
Submitted:
06 November 2024
Posted:
07 November 2024
You are already at the latest version
Abstract
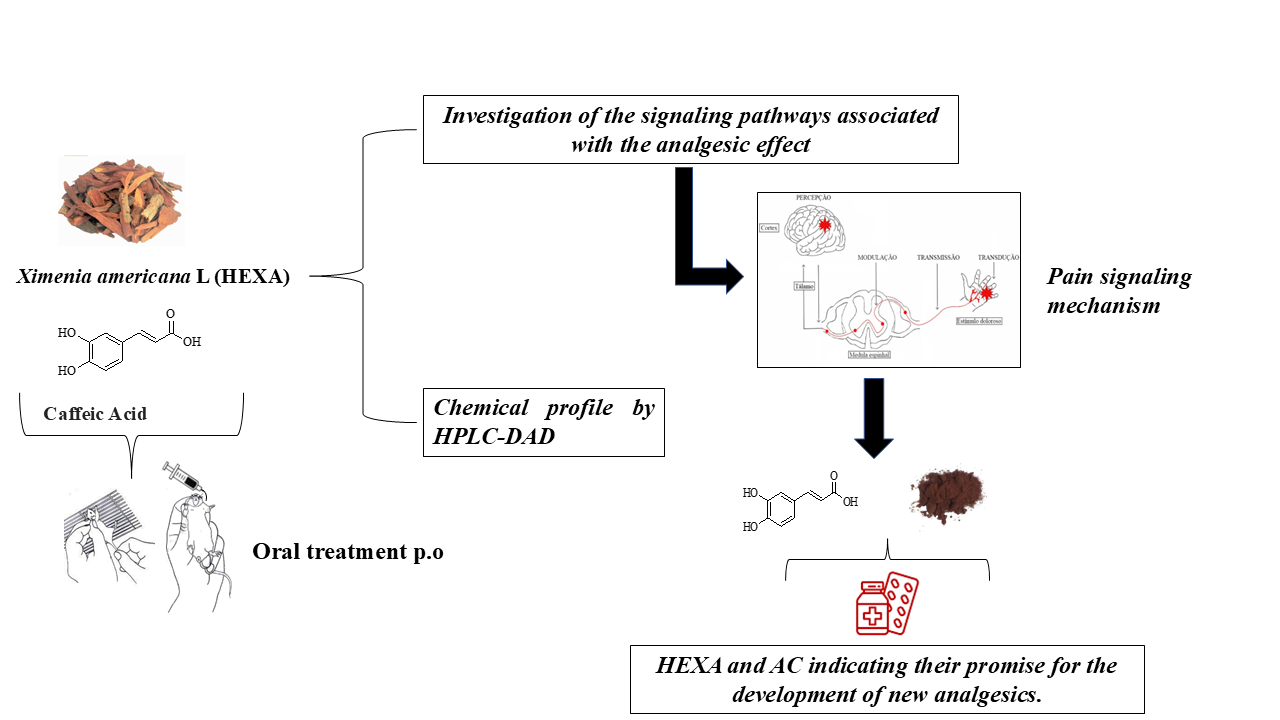
Ximenia americana L. (Olacaceae) has traditionally been used in folk medicine in the African continent and Brazilian Northeast to treat diseases such as diarrhea, fever, wounds, and pain. This study evaluated the antinociceptive effect of the Ximenia americana L. bark extract (HEXA) and its primary component, caffeic acid (CA), through in vivo assays. The antinociceptive properties were assessed using abdominal writhing, hot plate, and Von Frey tests. Additionally, the study investigated the modulation of various pain signaling pathways using a pharmacological approach. The results demonstrated that all doses of the HEXA extract significantly increased latency in the hot plate test, decreased the number of abdominal contortions, reduced hyperalgesia in the Von Frey test, and inhibited both phases of the formalin test. Caffeic acid reduced licking time in the first phase of the formalin test at all doses, with the highest dose showing significant effects in the second phase. The HEXA extract potentially modulated α2-adrenergic, nitric oxide, glutamatergic, vanilloid, cyclic guanosine monophosphate, and K+-ATP channel-dependent pathways. Conversely, caffeic acid influenced the opioid, glutamatergic, and vanilloid pathways while inhibiting nitric oxide and cyclic guanosine monophosphate. HEXA and CA exhibit significant antinociceptive effects due to their potential interference in multiple pain signaling pathways. While the molecular targets remain to be fully investigated, HEXA and CA demonstrate significant potential for the development of new analgesic drugs.
Keywords:
Ximenia americana L.
; caffeic acid
; antinociceptive
; anti-inflammatory
; mechanism of action
; pain
1. Introduction
Pain is a sensory and emotional experience that signals actual or potential harm to the body, thus providing an essential protective function against threats to the organism's health [1]. Pain can be classified based on duration as acute, subchronic, or chronic [2], and based on the origin of the signaling stimulus as neuropathic or inflammatory [1].
Acute pain is immediate and prevents damage and protects injured tissue [2]. On the other hand, chronic pain lasts more than ten days. This issue is a major global health concern that impacts people in physical, economic, emotional, and social ways [3,4]. In both cases, the pain usually requires management, which is mainly performed with the use of analgesic drugs.
Nonsteroidal anti-inflammatory drugs (NSAIDs) are the most used drugs in the treatment of acute, moderate, or chronic pain [5]. Their mechanism of action involves the peripheral and central inhibition of the cyclooxygenase enzymes (COX-1 and COX-2), which inhibits the production of lipid mediators such as prostaglandins, thromboxanes, and leukotrienes [6]. However, the long-term use of NSAIDs is associated with severe adverse effects, including nausea, dyspepsia, abdominal pain, skin reactions, and nephropathy, among others [7]. In addition, NSAIDs present significant therapeutic limitations, so in many cases, the use of opioid analgesics is necessary. However, these drugs pose even more significant risks for patients. Therefore, research targeting new therapeutic alternatives with fewer adverse effects can significantly improve the pharmacological management of pain disorders [8].
In this context, medicinal plants are an ancient source of therapeutic resources in treating various diseases [9]. The pharmacological properties associated with the traditional use of medicinal species are due to the presence of bioactive compounds, indicating that medicinal plants are sources of new molecules with therapeutic potential [10] and, as such, may help develop new drugs to treat pain [11]. Historically, bark is most common ingredient has been utilized for centuries in traditional medicine for treatment of various diseases. This part of plant contains different active compounds, such as tannins, salicylates, flavonoids, and alkaloids, contributing to their analgesic effects [12,13].
Ximenia americana L. is popularly known in Brazil as wild plum, "ambuí," wild plum, thorn plum, and Brazilian sandalwood [14,15]. Previous research has demonstrated that X. americana has biological activities such as antioxidant [16], gastroprotective [17], antiparasitic [18], wound healing [19], antiedematogenic [20], and antinociceptive [21]. Phytochemical studies have identified caffeic acid (CA) as a major constituent in X. Americana [18,22,23]. Also known as 3,4-hydroxycinnamic acid, this compound is usually found in numerous fruits and vegetables, such as prunes, artichokes, eggplant, and grapes. [24] Research in pharmacology has uncovered a range of biological activities associated with CA, such as neuroprotective [25], antimicrobial [26], anticancer [27], anti-inflammatory [28], antinociception [29] and antiviral [30].
Considering the ethnopharmacological evidence of X. americana as a promising medicinal plant, this study aimed to investigate the antinociceptive effect of Ximenia americana L. Bark Extract (HEXA) and its main constituent, caffeic acid (CA), in animal models, to understand how these compounds can act in modulating pain induced by different stimuli.
2. Results
In our previous paper, we have demonstrated that the HPLC analysis revealed the predominance of polyphenols in the hydroethanolic bark extract of X. americana, where nine compounds were identified (Figure 1), including caffeic acid and quercetin as the major constituents.
The ESI-MS Analysis revealed the MS/MS pattern for caffeic acid dominating the loss of a water molecule (18 Da), characterizing the fragment of m/z 161.05 ([M-H2O]-) and sequentially the decarboxylation of CO2 (44 Da), which can be explained by the Diels-Alder rearrangement reaction allowing the visualization of the fragment of m/z 135.05 ([M-CO2]-) as identified by [31] (Figure 1A/1B). The Molecular Networking tool with gold classification was successfully applied to correlate the fragmentation spectrum of caffeic acid and other compounds of the extract of Ximenia americana L. (Table 1), with the GNPS spectral library, allowing the confirmation of its molecular identity.
2.1. HEXA Failed in Inducing Central Nervous System Effects in Mice
The treatment with HEXA (200 mg/kg) did not alter the animals' motor coordination compared to the negative control group in the rotarod test. In addition, the number of falls and the time spent on the bar of the treated animals was similar to the negative control group. Concerning the open field test, the treatment with HEXA (200 mg/kg) did not affect behavioral parameters such as rearing (lifting) and grooming (self-cleaning), nor alter the number of crossings. Together, these results suggest that HEXA has no evident central nervous system activity.
2.2. HEXA Has Antinociceptive Activity In Vivo: Central and Peripheral Effects
The administration of acetic acid induces abdominal writhing in untreated mice. treatments with HEXA at doses of 50, 100, and 200 mg/kg caused a significant reduction in the number of abdominal writhings by 84.99%, 87.99%, and 89.49% (p < 0.0001) respectively, compared to the untreated group. (Figure. 2A).
In the hot plate model, treatments with HEXA at doses of 50, 100, and 200 mg/kg significantly increased the time the animals remained on the hot plate by 69.17%, 68.13%, and 71.98%, (p < 0.0001) respectively when compared to the negative control group. A time-point analysis revealed that the lowest dose (50 mg/kg) increased the latency by 66.67%, 56.89%, 48%, and 67.89% at 30, 60, 120, and 180 min. (p < 0.0001). In these respective time points, the dose of 100 mg/kg increased the permanence by 68%, 50.62%, 47.32%, and 65.27%, while the dose of 200 mg/kg increased the permanence of the animals by 70.67 %, 58.12%, 50% and 72.63% (p < 0.0001) (Figure. 2B) (F (15, 120) = 2,951).
The treatments with HEXA at 50, 100, and 200 mg/kg significantly increased the pain threshold by 75.30%, 83.32%, and 74.52%, respectively, thus demonstrating the antinociceptive activity of HEXA in this model when compared to the negative control group. Regarding the time intervals, treatment with HEXA at a dose of 50 mg/kg increased the pain threshold by 42.83% (day 01), 58.83% (day 02), 69.01% (day 03), 54% (day 04), 99.39% (day 05), 90% (day 06), 51.44% (day 07), 62.43% (day 08), 84% (day 09), 77 .22% (day 10), 72.49% (day 11), 76.05% (day 12), 87.46% (day 13), 93.44% (day 14), 69.67% (day 15), 64.18% (day 16), 89.42% (day 17), 89.42% (day 18), 70.50% (day 19), 74.70% (day 20) and 89, 89% (day 21) (p < 0.0001) when compared to the negative control group (Figure 2C).
On the other hand, treatment with HEXA at a dose of 100 mg/kg increased the threshold by 44.41% (day 01), 57.95% (day 02), 69.17% (day 03), 89.24% (day 04), 92.85% (day 05), 89.64% (day 06), 99.80% (day 07), 74.90% (day 08), 89.82% (day 09), 86.30% (day 10), 87.02% (day 11), 86.57% (day 12), 99.99% (day 13), 74.75% (day 14), 77.68% ( day 15), 68.94% (day 16), 93.06% (day 17), 93.06% (day 18), 96.40% (day 19), 86.31% (day 20) and 84 .37% (day 21) (p < 0.0001) respectively, when compared to the negative control group (Figure 2C).
Finally, treatment with HEXA at a dose of 200 mg/kg increased the threshold by 45.70% (day 01), 45.92% (day 02), 53.12% (day 03), 87.85% ( day 04), 97.11% (day 05), 87.09% (day 06), 93.83% (day 07), 64.70% (day 08), 71.28% (day 09), 77 .22% (day 10), 61.66% (day 11), 66.50% (day 12), 99.98% (day 13), 70.66% (day 14), 60.81% (day 15), 96.24% (day 16), 83.64% (day 17), 83.64% (day 18), 99.98% (day 19), 68.62% (day 20), 89, 15% (day 21), (p < 0.0001) respectively when compared to the negative control group (Figure 2C).
2.3. Formalin-Induced Nociception Test of HEXA and CA
Animals treated with HEXA showed a significant reduction in paw licking time in the first phase and the second phase compared to the negative control group. In the first phase (Figure 3A), treatment with doses of 50, 100, and 200 mg/kg caused a reduction of 77.63%, 59.37%, and 58.44% (p < 0.001, F=0.114), respectively, while in the second phase (Figure. 3B), the same doses caused reductions of 56.74%, 69.43 and 54.02% (p < 0.0001, F=15.08), respectively, compared to the control group. On the other hand, treatment with CA at doses of 0.8 and 1.8 mg/kg reduced paw licking time by 36.40% and 22.17% (p < 0.0001, F=46.26), respectively, in the first phase (Figure. 3C). However, only the 1.8 mg/kg dose reduced licking time significantly 47.97% (p < 0.0001, F=23.55) in the second phase (Figure 3D) when compared to the control group.
2.4. Analysis of Signaling Pathways Underlying the Analgesic Effect of HEXA and CA
2.4.1. Vanilloid System
The administration of HEXA (100 mg/kg), CA (1.8 mg/kg), and ruthenium red (a non-selective antagonist of transient potential receptors -TRP) significantly reduced paw licking time 39.84%, 34.42%, and 99.77% (p < 0.0001, F=131.5) respectively of animals challenged with capsaicin, a TRPV1 receptor agonist, when compared to the negative control group (Figure 4A). However, while the antagonist fully inhibited the nociceptive response, the natural products caused partial inhibition.
