sci U S A 1997, 94, 8807–8811. [Google Scholar] [CrossRef]
- Mizbani, A.; Taslimi, Y.; Zahedifard, F.; Taheri, T.; Rafati, S. Effect of A2 Gene on Infectivity of the Nonpathogenic Parasite Leishmania Tarentolae. Parasitol Res 2011, 109, 793–799. [Google Scholar] [CrossRef]
- Charest, H.; Matlashewski, G. Developmental Gene Expression in Leishmania Donovani: Differential Cloning and Analysis of an Amastigote-Stage-Specific Gene. Mol Cell Biol 1994, 14, 2975–2984. [Google Scholar] [CrossRef] [PubMed]
- Inbar, E.; Shaik, J.; Iantorno, S.A.; Romano, A.; Nzelu, C.O.; Owens, K.; Sanders, M.J.; Dobson, D.; Cotton, J.A.; Grigg, M.E.; et al. Whole Genome Sequencing of Experimental Hybrids Supports Meiosis-like Sexual Recombination in Leishmania. PLoS Genet 2019, 15. [Google Scholar] [CrossRef]
- Glans, H.; Karlberg, M.L.; Advani, R.; Bradley, M.; Alm, E.; Andersson, B.; Downing, T. High Genome Plasticity and Frequent Genetic Exchange in Leishmania Tropica Isolates from Afghanistan, Iran and Syria. PLoS Negl Trop Dis 2021, 15, e0010110. [Google Scholar] [CrossRef]
- Glans, H.; Karlberg, M.L.; Advani, R.; Bradley, M.; Alm, E.; Andersson, B.; Downing, T. High Genome Plasticity and Frequent Genetic Exchange in Leishmania Tropica Isolates from Afghanistan, Iran and Syria. PLoS Negl Trop Dis 2021, 15, e0010110. [Google Scholar] [CrossRef]
- Iantorno, S.A.; Durrant, C.; Khan, A.; Sanders, M.J.; Beverley, S.M.; Warren, W.C.; Berriman, M.; Sacks, D.L.; Cotton, J.A.; Grigg, M.E. Gene Expression in Leishmania Is Regulated Predominantly by Gene Dosage. mBio 2017, 8, e01393–17. [Google Scholar] [CrossRef]
- Bussotti, G.; Piel, L.; Pescher, P.; Domagalska, M.A.; Shanmugha Rajan, K.; Cohen-Chalamish, S.; Doniger, T.; Hiregange, D.G.; Myler, P.J.; Unger, R.; et al. Genome Instability Drives Epistatic Adaptation in the Human Pathogen Leishmania. Proc Natl Acad Sci U S A 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Ait Maatallah, I.; Akarid, K.; Lemrani, M. Tissue Tropism: Is It an Intrinsic Characteristic of Leishmania Species? Acta Trop 2022, 232, 106512. [Google Scholar] [CrossRef]
- Özbilgin, A.; Çulha, G.; Uzun, S.; Harman, M.; Topal, S.G.; Okudan, F.; Zeyrek, F.; Gündüz, C.; Östan, I.; Karakuş, M.; et al. Leishmaniasis in Turkey: First Clinical Isolation of Leishmania Major from 18 Autochthonous Cases of Cutaneous Leishmaniasis in Four Geographical Regions. Tropical Medicine and International Health 2016, 21, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Toz, S.O.; Culha, G.; Zeyrek, F.Y.; Ertabaklar, H.; Alkan, M.Z.; Vardarlı, A.T.; Gunduz, C.; Ozbel, Y. A Real-Time ITS1-PCR Based Method in the Diagnosis and Species Identification of Leishmania Parasite from Human and Dog Clinical Samples in Turkey. PLoS Negl Trop Dis 2013, 7, e2205. [Google Scholar] [CrossRef]
- Dinç, M. Proteomic Analyses of Biological Samples by Using Different Mass Spectrometric Strategies. ProQuest Dissertations & Theses 2018, 28476510. [Google Scholar]
- C. de Oliveira, T.; Rodrigues, P.T.; Menezes, M.J.; Gonçalves-Lopes, R.M.; Bastos, M.S.; Lima, N.F.; Barbosa, S.; Gerber, A.L.; Loss de Morais, G.; Berná, L.; et al. Genome-Wide Diversity and Differentiation in New World Populations of the Human Malaria Parasite Plasmodium Vivax. PLoS Negl Trop Dis 2017, 11. [Google Scholar] [CrossRef]
