
Submitted:
06 April 2025
Posted:
08 April 2025
You are already at the latest version
Abstract
Fomes fomentarius (L.) Fr., known for its medicinal properties since the 15th century, has been traditionally used across Europe, including the Balkan region, for tinder, spiritual practices, wound healing, and hemostasis. This study investigates the mycochemical profile of three Balkan strains, focusing on micro- and macroelements, polyamines (PAs), and phenolic compounds, alongside the bioactivity of hot water (H2O), chloroform (CHCl3), hydroethanolic (EtOH), and hydromethanolic (MeOH) extracts. Micro- and macroelements were analyzed via atomic absorption spectrophotometry (AAS), while PAs were quantified using HPLC-FD. Phenolic profiles were determined by LC-MS/MS, with total phenolic content (TPC) assessed using the Folin-Ciocalteu method. Antioxidant potential was evaluated through DPPH, ABTS, NO scavenging, lipid peroxidation inhibition, and FRAP assays, in addition to anti-acetylcholinesterase and antiproliferative activity assessments. This study represents the first investigation of polyamine profiles in F. fomentarius, with total PA levels ranging from 32.67 to 2910.09 nmol/g d.w. The Bosnian strain exhibited the highest PA levels, with spermidine (SPD) concentrations between 899.96 to 2910.09 nmol/g d.w. LC-MS/MS analysis identified several bioactive phenolics, including amentoflavone, baicalein, chrysoeriol, esculetin, and scopoletin, which are reported here for the first time in this fungus. H2O and EtOH extracts from Croatia and Serbia showed higher TPC content, correlating with notable antioxidant activity. EtOH and MeOH extracts demonstrated significant anti-acetylcholinesterase and antiproliferative activities, emphasizing their medicinal potential. Our findings highlight the therapeutic potential of polar extracts from Balkan F. fomentarius as natural antioxidants with applications in oxidative stress-related chronic diseases.
Keywords:
Balkan region
; bioactive compounds
; Fomes fomentarius
; neuroprotective agents
; oxidative stress
; phenolics
; polyamines
1. Introduction
Mushrooms have long been valued for their medicinal properties due to the presence of bioactive compounds with significant health-promoting effects [1,2]. These properties include antidiabetic, anti-inflammatory, antiproliferative, antimicrobial activities, antioxidant, and neuroprotective activities [3,4,5,6,7,8,9]. Over 2,000 species of edible and medicinal mushrooms have been documented, many of which have being used in traditional medicine across various regions, particularly in Asia. Key species like Fomes fomentarius, Cordyceps sinensis, Ganoderma lucidum, and Lentinula edodes, Lenzites betulinus have been integral to folk remedies in countries such as China, Japan, Korea, and India, for the treatment of ailments including inflammation, cancer, and neurodegenerative diseases [1,2,10,11,12,13].
Reactive oxygen species (ROS), known for their role in oxidative stress, have been linked to numerous diseases, including cancer and Alzheimer's disease [14]. Alzheimer's disease, characterized by a deficiency in acetylcholine, is often targeted using acetylcholinesterase (AChE) inhibitors. Mushrooms, due to their antioxidant potential, offer a promising alternative approach for managing oxidative stress and related pathological conditions [3]. Mushrooms, with their notable antioxidant potential, present a promising approach for managing oxidative stress and related conditions [3]. While their anti-AChE and antioxidant properties have been largely attributed to polysaccharides and phenolic compounds, secondary metabolites like polyamines (PAs) remain underexplored [8]. PA degradation contributes to oxidative stress by generating reactive aldehydes and ROS, partly due to reduced PA levels. However, PAs themselves act as potent antioxidants and ROS scavengers [15]. The age-related decline in PA levels has long been linked to cognitive and behavioral impairments [16]. This raises the hypothesis that maintaining SPD levels during aging may support longevity, with PA levels potentially increased through dietary supplementation [17].
Among these, F. fomentarius (L.) Fr., known as the "Iceman’s mushroom" due to its historical discovery alongside a 5,000-year-old mummy, has demonstrated various therapeutic properties, including antioxidant, anti-inflammatory, anti-AChE, and antiproliferative effects [10,18,19].
F. fomentarius is widely distributed across the northern hemisphere, growing on hardwood trees such as beech, birch, and oak [12,20,21]. The medicinal potential of its fruiting bodies and mycelia has been extensively explored, with recent studies highlighting its rich mycochemical composition, including phenolics, benzofurans, coumarins, triterpenoids, and polysaccharides like β-glucans [10,12,16]. These compounds are linked to the mushroom’s antiproliferative and antioxidant activities, with specific polyphenols and triterpenoids playing a crucial role in modulating oxidative stress and inhibiting cell proliferation [11,22]. Additionally, studies have shown that its bioactive compounds can reduce AChE activity, making F. fomentarius a potential candidate for neuroprotective therapies in diseases like Alzheimer's disease [3,23].
Ethnological studies reveal that Fomes fomentarius (tinder fungus or hoof fungus) has a longstanding role in traditional medicine and practical uses. It is among the earliest documented medicinal mushrooms, used for fire-starting, preservation, first aid, insect repellents, and spiritual rituals [10,24,25]. Fruiting bodies were employed in ceremonial smoking rituals in Germany and Austria and in West Siberia, where the Khanty burned them to ward off the deceased's influence [10].
F. fomentarius was used to treat bladder disorders, dysmenorrhoea, haemorrhoids, gastroenteric issues, inflammation, pain, and various cancers [10,22,25]. It featured in a Japanese anticancer beverage [18] and is noted in the traditional pharmacopeias of China, India, Korea, and Hungary for addressing cirrhosis, gastrointestinal disorders, inflammation, and oral ulcers [12,24]. Referred to as "mykes" in ancient Greece, it first appeared during Hippocrates’ era for cauterizing wounds and treating inflamed organs [24].
The Okanagan-Colville Indians used it for rheumatism, and it was widely applied as a styptic, earning the name “agaric of the surgeons” [25]. Hieronymus Bock recommended it as an emetic against mushroom poisoning in the 15th century. Moreover, it was used globally as an iodine-infused dressing for wounds and burns and sold in pharmacies as styptic bandages [10,25].
Despite its widespread use globally, data on its traditional usage in the Balkan region are limited, except one study conducted to gather information on harvesting methods, timing, storage techniques, preparations, as well as the ethnomycological utilization of its fruiting bodies [20]. Unique preparations of this species were traditionally prevalent in Corund, Lupeni, Odorheiu Secuiesc, and Sumuleu Ciuc in Transylvania (Romania). In other areas of the Balkan region, there is a lack of data indicating the use of this species for pharmacological purposes although it was noted that people in certain regions, such as Croatia and Bulgaria, were aware of its health benefits [26,27]. Alongside very limited information regarding the ethnomedicinal use of this species in Serbia, Živković et al. [28] conducted the first ethnomycological study, documenting the use of 85 fungal species from 28 families for food and medicinal purposes, among which, F. fomentarius (known as "trud" in Serbian) is used preventatively in the form of tea, (one spoon of dried and ground mushroom is boiled in 1 L of water for 30 min). Collectively, this suggests a mycophilic consumption behavior among people in the Balkan region, excluding Slovenia [29].
