
Submitted:
09 April 2025
Posted:
10 April 2025
You are already at the latest version
Abstract
Suboptimal temperatures severely repress the nutrient absorption, growth, and yield formation of vegetables in solar greenhouse during winter and early spring in China. Alginate oligosaccharides (AOS) are anionic acidic polysaccharides derived from brown algae, known for promoting plant growth and alleviating abiotic stress. In this study, we aimed to investigate the effects of different nutrient solution concentration combined with AOS on the growth and nutrient uptake of cucumber seedling under suboptimal temperatures (15/8 °C, day/night). Potted ‘Jinchun No. 4’ cucumber seedlings grown in coconut coir were treated with 0.5×, 1.0×, or 1.5× strength of Hogland solution alone (N0.5, N1, N1.5), or with 30 mg·L⁻¹ AOS (A0.5, A1, A1.5). The results showed that growth attributes, and nitrogen accumulation of cucumber plants of N1 and N1.5 were significantly higher than those of N0.5. And plants of A0.5 exhibited significantly higher plant height, chlorophyll a content, root surface area, root volume, root vitality, nitrogen metabolism enzyme (NR, GDH, GS) activities and N accumulation, than those under N0.5, N1, or N1.5. Moreover, compared to A0.5, the net photosynthetic rate, total root length, root surface area, root N content, leaf nitrate reductase activity, root glutamate dehydrogenase activity and N accumulation of A1 and A1.5 were significantly higher than those of A0.5. Correlation analysis indicated strong positive relationships between root surface area, root volume, root vitality, and N content in plant organs. In summary, AOS enhanced the nutrient uptake and growth of cucumber seedlings through promoting root surface area, root vitality, and nitrogen metabolism enzyme activities under suboptimal temperatures conditions, therefor it has a good application prospect and great ecological value for increasing nutrient use efficiency and reducing the use of chemical fertilizers.
Keywords:
Sodium alginate oligosaccharides
; Cucumber
; Suboptimal temperature
; Nitrogen metabolism
; Root morphology
1. Introduction
Suboptimal temperature is one of the major limiting factors for horticultural production in solar greenhouse during cold seasons in northern China, which severely impairing nutrient absorption and consequently stunting plant growth and yield formation [1,2]. Enhanced nutrient supply under low-temperature conditions promoted plant growth [3,4,5]. At present stage, China is promoting green agricultural development, which emphasizing eco-friendly farming practices and advocating for reducing the use of chemical fertilizers. Biostimulants are natural or synthetic substances which can improve plant growth, stress tolerance, or nutrient use efficiency [6,7,8,9,10,11]. Alginate oligosaccharides (AOS), a type of biostimulant, are produced from natural polysaccharides, which extracted from brown algae and composed of dimeric units of mannuronic acid (M) and guluronic acid (G), through enzymatic or acid hydrolysis methods [12]. AOS are biodegradable and have the advantages of environment-friendliness [13]. Xu et al. found that AOS application under normal temperatures significantly expanded root surface area and root volume, improved nitrogen uptake, and increased the tissue nitrogen concentration in cucumber [14]. Additionally, AOS application under suboptimal temperatures significantly enhanced cucumber plant growth, chlorophyll content and photosynthetic capacity [15]. Therefore, it is interesting to investigate whether AOS can reduce the chemical fertilizer usage by prompting growth and nutrient utilization efficiency of horticultural crops at suboptimal temperatures. Cucumber (Cucumis sativus L.) is a widely cultivated horticultural crop that thrives at a temperature range of 22-32 ℃/15-18 ℃ (day/night) [16,17]. In this study, we employed seedling experiments to explore the combined effects of different concentrations of Hoagland nutrient solutions and exogenous AOS on the growth, root configuration, nutrient uptake, and nitrogen metabolism enzyme activities of cucumber seedlings under suboptimal temperature stress, and found that the combined application of exogenous AOS promoted root surface, root activities, nitrogen metabolism enzyme activities, and nutrient accumulation of cucumber seedlings in each concentration of nutrient solution under 15/8 °C temperatures (day/night). These findings provide theoretical support for improving nutrient absorption efficiency and mitigating the adverse effects of suboptimal temperatures on greenhouse vegetable production.
2. Materials and Methods
2.1. Experimental Materials
AOS were purchased from HEHAI Biotech (Qingdao, China). The cucumber seeds of Cv. Jinchun 4 were used in this study. Seeds were soaked in water at 55 °C for 15 minutes. When the water cooled to room temperature, seeds were transferred into a constant-temperature incubator and germinated at a temperature of 28°C for 24 hours. Germinated seeds were sowed into 72 holes seedling-raising plates filled with a mixture of coir-coconut, vermiculite, and perlite (2:1:1, v:v:v). Seedlings were irrigated with 0.5 strength of Hogland solution and raised under natural temperature and light conditions in solar greenhouse. Seedlings were then transplanted into pots containing coir-coconut, when first true leaves started to develop. After a 5-day rejuvenation period, seedlings were transferred and acclimated in the climate chambers for 3 days under conditions of 25±1 °C/18±1 °C (day/night) with a 12-hour photoperiod (500±20 μmol·m⁻²·s⁻¹).
2.2. Experimental Design
Suboptimal temperatures were set at 15±1 °C during the day and 8±1 °C at night, with a 12-hour photoperiod (400±20 μmol·m⁻²·s⁻¹). Cucumber seedlings were treated with 0.5×, 1.0× and 1.5× strength of Hogland solution without AOS (N0.5, N1, and N1.5), or with 30 mg·L⁻¹ AOS (A0.5, A1, and A1.5). Seedlings were irrigated every 3 days (100 mL per plant) and 5 times during the experiment. Sampling and measurements were conducted on day 18 after treatment. Each treatment was replicated three times.
2.3.1. Biomass Measurement
Plant height was defined as the height from the cotyledon node to the apex of the stem and determined with a ruler. Stem diameter was measured at the midpoint between the cotyledon and the first true leaf using a digital Vernier caliper. Leaf area was determined with LA-S analyze system (WanShen, Hangzhou, China).
Seedling plants were separated into roots, stems, and leaves. Then fresh organ weights were weighted after rinsed 3 times with distilled water and blotted dry with filter paper. And samples were dried at 105 °C for 15 minutes, then at 80 °C until constant weight in an oven to determine dry weights.
2.3.2. Measurement of Photosynthetic Characteristics
The net photosynthetic rate (Pn) of the second fully expanded leaf from top was measured with Li-6800XT photosynthesis system (LI-COR Biosciences, Lincoln, NE, USA) under a photosynthetic photon flux density (PPFD) of 400 μmol·m⁻²·s⁻¹.
2.3.3. Measurement of Physiological Indicators
Roots were carefully washed, scanned using V850 Pro scanner (EPSON, Nagano, Japan), and analyzed with WinRHIZO root analysis software (Regent, Quebec, Canada).
2.3.4. NPK Content Estimation
The oven-dried roots, stems, and leaves were ground to pass a 100 mesh sievedigested and analyzed for total N, P and K according to the methods of Thomas et al. [18].
Nutrient accumulation = nutrient content × dry biomass weight;
Nutrient distribution rate (%) = nutrient accumulation per organ/total plant nutrient accumulation × 100%.
2.3.5. Biochemical Analysis Assays
Leaf samples were collected from the second fully expanded, healthy leaf without pest or disease damage. Root samples were collected from the primary root zone near the root tip. The chlorophyll content and the root activity were determined as previously described [19,20]. Samples for enzyme activity analyzing were rapidly frozen in liquid nitrogen and then stored at -80 °C. Enzyme activities of nitrate reductase (NR), glutamine synthetase (GS), glutamate synthase (GOGAT), and glutamate dehydrogenase (GDH), were determined using corresponding assay kits (Solarbio, Beijing, China) following the manufacturer’s instructions.
2.4. Data Analysis
Experimental data were processed using Microsoft Excel and visualized with GraphPad Prism 8 (GraphPad Software Inc., San Diego, CA), Correlation analysis of the heat map using ChiPlot (https://www.chiplot.online/) drawn intraclass correlation analysis. Statistical analysis was performed with the Data Processing System (DPS) version 17.0 (Refine, Hangzhou, China). Comparison between groups was made by two-way ANOVA, followed by Duncan’s new multiple range test for multiple comparisons (p < 0.05). Data were presented as mean±SD (n = 3).
