
Submitted:
10 April 2025
Posted:
10 April 2025
You are already at the latest version
Abstract
Background/Objectives: Carvacrol is a naturally occurring phenolic monoterpene that is one of the major constituents of the essential oils of oregano (Origanum vulgare), thyme (Thymus vulgaris) and other herbs. Carvacrol has anti-inflammatory and antinociceptive effects. At the systemic level, carvacrol is able to activate and inhibit several second messengers and ion channels. However, there is no evidence on the peripheral antinociception of carvacrol and its probable mechanism of action. The aim of this study was to investigate whether the local antinociception of carvacrol involves the opioid receptor-nitric oxide (NO)-GMP-K+ channel pathway. Methods: Formalin (1%) was injected into the dorsal surface of the right hind paw of Wistar rats. Rats received a subcutaneous injection into the dorsal surface of the paw of vehicle or carvacrol (100-300 µg/paw). To determine whether the local antinociception induced by carvacrol was mediated by the opioid receptor-NO-cGMP-K+ channel pathway and a biguanide-dependent mechanism, the effect of pretreatment (10 min before formalin injection) with the corresponding vehicles, naltrexone, L-NAME, 1 H-(1,2,4)-oxadiazolo (4,2-a) quinoxalin-1-one (ODQ), K+ channel blockers, and metformin on the antinociceptive effects induced by local carvacrol (300 µg/paw) were evaluated. Results: Carvacrol produced antinociception during both phases of the formalin test. Naltrexone, L-NAME, ODQ, glibenclamide and glipizide (both ATP-sensitive K+ channel blockers), tetraethylammonium and 4-aminopyridine (both voltage-gated K+ channel blockers), apamin and charybdotoxin (both Ca2+-activated K+ channel blockers), and metformin reversed the carvacrol-induced peripheral antinociception. Conclusions: Local peripheral administration of carvacrol was able to produce significant antinociception and activate the opioid receptor-NO-cGMP-K+ channel pathway. In addition, carvacrol activated a metformin-dependent mechanism at the peripheral level.
Keywords:
carvacrol
; antinociception
; nitric oxide
; cGMP
; K+ channels
1. Introduction
Carvacrol, or isothymol (IUPAC name: 2-methyl-5-propan-2-ylphenol), is a naturally occurring phenolic monoterpene derivative of cymene [1]. A colorless to pale yellow liquid with a characteristic pungent odor, carvacrol is widely used as a flavoring and food additive and is classified as non-toxic for human consumption [1,2]. Carvacrol is one of the major constituents of the essential oils of oregano (Origanum vulgare), thyme (Thymus vulgaris), pepperwort (Lepidium flavum), and other herbs [1,2]. Studies have shown that carvacrol has hepatoprotective, antioxidant, neuroprotective, antidiabetic, anti-inflammatory, and antinociceptive effects [1,2,3,4]. The involvement of nitric oxide (NO), cGMP, ATP-sensitive K+ channel and voltage-gated- K+ channels has been implicated in some biological effects of carvacrol [5,6,7,8]. Other in situ and in vitro studies demonstrated the ability of carvacrol to activate TRPA1 and TRPV3 channels and to inhibit TRPM7, TRPV1, nicotinic, Ca2+ and voltage-gated Na+ channels [9,10,11,12,13,14,15,16].
In the antinociceptive activity, carvacrol was able to reduce the nociception produced in several tests [17,18,19,20]. As far as opioid receptors are concerned, there is both positive and negative evidence regarding their activation by carvacrol in models of nociception [18,20,21]. Carvacrol has also been suggested to activate the opioid receptor-NO-cGMP-K+ channel pathway at the systemic level [20]. These previous studies on antinociception have investigated the mechanisms of action of carvacrol at the systemic level and in isolated models. However, potential mechanisms of action of carvacrol at the peripheral level, such as the opioid receptor-NO-cGMP-K+ channel pathway [20,22,23,24,25,26,27,28,29,30,31], have not been investigated. Potassium (K+) and calcium (Ca2+) channels are the ultimate targets for reducing neuronal or cell excitability [32]. The K+ channels can be modulated by the NO-cGMP pathway as well as by other second messengers and drugs [32,33]. Therefore, our study was conducted to investigate the possible activation of opioid receptors, the NO-cGMP pathway, K+ channels, and a biguanide-dependent mechanism in the peripheral antinociceptive effects of carvacrol in the rat formalin test.
2. Materials and Methods
2.1. Animals
Male Wistar rats aged 7-9 weeks from our own breeding facilities were used in this study. Each rat was used in only one experiment and was killed in a CO2 chamber at the end of the experiments. The Institutional Animal Care and Use Committee (CINVESTAV, IPN, Ciudad de México, Mexico) approved the study protocol with registration number 0169-15, after which the animals were treated according to the Guiding Principle on Ethical Standards for Investigation in Animals [34].
2.2. Drugs
Carvacrol, naltrexone (opioid receptor blocker), metformin (hypoglycemic biguanide), NG-L-nitro-arginine methyl ester (L-NAME)(a NO synthase inhibitor), 1 H-(1,2,4)-oxadiazolo (4,2-a) quinoxalin-1-one (ODQ)(a NO-sensitive soluble guanylyl cyclase inhibitor), glibenclamide and glipizide (both ATP-sensitive K+ channel blockers; Kir6.1-2), tetraethylammonium chloride (TEA) and 4-aminopyridine (4-AP) (both voltage-gated K+ channel blockers; KV), apamin (a small conductance Ca2+-activated K+ channel blocker; KCa2.1-3), and charybdotoxin (a big conductance Ca2+-activated K+ channel blocker; KCa1.1), were purchased from Sigma-Aldrich (Toluca, Mexico). Carvacrol was diluted in a 2% Tween solution. Glibenclamide, glipizide and ODQ were dissolved in a 20% DMSO solution. L-NAME, TEA, 4-AP, apamin, charybdotoxin, naloxone, and metformin were dissolved in saline solution.
2.3. Measurement of Nociceptive Activity
The rat paw 1% formalin test was used to assess nociceptive and antinociceptive effects [26,27,28]. Nociceptive behavior was induced by the subcutaneous injection of 50 µL of 1% formalin into the dorsal region of the right hind paw, and then the nociceptive behavior was quantified as the number of one-minute flinches of the treated paw every 5 min up to 60 min after injection. The flinching behavior of the formalin-injected paw is biphasic [26,27,28].
2.4. Study Design
To determine the peripheral antinociceptive effect of carvacrol, rats in several independent groups were administered either the corresponding vehicle or increased doses of carvacrol (30-300 µg/paw) subcutaneously in the right hind paw of the rat 20 minutes before administration of 50 µL of 1% formalin in the same paw. To demonstrate that the effect of carvacrol was localized to the injection site, a dose of 300 µg/paw of carvacrol was administered to the contralateral (left) paw 20 minutes before formalin was administered to the right paw and the response was assessed.
In order to determine the possible involvement of opioid receptors, a biguanide-dependent mechanism, and NO and cGMP production in the peripheral carvacrol-induced antinociceptive effects, independent groups of rats were treated with the corresponding vehicles or naltrexone (50 µg /paw), metformin (400 µg/paw), L-NAME (100 µg/paw), or ODQ (100 µg/paw). Blockers were administered 10 min after the administration of carvacrol (300 µg/paw). Ten minutes after the administration of the blockers, the formalin solution was administered, and then the responses were scored as described above. To determine whether carvacrol-induced antinociception is mediated by activation of K+ channels, independent groups of rats were first treated with carvacrol (300 µg/paw). After carvacrol and 10 min before formalin, animals were treated with the corresponding vehicles or glibenclamide (100 µg/paw), glipizide (100 µg/paw), apamin (2 µg/paw), charybdotoxin (2 µg/paw), 4-AP (100 µg/paw), and TEA (100 µg/paw). Then, responses were evaluated. Doses and drug administration schedule for peripheral administration were selected based on previous reports [20,22,23,26,27,28,29,30,31] and on pilot experiments in our laboratory. Rats in all experimental groups were evaluated for possible side effects.
