
Submitted:
29 January 2025
Posted:
30 January 2025
You are already at the latest version
Abstract
Leishmaniases are zoonotic vector-borne diseases caused by a wide variety of Leishmania species with complex transmission cycles involving different reservoirs, potential new hosts and vectors. Similarly, to other eukaryotes, Leishmania release extracellular vesicles (LEVs) to play important initial interactions that are crucial to modulate the subsequent systemic immune response on the establishment of infection in humans and others hosts like dogs. Recent studies in endemic areas of Brazil concluded that canine infections were predominantly due to L. amazonensis and not restricted to L. infantum (syn. Leishmania chagasi). Under these premises, the diagnosis of leishmaniasis needs to be improved with the identification of current etiological agent and the histopathologic features, highlighting the differential diagnosis and the molecular components of LEVs. In this way, the dual aim of that study is to register collected observations of natural canine infections, enhancing the relevance of differential diagnosis in companion animals and inserting in vitro results in the field of LEVs that still research gaps to be filled to understand the mechanisms and biological aspects involving the parasite-host interactions. Therefore, improve these studies of Parasitology research is important for diagnostic, prognostic, treatment advances and continuing need for global prevention, control, elimination/eradication of these parasitic infections.
Keywords:
Leishmania infectivity in mammalian hosts
; inserts of leishmaniases and coinfections in companion animals
; clinical and histopathologic features of natural zoonotic canine leishmaniasis
; cell communication by parasite-derived extracellular vesicles
; in vitro models advance to target parasite-secreted lipid bound vesicles
1. Introduction
Leishmania is an important genus of parasitic protozoa due to their medical and veterinary significance. They have made efficient adaptations to survive in a large diversity of hosts or reservoirs. Therefore, they survive in a variety of vertebrates such as dogs (the main reservoir of these parasites, though the causative agent of canine leishmaniasis identified is not restricted to Leishmania infantum), domestic cats, wild animals, and synanthropic animals, while being transiently infectious to humans (Figure 1) [1]. According to the World Health Organization (WHO), the ability of Leishmania to infect multiple hosts has facilitated their spread through phlebotomine sandflies. During their digenetic life cycle, female phlebotomine sandflies of several species of Phlebotomus and Lutzomyia act as vectors in the zoonotic transmission of Leishmania between mammals and sandflies (Figure 1) [1,2,3,4,5,6].
It is, therefore, important to draw the attention of veterinarians to transmission and the risk of transmission as clinical keys to establish the signs and pathological abnormalities of Leishmania infection, despite the difficulty in determining the incubation period. This is particularly true given that intracellular parasites may appear only after several weeks or months, typically with shorter incubation periods for cutaneous forms (Figure 2) [11,12]. Concurrently, with the spread of leishmaniasis, recent studies reveal that the most popular pet animals worldwide are at risk of acquiring leishmaniasis coinfection with other emerging zoonotic parasites, viruses, and other infectious agents. These findings underscore the need for further studies on extracellular vesicle production and the simultaneous cascade effects of molecular interactions on the host immune system (Section 1, Supplementary material) [1,13,14,15]. In this sense, coinfected companion animals (e.g., dogs and cats) are important elements of the Leishmania life cycle, and at the same time, they contribute to the transmission of other disease agents of medical and veterinary importance. These infections can cause sensible health deterioration (e.g., weight loss, anemia, and low immune resistance), leading to secondary infections, increased morbidity, and even the death of infected animals [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19]. What hypotheses should still be considered in the Leishmania zoonotic cycle? a. Hosts have different biochemical, immunological, behavioral, and other defense mechanisms to ward off parasites, and many features of parasites are adaptations to overcoming these defenses. b. The different levels of Leishmania virulence depend on the evolution of both the host and the parasite. c. The susceptibility of potential new hosts and vectors can vary under interconnected factors. d. Even more impressive than the life cycles of Leishmania are their many defenses against the antibodies of vertebrate hosts. These antibodies are proteins that recognize and bind to foreign proteins and other molecules in polydisperse complex biological samples (e.g., extracellular vesicles surrounded by a lipid layer of the parasite membrane) [1,14,15,17,19]. The clinical signs of leishmaniasis are directly related to the immune response of the infected dog; however, it is estimated that more than half of infected dogs do not manifest clinical signs of the disease [20]. Despite the development of some veterinary vaccines, the lack of effective vaccines and drug treatments for humans has led to the increase of this major health problem worldwide [1,2,12]. These parasites exhibit a high level of genetic variability in vivo and a propensity for rapid evolution in vitro. Infection is established through heterogeneous extracellular vesicles released by the parasite, which contain many molecules that modulate the host immune responses. These molecules were generically referred to as Leishmania extracellular vesicles or LEVs by Gabriel et al. (2021) [1,21].
In recent years, there has been increasing interest in the isolation and biological and functional characterization of the lipoproteic vesicles released by Leishmania (LEVs). This is due to their apparent potential for the development of effective diagnostic and therapeutic approaches, including the prediction of the outcome of the interaction between cells [1,21,22]. Technology advances have led to the production of new tools that make progress in the study of extracellular vesicle biology and its applications. However, mechanisms of the selective packaging of LEVs are still poorly understood. Moreover, there is no consensus on the differential isolation and characterization of these extra-cellular vesicles or the ultrasensitive detection of specific sub-types, biomarkers, and biogenesis [1,21]. Recent studies indicated that the lipid composition of extracellular vesicles influences their biogenesis, cargo sorting, interactions with target cells, and functional effects on recipient cells [22,23,24,25]. In this way, there is an emerging focus on lipids in LEVs. Nevertheless, still there are gaps in the research for understanding the mechanisms and biological aspects involved in Leishmania–host interactions [1,21,22]. We note that lipid bodies are organelles distributed in the cytoplasm of eukaryotic cells, largely associated with lipid storage in the past. However, they are now recognized as dynamic and functionally active organelles, involving in a variety of functions such as lipid metabolism, cell signaling, and inflammation. These lipid bodies have been identified inside dermal macrophages and are actively formed within heart macrophages following infection of another protozoan parasite, Trypanosoma cruzi [22,23]. The study emphasizes the importance of diagnosing natural canine leishmaniasis. In addition, it compares the findings with in vitro results related to the endocytosis and exocytosis processes that occur during the disease’s pathogenesis. This comparison complements the authors’ previous research on lipids in extracellular vesicles (LEVs). The ultimate goal is to contribute to future nanotechnological research within the One Health framework. This will further contribute to the development of new management approaches that improve prophylactic measures, ensure effective diagnosis and prognosis for both acute and chronic leishmaniases, and advance treatment options.
In essence, this study aims to bridge the gap between observations of natural infection and laboratory findings to enhance the overall understanding and management of leishmaniasis.
2. Materials and Methods
2.1. Quantification Method of Lipid Bodies in the Promastigote of Leishmania Amazonensis
BODIPY™ 493/503 (4,4-Difluoro-1,3,5,7,8-Pentamethyl-4-Bora-3a, 4a-Diaza-s-Indacene Molecular Probes Invitrogen™, Thermo Fisher Scientific, Waltham, MA) is a lipophilic fluorophore used for the identification of neutral lipids (reserve lipids) present in abundance in the form of intracellular lipid bodies. It is used for the quantification and analysis of these structures by flow cytometry [24] and for morphological observation by fluorescence microscopy. For lipid quantification, promastigotes of L. amazonensis (strain MHOM/BR/26361) were maintained in culture in RPMI 1640 medium supplemented with 10% SBF at 27ºC. After 24, 48, and 72 hours of culture, the cells were washed with PBS (pH 7.2) and incubated with BODIPY™ 493/503 at a concentration of 10 μg/mL for 30 minutes in the absence of light [25]. After incubation, the cells were analyzed using the BD FACSCanto IITM flow cytometer (BD Biosciences, San Jose, CA). A total of 10,000 events were collected for each sample, and the average flowering intensity data were analyzed using the GraphPad Prism program (version 6.01).
2.2. Transmission Electron Microscopy of Macrophages Infected with Leishmania Amazonensis
Peritoneal macrophages from mice (2x106/mL) were cultured and infected with L. amazonensis promastigotes (1:10 ratio) in the stationary phase [26]. After 72 hours of infection, the cells were washed with PBS (pH 7.2). Then, the cells were fixed for one hour with 2.5% glutaraldehyde type II (70%), 4% paraformaldehyde, and 2.5% sucrose in 0.1 M sodium cacodylate buffer (pH 7.2). After fixation, the cells were incubated in a solution containing 1% osmium tetroxide and 0.8% potassium ferrocyanide for one hour. Following this, the cells were dehydrated in a graded series of acetone for ten minutes at concentrations of 50%, 70%, 90%, and three times in 100%. After dehydration, the cells were slowly impregnated with Epon® resin (2:1, 1:1, and 1:2 ratios of 100% acetone: Epon®, Westlake Epoxy, Houston, TX). Subsequently, the material was placed in pure Epon® for six hours and finally polymerized at 60°C for 48 hours. The blocks were cut using an ultramicrotome (Leica EM UC6). The sections obtained were contrasted with 5% uranyl acetate and lead citrate, and then observed using a JEOL Transmission Electronic Microscope (TEM, JEOL Ltd., Akishima, Tokyo, Japan).
2.3. Transmission Electron Microscopy of Tissues from Male BALB/c Mice Infected with Leishmania Amazonensis
Male BALB/c mice (8–10 weeks old) [27] were infected subcutaneously with approximately 1×106 L. amazonensis promastigotes in their stationary form. After six weeks, the mice were euthanized, and the lesion tissues were processed for TEM analysis. The tissues were fixated for one hour with 2.5% glutaraldehyde type II (70%), 4% paraformaldehyde, and 2.5% sucrose in 0.1 M sodium cacodylate buffer (pH 7.2). Following fixation, the cells were incubated in a solution containing 1% osmium tetroxide and 0.8% potassium ferrocyanide for one hour. After this period, the cells were then dehydrated in a growing series of acetone for ten minutes at concentrations of 50%, 70%, 90%, and three times in 100%. After dehydration, the cells were slowly impregnated with Epon® resin at ratios of 2:1, 1:1, and 1:2 (100% acetone: Epon®). Subsequently, the material was placed in pure Epon® for six hours and finally polymerized at 60ºC for 48 hours. The blocks were cut using an ultramicrotome (Leica EM UC6, Leica Microsystems, Wetzlar, Hesse, Germany). The obtained sections were contrasted with 5% uranyl acetate and lead citrate and then observed using JEOL TEM.
2.4. Leishmania Diagnosis: Some Findings of Natural Canine Infection
In this section, we present information from the case report of “Jimmy”, a healthy, male, one-year-old dog (Shih Tzu breed), who exhibited an eye infection during clinical examination.
2.4.1. Careful Anamnesis and Clinical and Routine Laboratory Tests
To set the standards, monitoring of the following recommended clinicopathological parameters was considered: clinical history, complete physical examination, identification of clinical signs according to the four clinical stages (i.e., mild, moderate, severe, and very severe disease), and laboratory tests (including a complete blood count (CBC), biochemical profile, serology, and molecular techniques) [11].
2.4.2. Impression Cytology of the Infected Ocular Surface
The superficial layers of the ocular surface epithelium were removed using a minimally invasive, easy-to-perform method of impression cytology. Reliable information was obtained about the sampled area with minimal discomfort to the animal. Conjunctival cytological sample was obtained and analyzed using a light microscopy [28].