In this study, we conducted a comparative analysis of three strains of F. fomentarius collected from the Balkan region, focusing on their mycochemical composition and biological activities. This research aims to provide a comprehensive understanding of the chemical diversity and biological potential of these unexplored strains, which could offer significant benefits in the prevention and treatment of oxidative stress-related diseases such as cancer and neurodegenerative disorders.
2. Materials and Methods
2.1. Mushroom Material
Fruiting bodies of F. fomentarius (Ph. Basidiomycota, Cl. Agaricomycetes, O. Polyporales, Fam. Polyporaceae, Gen. Fomes) were collected in January of 2018 (National Park “Štrbački Buk”, Donji Lapac, Croatia, FC), April of 2018 (National Park “Fruška Gora”, Serbia, FS) and October of 2018 (Vrelo Bosne, Sarajevo, Bosnia and Herzegovina, FB) (Figure 1).
Identification of mushroom species was carried out by studying the fungal morphology macroscopically (color, shape, size and hyphae) and microscopically, using specific identification keys as described in Rašeta et al. [6]. The samples were identified and authenticated by Dr. Eleonora Čapelja, and stored and deposited at the ProFungi Laboratory, Department of Biology and Ecology, Faculty of Sciences, University of Novi Sad under the following numbers (FC 12-00725, FS 12-00726 and FB 12-00662).
2.2. Extracts Preparation
The fruiting bodies were sectioned, air-dried, and stored in dark containers at room temperature. The detailed procedure for extract preparation can be found in Rašeta et al. [30] where hot water (H2O), hydroethanolic (EtOH), chloroform (CHCl3), and hydromethanolic (MeOH) extracts were prepared according to the methods outlined by Rašeta et al. [4] and Gąsecka et al. [31]. The powdered fruiting bodies were mixed with the solvents at ratio of 1:10. Samples were macerated (IKA KS 4000i control) for 8 h, centrifuged (Sigma 3–30K, Burlington, Massachusetts, United States) at 3999 ×g for 15 min at 4 °C, then filtered through Whatman No.4 paper (Maidstone, Kent, UK) and evaporated under vacuum (Büchi R-210; Buchi Labortechnik AG, Flawil, Switzerland) at 40 ºC to dryness, except for the H2O extract, which was lyophilized (ChristAlpha, Martin Christ Gefriertrocknungsanlagen GmbH, Osterode am Harz, Germany). The dry residues were dissolved in dimethyl sulfoxide (DMSO). All prepared extracts were stored at 4 ºC for further use.
2.3. Mycochemical Characterization
2.3.1. Atomic Absorption Spectrophotometry (AAS)
Macro- (Ca and Mg) and microelements (Cr, Cu, Mn, Fe, and Zn) powdered fungal samples were quantified using atomic absorption spectrophotometry (FS AAS240/GTA120, Agilent) following the method described by Kebert et al. [32].
2.3.2. HPLC-FD Determination of Selected Polyamines Content
The PA profile and content of three main PAs, putrescine (PUT), SPD, and spermine (SPM) were determined in mushroom material by high-performance liquid chromatography coupled with fluorescent detector (HPLC-FD) using a method described by Scaramagli et al. [33], after derivatization of PAs with dansyl-chloride as a pre-treatment.
2.3.3. LC/MS-MS Quantification of Phenolic Compounds and Total Phenolic Content (TPC) Determination
To quantify the selected phenolic compounds and quinic acid in the analyzed extracts, we utilized the liquid chromatography coupled with tandem mass-spectrometric detection (LC-MS/MS) technique in accordance with the protocol established by Orčić et al. [34]. The concentrations of standard phenolic compounds and quinic acid were determined by referencing a calibration curve generated from a series of dilutions of a standard mixture. Data were collected in dynamic MRM mode, with peak areas quantified using MassHunter software. Calibration curves were generated using OriginPro. Detailed retention times and MS parameters are provided in Table S1.
The total phenolic content (TPC) was measured using the Folin–Ciocalteu (FC) method, as described by Singleton et al. [35]. This approach relies on the spectrophotometric detection of phenolic compounds through their reaction with the FC reagent, forming a colored complex. Absorbance was measured at 760 nm, and TPC was expressed as milligrams of gallic acid equivalents per gram of dry weight (mg GAE/g d.w.), based on a standard calibration curve.
2.3.4. Determination of Total Carbohydrate Content
The total carbohydrate content (TCC) was determined using the phenol–sulfuric acid method, following the procedure outlined by Rašeta et al. [6]. This method involved the use of concentrated H₂SO₄ and 5% phenol. Absorbance was recorded at 490 nm for glucose, and a calibration curve was generated to quantify the TC in the fungal extracts. Results were expressed as milligrams of glucose equivalents per gram of dry weight (mg GluE/g d.w.).
2.4. Biological Activities
2.4.1. Determination of the Antioxidant Activity
Antioxidant activity was evaluated through a series of standard radical scavenging assays, including DPPH [36], ABTS [37], and NO radical scavenging [38]. Additionally, Fe²⁺/ascorbate-induced lipid peroxidation was assessed [39], and the reducing power of the fungal extracts was measured using the FRAP assay [40].
In the DPPH assay, a mixture of 60 μL of 90 µM DPPH reagent, 180 μL of MeOH, and 10 μL of extract/standard was incubated in the dark at room temperature for 30 minutes, and absorbance was measured at 515 nm [36]. For the ABTS assay, 290 μL of ABTS reagent (7 mM ABTS, 2.45 mM K₂S₂O₈) was mixed with 10 μL of extract/standard, incubated at room temperature for 5 minutes, and absorbance was recorded at 734 nm [37]. The NO radical scavenging assay involved mixing 15 μL of extract/standard, 250 μL of sodium nitroprusside (10 mmol/L), and 250 μL of phosphate buffer (pH 7.4). After 90 minutes of light exposure, 500 μL of Griess reagent was added, and the absorbance was measured at 546 nm [38]. Lipid peroxidation inhibition was assessed using a TBA assay with polyunsaturated fatty acids derived from linseed oil (69.7% linolenic acid, 13.5% linoleic acid). Fatty acids were emulsified in phosphate buffer containing 0.25% Tween-80, sonicated for 1 hour, and mixed with FeSO₄, ascorbic acid, and test samples (0.13–232 mg/mL). After a 1-hour incubation at 37°C, EDTA and a TBA mixture were added, followed by heating, centrifugation, and absorbance measurement of malondialdehyde (MDA) at 532 nm [39]. The FRAP assay involved adding 10 μL of extract/standard to 225 μL of FRAP reagent (TPTZ, FeCl₃, acetate buffer) and 22.5 μL of distilled water (dH2O), with absorbance recorded at 593 nm after 6 min [40].
The radical scavenging capacity (RSC) for DPPH and lipid peroxidation inhibition is expressed as IC₅₀ values (μg/mL), whereas for NO radicals, RSC is expressed as IC₂₅ values (μg/mL). For the ABTS assay, results were derived from a Trolox calibration curve and expressed as milligrams of Trolox equivalents per gram of dry weight (mg TE/g d.w.). For the FRAP assay, the reduction potential was calibrated using ascorbic acid, with results expressed as milligrams of ascorbic acid equivalents per gram of dry weight (mg AAE/g d.w.). Propyl gallate (PG) was used as the positive control in all assays.