- Rao, P.N.; Uplekar, S.; Kayal, S.; Mallick, P.K.; Bandyopadhyay, N.; Kale, S.; Singh, O.P.; Mohanty, A.; Mohanty, S.; Wassmer, S.C.; et al. A Method for Amplicon Deep Sequencing of Drug Resistance Genes in Plasmodium Falciparum Clinical Isolates from India. J Clin Microbiol 2016, 54, 1500–1511. [Google Scholar] [CrossRef]
- Šlapeta, J.; Dowd, S.E.; Alanazi, A.D.; Westman, M.E.; Brown, G.K. Differences in the Faecal Microbiome of Non-Diarrhoeic Clinically Healthy Dogs and Cats Associated with Giardia Duodenalis Infection: Impact of Hookworms and Coccidia. Int J Parasitol 2015, 45, 585–594. [Google Scholar] [CrossRef]
- Mebrahtu, Y.; Lawyer, P.; Githure, J.; Were, J.B.; Muigai, R.; Hendricks, L.; Leeuwenburg, J.; Koech, D.; Roberts, C. Visceral Leishmaniasis Unresponsive to Pentostam Caused by Leishmania Tropica in Kenya. Am J Trop Med Hyg 1989, 41, 289–294. [Google Scholar] [CrossRef]
- Magill, A.J.; Grogl, M.; Gasser, R.A.; Sun, W.; Oster, C.N. Visceral Infection Caused by Leishmania Tropica in Veterans of Operation Desert Storm. N Engl J Med 1993, 328, 1383–1387. [Google Scholar] [CrossRef]
- Alborzi, A.; Pouladfar, G.R.; Fakhar, M.; Motazedian, M.H.; Hatam, G.R.; Kadivar, M.R. Isolation of Leishmania Tropica from a Patient with Visceral Leishmaniasis and Disseminated Cutaneous Leishmaniasis, Southern Iran. Am J Trop Med Hyg 2008, 79, 435–437. [Google Scholar] [CrossRef]
- SARKARI, B.; AHMADPOUR, N.B.; MOSHFE, A.; HAJJARAN, H. Molecular Evaluation of a Case of Visceral Leishmaniasis Due to Leishmania Tropica in Southwestern Iran. Iran J Parasitol 2016, 11, 126. [Google Scholar]
- Ghatee, M.A.; Mirhendi, H.; Karamian, M.; Taylor, W.R.; Sharifi, I.; Hosseinzadeh, M.; Kanannejad, Z. Population Structures of Leishmania Infantum and Leishmania Tropica the Causative Agents of Kala-Azar in Southwest Iran. Parasitol Res 2018, 117, 3447–3458. [Google Scholar] [CrossRef] [PubMed]
- Nafchi, H.R.; Kazemi-Rad, E.; Mohebali, M.; Raoofian, R.; Ahmadpour, N.B.; Oshaghi, M.A.; Hajjaran, H. Expression Analysis of Viscerotropic Leishmaniasis Gene in Leishmania Species by Real-Time RT-PCR. Acta Parasitol 2016, 61, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, D.; Carter, P.M.; Nation, C.S.; Pizarro, J.C.; Guidry, J.; Aiyar, A.; Kelly, B.L. LACK, a RACK1 Ortholog, Facilitates Cytochrome c Oxidase Subunit Expression to Promote Leishmania Major Fitness. Mol Microbiol 2015, 96, 95. [Google Scholar] [CrossRef] [PubMed]
- Hajjaran, H.; Mousavi, P.; Burchmore, R.; Mohebali, M.; Mohammadi Bazargani, M.; Hosseini Salekdeh, G.; Kazemi-Rad, E.; Khoramizadeh, M.R. Comparative Proteomic Profiling of Leishmania Tropica: Investigation of a Case Infected with Simultaneous Cutaneous and Viscerotropic Leishmaniasis by 2-Dimentional Electrophoresis and Mass Spectrometry. Iran J Parasitol 2015, 10, 366. [Google Scholar]
- Cytochrome c Oxidase Subunit IV - Leishmania Donovani | UniProtKB | UniProt. Available online: https://www.uniprot.org/uniprotkb/A0A3S5H6L2/entry (accessed on 31 March 2025).