3. Results
3.1. Effects of Different Nutrient Solution Concentration with AOS on the Growth of Cucumber Seedlings under Suboptimal Temperature
Two-way ANOVA revealed that exogenous AOS and the interaction between nutrient level (NL) and AOS (NL×AOS) significantly affected all nine plant growth parameters measured (Figure 1). In contrast, NL alone significantly influenced only plant height, stem diameter, leaf area, and stem fresh weight (FW). The root dry weight (DW) in A1.5 was 35.50% higher than in N1.5. Similarly, leaf DW in A1 and A1.5 increased by 34.91% and 31.09%, respectively, compared to N1 and N1.5.
Two-way ANOVA revealed that both AOS and the NL × AOS interaction significantly influenced all six root traits, while NL alone significantly affected root tip numbers, total root length, root forks, and root surface area (Figure 2). A0.5 significantly increased the number of root tips compared to N0.5 and N1. Furthermore, A1.5 showed a 36.17% increase in root tips compared to A0.5. Root diameter and total root length were significantly greater in A0.5 than in N0.5. Moreover, root average diameter in A1.5, total root length in A1.0 and A1.5 were significantly greater than those in A0.5. A0.5 treatment resulted in significantly more root forks, larger root surface area, and greater root volume than N0.5, N1, and N1.5. Moreover, A1.5 increased root forks by 13.87% relative to A0.5. Meanwhile, A1 and A1.5 significantly increased root surface area by 26.81% and 36.72%, respectively, compared with that of A0.5. Finally, root volume was significantly greater in AOS-treated plants than in non-AOS treatments.
3.2. Effects of AOS and Different Nutrient Solution Levels on the Chlorophyll Content and Net Photosynthetic rate of Cucumber Seedlings Under Suboptimal Temperature
The chlorophyll a and b contents in N1.5 were 14.33% and 46.28% higher than those in N0.5, respectively (Figure 3a-b). The A0.5 treatment significantly increased chlorophyll a content compared to N0.5, N1, and N1.5. furthermore, A1 and A1.5 further increased it by 9.10% and 11.46%, respectively. Similarly, chlorophyll b content was significantly higher in A0.5 than in N0.5. Two-way ANOVA analysis demonstrated that AOS, and NL × AOS have significant effect on chlorophyll a and b contents, while NL has significant effect on chlorophyll a content.
The Pn of N1.5 was 26.55% and 16.45% higher than that of N0.5 and N1, respectively (Figure 3c). Compared to A0.5, A1 and A1.5 significantly increased Pn by 10.40% and 23.06%, respectively. Additionally, Pn was significantly higher in nutrient solutions containing AOS than in those without it. Two-way ANOVA revealed that Pn was significantly influenced by NL, AOS, and their interaction (NL × AOS).
3.3. Effects of AOS and Different Nutrient Solution Levels on the Root Activity of Cucumber Seedlings Under Suboptimal Temperature
Plants in A0.5 exhibited significantly higher root activity compared to those receiving N0.5, N1, and N1.5 treatments (Figure 4). Furthermore, the A1.5 treatment enhanced root activity by 19.97% relative to A0.5. Statistical analysis revealed that root activity was significantly influenced by NL, AOS, and their interaction (NL×AOS).
3.4. Effects of AOS and Different Nutrient Solution Levels on the Nitrogen Metabolism Enzymes Activities in Cucumber Seedlings Under Suboptimal Temperature
Statistical analysis revealed significant effects of AOS and the NL×AOS interaction on all four nitrogen metabolism enzymes in both leaves and roots (Figure 5). While NL has significant effects on the activities of GDH in leaf, as well as GOGAT, GDH, and GS in root. With the exception of root GOGAT activity, the activities of nitrogen metabolism enzymes were significantly greater for plants in A0.5 than those grown in nutrient solutions without AOS in both the leaf and the root. Moreover, the activities of NR, GDH, and GS in leaf, as well as GOGA, GDH, and GS in root in A1.5 were significantly greater than those in A0.5.
3.5. Effects of AOS and Different Nutrient Solution Levels on Nitrogen Content, Accumulation, and Distribution in Different Cucumber Organs Under Suboptimal Temperature
Two-way ANOVA analysis showed that NL, AOS, and NL×AOS all significantly affected tissue N content and plant N accumulation (Figure 6a-b). N content in root and stem were significantly higher for plants in A0.5 than those grown in nutrient solutions without AOS. Moreover, N content in root and leaf in A1 and A1.5 were significantly greater than those in A0.5. N accumulation in root and stem were significantly higher for plants in A0.5 than those grown in nutrient solutions without AOS. N accumulation in leaf of A0.5 increased by 28.80% and 19.69%, respectively, compared with that in N0.5 and N1. Furthermore, N accumulation in plant organs in A1 and A1.5 were significantly more than those in A0.5.
Neither NL and AOS, nor NL×AOS significantly affected distribution of N in cucumber seedlings (Figure 6c). The nitrogen distribution ratio in cucumber roots decreased slightly with increasing nutrient solution concentration, while leaf nitrogen distribution increased slightly without significance. However, exogenous AOS significantly enhanced nitrogen allocation to leaves, indicating that AOS application promoted nitrogen transport from roots to shoots, enhancing overall plant growth.
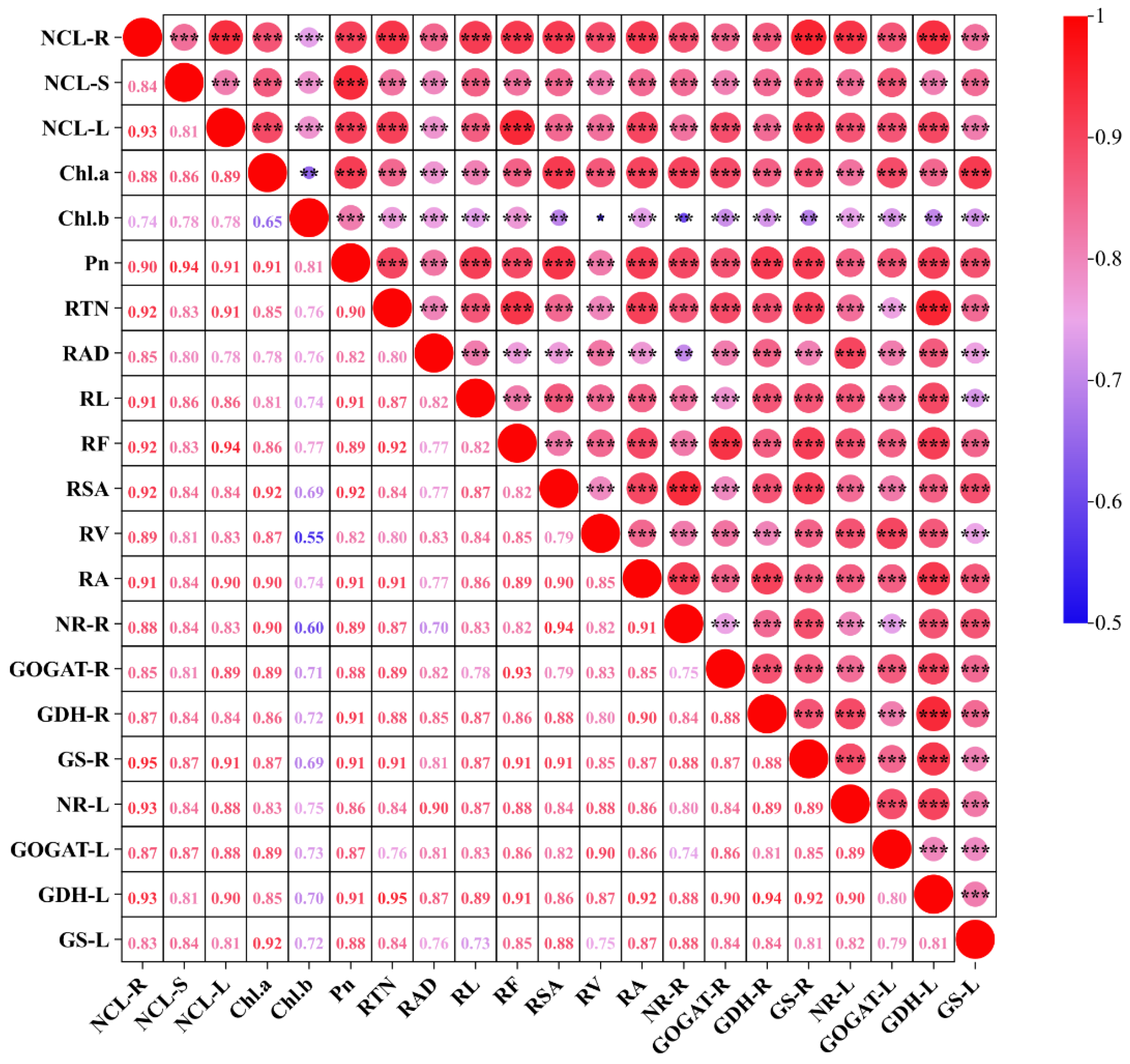
3.6. Correlation Analysis Between Different Traits
Correlation analysis revealed significant positive relationships between tissue nitrogen content and various physiological indices (Figure 7).Root N content was positively correlated with several indices, ranked from highest to lowest as follows: GS-R, GDH-L, NR-L, RTN, RF, RSA, RL, RA, Pn, RV, NR-R, Chl.a, GOGAT-L, GDH-R, GOGAT-R, RAD, GS-L and Chl.b. Stem N content exhibited positive correlations with the following indices, in descending order: Pn, GS-R, GOGAT-L, Chl.a, RL, RSA, RA, GDH-R, GS-L, NR-R, NR-L, RF, RTN, RV, GDH-L, GOGAT-R, RAD and Chl.b. Leaf N content was positively correlated with various indices, ordered from highest to lowest as follows: RF, RTN, Pn, GS-R, RA, GDH-L, Chl.a, GOGAT-R, NR-L, GOGAT-L, RL, RSA, GDH-R, RV, NR-R, GS-L, Chl.b and RAD.