2.5. Molecular Analysis of Canine Clinical Samples by Polymerase Chain Reaction
Following the protocols tested by Galvão et al., 5 mL of canine blood samples were collected via venipuncture in dry and ethylenediaminetetraacetic acid (EDTA)-containing tubes for PCR analysis. Moreover, secretions from the auditory duct, mucous membrane of the anus, oral mucosa, whole blood, preputial area, and ocular of “Jimmy” were collected using sterile cotton swabs (NeoLab®, neoLab Migge GmbH, Heidelberg, Baden-Württemberg, Germany) and frozen until DNA extraction. Preliminary studies included blood samples and ocular swabs from 130 dogs: 67 males (51.5%), 60 females (46.1%), and three animals (2.4%) with no gender information. After adding the samples from “Jimmy,” the final total was 131 animals from endemic areas of the State of Pará in the Amazon Region of Brazil. This area covers 1,284,042 km2 and is situated between the coordinates 04°20′45″ N and 09°50′27″ S and 46°03′18″ W [29,30]. For molecular diagnosis, DNA was extracted from the canine blood samples according to recommendations of the manufacturer of the AxyPrep™ Blood Genomic DNA Miniprep Kit (Axygen, Axygen Scientific, Inc., a subsidiary of Corning Incorporated, Union City, CA). For the extraction of DNA from secretions collected using swabs, a technique with NaCl was employed, yielding a final volume of 20 μL. After extraction, molecular tests were carried out using polymerase chain reaction (PCR), targeting the small subunit ribosomal ribonucleic acid (SSU rRNA) gene and extra-chromosomal kinetoplastid DNA (kDNA), both specific to Leishmania sp. and L. infantum (Table 1). For the detection of the SSU rRNA gene specific to Leishmania sp., the first PCR was performed with primers S4 (5’-GATCCAGCTGCAGGTTCACC-3’) and S12 (5’-GGTTGATTCCGTCAACGGAC-3’). Reactions were performed with a final volume of 25 µL containing 1× PCR buffer, 2 mM MgCl2, 0.2 mM dNTPs, 0.2 µM of each primer, and 1 U-Taq DNA polymerase. DNA was first denatured at 94oC for three minutes and then cycled 35 times at 94oC (one minute), 50oC (one minute), and 72oC (one minute). A final extension of seven minutes was performed at 72oC. The fragment (520 pb) produced by the S4/S12 PCR was used in a nested PCR with primers S17 (5’-CCAAGCTGCCCAGTAGAAT-3’) and S18 (5’-TCGGGCGGAT AAAACCC-3’), which is specific to the genus Leishmania. The reaction was performed under the same conditions as described earlier. The S4/S12 PCR product was denatured at 94oC (four minutes) and cycled 30 times; each cycle took place at 94oC (one minute), 55oC (one minute), and 72oC (30 seconds). Oligonucleotides S17 and S18 produced a 490-base-pair (bp) fragment. L. infantum-specific primers Leish 1 (5’-AACTTTTCTGGTC CTCCGGGTAG-3’) and Leish 2 (5’-ACCCCCAGTTTCCCGCC-3’) were used to amplify a 120-bp fragment of the Leishmania kinetoplast DNA minicircle. PCR was conducted in a 25 mL final reaction mixture containing PCR buffer 1, 0.150 mM dNTPs, 2 mM MgCl2, 0.2 mM of each primer, and 1 U-Taq DNA polymerase. The thermal cycling profile was as follows: 94oC (three minutes), followed by 35 cycles at 94oC (30 seconds), 60oC (30 seconds), and 72oC (30 seconds), with a final extension at 72oC (five minutes). Products of the SSU rRNA gene (490 bp) and kDNA (120 bp) were applied to 1.5% and 2.5% agarose gels, respectively, in tris-acetate-EDTA (TAE) buffer, stained with Diamond™ Nucleic Acid Dye (Promega Corporation, Madison, WI), and later submitted to electrophoresis and visualized under ultraviolet light. An Applied Biosystems thermocycler (Model 2720, Thermo Fisher Scientific) was employed for the PCR reactions [29,30].
After obtaining the amplicons, the PCR products were purified for the kDNA gene and nested PCR for the SSU-rDNA gene using ExoSAP-IT™ PCR Product Cleanup Reagent (GE Healthcare, Fisher Scientific, Waltham, MA). Sequencing was performed with the aid of the ABI 3500XL automatic DNA analyzer (Applied Biosystems™, Fisher Scientific) in combination with BigDye® Terminator v3.1 (Fisher Scientific). The nucleotide sequences obtained were edited and aligned using the BioEdit software (Technelysium Pty Ltd, Brisbane, Queensland, Australia). After comparison with other GenBank sequences (BLASTn: basic local alignment tool survey for nucleotides), the SSU-rDNA sequence was aligned with the set of Leishmania sequences available at GenBank NCBI NIH using the Clustal program W 1.6 (European Molecular Biology Laboratory (EMBL), Heidelberg, Germany). This was done to identify the single nucleotide polymorphisms (SNPs) and insertions–deletions (INDELs). Phylogenetic relationships were constructed with the Neighbor-Joining method, and the p-distance method was used to calculate genetic distances. The monophyletic groups were supported by the Bootstrap 2000 method using the MEGA 6 program. To test whether a relationship exists between the diagnostic methods, a contingency table was used, and the BioEstat 5.0 software (Belém, Pará, Brazil) was employed for the results considered as relevant (p<0.05) [29,30].
3.Results
3.1. Endocytosis and Exocytosis Processes of Leishmania: Macrophages Infected by Promastigotes, Mice Tissues Infected by Amastigotes, and the Quantification of Lipid Bodies in Promastigotes of Leishmania Amazonensis
The knowledge of specific cargo molecules identified within LEVs suggests that their function as adjuvants—like in immune responses—is a possible key advantage for infection establishment and disease progression. They induce, at the same time, quantitative and qualitative changes in the protein content of infected host cell extracellular vesicles (Figure 2 and Figure 3). However, many challenges remain in the extraction, purification, and analysis of polydisperse isolates containing LEVs.
3.2. Results of Polymerase Chain Reaction Tests for the Biological Samples Collected from 130 Dogs in the Amazonian Endemic Area and the Results Obtained from Our Case Report “Jimmy”
The variables of sex, age, and breed were analyzed regarding the presence of infection. None of these showed a statistical association; however, 47 dogs aged 1–8 years (36.2%) had positive results for Leishmania infection (Table 2).
When the infected animals were evaluated for the presence of at least two symptoms suggestive of leishmaniasis, it was observed that 65/72 (90.3%) of the dogs were symptomatic and 7/72 (9.7%) were asymptomatic, in addition to the presence of 28 symptomatic animals without infection (Table 3).
Regarding the detection of the parasite’s DNA in the different biological samples (blood and swabs), 58 (44.61%) dogs were positive in blood and 54 (41.53%) dogs were positive in the swab samples. Thus, 43/130 (33.1%) samples were positive in both types of samples; 15/58 (25.9%) of the blood samples and 11/72 (15.3%) of the swab samples were detected as positive (Table 4).
Figure 3.
Quantification of lipid bodies in Leishmania amazonensis at different culture times: (a) histogram of each experimental group; (b) overlay histogram demonstrating increased fluorescence intensity of BODIPY™ 493/503-labeled cells at 24, 48, and 72 hours of culture; and (c) graphic representation of BODIPY™ 493/503 labeling in Leishmania promastigotes. ****p=0.0001, ***p<0.001 (ANOVA and Tukey’s post hoc test).
Figure 3.
Quantification of lipid bodies in Leishmania amazonensis at different culture times: (a) histogram of each experimental group; (b) overlay histogram demonstrating increased fluorescence intensity of BODIPY™ 493/503-labeled cells at 24, 48, and 72 hours of culture; and (c) graphic representation of BODIPY™ 493/503 labeling in Leishmania promastigotes. ****p=0.0001, ***p<0.001 (ANOVA and Tukey’s post hoc test).
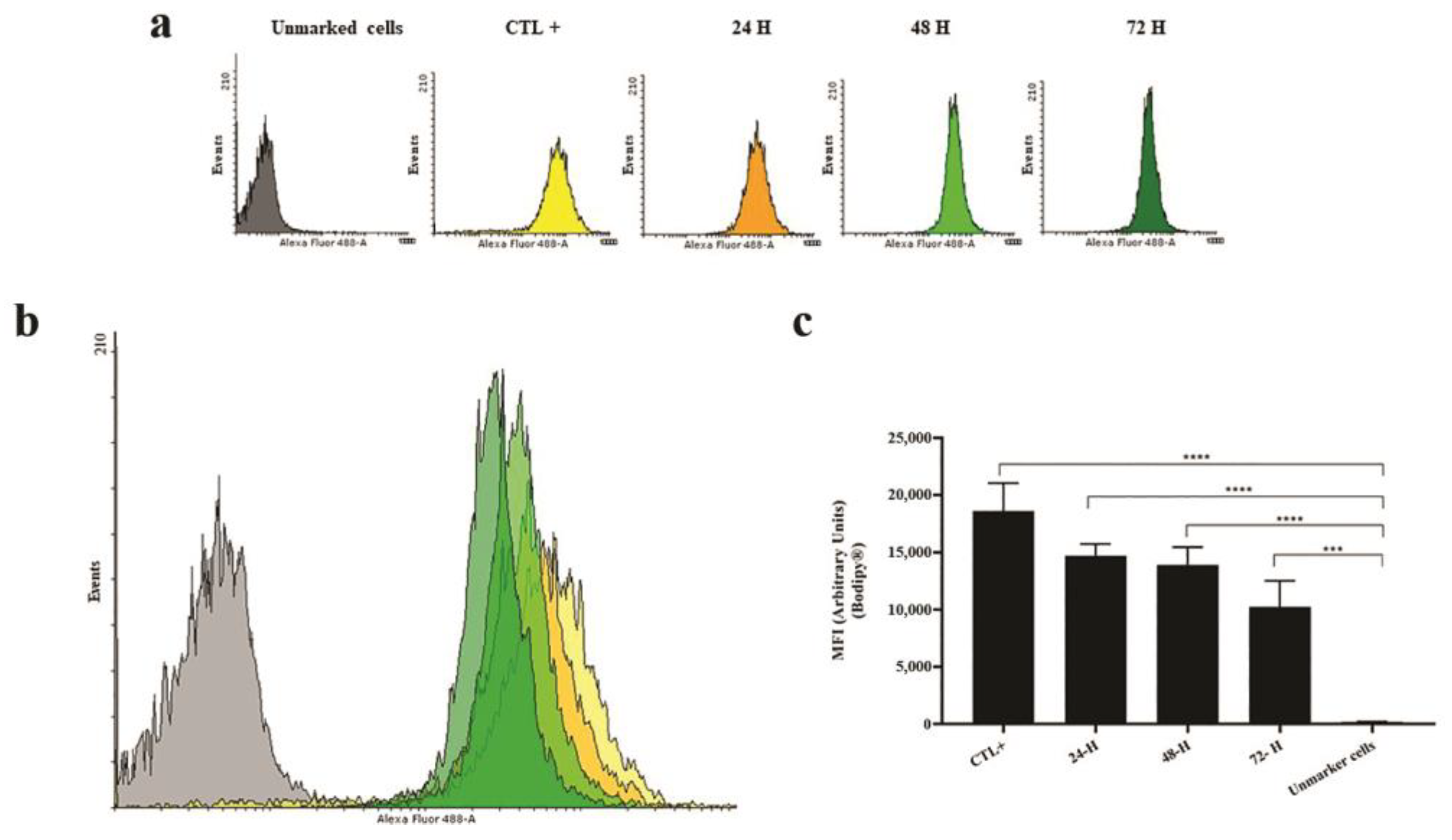

Table 2.
Distribution of Leishmania sp. across different age groups.
| Age | Infection (%) | Total | |
| Present | Absent | ||
| 0–12 months | 14 (10.7) | 10 (7.7) | 24 |
| 1–8 years | 47 (36.2) | 40 (30.8) | 87 |
| Over eight years | 7 (5.3) | 6 (4.6) | 13 |
| Not informed | 4 (3.1) | 2 (1.5) | 6 |
| Total animals | 72 | 58 | 130 |
Table 3.
