
Submitted:
07 April 2025
Posted:
08 April 2025
You are already at the latest version
Abstract
Bacteria can sense changes in the environment and adapt to survive under different conditions. The two-component signal transduction system EnvZ/OmpR is described to mediate response to osmotic stress. This system also regulates genes involved in other processes such as virulence, fatty acid uptake, exopolysaccharide production, peptide transportation and flagella production. Considering that some of these processes are known to be important for a successful symbiosis, the present study addresses the role of EnvZ in rhizobia-legume interaction, evaluating the effects of extra envZ-like gene copies in the Mesorhizobium-chickpea symbiosis. Five Mesorhizobium-transformed strains, expressing the envZ-like gene from M. mediterraneum UPM-Ca36T, were evaluated in terms of symbiotic performance. Chickpea plants inoculated with envZ-transformed strains (PMI6envZ+ and EE7envZ+) showed significant higher symbiotic effectiveness as compared to the corresponding empty-vector strains. In plants inoculated with PMI6envZ+, a higher number of infection threads was observed, and nodules were visible 4 days earlier. Overall, our results showed that the expression of extra-copies of the envZ-like gene influenced the symbiotic process at different stages, leading to strain-dependent effects. This study represents the first report on the involvement of an EnvZ-like protein in the rhizobia-legume symbiosis and contributes to elucidate the role of these systems in bacteria-host interaction.
Keywords:
Legume
; Nodulation
; Nitrogen fixation
; Overexpression
; Rhizobia
; Symbiosis
1. Introduction
The ability to sense changes in the external environment is fundamental to bacterial survival [1,2,3]. Changes in the surroundings can lead to stress conditions, limiting bacterial growth and influencing the interactions with other prokaryotic or eukaryotic organisms. Bacteria respond to these fluctuations using a set of versatile systems that allow the detection of information about the environment and accordingly modulate gene expression [4].
Bacteria can mediate responses to environmental stress through two-component regulatory systems that comprise, a histidine kinase (HK) and a cognate response regulator (RR) [5,6]. As general mechanism, the sense of an environmental stress by the HK allows the autophosphorylation of a conserved histidine residue, and this phosphoryl group is subsequently transferred to a conserved aspartic acid residue of the RR. In its phosphorylated state, the RR usually binds to DNA target sequences stimulating the transcription of appropriate target genes [6]. The well characterised two-component regulatory system EnvZ/OmpR is activated in response to osmotic stress and regulates the expression of outer membrane porins OmpF and OmpC according to the osmolarity level [6,7]. At high osmolarity conditions, EnvZ is autophosphorylated and further transfers the phosphoryl group to OmpR, which in turn binds to the promoter regions of ompF and ompC genes that encode outer membrane porins [8,9]. Both porins act in the nutrient exchange, but at high osmolarity OmpC becomes the major porin in the outer membrane, while at low osmolarity conditions OmpF is more abundant [6]. When the osmotic stress is relieved, EnvZ is able to act as a phosphatase and dephosphorylates the OmpR-P [10].
In Escherichia coli, envZ/ompR mutations affect directly or indirectly the expression of more than 100 genes, including genes involved in amino acid biosynthesis, such as isoleucine and cysteine, iron and maltose transport, and flagellar synthesis [11]. ompR mutation in Yersinia pestis affected the expression of 224 genes, indicating a global regulatory role for OmpR [12]. Besides its relation to osmotolerance, EnvZ/OmpR system has also been associated with other functions, such as virulence in pathogens, fatty acid uptake, exopolysaccharide (EPS) production, peptide transport and flagella production [9,13,14,15,16,17,18,19], even in mammalian cells [20]. In addition, EnvZ/OmpR regulates the type III secretion system genes in pathogenic bacteria, such as Salmonella typhimurium, Yersinia enterocolitica, and Pseudomonas syringae [14,21,22].
Rhizobia are diazotrophic soil bacteria that can colonize the roots of many legumes species and establish mutualistic symbioses (for example [23]). Inside the root nodules, rhizobia reduce atmospheric nitrogen into ammonium, which is an N-compound that the host plant can use. In exchange, the host plant provides organic compounds resulting from photosynthesis, which are used as nutrient source by the microsymbiont. This biological nitrogen fixation process is particularly important in the context of agroecosystems, since it represents a natural way to generate an N-input into the crop, and ultimately to the soil, thus allowing the reduction of synthetic N-fertilizers applications. These N-fertilizers have relevant environmental costs, since their synthesis requires high energetic input, which is largely dependent on fossil fuels [24]. In addition, N-fertilizers are easily leached to groundwater, leading to problems of nitrate contamination. For all these reasons, the optimization of rhizobia-legume symbiosis is vital in the context of sustainable agriculture.
Rhizobia genomes are typically large and usually comprise several plasmids, which harbor mainly accessory genes. The genes directly involved in nodule development and nitrogen fixation, as for example nod, nif or fix genes, are encoded in symbiotic plasmids or in chromosomal symbiosis islands (for review see [25]). Besides the well-characterized symbiosis genes, other genes associated to different molecular mechanisms, as for example stress response or secretion system genes, may influence the symbiotic plant-rhizobia interaction [26,27].
As mentioned previously, several studies have addressed the function of EnvZ/OmpR in numerous pathogenic bacteria, mostly Gammaproteobacteria. In rhizobia, which belong to Alpha- and Betaproteobacteria, there are few reports mentioning envZ and/or envZ-like genes [28,29,30]. To the best of our knowledge, there are no studies on the function of EnvZ/OmpR two-component system in symbiotic bacteria. Nevertheless, most of the mechanisms that the EnvZ/OmpR system has been reported to influence in other bacteria, are important in rhizobia-legume interaction. For example, the role of EPS has been studied using different rhizobia-legume symbiosis and its production is known to be important for root colonization and host interaction (for review see [31]).
The aim of this study was to evaluate the effects of overexpressing an envZ-like gene in the rhizobia-host interaction, namely to evaluate if a higher expression of this gene could lead to an improvement of the symbiotic performance in chickpea mesorhizobia strains. Our results show that the deregulation of the two-component regulatory system EnvZ/OmpR, by expression of extra envZ-like copies, may influence the symbiotic process by means of different mechanisms, in a strain-dependent way.
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
All bacterial strains and plasmids used in this study are listed in Table 1. A group of four chickpea Mesorhizobium strains previously isolated from Portuguese soils was used: V-15b-Viseu, ST-2-Setúbal, EE-7-ENMP, PMI-6-Portimão [32,33,34]. In addition, the type strain Mesorhizobium mediterraneum UPM-Ca36T was also used [35].
Tryptone-yeast (TY) or yeast-mannitol agar (YMA) medium [36] at 28 °C were used to grow mesorhizobia strains and tetracycline (15 μg.ml−1) was added into the medium for pRK415-modified mesorhizobia strains. For bacteria modified with plasmids carrying the gfp and rfp genes, kanamycin (50 μg.ml−1) and gentamycin (15 μg.ml−1) were added to TY medium, respectively. E. coli strains were grown in Luria-Bertani (LB) medium [37] at 37 °C. For E. coli strains harboring pRK415, LB was supplemented with tetracycline (15 μg.ml−1), and for the strain MT616 carrying pRK600, chloramphenicol (25 μg.ml−1) was added.

Table 1.
Bacterial strains and plasmids used in this work.
| Plasmids/Strains | Characteristics | Reference |
|---|---|---|
| pRK600 | pRK2013 npt::Tn9. Cmr | [38] |
| pRK415 | Broad host-range vector; Tcr | [39] |
| pRKenvZ | Plasmid pRK415 containing the envZ-like gene (locus tag CIT25_RS09080) from M. mediterraneum UPM-Ca36T; Tcr | This work |
| pMRGFP | Plasmid containing the gfp gene; Kmr | [40] |
| pMP4661 | Plasmid containing the rfp gene; Gmr | [41] |
| E. coli MT616 | Strain harboring the helper plasmid pRK600 | [38] |
| E. coli DH5α | Competent cells | NZYTech |
| Mesorhizobium sp. V-15b | isolated from chickpea root nodules (Portugal) | [32] |
| Mesorhizobium sp. ST-2 | isolated from chickpea root nodules (Portugal) | [32] |
| Mesorhizobium sp. PMI-6 | isolated from chickpea root nodules (Portugal) | [32] |
| Mesorhizobium sp. EE-7 | isolated from chickpea root nodules (Portugal) | [32] |
| M. mediterraneum UPM-Ca36T | isolated from chickpea root nodules (Spain) | [32] |
| V15benvZ+ | Mesorhizobium sp. V-15b harboring pRKenvZ | This work |
| ST2envZ+ | Mesorhizobium sp. ST-2 harboring pRKenvZ | This work |
| PMI6envZ+ | Mesorhizobium sp. PMI-6 harboring pRKenvZ | This work |
| PMI6envZ+gfp | Mesorhizobium sp. PMI-6 harboring pRKenvZ and pMRGFP | This work |
| EE7envZ+ | Mesorhizobium sp. EE-7 harboring pRKenvZ | This work |
| EE7envZ+gfp | Mesorhizobium sp. EE-7 harboring pRKenvZ and pMRGFP | This work |
| Ca36envZ+ | M. mediterraneum UPM-Ca36T harboring pRKenvZ | This work |
| V15bpRK | Mesorhizobium sp. V-15b harboring pRK415 | [42] |
| ST2pRK | Mesorhizobium sp. ST-2 harboring pRK415 | [42] |
| PMI6pRK | Mesorhizobium sp. PMI-6 harboring pRK415 | [42] |
| PMI6pRKgfp | Mesorhizobium sp. PMI-6 harboring pRK415 and pMRGFP | [42] |
| PMI6pRKrfp | Mesorhizobium sp. PMI-6 harboring pRK415 and pMP4661 | This work |
| EE7pRK | Mesorhizobium sp. EE-7 harboring pRK415 | This work |
| EE7pRKgfp | Mesorhizobium sp. EE-7 harboring pRK415 and pMRGFP | This work |
| EE7pRKrfp | Mesorhizobium sp. EE-7 harboring pRK415 and pMP4661 | This work |
| Ca36pRK | M. mediterraneum UPM-Ca36T harboring pRK415 | This work |
2.2. Phylogenetic Analysis
As an initial approach, BLAST analyses of the envZ-like gene from M. mediterraneum UPM-Ca36T were performed to search for homologous genes in other rhizobia. In order to better characterise the envZ gene in rhizobia, the genomic context of envZ or envZ-like genes from Mesorhizobium species nodulating different hosts, was compared to that of E. coli, using data available in the NCBI genome database. In addition, a protein domain analysis of all envZ and envZ-like genes was performed with ScanProsite (https://prosite.expasy.org/scanprosite/), an Interpro database member [43]. An envZ phylogenetic analysis was performed with a final set of 11 Mesorhizobium strains, together with two other Alphaproteobacteria strains (Ensifer fredii and Agrobacterium fabrum), one Betaproteobacteria (Burkholderia mallei) and a group of Gammaproteobacteria, including E. coli (Table S1). For comparison purposes, a phylogenetic analysis of the 16S rRNA gene sequence was also performed, using the same set of strains. Sequences were aligned using the ClustalW algorithm and MEGA X [44] was used to perform the phylogenetic analysis, including the determination of the best nucleotide substitution model using Modeltest [45]. The phylogenetic tree was generated using the Neighbour-Joining algorithm with 1000 bootstrap replications.