2.4.2. Glutamatergic Pathway
The treatment with HEXA (100 mg/kg), CA (1.8 mg/kg), and ascorbic acid reduced the licking time by 33.66%, 53.60%, and 68.44% (p < 0.0001, F=12.71) respectively, after intraplantar injection of glutamate, compared to challenged and non-treated animals. These results demonstrate the potential participation of the glutamatergic pathway in the antinociceptive effect of the compounds (Figure 4B).
2.4.3. Opioid Pathway
The administration of HEXA (100 mg/kg), CA (1.8 mg/kg), and morphine significantly reduced nociception time by 57.28%, 51.21%, and 94.20 %, (p < 0.0001, F=26.36) respectively when compared to the negative control group (Figure. 4C). Notably, the magnitude of inhibition was significantly higher in the morphine group. When the animals received pretreatment with naloxone, the antinociceptive effect of HEXA and CA was reversed, suggesting that both treatments could modulate nociception by interfering with the opioid system.
2.4.4. L-arginine/Nitric Oxide/cGMP Pathway
Swiss mice treated with HEXA (100 mg/kg), CA (1.8 mg/kg), and L-NOARG had significantly reduced nociception time 57.77%, 52.99%, and 51.68%, (p < 0.0001, F=36.14) respectively in comparison with the negative control group. The pretreatment with L-arginine was found to reverse the analgesic effect of HEXA and CA, suggesting an interference of HEXA and CA on the NO signaling pathway (Figure 4D).
2.4.5. Cyclic Guanosine Monophosphate (cGMP) Pathway
The treatment with HEXA (100 mg/kg), CA (1.8 mg/kg), and methylene blue reduced the animals' licking time by 56.11%, 38.98%, and 98.8% (p < 0.0001, F=17.14) respectively, when compared to untreated animals. However, pretreatment with methylene blue did not affect the analgesic effect of HEXA and CA (Figure 4E). Nevertheless, the participation of this pathway in the mechanisms underlying the antinociceptive effect of the compounds cannot be ruled out.
2.4.6. Involvement of the α2-Adrenergic Receptor
The groups of mice treated with HEXA (100 mg/kg), CA (1.8 mg/kg), and clonidine (α2 receptor agonist) had significantly reduced nociception time 52.99%, 53.27%, and 92.99 %, (p < 0.0001, F=54.83) respectively in comparison with the negative control group. The pretreatment with yohimbine (α2 receptor antagonist) was found to reverse the analgesic effect of HEXA and CA, suggesting that these products interfere with α2-adrenergic receptor signaling (Figure 4F).
2.4.7. K+ATP Channel-Dependent Signaling
When treated with HEXA at a dose of 100 mg/kg and CA at a dose of 1.8 mg/kg, the animals showed a decrease in paw licking time by 38.70% and 36.41% (p < 0.0001, F=10.04) respectively, compared to the control group. In the HEXA group, when the animals received pretreatment with glibenclamide, the pain-relieving effect was reversed. However, this effect was not observed in the combined treatment with glibenclamide and CA (see Figure 4G) sed, suggesting that HEXA components could partially act on K+ATP channels. However, this effect was not observed in the combined treatment with glibenclamide and CA (Figure 4G).
2.4.8. Cholinergic Pathway
The treatment with HEXA (100 mg/kg), CA (1.8 mg/kg), and acetylcholine reduced nociception time by 53.33%, 39.55%, and 62.67% (p < 0.0001, F=17.15) respectively, when compared to the control group. When the animals in the HEXA and CA groups received pretreatment with atropine, the antinociceptive effect was not affected, indicating that there is no clear relationship between this mechanism and the analgesic effect of the natural products investigated in this study (Figure 5A).
Figure 2.
Participation of (A) cholinergic, (B) adenosinergic, (C) dopaminergic pathway and (D) serotonergic system for the antinociceptive response of HEXA (100 mg/kg) and CA (1.8 mg/kg) against formalin-induced pain in mice.
Figure 2.
Participation of (A) cholinergic, (B) adenosinergic, (C) dopaminergic pathway and (D) serotonergic system for the antinociceptive response of HEXA (100 mg/kg) and CA (1.8 mg/kg) against formalin-induced pain in mice.
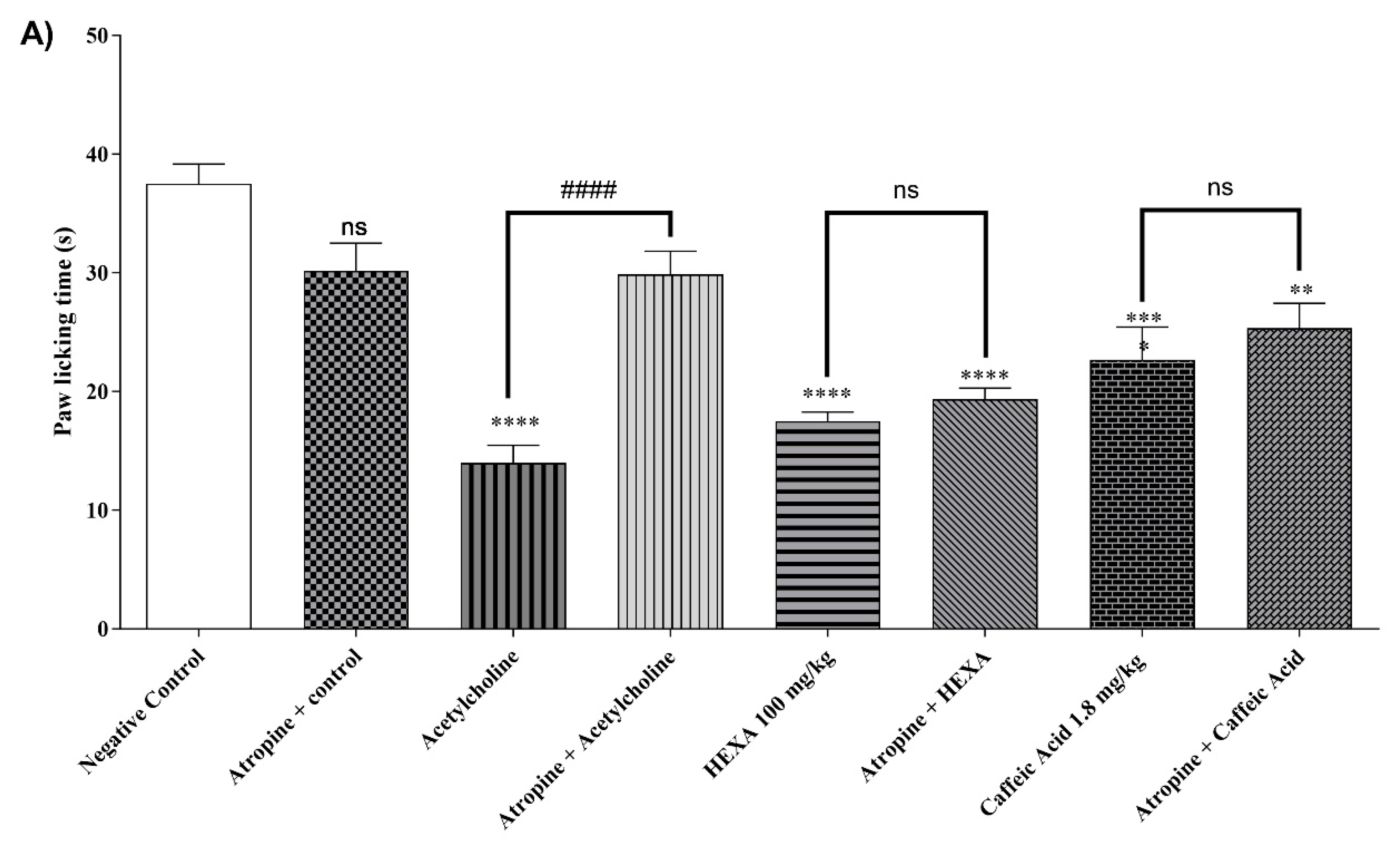
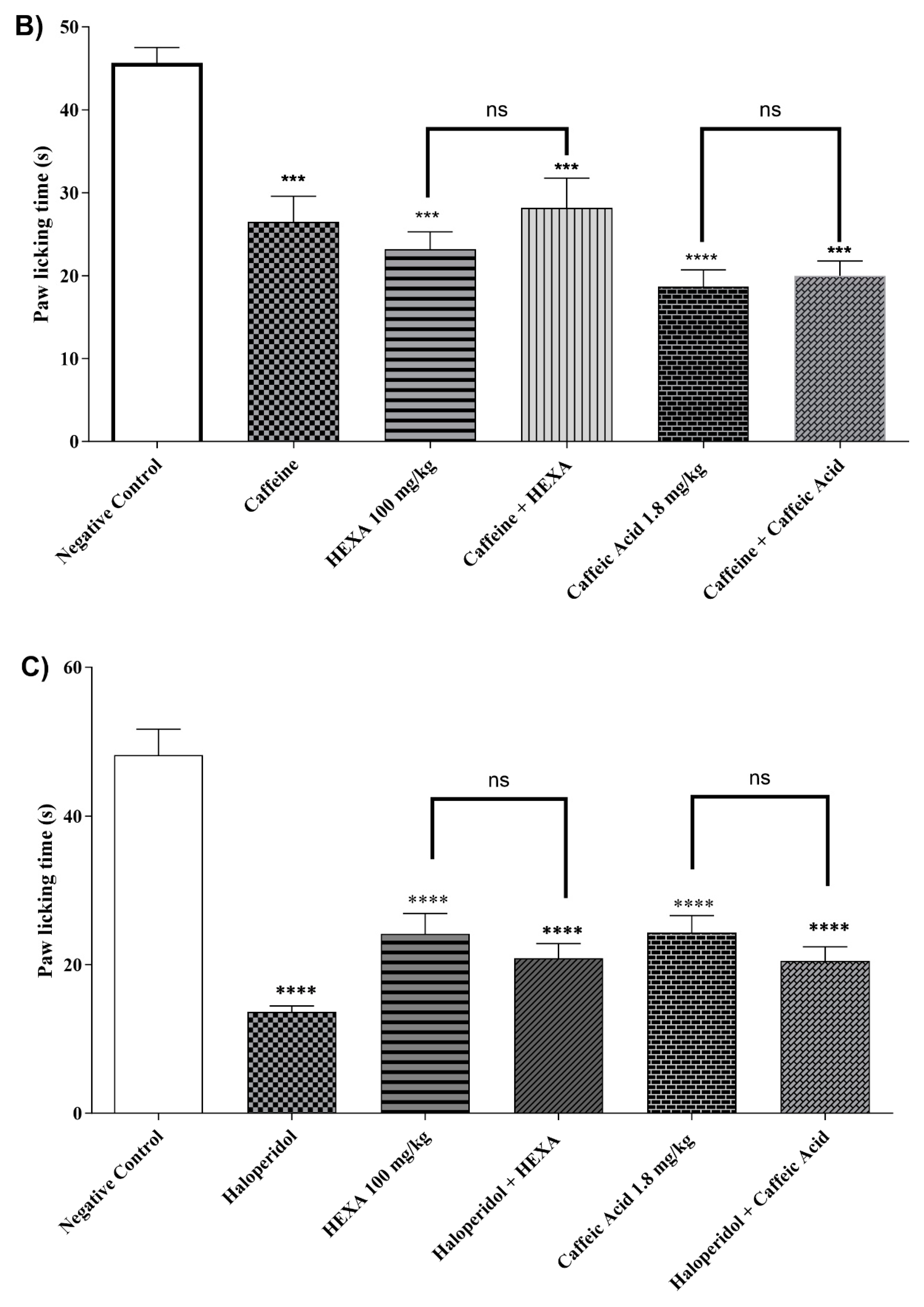
2.4.9. Adenosinergic Pathway
The treatment with HEXA (100 mg/kg) and CA (1.8 mg/kg) significantly reduced the animals' licking time by 49.27% and 59.12% (p < 0.0001, F=15.37) respectively, after intraplantar formalin injection compared to the control group. However, there was no statistical difference when the animals in the HEXA and CA groups received pretreatment with caffeine, a non-specific inhibitor of the pathway. (see Figure 5B).
2.4.10. Dopaminergic System
The treatment with HEXA (100 mg/kg) and CA (1.8 mg/kg) significantly reduced by 41.87% and 49.49%, (p < 0.0001, F= 23.70) respectively, the animals' licking time after when compared to the negative control group. However, no statistical difference was observed when the animals of the HEXA and CA groups received pretreatment with haloperidol, thus suggesting that none of these substances inhibit the adenosinergic signaling pathway (Figure 5C).