Prevalence of Leishmania sp. in dogs with symptomatic and asymptomatic leishmaniases.
| Infection | Symptomatic (%) | Asymptomatic (%) | Total |
|---|---|---|---|
| Present | 65 (50) | 7 (5.4) | 72 |
| Absent | 28 (21.5) | 30 (23.1) | 58 |
| Total | 93 | 37 | 130 |
χ2 = 25.80, p<0.001; odds ratio, 9.95; p<0.001, 95% CI: 3.9–25.3.
Table 4.
Detection of Leishmania sp. DNA in different types of biological samples.
| Biological sample | Blood (+) | Blood (-) | Total |
|---|---|---|---|
| Swab (+) | 43 (33.1%) | 11 (8.5%) | 54 |
| Swab (-) | 15 (11.5%) | 61 (46.9%) | 76 |
| Total | 58 | 72 | 130 |
(+) Positive; (-) negative. χ2= 43.44; p<0.001.
For the detection of Leishmania sp. in the animals studied, different types of oligonucleotides were used. One of these is specific for the detection of kDNA and the others for SSUr-rDNA. In the present study, when assessing the sensitivity of detection of the parasite’s DNA using the different oligonucleotides, a higher number of positive samples were detected using the kDNA marker than using SSUr-rDNA. The success in detecting Leishmania infection was comparable between the markers, and there was no statistical difference regarding the efficiency of the markers relative to each other, indicating that both have similar sensitivity (Table 5).
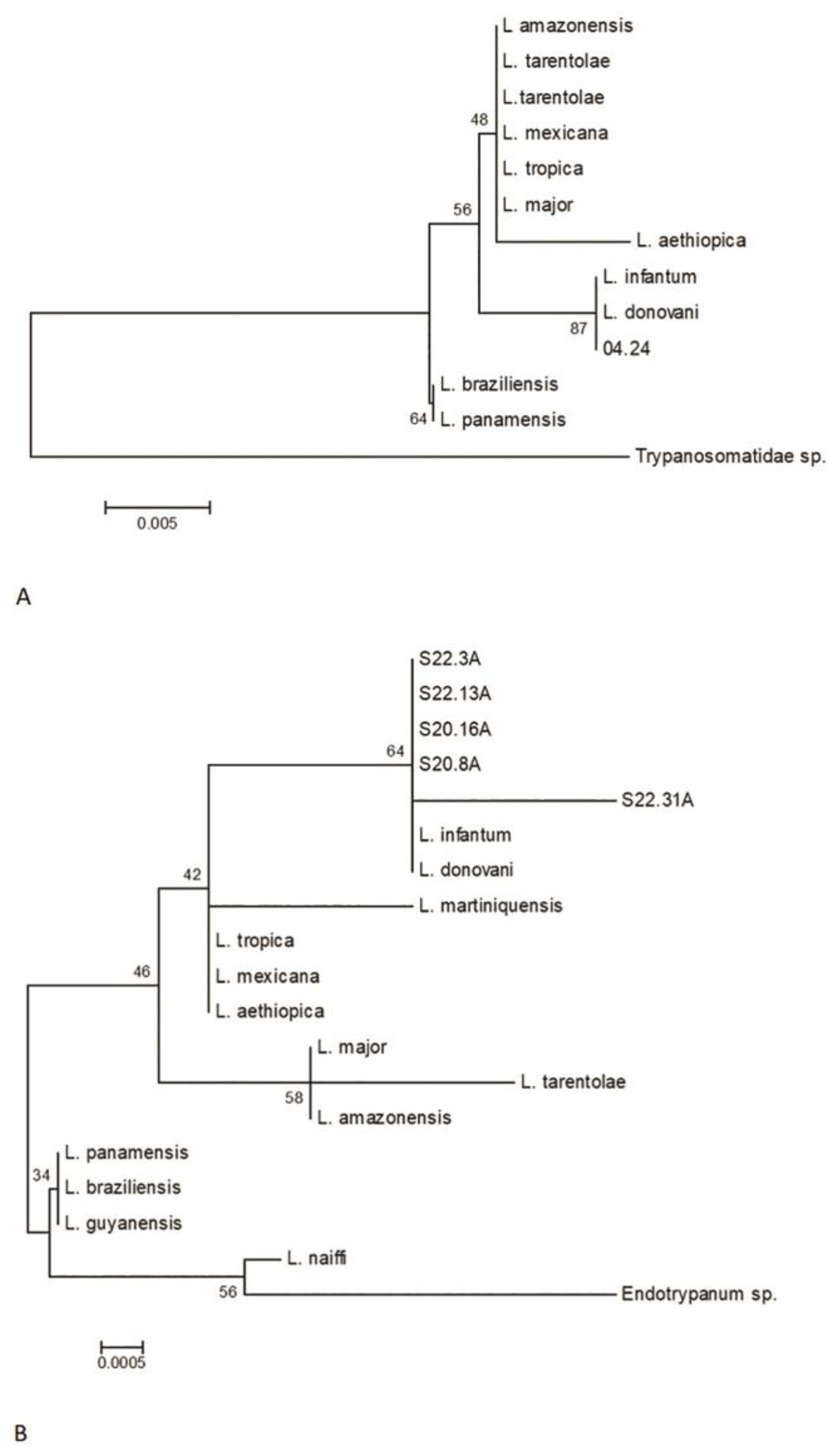
The amplified products of the SSUr-rDNA segment of the 18S RNA obtained were sequenced to determine the phylogenetic relationships of the samples. Using the initiators R223/R333, 26 out of 72 samples yielded the best sequences, all of which grouped within the donovani complex, resembling L. infantum (Figure 4).
The samples of the study showed nucleotide variation in the multiple alignments, including the presence of degenerate nucleotides (Y = C/T and R = A/G). These variations occurred at a frequency of 1–3 within the analyzed segment, located between positions 121 and 316 of the sequence (Table 6). Only one sample showed a pyrimidine transition mutation (C→T) at position 202. At this point, it is important to highlight that these samples come from different areas of the State of Pará. Evolutionary analyses were conducted using MEGA 6, with the augmentation products from three primers with A-markers R223/R333 and B-markers S17/S18 (Figure 5).
Figure 4.
The Neighbor-Joining method is proposed for reconstructing phylogenetic trees from evolutionary distance data. (a) For constructing the phylogenetic tree, the obtained sequence was compared with sequences from GenBank. (b) The amplified products of the S17/S18 initiators included 47 out of the 72 sequences analyzed, most of which grouped together and resembled Leishmania infantum. Additionally, it was observed that four samples (S20.16, S22.3, S22.13, and S22.31) show genetic variability. Notably, the S22.31 sample showed a higher genetic distance compared to the others.
Figure 4.
The Neighbor-Joining method is proposed for reconstructing phylogenetic trees from evolutionary distance data. (a) For constructing the phylogenetic tree, the obtained sequence was compared with sequences from GenBank. (b) The amplified products of the S17/S18 initiators included 47 out of the 72 sequences analyzed, most of which grouped together and resembled Leishmania infantum. Additionally, it was observed that four samples (S20.16, S22.3, S22.13, and S22.31) show genetic variability. Notably, the S22.31 sample showed a higher genetic distance compared to the others.
3.3. Leishmania Diagnosis: Some Findings from a Natural Canine Infection After Anamnesis and Clinical and Routine Laboratory Tests
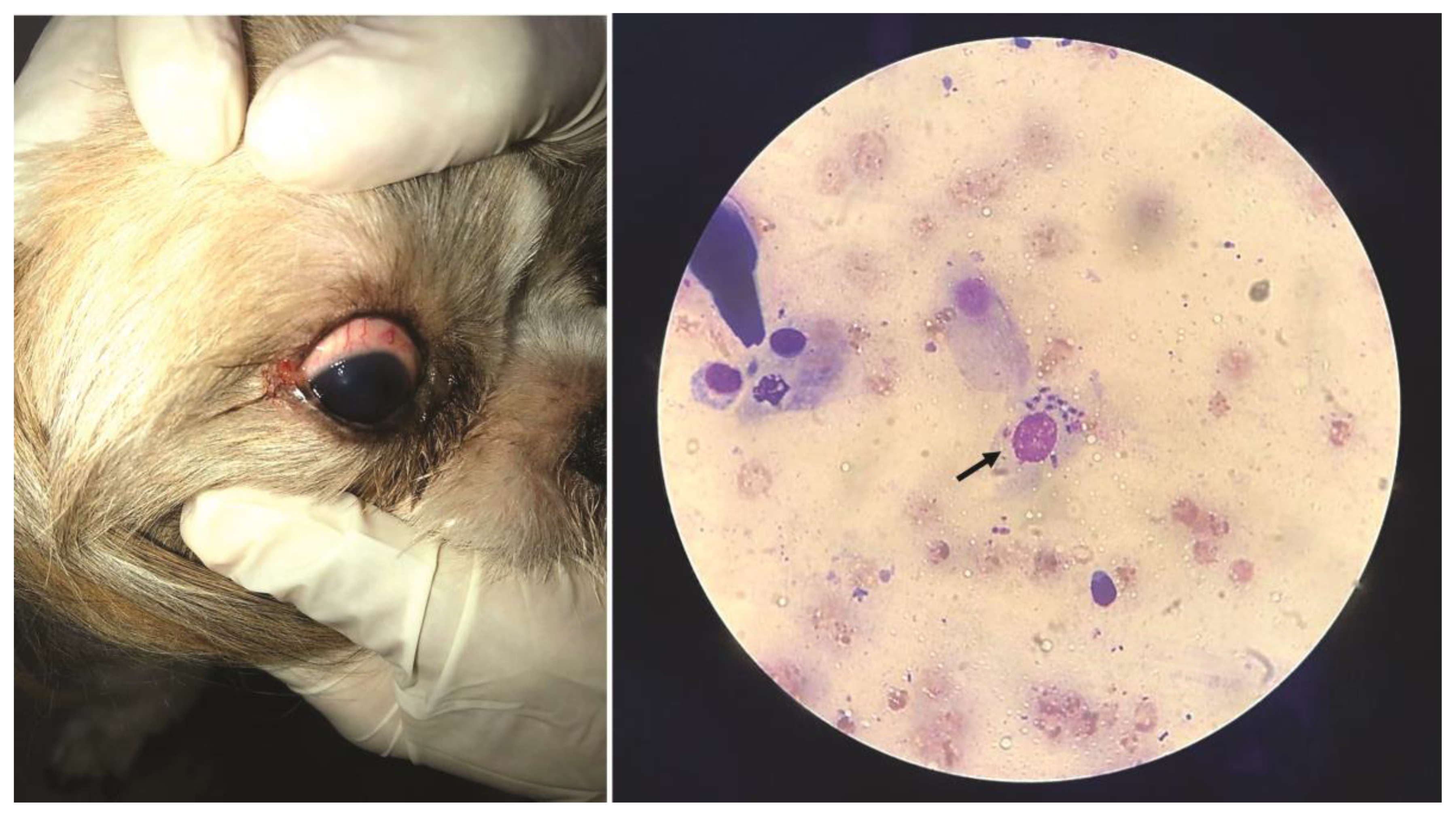
The selected case report pertains to “Jimmy,” a young, healthy dog with a single-eye infection detected during clinical examination (Figure 6). Despite the absence of other symptoms, conjunctival cytology and additional biological samples analyzed through routine laboratory tests confirmed the diagnosis of leishmaniasis. It is important to note that there was no record of previous vaccination against leishmaniasis in this clinical case; however, the region is endemic for the disease (Figure 6). The results obtained from the samples of the infected animal confirmed the Leishmania infection (Section 2, Supplementary material).
Table 6.