- Castro, H.; Teixeira, F.; Romao, S.; Santos, M.; Cruz, T.; Flórido, M.; Appelberg, R.; Oliveira, P.; Ferreira-da-Silva, F.; Tomás, A.M. Leishmania Mitochondrial Peroxiredoxin Plays a Crucial Peroxidase-Unrelated Role during Infection: Insight into Its Novel Chaperone Activity. PLoS Pathog 2011, 7, e1002325. [Google Scholar] [CrossRef]
- Isaza, C.E.; Zhong, X.; Rosas, L.E.; White, J.D.; Chen, R.P.Y.; Liang, G.F.C.; Chan, S.I.; Satoskar, A.R.; Chan, M.K. A Proposed Role for Leishmania Major Carboxypeptidase in Peptide Catabolism. Biochem Biophys Res Commun 2008, 373, 25–29. [Google Scholar] [CrossRef]
- Sarkar, A.; Ghosh, S.; Pakrashi, S.; Roy, D.; Sen, S.; Chatterjee, M. Leishmania Strains Causing Self-Healing Cutaneous Leishmaniasis Have Greater Susceptibility towards Oxidative Stress. Free Radic Res 2012, 46, 665–673. [Google Scholar] [CrossRef]
- Alvarez, V.E.; Niemirowicz, G.T.; Cazzulo, J.J. The Peptidases of Trypanosoma Cruzi: Digestive Enzymes, Virulence Factors, and Mediators of Autophagy and Programmed Cell Death. Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics 2012, 1824, 195–206. [Google Scholar] [CrossRef]
- Swenerton, R.K.; Zhang, S.; Sajid, M.; Medzihradszky, K.F.; Craik, C.S.; Kelly, B.L.; McKerrow, J.H. The Oligopeptidase B of Leishmania Regulates Parasite Enolase and Immune Evasion. Journal of Biological Chemistry 2011, 286, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Goyal, S.; Grover, S.; Dhanjal, J.K.; Goyal, M.; Tyagi, C.; Chacko, S.; Grover, A. Mechanistic Insights into Mode of Actions of Novel Oligopeptidase B Inhibitors for Combating Leishmaniasis. J Mol Model 2014, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ashrafmansouri, M.; Amiri-Dashatan, N.; Ahmadi, N. Identification of Protein Profile in Metacyclic and Amastigote-like Stages of Leishmania Tropica: A Proteomic Approach. AMB Express 2022, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gene Ontology and GO Annotations. Available online: https://www.ebi.ac.uk/QuickGO/ (accessed on 9 May 2023).
- Frasch, A.P.; Carmona, A.K.; Juliano, L.; Cazzulo, J.J.; Niemirowicz, G.T. Characterization of the M32 Metallocarboxypeptidase of Trypanosoma Brucei: Differences and Similarities with Its Orthologue in Trypanosoma Cruzi. Mol Biochem Parasitol 2012, 184, 63–70. [Google Scholar] [CrossRef]
Figure 1.
Clinical presentations of cutaneous leishmaniasis (CL) lesions. C1: Nodular dry-type lesion on the right cheek persisting for 3 months; C2: Plaque-type lesion with increased vascularization on the right zygomatic region, persisting for 24 months; C3: Dry type lesion on the right arm for 3 months; C4: Dry type lesion on the right arm persisting for 3 months; C5: Dry-type lesion located on the right cheek persisting for 24 months; C6: Dry-type lesion on the right cheek for 12 months; C7: Dry, pruritic lesion located on the right arm for 12 months.
Figure 1.
Clinical presentations of cutaneous leishmaniasis (CL) lesions. C1: Nodular dry-type lesion on the right cheek persisting for 3 months; C2: Plaque-type lesion with increased vascularization on the right zygomatic region, persisting for 24 months; C3: Dry type lesion on the right arm for 3 months; C4: Dry type lesion on the right arm persisting for 3 months; C5: Dry-type lesion located on the right cheek persisting for 24 months; C6: Dry-type lesion on the right cheek for 12 months; C7: Dry, pruritic lesion located on the right arm for 12 months.
