
Submitted:
08 April 2025
Posted:
09 April 2025
You are already at the latest version
Abstract
The tomato russet mite (TRM) Aculops lycopersici, a destructive pest of tomato crops worldwide, is a significant challenge to growers, both in greenhouse and open-field conditions. Traditional chemical control methods are often ineffective and set up resistance and adverse environmental impact. This has prompted the exploration of alternative control methods, such as biological control and eco-friendly botanical pesticides. In this study, the acaricidal effects of essential oils (EOs) extracted by three officinal plants, Origanum vulgare L., Salvia rosmarinus Spenn., and Salvia officinalis L., cultivated using precision aromatic crop (PAC) techniques was evaluated against A. lycopersici under laboratory conditions. The chemical composition of EOs was ascertained by a solid-phase microextraction (SPME) coupled with mass spectrometry (GC-MS) analyses. Carvacrol (83.42%), ρ-Cymene (3.06%) and γ-Terpinene (2.93%) were the major components of O. vulgare, while α-Pinene (28.0%), 1,8-Cineole (11.00%) and Borneol (7.72%) were present in S. rosmarinus. 1,8-Cineole (27.67%) was the major constituent of S. officinalis EO, followed by Camphor (21.91%) and Crisantenone (12.87%). Multiple concentrations (320–5000 μL L⁻¹) and exposure times (1 to 4 days) were tested to assess mortality rates. Results showed both dose and time-dependent toxic activity with significant differences among EOs. Oregano EO was found to be the most toxic of the EOs (90% of mortality at 0.5% w/v concentration after 4 days), while rosemary and sage EOs showed limited effects (46% and 42% for the latter EOs respectively). The lethal concentration (LC50) values were 2,228 μL L⁻¹ for oregano, 5,835 μL L⁻¹ for rosemary, and 6,013 μL L⁻¹ for sage, demonstrating ef-ficacy similar to commercially available botanical pesticides. These findings support the potential of O. vulgare EO as a viable alternative for controlling A. lycopersici, contributing to integrated pest management (IPM) strategies, and highlight the need for further re-search for discovering botanical agents for an eco-friendly pest control.
Keywords:
Medicinal and Aromatic Crops (MAPs)
; precision agriculture
; essential oils
; tomato russet mite
1. Introduction
The tomato russet mite (TRM), Aculops lycopersici (Tryon, 1917) (Acari: Eriophyidae), is a worldwide distributed pest, present in both tropical and temperate regions [1,2]. It infests mainly solanaceous plants, including wild species like black nightshade Solanum nigrum L. as well as cultivated crops such as pepper (Capsicum annuum L.), eggplant (Solanum melongena L.), potato (Solanum tuberosum L.), and tomato (Lycopersicon esculentum Miller). Aculops lycopersici is especially damaging on tomato cultivation, affecting yields in greenhouses and open-field crops. The eriophyid feeds on leaf, stem, and fruit surfaces, resulting in significant cellular disruption. Mite feeding damages the adaxial and abaxial epidermal cells of leaves, causing the development of a dense layer of callous tissue near the parenchyma where cell death occurs [3]. The feeding behavior of A. lycopersici causes russeted leaves, stems and fruits and, in severe cases, can lead to plant death [4]. Recent studies suggest that A. lycopersici, similar to other eriophyid mites, may also transmit virus vector, increasing its overall negative impact on crops [5].
Thriving in warm, arid conditions, A. lycopersici populations can escalate rapidly, causing substantial damage. These mites typically cluster and feed where they initially land on the plant, only spreading upward once the population density increase. Infestations remain often undetected until visible symptoms emerge; at this point a large population has already built up, especially on drought-stressed plants that accelerate mite reproduction [6]. Recently, infestations by A. lycopersici have surged in European tomato crops, echoing a broader trend of increasing economic impact from eriophyoid mites globally [7].
Synthetic acaricides have long been the standard approach for controlling phytophagous mites; however, the development of resistance, along with harmful effects on native phytoseiid populations, has raised significant concerns [8]. These challenges have prompted increasing interest in biological alternatives for integrated pest management (IPM), as researchers seek more sustainable and environmentally friendly solutions.
Efforts to identify natural enemies for controlling tomato russet mites have led to a substantial list of potential predators, particularly within the Phytoseiidae family [9,10,11,12,13]. Several species of these predatory mites have been observed in natural association with tomato russet mites, able of feeding and reproducing on them, at least under laboratory conditions [14,15]. However, when tested under field conditions, predators often failed to suppress the eriophyid population or required several generations for adapting to tomato plants, limiting their impact towards the pest population [10,11,16,17,18]. Moreover, the effectiveness of these predators can be further hindered by trichome density and pollen availability [19]. Tomato trichomes hamper predator movement and provide shelter for russet mites, while toxic secondary metabolites of plants and prey can intoxicate the predators, making biological control even more complicated [7,19].
Botanical pesticides are receiving renewed attention as a potentially cost-effective alternative to synthetic chemicals in integrated pest control programs (IPM) [20]. They were commonly used before World War II, but their role diminished with the introduction of synthetic alternatives [20,21,22,23].
In contrast, botanical products usually have fewer or less severe negative effects on human health and the environment, support natural enemies of pests, and exhibit low environmental persistence [24]. Among the botanical solutions, essential oils (EOs) and their active compounds have emerged as promising biocides, particularly for their efficacy against arthropod pests, including tephritid flies, ambrosia beetles, mites, ticks, and even certain weeds [25,26]. The large-scale production of plant derived EOs for the perfume and flavouring industries also makes these substances commercially viable [26]. Current research on botanical pesticides frequently investigates plant families such as Apiaceae, Myrtaceae, Lamiaceae, Meliaceae, Annonaceae, Simaroubaceae, and, more recently, Asteraceae [23,27].
The present study aimed to assess the acaricidal effects of essential oils from Origanum vulgare L., Salvia rosmarinus Spenn., (= syn. Rosmarinus officinalis L.) and Salvia officinalis L. (Lamiaceae) cultivated using PAC techniques, on A. lycopersici under laboratory conditions.
2. Results
2.1. Precision Aromatic Crop (PAC) Techniques
During the spring season, oregano was in its growth phase, while rosemary remained bare and sage exhibited low vigour. On September, oregano had been harvested, leaving bare soil with visible clumps, whereas rosemary displayed medium vegetative vigour and sage reached full vigour. On October, sage maintained full vigour, and rosemary continued developing, with vigour classes increasing to 0.60–0.80 (oregano had resumed growth following the rains). More than 50% of the sage surface showed NDVI values greater than 0.40 (Figure 1 and Figure 2).
Multispectral analysis enabled the identification of the optimal harvest period, corresponding to mean NDVI values of 0.85 for rosemary (in May), 0.80 for oregano (in June), and 0.75 for sage (in May and October 2024).
2.2. Analyses of Essential Oils
The essential oils extracted from the three species had a light-yellow colour and a characteristic aroma. The volatile profile of Oregano EO, includes 27 volatile compounds belonging to the following phytochemical groups: monoterpene phenol (84.31%), monoterpene hydrocarbons (9.53%), Oxygenated monoterpenes (1.91%), sesquiterpene hydrocarbons (2.87%), ethers, ketones, and alcohols each contributing less than 1%. The principal component of this EO was Carvacrol, followed by ρ-Cymene and γ-Terpinene. Nineteen main components representing 89.3% of the Rosemary essential oil were detected: 47.53% of Monoterpene hydrocarbons, 33.37% of Oxygenated monoterpenes, and 8.27% of Sesquiterpene hydrocarbons (Table 1). As shown in Table 1, the sage EO yield was 0.29%, a total of 23 volatile compounds were identified by GC-MS. These compounds were categorised into monoterpene hydrocarbons (29.64%), oxygenated sesquiterpenes (2.81%), oxygenated monoterpenes (59.57%), and sesquiterpene hydrocarbons (2.72%). Additionally, other compounds not classified in the previous categories were also identified. Monoterpene hydrocarbons and oxygenated monoterpenes were the most abundant compounds in all sage leaf samples, accounting for more than 90% of the total identified compounds. Among the monoterpene hydrocarbons, camphene and crisantenone were the most prevalent (average of 9.26% and 12.87%, respectively), while eucalyptol and camphor were the dominant compounds among the oxygenated monoterpenes (average of 27.67% and 21.91%, respectively) [28].