4. Discussion
While low temperatures significantly inhibit nutrient uptake, particularly nitrogen assimilation, resulting in marked growth reductions, supplemental nutrient application under these conditions substantially enhanced plant biomass production [3,4,5]. Previous studies have demonstrated that exogenous trehalose as well as low-molecular-weight sodium alginate modulates plant nutrient uptake, and boosted N, P, and K content of crop plants [21,22]. In our study, elevated nutrient solution concentrations resulted in significantly higher N, P, and K content and accumulation across all examined tissues (roots, stems, and leaves) of cucumber seedlings (Figure 6, S1-S2). Notably, the co-application of exogenous AOS further enhanced these nutrient parameters, demonstrating a synergistic effect on mineral nutrient uptake and distribution. These findings were consistent with previous findings.
Root architecture is of great importance for the ability of crops to absorb mineral nutrients [23]. And modification of root architecture represents an effective strategy to enhance nutrient use efficiency and crop yield [24]. Previous studies indicated that exogenous AOS could elevate the levels of auxins and gibberellins in roots, stimulate cell division, and promote root elongation, thereby expanding the root absorption area and enhancing nitrogen uptake [25,26,27,28]. In this study, the combined application of varying nutrient solution concentrations and exogenous AOS enhanced cucumber root architecture, significantly increasing root tip number, average root diameter, total root length, root volume, and root surface area. Additionally, these root morphological characteristics exhibited strong positive correlations with organ-level nitrogen content (Figure 7), which align with established theories connecting root system architecture to nutrient uptake capacity [24,29]. Root vitality is closely correlated with mineral nutrient uptake [20,30]. Previous studies have reported that exogenous AOS increased not only the root absorption area but also root vitality in Chinese cabbage [28]. Our results also demonstrate that exogenous AOS, nutrient solution, and their synergistic interaction significantly enhanced root activity in cucumber seedlings. These findings suggest that exogenous AOS improves root vitality and architecture to facilitate more efficient nutrient uptake, thereby promoting plant growth under suboptimal temperature conditions.
Crop nitrogen use efficiency (NUE) is governed by four key physiological processes: uptake, transport, assimilation, and remobilization. The assimilation phase involves several critical enzymes, including nitrate reductase (NR), glutamine synthetase (GS), glutamate synthase (GOGAT), and glutamate dehydrogenase (GDH). Among these, NR initiates inorganic N utilization, while the GS/GOGAT cycle drives the conversion of inorganic N to organic forms, ultimately determining NUE. Genetic evidence underscores the functional importance of these enzymes in NUE [31]. In Arabidopsis, the nia1 nia2 double mutant retains merely 0.5% of WT NR activity and exhibits severe growth defects under nitrate supply [32]. Similarly, knockdown of OsNR2 in rice significantly reduces grain yield [33].GS and GOGAT encoding genes in roots are transcriptionally activated by NO₃⁻ supply [34,35,36]. The Arabidopsis gdh1 mutant shows impaired growth when supplemented with inorganic N [37]. Increased nitrogen metabolism enzyme activities facilitate rapid nitrogen assimilation, promoting growth and development [38]. Exogenous chitosan and chitosan oligosaccharides significantly enhanced activities of NR, GS and GOGAT, which enables plants to efficiently convert nitrite into amino acids, facilitating nitrogen uptake [39]. Moreover, Kchikich et al. showed that seaweed extract significantly increased the GS and GDH activities in leaves, and promoted the effective absorption and assimilation of nitrogen in sorghum plants under cadmium stress [40]. Low-molecular-weight sodium alginate fragments produced by radiation degradation was also reported to enhance nitrogen metabolic enzyme activities in various crop species [22,41,42]. Studies by Zhang et al. confirmed that AOS stimulates NR enzymatic activity via coordinated gene expression induction and post-transcriptional control mechanisms in wheat roots [28]. They also reported that the Ca²⁺ metabolism inhibitors attenuated the promoting effects of AOS on NR and GS activities, as well as total nitrogen concentration in flowering Chinese cabbage, which suggested Ca²⁺ signaling participates in mediating AOS-triggered nitrogen metabolic regulation [43]. In this study, exogenous AOS combined with nutrient solutions significantly enhanced NR, GOGAT, GDH and GS activities in plants, and showing a positive correlation with stem nitrogen content.
Photosynthesis critically influences nitrogen assimilation through both direct and indirect mechanisms. In roots, inorganic nitrogen assimilation relies on respiratory energy and carbon skeletons derived from sucrose imported from source leaves. Alternatively, nitrate and ammonium can be transported via the xylem to leaves for assimilation. Notably, leaf-based nitrogen assimilation occurs at significantly higher rates in the light than in the dark [44,45,46], underscoring photosynthesis directly supplies the reducing power and ATP required for nitrogen assimilation [47]. On the other side, Nitrogen plays a critical role in amino acid synthesis, which is essential for forming key components of the photosynthetic machinery, including chlorophyll, light-harvesting complexes, and the Rubisco enzyme. By supporting these structures, nitrogen enables the light-dependent assimilation of carbon dioxide, water, and inorganic nitrogen into sugars, organic acids, and amino acids—the fundamental building blocks of biomass [48]. Across the effective nitrogen application range, elevated N supply consistently improved photosynthetic performance, as evidenced by higher values of Pn, Fv/Fm and ΦPSII, thus improved crop yield [49]. Wang et al. reported that Achnatherum inebrians plants infected with Epichloë gansuensis exhibited enhanced nitrogen metabolism enzyme activity, and improved nitrogen use efficiency, accompanied with greater photosynthetic capacity compared to uninfected plants [50]. Zhang et al. found that low-molecular-weight sodium alginate boosts photosynthetic carbon metabolism through altering the homeostasis of plant hormones in leaves [51]. Moreover, AOS was reported to significantly increased chlorophyll content, Rubisco activity, photosynthetic efficiency and nitrogen metabolism enzyme activity, thereby improving nitrogen absorption and utilization [41,42,43]. In this study, exogenous AOS combined with nutrient solutions significantly enhanced photosynthetic performance, thereby potentially improving nitrogen metabolism and promoting cucumber plant growth under suboptimal temperature stress.
5. Conclusions
Cucumber seedling growth was positively correlated with Hoagland nutrient solution concentration within a defined range under suboptimal temperatures. Notably, supplementation of nutrient solutions with exogenous AOS improved root architecture, enhanced nitrogen metabolism enzyme activity, and consequently promoted nitrogen absorption and utilization. Remarkably, AOS application under reduced nutrient conditions achieved growth performance comparable to those under high-concentration nutrient solutions, suggesting AOS as a fertilizer-saving strategy.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Effects of AOS and different nutrient solution levels on phosphorus content, accumulation, and distribution in different cucumber organs under suboptimal temperature; Figure S2: Effects of AOS and different nutrient solution levels on potassium content, accumulation, and distribution in different cucumber organs under suboptimal temperature.
Author Contributions
Conceptualization, L. B. and Y. L. (Yaling Li); methodology, Y. L. (Yun Li), M. S. and Y. M.; investigation, X. G., K. F., and L. G.; data curation, X. G., Y. L. (Yun Li) and L. B.; writing—original draft preparation, X. G. and Y. Y.; writing—review and editing, L. B., Y. L. (Yun Li) and Y. L. (Yaling Li); funding acquisition, L. H. and C. C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Shanxi Province Key R&D Plan (202102140601013, 202302010101003).