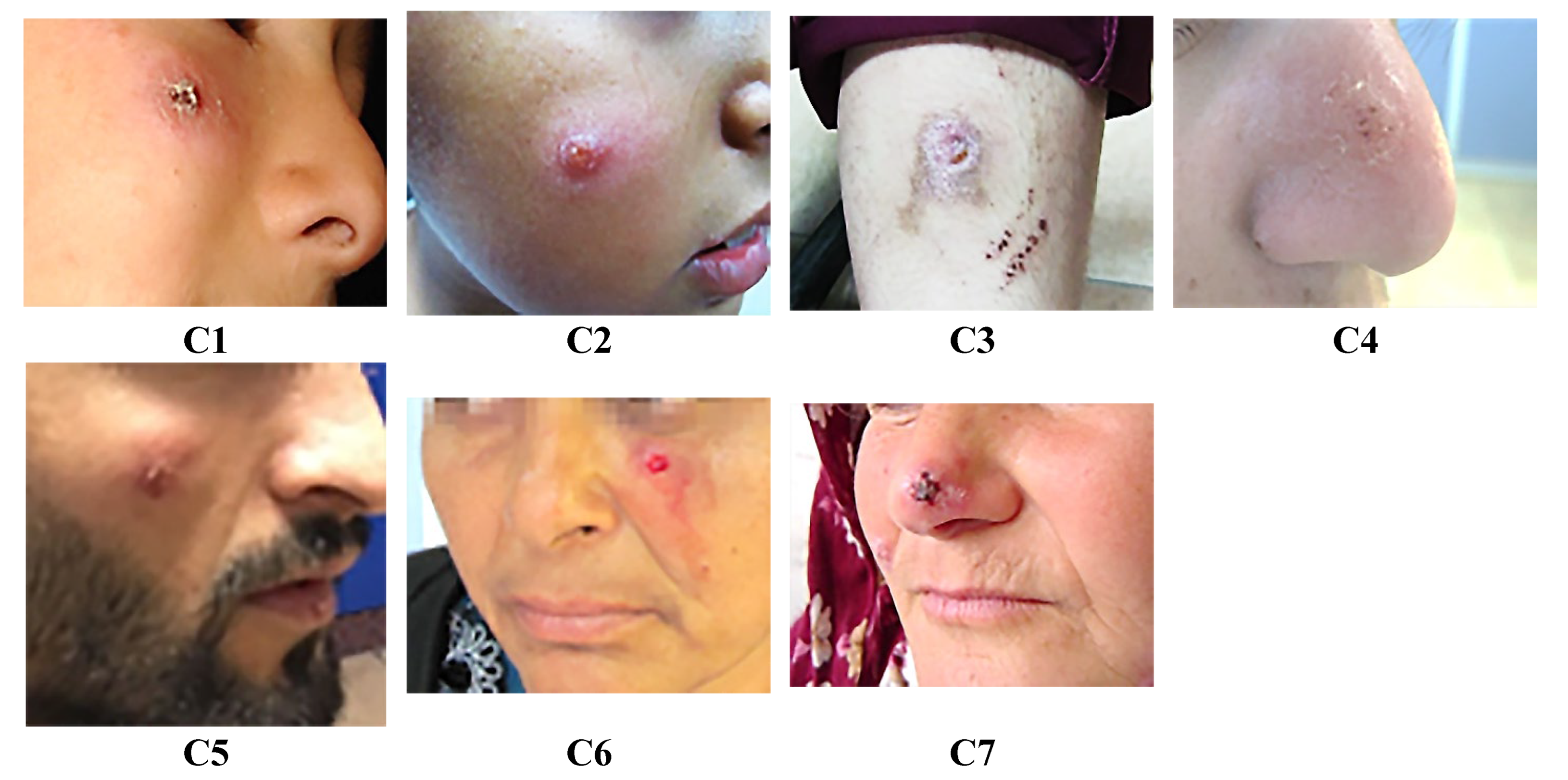
Figure 2.
Melting curves from quantitative polymerase chain reaction (qPCR) analyses of clinical specimens and promastigote cultures derived from leishmaniasis patients. Genotyping was performed using reference strains: Leishmania tropica (MHOM/AZ/1974/SAF-K27), Leishmania major (MHOM/SU/1973/5ASKH), Leishmania infantum (MHOM/TN/1980/IPT1), and Leishmania donovani (MHOM/IN/1980/DD8).
Figure 2.
Melting curves from quantitative polymerase chain reaction (qPCR) analyses of clinical specimens and promastigote cultures derived from leishmaniasis patients. Genotyping was performed using reference strains: Leishmania tropica (MHOM/AZ/1974/SAF-K27), Leishmania major (MHOM/SU/1973/5ASKH), Leishmania infantum (MHOM/TN/1980/IPT1), and Leishmania donovani (MHOM/IN/1980/DD8).