2.4.2. Determination of Anti-Acetylcholinesterase Activity (Anti-AChE)
2.4.3. Determination of Antiproliferative Activity
Cell culturing
Human breast cancer cell lines (MCF7, T47D, and MDA-MB-231), a cervical (HeLa), and an ovarian cancer cell line (A2780) were purchased from ECACC (European Collection of Cell Cultures, Salisbury, UK), while SiHa cells (cervical cancer) were obtained from ATCC (American Tissue Culture Collection, Manassas, Virginia, USA). Cells were cultivated in a minimal essential medium supplemented with 10% fetal bovine serum, 1% nonessential amino acids, and an antibiotic-antimycotic mixture. All media and supplements were purchased from Lonza Group Ltd. (Basel, Switzerland). The cells were cultivated at 37 °C in a humidified atmosphere containing 5% CO2.
Antiproliferative activity (MTT assay)
The growth-inhibitory activity of the extracts was determined by the MTT method against a panel of human cancer cell lines of gynecological origin [40].
2.5. Statistical Analysis
Three different strains of F. fomentarius were utilized, and each experiment was conducted three times, except for the examination of antiproliferative activity, which was performed twice. The outcomes were reported as average values with either standard deviation (SD) or standard error of the mean (SEM), depending on the antiproliferative assay. One-way analysis of variance (ANOVA) and Tukey's HSD Test were carried out to test for any significant differences between the means; the mean values of antioxidant activities between two extracts or two treatments were analyzed by an independent-samples t-test. Correlations were obtained by Pearson correlation coefficient in bivariate correlations. Differences between means at 5% (p < 0.05) level were considered significant. For defining the variability of chemical composition and biological activity between the investigated extracts, principal component analysis (PCA) was applied by PAST version 3.16 [44].
3. Results and Discussion
3.1. Mycochemical Profile
3.1.1. Macro- and Microelements Content
Mineral composition analysis of F. fomentarius included two macroelements (Mg and Ca) and five microelements (Cr, Cu, Mn, Fe and Zn). The results are presented in Figure 2a,b.
The Serbian strain (FS) exhibited the highest mean concentrations of macroelements, with Ca at 9.93 ± 0.21 mg/g d.w. and Mg at 2.28 ± 0.17 mg/g d.w. Across all strains, Ca was found in significantly higher concentrations (7.30–9.93 mg/g d.w.) compared to Mg, which ranged from 1.42 to 2.28 mg/g d.w. This pattern suggests that Ca, abundant in the wood substrate of this parasitic/saprotrophic species, is more readily absorbed and utilized by F. fomentarius strains. These findings are consistent with the work of Karaman and Matavulj [45], who reported higher Ca concentrations in lignicolous mushroom substrates compared to wild-growing fungi.
Significant accumulation of Zn, Fe, and Cu was observed in the analyzed F. fomentarius strains, while Mn and Cr were present at lower concentrations (Mn: 5.24–8.05 mg/kg d.w.; Cr: 2.05–3.12 mg/kg d.w., p < 0.01). Among the microelements, Fe was the most abundant, with the Croatian strain (FC) containing the highest concentration (48.07 ± 0.54 mg/kg d.w.), followed by similar levels in the FB and FS strains (27.36–27.78 mg/kg d.w.). The significant Fe accumulation aligns with previous reports of hyperaccumulation in lignicolous species such as Meripilus giganteus and F. fomentarius [45]. The second highest concentration was observed for Cu (25.90–34.27 mg/kg d.w.), with Zn also present in substantial amounts, vital for human physiological functions [46]. The FC strain exhibited the highest levels of both Cu and Zn, consistent with prior findings on Cu and Zn accumulation in fungi, particularly lignicolous and tericolous species.45 Mn and Cr were detected in lower amounts, ranging from 5.24 to 8.05 mg/kg d.w. for Mn and 2.05 to 3.12 mg/kg d.w. for Cr (p < 0.01). These levels are comparable to previously published data on edible mushrooms from Serbia [45] and India [46].
The higher concentrations of Ca and Mg in Balkan F. fomentarius compared to Turkish strains [47], suggest its potential as a valuable source of essential minerals crucial for bone health, particularly in mitigating osteoporosis risk [48]. Additionally, the Fe content, aligning with recommended daily intake levels, highlights its potential as a dietary supplement [46].
Although Serbian researchers have extensively studied mineral compositions in various lignicolous and tericolous mushroom species [4,45,49,50,51], this study represents the first comparative analysis of F. fomentarius strains from the Balkan region. These results underscore the role of F. fomentarius as an effective accumulator of essential biogenic minerals, particularly Fe, while highlighting significant inter-strain variability influenced by substrate and geography.
Environmental factors, including habitat, geographical position, and substrate type, play a significant role in the mineral composition of F. fomentarius [4,45]. Additional factors, such as ecotype, genotype, growth conditions, and age of the mycelium, also influence mineral uptake [46,52]. For example, Balkan strains exhibited significantly higher macroelement concentrations than strains from Uşak, Turkey, where Ca and Mg levels were reported at 0.1814 mg/g d.w. and 0.1816 mg/g d.w., respectively [47].
These findings emphasize the adaptability of F. fomentarius to absorb minerals directly from the wood substrate and indirectly from the surrounding soil, supporting its reputation as one of the most effective accumulators of Fe among fungi [45,53]. The high mineral content of F. fomentarius further validates its potential as a source of bioactive compounds and food supplements, particularly for mitigating mineral deficiencies and promoting bone health. However, its hard texture makes it unsuitable for direct consumption, necessitating its use in processed forms such as extracts or supplements [25].
3.1.2. Polyamines Content
Despite extensive research on PAs in plant- and animal-derived foods, data on PAs in wild-growing mushrooms has been notably lacking [54]. In mushrooms, SPD is generally the dominant PA, while putrescine (PUT) and SPD are associated with a range of morphological, physiological, and metabolic processes. SPM, however, is not consistently present in mushrooms [15].
This study is the first to investigate the PA profiles in F. fomentarius, analyzing three strains collected from the Balkan region. The results confirm the presence of all three PAs, with total levels ranging from 32.67 to 2910.09 nmol/g d.w. (Figure 3). Among the strains, the FB strain exhibited the highest concentrations, while the FC strain had the lowest.
SPD was found to be the predominant PA across the strains, with concentrations ranging from 899.96 to 2910.09 nmol/g d.w., except in the FC strain, where it was significantly lower (270.65 ± 14.72 nmol/g d.w.). The FB strain had the highest SPD levels (2910.09 ± 196.45 nmol/g d.w.), suggesting that SPD is ubiquitous in F. fomentarius, though its concentration varies significantly across strains. SPM was present in the lowest concentrations (32.67–54.35 nmol/g d.w.), with FB again showing the highest levels (54.35 ± 3.02 nmol/g d.w.). In contrast, PUT was predominant in the FC strain (278.56 ± 5.58 nmol/g d.w.), with concentrations higher than those reported for some edible wild mushrooms [54,55].
The results align with the observations by Dadáková et al. [54], who noted significantly higher levels of SPD in mushrooms compared to other foods. The SPD concentrations in F. fomentarius determined in this study far exceed those reported for other mushroom species, such as A. bisporus, L. edodes, and Pleurotus spp., which range between 32.30–71.30 mg/kg d.w. [55], and A. bisporus and X. badius, with values of 404–604 mg/kg d.w [54]. SPD plays a crucial role in cellular metabolism and growth, and its abundance in mushrooms has been previously documented as among the highest found in food [54,56].