The canine samples with nucleotidic variation.
| Code | Position | Degenerate nucleotide | Nucleotides |
|---|---|---|---|
| S20.16 | 262 | Y | C/T |
| S22.3 | 251, 316 | Y | C/T |
| S22.13 | 122, 228, 313 | R | A/G |
| S22.31 | 202 | T |
3.4. Results of Polymerase Chain Reaction for the Biological Samples Collected from “Jimmy”
PCR amplifies the number of copies of fragment(s) of DNA and/or RNA of the microorganism being studied (e.g., Leishmania and others). The POSITIVE result indicates the presence of the microorganism in the biological material analyzed, while the NEGATIVE result indicates its absence. This test may present, although rarely, false-negative or false-positive results, which is a characteristic limitation of the method. Animals with low parasitemia are more prone to false-negative results, particularly when the test is performed using a peripheral blood sample.
The biological samples were collected and analyzed following the established protocols of the Molecular Biology section of the Biomolecular Technology Laboratory at the Institute of Biological Sciences, Federal University of Pará (UFPA), Brazil. The results obtained are as follows:
- Auditive duct swabs: (PCR) Leishmania sp.—Negative
- Mucous membrane of the anus swabs: (PCR) Leishmania sp.—Positive
- Oral mucosa swabs: (PCR) Leishmania sp.—Positive
- Whole blood*: (PCR) Anaplasma platys—Negative
- Whole Blood*:(PCR) Babesia vogeli—Negative
- Whole blood*: (PCR) Ehrlichia canis—Negative
- Whole blood*: (PCR) Leishmania sp.—Positive
- Whole blood*: (PCR) Mycoplasma sp.—Negative
- Whole blood*: (PCR) Rangelia vitalii—Negative
- Preputial secretion swabs: (PCR) Leishmania sp.—Positive
- Ocular swab: (PCR) Leishmania sp.—Positive
*The whole blood sample was collected in tubes containing EDTA. PCR test results that are at odds with the clinical status of the animal should be repeated with a new biological sample. Indirect immunofluorescence assay (IFAT) and quantitative real-time PCR (qPCR) are alternative methods that aid diagnosis.
Figure 5.
Multiple alignment of sequences showing nucleotide variations in the amplified products of the S17/S18 primers.
Figure 5.
Multiple alignment of sequences showing nucleotide variations in the amplified products of the S17/S18 primers.

Figure 6.
Ocular infection in canine leishmaniasis and light microscopy examination of a conjunctival cytological sample showing a macrophage containing multiple Leishmania amastigotes (arrow) (author’s photos, G.R.G.).
Figure 6.
Ocular infection in canine leishmaniasis and light microscopy examination of a conjunctival cytological sample showing a macrophage containing multiple Leishmania amastigotes (arrow) (author’s photos, G.R.G.).
4. Discussion
The clinical signs of leishmaniasis in dogs are directly related to their immune response and follow four stages based on serological status, clinical signals, laboratory findings, and type of therapy and prognosis. These stages result from the interactions between lipids in extracellular vesicles (LEVs) and host cells, which are not well understood. In susceptible animals, parasites can spread from the skin to the local lymph node, spleen, and bone marrow within hours, providing a valuable area for molecular studies of host–parasite interactions [12,31]. According to the present molecular results, L. infantum infection is not associated with the gender or age of the dogs nor were coinfections considered in the positive cases. No difference was found between the types of biological samples studied (blood and conjunctival swab) in the detection of Leishmania sp. However, for greater reliability of the results, PCR is indicated for various types of samples, given the importance of Leishmania infection, mainly due to its high zoonotic potential.
Detection of infection by amplifying kDNA segments and SSUr-rDNA showed similar sensitivity, indicating that both can be used for the diagnosis of canine leishmaniasis [29,30]. The sequencing of 18S rRNA segments allowed the identification of L. infantum in all cases, and four animals were infected by strains with genetic variability, including three heterozygous samples and one with a pyrimidine (C→T) transition-type mutation. According to a clinical study, in resistant dogs, the parasite remains restricted to the skin and draining lymph nodes [31]. Based on these considerations, the first-choice samples for PCR should be bone marrow, lymph node, spleen, skin, and conjunctival swabs. Other samples, such as blood, buffy coat, and urine, are considered less sensitive [12]. New nanotechnology-based approaches to extracellular vesicles have gained significant interest in the scientific community for their potential in therapeutic and diagnostic innovation in parasitology [1]. Despite advances in the Leishmania research, the selective mechanisms of lipids in extracellular vesicles (LEVs) remain poorly understood. There is no consensus on their differential characterization, ultrasensitive detection, specific subtypes, biomarkers, or biogenesis. Understanding the molecular mechanisms and strategies parasites use (e.g., lipids and proteins) to ensure survival could help develop novel antiparasitic drug targets, therapies, and early diagnostic methods [1,32]. Actually, the diagnostic methods for canine leishmaniasis include parasitological (cytology or histology, immunohistochemistry, and culture), molecular (conventional, nested, and real-time PCR, which is considered the most sensitive technique), and serological (quantitative IFAT and enzyme-linked immunosorbent assay (ELISA) and quantitative rapid tests) methods [12]. However, individual case reports can add new parameters for the accuracy of diagnosis, confirming coinfection and the range of differential diagnoses—or if the animal remains healthy or develops a mild, self-limiting illness [31]. These hardy dogs mount a weak antibody response against canine leishmaniasis, but a strong and effective Th1 response. They may have low antibody titers but produce IFN-γ in response to parasitic antigens, generate type I granulomas, mount a strong late-type hypersensitivity response, and eventually destroy the parasites [31]. Resistance to Leishmania has a strong genetic component. For example, Ibizan Hounds, an ancient rabbit-hunting breed, appear resistant to Leishmania. This resistance may be associated with specific major histocompatibility complex (MHC) class II haplotypes and certain Slc11a1 (Nramp) alleles. Therefore, Ibizan Hounds could serve as a good canine model for studying protective anti-Leishmania immune responses [33].
Recent research shows significant differences in the cytokine serum profile and genetic variants related to immune response among canine breeds. Notable genes with fixed polymorphisms include IFNG and IL6R, while other variants with frequencies of 0.7 or higher were found in ARHGAP18, DAPK1, GNAI2, MITF, IL12RB1, LTBP1, SCL28A3, SCL35D2, PTPN22, CIITA, THEMIS, and CD180. Epigenetic regulatory genes, like HEY2 and L3MBTL3, also show intronic polymorphisms [34]. Future studies should further explore why immune response regulation differs in Ibizan Hounds compared to other breeds [34]. Some dogs develop severe and generalized nodular dermatitis, granulomatous lymphadenitis, splenomegaly, and hepatomegaly, exhibiting activation of polyclonal (occasionally monoclonal) B lymphocytes involving all four classes of IgG, as well as hypergammaglobulinemia. They may develop lesions associated with hypersensitivity types II and III [31]. Excessive immunoglobulin production can lead to immune-mediated hemolytic anemia, thrombocytopenia, and antinuclear antibodies [31]. Chronic immune complex deposition can result in glomerulonephritis, uveitis, and synovitis, leading to renal failure and death [31]. Elevated anti-histone antibodies are a feature of some dogs with glomerulonephritis associated with leishmaniasis, correlating with the protein/creatinine ratio and increasing the likelihood of glomerulonephritis [31]. In susceptible animals, parasites can spread from the skin to the local lymph node, spleen, and bone marrow within hours. In resistant dogs, the parasite remains restricted to the skin and draining lymph node, resulting in either a healthy state or a mild, self-limiting disease. Susceptible dogs mount a Th2 response characterized by high antibody levels but poor cell-mediated immunity, attributed to IL-10-producing Treg lymphocytes. The parasite can suppress IL-12 gene transcription, ensuring that the Th2 response predominates. These immune challenges affect the balance between progression to clinical disease and maintaining subclinical disease during chronic infection or progressive disease in susceptible dogs [31].
Following these considerations, vaccines and immunotherapies targeted at recovering or maintaining T and B cell functions can be important in mending the immune balance of parasite–host interactions required for the pathogenesis of canine leishmaniasis [35]. In veterinary practice, infected dogs may show signs of disease, but some positive dogs with subclinical infection or those infected but clinically healthy may exhibit clinicopathological abnormalities or no clinical signs of leishmaniasis. Anti-Leishmania therapeutic protocols can reduce parasite load and infectiousness in treated animals, though their efficacy is only temporary [3,12]. Dogs exhibit extraordinary phenotypic diversity due to selective breeding over the past 200 years, leading to significant variation in immune system function between breeds. This diversity is evident in the unique susceptibility of different breeds to immune-mediated and infectious diseases. Understanding the mechanisms of adaptive and innate immunity in dogs is crucial for elucidating the progression of diseases and the immune response. This knowledge is essential for comprehending how the immune system functions during disease progression in individual dogs [35,36,37,38,39]. In veterinary immunology, pattern recognition receptors (PRRs) are receptors that recognize specific molecular structures on pathogens. Mannose-binding lectin (MBL), a key soluble C-type lectin, is present at high levels in serum and binds to various oligosaccharides. Despite weak individual binding, multiple sites confer high functional activity, allowing MBL to strongly bind pathogens like Leishmania and activate the complement system [31]. Phagocytic cells have many PRRs that interact with infectious agents. Pathogens can bind to neutrophils in plasma, leading to quick ingestion by phagocytosis. Neutrophils can undergo NETosis, releasing nuclear contents into the extracellular fluid after activation by CXCL8 or lipopolysaccharides [31]. This forms networks of extracellular fibers called “neutrophil extracellular traps” (NETs) [29]. NETs are abundant at the sites of acute inflammation. These networks trap and kill several pathogens such as L. amazonensis [31]. NETs can be very important in containing microbial invaders by acting as physical barriers, capturing large numbers of parasites and thus preventing their spread [31]. When promastigote forms of this parasite are injected by sandflies into the skin of a dog, they are quickly phagocytosed by the neutrophils [31]. When neutrophils go into NETosis, parasites are released and then engulfed by macrophages and dendritic cells, in which organisms differentiate into amastigotes. Leishmania amastigotes are obligate intracellular parasites that divide within macrophages until the cells rupture, and when released into the body, they are phagocytosed by adjacent cells [31]. Depending on the degree of host immunity, parasites can be restricted to the skin (skin disease); alternatively, dendritic cells may migrate to the lymph nodes or enter the circulation and lodge in the internal organs, leading to the visceral spread of the disease.