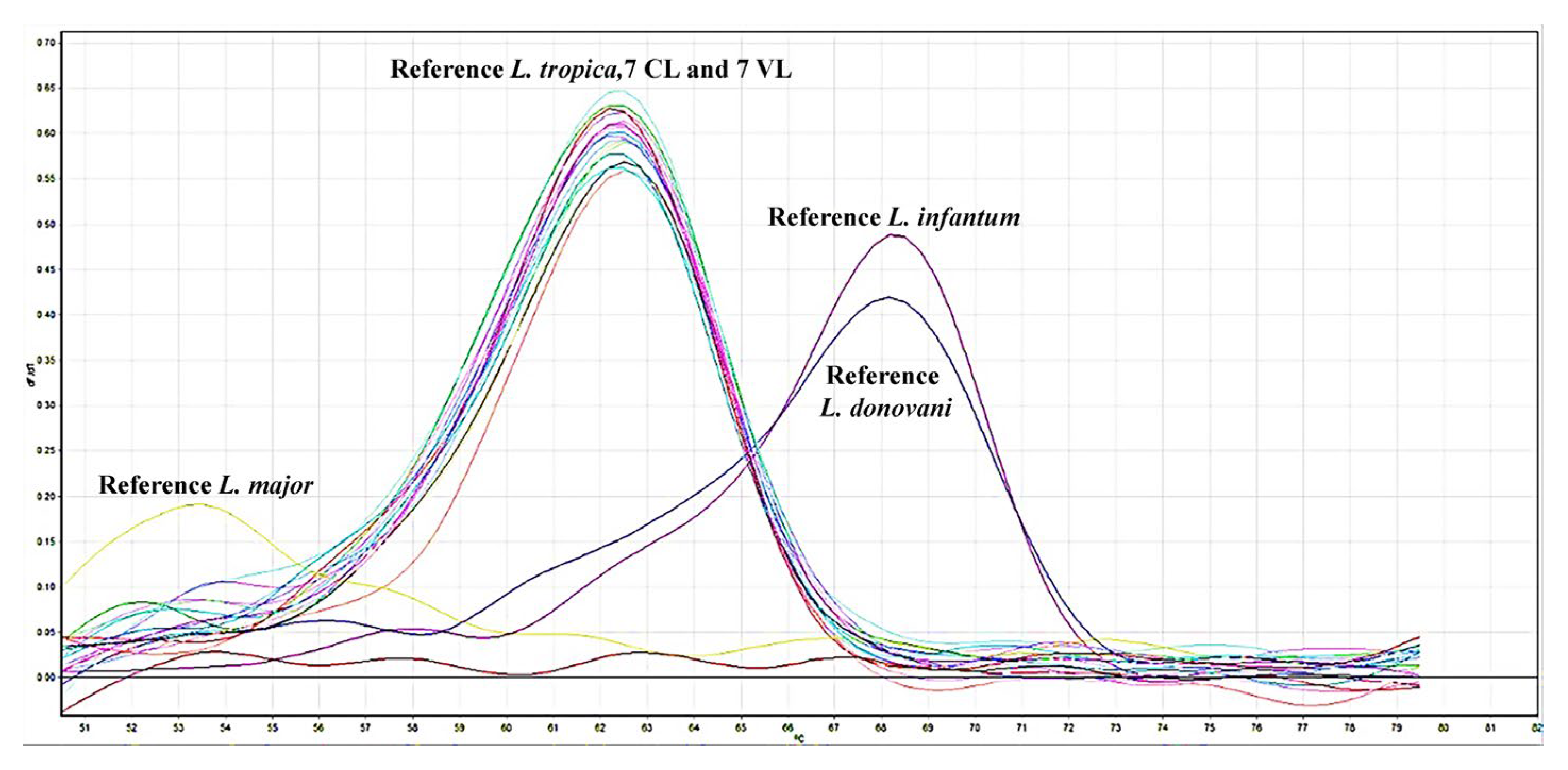
Figure 3.
mRNA expression fold changes of genes associated with viscerotropism. Significant increases in mRNA expression were observed in visceral leishmaniasis (VL) isolates compared to reference L. tropica. Control: reference isolate L. tropica, reference isolate L. infantum, VL: visceral leishmaniasis group, CL: cutaneous leishmaniasis group, PRDX1/2: Peroxidoxin 1 and 2, OPB: Oligopeptidase B, M32: Metallo-peptidase, Clan MA(E), family M32, COX IV: Cytochrome C Oxidase subunit IV, SCOT: Succinyl-CoA:3-ketoacid-coenzyme A transferase, PK: Pyruvate kinase, ns: not significant, *: 0.0133, ***: 0.0006, ****: <0.0001.