To the best of our knowledge, the SPD levels detected in this study represent the highest recorded in mushrooms. Based on these findings, F. fomentarius can be considered a valuable and untapped source of PAs, particularly SPD. According to the classification proposed by Kalač [56], the analyzed strains represent a significant reservoir of bioactive PAs. However, further studies are necessary to fully elucidate the roles of these compounds and to explore their potential applications as dietary supplements or therapeutic agents.
3.1.3. Phenolic Content Determined by LC-MS/MS
The LC-MS/MS analysis of F. fomentarius extracts revealed quinic acid (379.28–996.78 μg/g d.w.) and scopoletin (209.05–853.07 μg/g d.w.) as the predominant compounds, particularly in the FS and FB strains (Table 1).
These compounds were identified in F. fomentarius for the first time. The highest concentrations were found in the FB strain's MeOH extract (quinic acid, 996.78 μg/g d.w.) and the FS strain's EtOH extract (scopoletin, 853.07 μg/g d.w.). Quinic acid was detected in almost all extracts, except for the unpolar CHCl₃ extracts and the FS EtOH extract. Additionally, chlorogenic acid, a derivative of quinic acid, was identified at 6.74 μg/g d.w. in the MeOH extract, although its concentration was much lower than previously reported by Bal et al. [57].
Among the flavonoids, baicalein was found in the highest concentrations in the FS and FC strains (44.63–213.87 μg/g d.w.), while chrysoeriol and amentoflavone were also detected, marking their first identification in F. fomentarius. The FC strain's CHCl₃ extract was notable for having the highest concentration of baicalein and the exclusive presence of chrysoeriol and amentoflavone, indicating a unique phenolic profile.
To identify strain-specific compounds and evaluate the impact of solvents, Principal Component Analysis (PCA) was applied (Figure 4).
PCA revealed a variance of 34.5% for PC1 and 22.1% for PC2. Baicalein significantly contributed to the negative part of PC1, while protocatechuic acid had the greatest loading in the positive part. The FC strain clustered uniquely in the negative part of both axes, with the highest baicalein content, especially in the CHCl₃ extract, and exclusive presence of chrysoeriol and amentoflavone. In contrast, the FB MeOH extract, which was located in the positive parts of both axes, had the highest concentrations of all tested phenolic compounds, including quinic acid.
Hydroxycinnamic acids were more prevalent than hydroxybenzoic acids in the extracts, often found as glycosylated derivatives or esters of quinic acid, shikimic acid, or tartaric acid [58]. The presence of quinic acid and scopoletin in F. fomentarius was notable, as quinic acid had not been detected in previous studies [59]. Compared to other mushrooms such as Coprinus comatus and C. truncorum from Serbia [60], F. fomentarius contained significantly higher quinic acid levels, ranging from 1.59 to 215 μg/g d.w. Similarly, the scopoletin concentrations in F. fomentarius were much higher than those reported for C. comatus and C. truncorum (1.11–1.97 μg/g d.w.) in previous studies using the same method [34].
A total of six novel phenolic compounds were identified in F. fomentarius extracts—amentoflavone, baicalein, chrysoeriol, cinnamic acid, esculetin, and scopoletin, along with quinic acid—bringing the total number of phenolic compounds reported for this species to 19, as documented in various studies [23,57,59,61]. Comparative analyses indicate that the Serbian strain exhibited a greater diversity and quantity of phenolics compared to strains from Poland [61] and Turkey [23,57].
The phenolic content was highest in the FB and FS strains, while FC strain exhibited relatively lower phenolic levels. This disparity could be attributed to the presence of other bioactive or unexamined phenolics in the FC strain, along with factors such as geographical origin, climate differences, sampling periods, and the age of the fruiting bodies. For example, the FC strain was collected during the winter, which may have influenced its phenolic profile.
3.1.4. TPC and TCC Content
The total phenolic content (TPC) and total carbohydrate content (TCC) of F. fomentarius strains were evaluated across various extracts (Figure 5a,b).
Among the extracts, the highest TPC was observed in the FS strain's EtOH extract, with a value of 441.16 ± 1.85 mg GAE/g d.w., while the CHCl₃ extracts exhibited the lowest TPC, ranging from 14.30 to 72.55 mg GAE/g d.w. Similarly, the highest TCC was found in the FS strain's MeOH extract, with a concentration of 227.2 ± 0.116 mg GluE/g d.w., while the lowest values were recorded in CHCl₃ extracts (10.24–20.26 mg GluE/g d.w.). These results highlight the efficiency of polar solvents (EtOH and MeOH) in extracting phenolic compounds and carbohydrates, consistent with prior findings that non-polar extracts, primarily composed of triterpenoids and other non-polar compounds, contain lower levels of phenolics and carbohydrates.
When comparing the TPC values obtained in this study with the existing literature, the results were significantly higher. For example, Karaman et al. [59] reported TPC values of 43.06 ± 0.19 mg GAE/g d.w. and 82.54 ± 0.12 mg GAE/g d.w. for Serbian F. fomentarius samples prepared using EtOH and MeOH extracts [59], respectively, while the current study recorded considerably higher amounts in polar extracts. Similarly, Orhan and Üstün [3] reported a TPC of 47.29 ± 1.37 mg GAE/mg extract for 85% EtOH extracts of F. fomentarius. This study's findings align with Karadeniz et al. [62], who noted that EtOH is highly effective in phenolic extraction. However, disparities in TPC among studies may be attributed to various factors, including geographical origin, climatic conditions, harvesting location, extraction methods, mushroom developmental stage, and whether results are expressed on a dry or fresh weight basis [61].
Regarding TCC, limited data are available for F. fomentarius. The TCC values observed in this study were lower than those reported in previous studies. Vazirian et al. [18] recorded a total polysaccharide content of 533.0 ± 2.0 mg/g, while Deveci et al. [63] reported a total carbohydrate content of 73.31 ± 6.35%. Despite these discrepancies, the MeOH extracts in this study consistently exhibited the highest carbohydrate concentrations. The variability in TCC can be influenced by environmental factors such as geographical origin, climate, and the developmental stage of fruiting bodies [64]. Additionally, certain mushroom species are specific to particular regions due to variations in climate, soil composition, rainfall patterns, and seasonal changes. These environmental factors affect their growth, ultimately influencing both their fruiting bodies and mycelial development [65].
The results highlight the superior efficiency of polar solvents (MeOH and EtOH) in extracting both TPC and TCC, corroborating prior research [4,59,60]. This underscores the influence of solvent polarity on phenolic and carbohydrate extraction from F. fomentarius. Furthermore, the notable differences in TPC and TCC across strains and extraction methods suggest that geographic origin and substrate composition significantly impact the composition of phenolics and carbohydrates in F. fomentarius.
3.2. Antioxidant Activity
The antioxidant activities of F. fomentarius extracts were evaluated using DPPH, ABTS, NO scavenging, lipid peroxidation (LP) inhibition, and FRAP assays (Figure 6a–e).