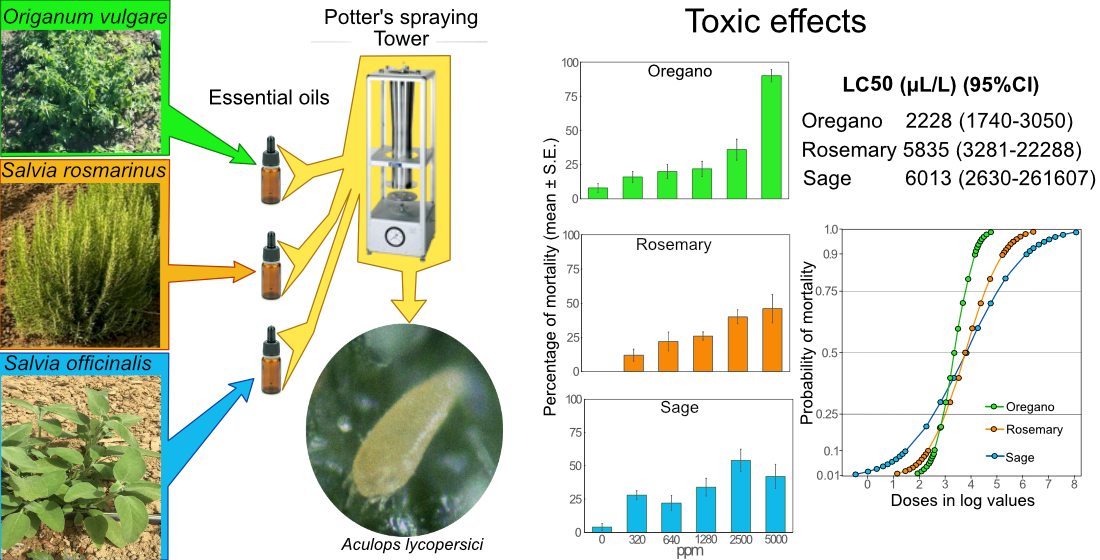
2.3. Toxicity of O. vulgare, S. rosmarinus, and S. officinalis EOs Against A. lycopersici
The three factors compared in the GLM analysis were EO (Oregano, Rosemary, Sage), Concentration (0, 320, 640, 1280, 2500, 5000) and Time (1, 2, 3, 4 days). The three EOs tested affected in a different way A. lycopersici adults (F2, 648=5.75, p=0.003), and a significant different effect was registered both for the factor Concentration (F5, 648=124.45, p<0.001) and Time (F3, 648=61.67, p<0.001). The interaction between the first two factors (F6, 648=18.42, P<0.001) and between Concentration and Time (F15, 648=2.14, p=0.007) indicate that each EO concentration caused different toxic effects, and that the mortality had a different trend in the three adopted EOs during the test period.
The highest concentration of Oregano EO (5000 μL L⁻1) hardly affected A. lycopersici after 24 hours (66% of mortality), and high mortality (90%) after 4 days was registered, while it was significantly lower with concentration of 2500 μL L⁻1 and almost null in all the remaining concentrations (Table 2). For Rosemary and Sage, the acaricidal effect remained scarce, with marginal increases during time even at the highest concentrations. No statistical differences were noted between each considered concentration for the latter extracts. As a matter of fact, only the highest concentration of oregano EO can be classified as highly toxic (class 4) according to the toxicity categories proposed by Hardman et al. [29]. At the concentration of 2500 μL L⁻1, the effect was moderate (class 2), while at lower concentrations (1280, 640, 320 μL L⁻1) the efficacy was classified as non-toxic (Class 1). Rosemary and Sage EOs caused similar results, falling in classes 1 and 2 (Table 2).
The analysis of mean survival times further corroborated these trends, indicating that at the highest concentration of 5000 μL L⁻1 of oregano EO, the mean survival time of A. lycopersici adults was about one day after treatment. A higher survival time was recorded using 2500 μL L⁻1 of the latter EO, although it was statistically lower compared to the control (Table 2). In contrast, survival times at lower concentrations did not differ to those observed in the control tests. This highlights that the acaricidal efficacy of oregano EO is evident only surpassing the critical concentration threshold of 2500 μL L⁻1.
The lethal concentrations of the three essential oils were calculated by probit analysis. The Pearson goodness of fit test indicated that LC values obtained by oregano EO, does not fit the linear model (χ² = 21.62; p< 0.001), because of the high discrepancies of toxic effects registered at the highest dose (90% of mortality) and the lower doses (less than 36% of mortality). On the other hand, LC values calculated for the other two EOs well fitted to the linear model (Table 3).
Different letters indicate significant differences among extracts and concentrations for survival time and corrected mortality. Tukey’s multiple comparison tests (p<0.05) were applied after GLM analysis. * Toxicity classes were defined on corrected mortality after Hardman et al. [29].
The LC50 values were 2228.90 μL L⁻1 for oregano EO, 5835.39 μL L⁻1 for rosemary EO, and 6013.81 μL L⁻1 for sage EO. The probability plot built on probit data indicates a more sensitive response for mortality in oregano EO doses, in comparison to the other two EOs (IQR=0.864). The probability of 50% of mortality was almost the same for rosemary and sage EOs but the different values of IQR (1.526 and 2.478 for the latter EOs respectively) indicate a more immediate effect of rosemary EO than of the sage one (Figure 3).
3. Discussion
The three Lamiaceae species used in the present study are wide-spread in the Mediterranean basin and used from millennia for their therapeutic and culinary properties [30,31]. Moreover, the three species are included in the BELFRIT list [32], making easier a potential future biopesticide registration.
Oregano EO is mainly composed by carvacrol, thymol and monoterpenes [33]. However, the predominant compound identified in this study, was carvacrol, accounting for about 83% of the total composition. This confirms monoterpenoids as the dominant class of volatile organic compounds (VOCs) in oregano EO. This composition places this EO within the carvacrol oregano chemotype, primarily due to its high carvacrol content (~80%). Similar volatile profiles have been reported for oregano from other Mediterranean regions, suggesting that the essential oil composition is more strongly influenced by the oregano variety than by the geographical origin of cultivation [34].
Recent research has shown that carvacrol, along with other monoterpenoids, is highly toxic to a range of invertebrate pests, including insects, acari, and nematodes [35,36,37]. Furthermore, these compounds are environmentally friendly, as they biodegrade or dissipate quickly, and exhibit low toxicity to mammals, fish, and other non-target organisms [38]. These properties make monoterpenoids like carvacrol a viable alternative to synthetic pesticides for pest management.
The percentage composition of key components in rosemary EO indicates that α-pinene, 1,8-cineole and borneol are the major constituents, as previously reported [39]. Furthermore, other researchers have noted that each rosemary EO typically contains nine major compounds, collectively accounting for over 90% of its composition, with 1,8-cineole being the predominant component (>52%) [40].
From a chemical perspective, rosemary EO can be classified into distinct chemotypes based on the relative abundance of its primary constituents. The main chemotypes identified include cineoliferum, characterised by a high 1,8-cineole content, camphoriferum, with camphor levels exceeding 20%, and verbenoniferum, where verbenone exceeds 15%. Additionally, chemotypes with high levels of α-pinene have been identified in specific regions, including Italy and Morocco [41].
The biological activity of α-pinene has been extensively investigated. In addition to its well-documented antifungal, antibacterial, and antiviral properties, α-pinene exhibits insecticidal and nematocidal effects [42], underscoring its potential for various control activities. According to a review by Jankowska et al. [43], α-pinene was found to be one of the most effective volatile organic compounds (VOCs) in inhibiting acetylcholinesterase (AChE), which may explain the higher entomotoxic activity of rosemary EO, in which α-pinene is generally the second most abundant compound. Moreover, several other constituents of rosemary EO, such as camphor, eucalyptol, and α-pinene, have been reported to exert a cytotoxic mode of action, leading to cell membrane damage [44,45].
The Salvia genus comprises about 900 species mainly distributed throughout the world, and some of which are used also in perfumery and cosmetics. The essential oil of many species of sage is characterized by the presence of 1,8 cineole (eucalyptol), β-thujone, camphor, borneol, and ρ-cymene which is attributed with the antimicrobial activity against many microorganisms [46,47].
The insecticidal efficacy of sage EO is attributed to its richness in monoterpenoids, which are widely recognised for their potent insecticidal effects against a broad spectrum of insects. For example, 1,8-cineole and α-pinene, two key monoterpenoid constituents of this EO, have been shown to inhibit erythrocyte acetylcholinesterase activity [48]. Moreover, it has been demonstrated that sage EO exhibits significant insecticidal activity on Spodoptera littoralis Boisduval, with mortality increasing proportionally to the concentration after 24 hours of exposure [49]. It has also been demonstrated that essential oils of sage EO possess both contact toxicity and repellent effects on Tetranychus urticae. Generally, the terpenoids in essential oils exert various effects on insects, including toxicity, reduced maturity, and diminished reproductive capacity. These oils are also a complex mixture of neurotoxic compounds with acute effects on insects, acting by interfering with the octopaminergic transmitters in arthropods. The action of these essential oils is likely due to the synergy or antagonism between the major compounds [50].
Interest in the use of plant extracts and essential oils as tools for controlling phytophagous populations has recently increased considerably, due to their ability to reduce environmental impact and preserve non-target organisms [24,26,51]. However, research has mainly focused on a limited number of phytophagous species, neglecting others that, although less studied, may be just as harmful or even more damaging. Regarding Acari, studies on vegetal products have mainly focused on T. urticae [23,52,53,54], while the knowledge available on the effectiveness of biopesticides against other damaging pests such as the tomato rust mite is still very limited. To the best of our knowledge, this is the first study on the effects of essential oils against A. lycopersici.