Figure 3.
mRNA expression fold changes of genes associated with viscerotropism. Significant increases in mRNA expression were observed in visceral leishmaniasis (VL) isolates compared to reference L. tropica. Control: reference isolate L. tropica, reference isolate L. infantum, VL: visceral leishmaniasis group, CL: cutaneous leishmaniasis group, PRDX1/2: Peroxidoxin 1 and 2, OPB: Oligopeptidase B, M32: Metallo-peptidase, Clan MA(E), family M32, COX IV: Cytochrome C Oxidase subunit IV, SCOT: Succinyl-CoA:3-ketoacid-coenzyme A transferase, PK: Pyruvate kinase, ns: not significant, *: 0.0133, ***: 0.0006, ****: <0.0001.
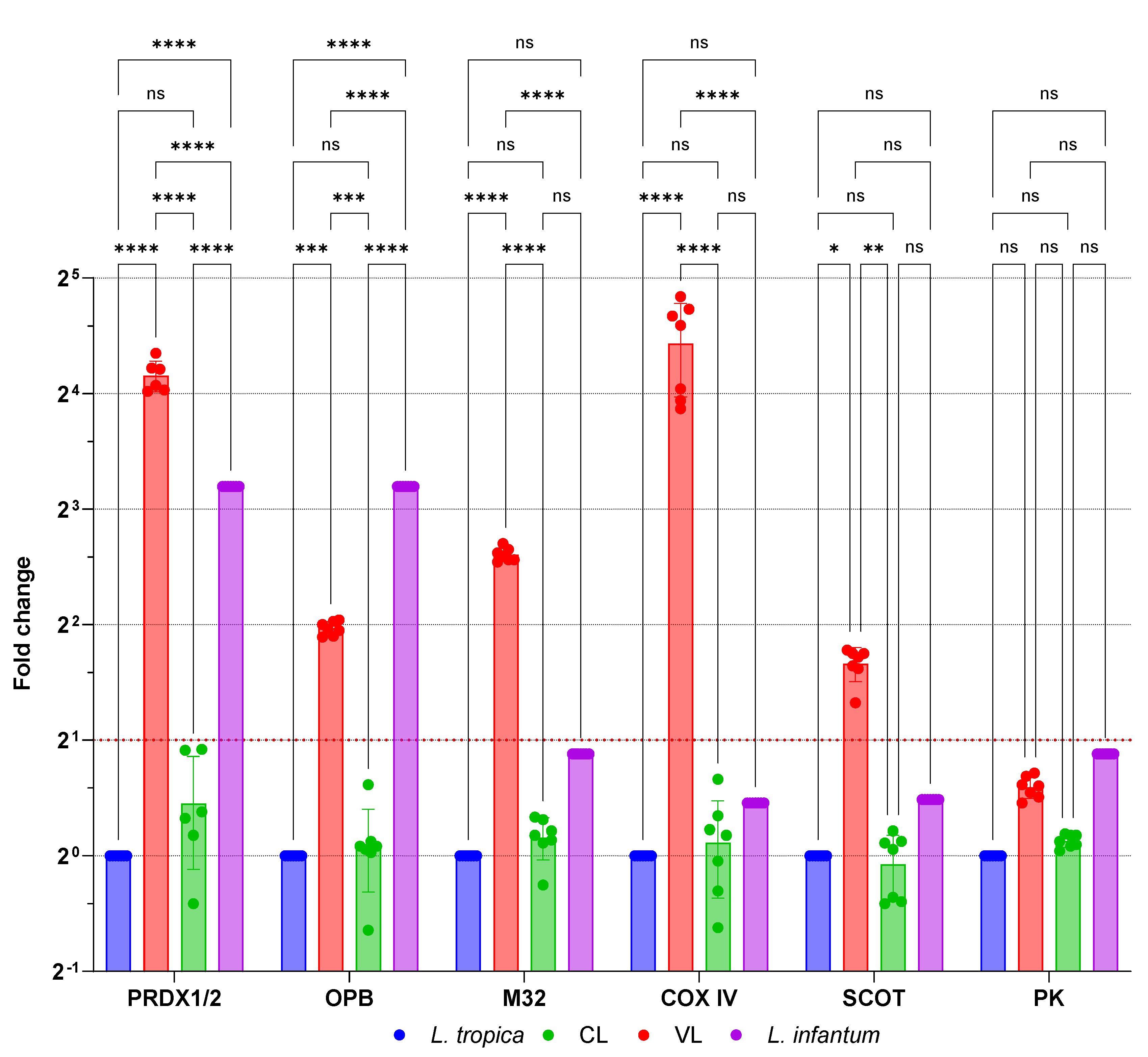
Figure 4.