The results, compared to the standard antioxidant propyl gallate (PG), revealed that polar extracts (EtOH, MeOH) demonstrated significantly higher antioxidant activity than nonpolar CHCl₃ extracts, with one notable exception: the FB strain’s CHCl₃ extract showed the strongest LP inhibition activity (IC50 = 11.88 ± 0.50 μg/mL) (Figure 6e). The FS strain’s EtOH extract exhibited the strongest DPPH scavenging activity (IC50 = 0.783 ± 0.021 μg/mL) (Figure 6a), while the FC strain’s EtOH extract demonstrated remarkable ABTS scavenging capacity (176.5 ± 0.464 mg TE/g d.w.) (Figure 6b). The FB strain’s MeOH extract showed the highest NO radical scavenging activity (IC25 = 0.267 ± 0.006 μg/mL) (Figure 6c), surpassing the activity of PG (IC25 = 0.957 ± 0.023 μg/mL). The FRAP assay results highlighted MeOH extracts as having the strongest reducing power (352.2–423.8 mg AAE/g d.w.), closely followed by the FB strain’s EtOH extract (362.8 mg AAE/g d.w.) (Figure 6d).
The DPPH scavenging activities of polar extracts (IC50 ranging from 0.783 to 31.40 μg/mL) were notably stronger than values reported in previous studies. For instance, Karaman et al. [59] reported a slightly higher IC50 for EtOH extracts (10.87 ± 0.11 μg/mL). Orhan and Üstün [3] recorded moderate DPPH inhibition (24.32–26.14%) in 85% EtOH extracts at higher concentrations (250–5000 μg/mL), while Ilyashenka [66] found considerably weaker activity in ethyl acetate (IC50 = 515.6 ± 100.5 μg/mL) and EtOH (IC50 = 751.2 ± 19.3 μg/mL) extracts. Similarly, the ABTS scavenging capacity of the FC strain’s EtOH extract (176.5 ± 0.464 mg TE/g d.w.) aligned with findings by Darkal et al. [67], who reported excellent ABTS activity (IC50 = 0.005–0.04% w/v) in F. fomentarius. In contrast, melanins extracted from F. fomentarius exhibited weaker antioxidant activity [68].
In the FRAP assay, MeOH extracts consistently exhibited the strongest reducing power, reflecting their robust electron-donating ability. These results corroborate findings by Darkal et al. [67], who demonstrated superior FRAP activity in milled F. fomentarius samples. Interestingly, the exceptional LP inhibition by the FB strain’s CHCl₃ extract, typically uncharacteristic for nonpolar extracts, warrants further investigation.
PCA of the antioxidant results (Figure 7 and Figure 8) revealed that the sample groups were influenced by the extraction solvents used.
MeOH extracts from all three strains clustered in the first quadrant, indicating high NO scavenging and FRAP activity alongside the highest TCC levels. CHCl₃ extracts grouped in the negative part of PC1, showing the lowest antioxidant potential and TPC/TCC values. FS and FC H₂O and EtOH extracts grouped together, reflecting moderate antioxidant activity.
The results highlight a strong correlation between antioxidant activity and the TPC and TCC of the extracts. This is consistent with prior research showing that the antioxidant potential of mushrooms is strongly associated with their phenolic content [4,5,58,59]. Phenolic compounds, such as quinic acid and scopoletin, play key roles in scavenging free radicals, with quinic acid known as a pro-metabolite of nicotinamide and tryptophan [69] and scopoletin acting as a superoxide scavenger [70].
In addition to phenolic compounds, carbohydrates, particularly polysaccharides, significantly contribute to the antioxidant potential of F. fomentarius. Polysaccharides are known to enhance immune responses and reduce oxidative stress through NO scavenging, as observed in samples with the highest TCC [6]. Their antioxidant activity is influenced by factors such as monosaccharide composition, molecular weight, and protein content [71].
Interestingly, PAs, particularly SPD, may also contribute to antioxidant effects (Figure 7a,b). The FB strain, with the highest PA content, exhibited notable NO scavenging activity. PAs act as free radical scavengers and metal chelators, mitigating oxidative damage, but can also generate pro-oxidative effects by producing hydrogen peroxide during catabolism [54,72]. This dual role underscores their complexity, as PAs may prevent chronic diseases while posing risks for cancer patients [56,73].
3.3. Anti-Acetylcholinesterase Activity
The neuroprotective potential of F. fomentarius extracts was evaluated by their potential to inhibit AChE, a critical therapeutic target for the treatment of Alzheimer's disease and other neurodegenerative disorders. AChE inhibition enhances acetylcholine levels in the brain, improving cholinergic functions and alleviating Alzheimer's disease symptoms [74]. FDA-approved AChE inhibitors, such as galantamine and rivastigmine, are primarily derived from natural sources [75,76,77], highlighting the importance of exploring bioactive compounds from fungi and other natural materials for therapeutic applications.
Among the analyzed extracts, the MeOH extract of the FB strain exhibited the strongest AChE inhibitory activity (IC50 = 0.099 mg/mL), followed by the EtOH (0.351 mg/mL) and CHCl₃ (1.003 mg/mL) extracts, while the H₂O extract had the weakest effect (IC50 = 2.881 mg/mL) (Figure 6f). These findings are consistent with prior studies showing weaker AChE inhibition in nonpolar extracts [8]. The potent activity of FB strain extracts strongly correlated with their higher levels of PAs, particularly SPD and SPM. PAs have been linked to neuroprotective effects, memory enhancement, and the regulation of acetylcholine metabolism in low doses [78,79].
The inhibitory activity of F. fomentarius extracts in this study was superior to previously reported values for related mushroom species. For example, Orhan and Üstün [3] reported modest AChE inhibition (15 ± 1.82%) in 85% EtOH extracts of F. fomentarius from Turkey, and Dundar et al. [23] found little to no activity in other Turkish strains. Similarly, extracts of Ganoderma lucidum from Slovenia, including MeOH and EtOH extracts, exhibited lower activity compared to the MeOH extracts in this study [80]. Recent findings by Rašeta et al. [8] also highlight significant AChE inhibition in polar extracts of other fungal species from Serbia, further supporting the potential of F. fomentarius as a source of AChE inhibitors.
PCA revealed strong associations between the levels of PAs, TPC, TCC, and AChE inhibition (Figure 8). PC1 and PC2 explained 93.19% of the variance, with distinct clustering of MeOH extracts in the first and third quadrants, alongside variables related to anti-AChE activity, DPPH, and FRAP assays. PUT and SPM clustered with anti-AChE activity, while SPD aligned with LP inhibition in the positive sections of both PCs. These patterns highlight the complementary roles of PAs, phenolic compounds, and carbohydrates in contributing to the neuroprotective potential of the extracts.
The MeOH extracts, particularly those from the FB strain, were rich in phenolic compounds such as quinic, p-hydroxybenzoic, cinnamic, p-coumaric, and caffeic acids, all of which have been identified as effective AChE inhibitors [81,82]. The higher TCC levels in MeOH extracts also contribute to their observed activity, as carbohydrates and polysaccharides have been shown to play roles in neuroprotection and antioxidant activity [83,84].
The role of PAs in AChE inhibition remains an area of growing interest. Studies suggest that PAs such as SPD and SPM can either inhibit or stimulate AChE activity depending on acetylcholine concentrations [85]. Low doses of PAs have been shown to support memory enhancement and regulate NMDA receptors, while higher concentrations may result in pro-oxidative effects through hydrogen peroxide production [86]. Furthermore, SPD and SPM have demonstrated neuroprotective properties by modulating acetylcholine metabolism and receptor assembly [87,88].
3.4. Antiproliferative Activity
The antiproliferative potential of F. fomentarius extracts was assessed using the MTT assay on human cancer cell lines, including breast (MDA-MB-231, T47D, MCF7), cervical (HeLa, SiHa), and ovarian (A2780) cancer cells. Extracts prepared with different solvents from the FB strain showed varied effects on the cancer cells (Table 2).