Among the tested oils, oregano EO exhibited the highest acaricidal activity. However, significant efficacy was observed only at the highest concentration (5000 µl/L), suggesting that its toxic effect manifests only beyond a specific concentration threshold. Probably, the main toxic effect of oregano EO is attributed to carvacrol, as it exhibits various bioactive properties, including antioxidant effects, inhibition of antibiotic-resistant bacteria, suppression of microbial and fungal toxins, and potential anticancer activity [34]. However, the primary mode of action of carvacrol remains unclear, although it has shown limited acetylcholinesterase inhibition in certain insect species, such as house flies, ticks, and cockroaches [48].
Regarding rosemary and sage EOs, Laborda et al. [53] assessed high toxic effects after 24 h exposure (79 to 100% of mortality), against T. urticae at concentrations ranging from 1500 to 2500 µl/L. Our results on A. lycopersici revealed very low toxicity after 24 h (2 to 12%) and also after 4-days exposure (26 to 54% of mortality), at comparable concentrations (1280 and 2500 µl/L). However, the sage EO used by the abovementioned authors had a different chemical profile: the main component was α-Thujone (42.3%), followed by Camphor (11.0%) and 1,8-Cineole (10.3%); the latter two were the principal components of our sage EO (21.91 and 27.67% for the two components respectively), while Camphor was not detected in our sage EO. The rosemary EO used by Laborda et al. [53] showed also a different compound composition to that used in our experiments. The four main components of their rosemary EO were 1,8-cineole, α-pinene, camphor and camphene (26.7, 18.6, 17.5 and 11.8 for the four compounds respectively). However, concentration of the above compounds in our rosemary EO was quite different (11.0, 28.0, 6.2 and 7.0 for the above components respectively). The lower toxic effects on A. lycopersici could be attributed to the different concentrations of the components of the essential oils adopted but also to a different susceptibility of the two mite species.
4. Materials and Methods
4.1. Cultivation of Officinal Plants with Precision Aromatic Crops (PAC) Techniques, and Essential Oil Extraction Methods
4.1.1. Plants Cultivation Method
Cultivation of O. vulgare (Oregano), S. rosmarinus (Rosemary), and S. officinalis (Sage) carried out at Morreale’s Farm in Grotte (Agrigento Province, Italy) (37°22'52.284"N 13°40'24.067"E) (World Geodetic Coordinate System 1984). The soil moisture regime is xeric, bordering on aridic, and the temperature regime is thermic. The experimental field includes 1,520 plants of O. vulgare, 1,980 plants of S. rosmarinus and 2,485 plants of S. officinalis, arranged with 35 cm spacing along the rows and 180 cm between the rows. Periodically, inter-row surface tillage was performed to control weed growth and disrupt soil capillarity. No organic or mineral fertilization was carried out during the growth period.
4.1.2. Precision Aromatic Crop (PAC) Techniques
Precision Aromatic Crop (PAC) techniques were applied to optimize the cultivation and monitoring of Medicinal and Aromatic Plants (MAPs). Specifically, unmanned aerial vehicles (UAVs) equipped with multispectral cameras, combined with post-processing software, have become a widely adopted technique for assessing vegetation indices (VIs) in the management of MAPs. This method seeks to modernize agricultural practices by minimizing resource use and boosting productivity [55,56,57].
Advanced technologies for spatially variable crop condition monitoring, focusing on the use of UAVs equipped with multispectral cameras and a spectroradiometer to assess MAPs were adopted.
The DJI Mavic 3 Multispectral (M3M) drone was used to capture high-resolution images across visible and near-infrared bands, including Green (G), Red (R), Red Edge (RE), and Near-Infrared (NIR) (Figure 4). The Hand Held 2 ASD spectroradiometer used in the tests have a sensitivity in the region 325-1075 nm. A solar irradiance sensor and a GNSS system with RTK correction (< 2 cm accuracy) ensure precise data collection. Flight parameters were carefully planned to use DJI GS Pro software to avoid issues like shadows, with flights scheduled at noon for optimal lighting conditions in the period from March to October 2024.
Ground Control Points (GCPs) were placed and georeferenced using an RTK-enabled GNSS receiver. After image collection, data were processed in Agisoft Metashape Professional to create a multiband ortho-mosaic, which was then calibrated and orthorectified. Spectral canopy data were extracted and analysed using QGIS software, calculating NDVI values to monitor vegetation health and optimize harvest timing for MAPs [55,56,57]. To assess vegetation dynamics and health, NDVI values were calculated for three key periods: March, September, and October, to identify vegetation areas. Zonal statistics in QGIS were used to determine the surface area corresponding to different NDVI classes, providing valuable data for effective land management. Spectral data also facilitated the generation of false-colour images, enhancing the visualisation of vegetation and offering critical insights into the health and vigour of rosemary, oregano, and sage. The collected biomass was intended for the extraction of EOs process, so it was necessary to identify the time of highest vegetative vigor for MAPs. The NDVI index is sensitive toward crop biophysical properties, like nitrogen, chlorophyll, vigor, biomass etc. In order to choose the optimal harvest time, the MAPs NDVI values were calculated in the period from 1st March and 30th October 2024 (Figure 5)
4.1.3. Extraction and Analyses of Essential Oils
The oregano EO was extracted from 500 g of dried leaves and flowers [55,56,58], using a 12 L conventional steam distiller for aromatic plants (Spring Extractor, Albrigi Luigi, Verona, Italy). After the distillation process, the EO was separated from its vegetation water and stored at 4°C. Yield of EO was expressed in percentage (volume/weight).
Volatile organic compounds (VOCs) in the oregano EO were analyzed using solid-phase microextraction (SPME) coupled with GC-MS, with a dilution of the oil in hexane at a ratio 1:100. The SPME fiber (DVB/CAR/PDMS, 50 mm, Supelco) was exposed to the diluted oils while stirring at 60°C. After 5 minutes of extraction, the fiber was placed in a GC splitless injector, and the VOCs were desorbed for 1 minute at 250°C. Chromatographic separation was achieved using a DB-624 capillary column (Agilent Technologies, 60 m length, 0.25 mm diameter, and 1.40 μm film thickness). The oven temperature was initially set at 40°C for 5 minutes, then increased linearly by 5°C per minute up to 200°C, where it was held for 2 minutes. Helium was used as the carrier gas at a flow rate of 1 mL/min, and the interface temperature was set to 230°C. Mass spectra were recorded in the m/z range of 40-400 amu using full-scan acquisition mode. Individual VOCs were identified by comparing the mass spectra with the NIST05 commercial library. The analysis was performed in triplicate, and the results are expressed as percentages relative to the most significant peak [34].
The extraction of EO from Rosemary is the same as that used for oregano, using 300 g of fresh plant material consisting mainly of flowering tops and leaves. After 3 hours of distillation, a light yellow EO with camphor odor was obtained. Chemical composition was determined by a GC/MS method using an Agilent 6890 Gas-Chromatography coupled with an Agilent 5973 Mass Spectrometer equipped with silica capillary column HP5-MS (30 m x 0.25 mm, film thickness 0.25 mm). The oven temperature was held at 60°C for 8 min, increased to 180°C with a gradient of 4°C/min, and then held for 2 min at 180°C. The components were identified by helium as carrier gas (1 ml/min) and injector temperature and ion-source temperature were 250°C and 280°C, respectively. The identification of Rosemary EO compounds was made by comparison with their relative retention time (RT) with those of original samples or by comparison with their relative retention index (RI) to the series of n-hydrocarbons and computer matching against commercial library and homemade library mass spectra made up from pure substances and components of known oils and MS literature data (NIST90). The Kovats index was calculated agreed with that reported by Adams [59].
For sage EO extraction and analysis, 200 g of leaves were manually chopped into small pieces and subjected to water distillation for three hours using a Clevenger apparatus. The extracted EO was dried with anhydrous sodium sulfate and stored in the dark at 4°C until analysis. The EO was analyzed using a gas chromatograph-mass spectrometer (Shimadzu GC-MS QP2010 Ultra), following the method outlined by Zito et al. [60]. The GC-MS system was equipped with an AOC-20i self-injector (Shimadzu, Kyoto, Japan) and a ZB-5 column (5% phenyl-polysiloxane; 30 m length, 0.32 mm internal diameter, and 0.25 µm film thickness, Phnomenex). For each analysis, 1.3 µL of the sample was injected at 280°C in a 1:1 ratio, with a helium carrier gas flow rate set to 3 mL/min. The oven temperature was held at 60°C for 1 minute, then increased at a rate of 10°C/min until reaching 300°C, where it was maintained for 5 minutes. The MS interface was set to 300°C, and the ion source operated at 200°C. Mass spectra were recorded at 70 eV (EI mode) from 30 to 450 m/z. The GC-MS data were processed using the GC-MS Solution software, version 4.11 [28]. The yields of EOs extracted from the species under study were expressed as a percentage (volume/weight)
4.2. Aculops lycopersici Experimental Set-Up
4.2.1. Solanum nigrum Seedlings for A. lycopersici Breeding
Seeds of S. nigrum (black nightshade) were isolated from fruits gathered from field (38° 6'25.03"N, 13° 21' 0.19"E), in autumn 2023, dried on filter paper for three days at room temperature and stored in a glass container in a refrigerator at 9°C, 35% of RH and photoperiod of 0:24 (light:dark) for 6 months. In April 2024, seeds were sown in plastic pots (22x22x26 cm) using a substrate mixture of blonde peat and perlite and placed in a greenhouse.