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AOS | Alginate oligosaccharides |
| NR | Nitrate reductase |
| GOGAT | Glutamate synthase |
| GS | Glutamine synthetase |
| GDH | Glutamate dehydrogenase |
| NL | Nutrient level |
| NCL-R | Nitrogen content in root |
| NCL-S | Nitrogen content in stem |
| NCL-L | Nitrogen content in leaf |
| Chl. a | Chlorophyll a content |
| Chl. b | Chlorophyll b content |
| Pn | Net photosynthetic rate |
| RTN | Root tip number |
| RAD | Root average diameter |
| RL | Total root length |
| RF | Root forks |
| RSA | Root surface area |
| RV | Root volume |
| RA | Root activity |
| NR-R | Nitrate reductase activity in root |
| GOGAT-R | Glutamate synthase activity in root |
| GDH-R | Glutamate dehydrogenase activity in root |
| GS-R | Glutamine synthetase in root. |
| NR-L | Nitrate reductase activity in leaf |
| GOGAT-L | Glutamate synthase activity in leaf |
| GDH-L | Glutamate dehydrogenase activity in leaf |
| GS-L | Glutamine synthetase in leaf |
| NUE | Nitrogen use efficiency |
References
- Aslam, M.; Fakher, B.; Ashraf, M.A.; Cheng, Y.; Wang, B.; Qin, Y. Plant low-temperature stress: Signaling and response. Agronomy 2022, 12, 702. [Google Scholar] [CrossRef]
- Hajihashemi, S.; Noedoost, F.; Geuns, J.M.C.; Djalovic, I.; Siddique, K.H.M. Effect of cold stress on photosynthetic traits, carbohydrates, morphology, and anatomy in nine cultivars of stevia rebaudiana. Frontiers in Plant Science 2018, 9, 1430. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Duan, Z.; Mao, J.; Li, X.; Dong, F. Effects of root-zone temperature and N, P, and K supplies on nutrient uptake of cucumber (Cucumis sativus L.) seedlings in hydroponics. Soil Science and Plant Nutrition 2012, 58, 707–717. [Google Scholar] [CrossRef]
- Haghighi, M.; Abdolahipour, B. Rootzone temperature on nitrogen absorption and some physiological traits in cucumber. Journal of Plant Process and Function 2020, 8, 51–59. Available online: https://sid.ir/paper/768371/en.
- Yan, Q.; Duan, Z.; Mao, J.; Li, X.; Dong, F. Low root zone temperature limits nutrient effects on cucumber seedling growth and induces adversity physiological response. Journal of Integrative Agriculture 2013, 12, 1450–1460. [Google Scholar] [CrossRef]
- D'Addabbo, T.; Laquale, S.; Perniola, M.; Candido, V. Biostimulants for plant growth promotion and sustainable management of phytoparasitic nematodes in vegetable crops. Agronomy 2019, 9, 616. [Google Scholar] [CrossRef]
- Li, P.; Yang, X.; Wang, H.; Pan, T.; Yang, J.; Wang, Y.; Xu, Y.; Yang, Z.; Xu, C. Metabolic responses to combined water deficit and salt stress in maize primary roots. Journal of Integrative Agriculture 2021, 20, 109–119. [Google Scholar] [CrossRef]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Khan, E.A. 24-Epibrassinolide application in plants: An implication for improving drought stress tolerance in plants. Plant Physiology and Biochemistry 2019, 135, 295–303. [Google Scholar] [CrossRef]
- Khan, W.; Aziz, T.; Hussain, I.; Ramzani, P.M.A.; Reichenauer, T.G. Silicon: a beneficial nutrient for maize crop to enhance photochemical efficiency of photosystem II under salt stress. Archives of Agronomy and Soil Science 2016, 63, 599–611. [Google Scholar] [CrossRef]
- Munir, N.; Hanif, M.; Abideen, Z.; Sohail, M.; El-Keblawy, A.; Radicetti, E.; Mancinelli, R.; Haider, G. Mechanisms and strategies of plant microbiome interactions to mitigate abiotic stresses. Agronomy 2022, 12, 2069. [Google Scholar] [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicology and Environmental Safety 2018, 156, 225–246. [Google Scholar] [CrossRef]
- Liu, M.; Liu, L.; Zhang, H.; Yi, B.; Everaert, N. Alginate oligosaccharides preparation, biological activities and their application in livestock and poultry. Journal of Integrative Agriculture 2021, 20, 24–34. [Google Scholar] [CrossRef]
- Zhang, C.; Li, M.; Rauf, A.; Khalil, A. A.; Shan, Z.; Chen, C.; Rengasamy, K. R.R.; Wan, C. Process and applications of alginate oligosaccharides with emphasis on health beneficial perspectives. Critical Reviews in Food Science and Nutrition 2021, 63, 303–329. [Google Scholar] [CrossRef]
- Xu, Y.; Han, Y.; Han, W.; Yang, Y.; Saito, M.; Lv, G.; Song, J.; Bai, W. Different oligosaccharides induce coordination and promotion of root growth and leaf senescence during strawberry and cucumber growth. Horticulturae 2024, 10, 627. [Google Scholar] [CrossRef]
- Tan, C.; Li, N.; Wang, Y.; Yu, X.; Yang, L.; Cao, R.; Ye, X. Integrated physiological and transcriptomic analyses revealed improved cold tolerance in cucumber (Cucumis sativus L.) by exogenous chitosan oligosaccharide. Int. J. Mol. Sci. 2023, 24, 6202. [Google Scholar] [CrossRef] [PubMed]
- Shibaeva, T.G.; Sherudilo, E.G.; Titov, A.F. Response of cucumber (Cucumis sativus L.) plants to prolonged permanent and short-term daily exposures to chilling temperature. Russian Journal of Plant Physiology 2018, 65, 286–294. [Google Scholar] [CrossRef]
- Bai, L.; Deng, H.; Zhang, X.; Yu, X.; Li, Y. Gibberellin is involved in inhibition of cucumber growth and nitrogen uptake at suboptimal root-zone temperatures. Plos One 2016, 11, e0156188. [Google Scholar] [CrossRef]
- Thomas, R.L.; Sheard, R.W.; Moyer, J.R. Comparison of conventional and automated procedures for nitrogen, phosphorus, and potassium analysis of plant material using a single digestion. Agron. J. 1967, 59, 240–243. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. Journal of Plant Physiology 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Clemensson-Lindell, A. Triphenyltetrazolium chloride as an indicator of fine-root vitality and environmental stress in coniferous forest stands: Applications and limitations. Plant Soil 1994, 159, 297–300. [Google Scholar] [CrossRef]
- Elkelish, A.; Alhudhaibi, A.M.; Hossain, A.S.; Haouala, F.; Alharbi, B.M.; El, B.M.F.; Badji, A.; AlJwaizea, N.I.; Sayed, A.A.S. Alleviating chromium-induced oxidative stress in Vigna radiata through exogenous trehalose application: Insights into growth, photosynthetic efficiency, mineral nutrient uptake, and reactive oxygen species scavenging enzyme activity enhancement. BMC Plant Biol. 2024, 24, 460. [Google Scholar] [CrossRef]
- Moussa, H.R.; Taha, M.A.; Dessoky, E.S.; Selem, E. Exploring the perspectives of irradiated sodium alginate on molecular and physiological parameters of heavy metal stressed Vigna radiata L. plants. Physiol Mol Biol Plants. 2023, 29, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, Y.; Xu, J.; Qi, J.; Liu, X.; Guo, L.; Zhang, H. Research on the mechanisms of phytohormone signaling in regulating root development. Plants 2024, 13, 3051. [Google Scholar] [CrossRef]
- Li, X.; Zeng, R.; Liao, H. Improving crop nutrient efficiency through root architecture modifications. Journal of Integrative Plant Biology 2016, 58, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.; Matsubara, Y. Purification of alginate oligosaccharides with root growth-promoting activity toward lettuce. Biosci Biotechnol Biochem. 2000, 64, 1067–1070. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, H.; Zhao, X.; Wang, W.; Du, Y.; He, A.; Sun, K. The promoting effects of alginate oligosaccharides on root development in Oryza sativa L. mediated by auxin signaling. Carbohydr Polym. 2014, 113, 446–454. [Google Scholar] [CrossRef]