The MeOH extracts of FC and FS strains were further tested and found to significantly inhibit cancer cell viability, particularly in the A2780 (ovarian cancer) and MCF-7 (breast cancer) lines, although the FS extract exhibited generally stronger activity than the FC extract (Table 2). These results corroborate earlier studies, such as Shnyreva et al. [89], who demonstrated that F. fomentarius MeOH–CHCl3 extracts exhibited antiproliferative activity on solid tumor cell lines.
In comparison with previous studies, the F. fomentarius extracts demonstrated higher activity against the HeLa cell line than reported by Dundar et al. [23], with their MeOH extract showing an IC50 of 22.45 mg/mL. However, the results were lower compared to those reported by Kolundžić et al. [19], where cyclohexane, dichloromethane, MeOH, and H₂O extracts exhibited an IC50 range of 8.31–51.87 μg/mL. These findings suggest that F. fomentarius extracts from Serbia might have a specific mechanism of action that selectively targets cancer cells while being non-toxic to normal cells, as demonstrated in a 2016 study [19].
The antiproliferative activity of these extracts correlated with their TPC and TCC as presented in Figure S1. Flavonoids such as baicalein, chrysoeriol, and amentoflavone did not correlate with this activity, but coumarins like scopoletin and esculetin showed a low positive correlation. Cinnamic acid exhibited a negative correlation. However, other phenolic compounds, such as quinic acid, showed a moderate to high positive correlation with antiproliferative activity. Specifically, the MeOH FS extract, which had a high concentration of quinic acid, showed strong activity against the MDA-MB-231 breast cancer line, with a high correlation coefficient of 0.92.
Caffeic acid displayed the highest positive correlation (1.0) with the inhibition of MDA-MB-231 cell viability (Figure S1), underscoring its role as a potent antitumor agent, consistent with findings by Rosendahl et al. [90]. Conversely, flavonoids such as baicalein, chrysoeriol, and amentoflavone showed no significant correlation with antiproliferative activity, while coumarins (scopoletin and esculetin) demonstrated a low positive correlation. Interestingly, cinnamic acid exhibited a negative correlation with this activity.
The observed antiproliferative activity of F. fomentarius extracts may be attributed to the synergistic effects of their bioactive components. Phenolic acids, particularly caffeic and quinic acids, have well-documented antitumor properties. For instance, quinic acid is recognized for its ability to enhance immune responses and induce apoptosis in cancer cells [69]. Caffeic acid is known to inhibit key signaling pathways involved in tumor progression [90].
Moreover, the selectivity of F. fomentarius extracts in targeting cancer cells while sparing normal cells is of particular interest for therapeutic applications. These findings align with recent studies highlighting the potential of fungal melanins from F. fomentarius to display protective and differentiative properties in neuroblastoma cells [68].
4. Conclusions
This study provides the first global documentation of PA levels in F. fomentarius and identifies six previously unreported phenolic compounds along with quinic acid. The extracts of F. fomentarius demonstrated remarkable biological activities, including strong neuroprotective and antiproliferative effects, with MeOH extracts showing the highest potency. Rich in bioactive compounds such as PAs and phenolic acids, F. fomentarius strains and their polar extracts demonstrated significant AChE inhibitory activity, underscoring their potential for the treatment of Alzheimer’s disease and other neurodegenerative disorders. The presence of compounds like quinic, p-hydroxybenzoic, and caffeic acids significantly contributed to their neuroprotective effects.
These findings suggest that F. fomentarius could serve as a valuable source of natural bioactive compounds, with the potential to develop novel therapeutic agents for neurodegenerative diseases.
Additionally, the extracts showed significant antiproliferative activity against several human cancer cell lines, with the strongest effects observed in ovarian (A2780) and breast (MCF-7, MDA-MB-231) cancer cells. The strong correlation between the phenolic content, particularly quinic and caffeic acids, and the inhibition of cancer cell viability underscores the potential of F. fomentarius as a natural source of anticancer agents which supported its traditional usage in treatment of different cancer types. Furthermore, the selective action of these extracts, coupled with their non-toxic nature toward normal cells, warrants further research into their mechanisms of action and their potential development into targeted cancer therapies. Evidence-based ethnopharmacological research using modern approaches is essential to validate the traditional uses of F. fomentarius and promote its acceptance as a herbal medicine, potentially leading to innovative treatments for neurodegenerative disorders and cancer. Its use as a food supplement can also be recommended to reduce oxidative stress, support neuroprotection, and potentially lower the risk of certain cancers due to its mineral composition, bioactive compounds, including phenolic acids and polyamines.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1: Optimised dynamic MRM parameters for all quantified compounds in the analyzed F. fomentarius extracts. Figure S1: Pearson’s correlation matrix for the examined parameters in three tested F. fomentarius strains, showing the relationships between total carbohydrate content (TCC), total phenolic content (TPC), the quantified levels of 14 polyphenolics and quinic acid, and the antiproliferative activity assessed using the MTT assay.
Author Contributions
Conceptualization, M.R.; methodology, M.R., M.K. (Marko Kebert), D.P.Š., S.B., Á.E.K., and I.Z.; software, M.R., D.P.Š., M.K. (Marko Kebert), S.B., Á.E.K., and I.Z.; validation, M.R. and M.K. (Marko Kebert); formal analysis, M.R., M.K. (Marko Kebert), D.P.Š., S.B., and Á.E.K.; investigation, M.R.; resources, M.R., M.K. (Marko Kebert), I.Z., and S.J.Š.; data curation, M.R., M.K. (Marko Kebert), D.P.Š., Á.E.K., and I.Z.; writing—original draft preparation, M.R., M.K. (Marko Kebert), D.P.Š., J.M., and I.Z.; writing—review and editing, M.R., M.K. (Marko Kebert), D.P.Š., J.M., I.Z., M.K. (Maja Karaman), and S.J.Š.; visualization, M.R.; supervision, M.R.; project administration, M.R.; funding acquisition, M.R., and I.Z. All authors have read and agreed to the published version of the manuscript.
Funding
The authors gratefully acknowledge the financial support of the Ministry of Science, Technological Development and Innovation of the Republic of Serbia (Grants No. 451-03-137/2025-03/ 200125 & 451-03-136/2025-03/ 200125).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available within manuscript.