Gene Ontology (GO) classification of genes associated with viscerotropism in L. tropica based on their molecular functions. Categories include catalytic activity, transferase activity, hydrolase activity, oxidoreductase activity, and antioxidant activity. (a) Biological Process, (b) Cellular Component, (c) Molecular Function.
Figure 4.
Gene Ontology (GO) classification of genes associated with viscerotropism in L. tropica based on their molecular functions. Categories include catalytic activity, transferase activity, hydrolase activity, oxidoreductase activity, and antioxidant activity. (a) Biological Process, (b) Cellular Component, (c) Molecular Function.
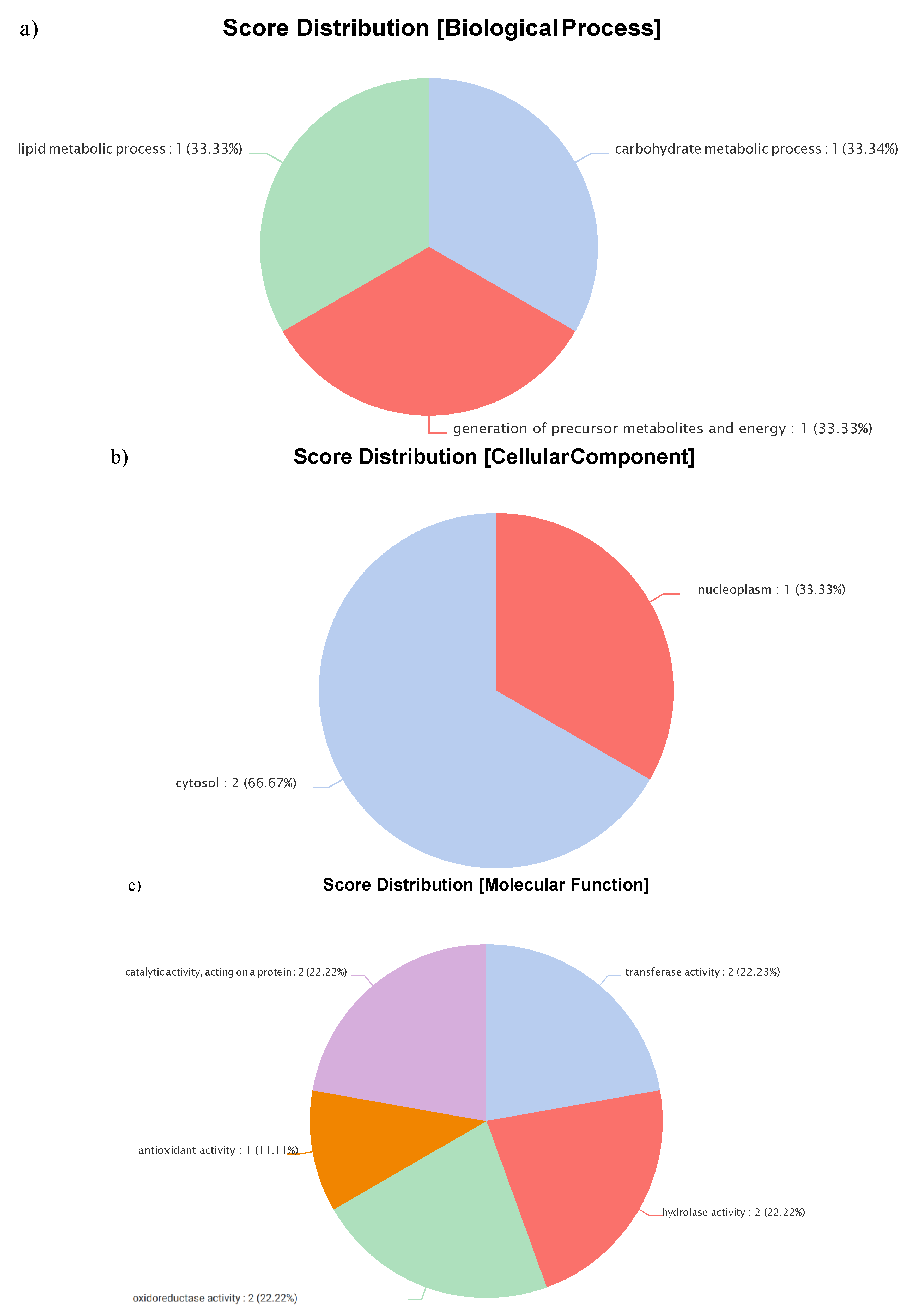
Table 1.