- Yang, J.; Shen, Z.; Sun, Z.; Wang, P.; Jiang, X. Growth stimulation activity of alginate-derived oligosaccharides with different molecular weights and mannuronate/guluronate ratio on Hordeum vulgare L. J Plant Growth Regul 2021, 40, 91–100. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, H.; Heng, Y.; Wang, W.; Zhao, X.; Du, Y. Nitric oxide mediates alginate oligosaccharides-induced root development in wheat (Triticum aestivum L.). Plant Physiol Biochem. 2013, 71, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Zhang, X.; Chen, P.; Du, Q.; Zhou, Y.; Yang, H.; Wang, X.; Yang, F.; Yong, T.; Yang, W. Improving maize's N uptake and N use efficiency by strengthening roots' absorption capacity when intercropped with legumes. Peer J 2021, 9, e11658. [Google Scholar] [CrossRef]
- Xie, X.; Weng, B.; Cai, B.; Dong, Y.; Yan, C. Effects of arbuscular mycorrhizal inoculation and phosphorus supply on the growth and nutrient uptake of Kandelia obovata (Sheue, Liu & Yong) seedlings in autoclaved soil. Applied Soil Ecology 2014, 75, 162–171. [Google Scholar] [CrossRef]
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: Current status and future prospects. J Genet Genomics 2022, 49, 394–404. [Google Scholar] [CrossRef]
- Wilkinson, J.Q.; Crawford, N.M. Identification and characterization of a chlorate-resistant mutant of Arabidopsis thaliana with mutations in both nitrate reductase structural genes NIA1 and NIA2. Molec. Gen. Genet. 1993, 239, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Wang, Y.; Chen, G.; Zhang, A.; Yang, S.; Shang, L.; Wang, D.; Ruan, B.; Liu, C.; Jiang, H.; Dong, G.; Zhu. L.; Hu, J.; Zhang, G.; Zeng, D.; Guo, L.; Xu, G.; Teng, S.; Harberd, N.P.; Qian, Q. The indica nitrate reductase gene OsNR2 allele enhances rice yield potential and nitrogen use efficiency. Nat Commun 2019, 10, 5207. [CrossRef]
- Redinbaugh, M.G.; Campbell, W.H. Glutamine synthetase and ferredoxin-dependent glutamate synthase expression in the maize (Zea mays) root primary response to nitrate (evidence for an organ-specific response). Plant Physiol. 1993, 101, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Prinsi, B.; Espen, L. Mineral nitrogen sources differently affect root glutamine synthetase isoforms and amino acid balance among organs in maize. BMC Plant Biol. 2015, 15, 96. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Wang, X.; Zhang, Z.; Xiong, S.; Meng, X.; Zhang, J.; Wang, L.; Zhang, X.; Yu, M.; Ma, X. Nitrogen regulating the expression and localization of four glutamine synthetase isoforms in wheat (Triticum aestivum L.). Int J Mol Sci. 2020, 21, 6299. [Google Scholar] [CrossRef]
- Melo-Oliveira, R.; Oliveira, I.C.; Coruzzi, G.M. Arabidopsis mutant analysis and gene regulation define a nonredundant role for glutamate dehydrogenase in nitrogen assimilation. Proc Natl Acad Sci U S A. 1996, 93, 4718–23. [Google Scholar] [CrossRef]
- Li, S.; Jiao, B.; Wang, J.; Zhao, P.; Dong, F.; Yang, F.; Ma, C.; Guo, P.; Zhou, S. Identification of wheat glutamate synthetase gene family and expression analysis under nitrogen stress. Genes 2024, 15, 827. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, J.; Gao, W.; Chen, Y.; Li, H.; Lawlor, D.W.; Paul, M.J.; Pan, W. Exogenous trehalose improves growth under limiting nitrogen through upregulation of nitrogen metabolism. BMC Plant Biol. 2017, 17, 247. [Google Scholar] [CrossRef]
- Kchikich, A.; Roussi, Z.; Krid, A.; Nhhala, N.; Ennoury, A.; Benmrid, B.; Kounnoun, A.; Maadoudi, M.E.; Nhiri, N.; Mohamed, N. Effects of mycorrhizal symbiosis and Ulva lactuca seaweed extract on growth, carbon/nitrogen metabolism, and antioxidant response in cadmium-stressed sorghum plant. Physiol Mol Biol Plants. 2024, 30, 605–618. [Google Scholar] [CrossRef]
- Aftab, Tariq; Khan, M. Masroor A.; Idrees, M.; Naeem, M.; Moinuddin; Hashmi, Nadeem; Varshney, Lalit. Enhancing the growth, photosynthetic capacity and artemisinin content in Artemisia annua L. by irradiated sodium alginate. Radiation Physics and Chemistry 2011, 80, 833-836. [CrossRef]
- Sarfaraz, Adeeba; Naeem, M.; Nasir, Shafia; Idrees, Mohd; Aftab, Tariq; Hashmi, Nadeem; Khan, M. Masroor; Moinuddin; Varshney, Lalit. An evaluation of the effects of irradiated sodium alginate on the growth, physiological activities and essential oil production of fennel (Foeniculum vulgare Mill.). J. Med. Plant. Res. 2011, 5, 15-21. Available online: https://academicjournals.org/journal/JMPR/article-full-text-pdf/3F390BE15764.
- Zhang, Y.; Yin, H.; Liu, H.; Wang, W.; Wu, L.; Zhao, X.; Du, Y. Alginate oligosaccharides regulate nitrogen metabolism via calcium in Brassica campestris L. var. utilis Tsen et Lee. The Journal of Horticultural Science and Biotechnology 2013, 88, 502-508. [CrossRef]
- Reed, A.J.; Canvin, D.T.; Sherrard, J.H.; Hageman, R.H. Assimilation of [15N] nitrate and of [15N] nitrite in leaves of five plant species under light and dark conditions. Plant Physiol 1983, 71, 291–294. [Google Scholar] [CrossRef]
- Matt, P.; Geiger, M.; Walch-Liu, P.; Engels, C.; Krapp, A.; Stitt, M. Elevated carbon dioxide increases nitrate uptake and nitrate reductase activity when tobacco is growing on nitrate, but increases ammonium uptake and inhibits nitrate reductase activity when tobacco is growing on ammonium nitrate. Plant Cell Environ 2001, 24, 1119–1137. [Google Scholar] [CrossRef]
- Matt, P.; Geiger, M.; Walch-Liu, P.; Engels, C.; Krapp, A.; Stitt, M. The immediate cause of the diurnal changes of nitrogen metabolism in leaves of nitrate-replete tobacco: a major imbalance between the rate of nitrate reduction and the rates of nitrate uptake and ammonium metabolism during the first part of the light period. Plant Cell Environ 2001, 24, 177–190. [Google Scholar] [CrossRef]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol Plant 2010, 3, 973–96. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Long, S.; Ort, D.R. What is the maximum efficiency with which photosynthesis can convert solar energy into biomass? Curr. Opin. Biotechnol 2008, 19, 153–159. [Google Scholar] [CrossRef]
- Nasar, J.; Khan, W.; Khan, M.Z.; Gitari, H.I.; Gbolayori, G.F.; Moussa, A.A.; Mandozai, A.; Rizwan, N.; Anwari, G.; Maroof, S.M. Photosynthetic activities and photosynthetic nitrogen use efficiency of maize crop under different planting patterns and nitrogen fertilization. J Soil Sci Plant Nutr 2021, 21, 2274–2284. [Google Scholar] [CrossRef]
- Wang, J.; Tian, P.; Christensen, M.J.; Zhang, X.; Li, C.; Nan, Z. Effect of Epichloë gansuensis endophyte on the activity of enzymes of nitrogen metabolism, nitrogen use efficiency and photosynthetic ability of Achnatherum inebrians under various NaCl concentrations. Plant Soil 2019, 435, 57–68. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, H.; Wang, W.; Zhao, X.; Du, Y.; Wu, L. Enhancement in photosynthesis characteristics and phytohormones of flowering Chinese cabbage (Brassica campestris L. var. utilis Tsen et Lee) by exogenous alginate oligosaccharides. Journal of Food, Agriculture & Environment 2013, 11, 669-675. Available online: http://pascal-francis.inist.fr/vibad/index.php?action=getRecordDetail&idt=27243252.
Figure 1.
Effects of AOS and different nutrient solution levels on the growth of cucumber seedlings under suboptimal temperature. Bar = 5 cm.(a) plant phenotype; (b) plant height; (c) stem thickness; (d) leaf area; (e) fresh weight of root; (f) fresh weight of stem; (g) fresh weight of leaf; (h) dry weight of root; (i) dry weight of stem; (j) dry weight of leaf. Different lowercase letters indicate significant differences at 0.05 level. The same below.
Figure 1.
Effects of AOS and different nutrient solution levels on the growth of cucumber seedlings under suboptimal temperature. Bar = 5 cm.(a) plant phenotype; (b) plant height; (c) stem thickness; (d) leaf area; (e) fresh weight of root; (f) fresh weight of stem; (g) fresh weight of leaf; (h) dry weight of root; (i) dry weight of stem; (j) dry weight of leaf. Different lowercase letters indicate significant differences at 0.05 level. The same below.