2.5. Data Analysis and Statistics
All experimental results are presented as the means ± S.E.M. for five animals per group. Curves were constructed by plotting the number of flinches versus time. The area under the curve of the number of flinches verses time (AUC) was calculated using the trapezoidal rule. Both phases of the test are reported. The results were analyzed by one-way analysis of variance using SPSS version 20 for Windows. Tukey’s test was then used to compare differences between the different groups. Differences were considered statistically significant when p <0.05.
3. Results
3.1. Antinociceptive Activity Produced by Carvacrol
Local peripheral injection of 1% formalin produced paw-flinching behavior representative of nociception. Local peripheral administration of carvacrol to the right hind paw significantly reduced the number of formalin-induced flinches, demonstrating a significant antinociceptive effect (p < 0.05; Figure 1a and b). The antinociceptive effect of carvacrol was dose-dependent. Carvacrol administration to the left (contralateral) paw did not significantly alter the nociception produced by formalin injection into the right paw (p > 0.05; Figure 1a and b). Carvacrol-induced antinociception was statistically significant in both phases of the 1% formalin test in rats (p < 0.05; Figure 1a and b).
3.2. Effect of Naltrexone, Metformin, and NO-cGMP Pathway Inhibitors on Carvacrol Antinociception
Naltrexone modified carvacrol-induced antinociception in both phases of the formalin test, demonstrating the involvement of opioid receptors (p < 0.05; Figure 2a and b). Local administration of metformin significantly reduced the carvacrol-induced antinociception on the phase two (p < 0.05; Figure 2b) but not on phase one (p > 0.05; Figure 2a) of the formalin test. When given with carvacrol vehicle, naltrexone and metformin did not significantly modify formalin-induced nociceptive responses (p > 0.05; Figure 1a and b).
Local peripheral administration of L-NAME and ODQ after carvacrol administration significantly reduced their antinociceptive effects during both phases of the formalin test (p < 0.05; Figure 3a and b). These results demonstrate the production of NO and cGMP in the antinociception produced by carvacrol at the peripheral level. When administered with carvacrol vehicle, L-NAME and ODQ did not significantly alter formalin-induced nociceptive responses (p > 0.05; Figure 3a and b).
3.3. Effect of the Administration of K+ Channel Blockers on the Antinociception of Carvacrol
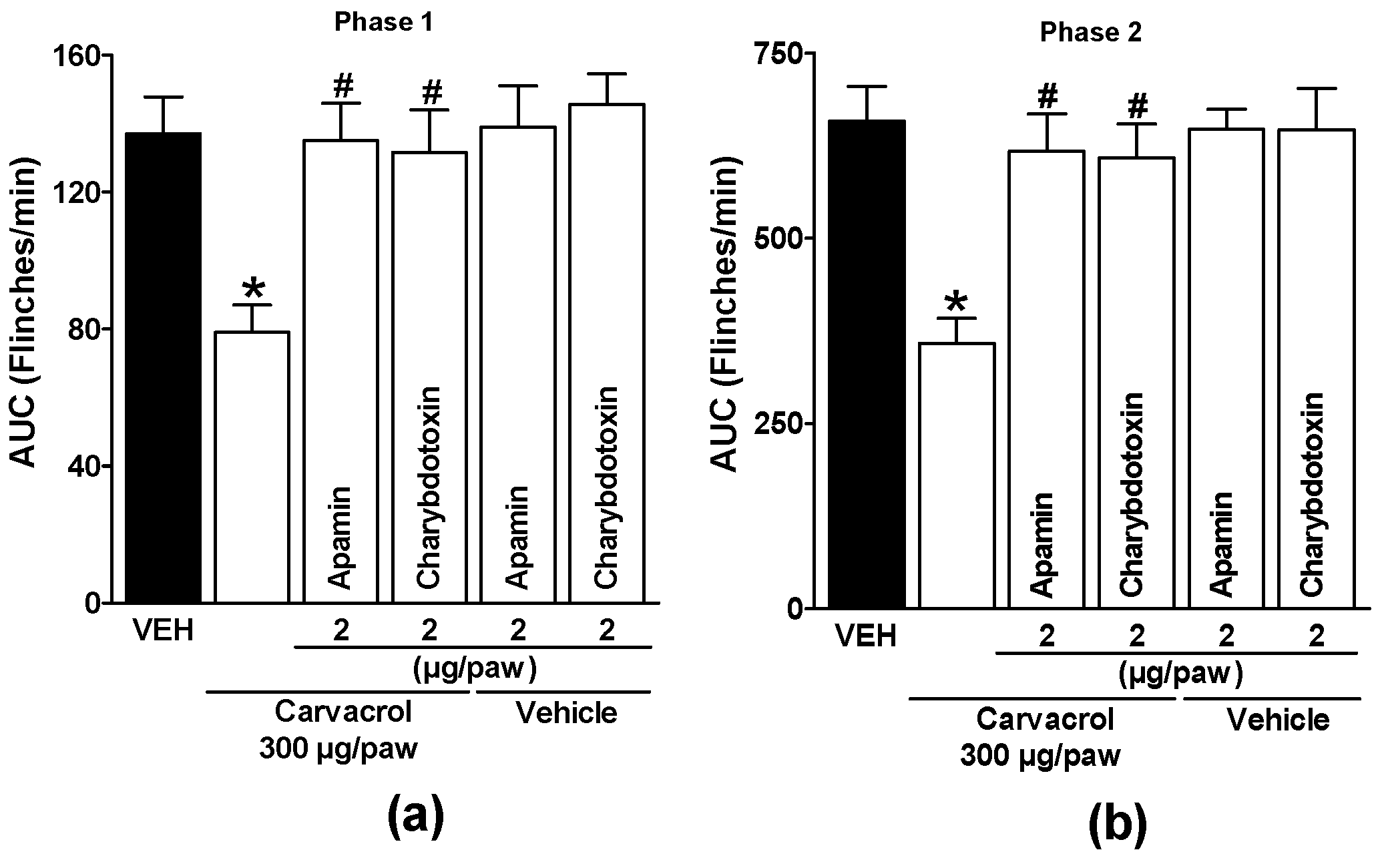
Subcutaneous administration of glipizide and glibenclamide significantly reduced carvacrol-induced antinociceptive effects in both phases of the test, demonstrating activation of ATP-sensitive K+ channels by carvacrol (p < 0.05; Figure 4a and b). Local peripheral injection of 4-AP and TEA significantly reduced carvacrol-induced antinociceptive effects during both phases of the formalin test, indicating activation of voltage-gated K+ channels by carvacrol (p < 0.05; Figure 5a and b). Subcutaneous peripheral administration of apamin and charybdotoxin significantly decreased the antinociceptive effects induced by carvacrol during both phases of the test, suggesting the activation of Ca2+-activated K+ channel by carvacrol (p < 0.05; Figure 6a and b). K+ channel blockers administered with the carvacrol vehicle did not significantly alter formalin-induced nociceptive behavior (p > 0.05: Figure 4, Figure 5 and Figure 6).