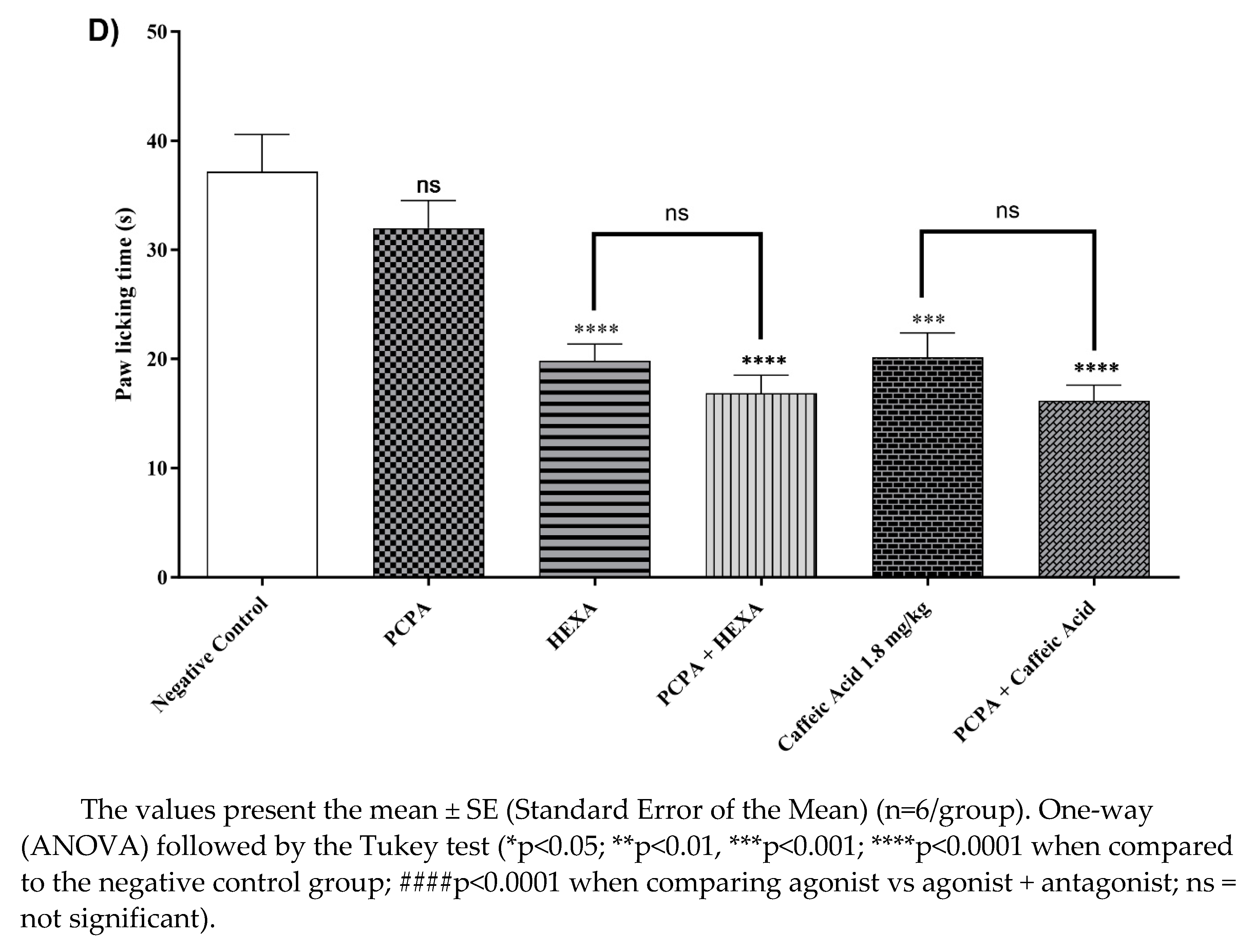
2.4.11. Involvement of the Serotonergic System
After receiving HEXA (100 mg/kg) and CA (1.8 mg/kg) treatments, the animals showed a 46.65% and 45.73% (p < 0.0001, F=15.33) reduction in licking time, compared to the control group. Additionally, when the HEXA and CA groups were given a pre-treatment of PCPA, the antinociceptive effect was sustained, indicating that both HEXA and CA do not affect the serotonergic signaling pathway (Figure 5D).
3. Discussion
This study contributed to new insights into the potential mechanisms of action underlying the antinociceptive effect of Ximenia americana L. bark extract (HEXA) and its major constituent caffeic acid (CA). In this context, significant reductions in pain responses were demonstrated through different assessment tests such as formalin, acetic acid, hot plate, and Complete Freund's Adjuvant, as well as through the pharmacological modulation of pain signaling pathways that to date have not been previously reported in the literature.
In our previously published data, our research group demonstrated that the extract of Ximenia americana L. used in this study (HEXA) has a unique chemical profile. It is characterized by the presence of alkaloids, steroids, saponins, glycosides, cyanogenic glycosides, anthraquinone, and, principally, flavonoids, terpenoids, and tannin derivatives The quantitative analysis by HPLC revealed rutin, gallic acid, quercetin, catechin, kaempferol, chlorogenic acid, ellagic acid, caffeic acid, and quercitrin as the main compounds [18,22,23,32]. The literature's comprehensive chemical profile, combined with the isolation of several other compounds from this species, including sambunigrin, gallic acid, catechin, and different stereoisomers of (epi)catechin, gallotannins quercetin, quercetin-3-O-β-xylopyranoside, quercetin-3-O-(6″-galloyl)-β-glucopyranoside quercitrin, avicularin, β-glucogalin, 1,6-digalloyl-β-glucopyranose, and kaempferol-3-O-(6″-galloyl)-β-glucopyranoside [16,33,34,35,36,37] underscores its potential for drug discovery, providing a solid foundation for further research in pharmacology.
Previous research by Coutinho et al, (2009) has demonstrated that quercitrin presents anti-inflammatory effects; epicatechin, isolated from ethanolic extracts of X. americana bark [39], and flavonoid-rich fractions obtained from the roots of this species showed antinociceptive activity and antioxidant potential [40]. In addition, caffeic acid has previously been shown to have biological activities such as antimicrobial[26], anticancer [41], muscle relaxant [42], antioxidant [43], gastroprotective, and antinociceptive. Corroborating with our results, the literature shows evidence that this antinociceptive effect is dose-dependent and may be mediated by nitric oxide[44] through the downregulation of NF-κB, α2-adrenergic and opioidergic receptors [29], but does not involve serotonergic pathways.
Thus, the antinociceptive activity of the HEXA can be justified by the presence of bioactive compounds with proven pharmacological properties. Our studies have demonstrated the therapeutic potential of various formulations obtained from different parts of X. americana. For instance, an extract from the stem of this species showed topical anti-inflammatory potential in ear edema models [22], while the ethanol extract showed anticonvulsant activity in models of acute inflammation [45]. Moreover, total polysaccharides obtained from X. americana showed anti-inflammatory and antinociceptive action in a model of acute pancreatitis. It was demonstrated that their pharmacological actions might occur through type 2 cannabinoid receptors [46].
Through the open field and rota rod tests, it was demonstrated that the HEXA, at the highest dose evaluated in the present study, did not promote central nervous system activity, suggesting that the antinociceptive effects observed in this study do not occur due to a change in behavioral parameters such as incoordination, stimulant or depressant, evaluated in the rota rod and open field models. The study found that HEXA, when orally administered at doses of 50, 100, and 200 mg/kg, demonstrated an antinociceptive effect by reducing nociception in mice models challenged with acetic acid and formalin, as evidenced by decreased abdominal writhing and paw licking. The results of the study showed that HEXA significantly reduced the number of writhing, indicating a possible peripheral analgesic effect by inhibiting the release of these mediators.
Regarding the formalin test, treatment with HEXA and CA significantly reduced nociception in both the first and second phases of the test, suggesting a possible inhibition of cell activation and mediator release. Zhen et al. (2015) demonstrated that an extract obtained from X. caffra, chemically characterized by the presence of phenolic compounds also identified in X. americana, including gallic acid, catechin, quercetin, and kaempferol and its derivatives, showed antioxidant, antiproliferative, and anti-inflammatory potential, reinforcing the potential action of the extract on the inflammatory component of the nociceptive response. In the hot plate test, the HEXA significantly increased the time the animals remained on the plate, which suggests the intriguing possibility that the extract may also act at a supraspinal level.
The Complete Freund's Adjuvant (CFA) is commonly used in chronic in vivo models. It triggers an inflammatory process that lowers nociceptive thresholds, leading to the development of allodynia and hyperalgesia in inflamed limbs. This compound induces an inflammatory response involving mediators such as histamine, serotonin, cytokines, glutamate, and prostaglandins [48,49], resulting in edema and persistent mechanical hyperalgesia in chronic models [50], as well as causing long-lasting mechanical allodynia in mice [51]. The latest findings demonstrate that HEXA significantly reduces allodynia and hyperalgesia, supporting the idea that this extract might inhibit the release of nociceptive process mediators, as previously suggested in this study. A study by Gamato et. al (2011) revealed that caffeic acid was more potent than its derivative rosmarinic acid in nociception models, with the 10 mg/kg dose showing the most significant antinociceptive potential. [52]
The extract of X. americana stem significantly reduced the chronic inflammatory process in a tendinitis model. [53]. The study conducted by Da Palma et al (2020) demonstrated that hydroalcoholic extracts obtained from the leaves and bark of X. americana presented wound-healing effects in acute, subchronic, and chronic stages of injury in mice. Thus, the therapeutic potential of this species in treating acute pain in both the acute and chronic phases is suggested. Given these results, the present study aims to investigate the potential signaling pathways underlying the analgesic effects of HEXA (hydroalcoholic extracts of X. americana) and its major constituent caffeic acid. For this purpose, we have chosen signaling pathways that are critically involved in the painful process, including the opioid, cholinergic, α2-adrenergic, adenosinergic, dopaminergic, glutamatergic, and vanilloid, serotoninergic pathways, as well as those mediated by cyclic guanosine monophosphate, nitric oxide, and K+ATP channels.
The opioid signaling pathway involves G protein-coupled receptors (Gi/0 subfamily), whose activation inhibits the enzyme adenylate cyclase, leading to the degradation of cyclic adenosine monophosphate (cAMP), blocking downstream events mediated by protein kinase A (PKA) [54]. Moreover, through this signaling pathway, opioid agonists open potassium channels and inhibit the opening of voltage-gated calcium channels [55], reducing the activity of nociceptor neurons and the release of neurotransmitters [54]. The results obtained in this study indicate that HEXA and CA can interfere with the opioid signaling pathway, which corroborates the results obtained in the hot plate assay.
Nitric oxide (NO) indirectly mediates the inflammatory process, contributing to vascular and cellular events. Since this mediator is produced from L-arginine under the action of NO-synthase (NOS), the inhibition of this synthesis pathway is associated with the inhibition of the pain process [56,57]. L-NOArg, an L-arginine analog, competitively inhibits NOS, blocking thus nitric oxide synthesis [58]. The present study showed that both HEXA and CA affect NO-mediated pain, corroborating the study by Kolgazi et al. (2021), who suggested that the gastroprotective potential of caffeic acid depends on the modulation of the nitric oxide pathway by regulation of NF-κB through NIK/IKK and c-Src/ERK signaling pathways [59].
The vanilloid receptor pathway or transient receptor potential vanilloid type 1 (TRPV1) is sensitive to a variety of harmful stimuli, such as temperatures elevated to ± 42ºC, lipid mediators, and vanilloid compounds such as capsaicin and resiniferatoxin[60,61]. The activation of these receptors results in neuron depolarization, leading to the release of multiple neuropeptides [62]. Ruthenium red, a non-selective antagonist of vanilloid receptors, is used to evaluate the potential interaction of substances with vanilloid receptors [63]. Glutamate induces a nociceptive response by activating glutamatergic receptors at supraspinal, spinal, and peripheral sites [64]. This neurotransmitter activates AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid) and NMDA (N-Methyl-D-Aspartic acid) receptors, thus allowing the passage of Na+ and Ca2+, facilitating the action potential and subsequent nociceptive responses [65].
Such inhibition also results in the availability of cGMP, providing peripheral antinociception through the opening of ATP-sensitive K+ channels [66]. On the other hand, the administration of methylene blue causes the activation of the guanylate cyclase enzyme, which catalyzes cGMP production [67]. In the present study, it was evidenced that both HEXA and CA act synergistically. The analgesic pathway mediated by α2 receptors, involves Gi-mediated inhibition of presynaptic Ca+2 channels, preventing glutamate (excitatory neurotransmitter) from being released from primary afferent fibers [68]. Simultaneously, the opening of K+ channels results in hyperpolarization, causing a decrease in the excitability of postsynaptic cells [69]. Agonists of these receptors have analgesic properties involving peripheral and central mechanisms [70]. In this study, the evaluation of HEXA in this pathway indicates that the antinociceptive effect of this extract can involve the participation of these receptors.
K+ATP channels are directly involved in cell hyperpolarization, consecutively decreasing intracellular concentrations of Ca2+ and reducing the release of neurotransmitters, thus inhibiting nociception. Experimentally, the administration of glibenclamide blocks these channels [71]. From the results obtained in the study, it is reasonable to hypothesize that the action of HEXA affects the activation of potassium channels, contributing to the observed analgesic effect.
The findings of this study indicate that HEXA exhibits antinociceptive effects both centrally and peripherally. These effects may be attributed to its ability to suppress the release of inflammatory mediators or regulate their interaction with receptors involved in pain perception. This research revealed that the components of the extract of X. americana can influence multiple pain pathways with the modulation of α2-adrenergic, nitric oxide, glutamatergic, vanilloid, cyclic guanosine monophosphate, and K+ATP channel-dependent pathways. On the other hand, caffeic acid specifically influences the opioid, glutamatergic, and vanilloid pathways by inhibiting nitric oxide and cyclic guanosine monophosphate.
4. Materials and Methods
4.1. Drugs, Reagents, and Doses
The substances used in this study were obtained from Sigma-Aldrich. The other substances used in the tests were the following: formalin (Pubchem CID = 712), morphine (5 mg/kg; s.c.) (Pubchem CID = 5288826), L-NOARG ((Pubchem CID= 440005), ruthenium red (Pubchem CID=117587625), capsaicin (Pubchem CID= 1548943), glutamate (Pubchem CID= 23689119), methylene blue (Pubchem CID= 6099), clonidine (Pubchem CID= 20179), yohimbine (Pubchem CID= 6169), glibenclamide (Pubchem CID= 3488), acetylcholine (Pubchem CID= 6060), atropine (Pubchem CID= 174174), caffeine (Pubchem CID= 2519), p-chlorophenyl alanine (Pubchem CID= 46520). Methanol, acetonitrile (LiChrosolv®, ≥99.9%), and ammonium hydroxide (NH4OH) were purchased from Sigma-Aldrich (St. Louis, U.S.A.).
HEXA was obtained as previously described by da Silva et al. (2018) [20] and administered at 50, 100, and 200 mg/kg orally. Caffeic acid (PubChem CID = 689043) was administered at 0.18 mg/kg and 1.8 mg/kg.
4.2. Chemical Profile by Electrospray Ionization-Mass Spectrometry (ESI-MS) Analysis
In our previous paper, Silva et al. (2018) determined the chemical profile of HEXA compounds, identifying 21 compounds using HPLC-DAD. However, in the current study, the analysis focuses specifically on the compound caffeic acid. For this purpose, Electrospray Ionization-Mass Spectrometry (ESI-MS) analysis was utilized.