Aculops lycopersici was collected from S. nigrum plants in the garden of the Department SAAF, in May 2024 and used for infesting S. nigrum potted plants placed inside entomological cages (150 x 150 mesh, 160 µm aperture), in a controlled conditions (25±1°C, 70±5% RH, 16:8 light:dark photoperiod).
4.2.2. Adult Cohort for the Experiments
To obtain coetaneous for the experiments, 200 adults of A. lycopersici were transferred onto the adaxial surface of four S. nigrum leaves placed in Petri dishes (Ø150 mm, h 10 mm) on cotton wool saturated with distilled water. Adults were allowed to oviposit, and after a 24-hour period, they were removed. The presence of juvenile stages was recorded daily, and the postembryonic development was monitored until attaining adulthood. Since it was not possible to distinguish females from males under the stereomicroscope, a mixed population of both sexes was used in the trials.
4.2.3. Experimental Units
The experimental unit (EU) consists in a leaf disc (Ø 1.6 cm) placed on cotton wool moistened with distilled water in a Petri dish (Ø100 mm, h 10 mm). The adaxial surface of the leaf was used as experimental surface.
4.2.4. Effects of Essential Oils on A. lycopersici
Five adults of A. lycopersici were transferred on each EU, using a specialized pen with micro clamping mandrel, into which a human eyelash was inserted at the tip. The flexibility of the eyelash and the presence of micro-sculptures on its surface enabled the delicate collection of the specimens without injure them.
Essential oils were tested at five different concentrations: 320, 640, 1280, 2500 and 5000 µL L-1. Each EO was initially dissolved in pure acetone to ensure homogeneous mixing and afterwards distilled water was added in a 3:2 ratio (for water and acetone, respectively). For each concentration, 10 replications were carried out for a total of 50 A. lycopersici adults per test. The negative control replications were treated with only water and acetone at ratio 3:2.
Each replicate was treated with 8 ml of the solution using a Potter Precision Spray Tower (Burkard Manufacturing Co. Limited, Woodcock Hill Industrial Estate, Rickmansworth, Hertfordshire WD3 1PJ, England), set to a pressure of 62.05 kPa. The EUs were checked at 24h intervals for 4 days after the treatment; the cotton wool was replenished daily with distilled water. Adult mortality was registered daily until the conclusion of the tests.
4.2.5. Statistical Analysis
The Johnson and Kotz [62] transformation was applied on the mortality data before the General linear model analysis (GLM). In the presence of significant differences between treatments, the averages were separated by the Tukey's HSD test (P<0.05). Mortality was corrected by Abbott's [61] formula before the Probit analysis. The lethal concentrations corresponding to the mortality of 10% (LC10), 30% (LC30), 50% (LC50) and 90% (LC90) were determined by means of a probit model implemented in the Minitab software, considering a 95% of confidence level. All analyses were performed using Minitab 19.0 software (Minitab Inc., State College, PA, USA).
Author Contributions
Conceptualization, Thomas Giordano, Santo Orlando, Carlo Greco and Haralabos Tsolakis; Data curation, Thomas Giordano, Ilaria Marotta, Mauro Conte, Santo Orlando and Haralabos Tsolakis; Formal analysis, Santo Orlando and Haralabos Tsolakis; Funding acquisition, Michele Massimo Mammano and Haralabos Tsolakis; Investigation, Thomas Giordano, Giuliano Cerasa, Ilaria Marotta, Mauro Conte and Adele Salamone; Methodology, Thomas Giordano, Michele Massimo Mammano, Carlo Greco and Haralabos Tsolakis; Resources, Michele Massimo Mammano; Supervision, Haralabos Tsolakis; Validation, Thomas Giordano, Giuliano Cerasa and Haralabos Tsolakis; Visualization, Giuliano Cerasa and Haralabos Tsolakis; Writing – original draft, Thomas Giordano, Giuliano Cerasa, Ilaria Marotta, Santo Orlando, Adele Salamone, Michele Massimo Mammano, Carlo Greco and Haralabos Tsolakis; Writing – review & editing, Thomas Giordano, Adele Salamone, Carlo Greco and Haralabos Tsolakis.
Funding
This research was partially funded by Rural Development Plan Sicily 2014–2020, Measure 16.1, as part of the Research Projects “PREVANIA - Prodotti ad elevato valore nutrizionale ed a impatto ambientale ridotto” and Val.Inn.P.O. (Validazione di protocolli innovativi per la produzione di piante officinali di interesse nutraceutico coltivate in Sicilia), bando PSR Sicilia 2014-2020, and by the University of Palermo (Fondo di Finanziamento per la Ricerca FFR 2024, D13, by Haralabos Tsolakis).
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Lindquist, E.E.; Sabelis, M.W.; Bruin, J. Eriophyoid Mites. Their Biology, Natural Enemies and Control; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996.
- Pfaff, A.; Gabriel, D.; Böckmann, E. Observation and Restriction of Aculops lycopersici Dispersal in Tomato Layer Cultivation. J. Plant Dis. Prot. 2024, 131, 155–166. [CrossRef]
- Royalty, R.N.; Per Ring, T.M. Morphological Analysis of Damage to Tomato Leaflets by Tomato Russet Mite (Acari: Eriophyidae). J. Econ. Entomol. 1988, 81, 816–820. [CrossRef]
- Haque, M.M. Population Growth of Tomato Russet Mite, Aculops Lycopersici (Acari: Eriophyidae) and Its Injury Effect on the Growth of Tomato Plants. J. Acarol. Soc. Jpn. 2002, 11, 1–10. [CrossRef]
- Luigi, M.; Tiberini, A.; Taglienti, A.; Bertin, S.; Dragone, I.; Sybilska, A.; Tarchi, F.; Goggioli, D.; Lewandowski, M.; Simoni, S. Molecular Methods for the Simultaneous Detection of Tomato Fruit Blotch Virus and Identification of Tomato Russet Mite, a New Potential Virus–Vector System Threatening Solanaceous Crops Worldwide. Viruses 2024, 16, 806. [CrossRef]
- Pfaff, A.L. Aculops Lycopersici Tryon (Acari: Eriophyoidea) Monitoring, Control Options and Economic Relevance in German Tomato Cultivation. Bad Hersfeld, Deutschland, April 2023.
- Duso, C.; Castagnoli, M.; Simoni, S.; Angeli, G. The Impact of Eriophyoids on Crops: Recent Issues on Aculus schlechtendali, Calepitrimerus vitis and Aculops lycopersici. Exp. Appl. Acarol. 2010, 51, 151–168. [CrossRef]
- Marcic, D. Acaricides in Modern Management of Plant-Feeding Mites. J. Pest Sci. 2012, 85, 395–408. [CrossRef]
- Hessein, N.A.; Perring, T.M. Feeding Habits of the Tydeidae with Evidence of Homeopronematus Anconai (Acari: Tydeidae) Predation on Aculops lycopersici (Acari: Eriophyidae). Int. J. Acarol. 1986, 12, 215–221. [CrossRef]
- Park, H.-H.; Shipp, L.; Buitenhuis, R. Predation, Development, and Oviposition by the Predatory Mite Amblyseius swirkii (Acari: Phytoseiidae) on Tomato Russet Mite (Acari: Eriophyidae). J. Econ. Entomol. 2010, 103, 563–569. [CrossRef]
- van Houten, Y.; Glas, J.; Hoogerbrugge, H.; Rothe, J.; Bolckmans, K.; Simoni, S.; Van Arkel, J.; Alba, J.; Kant, M.; Sabelis, M. Herbivory-Associated Degradation of Tomato Trichomes and Its Impact on Biological Control of Aculops lycopersici. Exp. Appl. Acarol. 2013, 60, 127–138. [CrossRef]
- Pijnakker, J.; Hürriyet, A.; Petit, C.; Vangansbeke, D.; Duarte, M.V.; Arijs, Y.; Moerkens, R.; Sutter, L.; Maret, D.; Wäckers, F. Evaluation of Phytoseiid and Iolinid Mites for Biological Control of the Tomato Russet Mite Aculops lycopersici (Acari: Eriophyidae). Insects 2022, 13, 1146. [CrossRef]
- Gard, B.; Bardel, A.; Douin, M.; Perrin, B.; Tixier, M.-S. Laboratory and Field Studies to Assess the Efficacy of the Predatory Mite Typhlodromus (Anthoseius) recki (Acari: Phytoseiidae) Introduced via Banker Plants to Control the Mite Pest Aculops lycopersici (Acari: Eriophyidae) on Tomato. BioControl 2024, 1–13. [CrossRef]
- Al-Azzazy, M.M.; Al-Rehiayani, S.M.; Abdel-Baky, N.F. Life Tables of the Predatory Mite Neoseiulus cucumeris (Acari: Phytoseiidae) on Two Pest Mites as Prey, Aculops lycopersici and Tetranychus urticae. Arch. Phytopathol. Plant Prot. 2018, 51, 637–648. [CrossRef]
- Amaral, F.S.; Ferreira, M.M.; Lofego, A.C. Neoseiulus tunus (De Leon, 1967)(Acari: Phytoseiidae): Is This a Potential Natural Enemy of Aculops lycopersici (Massee, 1937)(Acari: Eriophyidae)? Entomol. Commun. 2021, 3, ec03033–ec03033. [CrossRef]
- Castagnoli, M.; Simoni, S.; Liguori, M. Evaluation of Neoseiulus californicus (McGregor) (Acari: Phytoseiidae) as a candidate for the control of Aculops lycopersici (Tryon) (Acari: Eriophyoidea): A preliminary study. Redia 2003, 86, 97–100.