2.3. Modifying Mesorhizobia Strains with an Extra EnvZ-Like Gene
The complete sequence of the envZ-like gene from M. mediterraneum UPM-Ca36T (=USDA 3392T), with the locus tag CIT25_RS09080, was amplified by PCR using 0.5 μL of DNA, 1× GC Buffer, 0.2 mM of each dNTP, 7.5 pmol of each primer and 0.4 U of Phusion DNA Polymerase (Thermo Fisher Scientific) in a final volume of 25 μL. The primers designed for this PCR reaction were envZ-HindIII-F (5′-AAGCTTAATGAGACGTTTCCTGCCGCA-3′) and envZ-BamHI-R (5′-GGATCCCTACGTTGCCAGCGGCAAGC-3′). The amplification program was: 30 s at 98 °C, 30 cycles of 10 s at 98 °C, 20 s at 56 °C, 28 s at 72 °C, and a final extension of 5 min at 72 °C. The obtained fragment of 1407 bp was cloned in pCR-BluntTM vector (Thermo Fisher Scientific) and sequenced (Macrogen). Finally, the envZ-like gene from M. mediterraneum UPM-Ca36T was subcloned in the expression vector pRK415, using the restriction sites corresponding to HindIII and BamHI.
Five chickpea Mesorhizobium strains were modified with the plasmid pRKenvZ containing the envZ-like gene under the control of the lac promoter, as described below. pRK415 was used as expression vector in a previous study and shown to have a constitutive expression in mesorhizobia [42].
Triparental conjugation with E. coli DH5α cells harboring the plasmid pRKenvZ or pRK415 as donor, and the helper strain E. coli MT616 containing pRK600, were carried out as previously described [35]. The pRKenvZ-modified mesorhizobia strains were named V15benvZ+, ST2envZ+, PMI6envZ+, EE7envZ+ and Ca36envZ+. The mesorhizobia strains EE-7-ENMP and UPM-Ca36T were modified with plasmid pRK415 (control), and named EE7pRK and Ca36pRK, respectively. Mesorhizobium sp. strains V-15b-Viseu, ST-2-Setúbal and PMI-6-Portimão modified with pRK415 were already available from a previous study [32].
To confirm the mesorhizobia acquisition of pRKenvZ and pRK415, total DNA was extracted according to [46] and used to amplify the multiple cloning site region. Using the universal primers M13F and M13R-pUC, a DNA fragment of 1529 bp will be amplified from pRKenvZ and 122 bp will be amplified from pRK415. The PCR reaction was performed using 5 μL of total DNA, 1X Green GoTaq® Flexi buffer, 0.2 mM of each dNTP, 1.5 mM MgCl2, 15 pmol of each universal primer and 0.625U of GoTaq® G2 Flexi DNA Polymerase (Promega), in a final volume of 50 μL. The amplification program included an initial denaturation step of 2 min at 95 °C, followed by 30 cycles of 60 s at 95 °C, 45 s at 56 °C, 85 s at 72 °C, and a final extension of 5 min at 72 °C. Amplification fragment size was verified by agarose gel electrophoresis.
2.4. Evaluation of the Symbiotic Performance
To evaluate if additional envZ-like gene copies improved the symbiotic effectiveness of chickpea mesorhizobia strains, a pot trial was performed using chickpea pre-germinated seeds (variety ELIXIR, cultivar CHK3236) inoculated with the wild-type, the strains harboring pRKenvZ and the strains harboring pRK415 [47].
For inocula preparation, rhizobia strains were grown in liquid TY medium, with the appropriate antibiotics, at 28 °C for 72 h. Cell cultures were centrifuged at 10.000 × g and resuspended in fresh TY medium to an optical density (OD) at 540 nm of 1.0. Each chickpea seedling was transferred to a plastic pot with sterile vermiculite and inoculated with 1 mL of this bacterial suspension. Five replicates per treatment were performed. A nitrogen-free nutrient solution [48] was applied three times a week. Uninoculated chickpea plants were used as negative control and uninoculated plants receiving the nutrient solution supplemented with 0.1% of KNO3 were used as positive control. Plants were kept in a growth chamber using a cycle of 16 h-light/8 h-dark and temperatures of 24 –day/18 –night, with a constant relative humidity of 65%.
A second pot trial was performed to evaluate the performance of plants inoculated with strains harboring pRKenvZ under salinity stress. Plants inoculated the strains harboring the empty expression vector (pRK415), as well as the abovementioned negative and positive control treatments were also included in this second pot trial. All the procedures were the same as described above, with the only difference being the supplementation of the watering solution with 0.15% NaCl to impose the salinity stress [49]. The watering with the nutrient solution supplemented with NaCl started at 15 days after inoculation and was alternated with normal nutrient solution watering.
After 7 weeks, plants were harvested and several parameters were measured, namely number of nodules (NN) and average weight per nodule (AWN) as well as shoot dry weight (SDW) and root dry weight (RDW). In addition, the symbiotic effectiveness (SE) was estimated using the shoot dry weight values, including those from the positive and negative controls [50]. Statistical analysis was performed using the software SPSS statistics V.21 (SPSS Inc; IBM New York, USA) and included analysis of variance, namely one-way ANOVA, and the Tukey’s Multiple Range Test (p < 0.05).
Considering the results obtained in this symbiotic performance trial, a subset of strains was selected for further analyses: PMI6envZ+ and EE7envZ+ as well as the corresponding control strains, PMI6pRK and EE7pRK.
2.5. Bacterial Growth in Salt Stress Conditions
To evaluate the effect of extra envZ-like copies in the tolerance to salinity stress, strains were grown in 5 mL of liquid TY medium supplemented with 0.75% NaCl (three replicates for each strain). The optical densities at 540 nm were measured every 24 h, for a total of 168 h. T-test (p < 0.05) was used to compare the growth of different strains, at the same timepoint. Bacterial growth was also evaluated on YMA medium supplemented with 1.5% NaCl. Cell density was adjusted to an initial OD540 nm of 0.2 from which decimal dilutions up to 10−5 were performed. A volume of 10 μL from each dilution was inoculated in YMA plates with and without NaCl supplementation and growth was observed after 6 days of incubation at 28 °C.
2.6. Motility Assay and Mucoid Phenotype
The swimming ability was compared between the strains carrying the empty vector (pRK415) and those modified with extra envZ-like gene copies, using TY plates containing 0.25% agar [51]. A volume of 10 μL of cell suspension (OD540 nm of 0.1) was used to inoculate the center of a TY plate and the diameter of the growth was evaluated, as previously described [42].
In order to evaluate differences in the mucoid phenotype between the strains harboring pRK415 or pRKenvZ, bacterial growth was observed in YMA plates supplemented with 25 mg Congo Red l-1 [52]. All tests were performed in triplicate.
2.7. Evaluation of Nodulation Kinetics
The time-course of nodulation was monitored using chickpea seeds (variety ELIXIR, cultivar CHK3236) inoculated with PMI6pRK, PMI6envZ+, EE7pRK and EE7envZ+ strains (OD 540 nm of 0.6) and grown under hydroponic conditions, as previously described [53]. This trial was performed using eight plants per treatment and the same plant growth chamber settings described above for the evaluation of symbiotic performance. The number of nodules was evaluated every three days, for a total of 33 days. T-test (p < 0.05) was used to compare different treatments at the same timepoint.
2.8. Analysis of Rhizobia Infection Process
The infection process was investigated using confocal microscopy and fluorescent proteins to tag the different rhizobia strains, namely green fluorescent protein (GFP) and red fluorescent protein (RFP). For these analyses, strains PMI6envZ+ and EE7envZ+ were modified with plasmid pMRGFP [40], which includes the gfp gene, while strains carrying PMI6pRK and EE7pRK were modified with plasmid pMRGFP or pMP4661, which includes the rfp gene [41]. [32,47]These plasmids were mobilized into mesorhizobia cells by triparental conjugation as described above, and its presence was confirmed by antibiotic resistance and fluorescence microscopy observations of cell cultures.
Pre-germinated chickpea seeds were inoculated with mesorhizobia strains tagged with GFP (single inoculation experiments) or RFP and GFP (co-inoculation experiments) [53]. Chickpea roots were stained using 10 μM propidium iodide (Sigma-Aldrich) or 50 mg calcofluor white L−1 (Sigma- Aldrich) and 10% potassium hydroxide [54]. At 4 or 6 days after inoculation, analyses of the infection process were performed using a Confocal Laser Scanning Microscope (Leica TCS SPE), which allowed the visualization of fluorescence from GFP at 488 nm, RFP and propidium iodide at 532 nm and calcofluor white at 405 nm. Using the Leica LASX software, projections were made by stacks accumulation and adjusting individual channels.
3. Results
3.1. Phylogenetic Analysis of EnvZ/envZ-Like Genes in the Genus Mesorhizobium
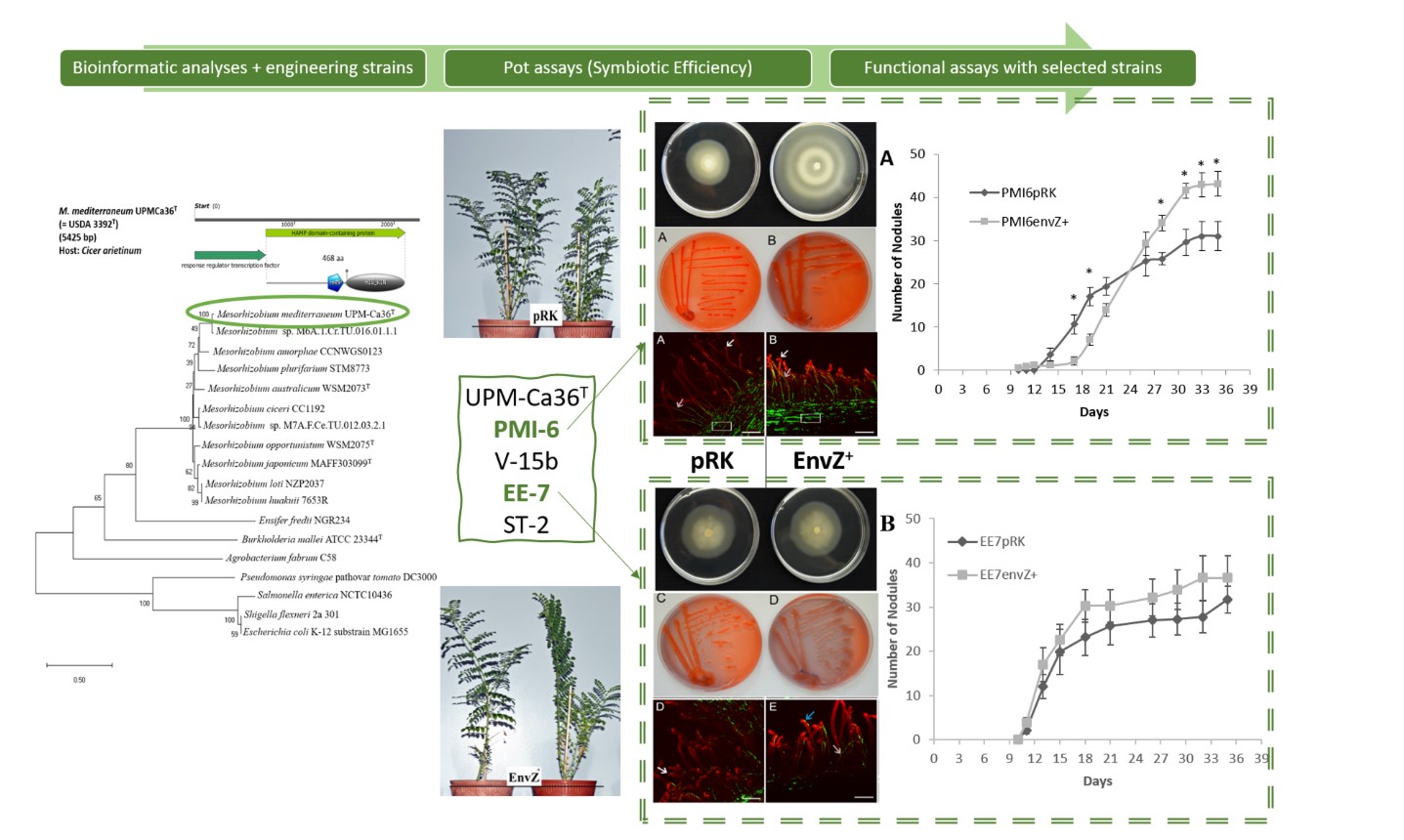
Analysis of the envZ-like gene from M. mediterraneum UPM-Ca36T using BLAST allowed the identification of highly similar sequences in several other Mesorhizobium genomes, comprising different species and isolated from distinct host legumes. Since most of these sequences are not annotated as envZ, the designation envZ-like genes was used in these cases. Analysis of the genomic context of envZ and envZ-like genes in Mesorhizobium strains showed that envZ and the corresponding response regulator, located immediately upstream, are usually positioned next to genes encoding ABC transporters (Figure 1 and Figure S1). In terms of the presence of conserved protein domains, all analysed EnvZ and EnvZ-like proteins present a HAMP linker domain of 53 amino acids and a histidine kinase domain of approximately 200 amino acids at the C-terminal (Figure 1 and Figure S1).