The sample was prepared by dissolving the dried extract in 1 mL of a methanol: acetonitrile mixture (2:1, v:v) containing 10 μL of ammonium hydroxide. The ESI-MS analyses were performed on a high-resolution Quadrupole-Orbitrap hybrid mass spectrometer (Q-Exactive, Thermo Scientific). The E.S.I. source conditions were set as follows: negative ionization mode, spray voltage of 3.2 kV, capillary temperature of 320°C, S-Lens RF level at 45 V, sheath gas flow rate at 8 L.min-1, and auxiliary gas flow rate at 5 L.min-1, and for MSMS a collision energy of 15 eV was used.
4.2.1. Compound Annotation
The mass spectrometer data files were converted from (.raw) to (.mzML) format using MSconvert software (ProteoWizard, Palo Alto, CA, U.S.A.) and processed using the online platform (https://gnps.ucsd.edu/ProteoSAFe/static/gnps-splash.jsp) to retrieve compound information. Molecular annotations were based on the comparison of experimental MS/MS spectra with the GNPS spectral library using tools such as Molecular Networking [72].
4.3. Animals
Swiss mice (Mus musculus), both sexes, 20-30g, housed in polypropylene cages (410 x 282 x 150 mm; n=6), at 24 ± 2°C, 12-hour light/dark cycle, with free access to water and specific chow (Labina, Presence®). Mice were fasted from solids for 6-8 hours before experiments.
4.4. Ethical Information
The study adhered to guidelines from CONCEA, U.K. Animals (Scientific Procedures) Act, 1986, EU Directive 2010/63/EU, and NIH guidelines. Protocols were approved by the Ethics Committee on Animal Use of URCA (approval number 180/2020.2).
4.5. Evaluation of the effects of HEXA on the Central Nervous System
4.5.1. Rota Rod Performance Test
Mice (n=3/group) were treated with water (10 mL/kg; p.o.) or HEXA (200 mg/kg; p.o.). After 1 hour, the number of falls and time on the rotating rod were recorded. [73]
4.5.2. Open Field Test
Mice (n=3/group) were treated with water (10 mL/kg; p.o.) or HEXA (200 mg/kg; p.o.). After 1 hour, the number of fields explored in a 30 cm2 open field was observed for 5 minutes [74].
4.6. Screening for the Antinociceptive Effect
The antinociceptive effect was assessed using acetic acid-induced writhing, formalin, hot plate, and Von Frey tests. Mice were divided into groups (n=6) and treated with water (10 mL/kg; p.o.) or HEXA (50, 100, 200 mg/kg; p.o.)[75]. The experimental timeline is shown in Figure 1
4.6.1. Acetic Acid-Induced Abdominal Contortions
4.6.2. The Formalin Test
Mice (n=6/group) were treated with water (10 mL/kg; p.o.), HEXA (50, 100, 200 mg/kg; p.o.), or CA (0.18, 1.8 mg/kg). After 1 hour, 2.5% formalin (20 μL) was administered, and the nociceptive response was observed at 0-5 min and 15-30 min post-injection [77].
4.7. Evaluation of Central and Peripheral Antinociceptive Responses
4.7.1. Hot Plate Test
4.7.2. Mechanical Pressure Hypernociception (Von Frey Test)
Mice (n = 6/group) were subjected to mechanical stimulation in the right paw at 24 h and 30 min (basal measure) before the test. After 30 min, the animals received an injection (20 μl/paw) of complete Freund's adjuvant (CFA) (1 mg/ml), in the right hind paw. The animals were treated with water (10 mL/ kg; p.o.) or HEXA (50 or 100, or 200 mg/kg, p.o.). Subsequently, treatments were carried out on the 5th, 9th, 13th, 17th, and 21st days after induction. The test was performed using the Von Frey apparatus. Briefly, the animals were placed individually in glass boxes (12 x 20 x 20cm) supported on an elevated surface and covered with a wire mesh. After 1h of acclimatization, the stimulus was applied with a rod with a constant force on the plantar surface of the right hind paw (to assess hypernociception to the lesion in injured mice) using the Von Frey filament resistance apparatus for 21 consecutive days. The force in (g) was determined by removing the paw from the filament. The force was applied six times, and the average of the three most similar values was considered [79,80].
4.8. Investigation of the Signaling Pathways Associated with the Analgesic Effect of HEXA and AC
The effect of HEXA (100 mg/kg; b.w.), or CA (1.8 mg/kg; b.w.) on pain signaling pathways was evaluated using the formalin assay through the pharmacological signaling pathways including opioid receptors, nitric oxide, vanilloid, glutamatergic, cyclic guanosine monophosphate pathway, adrenergic α-2, K+ATP channels, cholinergic, adenosinergic, dopaminergic and the serotonergic pathway.
In the formalin model, mice (n = 6) were pretreated with HEXA (100 mg/kg; p.o.), CA (1.8 mg/kg; p.o.), or, according to the signaling pathway: Opioid - morphine (agonist - 5 mg/kg; s.c.) and naloxone (antagonist - 4 mg/kg; i.p.) [81]; Nitric Oxide - L-NOARG (nitric oxide synthase (NOS) inhibitor - 75 mg/kg; i.p.) and L-Arginine (NOS substrate - 600 mg/kg; i.p.) [82,83]. Vanilloid - Ruthenium red (non-selective TRP antagonist - 3 mg/kg; i.p.) and capsaicin (TRPV1 receptor agonist - 5.2 nmol/paw) [64]; Glutamatergic - Ascorbic acid (100 mg/kg; i.p.) and buffered glutamate (20 μmol/paw) (Lam et al., 2005; Majewska et al., 1990; cGMP - Methylene blue (guanylate cyclase inhibitor - 20 mg/kg; i.p.) [64]; α2-adrenergic - clonidine (agonist - 0.1 mg/kg; i.p.) and yohimbine (antagonist - 0.15 mg/kg; i.p.) [88]; K+ATP channels - glibenclamide (K+ATP channel blocker - 3 mg/kg; i.p.) [64,89]; Cholinergic - acetylcholine (agonist - 1 mg/kg; i.p.) and atropine (antagonist - 1 mg/kg; i.p.) [83]; Adenosinergic - caffeine (10 mg/kg; i.p.) [90]; Dopaminergic - haloperidol (non-selective dopamine receptor antagonist - 0.2 mg/kg; i.p.) [91,92]; and Serotoninergic - p-chlorophenyl alanine (PCPA) (serotonin receptor antagonist - 100 mg/kg; i.p.) [93,94].
4.9. Statistical Analysis
The values were expressed as mean ± SEM. Data were analyzed by One-way ANOVA followed by Tukey's test, or two-way ANOVA followed by Dunnett's or Tukey's posthoc, using the GraphPad Software version 6.1. p values < 0.05 were considered significant.
5. Conclusions
In conclusion, HEXA and CA have significant antinociceptive effects in the absence of evident SNC toxicity in mice and, as such, have the potential to be used in the development of new analgesic drugs. The present results brought new insights into the understanding of the antinociceptive effects of HEXA and CA, as well as the main mechanisms underlying their effects in pain management, shedding light on the search for new therapeutic alternatives.
Supplementary Materials
Not applicable.
Author Contributions
Conceptualization, Resources, RTP; Methodology, Resources, LYSS and ISA; Validation, formal analysis, TMS; Investigation, Data curation, ESS; Formal analysis, Visualization, RHSC; Investigation; Visualization, ABS; Writing - original draft; Supervision, JRF; Methodology , Writing - original draft, AOBPBM; Writing - review & editing, Visualization, HDM; Methodology, Validation, JCPS; Methodology, Investigation, ARC; Methodology, Validation, RNM; Project administration , Writing - review & editing IRAM; All authors have read and agreed to the published version of the manuscript.
Funding
Acknowledges financial support from CNPq-PQ (Grant 303438/2021-5) and FUNCAP-UNIVERSAL (Grant 404780/2021-0) and CNPq (Grant 406522/2021-9-1).
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Ethics Committee on Animal Use of Regional University of Cariri - URCA (approval number 180/2020.2).
Informed Consent Statement
Not applicable.
Acknowledgments
The authors would like to thank the financial support provided by Coordination for the Improvement of Higher Education Personnel - Brazil (CAPES), Cearense Foundation to Support Scientific and Technological Development (FUNCAP) - finance code BPI, National Council for Scientific and Technological Development (CNPq) – finance code 304291/2017-0, and Financier of Studies and Projects - Brasil (FINEP). This article is a contribution of the National Institute of Science and Technology - Ethnobiology, Bioprospecting and Nature Conservation/CNPq/FACEPE and Excitotoxicity and Neuroprotection (INCT-EN); National Institute of Science and Technology of the Health Economic-Industrial Complex -iCEIS and National Institute of Science and Technology in Analytical Chemistry of Food: Food Safety and Characterization of Functional Foods (INCT-ALIM) for the support offered to carry out this work.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper
References
- Baral, P.; Udit, S.; Chiu, I.M. Pain and Immunity: Implications for Host Defence. Nat Rev Immunol 2019, 19, 433–447. [CrossRef]
- Matsuda, M.; Huh, Y.; Ji, R.R. Roles of Inflammation, Neurogenic Inflammation, and Neuroinflammation in Pain. J Anesth 2019, 33, 131–139. [CrossRef]
- Arthur, J.; Bruera, E. Balancing Opioid Analgesia with the Risk of Nonmedical Opioid Use in Patients with Cancer. Nat Rev Clin Oncol 2019, 16, 213–226. [CrossRef]
- Cohen, S.P.; Vase, L.; Hooten, W.M. Chronic Pain: An Update on Burden, Best Practices, and New Advances. The Lancet 2021, 397, 2082–2097. [CrossRef]
- Day, R.O.; Graham, G.G. Non-Steroidal Anti-Inflammatory Drugs (NSAIDs). BMJ 2013, 346, 1396.2-1396. [CrossRef]
- Lima, C.P.; Silva, H.R.D.O.; Pogian, V.B.; Santos, V.G. Pharmaceutical Evaluation of the Risks of the Use of Non-Steroid Anti-Inflammatories. Unisanta Health Science 2020, 4, 1–20.
- Silva, L.A.; Gerica, O, Al, H. Review on Non-Steroid Antiinflammatory: Acetylsalicylic Acid. Rev Inic Cient e Ext 2018, 1, 169–174.
- Mohammad, F.; Hasan, E.; Wedad, A.; Mohamed, E.G. Natural Antioxidant Flavonoids in Formalin-Induced Mice Paw Inflammation; Inhibition of Mitochondrial Sorbitol Dehydrogenase Activity. J Biochem Mol Toxicol 2017, 31, 1–8. [CrossRef]
- Mussin, J.; Giusiano, G. Ethno–Phytopharmacology: Product Validation Process Based on Traditional Knowledge of Medicinal Plants. In Agricultural, Forestry and Bioindustry Biotechnology and Biodiscovery; Springer, 2020; pp. 331–353.