- Trottin-Caudal, Y.; Fournier, C.; Leyre, J.M. Biological control of Aculops lycopersici (Massee) using the predatory mites Neoseiulus californicus McGregor and Neoseiulus cucumeris (Oudemans) on tomato greenhouse crops. In Colloque international tomate sous abri, protection intégrée - agriculture biologique, Avignon, France, 17-18 et 19 septembre 2003; Roche, L.; Edin, M.; Mathieu, V.; Laurens, F. Eds.; Centre de Balandran, BP 32, 30127 Bellegarde, France, 2003; pp. 153–157.
- Fischer, S.; Klötzli, F.; Falquet, L.; Celle, O. An Investigation on Biological Control of the Tomato Russet Mite Aculops lycopersici (Massee) with Amblyseius andersoni (Chant). In Proceedings of the IOBC/WPRS Working Group "Integrated Control in Protected Crops, Temperate Climate", Turku, Finland, 10-14 April, 2005; Enkegaard, A. Ed.; Finland 2005 IOBC/WPRS Bull 28:99–102.
- Aysan, E.; Nabi, A.K. Tritrophic Relationships among Tomato Cultivars, the Rust Mite, Aculops lycopersici (Massee)(Eriophyidae), and Its Predators. Acarologia 2018, 58, 5–17. 10.24349/acarologia/20184283.
- Isman, M.B.; Grieneisen, M.L. Botanical Insecticide Research: Many Publications, Limited Useful Data. Trends Plant Sci. 2013, 19, 140–145. 10.1016/j.tplants.2013.11.005.
- Jacobson, M. Botanical pesticides. Past, present, and future. In Insecticides of Plant Origin; Arnason, J.T., Philogène, B.J.R., Morand, P., Eds.; American Chemical Society: Washington, DC, USA, 1989; pp. 1–10. Available online: https://pubs.acs.org/ doi/pdf/10.1021/bk-1989-0387.ch001.
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential Oils in Insect Control: Low-Risk Products in a High-Stakes World. Annu. Rev. Entomol. 2012, 57, 405–424. [CrossRef]
- Rizzo, R.; Ragusa, E.; Benelli, G.; Lo Verde, G.; Zeni, V.; Maggi, F.; Petrelli, R.; Spinozzi, E.; Ferrati, M.; Sinacori, M. Lethal and Sublethal Effects of Carlina Oxide on Tetranychus urticae (Acari: Tetranychidae) and Neoseiulus californicus (Acari: Phytoseiidae). Pest Manag. Sci. 2023, 80, 967–977. [CrossRef]
- Marčić, D.; Döker, I.; Tsolakis, H. Bioacaricides in Crop Protection—What Is the State of Play? Insects 2025, 16, 95. [CrossRef]
- Park, Y.-L.; Tak, J.-H. Chapter 6—Essential oils for arthropod pest management in agricultural production systems. In Essential Oils in Food Preservation, Flavor and Safety, 1st ed.; Preedy, V.R., Ed.; Academic Press: Cambridge, MA, USA; Elsevier: San Diego, CA, USA, 2016; pp. 61–70. [CrossRef]
- Koul, O.; Walia, S.; Dhaliwal, G. Essential Oils as Green Pesticides: Potential and Constraints. Biopestic. Int 2008, 4, 63–84.
- Tsolakis, H.; Ragusa Di Chiara, S. Laboratory Evaluation of the Effect of Plant Extracts on Tetranychus urticae Koch (Acariformes, Tetranychidae). In Acarina biodiversity in the natural and human sphere; Luxograph, 2004; pp. 539–548.
- Greco, C.; Comparetti, A.; Fascella, G.; Febo, P.; La Placa, G.; Saiano, F.; Mammano, M.M.; Orlando, S.; Laudicina, V.A. Effects of Vermicompost, Compost and Digestate as Commercial Alternative Peat-Based Substrates on Qualitative Parameters of Salvia officinalis. Agronomy 2021, 11, 98. [CrossRef]
- Hardman, J.M.; Franklin, J.L.; Moreau, D.L.; Bostanian, N.J. An Index for Selective Toxicity of Miticides to Phytophagous Mites and Their Predators Based on Orchard Trials. Pest Manag. Sci. Former. Pestic. Sci. 2003, 59, 1321–1332. [CrossRef]
- Sharifi-Rad, M.; Berkay Yılmaz, Y.; Antika, G.; Salehi, B.; Tumer, T.B.; Kulandaisamy Venil, C.; Das, G.; Patra, J.K.; Karazhan, N.; Akram, M. Phytochemical Constituents, Biological Activities, and Health-promoting Effects of the Genus Origanum. Phytother. Res. 2020, 35, 95–121. [CrossRef]
- Lombrea, A.; Antal, D.; Ardelean, F.; Avram, S.; Pavel, I.Z.; Vlaia, L.; Mut, A.-M.; Diaconeasa, Z.; Dehelean, C.A.; Soica, C. A Recent Insight Regarding the Phytochemistry and Bioactivity of Origanum vulgare L. Essential Oil. Int. J. Mol. Sci. 2020, 21, 9653. [CrossRef]
- Cousyn, G.; Dalfrà, S.; Scarpa, B.; Geelen, J.; Anton, R.; Serafini, M.; Delmulle, L. Harmonizing the Use of Plants in Food Supplements in the European Union: Belgium, France and Italy-a First Step. Eur. Food Feed Law Rev. 2013, 3, 187–196.
- Orhan, F.; Ölmez, M. Effect of Herbal Mixture Supplementation to Fish Oiled Layer Diets on Lipid Oxidation of Egg Yolk, Hen Performance and Egg Quality. Ank. Üniversitesi Vet. Fakültesi Derg. 2011, 58, 33–39. [CrossRef]
- Liguori, G.; Greco, G.; Salsi, G.; Garofalo, G.; Gaglio, R.; Barbera, M.; Greco, C.; Orlando, S.; Fascella, G.; Mammano, M.M. Effect of the Gellan-Based Edible Coating Enriched with Oregano Essential Oil on the Preservation of the ‘Tardivo Di Ciaculli’Mandarin (Citrus reticulata Blanco Cv. Tardivo Di Ciaculli). Front. Sustain. Food Syst. 2024, 8, 1334030. [CrossRef]
- Cetin, H.; Cilek, J.E.; Aydin, L.; Yanikoglu, A. Acaricidal Effects of the Essential Oil of Origanum minutiflorum (Lamiaceae) against Rhipicephalus turanicus (Acari: Ixodidae). Vet. Parasitol. 2009, 160, 359–361. [CrossRef]
- Dolan, M.C.; Jordan, R.A.; Schulze, T.L.; Schulze, C.J.; Cornell Manning, M.; Ruffolo, D.; Schmidt, J.P.; Piesman, J.; Karchesy, J.J. Ability of Two Natural Products, Nootkatone and Carvacrol, to Suppress Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae) in a Lyme Disease Endemic Area of New Jersey. J. Econ. Entomol. 2009, 102, 2316–2324. [CrossRef]
- Tong, F.; Gross, A.D.; Dolan, M.C.; Coats, J.R. The Phenolic Monoterpenoid Carvacrol Inhibits the Binding of Nicotine to the Housefly Nicotinic Acetylcholine Receptor. Pest Manag. Sci. 2012, 69, 775–780. [CrossRef]
- Isman, M.B. Plant Essential Oils for Pest and Disease Management. Crop Prot. 2000, 19, 603–608. [CrossRef]
- Aziz, E.; Batool, R.; Akhtar, W.; Shahzad, T.; Malik, A.; Shah, M.A.; Iqbal, S.; Rauf, A.; Zengin, G.; Bouyahya, A. Rosemary Species: A Review of Phytochemicals, Bioactivities and Industrial Applications. South Afr. J. Bot. 2022, 151, 3–18. [CrossRef]
- Wanna, R.; Bozdoğan, H. Activity of Rosmarinus officinalis (Lamiales: Lamiaceae) Essential Oil and Its Main Constituent, 1, 8-Cineole, against Tribolium castaneum (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2025, 60, 86–106.