Figure 1.
Genomic context of envZ or envZ-like genes in E. coli and in two mesorhizobia species (M. mediterraneum UPM-Ca36T and M. ciceri CC1192) nodulating chickpea (Cicer arietinum). The annotation available on the NCBI database was used. envZ or envZ-like as well as the upstream response regulator are marked in green. For the envZ or envZ-like genes, the protein domain analysis is shown, with HAMP standing for HAMP domain and HIS_KIN standing for histidine kinase domain.
Figure 1.
Genomic context of envZ or envZ-like genes in E. coli and in two mesorhizobia species (M. mediterraneum UPM-Ca36T and M. ciceri CC1192) nodulating chickpea (Cicer arietinum). The annotation available on the NCBI database was used. envZ or envZ-like as well as the upstream response regulator are marked in green. For the envZ or envZ-like genes, the protein domain analysis is shown, with HAMP standing for HAMP domain and HIS_KIN standing for histidine kinase domain.
The phylogenetic analysis of the envZ and envZ-like genes (Figure 2) showed that all the Mesorhizobium strains form a monophyletic group, since the remaining Alphaproteobacteria, with an annotated envZ gene, form different lineages, including E. fredii NGR234 (a broad host range rhizobium). The Gammaproteobacteria included in the phylogeny grouped in a separate cluster, as expected, since the sequence of the envZ gene of these species shows low identity to the Mesorhizobium sequences. Using this same set of species, the 16S rRNA gene-based phylogeny (Figure S2) shows that the clusters generated reflect the different classes of phylum Pseudomonadota, with all the Alphaproteobacteria grouping in the same cluster. Overall, these analyses support the annotation of the gene from M. mediterraneum UPM-Ca36T (=USDA 3392T) with the locus tag CIT25_RS09080 as envZ-like and for simplicity, this gene will be from now on referred as envZ. Despite the presence of genes encoding the EnvZ/OmpR two-component system in the genome of UPM-Ca36T, the two genes typically regulated by this sensor system (ompF and ompC) were not identified. This genome is still in the draft stage, so the presence of ompF and ompR cannot be completely ruled out.
Figure 2.
Neighbour-Joining phylogenetic tree based on envZ sequences. Tamura 3-parameter model with gamma distributed rate among sites was used. Percentage of bootstrap support (1000 replicates) is indicated on internal branches. Scale bar indicates 0.5 substitutions per site.
Figure 2.
Neighbour-Joining phylogenetic tree based on envZ sequences. Tamura 3-parameter model with gamma distributed rate among sites was used. Percentage of bootstrap support (1000 replicates) is indicated on internal branches. Scale bar indicates 0.5 substitutions per site.

3.2. Evaluation of the Symbiotic Performance
Five mesorhizobia strains overexpressing envZ were obtained by transformation with an expression vector. A previous study confirmed that this expression vector is suitable for ectopic expression in chickpea mesorhizobia [42]. The mesorhizobia strains overexpressing envZ were tested under controlled conditions to assess if this higher expression levels could increase the symbiotic effectiveness. Despite the fact that envZ overexpression did not increase the number of nodules developed on chickpea plants for any of the five strains tested, the symbiotic effectiveness (SE) was improved for PMI6envZ+ and EE7envZ+ with an increase of 64% and 96%, respectively (Table 2). SDW, which is used to calculate the SE, was significantly higher for plants inoculated with PMI6envZ+ and EE7envZ+, than for those inoculated with PMI6pRK and EE7pRK, respectively. From those two strains with extra envZ copies, only EE7envZ+ showed statistical differences in the average weight per nodule (AWN), which indicates that larger nodules were formed in plants inoculated with EE7envZ+, when compared to plants inoculated with the corresponding strain harboring pRK415 (empty vector). Nodules from plants inoculated with ST2envZ+ also showed a significant increase on AWN, yet in this case, no differences on SE or SDW were detected. In terms of root dry weight (RDW), significantly higher values were obtained in plant inoculated with EE7envZ+, PMI6envZ+ and ST2envZ+, when compared to the corresponding strains harboring pRK415. These results showed that an increase on AWN or RDW is not directly reflected in an improvement of SE.
The homologous overexpression of envZ on UPM-Ca36T as well as the heterologous overexpression on V-15b did not improve any of the above-mentioned symbiotic parameters. For the five strains used in this study, the SE of the wild type strains was similar to that of the corresponding strains containing pRK415 (no statistically significant differences were obtained).
The second pot trial carried out under salinity conditions did not a better performance of plant inoculated with envZ+ strains, as compared to the corresponding strains harboring pRK415.
Taking these results in account, further analyses were performed using only the two strains that showed significant improvement in the SE, namely PMI6envZ+ and EE7envZ+ strains, and their corresponding control (empty vector) strains.
Table 2.
Plant parameters obtained from pot assays of chickpea plants inoculated with transformed mesorhizobia strains.
Table 2.
Plant parameters obtained from pot assays of chickpea plants inoculated with transformed mesorhizobia strains.
| Strain | SDW (g) | RDW (g) | NN | AWN (mg) | SE (%) |
|---|---|---|---|---|---|
| V15bpRK | 0.699 + 0.062 a | 0.335 + 0.039 a | 67 + 17 a | 1.567 + 0.246 a | 32.22 + 6.1 a |
| V15benvZ+ | 0.756 + 0.044 a | 0.432 + 0.027 a | 73 + 11 a | 1.332 + 0.123 a | 37.78 + 4.3 a |
| Ca36pRK | 0.789 + 0.085 a | 0.342 + 0.035 a | 57 + 7 a | 1.994+ 0.256 a | 41.07 + 8.4 a |
| Ca36envZ+ | 0.848 + 0.036 a | 0.296 + 0.021 a | 77 + 3 a | 1.333+ 0.054 a | 46.78 + 3.5 a |
| ST2pRK | 1.469 + 0.096 a | 0.478 + 0.033 b | 52 + 6 a | 2.546 + 0.314 b | 38.76 + 4.6 a |
| ST2envZ+ | 1.724 + 0.132 a | 0.603 + 0.023 a | 47 + 4 a | 3.754 + 0.257 a | 51.15 + 6.4 a |
| PMI6pRK | 1.325 + 0.113 b | 0.436 + 0.032 b | 66 + 13 a | 2.411 + 0.440 a | 31.78 + 5.5 b |
| PMI6envZ+ | 1.728 + 0.079 a | 0.527 + 0.020 a | 55 + 8 a | 3.405 + 0.734 a | 51.40 + 3.8 a |
| EE7pRK | 0.680 + 0.041 b | 0.319 + 0.011 b | 65 + 9 a | 1.099 + 0.440 b | 30.3 + 4.1 b |
| EE7envZ+ | 0.973 + 0.027 a | 0.556 + 0.034 a | 63 + 6 a | 1.707 + 0.734 a | 59.1 + 2.7 a |
Average of five replicates per treatment are shown ± standard error. Different letters in the same group of strains indicate statistically significant differences by independent sample t test (p < 0.05); significant differences are highlighted in bold. SDW—Shoot Dry Weight; RDW—Root Dry Weight; NN—Number of Nodules; AWN—Average Weight per Nodule; SE—Symbiotic Effectiveness.
3.3. Bacterial Growth Under Salt Stress Conditions
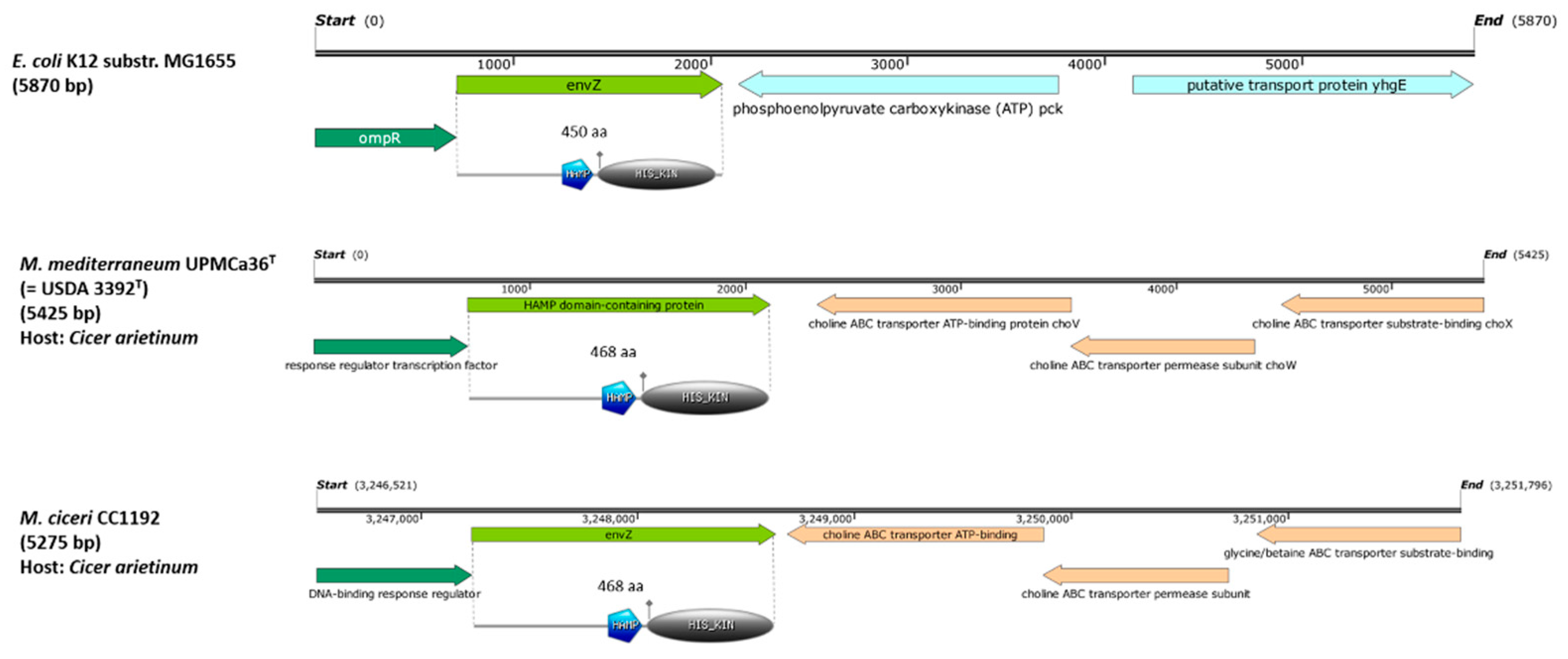
Since the envZ gene typically responds to an increase in osmolarity [6], the tolerance to salinity of the modified strains was evaluated on plates using YMA medium supplemented with 1.5% NaCl. This test showed that PMI6envZ+ was clearly more tolerant to salinity than the PMI6pRK strain, since the strain with extra envZ copies was able to grow even in the highest dilution (10−5), whereas PMI6pRK was not able to grow in any of the dilutions (Figure 3a). In addition, under control conditions (top panel of Figure 3a), it is evident an alteration of the mucoid phenotype of strain PMI6envZ+ (further discussed on section “Exopolysaccharides production”). Under these same salinity conditions, both EE7pRK and EE7envZ+ strains were only able to grow until the 10−2 dilution, showing low tolerance to this salinity stress and no effect of the envZ extra copies (Figure 3b).