4. Discussion
There are currently a number of different herbal preparations used as complementary or alternative medicines. These herbal preparations contain a variety of chemical substances that produce a pharmacological effect (either therapeutic or undesirable). Carvacrol is a phenol with multiple biological activities, including antinociceptive and anti-inflammatory effects [4,17,18,19,20]. In the present study, local peripheral administration of carvacrol was able to significantly reduce the phase one (non-inflammatory phase) and the phase two (inflammatory phase) of the formalin test. These effects were localized to the injection site, as carvacrol administration in the contralateral paw was ineffective. There is evidence for activation of TRPA1 located in the primary afferent neurons during the phase 1 of the formalin test. During the phase 2, the activation of TRPA1 channels and the production of prostaglandins and interleukins are involved in a combination of peripheral depolarization and inflammatory sensitization in the spinal cord dorsal horn tissue [35,36]. Carvacrol has been shown to be a blocker of cation channels and an activator of TRPV3 channels [9,10,11,12,13,14,15,16]. Similarly, carvacrol was able to activate and subsequently desensitize TRPA1 channels in HEK293 cells [9]. In this sense, the probable inactivation of TRPA1 and voltage-gated cation channels produced by carvacrol in its antinociceptive effects determined in the present study cannot be excluded [10,14,16]. As noted above, carvacrol inhibits inflammatory pain in several experimental models, including the second phase of the formalin test. However, a previous study found that carvacrol was unable to block the arachidonic inflammatory cascade (involving the production of prostaglandins, leukotrienes, or epoxyeicosatrienoic acid derivatives) in isolated rat aortic rings [7]. Therefore, further studies in other experimental models are needed to determine the ability of carvacrol to inhibit pro-inflammatory mediators such as prostaglandins, cytokines, chemokines, proteases, neuropeptides, and growth factors.
Opioid drugs and some phytoconstituents are able to reduce both phases of the formalin test [29,30,36]. On the other hand, nonsteroidal anti-inflammatory drugs (NSAIDs) suppress only the second phase [26,31,36,37]. In the present study, local peripheral injection of naltrexone significantly reversed carvacrol-induced antinociception. This result is consistent with the ability of naloxone to block the systemic antinociception induced by the Ziziphora clinopodioides (carvacrol in 65.2%) and Thymus persicus (carvacrol in 32%) essential oils in the formalin test [20,21]. In contrast, opioid receptors are not involved in the systemic antinociceptive effect induced by carvacrol on both phases of the formalin test in mice [18]. It is likely that these discrepancies are due to the different species used and the different routes and times of administration of the drugs.
Nitric oxide (NO) is a gas that is synthesized in tissues by the enzyme NO synthase (NOS), which uses the semi-essential amino acid L-arginine to form citrulline and NO in a 1:1 ratio. The released NO travels to target cells and activates the enzyme soluble guanylate cyclase (sGC) to synthesize cGMP from guanosine triphosphate (GTP). cGMP acts as a second messenger, either directly or through protein kinase (PKG) activation, to open potassium channels and modulate calcium channels. The action of cGMP is terminated by its inactivation by means of a phosphodiesterase enzyme [38,39,40]. The production of NO and cGMP is involved in immunity, inflammation, platelet aggregation, nociception, smooth muscle relaxation, and other pathological and physiological processes [38,39,40]. In some models of nociception, the production of NO and cGMP is involved in the antinociceptive effects of several drugs, including NSAIDs, phytoconstituents, opioids, and neuromodulators [20,22,23,24,25,26,27,28,29]. Local peripheral administration of L-NAME, a NO synthesis inhibitor and ODQ, a NO-sensitive soluble guanylyl cyclase inhibitor [38,39,40], significantly reversed the antinociceptive effect of carvacrol in the present study. These results are consistent with the production of NO and cGMP in the gastroprotection produced by carvacrol in rats and in the antinociception produced by of Lippia origanoides (carvacrol in 53.9%) and Thymus persicus aerial parts (carvacrol in 32.2%) essential oils [5,8,20]. In contrast, neither L-NAME nor ODQ blocked the endothelium-dependent relaxation of carvacrol on rat aorta [7]. In the latter case, the discrepancy is mainly due to the difference in the experimental model used. To our knowledge, this is the first report on the ability of locally administered carvacrol to activate the NO-cGMP pathway to produce peripheral antinociception.
Potassium channels are integral membrane proteins that regulate the transport of K+ ions across the membrane. In excitable cells, such as neurons, K+ channel activity always reduces excitability by moving the membrane potential away from the firing threshold (hyperpolarization) [32]. There are four different types of K+ channels according to their structure and functional properties [32]. K+ channels can be modulated by membrane depolarization, intracellular calcium, and second messengers such as NO and cGMP and many drugs [32,33]. Abundant evidence has demonstrated the involvement of some K+ channels in systemic or local antinociception induced by various types of analgesic and non-analgesic drugs [22,24,25,26,27,28,29,30,31,41]. In the present study, local peripheral injection of glibenclamide and glipizide was able to block the effect of carvacrol, suggesting that carvacrol is able to activate, in addition to the NO-cGMP pathway, the ATP-sensitive K+ channels to produce its peripheral antinociceptive effect. These results with sulfonylureas are consistent with the involvement of the ATP-sensitive K+ channel in the gastroprotection produced by carvacrol, the prevention of apoptosis on rat testis by carvacrol, and the antinociception of the essential oil of Thymus persicus aerial parts (carvacrol in 32%) in the formalin test [5,6,20]. In other experimental groups of our study, the voltage-gated K+ channel inhibitors 4-AP and TEA and the Ca2+-activated K+ channel blockers apamin and charybdotoxin blocked the effects of carvacrol, suggesting the activation of these channels in its antinociception in the periphery. These results are consistent with the carvacrol-induced relaxation of isolated rat aortic rings by activation of voltage-gated K+ channels and the relaxant effects of Lippia origanoides essential oil (containing 53.9% carvacrol) in tracheal tissue by activation of voltage-gated and Ca2+-activated K+ channels [7,8].
Liver tissue is the primary site of action for the hypoglycemic drug metformin. Metformin is a first-line treatment for type 2 diabetes. Metformin has been shown to be involved in nociceptive and antinociceptive responses in some experimental models. In the present study, metformin was able to reverse the antinociceptive effect produced by carvacrol. This latter result is consistent with the ability of metformin to block the antinociceptive effects produced by citral, pamabrom, diclofenac, indomethacin, and metamizole [30,31,37,42]. Therefore, preclinical and clinical studies to evaluate the potential for metformin to interact with terpenoids or NSAIDs are recommended.
5. Conclusions
Local peripheral administration of carvacrol was able to produce significant antinociception in both phases of the 1% rat formalin test. The mechanism of action of the peripheral antinociceptive activity of carvacrol involves activation of the opioid receptor-NO-cGMP pathway and K+ channels. A metformin-dependent mechanism is also involved in carvacrol-induced antinociception.
Author Contributions
Conceptualization, M.I.O., R.C.C., E.F.M., V.M.M.P., G.C.H., and M.P.G.G.; methodology, M.I.O. and M.P.G.G.; formal analysis, M.I.O., R.C.C., E.F.M., V.M.M.P., G.C.H., and M.P.G.G.; investigation, M.I.O., R.C.C., and G.C.H.; resources, M.I.O., G.C.H. and M.P.G.G.; data curation, M.I.O., R.C.C., E.F.M., V.M.M.P., G.C.H., and M.P.G.G.; writing—original draft preparation, M.I.O., R.C.C., E.F.M., V.M.M.P., G.C.H., and M.P.G.G.; writing—review and editing, M.I.O., R.C.C., E.F.M., V.M.M.P., G.C.H., and M.P.G.G.; supervision, M.I.O., R.C.C. and G.C.H.; project administration, M.I.O., G.C.H. and M.P.G.G.; funding acquisition, M.I.O., R.C.C., E.F.M., V.M.M.P., G.C.H., and M.P.G.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The Institutional Animal Care and Use Committee (CINVESTAV, IPN, Ciudad de México, Mexico) approved the study protocol with registration number 0169-15, after which the animals were treated according to the Guiding Principle on Ethical Stand-ards for Investigation in Animals [34].