- Napoli, E.M.; Curcuruto, G.; Ruberto, G. Screening of the Essential Oil Composition of Wild Sicilian Rosemary. Biochem. Syst. Ecol. 2010, 38, 659–670. [CrossRef]
- Allenspach, M.; Steuer, C. α-Pinene: A Never-Ending Story. Phytochemistry 2021, 190, 112857. [CrossRef]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular Targets for Components of Essential Oils in the Insect Nervous System—a Review. Molecules 2017, 23, 34. [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological Effects of Essential Oils–a Review. Food Chem. Toxicol. 2008, 46, 446–475. [CrossRef]
- Dunan, L.; Malanga, T.; Benhamou, S.; Papaiconomou, N.; Desneux, N.; Lavoir, A.-V.; Michel, T. Effects of Essential Oil-Based Formulation on Biopesticide Activity. Ind. Crops Prod. 2023, 202, 117006. [CrossRef]
- Pierozan, M.K.; Pauletti, G.F.; Rota, L.; Santos, A.C.A. dos; Lerin, L.A.; Di Luccio, M.; Mossi, A.J.; Atti-Serafini, L.; Cansian, R.L.; Oliveira, J.V. Chemical Characterization and Antimicrobial Activity of Essential Oils of Salvia L. Species. Food Sci. Technol. 2009, 29, 764–770. [CrossRef]
- Delamare, A.P.L.; Moschen-Pistorello, I.T.; Artico, L.; Atti-Serafini, L.; Echeverrigaray, S. Antibacterial Activity of the Essential Oils of Salvia officinalis L. and Salvia triloba L. Cultivated in South Brazil. Food Chem. 2007, 100, 603–608. [CrossRef]
- Anderson, J.A.; Coats, J.R. Acetylcholinesterase Inhibition by Nootkatone and Carvacrol in Arthropods. Pestic. Biochem. Physiol. 2012, 102, 124–128. [CrossRef]
- Rguez, S.; Daami-Remadi, M.; Cheib, I.; Laarif, A.; Hamrouni, I. Composition Chimique, Activité Antifongique et Activité Insecticide de l’huile Essentielle de Salvia officinalis. Tunis J Med Plants Nat Prod 2013, 9, 65–76.
- Aissaoui, A.B.; Zantar, S.; Elamrani, A. Chemical Composition and Potential Acaricide of Salvia officinalis and Eucalyptus globulus on Tetranychus urticae Koch (Acarina: Tetranychidae). J. Appl. Chem. Environ. Prot. 2019, 4, 1–15.
- Chandler, D.; Bailey, A.S.; Tatchell, G.M.; Davidson, G.; Greaves, J.; Grant, W.P. The Development, Regulation and Use of Biopesticides for Integrated Pest Management. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1987–1998. [CrossRef]
- Chiasson, H.; Bélanger, A.; Bostanian, N.; Vincent, C.; Poliquin, A. Acaricidal Properties of Artemisia absinthium and Tanacetum vulgare (Asteraceae) Essential Oils Obtained by Three Methods of Extraction. J. Econ. Entomol. 2001, 94, 167–171. [CrossRef]
- Laborda, R.; Manzano, I.; Gamon, M.; Gavidia, I.; Perez-Bermudez, P.; Boluda, R. Effects of Rosmarinus officinalis and Salvia officinalis Essential Oils on Tetranychus urticae Koch (Acari: Tetranychidae). Ind. Crops Prod. 2013, 48, 106–110. [CrossRef]
- da Camara, C.A.; Akhtar, Y.; Isman, M.B.; Seffrin, R.C.; Born, F.S. Repellent Activity of Essential Oils from Two Species of Citrus against Tetranychus urticae in the Laboratory and Greenhouse. Crop Prot. 2015, 74, 110–115. [CrossRef]
- Greco, C.; Catania, P.; Orlando, S.; Vallone, M.; Mammano, M.M. Assessment of Vegetation Indices as Tool to Decision Support System for Aromatic Crops.; Springer, 2024; pp. 322–331.
- Greco, C.; Gaglio, R.; Settanni, L.; Sciurba, L.; Ciulla, S.; Orlando, S.; Mammano, M.M. Smart Farming Technologies for Sustainable Agriculture: A Case Study of a Mediterranean Aromatic Farm. 2025.
- Greco, C.; Catania, P.; Orlando, S.; Calderone, G.; Mammano, M.M. Rosemary Biomass Estimation from UAV Multispectral Camera. In Biosystems Engineering Promoting Resilience to Climate Change - AIIA 2024 - Mid Term.; Lecture Notes in Civil Engineering; Springer Cham, 2025; Vol. 586.
- Garofalo, G.; Ponte, M.; Greco, C.; Barbera, M.; Mammano, M.M.; Fascella, G.; Greco, G.; Salsi, G.; Orlando, S.; Alfonzo, A. Improvement of Fresh Ovine “Tuma” Cheese Quality Characteristics by Application of Oregano Essential Oils. Antioxidants 2023, 12, 1293. [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography /Mass Spectrometry, ed. 4.1. 2017. Available online: http://essentialoilcomponentsbygcms.com/ (accessed on 01 February 2025).
- Zito, P.; Sajeva, M.; Bruno, M.; Rosselli, S.; Maggio, A.; Senatore, F. Essential Oils Composition of Two Sicilian Cultivars of Opuntia ficus-indica (L.) Mill.(Cactaceae) Fruits (Prickly Pear). Nat. Prod. Res. 2013, 27, 1305–1314. [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Am. Mosq. Control Assoc. 1987, 3, 302–303.
- Johnson, N.; Kotz, S. Discrete Distributions. Their Distributions in Statistics. 1969, Boston, Houghton Mifflin. xvi, 328 p.
Figure 1.
MAPs NDVI values during vegetative period.

Figure 2.
Percentage surface area of rosemary, oregano, and sage based on NDVI classes during the three considered periods.
Figure 2.
Percentage surface area of rosemary, oregano, and sage based on NDVI classes during the three considered periods.
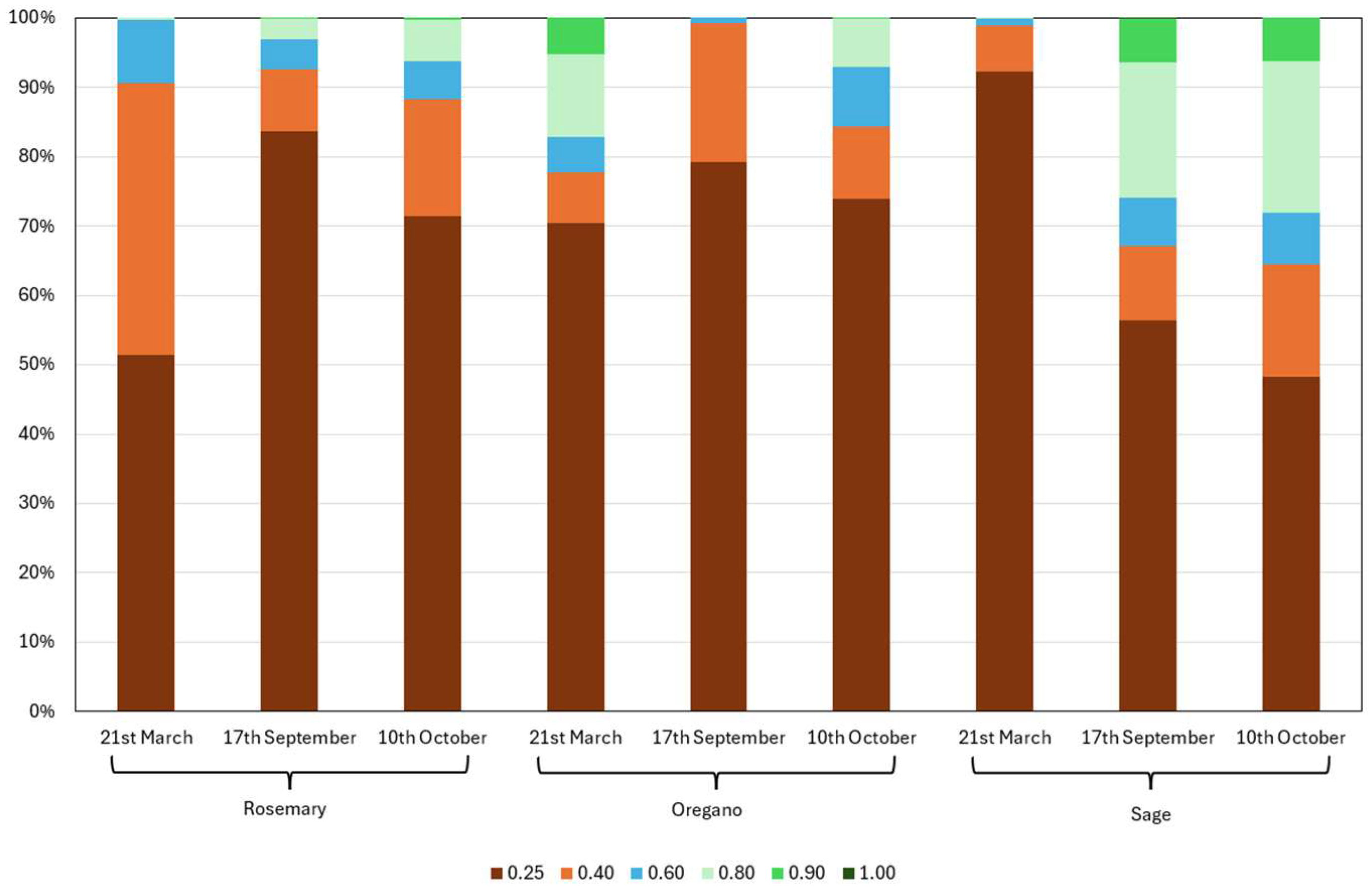
Figure 3.