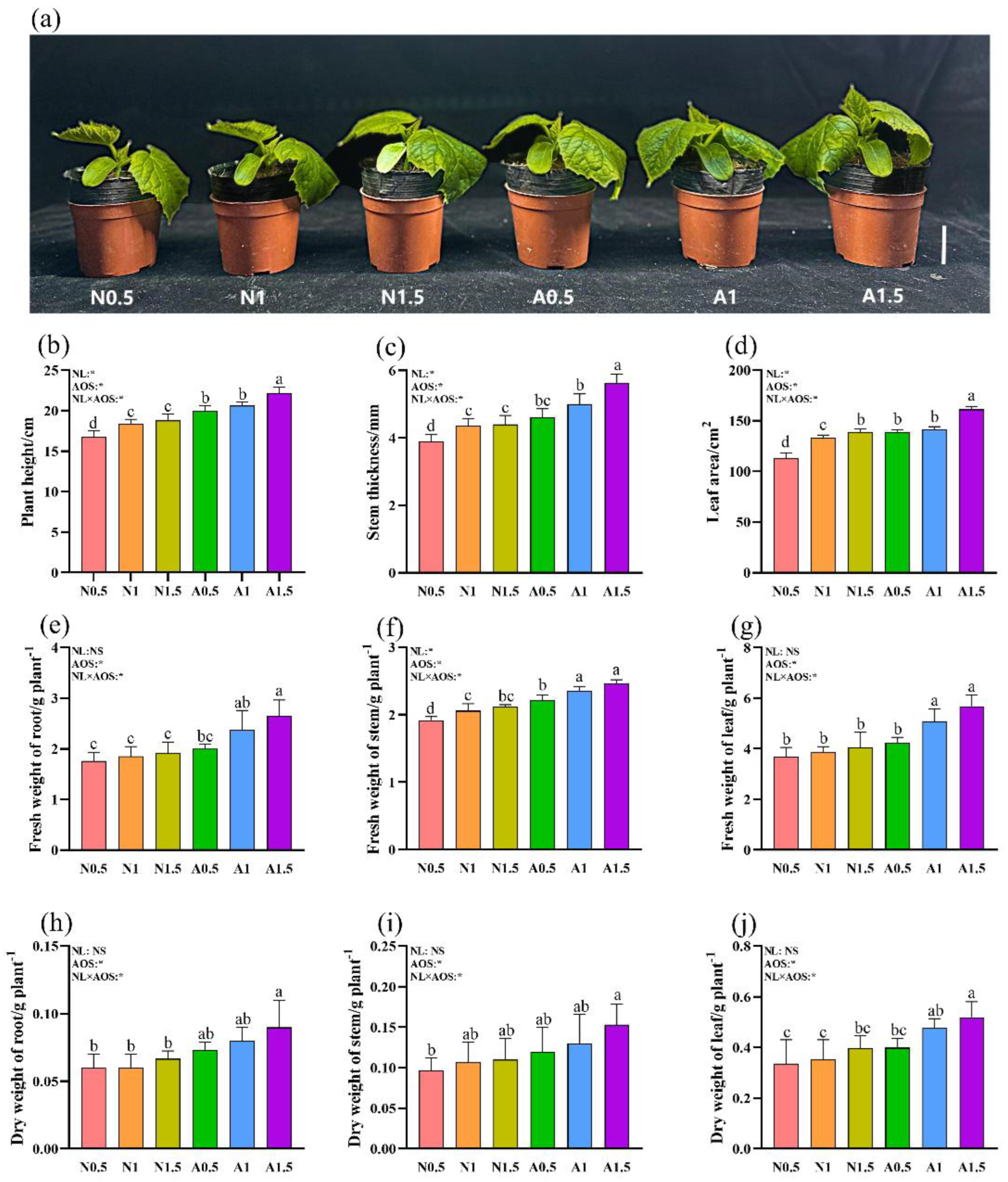
Figure 2.
Effects of AOS and different nutrient solution levels on the root morphology of cucumber seedlings under suboptimal temperature. (a) root tip number; (b) root average diameter; (c) total root length; (d) root forks; (e) root surface area; (f) root volume.
Figure 2.
Effects of AOS and different nutrient solution levels on the root morphology of cucumber seedlings under suboptimal temperature. (a) root tip number; (b) root average diameter; (c) total root length; (d) root forks; (e) root surface area; (f) root volume.
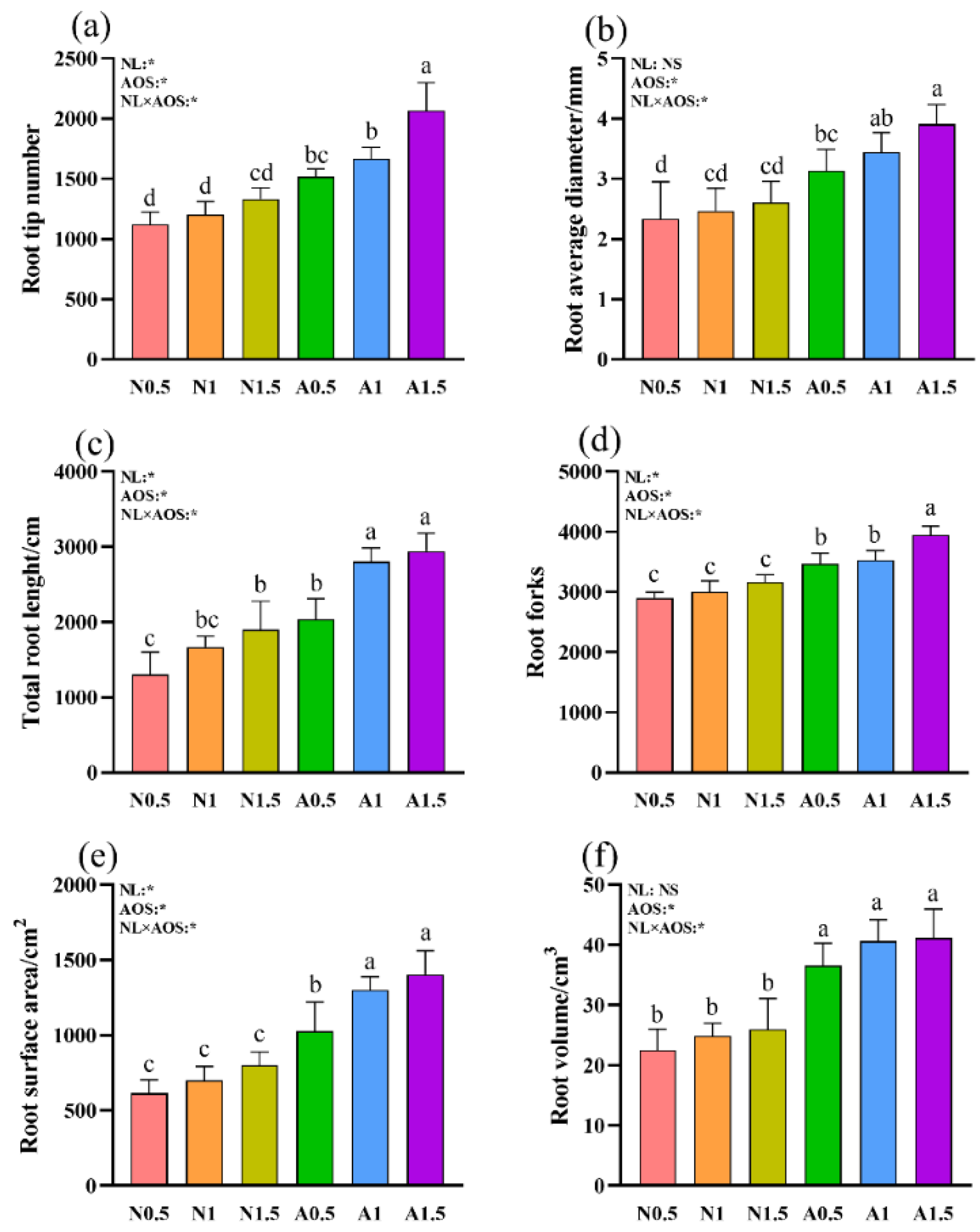
Figure 3.
Effects of AOS and different nutrient solution levels on chlorophyll content and net photosynthetic rate of cucumber seedlings under suboptimal temperature. (a) chlorophyll a content; (b) chlorophyll b content; (c) net photosynthetic rate.
Figure 3.