The growth of the modified strains was also evaluated in liquid TY medium supplemented with 0.75% NaCl. Under this salinity condition the PMI6envZ+ strain was able to grow faster in the exponential phase compared with the PMI6pRK strain (Figure 3c). Significant differences were found between the growth of PMI6pRK and PMI6envZ+ strains from 48 h onwards (p <0.05), using T-test. Similarly to what was observed in the plate assay, no differences were observed between EE7pRK and EE7envZ+ growth rate and both strains reached stationary phase at 120 h (Figure 3d).
Figure 3.
Evaluation of tolerance to salinity stress. Plate test with YMA medium (no NaCl added) or YMA supplemented with 1.5% NaCl: (a) PMI6pRK and PMI6envZ+ strains; (b) EE7pRK and EE7envZ+ strains. Growth curves in liquid TY medium supplemented with NaCl 0.75%: (c) PMI6pRK and PMI6envZ+; (d) EE7pRK and EE7envZ+. Bars in (c) and (d) represent standard deviation.
Figure 3.
Evaluation of tolerance to salinity stress. Plate test with YMA medium (no NaCl added) or YMA supplemented with 1.5% NaCl: (a) PMI6pRK and PMI6envZ+ strains; (b) EE7pRK and EE7envZ+ strains. Growth curves in liquid TY medium supplemented with NaCl 0.75%: (c) PMI6pRK and PMI6envZ+; (d) EE7pRK and EE7envZ+. Bars in (c) and (d) represent standard deviation.
3.4. Swimming
Since the motility of rhizobia strains may affect their infection ability [55,56], the effects of extra copies of envZ in the swimming ability was evaluated.
The strain PMI6envZ+ showed a migration zone larger than PMI6pRK (Figure S3a,b). This observation indicates a faster swimming ability of PMI6envZ+, which could contribute to improve the performance of this strain in the root hairs infection process. The same test was performed for EE7envZ+ and EE7pRK and no differences were observed between these strains (Figure S3c,d).
3.5. Exopolysaccharides Production
As the ability to form exopolysaccharides is a very important feature at the rhizobia-legume interaction [28], a plate assay was performed in order to verify whether envZ overexpression influences the mucoid phenotype, which is typical of most rhizobia. When growing in YMA, PMI6envZ+ showed a stronger mucoid phenotype than PMI6pRK (Figure S4a,b) or the wild type strain. Despite the fact that the EE-7 wild type strain is naturally very mucoid, the strain modified with extra-envZ copies showed a more accentuated mucoid phenotype than the empty vector strain (Figure S4c,d), although this change is not as evident as in the case of PMI-6. In addition, and since the YMA plates were supplemented with Congo Red, a pH indicator, it was possible to track changes in the medium pH. In both PMI6envZ+ and EE7envZ+, the acidification of the culture medium was detected by the change from red to blue-violet coloration of the culture media. This pH change was more evident for PMI6envZ+, since the empty-vector strain did not acidify the medium, while in the case of EE7envZ+, the empty-vector strain already shows some degree of medium acidification.
3.6. Nodule Formation
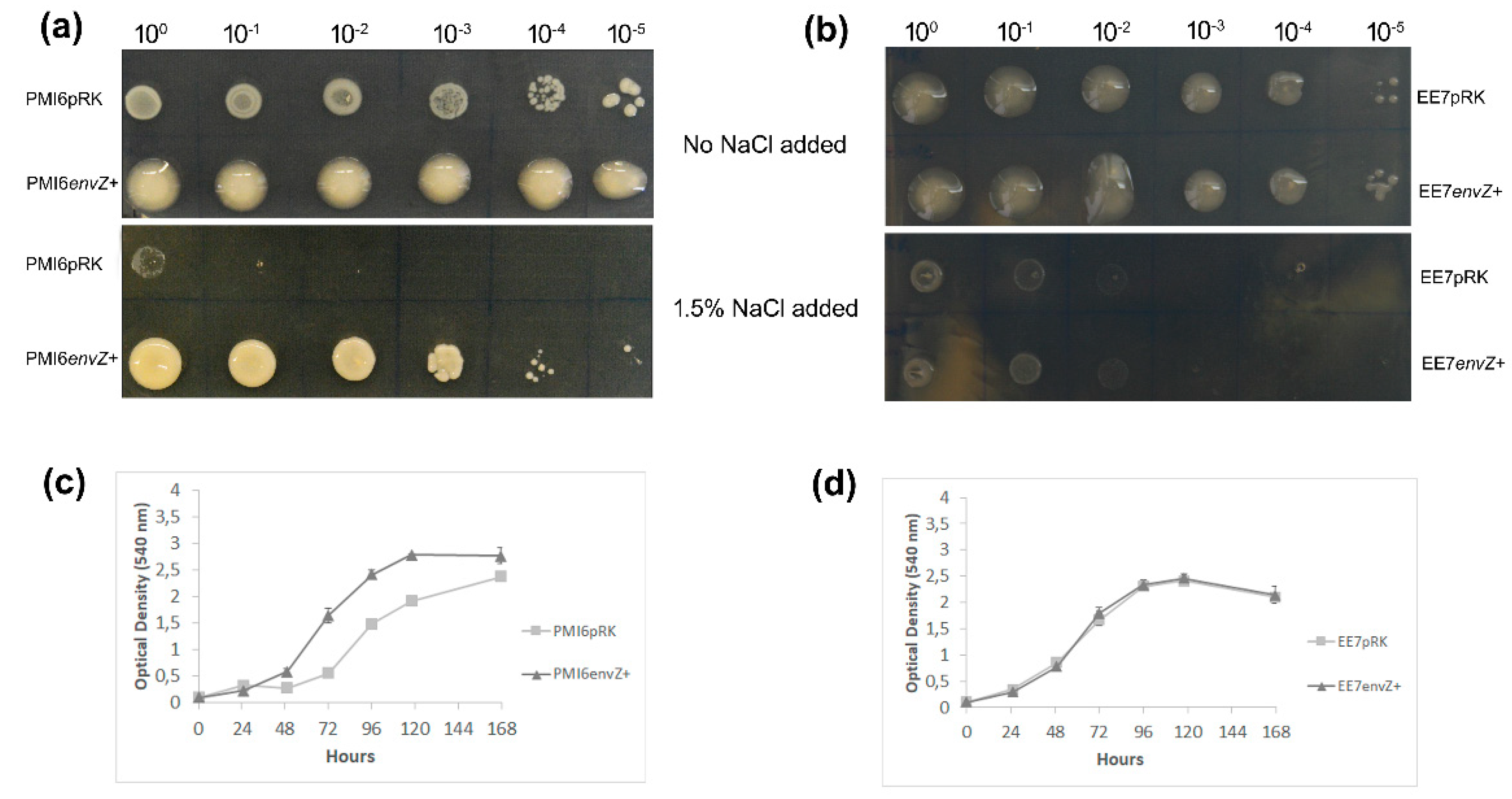
To investigate the effects of extra-envZ copies in the rate of nodule formation, an assay with chickpea plants grown in hydroponic conditions was performed. Plants inoculated with PMI6envZ+ showed the first nodules at 10 days after inoculation (DAI), while plants inoculated with PMI6pRK showed the first nodules only at 14 DAI (Figure 4). Although in an early phase of the assay, the number of nodules in plants inoculated with PMI6pRK is statistically higher (p <0.05), namely at 17 and 19 DAI. At 28 DAI the strain harboring extra envZ copies began to show a significantly higher (p <0.05) number of nodules and this number continues to be higher than in the plants inoculated with PMI6pRK until the end of the experiment (Figure 4a).
Similarly, plants that were inoculated with EE7envZ+ showed a higher number of nodules than plants inoculated with EE7pRK, however, in this case, the difference is not statistically significant (Figure 4b).
Figure 4.
Nodulation kinetics on chickpea plants. Average number of nodules for plants inoculated with (a) PMI6pRK or PMI6envZ+ strains (b) EE7pRK or EE7envZ+ strains. Asterisks indicate statistically significant differences by independent sample t test (p < 0.05). Bars represent standard error.
Figure 4.
Nodulation kinetics on chickpea plants. Average number of nodules for plants inoculated with (a) PMI6pRK or PMI6envZ+ strains (b) EE7pRK or EE7envZ+ strains. Asterisks indicate statistically significant differences by independent sample t test (p < 0.05). Bars represent standard error.