Parametric cumulative failure plot for probability of mortality based on Probit data and maximum likelihood method. * Interquartile range.
Figure 3.
Parametric cumulative failure plot for probability of mortality based on Probit data and maximum likelihood method. * Interquartile range.
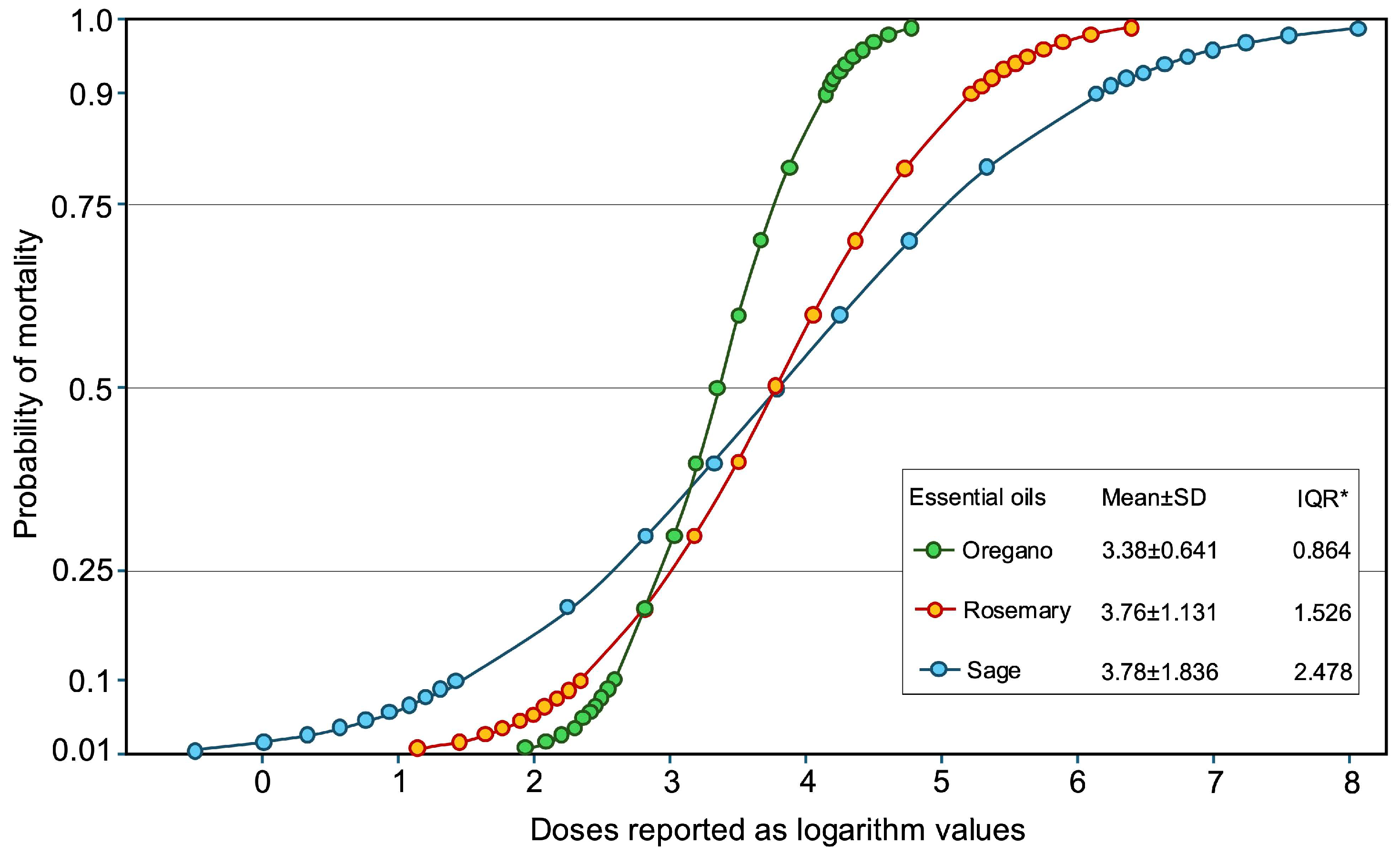
Figure 4.
Reflectance of Rosemary, Oregano and Sage in the 500-900 nm spectrum region measured with the spectroradiometer and spectral sensitivity of the M3M camera (G–Green, R–Red, RE–Red Edge and NIR–Near-Infrared).
Figure 4.
Reflectance of Rosemary, Oregano and Sage in the 500-900 nm spectrum region measured with the spectroradiometer and spectral sensitivity of the M3M camera (G–Green, R–Red, RE–Red Edge and NIR–Near-Infrared).
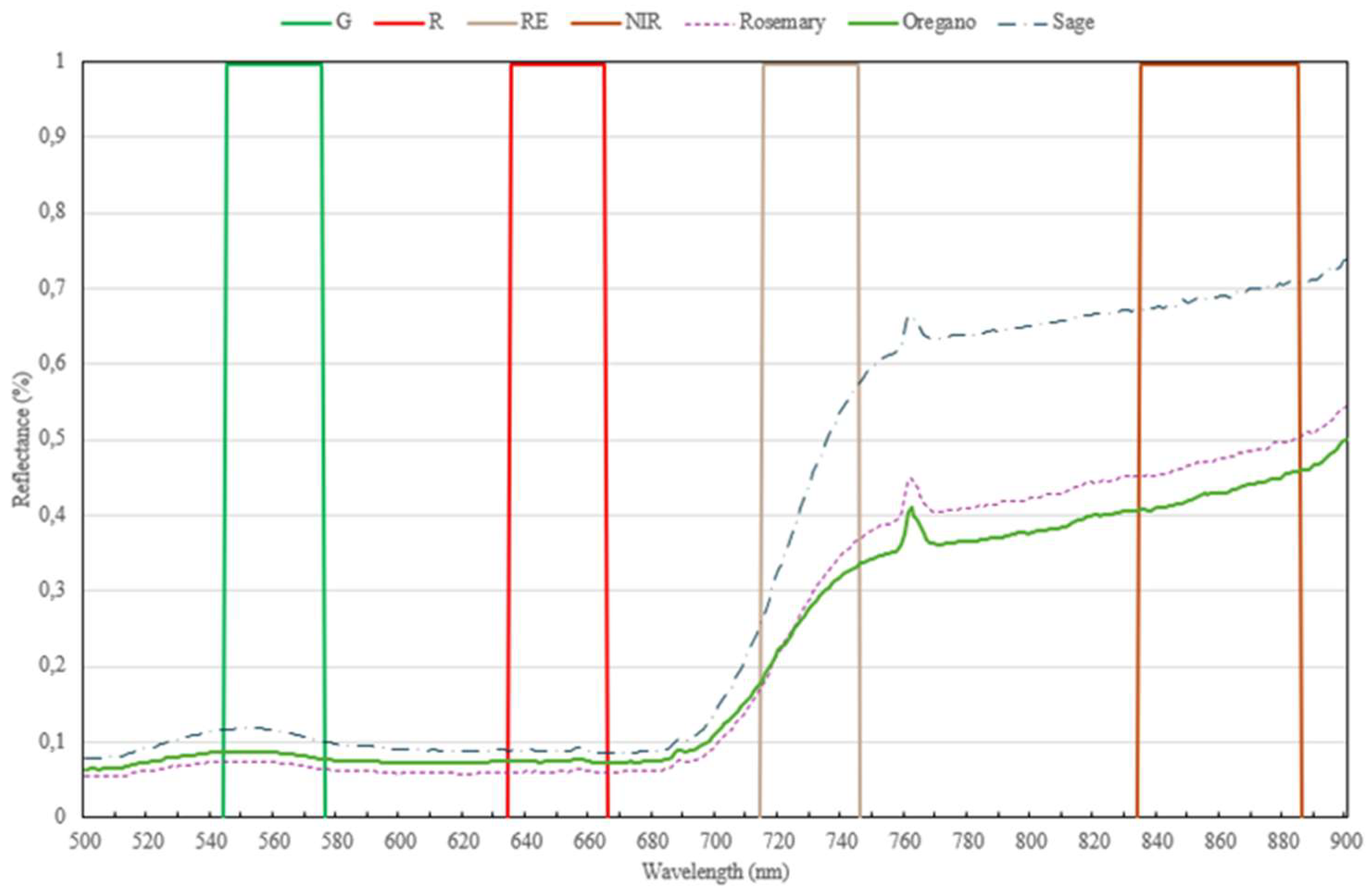
Figure 5.
MAPs monitoring through Precision Agriculture.


Table 1.
Percentage of components characterising the essential oils (EOs) of Oregano, Rosemary, and Sage, with their relative weights (RW %).
Table 1.