Effects of AOS and different nutrient solution levels on chlorophyll content and net photosynthetic rate of cucumber seedlings under suboptimal temperature. (a) chlorophyll a content; (b) chlorophyll b content; (c) net photosynthetic rate.
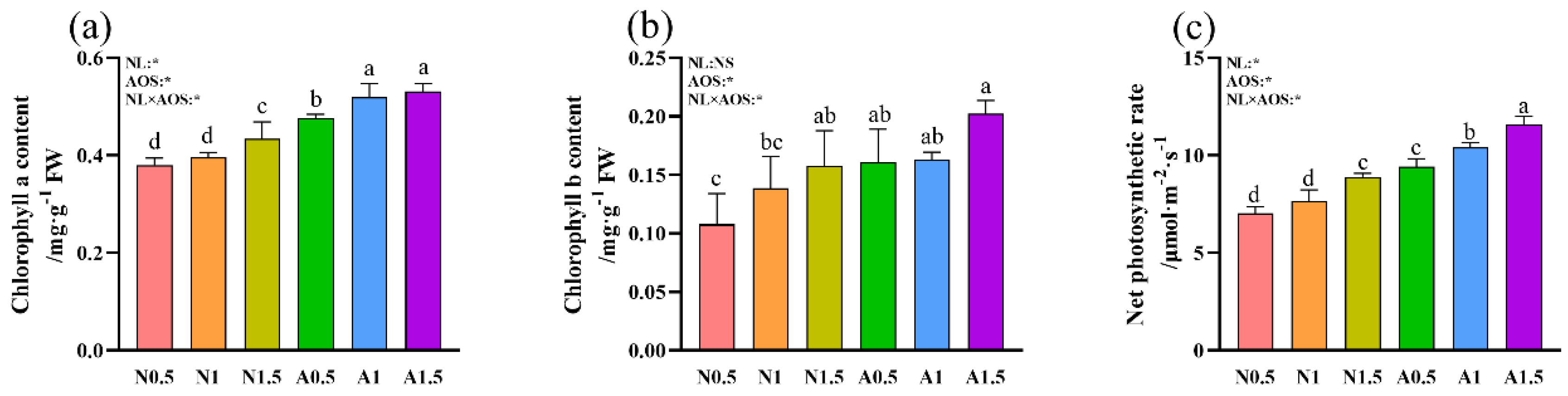
Figure 4.
Effects of AOS and different nutrient solution levels on root activity of cucumber seedlings under suboptimal temperature.
Figure 4.
Effects of AOS and different nutrient solution levels on root activity of cucumber seedlings under suboptimal temperature.
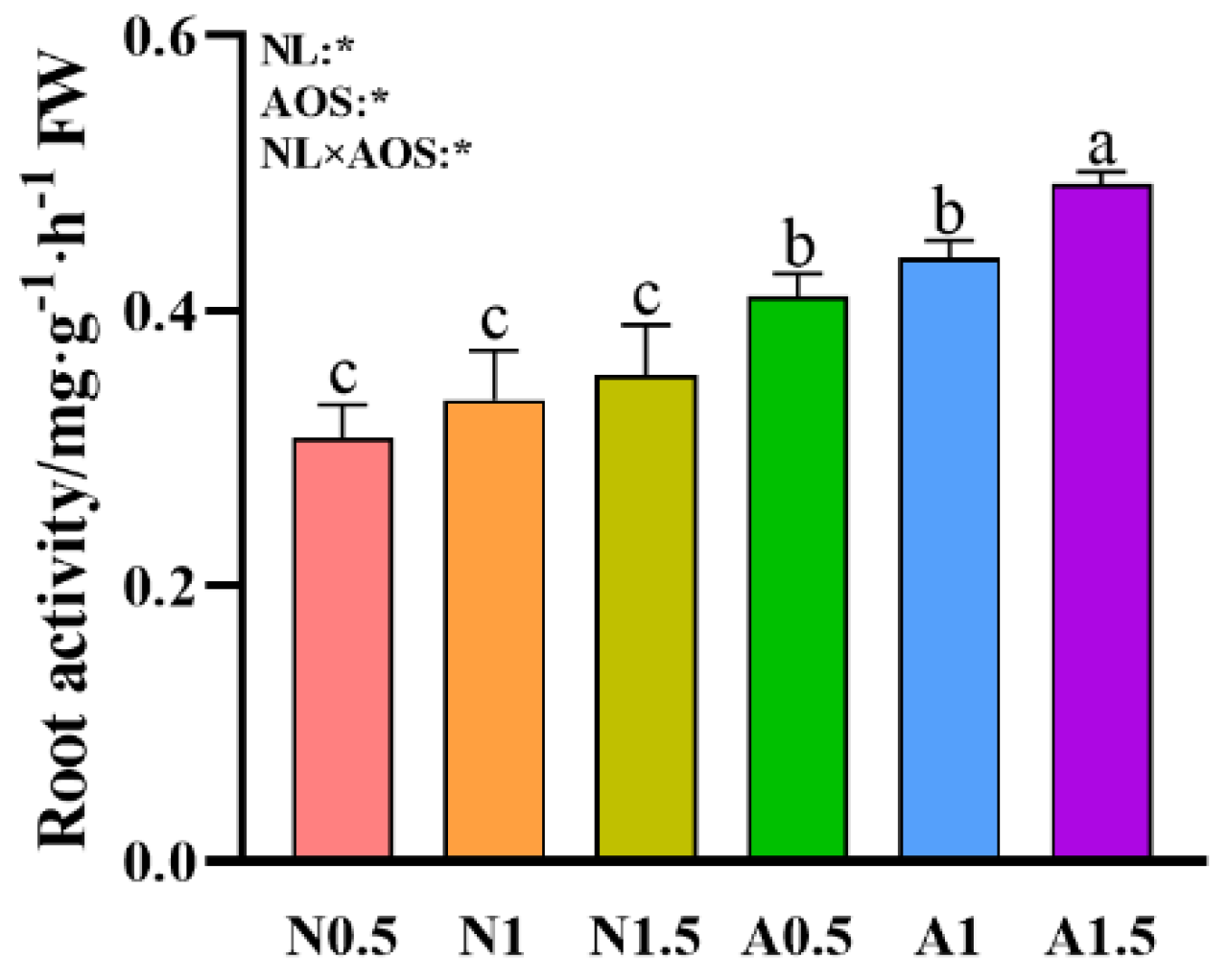
Figure 5.
Effects of AOS and different nutrient solution levels on the nitrogen metabolism enzymes activities in cucumber seedlings' leaves and roots under suboptimal temperature. (a) NR activity in leaf; (b) NR activity in root; (c) GS activity in leaf; (d) GS activity in root. (e)GOGAT activity in leaf; (f) GOGAT activity in root; (g) GDH activity in leaf; (h) GDH activity in root;.
Figure 5.
Effects of AOS and different nutrient solution levels on the nitrogen metabolism enzymes activities in cucumber seedlings' leaves and roots under suboptimal temperature. (a) NR activity in leaf; (b) NR activity in root; (c) GS activity in leaf; (d) GS activity in root. (e)GOGAT activity in leaf; (f) GOGAT activity in root; (g) GDH activity in leaf; (h) GDH activity in root;.