3.7. Colonization and Infection Thread Formation
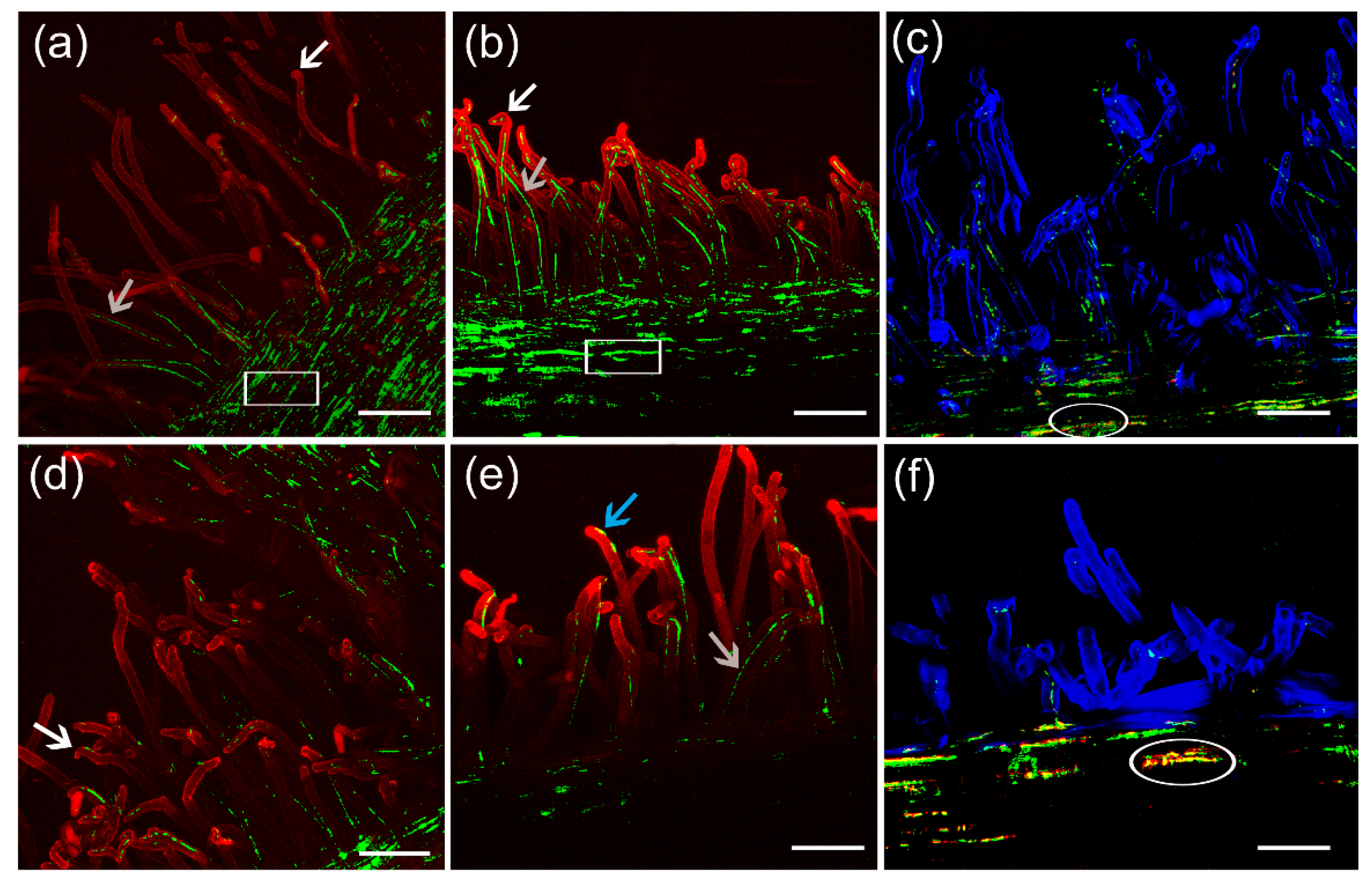
In order to investigate if the observed improvement of the symbiotic phenotype of PMI6envZ+ and EE7envZ+ relied on a higher competitiveness of these strains in the early stages of the interaction with the host roots, the infection process was evaluated using confocal microscopy. Considering the development and size of chickpea roots, it was not possible to accurately evaluate colonization and infection thread formation in a quantitative manner. Instead, experiments of single and co-inoculation were performed and analysed qualitatively at different time points.
At 4 DAI, no differences were detected between chickpea seedlings inoculated with EE7pRKgfp and EE7envZ+gfp strains. Important infection parameters such as curling, formation of infection threads and formation of caps on root hair tips were very similar between two strains (Figure 5d,e). Also at 4 DAI, chickpea seedlings inoculated with the PMI6envZ+gfp strain showed more infection threads than roots inoculated with PMI6pRKgfp (Figure 5a,b). In addition, colonization of the surface of the roots seemed more efficient for the strain with extra envZ copies than for PMI6pRKgfp. Although no differences were detected in the infection process when EE-7 derivative strains were analysed separately (Figure 5d,e), upon co-inoculation, a higher amount of green-tagged bacteria (EE7envZ+gfp) was observed in intracellular zones and in infection threads, suggesting a higher efficiency on these early symbiosis processes of the strain with extra envZ copies (Figure 5f). Similar observations resulted from the analysis of co-inoculation of PMI6envZ+gfp and PMI6pRKrfp strains (Figure 5c), which agree with the results obtained by the single inoculation analysis. Since strains were inoculated in a 1:1 mixture, the co-inoculation analysis indicates that competitiveness at root hair-infection stage is higher in the strains overexpressing envZ.
Figure 5.
Confocal laser scanning micrographs showing the initial infection process of chickpea roots inoculated with rhizobia tagged with green and red fluorescent protein (GFP and RFP, respectively). Single inoculation experiments: (a) PMI6pRKgfp, (b) PMI6envZ+gfp, (d) EE7pRKgfp and (e) EE7envZ+gfp strains on roots stained with propidium iodide, at 4 days after inoculation. Co-inoculation experiments: (c) PMI6pRKrfp and PMI6envZ+gfp; (f) EE7pRKrfp and EE7envZ+gfp strains on chickpea roots stained with calcofluor white, at 6 days after inoculation. Root hair curling (white arrows); infection threads (gray arrows); caps on root hairs tips (blue arrows), rhizobia attached to the roots (square) and empty vector strains sharing the same intercellular space with those harboring pRKenvZ (ellipses) are shown. Scale bars: 75 μm (a, b, d and e); 50 μm (c and f).
Figure 5.
Confocal laser scanning micrographs showing the initial infection process of chickpea roots inoculated with rhizobia tagged with green and red fluorescent protein (GFP and RFP, respectively). Single inoculation experiments: (a) PMI6pRKgfp, (b) PMI6envZ+gfp, (d) EE7pRKgfp and (e) EE7envZ+gfp strains on roots stained with propidium iodide, at 4 days after inoculation. Co-inoculation experiments: (c) PMI6pRKrfp and PMI6envZ+gfp; (f) EE7pRKrfp and EE7envZ+gfp strains on chickpea roots stained with calcofluor white, at 6 days after inoculation. Root hair curling (white arrows); infection threads (gray arrows); caps on root hairs tips (blue arrows), rhizobia attached to the roots (square) and empty vector strains sharing the same intercellular space with those harboring pRKenvZ (ellipses) are shown. Scale bars: 75 μm (a, b, d and e); 50 μm (c and f).
4. Discussion
The EnvZ/OmpR two-component regulatory system regulates expression of outer membrane proteins (porins) in response to osmotic stress [6,57,58]. In addition, several studies in different Gammaproteobacteria, showed that mutations in envZ or ompR alter the expression levels of several other genes related with a large range of functions, as for example genes related to virulence in pathogens, fatty acid uptake, exopolysaccharide (EPS) production, peptide transportation and flagella production [9,15,16]. To our knowledge, the potential role of this gene in the rhizobia-legume symbiosis was not previously investigated; nonetheless, the existence of this system is mentioned in previous works [29,30]. Since most of the enumerated functions regulated by this system are relevant for the rhizobia interaction with their hosts, the aim of present study was to evaluate the effects of extra envZ copies on the symbiotic performance.
The identification of the envZ-like gene from M. mediterraneum UPM-Ca36T was confirmed by bioinformatic analyses. The envZ genes from the Mesorhizobium species analysed share over 80% of sequence similarity and form a monophyletic group, in a distinct lineage from the well characterised envZ gene from E. coli. Nevertheless, for all these different species, the same conserved protein domains were found, namely a HAMP linker domain, which is a domain common to histidine kinases and other chemoreceptors [28,59] and a histidine kinase domain, which includes several characteristic motifs, including a histidine residue that acts as the sensor part of the protein, since it can be phosphorylated [60]. Interestingly, in most Mesorhizobium, the envZ/ompR genes are encoded near choline and glycine betaine ABC transporter genes, which probably also play a role in osmoprotection, since the uptake of these important osmoprotectants is a common strategy to respond to salinity conditions in rhizobia [61].
As a first approach, to evaluate if extra envZ copies could improve the symbiotic plant parameters, a pot assay was performed using chickpea plants inoculated with the five mesorhizobia strains modified with the expression vectors pRKenvZ and pRK415. Only two strains, PMI6envZ+ and EE7envZ+, showed a significant improvement in the symbiotic effectiveness, compared to the respective control strain, harboring the pRK415 plasmid. Plants inoculated with PMI6envZ+ and EE7envZ+ showed a higher shoot and root dry weights. Furthermore, EE7envZ+ induced the development of larger nodules (higher average weight per nodule) compared to EE7pRK. Under salinity conditions, this outperformance of chickpea plants inoculated with the strains overexpressing envZ was no longer observed. The fact that closely related rhizobia respond differently to the overexpression of the same gene, was previously reported in a study using highly conserved symbiosis genes [42]. This is probably due to differences in genomic background, since the expression levels using the pRK415 vector are likely to be consistence among different strains [42].
As the main function of EnvZ is to sense the fluctuations of osmotic conditions in the environment [6], the modified strains were submitted to salinity tolerance assays. In liquid medium, the PMI6envZ+ strain grew faster than the control. Accordingly, this strain also showed a higher ability to grow in solid medium supplemented with 1.5% NaCl. Contrary to PMI6envZ+, EE-7 harboring extra envZ copies did not show an enhanced tolerance to salinity. These finding support the same trend displayed by the symbiotic performance trial, i. e., the heterologous expression of envZ may affect different mechanisms in different strains.
To investigate which aspects of the symbiosis were affected by the expression of extra envZ copies, further studies were performed with EE-7 and PMI-6 modified strains. In general, a successful rhizobia penetration through the infection thread requires the biosynthesis of functional Nod factors and exopolysaccharides (EPS) [62,63]. EPS may be important in lowering the legume immune response during rhizobia invasion [64,65]. Nevertheless, it is known that different rhizobia strains may produce different EPS, not only in terms of composition but also in terms of quantity [66]. Since the mucoid phenotype indicates the ability of rhizobia to produce EPS [67], the highest mucoid phenotype observed in bacteria modified with extra envZ copies may have improved the ability of these bacteria to infect the root hairs, particularly in the case of PMI6envZ+. S. meliloti 1021 overexpressing exoR, which encodes an enzyme involved in succinoglycan biosynthesis, enhanced the symbiosis with Medicago truncatula [64]. In addition, the two-component regulatory system ExoS-ChvI, which positively regulates the synthesis of succinoglycan in S. meliloti, shares high similarity to the ChvG-ChvI, a member of the EnvZ-OmpR family [28]. EPS influence the symbiosis process in other rhizobia species as well; for example, R. leguminosarum bv trifolii strains overproducing EPS increased the shoot fresh and dry weight and the number of nodules on clovers roots as well as nodule occupancy [68]. In addition, the observed influence of EnvZ in EPS production is in line with previous studies reporting that the EnvZ/OmpR two-component system regulates polysaccharide production in Erwinia amylovora and Salmonella typhi and outer membrane proteins in Xenorhabdus nematophilus [16,18,69].