Clinical histories and Leishmania species genotyping results of patients included in the study.
Table 1.
Clinical histories and Leishmania species genotyping results of patients included in the study.
| Patient Code | Gender | Age | Region | Symptom | Genotype of Amastigotes from Clinical Samples | Genotype of Promastigotes Grown in Culture |
|---|---|---|---|---|---|---|
| V1 | M | 7 | Aegean | Fever, rapid weight loss, hepatosplenomegaly, pancytopenia, nausea, diarrhea | L. tropica | L. tropica |
| V2 | F | 10 | Aegean | Weight loss, weakness, fever, anorexia, pancytopenia, nose and tooth bleeding, hepatosplenomegaly, growth retardation | L. tropica | L. tropica |
| V3 | F | 12 | Aegean | Swelling in the left upper quadrant, night sweats, anorexia, rapid weight loss (5 kg in the last 1 month), splenomegaly, pancytopenia | L. tropica | L. tropica |
| V4 | M | 20 | Aegean | Fever, hepatosplenomegaly, anorexia, spleen infarction, general condition disorder, diarrhea, pancytopenia, nose and gum bleeding, general condition disorder | L. tropica | L. tropica |
| V5 | M | 50 | Aegean | Fever, diarrhea, nausea, anorexia, dizziness, weakness, thrombocytopenia, leukopenia, hepatosplenomegaly | L. tropica | L. tropica |
| V6 | M | 53 | Aegean | Fever, anorexia, pancytopenia, hepatosplenomegaly, weight loss, malaise, diarrhea | L. tropica | L. tropica |
| V7 | M | 55 | Mediterranean | Fever, weight loss, pancytopenia, hepatosplenomegaly, anorexia, nausea | L. tropica | L. tropica |
| C1 | M | 10 | Aegean | Nodular dry type lesion on the right cheek for 3 months | L. tropica | L. tropica |
| C2 | M | 11 | Aegean | Plaques with vascularity on the skin of the right zygomatic region of the face, dry type lesion present for 24 months | L. tropica | L. tropica |
| C3 | F | 17 | Aegean | Dry type lesion on the right arm for 3 months | L. tropica | L. tropica |
| C4 | F | 18 | Aegean | Dry type lesion on the right side of the nose for 7 months | L. tropica | L. tropica |
| C5 | M | 25 | Aegean | Dry type lesion on the right cheek for 8 months | L. tropica | L. tropica |
| C6 | F | 35 | Aegean | Dry type lesion under the eye on the left cheek for 6 months | L. tropica | L. tropica |
| C7 | F | 47 | Aegean | Dry type, itchy lesion on the tip of the nose for 12 months | L. tropica | L. tropica |
Table 3.
Summary of missense mutations identified through targeted next-generation sequencing (NGS) in Leishmania tropica isolates from VL and CL patients, highlighting differences potentially associated with disease tropism.
Table 3.
Summary of missense mutations identified through targeted next-generation sequencing (NGS) in Leishmania tropica isolates from VL and CL patients, highlighting differences potentially associated with disease tropism.
| Viscerotropism associated genes | Reference L. tropica | CL | VL |
|---|---|---|---|
| Peroxidoxin 1 | _ | _ | _ |
| Peroxidoxin 2 | _ | _ | _ |
| Oligopeptidase B | c.1031A>G (p.Asp344Gly) c.1306C>G (p.Pro436Ala) |
c.1031A>G (p.Asp344Gly) c.1306C>G (p.Pro436Ala) |
_ |
| Metallo-peptidase, Clan MA (E), M32 family protein | c.169G>T (p.Ala57Ser) | c.169G>T (p.Ala57Ser) | _ |
| Cytochrome C Oxidase subunit IV | _ | _ | _ |
| Succinyl-CoA:3-ketoacid-coenzyme A transferase | _ | _ | _ |
| Pyruvate kinase | c.1362G>A (p.Glu454Lys) | c.1362G>A (p.Glu454Lys) | c.1362G>A (p.Glu454Lys) |
Reference L. tropica (MHOM/AZ/1974/SAF-K 27).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