Although the disease is widespread in endemic areas, most dogs are resistant to Leishmania, and only 10%–15% develop the visceral form of the disease [31]. Macrophages are the major host cells for Leishmania, as well as the effector cells that limit or allow the adaptative growth of these parasites within infected macrophage phagolysosomes (intracellular form) [31]. Their resistance to intracellular destruction is the result of multiple mechanisms, including genetic factors. Comparative studies of 245 macrophage genes demonstrated that 37% were suppressed by Leishmania infection [31]. Leishmania lipophosphoglycans (LPGs) delay the maturation of the phagosome, preventing the production of nitric oxide (NO) and inhibiting the response of macrophages to cytokines [31]. These parasites also reduce the presentation of macrophage antigen by suppressing the expression of class II MHC when the parasites stimulate chronic inflammation. They are thus characterized by granulocytic invasion, followed by macrophages, lymphocytes, and natural killer (NK) cells that collectively form granulomas [31]. Additionally, one important factor that determines the success or failure of an infection is the availability of iron [31]. Innate resistance to many intracellular organisms, such as Leishmania, is controlled, in part, by a gene called Slc11a1 (short for solute carrier family 11, member 1a; formerly called Nramp1) [31]. Leishmania can evade the host’s immune response and ensure their survival and completion of their life cycles. In general, antibody-mediated immune responses protect against extracellular protozoa, while cell-mediated responses control intracellular protozoa [31]. Parasitic protozoa employ some sophisticated techniques to ensure their survival in the face of an animal’s immune response [31]. The Th1-mediated responses that result in macrophage activation are important in many diseases caused by protozoa, in which organisms are resistant to intracellular destruction [31]. One of the most significant routes of destruction in M1 cells is the production of NO [31]. The nitrogen radicals formed by the interaction of NO with oxidants are lethal to many intracellular protozoa [31]. However, protozoa, such as Leishmania and T. cruzi, are adept at surviving inside macrophages. They can migrate to safe intracellular vacuoles by blocking phagosome maturation and suppressing the production of oxidants or cytokines [31]. The development of canine immune organs was reviewed, highlighting that hematopoietic and immune cells originate from a common bone marrow stem cell. B cells mature in the fetal liver and bone marrow, acquiring B cell receptor (BCR) and undergoing selection to ensure that only functional BCR-expressing B cells survive. Immature T cells are exported to the thymus for final maturation. While puppies are considered immunocompetent between six and 12 weeks of age, the onset of immunocompetence varies due to the presence of maternal-derived antibodies (MDA) [40]. Increased lifespan has revealed age-related susceptibility to infectious, inflammatory, autoimmune, and neoplastic diseases. Age-related changes include impaired cell-mediated immune responses, reduced blood lymphocyte proliferation, and decreased cutaneous delayed-type hypersensitivity [40]. Additionally, there is a decline in the humoral immune response, likely due to decreased Th cell functionality. Despite this, the ability to mount humoral immune responses persists, as shown by protective vaccine antibody titers and responses to booster vaccinations. The triennial revaccination program provides adequate protection for young and adult dogs but may not protect geriatric dogs.
Older dogs often exhibit impaired immune responses to new antigenic challenges, likely due to a reduced pool of naïve T cells and lower T cell receptor diversity [40]. The key genetic elements of immune responsiveness are found in the MHC genes, present as the dog leukocyte antigen (DLA) and feline leukocyte antigen (FLA) systems [36]. This suggests that specific dog breeds have genetically determined immune functions, with recent studies showing breed-specific serological responses to vaccination. This genetic background likely affects immune system maturation. In dogs, C-reactive protein (CRP) is the main acute-phase protein, increasing significantly in infectious diseases, such as leishmaniasis, babesiosis, parvovirus, and colibacillosis [36]. Acute-phase protein levels also rise moderately in canine inflammatory bowel disease. CRP, haptoglobin, and serum amyloid A (SAA) protein levels are elevated in the cerebrospinal fluid and serum of dogs with corticosteroid-responsive arthritis and meningitis. Pregnant dogs show moderately higher levels of haptoglobin, ceruloplasmin, and fibrinogen [36]. Some studies have shown that dogs in the asymptomatic and symptomatic groups exhibit heterogeneity in copper (Cu), zinc (Zn), and iron (Fe) concentrations compared with the control group, emphasizing the important roles of trace elements (TEs) in leishmaniasis [40]. This suggests that TEs could be assessed as a prognostic factor in leishmaniasis and/or as an adjuvant for the treatment of leishmaniasis [41]. Susceptible dogs mount a Th2 response with high antibody levels but poor cell-mediated immunity, attributed to IL-10-producing Treg lymphocytes [31]. The parasite can suppress IL-12 gene transcription, ensuring Th2 response predominance. This leads to chronic, progressive disease with macrophages accumulating parasites and spreading throughout the body [31]. Despite their antigenicity, parasitic protozoa survive using multiple evasion mechanisms. Studies focus on blocking these mechanisms and developing vaccines for canine leishmaniasis [1,31]. Effective vaccines, such as those using purified Leishmania glycoproteins (fucose–mannose ligands), prevent disease and serve as immunotherapeutic agents [31]. An alternative vaccine with L. infantum promastigote excretory and secretory products and muramyl dipeptide also shows promise [31]. Experimental vaccines, including attenuated and DNA based, have shown promising results in veterinary medicine [31]. Additional Leishmania control mechanisms and tools are needed, including new drugs, vaccines, diagnostics, and vector control agents and strategies [1,42].
Considering these important points, for the future of prevention and treatment of leishmaniases, we can include all advances resulting from the LEVs research worldwide, which represent a variety of advantages over live biotherapeutics (next-generation therapies) [42]. Over the past years, the isolation and analysis of extracellular vesicles from Leishmania have been challenging. The protocols are not yet standardized, as referred to in the guidelines for extracellular production of Leishmania published by Gabriel et al. in 2021 [1,42], since many protocols with potential effects on the outcome are used and published. It is important to note that several protocols are labor-intensive, requiring costly equipment, increasing risks for the loss of heterogeneous extracellular vesicles, and not discriminating well between exosomes and contaminating structures such as lager vesicles and protein/lipid aggregates [43]. In vitro direct isolation performed with magnetic beads for multiomic analysis (size, concentration, and phenotype by Spectradyne particle analysis, western blot analysis, LC–MS analysis, and flow cytometry) can be applied for LEVs research, requiring minimal hands-on activities and providing highly pure exosomes with a minimal loss of material. It also enables future automation opportunities in different formats and other simple, rapid, and reliable bead-based extracellular vesicle isolation methods based on the strong anion exchange (SAX) principle (e.g., using both automated KingFisher for rapid and efficient isolation of exosomes compatible with the KingFisher Duo Prime, Flex, and Apex systems (Thermo Fisher Scientific) and improved manual protocols) (Figure 7) [44].
For medical parasitology, multidisciplinary cooperation in multiomic extracellular vesicle research is crucial to understanding Leishmania–host cell interactions. This includes a broad-scale analysis of extracellular molecules and the development of innovative applications based on Leishmania virulence factors, such as LPG, surface acid proteinase (GP63), glycoinositolphospholipids (GIPLs), proteophosphoglycan (PPG), A2 protein, kinetoplastid membrane protein-11 (KMP-11), nucleotidases, heat-shock proteins (HSPs), and transmembrane transporters, which support parasite survival and propagation in host cells [1]. Based on these findings, we propose a rapid bead-based isolation of LEVs for future multiomic research. This includes techniques for manual and automated bead-based target isolation workflows to achieve quick LEVs isolation (e.g., within ten minutes using Invitrogen™ Dynabeads™, Thermo Fisher Scientific), based on our observations and practice [1,44].
This study reinforces previous guidelines for LEVs research in vitro and in vivo, aiming to improve prevention, diagnosis, prognosis, and therapeutics. Our results emphasize the importance of focusing on lipids in LEVs and understanding their release mechanisms from parasites, and their role in sensing the lysosome-specific environment (pH and host temperature). LEVs have a lipid bilayer membrane that protects encapsulated materials (proteins, nucleic acids, lipids, and metabolites) from the extracellular environment. Exploring the mechanisms and functions of LEVs in parasitic production and signaling can help activate or deactivate barriers to prevent host infection and understand how LEVs interfere with disease progression [1]. Lipids Lipids are crucial components of extracellular vesicles, serving as energy sources, structural elements, and signaling mediators in Apicomplexa parasites. However, our knowledge about the lipid composition and the function of lipids in mechanisms involved in vesicular trafficking during leishmaniasis is limited [45,46]. Thus, analyzing the reproducible results for lipid bodies of Leishmania labeled with BODIPY™ 493/503 (Thermo Fisher Scientific) has shown that the crucial 72-hour period is a limit to the decrease of lipids released by parasites of genus Leishmania and subgenus Leishmania (e.g., species complex: Leishmania donovani, Leishmania tropica, Leishmania major, Leishmania aethiopica, and Leishmania mexicana) [1]. According to Zhang (2021), amastigotes likely acquire most of their lipids from host cells while retaining some capacity for de novo synthesis. In contrast, promastigotes rely on de novo synthesis to produce most of their lipids, including glycerophospholipids, sterols, and sphingolipids [45]. This balance is just the tip of the iceberg in lipid metabolism, playing a crucial role in Leishmania infection within the first 72 hours after transmission. It likely alters plasma membrane fluidity and could become a new focus for cell–cell communication research between trypanosomatids and their hosts [46]. Factors linked to these changes include the length of the fatty acid tail, temperature, cholesterol content of the bilayer, and the degree of saturation of fatty acid tails [47]. Therefore, new protocols are needed to improve prevention and clinical treatment, considering the relationship of lipids released from parasites [1].
Various laboratory tests are available to diagnose parasitic infections, including conventional gold standard methods and serological tests. In recent decades, several molecular diagnostic tools have been developed to detect parasites and new strains [48,49,50]. Accurate diagnosis of zoonotic infections involves collaboration between medical scientists, policymakers, and public health officials working to prevent the dissemination of these diseases and establish a worldwide network of surveillance for the coinfection of parasitic infections [49,50]. Advanced techniques for studying and diagnosing simultaneous infections indicate that multi-parasitism is more common than single infections [51,52]. Evolutionarily, multi-parasite systems are ecologically dynamic, involving key host species and contributing to transmission [52]. Understanding these complex relationships is a top priority for biomedical sciences in the 21st century [53]. Clinically, coinfection of zoonotic parasites in companion animals may appear in its classical form, with acute aggressive evaluation or as long-standing infection, which may be asymptomatic or sometimes nonspecific, making clinical diagnosis difficult [52,53,54]. Veterinarians and clinical researchers should consider the health status and background of the patients (animals or humans) to implement a better parasite management program [53]. Considering certain factors, such as resource-mediated processes, often influence how, where, and which coinfecting parasites interact. These factors may dictate the need for more intensive monitoring for effective treatment, while others may suggest a less aggressive approach [53,55,56]. Furthermore, innovative strategies applying knowledge about extracellular vesicles and their specific profiles (e.g., proteic and lipidic data basis) from studies of host–parasite mechanisms can be incorporated into immunotherapy. This incorporation can interfere in the dynamics of disease transmission and progression and the development of effective, safe, and available vaccines against leishmaniasis, thus helping protect puppies and dogs of different ages. Four vaccines against canine leishmaniasis are available on the market, Leishmine® and Leish-Tec® in Brazil and CaniLeish® and LetiFend® in Europe (the first vaccine based on purified excreted/secreted antigens of Leishmania has been licensed in Europe since 2011) [1,3,35,57]. Current vaccine recommendations require vaccinating seronegative dogs. In regions where the disease is endemic in both dogs and humans, accurately identifying healthy, uninfected animals is challenging due to limitations in diagnostics. Adverse events were mild and localized, suggesting that vaccinating healthy subclinical dogs may justify revising current vaccination and immunotherapy guidelines for infected but healthy animals [35]. Despite existing studies on licensed vaccines for canine leishmaniasis, they are still considered insufficient due to lack of standardization, methodological deficiencies, and differences in study populations.
Further research is needed in xenodiagnostic studies to assess the infectivity of Leishmania and the potential interference of vaccination in diagnosing its infection. Additionally, long-term pharmacological surveillance should be maintained post-licensing to provide reliable information to relevant organizations and the public [57]. As indicated in our previous publication on Guidelines for Exosomal Research, further advances in techniques and protocols are expected to improve the accuracy of isolating and characterizing LEVs and their activity on host immune responses. This includes understanding their lipid bilayer membrane, which protects encapsulated materials like lipids from the extracellular environment. Knowledge about the lipid composition and function of LEVs is limited, and we propose its significant molecular role during the pathogenesis of leishmaniasis. Changes in lipid profile and metabolism in both parasite and host during disease development depend on lipid bodies. Further research is needed to fully understand the in vivo relationship between host lipid metabolism and LEVs, immune responses, disease prognosis, and the necessary advances in the prevention and treatment of leishmaniasis [58].
5. Conclusions
According to the WHO, One Health is an integrated, unifying approach that aims to sustainably balance and optimize the health of people, animals, and ecosystems. There is a well-documented concern related to the close contact between companion animals and people, with the well-known positive aspects of human–animal interactions. However, new hypotheses and new models, as well as the reformulation of aspects of analytic techniques, have been put forward over time. It is claimed that it is an open field for future research in favor of solutions for public health concerning emerging or reemerging diseases.