To monitor the time-course of nodule development, chickpea seedlings were inoculated and grown in hydroponic conditions. Although the pot assay showed no significant difference in the number of nodules (NN) developed in chickpea plants inoculated with PMI6envZ+ compared to plants inoculated with PMI6pRK, the hydroponic conditions showed that the NN is significantly higher in plants inoculated with the strain overexpressing envZ in the later period of the experiment. This apparent discrepancy between the results is most likely due the shorter duration of the hydroponic assay (5 weeks), compared with the pot assay (8 weeks). These results suggest that the higher NN in plants inoculated with PMI6envZ+ might be temporary and yet benefit the plant in terms of N-fixation in this period and contribute to a better symbiotic performance. The nodulation kinetics was also evaluated in plants inoculated with EE-7 strains harboring pRK415 or pRKenvZ plasmids. Although plants inoculated with EE7envZ+ showed more nodules during the whole experiment, this difference was statistically not significant, and in agreement with the results obtained in the pot assay.
The qualitative analysis of the early steps of the infection process showed that roots inoculated with PMI6envZ+gfp displayed a higher number of infection threads than the ones inoculated with the corresponding control strain (harboring pRK415). These results suggest that EnvZ may contribute to a more effective infection and this difference at the beginning of the symbiotic process may account for the earlier nodulation and higher NN observed in hydroponic conditions in plants inoculated with PMI6envZ+. Upon single inoculation, no evident alteration in the number of infection threads was detected for EE7envZ+, when compared with the empty vector strain. Nevertheless, upon co-inoculation a higher density of EE7envZ+cells attached on the intracellular zone was observed and this may contribute to the higher NN observed in plants inoculated with this strain in hydroponic conditions.
Several studies in different Gammaproteobacteria species have shown that EnvZ/OmpR system may influence bacteria motility, biofilm formation and flagella production [9,11,16,70,71,72]. Several studies also showed that motile rhizobia are more efficient in nodulation [73,74] or more competitive on the plant root colonization [55,75] than non-motile ones. Therefore, improvement of motility, as observed in the swimming assay for PMI6envZ+ could account to the more pronounced enhancement of the infection ability and competitiveness of PMI6envZ+.
Previous studies revealed that EnvZ/OmpR may regulate secretion system (SS) genes in pathogenic bacteria, such as Salmonella typhimurium, Pseudomonas syringae and Yersinia enterocolitica [14,21,22]. Although there is no report showing the EnvZ/OmpR influence on SS in rhizobia, it is known that different types of SS are present in this group of bacteria and their importance for the symbiosis has been previously shown [76,77]. One possible explanation for the SE improvement of the strains overexpressing envZ could be the influence of EnvZ/OmpR in the regulation of SS in rhizobia.
5. Conclusions
This study represents the first report on the involvement of the envZ-like genes in the rhizobia-legume symbiosis. Nevertheless, another two-component system from the same family, ExoS/ChvI, has been previously reported to be essential to the establishment of an effective symbiosis in S. meliloti [78,79,80]. Extra copies of envZ in PMI-6 strain improved some important parameters, such as colonization and infection, number of nodules in an early phase of the symbiosis and mucoid phenotype (EPS production). These same parameters evaluated in EE7envZ+ and EE7pRK did not reveal differences as evident as the ones revealed by PMI6 and its derivative. Overall, these results showed that the envZ extra-copies influenced the mesorhizobia-chickpea symbiosis at different stages. Further studies are required to better understand all the mechanisms regulated by the EnvZ/OmpR system in the rhizobia-legume interactions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Genomic context of envZ or envZ-like genes in several Mesorhizobium species from different hosts, including other legume species from the Cicer genus. The annotation available on the NCBI database was used. envZ or envZ-like as well as the upstream response regulator are marked in green. For the envZ or envZ-like genes, the protein domain analysis is shown, with HAMP standing for HAMP domain and HIS_KIN standing for histidine kinase domain; Figure S2: Neighbour-Joining phylogenetic tree based on 16S rRNA gene sequences. Tamura-Nei model with gamma distributed rate among sites was used. Percentage of bootstrap support (1000 replicates) is indicated on internal branches. Scale bar indicates 0.02 substitutions per site; Figure S3: Swimming ability of (a) PMI6pRK, (b) PMI6envZ+, (c) EE7pRK and (d) EE7envZ+ strains in TY medium containing 0.25% agar; Figure S4: Mucoid phenotype in YMA plates supplemented with Congo Red. (a) PMI6pRK, (b) PMI6envZ+, (c) EE7pRK and (d) EE7envZ+. Table S1: List of bacterial strains used for phylogenetic analysis and the corresponding envZ or envZ-like gene and 16S rRNA gene locus tag. References [81,82,83,84,85,86,87,88,89,90,91,92,93] are cited in Supplementary Materials.
Author Contributions
Conceptualization, S.O.; methodology, J.R.d.-S., E.M. and A.A.; formal analysis, J.R.d.-S., E.M. and A.A.; writing—original draft preparation, J.R.d.-S.; writing—review and editing, E.M. and A.A.; funding acquisition, S.O. All authors have read and agreed to the published version of the manuscript, with the exception of S.O.†.
Funding
This research was funded by FEDER Funds through the Operational Program for Competitiveness Factors—COMPETE and by National Funds through FCT—Foundation for Science and Technology under the Project UIDB/05183 and the Strategic Project UID/AGR/00115/2013, Project nº FCOMP-01-0124-FEDER-028316 (PTDC/BIA-EVF/4158/2012), project POCI-01-0145-FEDER-016810 (PTDC/AGR-PRO/2978/2014) and InAlentejo ALENT-07-0262-FEDER-001871. This publication includes experimental work carried out in the scope of the PhD fellowship granted to J.R. da-Silva (1254-13-8), and the corresponding funding from CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) is acknowledged.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Acknowledgments
We thank Doctor Alvaro Peix from IRNASA-CSIC for providing pMP4661 and G. Mariano for technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tschauner, K.; Hörnschemeyer, P.; Müller, V.S.; Hunke, S. Dynamic interaction between the CpxA sensor kinase and the periplasmic accessory protein CpxP mediates signal recognition in E. Coli. PLoS ONE 2014, 9, e107383. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, M.; Epstein, B.; Badgley, B.D.; Unno, T.; Xu, L.; Reese, J.; Gyaneshwar, P.; Denny, R.; Mudge, J.; Bharti, A.K.; et al. Comparative genomics of the core and accessory genomes of 48 Sinorhizobium strains comprising five genospecies. Genome Biol 2013, 14, R17. [Google Scholar] [CrossRef] [PubMed]
- Tseng, T.-T.; Tyler, B.M.; Setubal, J.C. Protein secretion systems in bacterial-host associations, and their description in the gene ontology. BMC Microbiol 2009, 9, S2. [Google Scholar] [CrossRef] [PubMed]
- Boor, K.J. Bacterial Stress Responses: What doesn’t kill them can make Them stronger. PLoS Biol 2006, 4, e23. [Google Scholar] [CrossRef]
- Foo, Y.H.; Gao, Y.; Zhang, H.; Kenney, L.J. Cytoplasmic sensing by the inner membrane histidine kinase EnvZ. Progress in Biophysics and Molecular Biology 2015, 118, 119–129. [Google Scholar] [CrossRef]
- Wang, L.C.; Morgan, L.K.; Godakumbura, P.; Kenney, L.J.; Anand, G.S. The inner membrane histidine kinase EnvZ senses osmolality via helix-coil transitions in the cytoplasm: The mechanism of EnvZ osmosensing. The EMBO Journal 2012, 31, 2648–2659. [Google Scholar] [CrossRef]
- Pasqua, M.; Coluccia, M.; Eguchi, Y.; Okajima, T.; Grossi, M.; Prosseda, G.; Utsumi, R.; Colonna, B. Roles of two-component signal transduction systems in Shigella virulence. Biomolecules 2022, 12, 1321. [Google Scholar] [CrossRef]
- Yoshida, T.; Cai, S.J.; Inouye, M. Interaction of EnvZ, a sensory histidine kinase, with phosphorylated OmpR, the cognate response regulator. Molecular Microbiology 2002, 46, 1283–1294. [Google Scholar] [CrossRef]
- Yuan, J.; Wei, B.; Shi, M.; Gao, H. Functional assessment of EnvZ/OmpR two-component system in Shewanella oneidensis. PLoS ONE 2011, 6, e23701. [Google Scholar] [CrossRef]
- Mattison, K.; Kenney, L.J. Phosphorylation alters the interaction of the response regulator OmpR with its sensor kinase EnvZ. Journal of Biological Chemistry 2002, 277, 11143–11148. [Google Scholar] [CrossRef]
- Oshima, T.; Aiba, H.; Masuda, Y.; Kanaya, S.; Sugiura, M.; Wanner, B.L.; Mori, H.; Mizuno, T. Transcriptome analysis of all two-component regulatory system mutants of Escherichia coli K-12. Molecular Microbiology 2002, 46, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Zhang, Y.; Han, Y.; Yang, L.; Liu, X.; Guo, Z.; Tan, Y.; Huang, X.; Zhou, D.; Yang, R. Phenotypic and transcriptional analysis of the osmotic regulator OmpR in Yersinia pestis. BMC Microbiol 2011, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Gerken, H.; Shetty, D.; Kern, B.; Kenney, L.J.; Misra, R. Effects of pleiotropic ompR and envZ alleles of Escherichia Coli on envelope stress and antibiotic sensitivity. J Bacteriol 2024, 206, e00172–24. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Oropeza, R.; Kenney, L.J. Dual regulation by phospho-OmpR of ssrA/B Gene Expression in Salmonella Pathogenicity Island 2. Molecular Microbiology 2003, 48, 1131–1143. [Google Scholar] [CrossRef]