Percentage of components characterising the essential oils (EOs) of Oregano, Rosemary, and Sage, with their relative weights (RW %).
| Component/Chemical Class |
Oregano EO (RW%) |
Rosemary EO (RW%) |
Sage EO (RW%) |
| Monoterpene phenol | |||
| Carvacrol | 83.42 | - | - |
| Thymol | 0.89 | - | - |
| Subtotal | 84.31 | - | - |
| Monoterpene hydrocarbons | |||
| ρ-Cymene | 3.06 | 0.80 | - |
| γ-Terpinene | 2.93 | 2.10 | - |
| β-Myrcene | 1.01 | 1.21 | 2.19 |
| α-Terpinene | 0.99 | 0.45 | - |
| Limonene | 0.45 | 2.22 | - |
| Terpinolene | 0.41 | 0.53 | - |
| β-Ocimene | 0.42 | - | - |
| β-Pinene | 0.13 | 4.1 | 2.66 |
| Sabinene | 0.13 | - | - |
| α-Pinene | - | 28.0 | 2.66 |
| Linalyl Acetate | - | 1.12 | - |
| Crisantenone | - | - | 12.87 |
| Camphene | - | 7.00 | 9.26 |
| Subtotal | 9.53 | 47.53 | 29.64 |
| Oxygenated monoterpenes | |||
| Linalool | 0.32 | 3.45 | - |
| Camphor | 0.27 | 6.20 | 21.91 |
| Terpinen-4-ol | 0.18 | - | - |
| 1,8-Cineole (Eucalyptol) | - | 11.00 | 27.67 |
| Borneol | - | 7.72 | 2.59 |
| β-Thujone | - | 0.73 | - |
| α-Thujone | - | - | 5.32 |
| Bornyl Acetate | - | - | 1.45 |
| 4-Caranol | - | - | 0.09 |
| 4-Terpineol | 0.31 | - | 0.54 |
| α-Terpineol | - | 4.40 | - |
| α-Thujene | 0.23 | - | - |
| α-Phellandrene | 0.60 | - | - |
| Subtotal | 1.91 | 33.37 | 59.57 |
| Sesquiterpene hydrocarbons | |||
| β-Caryophyllene | 1.07 | 6.64 | 1.46 |
| α-Humulene | 0.70 | 0.75 | - |
| Germacrene | 0.07 | - | - |
| β-Bisabolene | 0.63 | - | - |
| Farnesene | 0.40 | - | - |
| α-Caryophyllene | - | - | 0.89 |
| Alloaromadendrene | - | - | 0.24 |
| α-Gurjunene | - | - | 0.13 |
| Carophyllene oxide | 0.88 | ||
| Subtotal | 2.87 | 8.27 | 2.72 |
| Oxygenated sesquiterpenes | |||
| Viridiflor | - | - | 1.22 |
| Palustrol | - | - | 0.75 |
| Ledol | - | - | 0.53 |
| Spathulenol | - | - | 0.31 |
| Subtotal | - | - | 2.81 |
| Other compounds | |||
| Camphol | 0.70 | - | - |
| 1-Octen-3-ol | 0.49 | - | - |
| 3-Octanone | 0.36 | - | - |
| Carvacrol Methyl Ether | 0.03 | - | - |
| Maool | - | - | 0.62 |
| Diethyl Phthalate | - | - | 0.53 |
| Naphthalene | - | - | 0.28 |
| Subtotal | 1.58 | - | 1.43 |
Table 2.
Susceptibility of adult stages of Aculops lycopersici to different concentrations of Origanum vulgare, Salvia rosmarinus and Salvia officinalis.
Table 2.
Susceptibility of adult stages of Aculops lycopersici to different concentrations of Origanum vulgare, Salvia rosmarinus and Salvia officinalis.
| Concentrations |
Cumulative mortality (%) (mean ± SE) |
Survival time (days) | Abbott’s corrected mortality | Toxicity class | ||||
| Plant extracts | (µL L-1) | Day 1 | Day 2 | Day 3 | Day 4 | (mean ± SE) | (%) | * |
| O. vulgare | 5000 | 66.00 ± 7.92 | 74.00 ± 8.46 | 80.00 ± 7.30 | 90.00 ± 4.47 a | 0.90 ± 0.203 a | 89.13 | 4 |
| 2500 | 14.00 ± 4.27 | 20.00 ± 2.98 | 34.00 ± 6.00 | 36.00 ± 7.77 bc | 2.96 ± 0.216 bcd | 30.43 | 2 | |
| 1280 | 0.00 ± 0.00 | 4.00 ± 2.67 | 12.00 ± 4.42 | 22.00 ± 5.54 efg | 3.62 ± 0.114 ed | 15.22 | 1 | |
| 640 | 2.00 ± 2.00 | 4.00 ± 2.67 | 12.00 ± 3.27 | 20.00 ± 5.16 ef | 3.62 ± 0.124 ed | 13.04 | 1 | |
| 320 | 2.00 ± 2.00 | 6.00 ± 3.06 | 10.00 ± 3.33 | 16.00 ± 4.00 efg | 3.66 ± 0.127 ed | 8.70 | 1 | |
| Control | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 8.00 ± 3.27 fg | 3.92 ± 0.038 e | 0.00 | - | |
| S. rosmarinus | 5000 | 16.00 ± 6.53 | 36.00 ± 9.33 | 42.00 ± 9.17 | 46.00±10.3 b | 2.60 ± 0.234 b | 46.00 | 2 |
| 2500 | 12.00 ± 4.42 | 24.00 ± 4.99 | 34.00 ± 4.27 | 40.00±5.16 bc | 2.90 ± 0.214 bcd | 40.00 | 2 | |
| 1280 | 2.00 ± 2.00 | 12.00 ± 4.42 | 20.00 ± 5.16 | 26.00±3.06 de | 3.40 ± 0.159 cde | 26.00 | 2 | |
| 640 | 2.00 ± 2.00 | 16.00 ± 4.99 | 16.00 ± 4.99 | 22.00±6.96 e | 3.44 ± 0.165 cde | 22.00 | 1 | |
| 320 | 4.00 ± 2.67 | 6.00 ± 3.06 | 6.00 ± 3.06 | 12.00±4.42 efg | 3.72 ± 0.128 cd | 12.00 | 1 | |
| Control | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00±0.00 g | 4.00 ± 0.00 e | 0.00 | - | |
| S. officinalis | 5000 | 12.00 ± 5.33 | 32.00 ± 9.52 | 42.00 ± 9.17 | 42.00 ± 9.17 bc | 2.72 ± 0.225 bc | 39.58 | 2 |
| 2500 | 12.00 ± 6.80 | 32.00 ± 6.80 | 40.00 ± 7.30 | 54.00 ± 8.46 b | 2.62 ± 0.216 bc | 52.08 | 3 | |
| 1280 | 10.00 ± 4.47 | 28.00 ± 6.11 | 32.00 ± 8.00 | 34.00 ± 6.70 bcd | 2.96 ± 0.218 bcd | 31.25 | 2 | |
| 640 | 0.00 ± 0.00 | 14.00 ± 4.27 | 22.00 ± 5.54 | 22.00 ± 5.54 cde | 3.42 ± 0.159 cde | 18.75 | 1 | |
| 320 | 4.00 ± 2.67 | 16.00 ± 4.00 | 18.00 ± 4.67 | 28.00±3.27 bcde | 3.34 ± 0.173 bcde | 25.00 | 1 | |
| Control | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 4.00±2.67 e | 3.96 ± 0.028 e | 0.00 | - | |
Table 3.
Lethal concentrations of the essential oils of Origanum vulgare, Salvia rosmarinus and Salvia officinalis against adult stages of Aculops lycopersici.
Table 3.
Lethal concentrations of the essential oils of Origanum vulgare, Salvia rosmarinus and Salvia officinalis against adult stages of Aculops lycopersici.
| Essential oils | LC10 µL L-1 (95% CI) |
LC30 µL L-1 (95% CI) |
LC50 µL L-1 (95% CI) |
LC90 µL L-1 (95% CI) |
LC95 µL L-1 (95% CI) |
Intercept ± SE | Slope ± SE | Goodness of fit χ2 (d.f.) |
| Origanum vulgare | 369.33 (208.57-530.18) |
1,068.17 (799.56-1362.01) |
2,228.90 (1740.28-3050.91) |
13,451.79 (8015.49-31804.60) |
22,391.85 (12084.54-63217.89) |
-5.49 ± 0.71 | 1.64 ± 0.22 | 21.62 (3) p = 0.000 |
| Salvia rosmarinus | 207.16 (33.57-428.25) |
1,488.81 (907.55-2451.9) |
5,835.39 (3281-22288.8) |
164,369.0 (35154.4-10573529.9) |
423,447.9 (67507.2-61877106.9) |
-3.32 ± 0.67 | 0.88 ± 0.21 | 0.53 (3) p = 0.911 |
| Salvia officinalis | 26.62 (0.002-146.38) |
654.53 (80.06-1292.05) |
6,013.81 (2630.4-261607.4) |
1,358,595.0 (74342.1-20,379,803,862,870.6) |
6,315,387.0 (184,714.1-3,651,741,272,548,380.0) |
-2.05 ± 0.61 | 0.54 ± 0.19 | 5.73 (3) p = 0.125 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