Acknowledgments
The authors would like to thank MSc Katarina Dragić for help in the experimental part of this study.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| FC | Fomes fomentarius strains sampled from Croatia |
| FB | Fomes fomentarius strains sampled from Bosnia and Herzegovina |
| FS | Fomes fomentarius strains sampled from Serbia |
| H2O | Hot water extract |
| EtOH | Hydroethanolic extract prepared with 70% ethanol |
| MeOH | Hydromethanolic extract prepared with 80% methanol |
| CHCl3 | Chloroform extract |
| ROS | Reactive oxygen species |
| AChE | Achetylcholinesterase |
| AAS | Atomic absorption spectrophotometry |
| HPLC-FD | High-performance liquid chromatography coupled with fluorescent detector |
| LC-MS/MS | Liquid chromatography coupled with tandem mass-spectrometric detection |
| PA | Polyamine |
| PUT | Putrescine |
| SPD | Spermidine |
| SPM | Spermine |
| TPC | Total phenolic content |
| TCC | Total carbohydrate content |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl radical, DPPH• |
| ABTS | 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid, ABTS•+ |
| NO | Nitric oxid radical, NO• |
| FRAP | Ferric reducing antioxidant power |
| FC | Folin-Ciocalteu |
| AAE | Ascorbic acid equivalents |
| GAE | Gallic acid equivalents |
| GluE | Glucose equivalents |
| TE | Trolox equivalents |
| d.w. | Dry weight |
| dH2O | Distilled water |
| PG | Propyl gallate |
| SD | Standard deviation |
| SEM | Standard error of the mean |
| PCA | Principal Component Analysis |
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three letter acronym |
| LD | Linear dichroism |
References
- Gafforov, Y.; Rašeta, M.; Rapior, S.; Yarasheva, M.; Wang, X.; Zhou, L.; Wan-Mohtar, W. A. A. Q. I.; Zafar, M.; Lim, Y. W.; Wang, M.; Abdullaev, B.; Bussmann, R. W.; Zengin, G.; Chen, J. Macrofungi as medicinal resources in Uzbekistan: biodiversity, ethnomycology, and ethnomedicinal practices. J. Fungi 2023, 9, 922. [Google Scholar] [CrossRef] [PubMed]
- Morales, D. Fomes fomentarius: an underexplored mushroom as source of bioactive compounds. Food Biosci. 2024, 61, 104781. [Google Scholar] [CrossRef]
- Orhan, I. E.; Üstün, O. Determination of total phenol content, antioxidant activity and acetylcholinesterase inhibition in selected mushrooms from Turkey. J. Food Compos. Anal. 2011, 24, 386–390. [Google Scholar] [CrossRef]
- Rašeta, M.; Karaman, M.; Jakšić, M.; Šibul, F.; Kebert, M.; Novaković, A.; Popović, M. Mineral composition, antioxidant and cytotoxic biopotentials of wild-growing Ganoderma species (Serbia): G. lucidum (Curtis) P. Karst vs. G. applanatum (Pers.) Pat. Int. J. Food Sci. Technol. 2016, 51, 2583–2590. [Google Scholar] [CrossRef]
- Rašeta, J.; Vrbaški, S.; Bošković, V. E.; Popović, M.; Mimica-Dukić, N. M.; Karaman, A. M. Comparison of antioxidant capacities of two Ganoderma lucidum strains of different geographical origins. Matica Srpska Journal of Natural Science 2017, 133, 209–219. [Google Scholar] [CrossRef]
- Rašeta, M.; Popović, M.; Beara, I.; Šibul, F.; Zengin, G.; Krstić, S.; Karaman, M. Anti-inflammatory, antioxidant and enzyme inhibition activities in correlation with mycochemical profile of selected indigenous Ganoderma spp. from Balkan region (Serbia). Chem. Biodivers. 2021, 18, e2000828. [Google Scholar] [CrossRef]
- Rašeta, M.; Mišković, J.; Čapelja, E.; Zapora, E.; Petrović Fabijan, A.; Knežević, P.; Karaman, M. Do Ganoderma species represent novel sources of phenolic based antimicrobial agents? Molecules 2023, 28, 3264. [Google Scholar] [CrossRef]
- Rašeta, M.; Kebert, M.; Mišković, J.; Rakić, M.; Kostić, S.; Čapelja, E.; Karaman, M. Polyamines in edible and medicinal fungi from Serbia: a novel perspective on neuroprotective properties. J. Fungi 2024, 10, 21. [Google Scholar] [CrossRef]
- Khojimatov, O.K.; Gafforov, Y.; Bussmann, R.W. Ethnobiology of Uzbekistan: Ethnomedicinal Knowledge of Mountain Communities, 1st ed.; Springer Nature, Basel, Switzerland, 2023; 1564 pages.
- Grienke, U.; Zöll, M.; Peintner, U.; Rollinger, J. M. European medicinal polypores-a modern view on traditional uses. J. Ethnopharmacol. 2014, 154, 564–583. [Google Scholar] [CrossRef]
- Kim, S. H.; Jakhar, R.; Kang, S. C. Apoptotic properties of polysaccharide isolated from fruiting bodies of medicinal mushroom Fomes fomentarius in human lung carcinoma cell line. Saudi J. Biol. Sci. 2015, 22, 484–490. [Google Scholar] [CrossRef]
- Gafforov, Y.; Kalitukha, L.; Tomšovský, M.; Angelini, P.; Venanzoni, R.; Angeles Flores, G.; Yarasheva, M.; Wan-Mohtar, W. A. A. Q. I.; Rapior, S. Fomes fomentarius (L.) Fr. – POLYPORACEAE. In Ethnobiology of Uzbekistan: Ethnomedicinal Knowledge of Mountain Communities, 1st ed.; Khojimatov, O. K., Gafforov, Y., Bussmann, R. W., Eds.; Springer Nature: Basel, Switzerland, 2023; pp. 1045–1063. [Google Scholar]
- Tubić Vukajlović, J.; Djordjević, K.; Tosti, T.; Simić, I.; Grbović, F.; Milošević-Djordjević, O. In vitro effect of Lenzites betulinus mushroom against therapy-induced DNA damage in peripheral blood lymphocytes of patients with acute coronary syndrome. J. Ethnopharmacol. 2024, 335, 118640. [Google Scholar] [CrossRef] [PubMed]
- Bardaweel, S. K.; Gul, M.; Alzweiri, M.; Ishaqat, A.; ALSalamat, H. A.; Bashatwah, R. M. Reactive oxygen species: The dual role in physiological and pathological conditions of the human body. Eurasian J. Med. 2018, 50, 193. [Google Scholar] [CrossRef] [PubMed]
- Rocha, R. O.; Wilson, R. Essential, deadly, enigmatic: polyamine metabolism and roles in fungal cells. Fungal Biol. Rev. 2019, 33, 45–57. [Google Scholar] [CrossRef]
- Makletsova, M. G.; Syatkin, S. P.; Poleshchuk, V. V.; Urazgildeeva, G. R.; Chigaleychik, L. A.; Sungrapova, C. Y.; Illarioshkin, S. N. Polyamines in Parkinson’s disease: their role in oxidative stress induction and protein aggregation. J. Neurol. Res. 2019, 9 (1–2), 1–7.