- Ji, X.; Shi, A.; Wang, J.; Zhang, B.; Hu, Y.; Lv, H.; Wu, J.; Sun, Y.; Liu, J.-M.; Zhang, Y.; et al. EnvZ/OmpR Controls Protein expression and modifications in Cronobacter sakazakii for virulence and environmental resilience. J. Agric. Food Chem. 2024, 72, 18697–18707. [Google Scholar] [CrossRef]
- Li, W.; Ancona, V.; Zhao, Y. Co-regulation of polysaccharide production, motility, and expression of type III secretion Genes by EnvZ/OmpR and GrrS/GrrA systems in Erwinia amylovora. Mol Genet Genomics 2014, 289, 63–75. [Google Scholar] [CrossRef]
- Mills, S.D.; Ruschkowski, S.R.; Stein, M.A.; Finlay, B.B. Trafficking of porin-deficient Salmonella typhimurium mutants inside HeLa cells: ompR and envZ Mutants are defective for the formation of Salmonella -Induced Filaments. Infect Immun 1998, 66, 1806–1811. [Google Scholar] [CrossRef]
- Pickard, D.; Li, J.; Roberts, M.; Maskell, D.; Hone, D.; Levine, M.; Dougan, G.; Chatfield, S. Characterization of defined ompR mutants of Salmonella typhi: ompR is involved in the regulation of Vi polysaccharide expression. Infect Immun 1994, 62, 3984–3993. [Google Scholar] [CrossRef]
- Shin, S.; Park, C. Modulation of flagellar expression in Escherichia coli by acetyl phosphate and the osmoregulator OmpR. J Bacteriol 1995, 177, 4696–4702. [Google Scholar] [CrossRef]
- Leopold, A.V.; Verkhusha, V.V. Engineering signalling pathways in mammalian cells. Nat. Biomed. Eng 2024, 8, 1523–1539. [Google Scholar] [CrossRef]
- Feng, X.; Walthers, D.; Oropeza, R.; Kenney, L.J. The response regulator SsrB activates transcription and binds to a region overlapping OmpR binding sites at Salmonella Pathogenicity island 2. Molecular Microbiology 2004, 54, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Nieckarz, M.; Jaworska, K.; Raczkowska, A.; Brzostek, K. The regulatory circuit underlying downregulation of a type III secretion system in Yersinia enterocolitica by transcription factor OmpR. IJMS 2022, 23, 4758. [Google Scholar] [CrossRef] [PubMed]
- Masson-Boivin, C.; Sachs, J.L. Symbiotic nitrogen fixation by Rhizobia — the roots of a success story. Current Opinion in Plant Biology 2018, 44, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.K.; Rajput, V.D.; Kumari, A.; Espinosa-Saiz, D.; Menendez, E.; Minkina, T.; Dwivedi, P.; Mandzhieva, S. Plant growth-promoting Rhizobacteria: A potential bio-asset for restoration of degraded soil and crop productivity with sustainable emerging techniques. Environ geochem Health 2023, 45, 9321–9344. [Google Scholar] [CrossRef]
- Geddes, B.; Kearsley, J.; Morton, R.; DiCenzo, C.; Finan, T. The genomes of Rhizobia. In Advances in Botanical Research; Frendo, P., Frugier, F., Masson-Boivin, C., Eds.; Elsevier: Canada, 2020; Volume 94, pp. 1–348. [Google Scholar]
- da-Silva, J.R.; Alexandre, A.; Brígido, C.; Oliveira, S. Can stress response genes be used to improve the symbiotic Performance of Rhizobia? AIMS Microbiology 2017, 3, 365–382. [Google Scholar] [CrossRef]
- Paço, A.; da-Silva, J.R.; Eliziário, F.; Brígido, C.; Oliveira, S.; Alexandre, A. traG Gene Is Conserved across Mesorhizobium Spp. able to nodulate the same host plant and expressed in response to root exudates. BioMed Research International 2019, 2019, 1–13. [Google Scholar] [CrossRef]
- Cheng, H.-P.; Walker, G.C. Succinoglycan production by Rhizobium Meliloti is regulated through the ExoS-ChvI two-component regulatory system. J Bacteriol 1998, 180, 20–26. [Google Scholar] [CrossRef]
- David, M.; Daveran, M.-L.; Batut, J.; Dedieu, A.; Domergue, O.; Ghai, J.; Hertig, C.; Boistard, P.; Kahn, D. Cascade regulation of nif gene expression in Rhizobium meliloti. Cell 1988, 54, 671–683. [Google Scholar] [CrossRef]
- Ronson, C.W.; Astwood, P.M.; Nixon, T.B.; Ausubel, F.M. Deduced products of c4-dicarboxylate transport regulatory genes of Rhizobium leguminosarum are homologous to nitrogen regulatory gene products. Nucl Acids Res 1987, 15, 7921–7934. [Google Scholar] [CrossRef]
- Acosta-Jurado, S.; Fuentes-Romero, F.; Ruiz-Sainz, J.-E.; Janczarek, M.; Vinardell, J.-M. Rhizobial Exopolysaccharides: Genetic Regulation of Their Synthesis and Relevance in Symbiosis with Legumes. IJMS 2021, 22, 6233. [Google Scholar] [CrossRef]
- Alexandre, A.; Brígido, C.; Laranjo, M.; Rodrigues, S.; Oliveira, S. Survey of chickpea Rhizobia diversity in Portugal reveals the predominance of species distinct from Mesorhizobium ciceri and Mesorhizobium Mediterraneum. Microb Ecol 2009, 58, 930–941. [Google Scholar] [CrossRef] [PubMed]
- Brígido, C.; Alexandre, A.; Oliveira, S. Transcriptional analysis of major chaperone Genes in salt-tolerant and salt-sensitive Mesorhizobia. Microbiological Research 2012, 167, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Laranjo, M.; Alexandre, A.; Rivas, R.; Velázquez, E.; Young, J.P.W.; Oliveira, S. Chickpea rhizobia symbiosis genes are highly conserved across multiple Mesorhizobium species: Mesorhizobia symbiosis genes are conserved. FEMS Microbiology Ecology 2008, 66, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Nour, S.M.; Cleyet-Marel, J.-C.; Normand, P.; Fernandez, M.P. Genomic heterogeneity of strains nodulating Chickpeas (Cicer Arietinum L.) and description of Rhizobium mediterraneum Sp. Nov. International Journal of Systematic Bacteriology 1995, 45, 640–648. [Google Scholar] [CrossRef]
- Somasegaran, P.; Hoben, H. Handbook for Rhizobia; Springer-Verlag: New York, 1994. [Google Scholar]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual; 3rd ed.; Cold Spring Harbor Laboratory: New York, 2001. [Google Scholar]
- Finan, T.M.; Kunkel, B.; De Vos, G.F.; Signer, E.R. Second symbiotic megaplasmid in Rhizobium meliloti carrying exopolysaccharide and thiamine synthesis genes. J Bacteriol 1986, 167, 66–72. [Google Scholar] [CrossRef]
- Keen, N.T.; Tamaki, S.; Kobayashi, D.; Trollinger, D. Improved broad-host-range plasmids for DNA cloning in gram-negative bacteria. Gene 1988, 70, 191–197. [Google Scholar] [CrossRef]
- García-Fraile, P.; Carro, L.; Robledo, M.; Ramírez-Bahena, M.-H.; Flores-Félix, J.-D.; Fernández, M.T.; Mateos, P.F.; Rivas, R.; Igual, J.M.; Martínez-Molina, E.; et al. Rhizobium promotes non-legumes growth and quality in several production steps: towards a biofertilization of edible raw vegetables healthy for humans. PLoS ONE 2012, 7, e38122. [Google Scholar] [CrossRef]
- Bloemberg, G.V.; Wijfjes, A.H.M.; Lamers, G.E.M.; Stuurman, N.; Lugtenberg, B.J.J. Simultaneous imaging of Pseudomonas Fluorescens WCS365 populations expressing three different autofluorescent proteins in the rhizosphere: New Perspectives for Studying Microbial Communities. MPMI 2000, 13, 1170–1176. [Google Scholar] [CrossRef]
- da-Silva, J.R.; Menéndez, E.; Eliziário, F.; Mateos, P.F.; Alexandre, A.; Oliveira, S. Heterologous expression of nifA or nodD genes improves chickpea-Mesorhizobium symbiotic performance. Plant Soil 2019, 436, 607–621. [Google Scholar] [CrossRef]
- De Castro, E.; Sigrist, C.J.A.; Gattiker, A.; Bulliard, V.; Langendijk-Genevaux, P.S.; Gasteiger, E.; Bairoch, A.; Hulo, N. ScanProsite: Detection of PROSITE signature matches and ProRule-Associated functional and structural residues in proteins. Nucleic Acids Research 2006, 34, W362–W365. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Molecular Biology and Evolution 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. MODELTEST: testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Rivas, R.; Velázquez, E.; Valverde, A.; Mateos, P.F.; Martínez-Molina, E. A two primers random amplified polymorphic DNA procedure to obtain polymerase chain reaction fingerprints of bacterial species. Electrophoresis 2001, 22, 1086–1089. [Google Scholar] [CrossRef]
- Alexandre, A.; Brígido, C.; Laranjo, M.; Rodrigues, S.; Oliveira, S. Survey of chickpea Rhizobia diversity in Portugal reveals the predominance of species distinct from Mesorhizobium ciceri and Mesorhizobium mediterraneum. Microb Ecol 2009, 58, 930–941. [Google Scholar] [CrossRef]
- Broughton, W.J.; Dilworth, M.J. Control of leghaemoglobin synthesis in snake beans. Biochemical Journal 1971, 125, 1075–1080. [Google Scholar] [CrossRef]
- Brígido, C.; Nascimento, F.X.; Duan, J.; Glick, B.R.; Oliveira, S. Expression of an exogenous 1-Aminocyclopropane-1-Carboxylate deaminase gene in Mesorhizobium Spp. reduces the negative effects of salt Stress in chickpea. FEMS Microbiol Lett, 2013. [Google Scholar] [CrossRef]
- Gibson, A. evaluation of nitrogen fixation by legumes in the greenhouse and growth chamber. In Symbiotic Nitrogen Fixation Technology; Elkan, G., Ed.; Marcel Dekker: New Yor, 1987; pp. 321–363. [Google Scholar]
- Rouws, L.F.M.; Simões-Araújo, J.L.; Hemerly, A.S.; Baldani, J.I. Validation of a Tn5 transposon mutagenesis system for Gluconacetobacter diazotrophicus through characterization of a flagellar mutant. Arch Microbiol 2008, 189, 397–405. [Google Scholar] [CrossRef]
- Robledo, M.; Rivera, L.; Jiménez-Zurdo, J.I.; Rivas, R.; Dazzo, F.; Velázquez, E.; Martínez-Molina, E.; Hirsch, A.M.; Mateos, P.F. Role of Rhizobium endoglucanase CelC2 in cellulose biosynthesis and biofilm formation on plant roots and abiotic surfaces. Microb Cell Fact 2012, 11, 125. [Google Scholar] [CrossRef]
- Brígido, C.; Robledo, M.; Menéndez, E.; Mateos, P.F.; Oliveira, S. A ClpB chaperone knockout mutant of Mesorhizobium Ciceri shows a delay in the root nodulation of chickpea plants. MPMI 2012, 25, 1594–1604. [Google Scholar] [CrossRef]
- Flores-Félix, J.; Menéndez, E.; Marcos-García, M.; Celador-Lera, L.; Rivas, R. Calcofluor white, an alternative to propidium iodide for plant tissues staining in studies of root colonization by fluorescent-tagged Rhizobia. JABB 2015, 2, 65–70. [Google Scholar] [CrossRef]
- Caetano-Anolles, G.; Wall, L.; De Micheli, A.; Macchi, E.; Baur, W.; Favelukes, G. Role of motility and chemotaxis in efficiency of nodulation by Rhizobium meliloti. Plant Physiol 1988, 1228–1235. [Google Scholar] [CrossRef]