- Ribeiro, D.A.; Oliveira, L.G.S. de; Macêdo, D.G. de; Menezes, I.R.A. de; Costa, J.G.M. da; Silva, M.A.P. da; Lacerda, S.R.; Souza, M.M. de A. Promising Medicinal Plants for Bioprospection in a Cerrado Area of Chapada Do Araripe, Northeastern Brazil. J Ethnopharmacol 2014, 155, 1522–1533. [CrossRef]
- Sreekeesoon, D.P.; Mahomoodally, M.F. Ethnopharmacological Analysis of Medicinal Plants and Animals Used in the Treatment and Management of Pain in Mauritius. J Ethnopharmacol 2014, 157, 181–200. [CrossRef]
- Da Silva-Leite, K.E.S.; Assreuy, A.M.S.; Mendonça, L.F.; Damasceno, L.E.A.; De Queiroz, M.G.R.; Mourão, P.A.S.; Pires, A.F.; Pereira, M.G. Polysaccharide Rich Fractions from Barks of Ximenia Americana Inhibit Peripheral Inflammatory Nociception in Mice Antinociceptive Effect of Ximenia Americana Polysaccharide Rich Fractions. Revista Brasileira de Farmacognosia 2017, 27, 339–345. [CrossRef]
- Jahromi, B.; Pirvulescu, I.; Candido, K.D.; Knezevic, N.N. Herbal Medicine for Pain Management: Efficacy and Drug Interactions. Pharmaceutics 2021, 13, 251. [CrossRef]
- Da Silva, G.G.; De Souza, P.A.; De Morais, P.L.D.; Dos Santos, E.C.; Moura, R.D.; Menezes, J.B. Caracterização Do Fruto de Ameixa Silvestre (Ximenia Americana L.). Rev Bras Frutic 2008, 30, 311–314. [CrossRef]
- Shettar, A.K.; Sateesh, M.K.; Kaliwal, B.B.; Vedamurthy, A.B. In Vitro Antidiabetic Activities and GC-MS Phytochemical Analysis of Ximenia Americana Extracts. South African Journal of Botany 2017, 111, 202–211. [CrossRef]
- Almeida, M.L.B.; Freitas, W.E.D.S.; Morais, P.L.D. De; Sarmento, J.D.A.; Alves, R.E. Bioactive Compounds and Antioxidant Potential Fruit of Ximenia Americana L. Food Chem 2016, 192, 1078–1082. [CrossRef]
- Aragão, T.P.; dos Prazeres, L.D.K.T.; Brito, S.A.; Neto, P.J.R.; Rolim, L.A.; da Silva Almeida, J.R.G.; Caldas, G.F.R.; Wanderley, A.G. alves Contribution of Secondary Metabolites to the Gastroprotective Effect of Aqueous Extract of Ximenia Americana L. (Olacaceae) Stem Bark in Rats. Molecules 2018, 23, 1–18. [CrossRef]
- de Menezes, I.R.A.; da Costa, R.H.S.; Augusti Boligon, A.; Rolón, M.; Coronel, C.; Vega, C.; Melo Coutinho, H.D.; da Costa, M.S.; Tintino, S.R.; Silva Pereira, R.L.; et al. Ximenia Americana L. Enhances the Antibiotic Activity and Inhibit the Development of Kinetoplastid Parasites. Comp Immunol Microbiol Infect Dis 2019, 64, 40–46. [CrossRef]
- Da Palma, A.F.M.; Marques, L.K.M.; Carneiro, R.D.S.; Carvalho, G.F.S.; Ferreira, D.C.L.; Sant’Ana, A.E.G.; Maia Filho, A.L.M.; Marques, R.B.; Alves, W.D.S.; Uchôa, V.T.; et al. Evaluation of Hydroalcoholic Extracts of Stem and Leaves of Ximenia Americana L. In the Healing of Excisional Acute Wounds in Mice. Revista Virtual de Quimica 2020, 12, 37–50. [CrossRef]
- da Silva, B.A.F.; da Costa, R.H.S.; Fernandes, C.N.; Leite, L.H.I.; Ribeiro-Filho, J.; Garcia, T.R.; Coutinho, H.D.M.; Wanderley, A.G.; de Menezes, I.R.A. HPLC Profile and Antiedematogenic Activity of Ximenia Americana L. (Olacaceae) in Mice Models of Skin Inflammation. Food and Chemical Toxicology 2018, 119, 199–205. [CrossRef]
- Soro, T.Y.; Zahoui, O.S.; Nene-bi, A.S. Analgesic Activity of the Fractions of the Aqueous Extract of Ximenia Americana (Linné) (Olacaceae). International Journal of Pharmacology and Toxicology 2016, 4, 1. [CrossRef]
- da Silva, B.A.F.; da Costa, R.H.S.; Fernandes, C.N.; Leite, L.H.I.; Ribeiro-Filho, J.; Garcia, T.R.; Coutinho, H.D.M.; Wanderley, A.G.; de Menezes, I.R.A. HPLC Profile and Antiedematogenic Activity of Ximenia Americana L. (Olacaceae) in Mice Models of Skin Inflammation. Food and Chemical Toxicology 2018, 119, 199–205. [CrossRef]
- da Costa, R.H.S.; Martins, A.O.B.P.B.; de Oliveira, M.R.C.; Alcântara, I.S.; Ferreira, F.F.; dos Santos, F.F.C.; de Araújo Delmondes, G.; da Cunha, F.A.B.; Coutinho, H.D.M.; de Menezes, I.R.A. Acaricide Activity of the Ximenia Americana L. (Olacaceae) Stem Bark Hydroethanolic Extract against Rhipicephalus (Boophilus) Microplus. Biologia (Bratisl) 2022, 77, 1667–1674. [CrossRef]
- Cizmarova, B.; Hubkova, B.; Bolerazska, B.; Marekova, M.; Birkova, A. Caffeic Acid: A Brief Overview of Its Presence, Metabolism, and Bioactivity. Bioact Compd Health Dis 2020, 3, 74–81. [CrossRef]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, M.I.; Pasupuleti, V.R. Therapeutic Implications of Caffeic Acid in Cancer and Neurological Diseases. Front Oncol 2022, 12. [CrossRef]
- Khan, F.; Bamunuarachchi, N.I.; Tabassum, N.; Kim, Y.M. Caffeic Acid and Its Derivatives: Antimicrobial Drugs toward Microbial Pathogens. J Agric Food Chem 2021, 69, 2979–3004.
- Nasr Bouzaiene, N.; Kilani Jaziri, S.; Kovacic, H.; Chekir-Ghedira, L.; Ghedira, K.; Luis, J. The Effects of Caffeic, Coumaric and Ferulic Acids on Proliferation, Superoxide Production, Adhesion and Migration of Human Tumor Cells in Vitro. Eur J Pharmacol 2015, 766, 99–105. [CrossRef]
- Zielińska, D.; Zieliński, H.; Laparra-Llopis, J.M.; Szawara-Nowak, D.; Honke, J.; Giménez-Bastida, J.A. Caffeic Acid Modulates Processes Associated with Intestinal Inflammation. Nutrients 2021, Vol. 13, Page 554 2021, 13, 554. [CrossRef]
- Park, S.H.; Sim, Y.B.; Kim, S.M.; Lee, J.K.; Jung, J.S.; Suh, H.W. The Effect of Caffeic Acid on the Antinociception and Mechanisms in Mouse. J Appl Biol Chem 2011, 54, 177–182. [CrossRef]
- Crevar-Sakač, M.; Vujić, Z.; Kotur-Stevuljević, J.; Ivanišević, J.; Jelić-Ivanović, Z.; Milenković, M.; Markelić, M.; Vujčić, Z. Uticaj Atorvastatina i Tinkture Lista Artičoke Na Oksidativni Stres Kod Pacova Sa Hiperholesterolemijom. Vojnosanit Pregl 2016, 73, 178–187. [CrossRef]
- Wu, Z.-J.; Ma, X.-L.; Fang, D.-M.; Qi, H.-Y.; Ren, W.-J.; Zhang, G.-L. Analysis of Caffeic Acid Derivatives from Osmanthus Yunnanensis Using Electrospray Ionization Quadrupole Time-of-Flight Mass Spectrometry. European Journal of Mass Spectrometry 2009, 15, 415–429. [CrossRef]
- Brandão, D.O.; Fernandes, F.H.A.; Júnior, F.J.L.R.; Silva, P.C.D.; Santana, C.P.; De Medeiros, F.D.; Véras, G.; Medeiros, A.C.D. Validation of UPLC Method for Determination of Gallic Acid from Ximenia Americana L. Planta Med 2014, 80, P2O60.
- de Araújo, M.R.S.; da Costa Assunção, J.C.; Dantas, I.N.F.; Costa-Lotufo, L.V.; Monte, F.J.Q. Chemical Constituents of Ximenia Americana. Nat Prod Commun 2008, 3, 1934578X0800300605.
- Le, N.H.T.; Malterud, K.E.; Diallo, D.; Paulsen, B.S.; Nergård, C.S.; Wangensteen, H. Bioactive Polyphenols in Ximenia Americana and the Traditional Use among Malian Healers. J Ethnopharmacol 2012, 139, 858–862. [CrossRef]
- Queiroz Monte, F.J.; de Lemos, T.L.G.; de Arajo, M.R.S.; Sousa Gomes, E. de Ximenia Americana: Chemistry, Pharmacology and Biological Properties, a Review. Phytochemicals - A Global Perspective of Their Role in Nutrition and Health 2012. [CrossRef]
- Siddaiah, M.; Jayavcera, K.N.; Mallikarjuna, R.P.; Ravindra, R.K.; Yasodha, K.Y.; Narender, R.G. Phytochemical Screening and Analgesic Activity of Methanolic Extract of Ximenia Americana. Journal of Pharmacy and Chemistry 2009, 3, 23–25.
- Hemamalini, K.; Srikanth, A.; Sunny, G.; Praneethkumar, H. Phytochemical Screening and Analgesic Activity of Methanolic Extract of Ximenia Americana. Journal of Current Pharma Research 2011, 1, 153.
- Coutinho, M.A.S.; Muzitano, M.F.; Costa, S.S. Flavonoids: Potential Therapeutic Agents for the Inflammatory Process. Revista Virtual de Química 2009, 1, 241–256. [CrossRef]
- Dias, T.L.M.F.; Melo, G.M.A.; Da Silva, Y.K.C.; Queiroz, A.C.; Goulart, H.F.; Alexandre-Moreira, M.S.; Santana, A.E.G.; Uchôa, V.T. Antinociceptive and Anti-Inflammatory Activities of the Ethanolic Extract, of Fractions and of Epicatechin Isolated from the Stem Bark of Ximenia Americana L. (Oleacaceae). Revista Virtual de Quimica 2018, 10, 86–101. [CrossRef]
- Kiessoun, K.; Ouattara, N.; Arsène, M.; Souza, A.; Sytar, O.; Brestic, M.; Mamoudou, D.H. Anti-Nociceptive and Anti-Inflammatory Properties of Polyphenol-Rich Fractions of Roots from Ximenia Americana L ., ( Olacaceae ), in Experimental Mice. Int J Pharm Pharm Res 2018, 12, 281–297.
- Matejczyk, M.; Świsłocka, R.; Golonko, A.; Lewandowski, W.; Hawrylik, E. Cytotoxic, Genotoxic and Antimicrobial Activity of Caffeic and Rosmarinic Acids and Their Lithium, Sodium and Potassium Salts as Potential Anticancer Compounds. Adv Med Sci 2018, 63, 14–21. [CrossRef]
- de Alencar Silva, A.; Pereira-de-Morais, L.; Rodrigues da Silva, R.E.; de Menezes Dantas, D.; Brito Milfont, C.G.; Gomes, M.F.; Araújo, I.M.; Kerntopf, M.R.; Alencar de Menezes, I.R.; Barbosa, R. Pharmacological Screening of the Phenolic Compound Caffeic Acid Using Rat Aorta, Uterus and Ileum Smooth Muscle. Chem Biol Interact 2020, 332. [CrossRef]
- Amorati, R.; Pedulli, G.F.; Cabrini, L.; Zambonin, L.; Landi, L. Solvent and PH Effects on the Antioxidant Activity of Caffeic and Other Phenolic Acids. J Agric Food Chem 2006, 54, 2932–2937. [CrossRef]
- Kolgazi, M.; Cilingir, S.; Yilmaz, O.; Gemici, M.; Yazar, H.; Ozer, S.; Acikel-Elmas, M.; Arbak, S.; Suyen, G.G. Caffeic Acid Attenuates Gastric Mucosal Damage Induced by Ethanol in Rats via Nitric Oxide Modulation. Chem Biol Interact 2021, 334, 109351. [CrossRef]
- Quintans-Júnior, L.J.; De Almeida, R.N.; Falcão, A.C.G.M.; Agra, M. de F.; De Sousa, M. de F.V.; Barbosa-Filho, J.M. Avaliação Da Atividade Anticonvulsivante de Plantas Do Nordeste Brasileiro. Acta Farmaceutica Bonaerense 2002, 21, 179–184.
- da Silva-Leite, K.E.S.; Girão, D.K.F.B.; de Freitas Pires, A.; Assreuy, A.M.S.; de Moraes, P.A.F.; Cunha, A.P.; Ricardo, N.M.P.S.; Criddle, D.N.; de Souza, M.H.L.P.; Pereira, M.G.; et al. Ximenia Americana Heteropolysaccharides Ameliorate Inflammation and Visceral Hypernociception in Murine Caerulein-Induced Acute Pancreatitis: Involvement of CB2 Receptors. Biomedicine and Pharmacotherapy 2018, 106, 1317–1324. [CrossRef]
- Zhen, J.; Guo, Y.; Villani, T.; Carr, S.; Brendler, T.; Mumbengegwi, D.R.; Kong, A.N.T.; Simon, J.E.; Wu, Q. Phytochemical Analysis and Anti-Inflammatory Activity of the Extracts of the African Medicinal Plant Ximenia Caffra. J Anal Methods Chem 2015, 2015. [CrossRef]
- Ferreira, J.; Campos, M.M.; Pesquero, J.B.; Araújo, R.C.; Bader, M.; Calixto, J.B. Evidence for the Participation of Kinins in Freund’s Adjuvant-Induced Inflammatory and Nociceptive Responses in Kinin B1 and B2 Receptor Knockout Mice. Neuropharmacology 2001, 41, 1006–1012. [CrossRef]
- Billiau, A.; Matthys, P. Modes of Action of Freund’s Adjuvants in Experimental Models of Autoimmune Diseases. J Leukoc Biol 2001, 70, 849–860. [CrossRef]
- Dutra, R.C.; Campos, M.M.; Santos, A.R.S.; Calixto, J.B. Medicinal Plants in Brazil: Pharmacological Studies, Drug Discovery, Challenges and Perspectives. Pharmacol Res 2016, 112, 4–29. [CrossRef]
- Sandkühler, J. Models and Mechanisms of Hyperalgesia and Allodynia. Physiol Rev 2009, 89, 707–758. [CrossRef]
- Gamaro, G.D.; Suyenaga, E.; Borsoi, M.; Lermen, J.; Pereira, P.; Ardenghi, P. Effect of Rosmarinic and Caffeic Acids on Inflammatory and Nociception Process in Rats. ISRN Pharmacol 2011, 2011, 1–6. [CrossRef]
- Leal, S.S.; Uchôa, V.T.; Figuerêdo-Silva, J.; Soares, R.B.; Mota, D.M.; De Alencar, R.C.; Filho, A.L.M.M.; Sant’Ana, A.E.G.; Beltrame Junior, M. Eficácia Da Fonoforese Com Ximenia Americana L. Na Inflamação de Tendão de Ratos. Revista Brasileira de Medicina do Esporte 2016, 22, 355–360. [CrossRef]
- Dhawan, B.N.; Cesselin, F.; Raghubir, R.; Reisine, T.; Bradley, P.B.; Portoghese, P.S.; Hamon, M. International Union of Pharmacology. XII. Classification of Opioid Receptors. Pharmacol Rev 1996, 48, 567–592.