Data Availability Statement
The raw data supporting the conclusion of this article will be made available by the author, without undue reservation.
Acknowledgments
Authors acknowledge to M.V.Z. Ricardo Gaxiola Centeno, M.V.Z. Rafael Leyva Muñoz, M.V.Z. Benjamín Chávez Álvarez, M.V.Z. Carlos Giovanni Sam Miranda and René Pánfilo Morales from Cinvestav-UPEAL for their technical assistance in this project.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ANOVA | Analysis of variance |
| ATP | Adenosine triphosphate |
| AUC | Area under the curve |
| cGMP | Cyclic Guanosine monophosphate |
| CINVESTAV | Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional |
| CL | Contralateral |
| IL | Ipsilateral |
| IPN | Instituto Politécnico Nacional |
| IUPAC | International Union of Pure and Applied Chemistry |
| L-NAME | NG-L-nitro-arginine methyl ester |
| MDPI | Multidisciplinary Digital Publishing Institute |
| NO | Nitric Oxide |
| NOS | Nitric Oxide synthase |
| NSAIDs | Nonsteroidal anti-inflammatory drugs |
| ODQ | 1 H-(1,2,4)-oxadiazolo (4,2-a) quinoxalin-1-one |
| PKG | Protein kinase |
| SEM | Standard error of the mean |
| sGC | Soluble guanylate cyclase |
| TEA | Tetraethylammonium chloride |
| VEH | Vehicle |
| 4-AP | 4-aminopyridine |
References
- Imran, M.; Aslam, M.; Alsagaby, S.A.; Saeed, F.; Ahmad, I.; Afzaal, M.; Arshad, M.U.; Abdelgawad, M.A.; El-Ghorab, A.H.; Khames, A.; et al. Therapeutic application of carvacrol: A comprehensive review. Food Sci. Nutr. 2022, 10, 3544–3561. [CrossRef]
- Suntres, Z.E.; Coccimiglio, J.; Alipour, M. The Bioactivity and Toxicological Actions of Carvacrol. Crit. Rev. Food Sci. Nutr. 2014, 55, 304–318. [CrossRef]
- Azizi, Z.; Majlessi, N.; Choopani, S.; Naghdi, N. Neuroprotective effects of carvacrol against Alzheimer's disease and other neurodegenerative diseases: A review. Avicenna J. Phytomed. 2022, 12, 371–387. [CrossRef]
- Khazdair, M.R.; Ghorani, V.; Boskabady, M.H. Experimental and clinical evidence on the effect of carvacrol on respiratory, allergic, and immunologic disorders: A comprehensive review. BioFactors 2022, 48, 779–794. [CrossRef]
- Oliveira, I.S.; da Silva, F.V.; Viana, A.F.; dos Santos, M.R.; Quintans-Júnior, L.J.; Martins Mdo, C.; et al. Gastroprotective activity of carvacrol on experimentally induced gastric lesions in rodents. Naunyn Schmiedebergs Arch. Pharmacol. 2012, 385, 899-908.
- Balci, C,N.; Firat, T.; Acar, N.; Kukner, A. Carvacrol treatment opens Kir6.2 ATP-dependent potassium channels and prevents apoptosis on rat testis following ischemia-reperfusion injury model. Rom. J. Morphol. Embryol. 2021, 62, 179-190.
- Testai, L.; Chericoni, S.; Martelli, A.; Flamini, G.; Breschi, M.C.; Calderone, V. Voltage-operated potassium (Kv) channels contribute to endothelium-dependent vasorelaxation of carvacrol on rat aorta. J. Pharm. Pharmacol. 2016, 68, 1177–1183. [CrossRef]
- Menezes, P.M.N.; Brito, M.C.; de Paiva, G.O.; dos Santos, C.O.; de Oliveira, L.M.; Ribeiro, L.A.d.A.; de Lima, J.T.; Lucchese, A.M.; Silva, F.S. Relaxant effect of Lippia origanoides essential oil in guinea-pig trachea smooth muscle involves potassium channels and soluble guanylyl cyclase. J. Ethnopharmacol. 2018, 220, 16–25. [CrossRef]
- Xu, H.; Delling, M.; Jun, J.C.; Clapham, D.E. Oregano, thyme and clove-derived flavors and skin sensitizers activate specific TRP channels. Nat. Neurosci. 2006, 9, 628–635. [CrossRef]
- Gonçalves, J.C.R.; Alves, A.d.M.H.; de Araújo, A.E.V.; Cruz, J.S.; Araújo, D.A.M. Distinct effects of carvone analogues on the isolated nerve of rats. Eur. J. Pharmacol. 2010, 645, 108–112. [CrossRef]
- Parnas, M.; Peters, M.; Dadon, D.; Lev, S.; Vertkin, I.; Slutsky, I.; Minke, B. Carvacrol is a novel inhibitor of Drosophila TRPL and mammalian TRPM7 channels. Cell Calcium 2009, 45, 300–309. [CrossRef]
- Schwarz, M.G.; Namer, B.; Reeh, P.W.; Fischer, M.J. TRPA1 and TRPV1 Antagonists Do Not Inhibit Human Acidosis-Induced Pain. J. Pain 2017, 18, 526–534. [CrossRef]
- Luo, Q.-T.; Fujita, T.; Jiang, C.-Y.; Kumamoto, E. Carvacrol presynaptically enhances spontaneous excitatory transmission and produces outward current in adult rat spinal substantia gelatinosa neurons. Brain Res. 2014, 1592, 44–54. [CrossRef]
- Joca, H.C.; Vieira, D.C.O.; Vasconcelos, A.P.; Araújo, D.A.M.; Cruz, J.S. Carvacrol modulates voltage-gated sodium channels kinetics in dorsal root ganglia. Eur. J. Pharmacol. 2015, 756, 22–29. [CrossRef]
- Lozon, Y.; Sultan, A.; Lansdell, S.J.; Prytkova, T.; Sadek, B.; Yang, K.-H.S.; Howarth, F.C.; Millar, N.S.; Oz, M. Inhibition of human α7 nicotinic acetylcholine receptors by cyclic monoterpene carveol. Eur. J. Pharmacol. 2016, 776, 44–51. [CrossRef]
- Muñoz-Pérez, V.M.; Ortiz, M.I.; Gerardo-Muñoz, L.S.; Cariño-Cortés, R.; Salas-Casas, A. Tocolytic Effect of the Monoterpenic Phenol Isomer, Carvacrol, on the Pregnant Rat Uterus. Chin. J. Physiol. 2020, 63, 204–210. [CrossRef]
- Guimarães, A.G.; Oliveira, G.F.; Melo, M.S.; Cavalcanti, S.C.; Antoniolli, A.R.; Bonjardim, L.R.; Silva, F.A.; Santos, J.P.A.; Rocha, R.F.; Moreira, J.C.F.; et al. Bioassay-guided Evaluation of Antioxidant and Antinociceptive Activities of Carvacrol. Basic Clin. Pharmacol. Toxicol. 2010, 107, 949–957. [CrossRef]
- Melo, F.H.C.; Rios, E.R.V.; Rocha, N.F.M.; Citó, M.D.C.d.O.; Fernandes, M.L.; de Sousa, D.P.; de Vasconcelos, S.M.M.; de Sousa, F.C.F. Antinociceptive activity of carvacrol (5-isopropyl-2-methylphenol) in mice. J. Pharm. Pharmacol. 2012, 64, 1722–1729. [CrossRef]
- de Oliveira, A.S.; Llanes, L.C.; Nunes, R.J.; Nucci-Martins, C.; de Souza, A.S.; Palomino-Salcedo, D.L.; Dávila-Rodríguez, M.J.; Ferreira, L.L.G.; Santos, A.R.S.; Andricopulo, A.D. Antioxidant Activity, Molecular Docking, Quantum Studies and In Vivo Antinociceptive Activity of Sulfonamides Derived From Carvacrol. Front. Pharmacol. 2021, 12. [CrossRef]
- Abed, D.Z.; Sadeghian, R.; Mohammadi, S.; Akram, M. Thymus persicus (Ronniger ex Rech. f.) Jalas alleviates nociceptive and neuropathic pain behavior in mice: Multiple mechanisms of action. J. Ethnopharmacol. 2021, 283, 114695. [CrossRef]
- Mohammadifard, F.; Alimohammadi, S. Chemical Composition and Role of Opioidergic System in Antinociceptive Effect of Ziziphora Clinopodioides Essential Oil. Basic Clin. Neurosci. J. 2018, 9, 357–365. [CrossRef]
- Alizamani, E.; Ghorbanzadeh, B.; Naserzadeh, R.; Mansouri, M.T. Montelukast, a cysteinyl leukotriene receptor antagonist, exerts local antinociception in animal model of pain through the L-arginine/nitric oxide/cyclic GMP/K(ATP) channel pathway and PPARgamma receptors. Int. J. Neurosci. 2020, 131, 1004-1011.