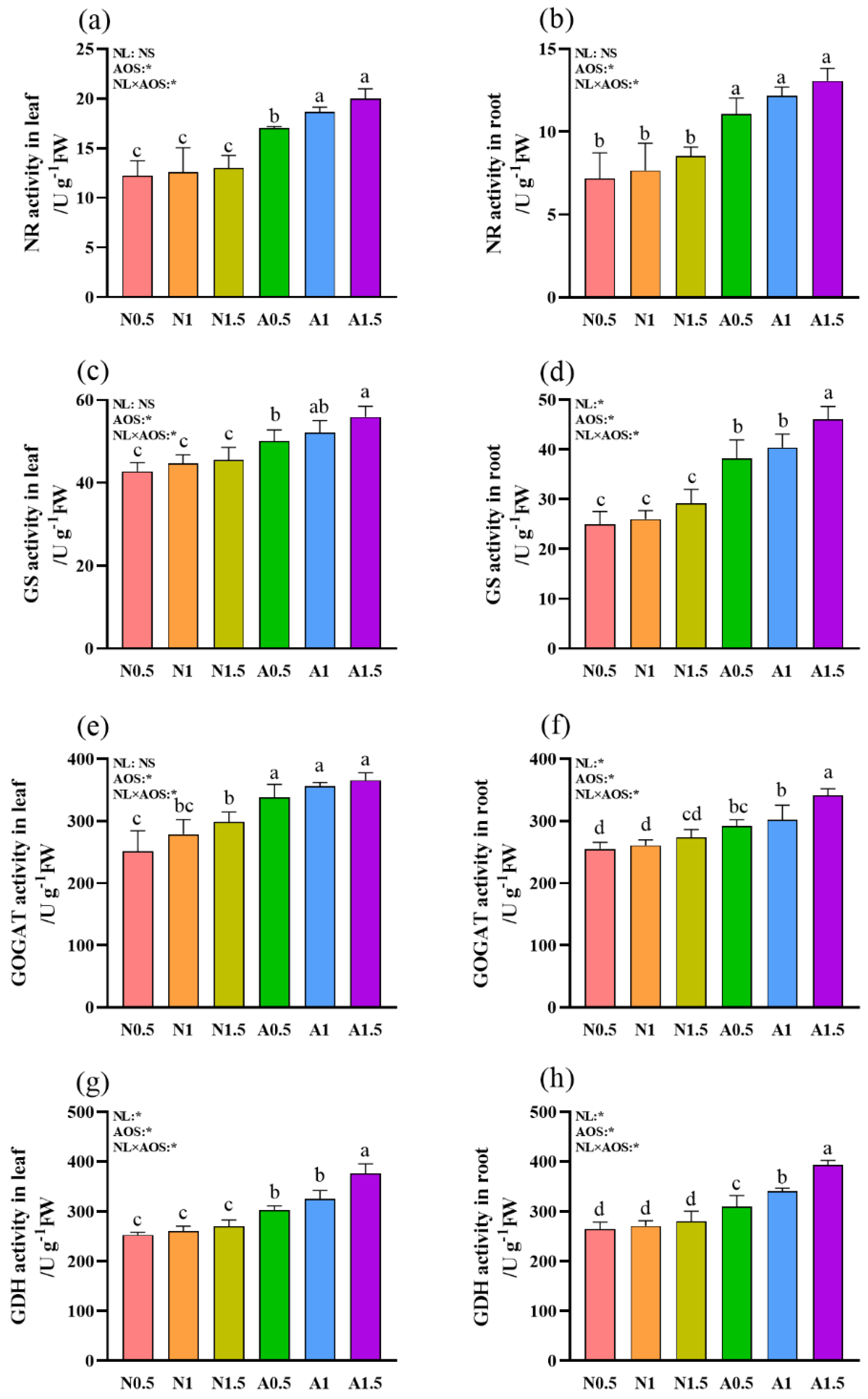
Figure 6.
Effects of AOS and different nutrient solution levels on nitrogen content, accumulation, and distribution in different cucumber organs under suboptimal temperature. (a) N content; (b) N accumulation; (c) N matter ratio.
Figure 6.
Effects of AOS and different nutrient solution levels on nitrogen content, accumulation, and distribution in different cucumber organs under suboptimal temperature. (a) N content; (b) N accumulation; (c) N matter ratio.
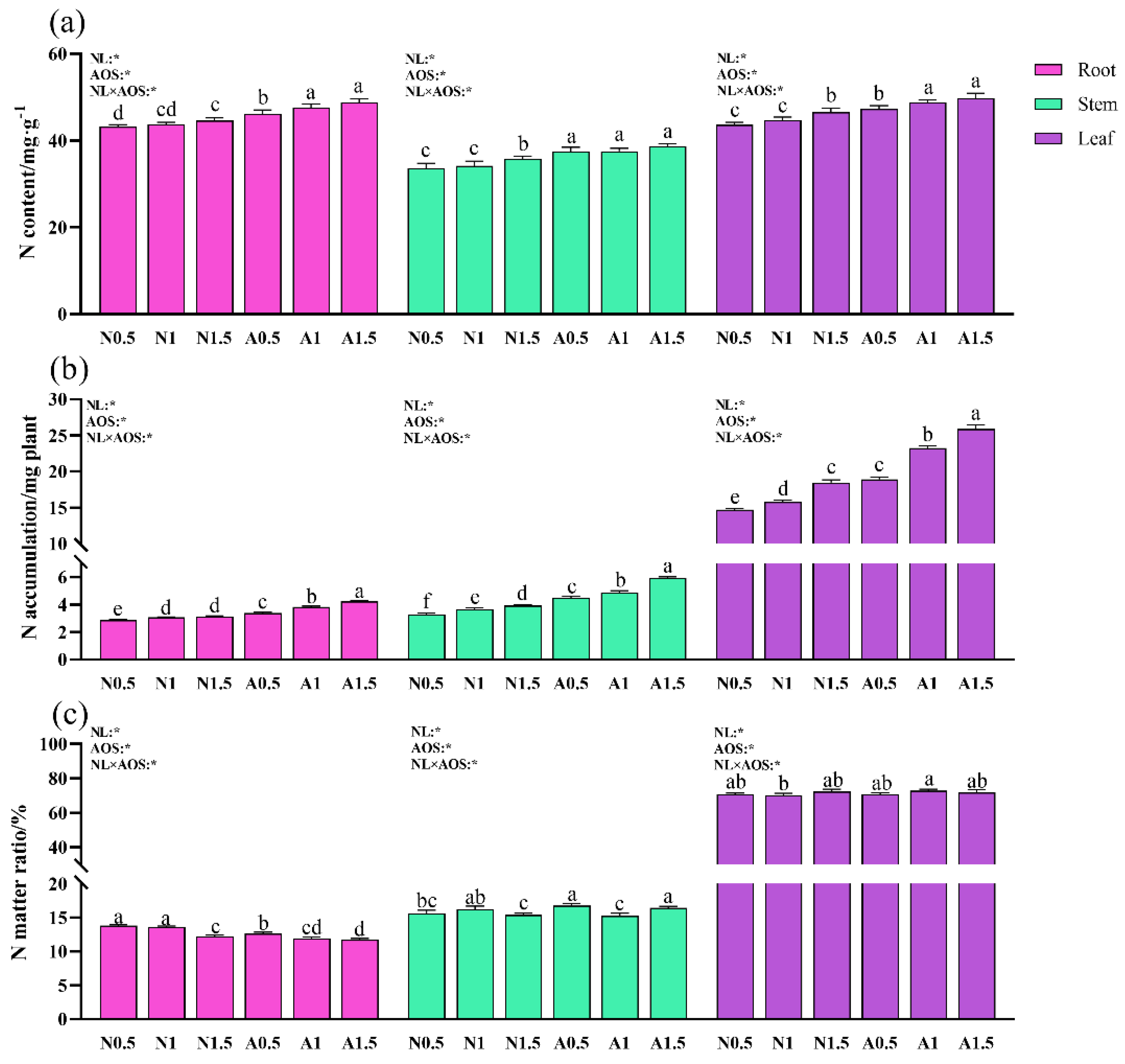
Figure 7.
Correlation analysis between physiological and biochemical parameters of cucumber seedlings (n=18). NCL-R, nitrogen content in root. NCL-S, nitrogen content in stem. NCL-L, nitrogen content in leaf. Chl. a, chlorophyll a content. Chl. b, chlorophyll b content. Pn, net photosynthetic rate. RTN, root tip number. RAD, root average diameter. RL, total root length. RF, root forks. RSA, root surface area. RV, root volume. RA, root activity. NR-R, nitrate reductase activity in root. GOGAT-R, glutamate synthase activity in root. GDH-R, glutamate dehydrogenase activity in root. GS-R, glutamine synthetase in root. NR-L, nitrate reductase activity in leaf. GOGAT-L, glutamate synthase activity in leaf. GDH-L, glutamate dehydrogenase activity in leaf. GS-L, glutamine synthetase in leaf. *, ** and *** indicate the statistical significance at 0.05, 0.01 and 0.001 levels, respectively.
Figure 7.
Correlation analysis between physiological and biochemical parameters of cucumber seedlings (n=18). NCL-R, nitrogen content in root. NCL-S, nitrogen content in stem. NCL-L, nitrogen content in leaf. Chl. a, chlorophyll a content. Chl. b, chlorophyll b content. Pn, net photosynthetic rate. RTN, root tip number. RAD, root average diameter. RL, total root length. RF, root forks. RSA, root surface area. RV, root volume. RA, root activity. NR-R, nitrate reductase activity in root. GOGAT-R, glutamate synthase activity in root. GDH-R, glutamate dehydrogenase activity in root. GS-R, glutamine synthetase in root. NR-L, nitrate reductase activity in leaf. GOGAT-L, glutamate synthase activity in leaf. GDH-L, glutamate dehydrogenase activity in leaf. GS-L, glutamine synthetase in leaf. *, ** and *** indicate the statistical significance at 0.05, 0.01 and 0.001 levels, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