- Stein, C.; Hassan, A.H.S.; Lehrberger, K.; Stein, C.; Giefing, J.; Yassouridis, A. Local Analgesic Effect of Endogenous Opioid Peptides. The Lancet 1993, 342, 321–324. [CrossRef]
- Jin, W.Y.; Liu, Z.; Liu, D.; Yu, L.C. Antinociceptive Effects of Galanin in the Central Nucleus of Amygdala of Rats, an Involvement of Opioid Receptors. Brain Res 2010, 1320, 16–21. [CrossRef]
- Shih, C.C.; Hwang, H.R.; Chang, C.I.; Su, H.M.; Chen, P.C.; Kuo, H.M.; Li, P.J.; Wang, H.M.D.; Tsui, K.H.; Lin, Y.C.; et al. Anti-Inflammatory and Antinociceptive Effects of Ethyl Acetate Fraction of an Edible Red Macroalgae Sarcodia Ceylanica. Int J Mol Sci 2017, 18, 1–15. [CrossRef]
- Iacopucci, A.P.; Mello, R.O.; Barbosa-Silva, R.; Melo-Thomas, L. L-NOARG-Induced Catalepsy Can Be Influenced by Glutamatergic Neurotransmission Mediated by NMDA Receptors in the Inferior Colliculus. Behavioural Brain Research 2012, 234, 149–154. [CrossRef]
- Kim, S.R.; Jung, Y.R.; Kim, D.H.; An, H.J.; Kim, M.K.; Kim, N.D.; Chung, H.Y. Caffeic Acid Regulates LPS-Induced NF-ΚB Activation through NIK/IKK and c-Src/ERK Signaling Pathways in Endothelial Cells. Arch Pharm Res 2014, 37, 539–547.
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The Cloned Capsaicin Receptor Integrates Multiple Pain-Producing Stimuli. Neuron 1998, 21, 531–543. [CrossRef]
- Devesa, I.; Planells-Cases, R.; Fernández-Ballester, G.; González-Ros, J.M.; Ferrer-Montiel, A.; Fernández-Carvajal, A. Role of the Transient Receptor Potential Vanilloid 1 in Inflammation and Sepsis. J Inflamm Res 2011, 4, 67–81. [CrossRef]
- Gamse, R.; Holzer, P.; Lembeck, F. Indirect Evidence for Presynaptic Location of Opiate Receptors on Chemosensitive Primary Sensory Neurones. Naunyn Schmiedebergs Arch Pharmacol 1979, 308, 281–285. [CrossRef]
- Maia, J.L.; Lima-Júnior, R.C.P.; Melo, C.M.; David, J.P.; David, J.M.; Campos, A.R.; Santos, F.A.; Rao, V.S.N. Oleanolic Acid, a Pentacyclic Triterpene Attenuates Capsaicin-Induced Nociception in Mice: Possible Mechanisms. Pharmacol Res 2006, 54, 282–286. [CrossRef]
- Beirith, A.; Santos, A.R.S.; Calixto, J.B. Mechanisms Underlying the Nociception and Paw Oedema Caused by Injection of Glutamate into the Mouse Paw. Brain Res 2002, 924, 219–228. [CrossRef]
- Vorobeychik, Y.; Willoughby, C.D.; Mao, J. NMDA Receptor Antagonists in the Treatment of Pain. In Comprehensive Treatment of Chronic Pain by Medical, Interventional, and Integrative Approaches; Springer New York: New York, NY, 2013; pp. 61–67.
- Soares, A.C.; Leite, R.; Tatsuo, M.A.K.F.; Duarte, I.D.G. Activation of ATP-Sensitive K+ Channels: Mechanism of Peripheral Antinociceptive Action of the Nitric Oxide Donor, Sodium Nitroprusside. Eur J Pharmacol 2000, 400, 67–71. [CrossRef]
- Garthwaite, J. Glutamate, Nitric Oxide and Cell-Cell Signalling in the Nervous System. Trends Neurosci 1991, 14, 60–67. [CrossRef]
- Kimura, M.; Saito, S.; Obata, H. Dexmedetomidine Decreases Hyperalgesia in Neuropathic Pain by Increasing Acetylcholine in the Spinal Cord. Neurosci Lett 2012, 529, 70–74. [CrossRef]
- Obata, H. Analgesic Mechanisms of Antidepressants for Neuropathic Pain. Int J Mol Sci 2017, 18. [CrossRef]
- Chan, A.K.M.; Cheung, C.W.; Chong, Y.K. Alpha-2 Agonists in Acute Pain Management. Expert Opin Pharmacother 2010, 11, 2849–2868. [CrossRef]
- Lopes, L.D.S.; Marques, R.B.; Fernandes, H.B.; Pereira, S.D.S.; Ayres, M.C.C.; Chaves, M.H.; Almeida, F.R.C. Mechanisms of the Antinociceptive Action of (-) Epicatechin Obtained from the Hydroalcoholic Fraction of Combretum Leprosum Mart & Eic in Rodents. J Biomed Sci 2012, 19, 2–7. [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V. V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with GNPS. Nat Biotechnol 2016, 34, 828. [CrossRef]
- Dunham, N.W.; Miya, T.S. A Note on a Simple Apparatus for Detecting Neurological Deficit in Rats and Mice. J Am Pharm Assoc 1957, 46, 208–209.
- Lapa, A.J.J.; Souccar, C.; Lima-Landman, M.T.R.; Castro, M.S. de A.; Lima, T.C.M. Pharmacological Activity Evaluation Methods of Medicinal Plants; Sociedade Brasileira de Plantas Medicinais, 2008;
- Pessoa, R.T.; Alcântara, I.S.; da Silva, L.Y.S.; da Costa, R.H.S.; Silva, T.M.; de Morais Oliveira-Tintino, C.D.; Ramos, A.G.B.; de Oliveira, M.R.C.; Martins, A.O.B.P.B.; de Lacerda, B.C.G.V.; et al. Ximenia Americana L.: Chemical Characterization and Gastroprotective Effect. Analytica 2023, 4, 141–158. [CrossRef]
- Koster, R. Acetic Acid for Analgesic Screening. In Proceedings of the Fed proc; 1959; Vol. 18, p. 412.
- Tjølsen, A.; Berge, O.-G.; Hunskaar, S.; Rosland, J.H.; Hole, K. The Formalin Test: An Evaluation of the Method. Pain 1992, 51, 5–17. [CrossRef]
- Bannon, A.; Malmberg, A. Models of Nociception: Hot-Plate, Tail-Flick, and Formalin Tests in Rodents; 2007; Vol. Chapter 8;
- Martinov, T.; Mack, M.; Sykes, A.; Chatterjea, D. Measuring Changes in Tactile Sensitivity in the Hind Paw of Mice Using an Electronic von Frey Apparatus. Journal of Visualized Experiments 2013. [CrossRef]
- Tena, B.; Escobar, B.; Arguis, M.J.; Cantero, C.; Rios, J.; Gomar, C. Reproducibility of Electronic von Frey and von Frey Monofilaments Testing. Clin J Pain 2012, 28, 318–323. [CrossRef]
- Stevenson, G.W.; Bilsky, E.J.; Negus, S.S. Targeting Pain-Suppressed Behaviors in Preclinical Assays of Pain and Analgesia: Effects of Morphine on Acetic Acid-Suppressed Feeding in C57BL/6J Mice. J Pain 2006, 7, 408–416. [CrossRef]
- Miller, E.C.; Swanson, A.B.; Phillips, D.H.; Fletcher, T.L.; Liem, A.; Miller, J.A. Structure-Activity Studies of the Carcinogenicities in the Mouse and Rat of Some Naturally Occurring and Synthetic Alkenylbenzene Derivatives Related to Safrole and Estragole. Cancer Res 1983, 43, 1124–1134.
- Schechtmann, G.; Song, Z.; Ultenius, C.; Meyerson, B.A.; Linderoth, B. Cholinergic Mechanisms Involved in the Pain Relieving Effect of Spinal Cord Stimulation in a Model of Neuropathy. Pain 2008, 139, 136–145. [CrossRef]
- Lam, D.K.; Sessle, B.J.; Cairns, B.E.; Hu, J.W. Neural Mechanisms of Temporomandibular Joint and Masticatory Muscle Pain: A Possible Role for Peripheral Glutamate Receptor Mechanisms. Pain Res Manag 2005, 10, 145–152. [CrossRef]
- Li, P.; Zhuo, M. Silent Glutamatergic Synapses and Nociception in Mammalian Spinal Cord. Nature 1998, 393, 695–698. [CrossRef]
- Majewska, M.D.; Bell, J.A.; London, E.D. Regulation of the NMDA Receptor by Redox Phenomena: Inhibitory Role of Ascorbate. Brain Res 1990, 537, 328–332. [CrossRef]
- Zeraati, F.; Araghchian, M.; Farjoo, M.H. Ascorbic Acid Interaction With Analgesic Effect of Morphine and Tramadol in Mice. Anesth Pain Med 2014, 4. [CrossRef]
- Santos, A.R.S.; Gadotti, V.M.; Oliveira, G.L.; Tibola, D.; Paszcuk, A.F.; Neto, A.; Spindola, H.M.; Souza, M.M.; Rodrigues, A.L.S.; Calixto, J.B. Mechanisms Involved in the Antinociception Caused by Agmatine in Mice. Neuropharmacology 2005, 48, 1021–1034. [CrossRef]
- Ocaña, M.; Cendán, C.M.; Cobos, E.J.; Entrena, J.M.; Baeyens, J.M. Potassium Channels and Pain: Present Realities and Future Opportunities. Eur J Pharmacol 2004, 500, 203–219. [CrossRef]
- Ferré, S.; Diamond, I.; Goldberg, S.R.; Yao, L.; Hourani, S.M.O.; Huang, Z.L.; Urade, Y.; Kitchen, I. Adenosine A2A Receptors in Ventral Striatum, Hypothalamus and Nociceptive Circuitry. Prog Neurobiol 2007, 83, 332–347. [CrossRef]
- Finan, P.H.; Smith, M.T. The Comorbidity of Insomnia, Chronic Pain, and Depression: Dopamine as a Putative Mechanism. Sleep Med Rev 2013, 17, 173–183. [CrossRef]
- Wood, P.B. Mesolimbic Dopaminergic Mechanisms and Pain Control. Pain 2006, 120, 230–234. [CrossRef]
- Maleki, N.; Nayebi, A.M.; Garjani, A. Effects of Central and Peripheral Depletion of Serotonergic System on Carrageenan-Induced Paw Oedema. Int Immunopharmacol 2005, 5, 1723–1730. [CrossRef]
- Suzuki, R.; Rygh, L.J.; Dickenson, A.H. Bad News from the Brain: Descending 5-HT Pathways That Control Spinal Pain Processing. Trends Pharmacol Sci 2004, 25, 613–617. [CrossRef]
Figure 1.
Chromatogram of ESI-MS scanning by UPLC-MS. 1A. ESI-MS scanning spectrum in negative ionization mode for the range of m/z 150-190. It is possible to observe the peak at m/z 179.05 referring to the caffeic acid product. 1B. ESI-MS/MS full scan spectrum in negative ionization mode, relevant to the peak at m/z 179.05 observed in the MS spectrum for the caffeic acid product.
Figure 1.
Chromatogram of ESI-MS scanning by UPLC-MS. 1A. ESI-MS scanning spectrum in negative ionization mode for the range of m/z 150-190. It is possible to observe the peak at m/z 179.05 referring to the caffeic acid product. 1B. ESI-MS/MS full scan spectrum in negative ionization mode, relevant to the peak at m/z 179.05 observed in the MS spectrum for the caffeic acid product.
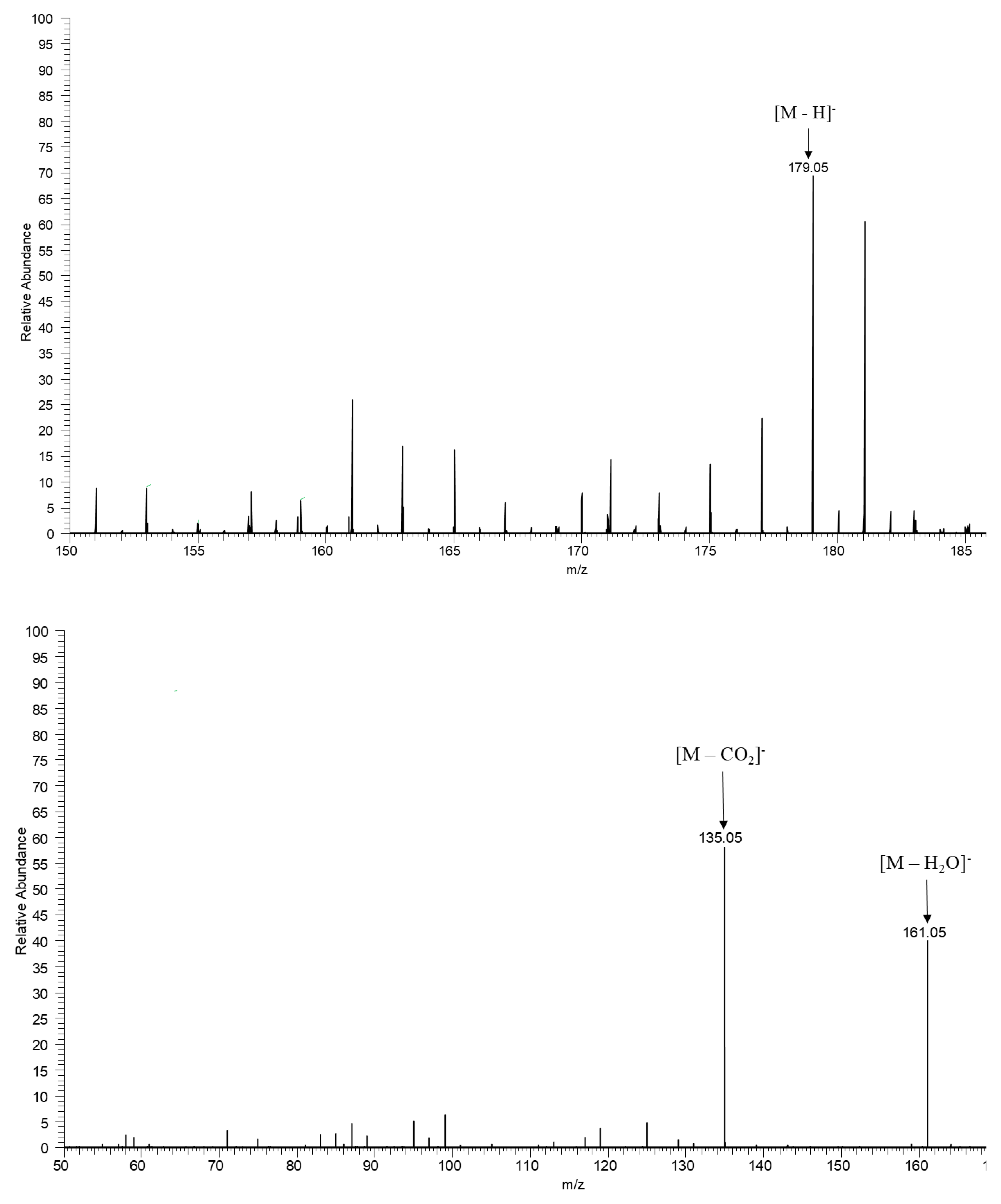
Figure 2.