- Amarante, L.H.; Duarte, I.D. The kappa-opioid agonist (+/-)-bremazocine elicits peripheral antinociception by activation of the L-arginine/nitric oxide/cyclic GMP pathway. Eur. J. Pharmacol. 2002, 454, 19-23.
- Ghorbanzadeh, B.; Kheirandish, V.; Mansouri, M.T. Involvement of the L-arginine/Nitric Oxide/Cyclic GMP/KATP Channel Pathway and PPARγ Receptors in the Peripheral Antinociceptive Effect of Carbamazepine. Drug Res. 2019, 69, 650–657. [CrossRef]
- Ghorbanzadeh, B.; Mansouri, M.T.; Naghizadeh, B.; Alboghobeish, S. Local antinociceptive action of fluoxetine in the rat formalin assay: role of l-arginine/nitric oxide/cGMP/KATP channel pathway. Can. J. Physiol. Pharmacol. 2018, 96, 165–172. [CrossRef]
- Ortiz, M.I.; Granados-Soto, V.; Castañeda-Hernández, G. The NO-cGMP-K+ channel pathway participates in the antinociceptive effect of diclofenac, but not of indomethacin. Pharmacol. Biochem. Behav. 2003, 76, 187-195.
- Ortiz, M.I.; Medina-Tato, D.A.; Sarmiento-Heredia, D.; Palma-Martínez, J.; Granados-Soto, V. Possible activation of the NO–cyclic GMP–protein kinase G–K+ channels pathway by gabapentin on the formalin test. Pharmacol. Biochem. Behav. 2006, 83, 420–427. [CrossRef]
- Ortiz, M.I.; Cariño-Cortés, R.; Castañeda-Hernández, G. Participation of the opioid receptor - nitric oxide - cGMP - K(+) channel pathway in the peripheral antinociceptive effect of nalbuphine and buprenorphine in rats. Can. J. Physiol. Pharmacol. 2020, 98, 753-762.
- Ortiz, M.I.; Cariño-Cortés, R.; Muñoz-Pérez, V.M.; Salas-Casas, A.; Castañeda-Hernández, G. Role of the NO-cGMP-K+ channels pathway in the peripheral antinociception induced by α-bisabolol. Can. J. Physiol. Pharmacol. 2021, 99, 1048–1056. [CrossRef]
- Ortiz, M.I.; Cariño-Cortés, R.; Muñoz-Pérez, V.M.; Medina-Solís, C.E.; Castañeda-Hernández, G. Citral inhibits the nociception in the rat formalin test: effect of metformin and blockers of opioid receptor and the NO-cGMP-K+ channel pathway. Can. J. Physiol. Pharmacol. 2022, 100, 306–313. [CrossRef]
- Ortiz, M.I.; Cariño-Cortés, R.; Castañeda-Hernández, G.; Medina-Solís, C.E. Effect of nitric oxide-cyclic GMP-K+ channel pathway blockers; naloxone and metformin; on the antinociception induced by the diuretic pamabrom. Can. J. Physiol. Pharmacol. 2023, 101, 41-51.
- Alexander, S.P.H.; Mathie, A.; Peters, J.A.; Veale, E.L.; Striessnig, J.; Kelly, E.; et al. The Concise Guide to Pharmacology 2019/20: Ion channels. Br. J. Pharmacol. 2019, 176, S142–S228.
- Dascal, N.; Kahanovitch, U. The Roles of Gβγ and Gα in Gating and Regulation of GIRK Channels. Int. Rev. Neurobiol. 2015, 123, 27–85.
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. PAIN® 1983, 16, 109–110. [CrossRef]
- McNamara, C.R.; Mandel-Brehm, J.; Bautista, D.M.; Siemens, J.; Deranian, K.L.; Zhao, M.; Hayward, N.J.; Chong, J.A.; Julius, D.; Moran, M.M.; et al. TRPA1 mediates formalin-induced pain. Proc. Natl. Acad. Sci. USA 2007, 104, 13525–13530. [CrossRef]
- Muley, M.M.; Krustev, E.; McDougall, .J.J. Preclinical Assessment of Inflammatory Pain. CNS Neurosci. Ther. 2016, 22, 88–101.
- Ortiz, M.I. Metformin and phenformin block the peripheral antinociception induced by diclofenac and indomethacin on the formalin test. Life Sci. 2012, 90, 8–12. [CrossRef]
- Mónica, F.; Bian, K.; Murad, F. The Endothelium-Dependent Nitric Oxide-cGMP Pathway. Adv. Pharmacol. 2016, 77, 1–27.
- Xiao, S.; Li, Q.; Hu, L.; Yu, Z.; Yang, J.; Chang, Q.; et al. Soluble Guanylate Cyclase Stimulators and Activators: Where are We and Where to Go? Mini Rev. Med. Chem. 2019, 19, 1544–1557.
- Koesling, D.; Mergia, E.; Russwurm, M. Physiological Functions of NO-Sensitive Guanylyl Cyclase Isoforms. Curr. Med. Chem. 2016, 23, 2653–2665. [CrossRef]
- Ocaña, M.; Cendán, C.M.; Cobos, E.J.; Entrena, J.M.; Baeyens, J.M. Potassium channels and pain: present realities and future opportunities. Eur. J. Pharmacol. 2004, 500, 203–219. [CrossRef]
- Guzmán-Priego, C.G.; Méndez-Mena, R.; Baños-González, M.A.; Araiza-Saldaña, C.I.; Castañeda-Corral, G.; Torres-López, J.E. Antihyperalgesic Effects of Indomethacin, Ketorolac, and Metamizole in Rats: Effects of Metformin. Drug Dev. Res. 2017, 78, 98–104. [CrossRef]
Figure 1.
Local peripheral antinociception of carvacrol during: (a) phase one and (b) phase two of the formalin test. Rats were pretreated with s.c. injection of vehicle (VEH) or carvacrol into either the right (ipsilateral, IL) or left (contralateral, CL) paw before formalin injection. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
Figure 1.