In this context, despite many advances, we continue fighting to develop human vaccines against Leishmania and other neglected diseases. Considering the risk of companion animals transmitting zoonoses to humans, multidisciplinary knowledge sharing is necessary between medical scientists and veterinarians, who are able to accurately diagnose (e.g., molecular nanodiagnostics) zoonotic protozoal and helminth infections. Prevention efforts may include a worldwide network of surveillance for coinfection, continuously adapting to global environmental changes. Unfortunately, Leishmania-infected dogs continue to be parasite reservoirs for sandfly vectors, and cofactors can improve the risk of coinfection of Leishmania with other pathogens. Therefore, further studies focusing on the clinical diagnosis and epidemiological aspects of coinfection with zoonotic diseases in endemic or non-endemic regions are necessary for better control of them in companion animals, as the source of infection is important and may be underrated. Highlighting crucial molecular mechanisms into this nanouniverse related to immunomodulation to understand how the extracellular vesicles of different disease agents interact with host cells is urgent.
Why should one scrutinize the evidence of extracellular vesicles in One Health? We propose a hypothesis for future opportunities to extend research on comparative studies considering multi-infections. Against this background and considering that lipids are essential metabolites necessary for the biogenesis, signaling, and energy storage of Apicomplexa parasite membranes, we propose, based on our findings, to broaden the research community’s interest in the lipids released by Leishmania and their role in host–parasite extracellular interactions (mediating communication between infected and uninfected cells). This follows the advancements in the expertise development of rapid bead-based isolation of LEVs using Invitrogen™ Dynabeads™ for a multiomic research. The research can reinforce our considerations of the previously proposed guidelines for the in vitro and in vivo research of Leishmania extracellular vesicles, using animal or human samples (e.g., infected tissues). It can embrace topics related to extracellular vesicles from the etiologic agent, host-specific interactions, consequences for host health (e.g., asymptomatic host), clinical and laboratory features for differential diagnosis, new approaches for nano-targeted therapies, production of vaccines against infection or coinfections, and synergistic effects of extracellular vesicles during the course of an infection within the host. Stray animals may unintentionally pose a public health hazard. Therefore, there is an open field of research to improve differential diagnosis in companion animals and highlight pathogen–host interactions. Moreover, there are still gaps in the research on extracellular vesicles, immunomodulation, and their interactions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Á.M.G.; formal analysis, Á.M.G., G.R.G., W.L.A.P., A.G.P., L.M.N.C., K.W.P., E.C.G., D.D.F.A. and E.O.d.S.; investigation, Á.M.G., G.R.G., W.L.A.P., A.G.P., L.M.N.C. and K.W.P.; resources, Á.M.G., G.R.G., W.L.A.P., E.C.G., D.D.F.A. and E.O.d.S.; writing—review and editing, Á.M.G., G.R.G., W.L.A.P., A.G.P., L.M.N.C., K.W.P., E.C.G., D.D.F.A. and E.O.d.S.; supervision, E.C.G., D.D.F.A. and E.O.d.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Coordination for the Improvement of Higher Education Personnel (CAPES) and the Portuguese Foundation for Science and Technology (FCT), through the research project GHTM–UID/Multi/04413/2013 and the Portugal–Brazil research project PTDC/SAU-PAR/28459/2017 EXOTRYPANO IHMT-NOVA/FMV-ULisboa/UFRN.
Institutional review board statement
All procedures performed involving animals were conducted in accordance with the ethical technical standards of the Animal Use Ethics Committee of the Federal University of Pará (UFPA-Brazil) (CEUA 8044181219, approved on April 30, 2020).
Data Availability Statement
Data supporting these findings are available within the article, at https://doi.org/10.20935/AcadBiol7491, or upon request.
Acknowledgments
All the authors of the manuscript acknowledge their respective laboratories, institutes, and universities. They also thank the anonymous reviewers for their thoughtful comments and efforts for improving the manuscript. Á.M.G. gratefully acknowledges Professor Gabriela Santos-Gomes from the Laboratory of IHMT-UNL, Portugal for her continuous support, scientific guidance, and thesis supervision. Á.M.G. also gratefully acknowledges the Graduate Program in Biology of Infectious and Parasitic Agents (PPG BAIP) at the Institute of Biological Sciences, Federal University of Pará, Brazil.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gabriel ÁM, Galué-Parra A, Pereira WLA, Pedersen KW, da Silva EO. Leishmania 360°: guidelines for exosomal research. Microorganisms. 2021;9:2081. [CrossRef]
- WHO. Control of neglected tropical diseases. [cited 2023 June 10]. Available from: https://www.who.int/teams/control-of-neglected-tropical-diseases/interventions/strategies.
- Manuela G, Antonio B, Valentina FM, Gioia B, Riccardo B, Daniela G, et al. Examining the relationship of clinical and laboratory parameters with infectiousness to Phlebotomus perniciosus and its potential infectivity in dogs with overt clinical leishmaniasis. Front Vet Sci. 2021;8: 667290. [CrossRef]
- Wijerathna T, Gunathilaka N. Diurnal adult resting sites and breeding habitats of phlebotomine sand sandflies flies in cutaneous leishmaniasis endemic areas of Kurunegala District, Sri Lanka. Parasites Vectors. 2020;13:284. [CrossRef]
- Cecílio P, Cordeiro-da-Silva A, Oliveira F. Sandflies: basic information on the vectors of leishmaniasis and their interactions with Leishmania parasites. Commun Biol. 2022;5:305. [CrossRef]
- Dodd RY. Transmission of parasites by blood transfusion. Vox Sang. 1998;74(Suppl 2):161–3. PMID: 9704440. [CrossRef]
- Pangrazio KK, Costa EA, Amarilla SP, Cino AG, Silva TMA, Paixão TA, et al. Tissue distribution of Leishmania chagasi and lesions in transplacentally infected fetuses from symp tomatic and asymptomatic naturally infected bitches, Vet Parasitol. 2009;165(3–4):327–31. ISSN 0304-4017. [CrossRef]
- Avila-García M, Mancilla-Ramírez J, Segura-Cervantes E, Farfan-Labonne B, Ramírez-Ramírez A, Galindo-Sevilla N. Transplacental transmission of cutaneous Leishmania mexicana strain in BALB/c mice. Am J Trop Med Hyg. 2013; 89(2):354–8. PMID: 23798582; PMCID: PMC3741259. [CrossRef]
- Karkamo V, Kaistinen A, Näreaho A, Dillard K, Vainio-Siukola K, Vidgrén G, et al. The first report of autochthonous non-vector-borne transmission of canine leishmaniasis in the Nordic countries. Acta Vet Scand. 2014;56:84. [CrossRef]
- Naucke TJ, Amelung S, Lorentz S. First report of transmission of canine leishmaniasis through bite wounds from a naturally infected dog in Germany. Parasites Vectors. 2016;9:256. [CrossRef]
- Miró G, López-Vélez R. Clinical management of canine leishmaniasis versus human leishmaniasis due to Leishmania infantum: Putting “One Health” principles into practice. Vet Parasitol. 2018;254:151–9. [CrossRef]
- Solano-Gallego L, Miró G, Koutinas A, Cardoso L, Pennisi MG, Ferrer L, et al. LeishVet guidelines for the practical management of canine leishmaniasis. Parasit Vectors. 2011; 4:86. PMID: 21599936; PMCID: PMC3125381. [CrossRef]
- Herrera-Mares A, Guzmán-Cornejo C, Ulloa-García A, Córdoba-Aguilar A, Silva-de la Fuente MC, Suzán G. Mites, rodents, and pathogens: a global review for a multi-species interaction in disease ecology. Acta Tropica. 2022; 232:106509. [CrossRef]
- Janeway CA Jr, Travers P, Walport M, Shlomchik, M. Immunobiology: the immune system in health and disease. 5th ed. New York: Garland Science; 2001.
- Murray JL, Connell JL, Stacy A, Turner KH, Whiteley M. Mechanisms of synergy in polymicrobial infections. J Microbiol. 2014;52(3):188–99. PMID: 24585050; PMCID: PMC7090983. [CrossRef]
- Baneth G, Thamsborg SM, Otranto D, Guillot J, Blaga R, Deplazes P, et al. Major parasitic zoonoses associated with dogs and cats in Europe. J Comp Pathol. 2016;155:S54–74. [CrossRef]
- Vilhena H, Martinez-Díaz VL, Cardoso L, Vieira L, Altet L, Francino O, et al. Feline vector-borne pathogens in the north and centre of Portugal. Parasit Vectors. 2013;6:99–104. [CrossRef]
- Dantas-Torres F, Otranto D. Dogs, cats, parasites, and humans in Brazil: opening the black box. Parasit Vectors. 2014; 7:22–47. [CrossRef]
- Futuyma DJ. Evolutionary biology. 3rd ed. Sunderland: Sinauer Association; 1998. p. 15, p. 763.
- Do Rosário CJRM, Dominici MF, Braga MSCO, Lima CAA, Pereira JG, Melo FA. Quantification of IL-10 and IFN-γ in dogs with or without clinical signs of Leishmania (Leishmania) chagasi infection symptoms. Pesq. Vet. Bras. 2018; 38(01). [CrossRef]
- Yakubovich EI, Polischouk AG, Evtushenko VI. Principles and problems of exosome isolation from biological fluids. Biochem (Mosc) Suppl Ser A Membr Cell Biol. 2022;16(2):115–26. PMID: 35730027; PMCID: PMC9202659. [CrossRef]
- Harayama T, Riezman H. Understanding the diversity of membrane lipid composition. Nat Rev Mol Cell Biol. 2018;19:281–96. [CrossRef]
- Fyfe J, Casari I, Manfredi M, Falasca M. Role of lipid signalling in extracellular vesicles-mediated cell-to-cell communication. Cytokine Growth Factor Rev. 2023;73:20–26. [CrossRef]
- Gocze PM, Freeman DA. Factors underlying the variability of lipid droplet fluorescence in MA-10 Leydig tumor cells. Cytom J Int Soc Anal Cytol. 1994;17(2):151–8. [CrossRef]
- Plotkowski MC, Brandão BA, de Assis MC, Feliciano LFP, Raymond B, Freitas C, et al. Lipid body mobilization in the ExoU-induced release of inflammatory mediators by airway epithelial cells. Microb Pathog. 2008;45(1):30–7. [CrossRef]
- Da Silva BJM, Souza-Monteiro JR, Rogez H, Crespo-López ME, do Nascimento JLM, Silva EO. Selective effects of Euterpe oleracea (açai) on Leishmania (Leishmania) amazonensis and Leishmania infantum. Biomed Pharmacother. 2018;97:1613–21. [CrossRef]
- Baptista-Fernandes T, Marques C, Rodrigues OR, Santos-Gomes GM. Intra-specific variability of virulence in Leishmania infantum zymodeme MON-1 strains. Comp Immunol Microbiol Infect Dis. 2007;30(1):41–53. [CrossRef]
- Singh R, Joseph A, Umapathy T, Tint NL, Dua HS. Impression cytology of the ocular surface Br J Ophthalmol. 2005; 89:1655–9. [CrossRef]
- Galvão GR, Gonçalves EC, Moura LGS, Virgolino RR, Neves AMP, Aguiar DCF. First molecular description of autochthonous urban cases of canine visceral leishmaniasis in the city of Belém, Pará, Brazil. Braz J Biol. 2023;83:e267617. [CrossRef]
- Galvão GR. Detecção e epidemiologia molecular de Leishmania sp. em três municípios do estado do Pará [Dissertação de Mestrado em Biologia de Agentes Infecciosos e Parasitários]. Belém: Universidade Federal do Pará; 2018. 58 p.