Antinociceptive effect of HEXA (50, 100, and 200 mg/kg) by acetic acid-induced abdominal writhing test (`2A); the hot plate test (2B) and the hypernociception meansure by Von Frey test induced by CFA (2C).
Figure 2.
Antinociceptive effect of HEXA (50, 100, and 200 mg/kg) by acetic acid-induced abdominal writhing test (`2A); the hot plate test (2B) and the hypernociception meansure by Von Frey test induced by CFA (2C).
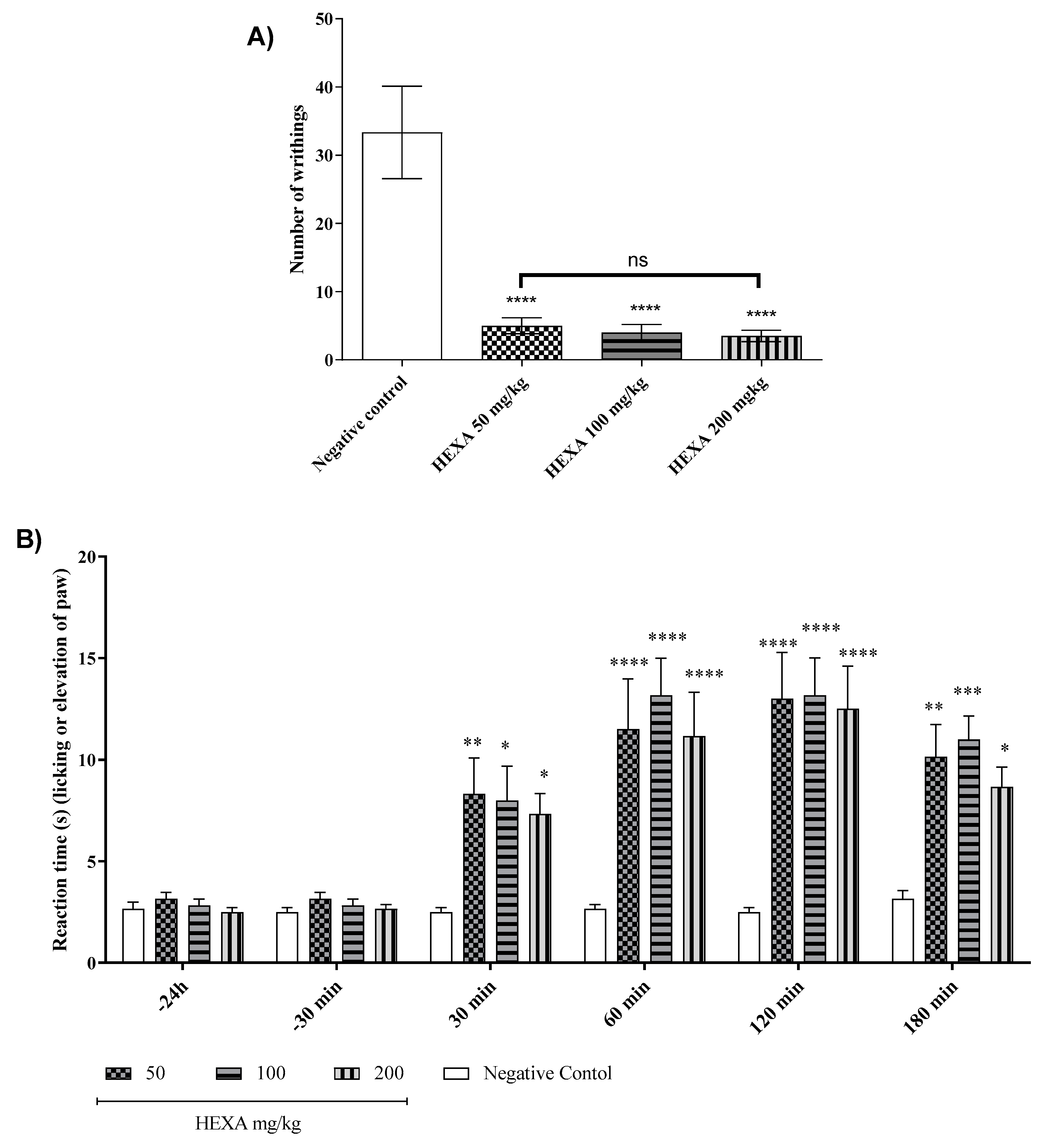
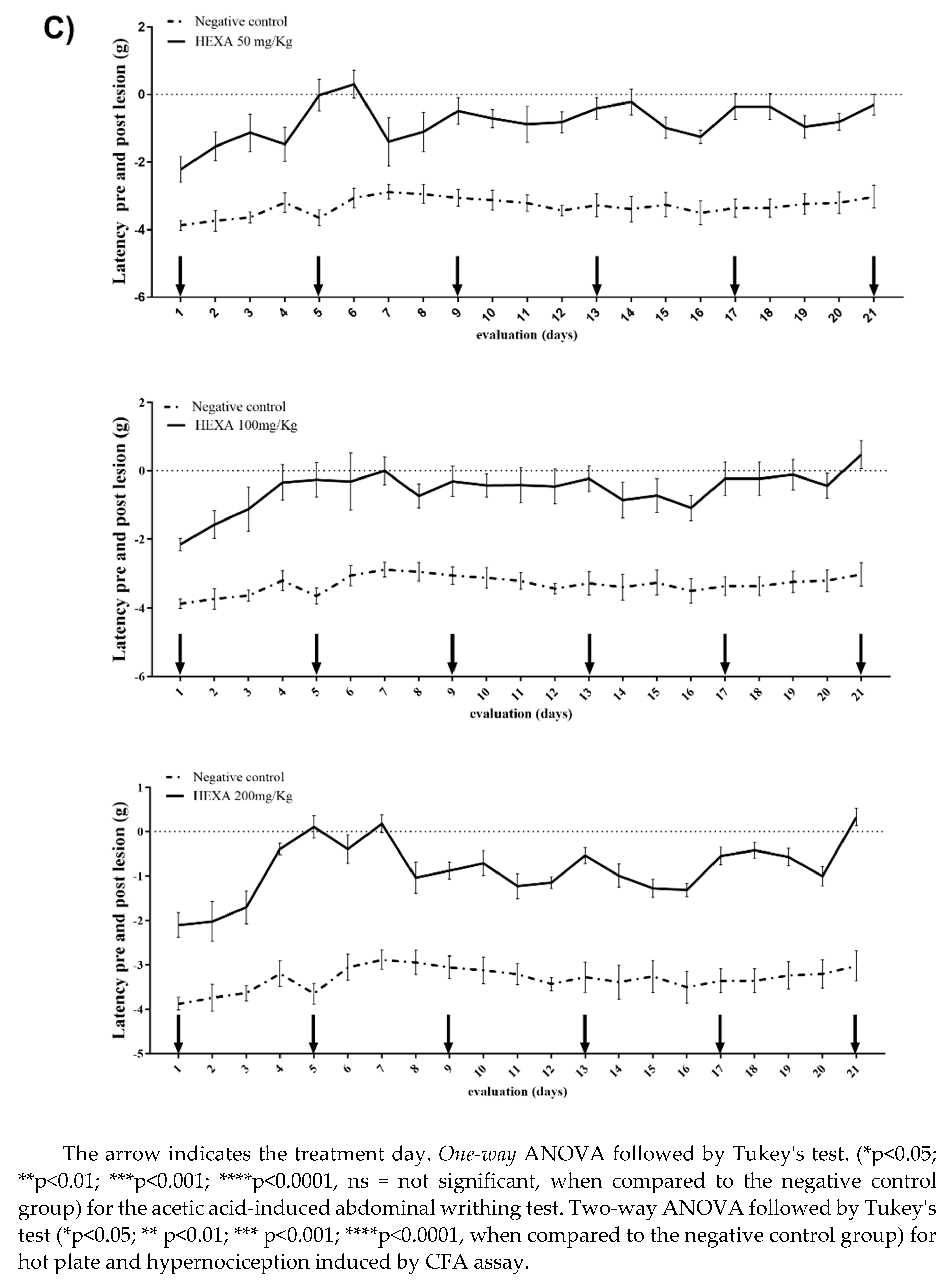
Figure 3.
Signaling pathways underlying the antinociceptive response of HEXA (100 mg/kg) and CA (1.8 mg/kg) in the antinociceptive response: (A) vanilloid; (B) glutamatergic; (C) opioid; (D) L-Arginine/Nitric Oxide/cGMP; (E) cyclic guanosine monophosphate; (F) Participation of α2-adrenergic receptors; (G) K+ATP channels against formalin-induced pain in mice.
Figure 3.
Signaling pathways underlying the antinociceptive response of HEXA (100 mg/kg) and CA (1.8 mg/kg) in the antinociceptive response: (A) vanilloid; (B) glutamatergic; (C) opioid; (D) L-Arginine/Nitric Oxide/cGMP; (E) cyclic guanosine monophosphate; (F) Participation of α2-adrenergic receptors; (G) K+ATP channels against formalin-induced pain in mice.
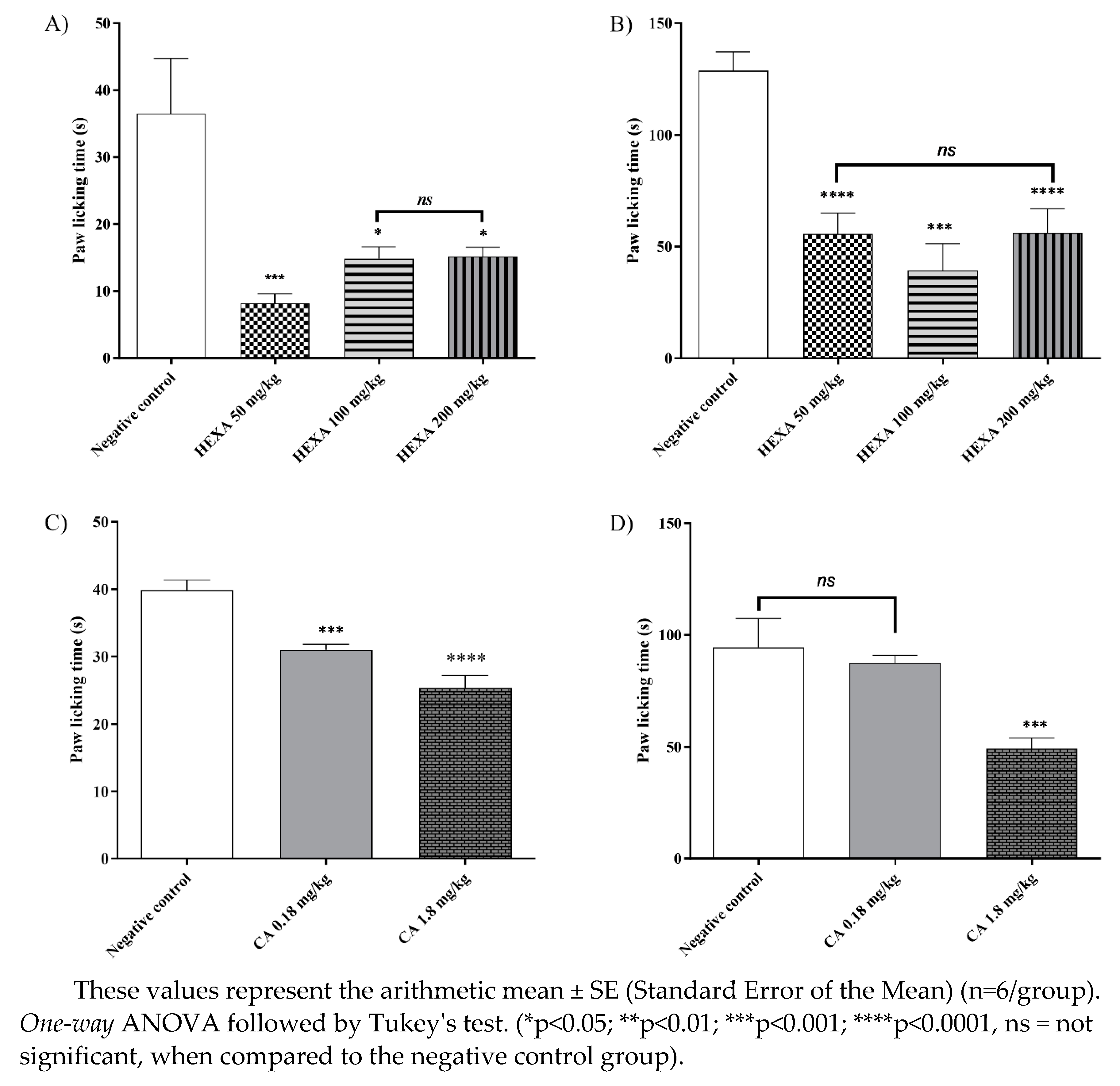
Figure 4.
Signaling pathways underlying the antinociceptive response of HEXA (100 mg/kg) and CA (1.8 mg/kg) in the antinociceptive response: (A) vanilloid; (B) glutamatergic; (C) opioid; (D) L-Arginine/Nitric Oxide/cGMP; (E) cyclic guanosine monophosphate; (F) Participation of α2-adrenergic receptors; (G) K+ATP channels against formalin-induced pain in mice.
Figure 4.