Local peripheral antinociception of carvacrol during: (a) phase one and (b) phase two of the formalin test. Rats were pretreated with s.c. injection of vehicle (VEH) or carvacrol into either the right (ipsilateral, IL) or left (contralateral, CL) paw before formalin injection. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
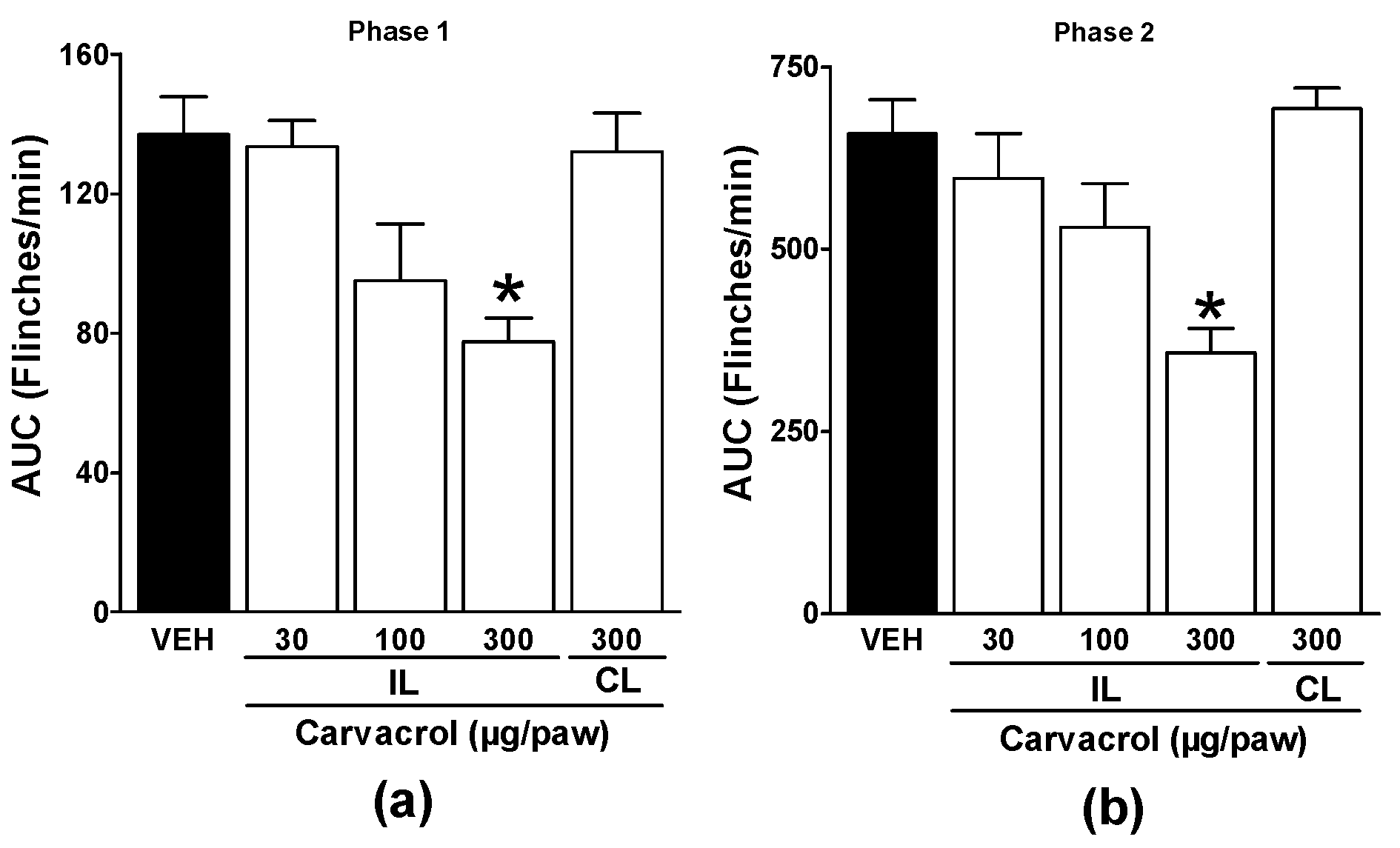
Figure 2.
Local peripheral effect of naltrexone and metformin on the carvacrol-induced antinociceptive activity during (a) phase one and (b) phase two of the formalin test. Rats received a local injection of carvacrol (-20 min) or vehicle and then naltrexone or metformin (-10 min) into the right paw. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) and #significantly different from the carvacrol group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
Figure 2.
Local peripheral effect of naltrexone and metformin on the carvacrol-induced antinociceptive activity during (a) phase one and (b) phase two of the formalin test. Rats received a local injection of carvacrol (-20 min) or vehicle and then naltrexone or metformin (-10 min) into the right paw. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) and #significantly different from the carvacrol group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
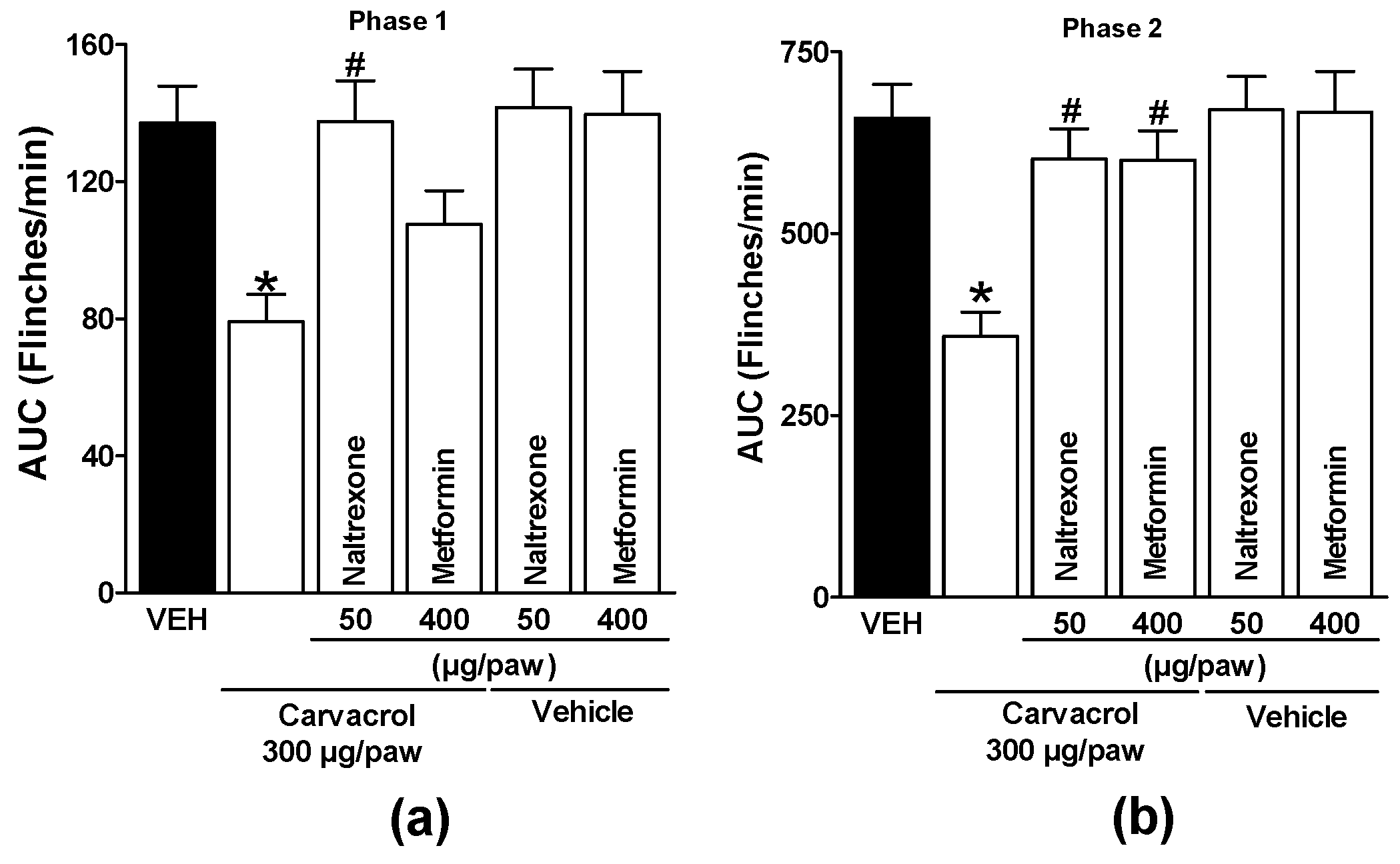
Figure 3.