- Tizard IR. Imunologia veterinária. 9th ed. Rio de Janeiro: Elsevier, 2014.
- Haase S, Temesvari L, Zhang K. Editorial: lipids in host and protozoan parasite interaction. Front Cell Infect Microbiol. 2023;13:1334002. PMID: 38045759; PMCID: PMC10693326. [CrossRef]
- Solano-Gallego L, Llull J, Ramos G, Riera C, Arboix M, Alberola J, et al. The Ibizian hound presents a predominantly cellular immune response against natural Leishmania infection. Vet Parasitol. 2000;90(1-2):37-45. PMID: 10828510. [CrossRef]
- Álvarez L, Marín-García PJ, Llobat L. Immunological and genomic characterization of Ibizan Hound dogs in an endemic Leishmania infantum region. Parasit Vectors. 2022; 15(1):445. PMID: 36443886; PMCID: PMC9706964. [CrossRef]
- Toepp AJ, Petersen CA. The balancing act: immunology of leishmaniasis. Res Vet Sci. 2020; 130:19–25. [CrossRef]
- Day MJ. Immune system development in dog and cat. J Comp Pathol. 2007;137 Suppl 1: S10–5. PMID: 17560591. [CrossRef]
- Chaplin DD. Overview of the immune response. J Allergy Clin Immunol. 2010;125(2 Suppl 2): S3–23. PMID: 20176265; PMCID: PMC2923430. [CrossRef]
- Hosein S, Blake D, Solano-Gallego L. Insights on adaptive and innate immunity in canine leishmaniasis. Parasitology. 2017;144(1):95–115. [CrossRef]
- Varadé J, Magadán S, González-Fernández Á. Human immunology and immunotherapy: main achievements and challenges. Cell Mol Immunol. 2021;18:805–28. [CrossRef]
- Pereira M, Valério-Bolas A, Saraiva-Marques C, Alexandre-Pires G, Pereira da Fonseca I, Santos-Gomes G. Desenvolvimento do Sistema Imunológico Canino: Do Útero ao Idoso. Vet Sci. 2019; 6(4): 83. [CrossRef]
- Taghipour A, Abdoli A, Ramezani A, Abolghazi A, Mofazzal Jahromi MA, Maani S, et al. Leishmaniasis and trace element alterations: a systematic review. Biol Trace Elem Res. 2021; 199:3918–38. [CrossRef]
- Hotez PJ, Pecoul B, Rijal S, Boehme C, Aksoy S, Malecela M, et al. Eliminating the neglected tropical diseases: translational science and new technologies. PLoS Negl Trop Dis. 2016; 10(3):e0003895. [CrossRef]
- Neurauter A, Kierulf B, Kullmann LAM, Manger I, Pedersen KW. From basic research to potential clinical applications: adapting methods for EV enrichment and analysis. [cited 2023 June 10]. Available from: https://www.esccap.org/guidelines/gl6/.
- Kullmann A, Reed BM, Pedersen KW. Rapid bead-based isolation of exosomes for multiomic research. 2023 [cited 2023 Aug 28]. Available from: https://assets.thermofisher. com/TFS-Assets/BID/Application-Notes/rapid-bead-based-isolation-exosomes-app-note.pdf.
- Zhang K. Balancing de novo synthesis and salvage of lipids by Leishmania amastigotes. Curr Opin Microbiol. 2021; 63:98–103. [CrossRef]
- Shunmugam S, Arnold C-S, Dass S, Katris NJ, Botté CY. The flexibility of Apicomplexa parasites in lipid metabolism. PLoS Pathog. 2022;18(3):e1010313. [CrossRef]
- Johnson A. How does cholesterol affect the fluidity of a membrane? 2024 [cited 2024 Oct 31]. Available from: https://scienceoxygen.com/how-does-cholesterol-affect-the-fluidity-of-a-membrane/.
- Cargnelutti DE, Borremans CG, Tonelli RL, Carrizo LC, Salomon MC. Diagnosis of Leishmania infection in a nonendemic area of South America. J Microbiol Immunol Infec. 2016;49:809–12. [CrossRef]
- Johansen MV, Lier T, Sithithaworn P. Progress in research and control of helminth infections in Asia Towards improve diagnosis of neglected zoonotic trematodes using a One Health approach. Acta Tropica. 2015;141(2):161–9. [CrossRef]
- Al Dahouk S, Sprague LD, Neubauer H. New developments in the diagnostic procedures for zoonotic brucellosis in humans. Rev Sci Tech Off Int Epiz. 2013;32(1):177–88. [CrossRef]
- Tavares RG, Staggemeier R, Borges ALP, Rodrigues MT, Castelan LA, Vasconcelos J, et al. Molecular techniques for the study and diagnosis of parasite infection. J Venom Anim Toxins Incl Trop Dis. 2011;17(3):239–48. [CrossRef]
- Steinmann P, Utzinger J, Du ZW, Zhou XN. Multiparasitism: a neglected reality on global, regional and local scale. Adv Parasitol.2010;73: 21–50. [CrossRef]
- Griffiths EC, Pedersen AB, Fenton A, Petchey OL. Analysis of a summary network of coinfection in humans reveals that parasites interact most via shared resources. Proc R Soc B. 2014;281:1–9.
- Laurenti MD, Rossi CN, da Matta VL, Tomokane TY, Corbett CE, Secundino NF, et al. Asymptomatic dogs are highly competent to transmit Leishmania (Leishmania) infantum chagasi to the natural vector. Vet Parasitol. 2013;196(3-4):296–300. [CrossRef]
- European Scientific Counsel Companion Animal Parasites. Control of intestinal protozoa in dogs and cats - guideline 6 1st edition. Malvern: ESCCAP; 2011 [cited 2023 June 10]. Available from: https://www.esccap.org/uploads/docs/3sbvfy71_ESCCAP_Guide_6_spanish_version_def.pdf.
- European Scientific Counsel Companion Animal Parasites. Control of vector borne diseases in dogs and cats - guideline 52rd edition. Malvern: ESCCAP; 2012 [cited 2023 June 10]. Available from: https://www.esccap.org/guidelines/.
- Velez R, Gállego M. Commercially approved vaccines for canine leishmaniasis: a review of available data on their safety and efficacy. Trop Med Int Health. 2020;25(5):540-557. PMID: 32034985. [CrossRef]
- Martínez CR, Ruiz CJ. Alterations in host lipid metabolism produced during visceral leishmaniasis infections. Curr Trop Med Rep. 2019;6:250–5. [CrossRef]
Figure 1.
One Health Approach to Zoonotic Leishmaniases caused by old- and new-world species in endemic areas, geographically distributed or segregated into four eco-epidemiological regions of the world: the Americas, East Africa, North Africa, and West and Southeast Asia. Green color indicates the spread of visceral leishmaniasis, and purple color indicates cutaneous leishmaniasis. Both types present rapid adaptation, expanding geographic ranges influenced by risk factors, with new epidemiological scenarios emerging in previously disease-free areas, and thus increasing transmission (also through transfused blood products from blood donors who are carriers of infection, by transplacental and venereal transmission, and perhaps through direct dog-to-dog Leishmania transmission via wounds or bites) [1,2,3,4]. The higher incidence of disease, as well as its emergence or reemergence in recent years, may be related to many factors. Among these factors, we highlight socioeconomic conditions; climate and environmental change; closer contact between humans, pets, and wild ecosystems; and the parasite’s resistance to drugs and their vectors’ resistance to insecticides [6,7,8,9,10].
Figure 1.
One Health Approach to Zoonotic Leishmaniases caused by old- and new-world species in endemic areas, geographically distributed or segregated into four eco-epidemiological regions of the world: the Americas, East Africa, North Africa, and West and Southeast Asia. Green color indicates the spread of visceral leishmaniasis, and purple color indicates cutaneous leishmaniasis. Both types present rapid adaptation, expanding geographic ranges influenced by risk factors, with new epidemiological scenarios emerging in previously disease-free areas, and thus increasing transmission (also through transfused blood products from blood donors who are carriers of infection, by transplacental and venereal transmission, and perhaps through direct dog-to-dog Leishmania transmission via wounds or bites) [1,2,3,4]. The higher incidence of disease, as well as its emergence or reemergence in recent years, may be related to many factors. Among these factors, we highlight socioeconomic conditions; climate and environmental change; closer contact between humans, pets, and wild ecosystems; and the parasite’s resistance to drugs and their vectors’ resistance to insecticides [6,7,8,9,10].
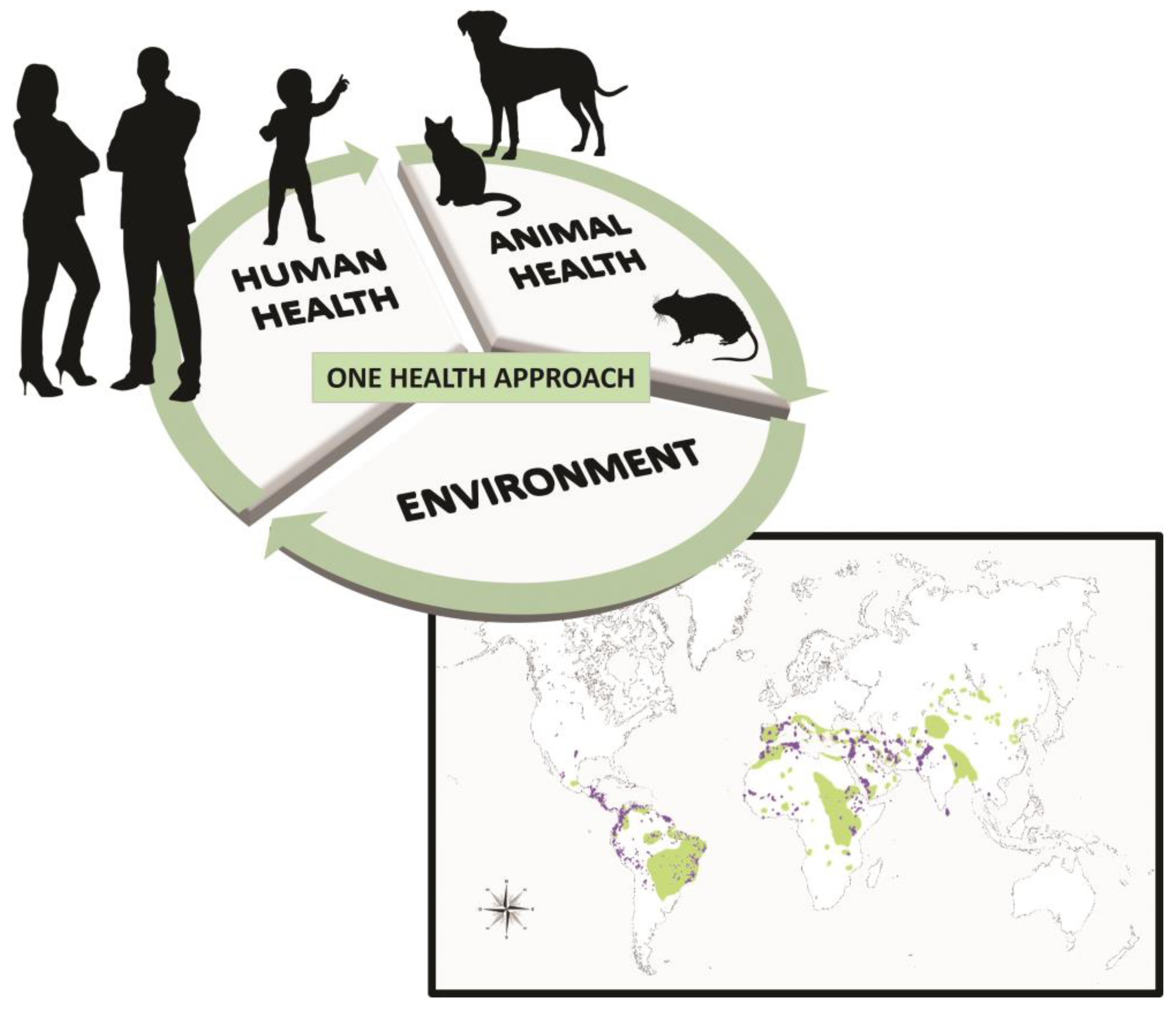
Figure 2.