- Gay-Fraret, J.; Ardissone, S.; Kambara, K.; Broughton, W.J.; Deakin, W.J.; Quéré, A. Cyclic-β-glucans of Rhizobium (Sinorhizobium) sp. strain NGR234 are required for hypo-osmotic adaptation, motility, and efficient symbiosis with host plants. FEMS Microbiol Lett 2012, 333, 28–36. [Google Scholar] [CrossRef]
- Alphen, W.V.; Lugtenberg, B. Influence of osmolarity of the growth medium on the outer membrane protein Pattern of Escherichia coli. J Bacteriol 1977, 131, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Bontemps-Gallo, S.; Madec, E.; Robbe-Masselot, C.; Souche, E.; Dondeyne, J.; Lacroix, J.-M. The opgC gene is required for OPGs succinylation and is osmoregulated through RcsCDB and EnvZ/OmpR in the phytopathogen Dickeya dadantii. Sci Rep 2016, 6, 19619. [Google Scholar] [CrossRef] [PubMed]
- Aravind, L.; Ponting, C.P. The cytoplasmic helical linker domain of receptor histidine kinase and methyl-accepting proteins is common to many prokaryotic signalling proteins. FEMS Microbiology Letters 1999, 176, 111–116. [Google Scholar] [CrossRef] [PubMed]
- West, A.H.; Stock, A.M. Histidine kinases and response regulator proteins in two-component signaling systems. Trends in Biochemical Sciences 2001, 26, 369–376. [Google Scholar] [CrossRef]
- Boncompagni, E.; Østerås, M.; Poggi, M.-C.; Le Rudulier, D. Occurrence of choline and glycine betaine uptake and metabolism in the family Rhizobiaceae and their roles in osmoprotection. Appl Environ Microbiol 1999, 65, 2072–2077. [Google Scholar] [CrossRef]
- Jones, K.M.; Kobayashi, H.; Davies, B.W.; Taga, M.E.; Walker, G.C. How Rhizobial symbionts invade plants: the Sinorhizobium–medicago model. Nat Rev Microbiol 2007, 5, 619–633. [Google Scholar] [CrossRef]
- Klein, S. Interaction of Nod and Exo Rhizobium Meliloti in Alfalfa Nodulation. MPMI 1988, 1, 94. [Google Scholar] [CrossRef]
- Jones, K.M. Increased production of the exopolysaccharide succinoglycan enhances Sinorhizobium meliloti 1021 symbiosis with the host plant Medicago truncatula. J Bacteriol 2012, 194, 4322–4331. [Google Scholar] [CrossRef]
- Jones, K.M.; Sharopova, N.; Lohar, D.P.; Zhang, J.Q.; VandenBosch, K.A.; Walker, G.C. Differential response of the plant Medicago truncatula to its symbiont Sinorhizobium meliloti or an exopolysaccharide-deficient mutant. Proc. Natl. Acad. Sci. U.S.A. 2008, 105, 704–709. [Google Scholar] [CrossRef]
- Ghosh, P.K.; Maiti, T.K. Structure of Extracellular Polysaccharides (EPS) produced by rhizobia and their functions in legume–bacteria symbiosis: — a review. Achievements in the Life Sciences 2016, 10, 136–143. [Google Scholar] [CrossRef]
- Staehelin, C.; Forsberg, L.S.; D’Haeze, W.; Gao, M.-Y.; Carlson, R.W.; Xie, Z.-P.; Pellock, B.J.; Jones, K.M.; Walker, G.C.; Streit, W.R.; et al. Exo-oligosaccharides of Rhizobium sp. strain NGR234 are required for symbiosis with various legumes. J Bacteriol 2006, 188, 6168–6178. [Google Scholar] [CrossRef] [PubMed]
- Janczarek, M.; Jaroszuk-Ściseł, J.; Skorupska, A. Multiple copies of rosR and pssA genes enhance exopolysaccharide production, symbiotic competitiveness and clover nodulation in Rhizobium leguminosarum bv. Trifolii. Antonie van Leeuwenhoek 2009, 96, 471–486. [Google Scholar] [CrossRef] [PubMed]
- Forst, S.; Tabatabai, N. Role of the histidine kinase, envz, in the production of outer membrane proteins in the symbiotic-pathogenic bacterium Xenorhabdus nematophilus. Appl Environ Microbiol 1997, 962–968. [Google Scholar] [CrossRef]
- Kim, D.; Boylan, B.; George, N.; Forst, S. Inactivation of ompR promotes precocious swarming and flhDC expression in Xenorhabdus Nematophila. J Bacteriol 2003, 185, 5290–5294. [Google Scholar] [CrossRef]
- Prüß, B.M. Involvement of two-component signaling on bacterial motility and biofilm development. J Bacteriol 2017, 199. [Google Scholar] [CrossRef]
- Tipton, K.A.; Rather, P.N. An ompR-envZ Two-component system ortholog regulates phase variation, osmotic tolerance, motility, and virulence in Acinetobacter baumannii strain AB5075. J Bacteriol 2017, 199. [Google Scholar] [CrossRef]
- Soby, S.; Bergman, K. Motility and chemotaxis of Rhizobium Meliloti in soil. Appl Environ Microbiol 1983, 46, 995–998. [Google Scholar] [CrossRef]
- Zheng, H.; Mao, Y.; Teng, J.; Zhu, Q.; Ling, J.; Zhong, Z. Flagellar-dependent motility in Mesorhizobium tianshanense is involved in the early stage of plant host interaction: Study of an flgE Mutant. Curr Microbiol 2015, 70, 219–227. [Google Scholar] [CrossRef]
- Mellor, H.Y.; Glenn, A.R.; Arwas, R.; Dilworth, M.J. Symbiotic and competitive properties of motility mutants of Rhizobium trifolii TA1. Arch. Microbiol. 1987, 148, 34–39. [Google Scholar] [CrossRef]
- Hubber, A.M.; Sullivan, J.T.; Ronson, C.W. Symbiosis-induced cascade regulation of the Mesorhizobium loti R7A VirB/D4 type IV Ssecretion system. MPMI 2007, 20, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, S.; Kaneko, T.; Sato, S.; Saeki, K. Hijacking of leguminous nodulation signaling by the Rhizobial type III secretion system. Proc. Natl. Acad. Sci. U.S.A. 2013, 110, 17131–17136. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, L.; Dimmick, K.A.; Fleming, J.S.; Charles, T.C. Null mutations in Sinorhizobium meliloti exoS and chvI demonstrate the importance of this two-component regulatory system for symbiosis. Molecular Microbiology 2009, 74, 1223–1237. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.J.; Fisher, R.F.; Perovich, V.M.; Sabio, E.A.; Long, S.R. Identification of direct transcriptional target genes of ExoS/ChvI two-component signaling in Sinorhizobium Meliloti. J Bacteriol 2009, 191, 6833–6842. [Google Scholar] [CrossRef]
- Soto, M.J.; Sanjuán, J.; Olivares, J. Rhizobia and plant-pathogenic bacteria: common infection weapons. Microbiology 2006, 152, 3167–3174. [Google Scholar] [CrossRef]
- Greenlon, A.; Chang, P.L.; Damtew, Z.M.; Muleta, A.; Carrasquilla-Garcia, N.; Kim, D.; Nguyen, H.P.; Suryawanshi, V.; Krieg, C.P.; Yadav, S.K.; et al. Global-level population genomics reveals differential effects of geography and phylogeny on horizontal gene transfer in soil bacteria. Proc. Natl. Acad. Sci. U.S.A. 2019, 116, 15200–15209. [Google Scholar] [CrossRef]
- Diouf, F.; Diouf, D.; Klonowska, A.; Le Queré, A.; Bakhoum, N.; Fall, D.; Neyra, M.; Parrinello, H.; Diouf, M.; Ndoye, I.; et al. Genetic and genomic diversity studies of Acacia symbionts in Senegal reveal new species of Mesorhizobium with a putative geographical pattern. PLoS ONE 2015, 10, e0117667. [Google Scholar] [CrossRef]
- Nandasena, K.; Yates, R.; Tiwari, R.; O’Hara, G.; Howieson, J.; Ninawi, M.; Chertkov, O.; Detter, C.; Tapia, R.; Han, S.; et al. Complete genome sequence of Mesorhizobium ciceri Bv. Biserrulae type strain (WSM1271T). Stand. Genomic Sci. 2013, 9, 462–472. [Google Scholar] [CrossRef]
- Wang, S.; Hao, B.; Li, J.; Gu, H.; Peng, J.; Xie, F.; Zhao, X.; Frech, C.; Chen, N.; Ma, B.; et al. Whole-genome sequencing of Mesorhizobium huakuii 7653R provides molecular insights into host specificity and symbiosis island dynamics. BMC Genomics 2014, 15, 440. [Google Scholar] [CrossRef]
- Kelly, S.; Sullivan, J.; Ronson, C.; Tian, R.; Bräu, L.; Davenport, K.; Daligault, H.; Erkkila, T.; Goodwin, L.; Gu, W.; et al. Genome sequence of the Lotus Spp. microsymbiont Mesorhizobium loti strain NZP2037. Stand in Genomic Sci 2014, 9, 7. [Google Scholar] [CrossRef]
- Wang, X.; Luo, Y.; Liu, D.; Wang, J.; Wei, S.; Zhao, L. Complete genome sequence of the Robinia pseudoacacia L. symbiont Mesorhizobium amorphae CCNWGS0123. Stand in Genomic Sci 2018, 13, 18. [Google Scholar] [CrossRef] [PubMed]
- Haskett, T.; Wang, P.; Ramsay, J.; O’Hara, G.; Reeve, W.; Howieson, J.; Terpolilli, J. Complete genome sequence of Mesorhizobium ciceri strain CC1192, an efficient nitrogen-fixing microsymbiont of Cicer Arietinum. Genome Announc 2016, 4, e00516–16. [Google Scholar] [CrossRef] [PubMed]
- Greenlon, A.; Chang, P.L.; Damtew, Z.M.; Muleta, A.; Carrasquilla-Garcia, N.; Kim, D.; Nguyen, H.P.; Suryawanshi, V.; Krieg, C.P.; Yadav, S.K.; et al. Global-level population genomics reveals differential effects of geography and phylogeny on horizontal gene transfer in soil bacteria. Proc. Natl. Acad. Sci. U.S.A. 2019, 116, 15200–15209. [Google Scholar] [CrossRef] [PubMed]
- Schmeisser, C.; Liesegang, H.; Krysciak, D.; Bakkou, N.; Le Quéré, A.; Wollherr, A.; Heinemeyer, I.; Morgenstern, B.; Pommerening-Röser, A.; Flores, M.; et al. Rhizobium sp. strain NGR234 possesses a remarkable number of secretion systems. Appl Environ Microbiol 2009, 75, 4035–4045. [Google Scholar] [CrossRef]
- Wood, D.W.; Setubal, J.C.; Kaul, R.; Monks, D.E.; Kitajima, J.P.; Okura, V.K.; Zhou, Y.; Chen, L.; Wood, G.E.; Almeida, N.F.; et al. The genome of the natural genetic engineer Agrobacterium Tumefaciens C58. Science 2001, 294, 2317–2323. [Google Scholar] [CrossRef]
- Buell, C.R.; Joardar, V.; Lindeberg, M.; Selengut, J.; Paulsen, I.T.; Gwinn, M.L.; Dodson, R.J.; Deboy, R.T.; Durkin, A.S.; Kolonay, J.F.; et al. The complete genome sequence of the Arabidopsis and tomato pathogen Pseudomonas Syringae Pv. Tomato DC3000. Proc. Natl. Acad. Sci. U.S.A. 2003, 100, 10181–10186. [Google Scholar] [CrossRef]
- Hayashi, K.; Morooka, N.; Yamamoto, Y.; Fujita, K.; Isono, K.; Choi, S.; Ohtsubo, E.; Baba, T.; Wanner, B.L.; Mori, H.; et al. Highly accurate genome sequences of Escherichia Coli K-12 Strains MG1655 and W3110. Molecular Systems Biology 2006, 2, 2006.0007. [Google Scholar] [CrossRef]
- Jin, Q. Genome Sequence of Shigella flexneri 2a: Insights into pathogenicity through comparison with genomes of Escherichia coli K12 and O157. Nucleic Acids Research 2002, 30, 4432–4441. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