Local peripheral effect of L-NAME and ODQ on the carvacrol-induced antinociceptive activity during (a) phase one and (b) phase two of the formalin test. Rats received a local injection of carvacrol (-20 min) or vehicle and then L-NAME or ODQ (-10 min) into the right paw. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) and #significantly different from the carvacrol group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
Figure 3.
Local peripheral effect of L-NAME and ODQ on the carvacrol-induced antinociceptive activity during (a) phase one and (b) phase two of the formalin test. Rats received a local injection of carvacrol (-20 min) or vehicle and then L-NAME or ODQ (-10 min) into the right paw. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) and #significantly different from the carvacrol group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
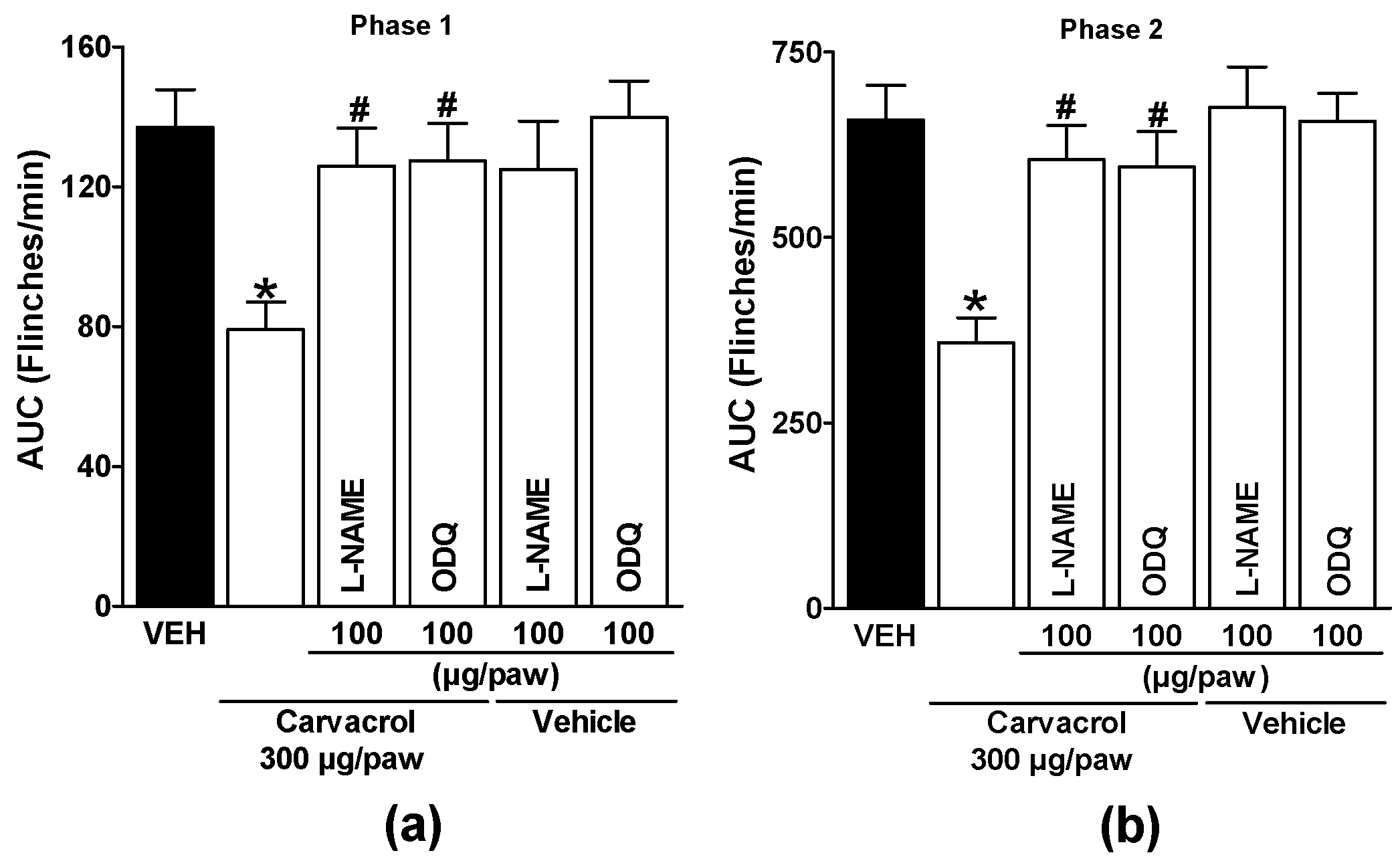
Figure 4.
Local peripheral effect of glibenclamide and glipizide on the carvacrol-induced antinociceptive activity during (a) phase one and (b) phase two of the formalin test. Rats received a local injection of carvacrol (-20 min) or vehicle and then glibenclamide or glipizide (-10 min) into the right paw. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) and #significantly different from the carvacrol group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
Figure 4.