Signaling pathways underlying the antinociceptive response of HEXA (100 mg/kg) and CA (1.8 mg/kg) in the antinociceptive response: (A) vanilloid; (B) glutamatergic; (C) opioid; (D) L-Arginine/Nitric Oxide/cGMP; (E) cyclic guanosine monophosphate; (F) Participation of α2-adrenergic receptors; (G) K+ATP channels against formalin-induced pain in mice.
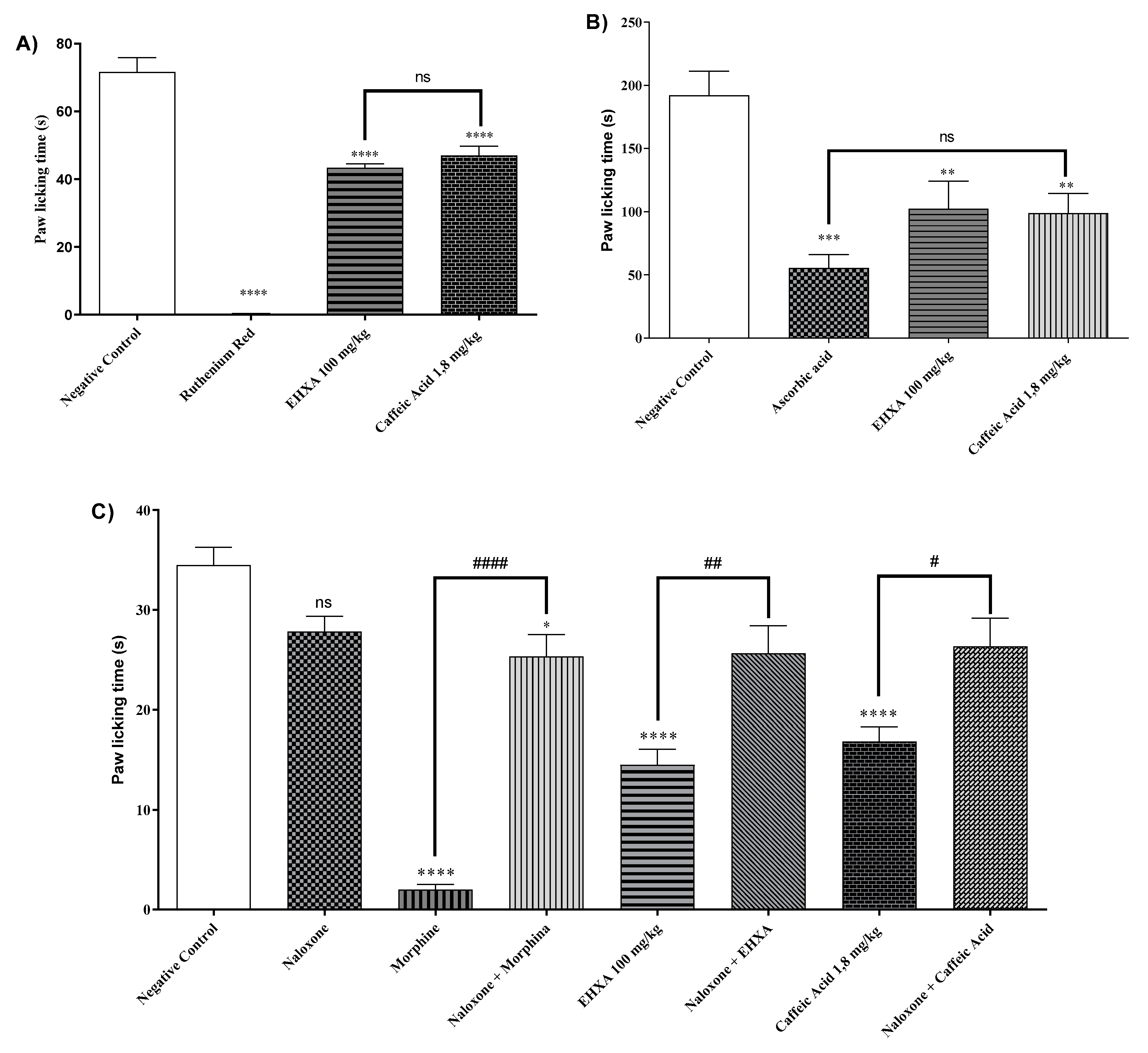
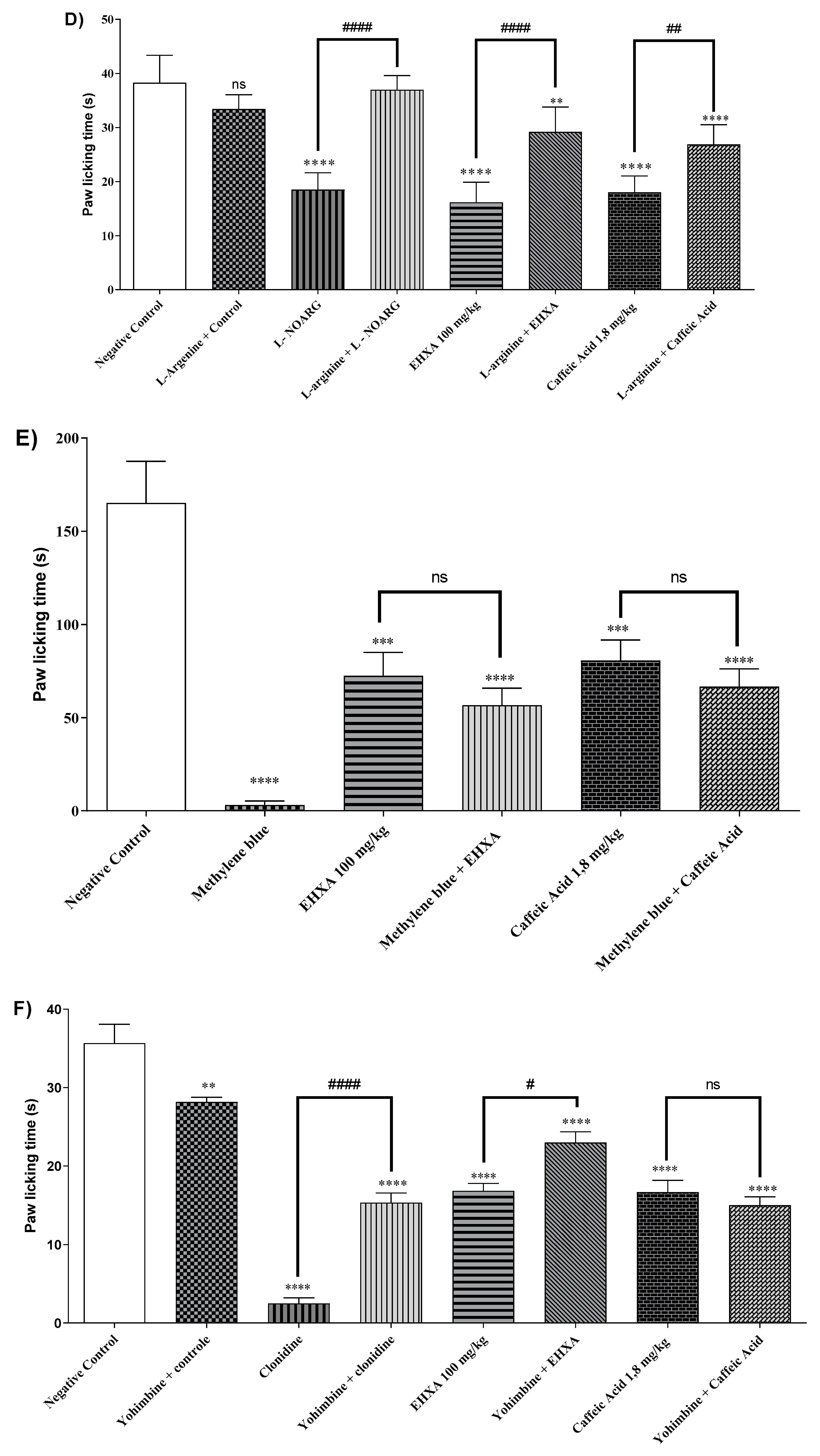
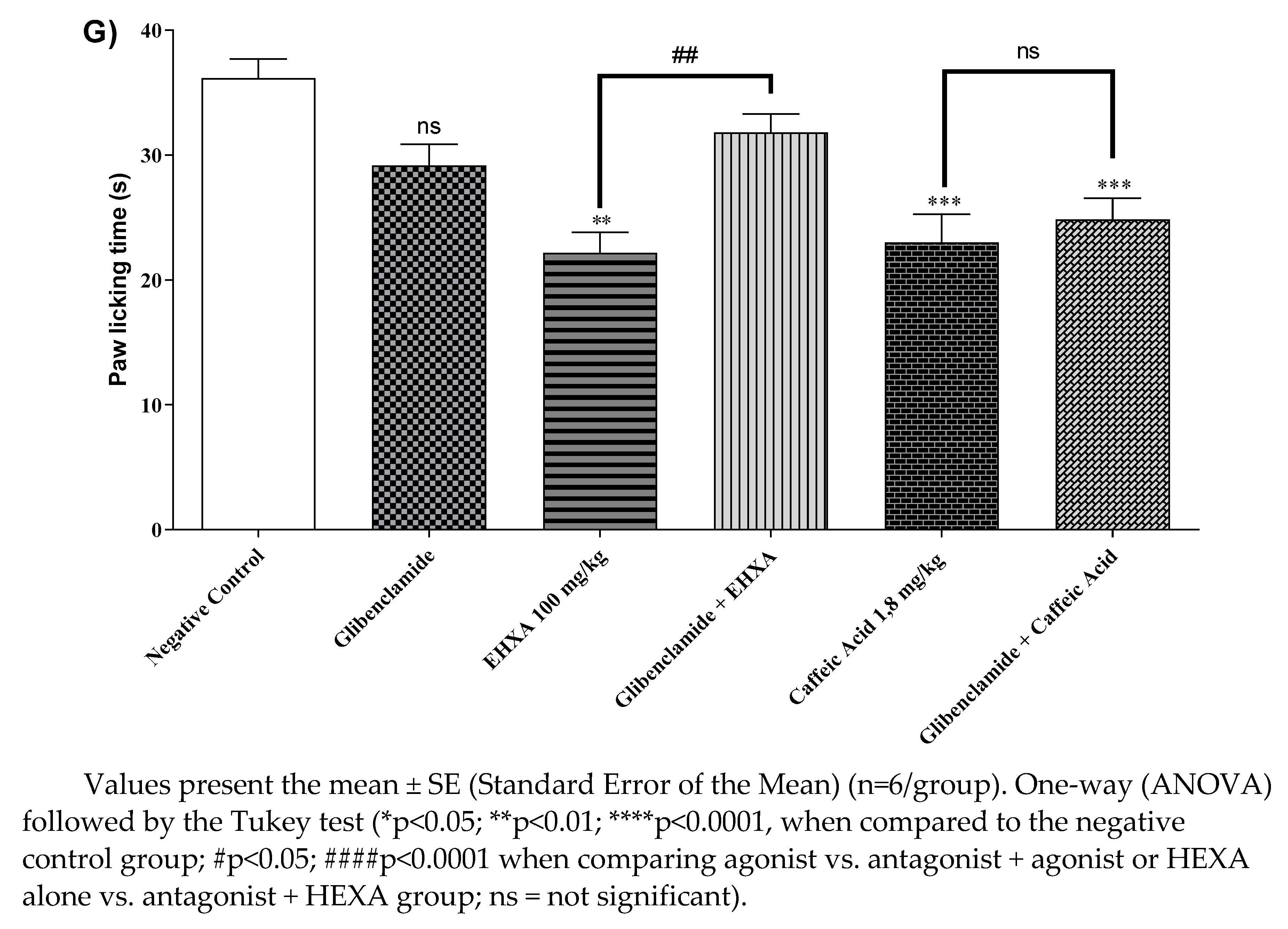

Table 1.
Results of compound annotation in the hydroethanolic extract of Ximenia americana L. through molecular network analysis.
Table 1.
Results of compound annotation in the hydroethanolic extract of Ximenia americana L. through molecular network analysis.
| Parent Mass | Adduct | Molecular Formula | Metabolite Name | Chemical Structure | Chemical Class |
|---|---|---|---|---|---|
| 169.014 | [M-H] - | C7H6O5 | Gallic acid | 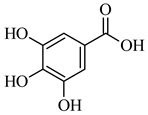 |
Organic acid |
| 179.034 | [M-H] - | C9H8O4 | Caffeic acid | 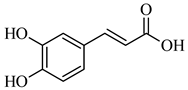 |
Organic acid |
| 195.050 | [M-H] - | C6H12O7 | Gluconic acid | 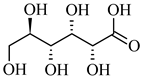 |
Organic acid |
| 193.034 | [M-H] - | C6H10O7 | D-Glucuronic acid | 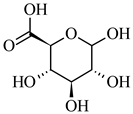 |
Organic acid |
| 285.039 | [M-H] - | C15H10O6 | Kaempferol | 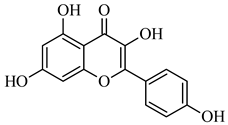 |
Flavonoid |
| 289.071 | [M-H] - | C15H14O6 | (-)-Catechin | 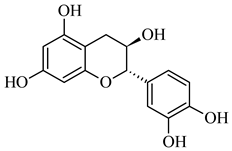 |
Flavonoid |
| 301.035 | [M-H] - | C15H10O7 | Quercetin |
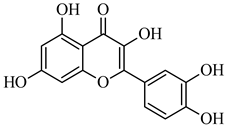 |
Flavonoid |
| 609.147 | [M-H] - | C27H30O16 | Rutin | 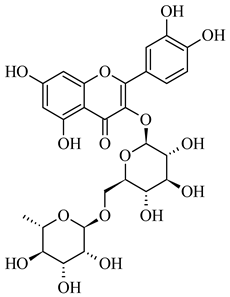 |
Flavonoid |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