(a) Transmission electron microscopy of macrophage infected with Leishmania amazonensis (MHOM/BR/2009/M26361 strain). Scale bar: 1 µ. (b) Tubular structures and vesicles (arrows) participating in the endocytosis and exocytosis processes can be seen in the flagellar pocket (FP) of the promastigote. Scale bar: 500 nm. (c and d) Transmission electron microscopy of tissues from male BALB/c mice infected with Leishmania amazonensis: lipid-like structures (arrows). Scale bar: 2 μm.
Figure 2.
(a) Transmission electron microscopy of macrophage infected with Leishmania amazonensis (MHOM/BR/2009/M26361 strain). Scale bar: 1 µ. (b) Tubular structures and vesicles (arrows) participating in the endocytosis and exocytosis processes can be seen in the flagellar pocket (FP) of the promastigote. Scale bar: 500 nm. (c and d) Transmission electron microscopy of tissues from male BALB/c mice infected with Leishmania amazonensis: lipid-like structures (arrows). Scale bar: 2 μm.
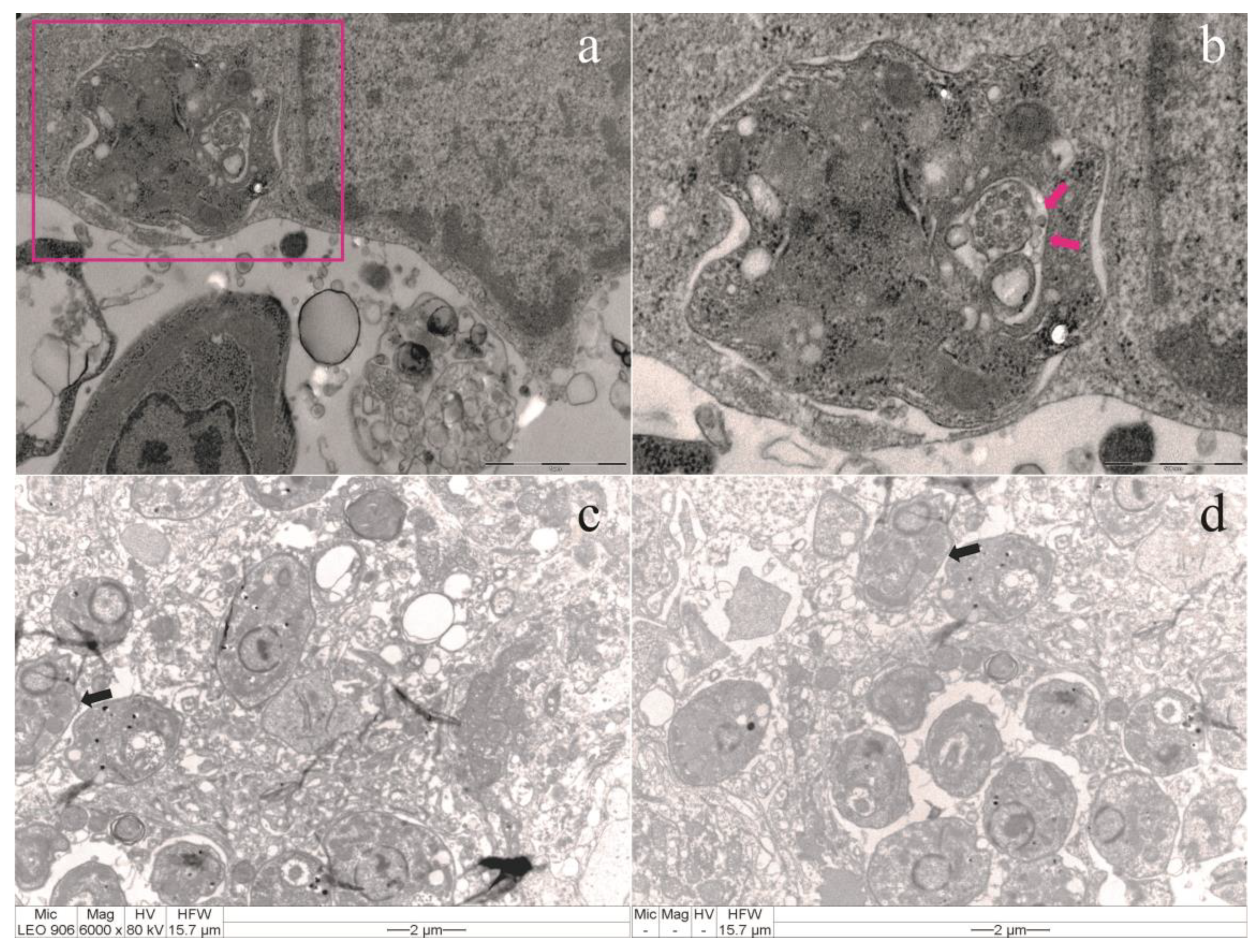
Figure 7.
Extracellular vesicles isolation workflows. Manual (a) and automated (b) methods can be applied for in vitro and in vivo LEV research to highlight their biogenesis, composition, and molecular interactions with host immune cells [44].
Figure 7.
Extracellular vesicles isolation workflows. Manual (a) and automated (b) methods can be applied for in vitro and in vivo LEV research to highlight their biogenesis, composition, and molecular interactions with host immune cells [44].
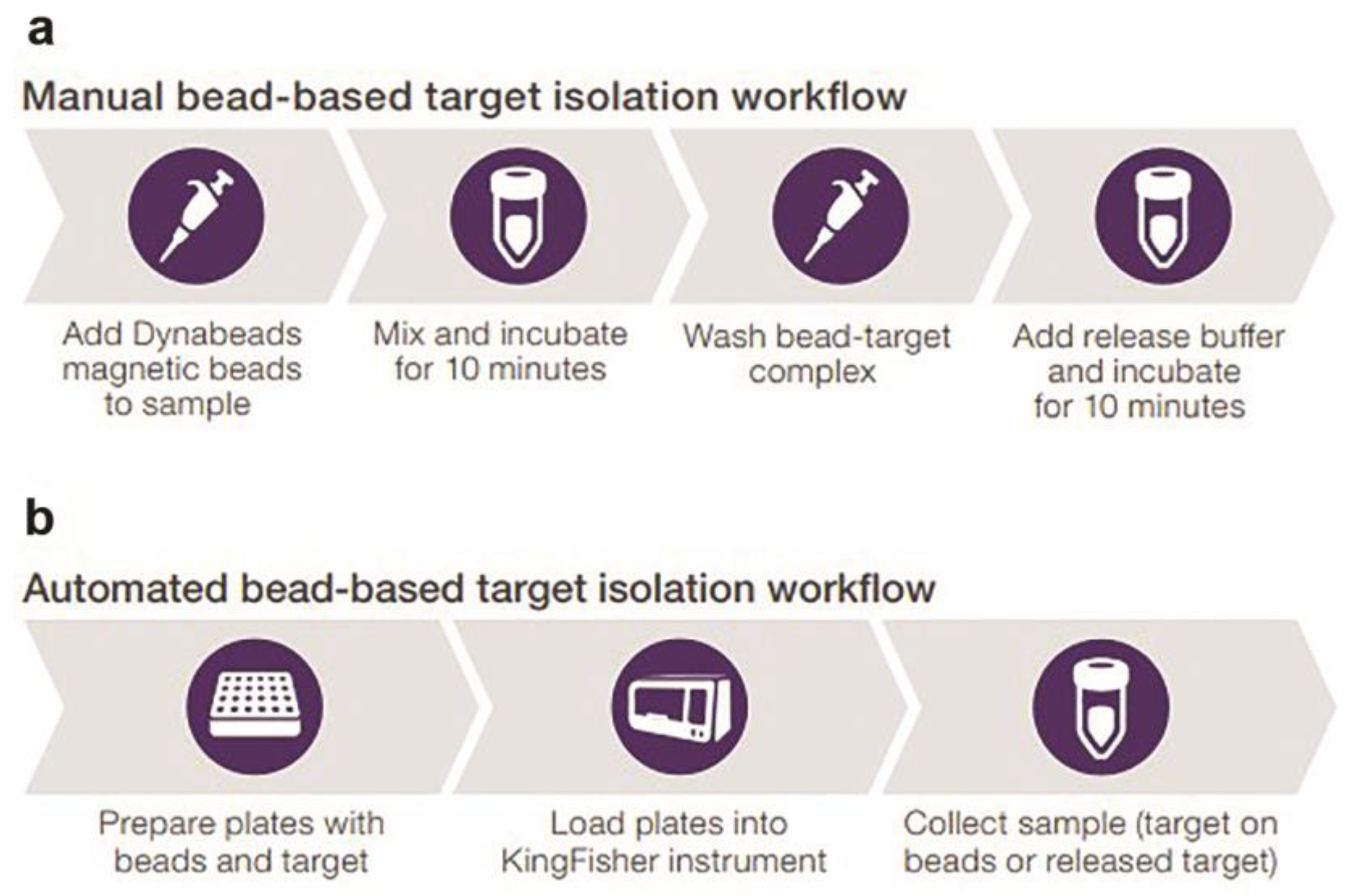
Table 1.
Oligonucleotides used for identifying amplified DNA fragment size and Leishmania species through molecular biology assays, including PCR-SSUr and PCR-kDNA.
Table 1.
Oligonucleotides used for identifying amplified DNA fragment size and Leishmania species through molecular biology assays, including PCR-SSUr and PCR-kDNA.
| Gene | Methods | Oligonucleotides | Base pair (bp) | Identification |
|---|---|---|---|---|
| SSUr |
PCR | R221—5’-GGTTCCTTTCCTGATTTACG-3’ R332—5’-GGCCCGTAAAGGCCGAATAG-3’ |
603 | Kinetoplastid |
| Nested | R223—5’-TCCCATCGCAACCTCGGTT-3’ R333—5’-AAAGCGGGCGCGGTGCTG-3’ |
490 | Leishmania sp. | |
| SSUs |
PCR | S4—5’-GAT CCA GCT GCA GGT TCA CC-3’ S12—5’-GGT TGA TTC CGT CAACGG AC-3’ |
520 | Trypanosomatids |
| Nested | S17—5’-CCA AGC TGC CCA GTA GAA T-3’ S18—5’-TCG GGC GGA TAA AAC ACC-3’ |
358 | Leishmania sp. | |
| kDNA | PCR |
Leish 1—5’-AACTTTTCTGGTCCTCCGGGTAG-3’ Leish 2—5’-ACCCCCAGTTTCCCGCC -3´ |
120 | Leishmania infantum |
PCR, polymerase chain reaction.
Table 5.
Detection of Leishmania sp. DNA in animals through markers for kDNA and SSUr-rDNA chromosomal.
Table 5.
Detection of Leishmania sp. DNA in animals through markers for kDNA and SSUr-rDNA chromosomal.
|
kDNA (minicircle) |
SSUr-rDNA (chromosomal) |
|||
|---|---|---|---|---|
| Blood | Swab | Blood | Swab | |
| Positive | 47 (36.2%) | 38 (29.2%) | 38 (29.2%) | 32 (24.6%) |
| Negative | 83 (63.8%) | 92 (70.8%) | 92 (70.8%) | 98 (75.4%) |
Phi coefficient rφ (kDNA blood × SSUr-rDNA swab) p-value = 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