Local peripheral effect of glibenclamide and glipizide on the carvacrol-induced antinociceptive activity during (a) phase one and (b) phase two of the formalin test. Rats received a local injection of carvacrol (-20 min) or vehicle and then glibenclamide or glipizide (-10 min) into the right paw. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) and #significantly different from the carvacrol group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
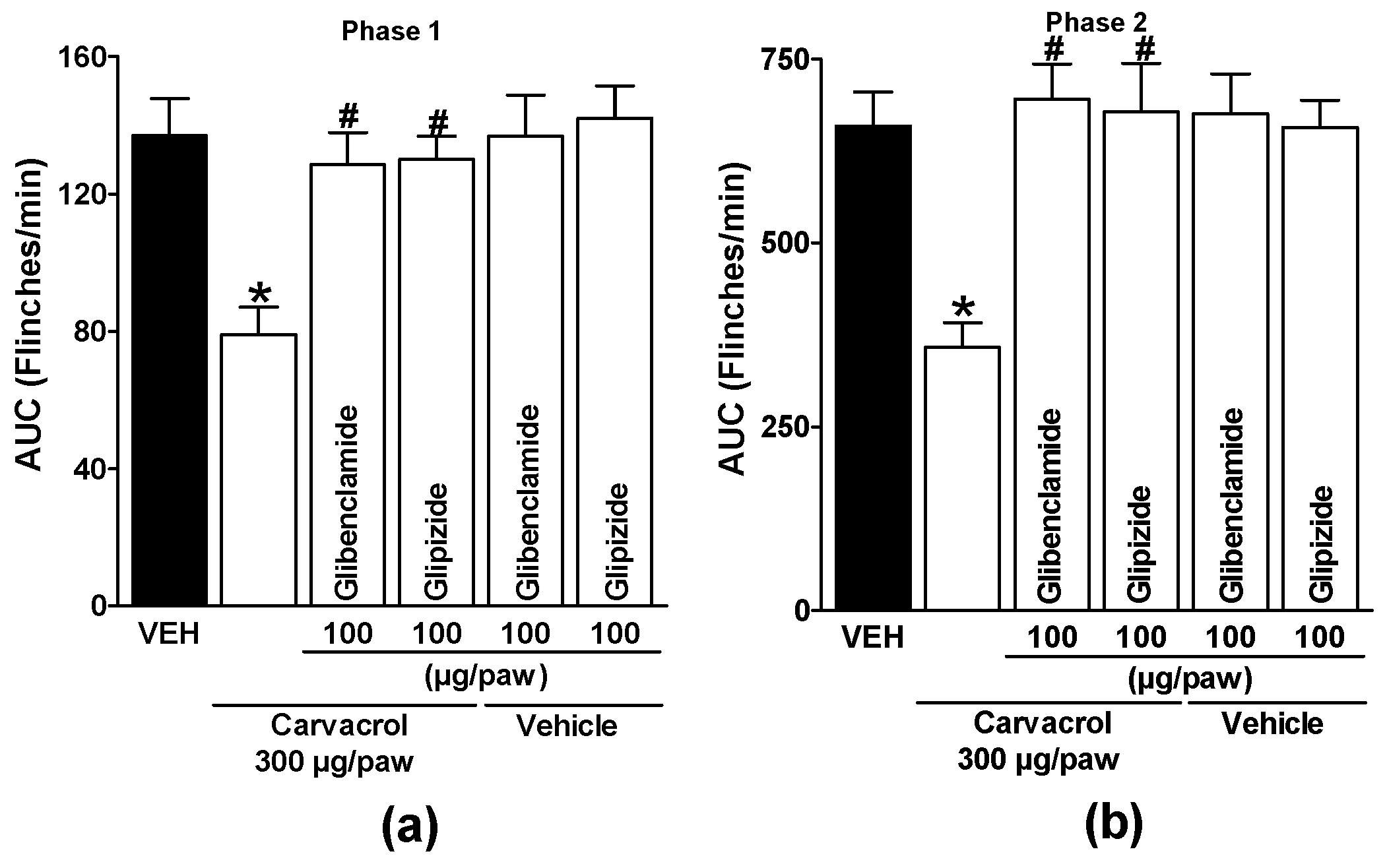
Figure 5.
Local peripheral effect of 4-AP and TEA on the carvacrol-induced antinociceptive activity during (a) phase one and (b) phase two of the formalin test. Rats received a local injection of carvacrol (-20 min) or vehicle and then 4-AP or TEA (-10 min) into the right paw. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) and #significantly different from the carvacrol group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
Figure 5.
Local peripheral effect of 4-AP and TEA on the carvacrol-induced antinociceptive activity during (a) phase one and (b) phase two of the formalin test. Rats received a local injection of carvacrol (-20 min) or vehicle and then 4-AP or TEA (-10 min) into the right paw. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) and #significantly different from the carvacrol group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
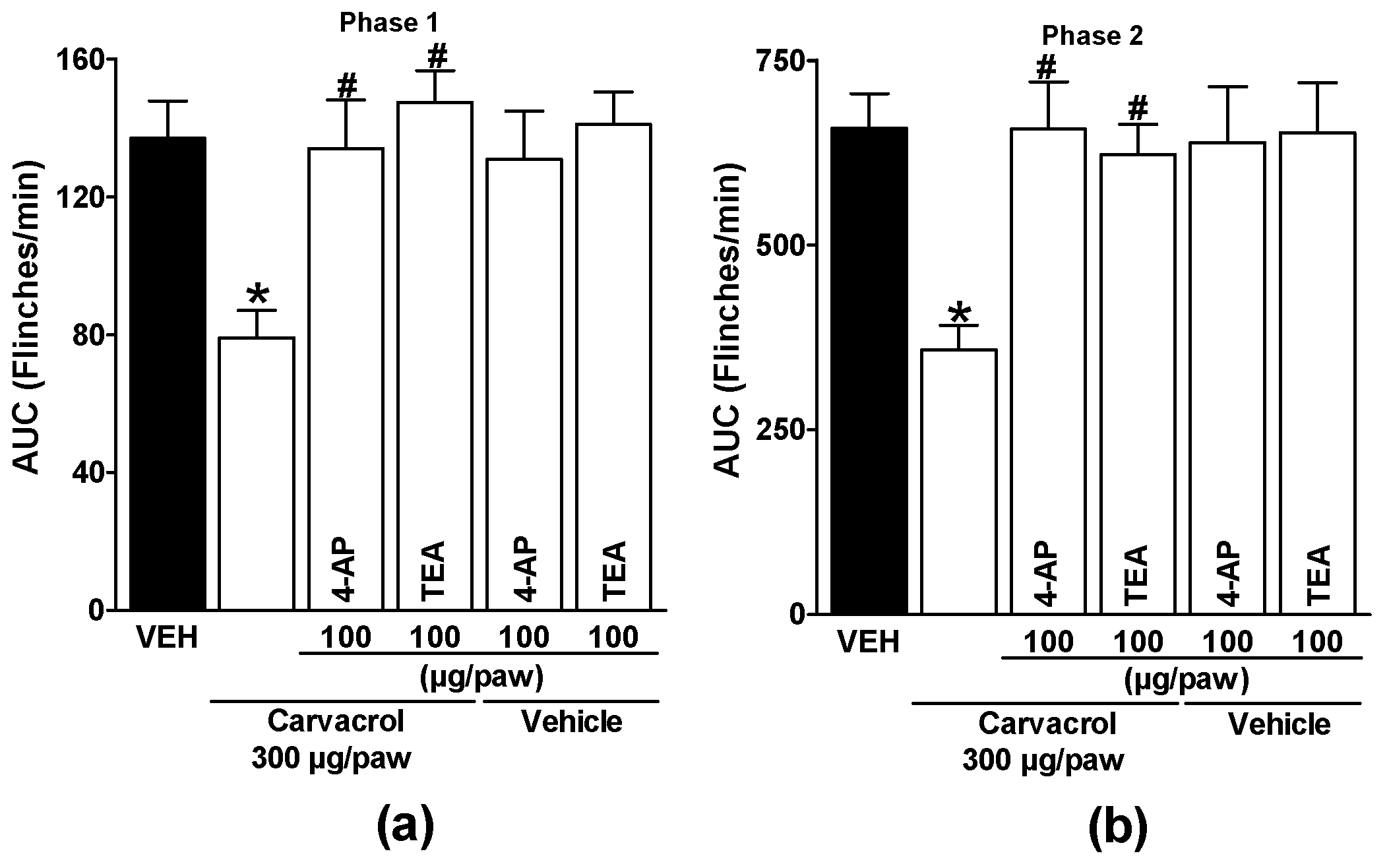
Figure 6.
Local peripheral effect of apamin and charybdotoxin on the carvacrol-induced antinociceptive activity during (a) phase one and (b) phase two of the formalin test. Rats received a local injection of carvacrol (-20 min) or vehicle and then apamin or charybdotoxin (-10 min) into the right paw. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) and #significantly different from the carvacrol group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
Figure 6.
Local peripheral effect of apamin and charybdotoxin on the carvacrol-induced antinociceptive activity during (a) phase one and (b) phase two of the formalin test. Rats received a local injection of carvacrol (-20 min) or vehicle and then apamin or charybdotoxin (-10 min) into the right paw. Data are expressed as the area under the curve of number of flinches versus time (AUC) for both phases. Each point represents to the mean ± S.E.M. of 5 animals. *Significantly different from the vehicle group (P < 0.05) and #significantly different from the carvacrol group (P < 0.05) as determined by a one-way analysis of variance (ANOVA) followed by Dunnett’s test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