- Đorđievski, S.; Vukašinović, E. L.; Čelić, T. V.; Pihler, I.; Kebert, M.; Kojić, D.; Purać, J. Spermidine dietary supplementation and polyamines level in reference to survival and lifespan of honey bees. Sci. Rep. 2023, 13, 4329. [Google Scholar] [CrossRef]
- Vazirian, M.; Dianat, S.; Manayi, A.; Ziari, R.; Mousazadeh, A. R.; Habibi, E.; Saeidnia, S.; Amanzadeh, Y. Anti-inflammatory effect, total polysaccharide, total phenolics content and antioxidant activity of the aqueous extract of three basidiomycetes. Res. J. Pharmacogn. 2014, 1, 15–21. [Google Scholar]
- Kolundžić, M.; Grozdanić, N. Đ.; Dodevska, M.; Milenković, M.; Sisto, F.; Miani, A.; Farronato, G.; Kundaković, T. Antibacterial and cytotoxic activities of wild mushroom Fomes fomentarius (L.) Fr., Polyporaceae. Ind. Crops Prod. 2016, 79, 110–115. [Google Scholar] [CrossRef]
- Papp, N.; Rudolf, K.; Bencsik, T.; Czégényi, D. Ethnomycological use of Fomes fomentarius (L.) Fr. and Piptoporus betulinus (Bull.) P. Karst. in Transylvania, Romania. Genetic Resour. Crop Evol. 2017, 64, 101–111. [Google Scholar] [CrossRef]
- Pasailiuk, M. V. Total flavonoid content, lipid peroxidation and total antioxidant activity of Hericium coralloides, Fomes fomentarius and Schizophyllum commune cultivated by the method of direct confrontation. Ital. J. Mycol. 2020, 49, 25–37. [Google Scholar]
- Chen, W.; Zhao, Z.; Chen, S.; Li, Y. Optimization for the production of exopolysaccharide from Fomes fomentarius in submerged culture and its antitumor effect in vitro. Bioresour. Technol. 2008, 99, 3187–3194. [Google Scholar] [CrossRef]
- Dundar, A.; Okumuş, V.; Ozdemir, S.; Çelik, K. S.; Boğa, M. S.; Ozcagli, E. Determination of cytotoxic, anticholinesterase, antioxidant and antimicrobial activities of some wild mushroom species. Cogent Food Agric. 2016, 2, 1178060. [Google Scholar] [CrossRef]
- Gáper, J.; Gáperová, S.; Pristaš, P.; Náplavová, K. Medicinal value and taxonomy of the tinder polypore, Fomes fomentarius (Agaricomycetes): a review. Int. J. Med. Mushrooms 2016, 18, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Gründemann, C.; Reinhardt, J. K.; Lindequist, U. European medicinal mushrooms: do they have potential for modern medicine? - An update. Phytomedicine 2020, 66, 153131. [Google Scholar] [CrossRef] [PubMed]
- Tortić, M. Gljive kao lijek. Priroda: Popularni časopis hrvatskog prirodoslovnog društva 1961, 9, 272–275. [Google Scholar]
- Uzunov, B. A.; Stoyneva-Gärtner, M. P. Mushrooms and lichens in Bulgarian ethnomycology. J. Mycol. 2015, 2015. [Google Scholar] [CrossRef]
- Živković, J.; Ivanov, M.; Stojković, D.; Glamočlija, J. Ethnomycological investigation in Serbia: astonishing realm of mycomedicines and mycofood. J. Fungi 2021, 75, 349. [Google Scholar] [CrossRef]
- Peintner, U.; Schwarz, S.; Mešić, A.; Moreau, P.; Moreno, G.; Saviuc, P. Mycophilic or mycophobic? Legislation and guidelines on wild mushroom commerce reveal different consumption behaviour in European countries. PLoS ONE 2013, 8, e63926. [Google Scholar] [CrossRef]
- Rašeta, M.; Mišković, J.; Kebert, M.; Berežni, S.; Krstić, S.; Gojgić-Cvijović, G.; Pirker, T.; Bauer, R.; Karaman, M. Mycochemical profiles and bioactivities of Fistulina hepatica and Volvopluteus gloiocephalus from Serbia: antioxidant, enzyme inhibition, and cytotoxic potentials. Food Biosci. 2025, 66, 106221. [Google Scholar] [CrossRef]
- Gąsecka, M.; Mleczek, M.; Siwulski, M.; Niedzielski, P. Phenolic composition and antioxidant properties of Pleurotus ostreatus and Pleurotus eryngii enriched with selenium and zinc. Eur. Food Res. Technol. 2016, 242, 723–732. [Google Scholar] [CrossRef]
- Kebert, M.; Kostić, S.; Vuksanović, V.; Gavranović Markić, A.; Kiprovski, B.; Zorić, M.; Orlović, S. Metal- and organ-specific response to heavy metal-induced stress mediated by antioxidant enzymes’ activities, polyamines, and plant hormones levels in Populus deltoides. Plants 2022, 11, 3246. [Google Scholar] [CrossRef]
- Scaramagli, S.; Biondi, S.; Torrigiani, P. Methylglyoxal(bis-guanylhydrazone) inhibition of organogenesis is not due to S-adenosylmethionine decarboxylase inhibition/polyamine depletion in tobacco thin layers. Physiol. Plant. 1999, 107, 353–360. [Google Scholar] [CrossRef]
- Orčić, D.; Francišković, M.; Bekvalac, K.; Svirčev, E.; Beara, I.; Lesjak, M.; Mimica-Dukić, N. Quantitative determination of plant phenolics in Urtica dioica extracts by high-performance liquid chromatography coupled with tandem mass-spectrometric detection. Food Chem. 2014, 143, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V. L.; Orthofer, R.; Lamuela-Raventos, R. M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Espín, J. C.; Soler-Rivas, C.; Wichers, H. J. Characterization of the total free radical scavenger capacity of vegetable oils and oil fractions using 2,2-diphenyl-1-picrylhydrazyl radical. J. Agric. Food Chem. 2000, 48, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M. B.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Green, C. E.; Wagner, D. A.; Glogowski, J.; Skipper, P. L.; Wishnok, J. S.; Tannenbaum, S. R. Analysis of nitrate, nitrite and nitrate in biological fluids. Anal. Biochem. 1982, 243, 709–714. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J. Iron and free radical reactions: two aspects of antioxidant protection. Trends Biochem. Sci. 1986, 11, 372–375. [Google Scholar] [CrossRef]
- Benzie, I. F. F.; Strain, J. J. Ferric reducing/antioxidant power assay: direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid and concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar]
- Ellman, G. L.; Courtney, K. D.; Andrés, V.; Featherstone, R. M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Pintać, D.; Četojević-Simin, D.; Berežni, S.; Orčić, D.; Mimica-Dukić, N.; Lesjak, M. Investigation of the chemical composition and biological activity of edible grapevine (Vitis vinifera L.) leaf varieties. Food Chem. 2019, 286, 686–695. [Google Scholar] [CrossRef]
- Mosmann, T. R. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65 (1–2), 55–63.
- Hammer, Ø.; Harper, D. A. T.; Ryan, P. D. PAST: paleontological statistics software package for education and data analysis version 2. 09. 2001, 4, 1–9. [Google Scholar]
- Karaman, M.; Matavulj, M. Macroelements and heavy metals in some lignicolous and tericolous fungi. Matica Srpska Journal of Natural Science 2005, 108, 255–267. [Google Scholar] [CrossRef]
- Mallikarjuna, S. E.; Ranjini, A.; Haware, D. J.; Vijayalakshmi, M. R.; Shashirekha, M.; Rajarathnam, S. Mineral composition of four edible mushrooms. J. Chem. 2013, 2013, 1–5. [Google Scholar] [CrossRef]
- Tel-Çayan, G.; Öztürk, M.; Duru, M. E.; Yabanlı, M.; Türkoğlu, A. Content of minerals and trace elements determined by ICP-MS in eleven mushroom species from Anatolia, Turkey. Chiang Mai J. Sci. 2017, 44, 939–945. [Google Scholar]
- Florencio-Silva, R.; Sasso, G. R.; Sasso-Cerri, E.; Simões, M. J.; Cerri, P. S. Biology of bone tissue: structure, function, and factors that influence bone cells. BioMed Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef]
- Stilinović, N.; Škrbić, B.; Živančev, J.; Mrmoš, N.; Pavlović, N.; Vukmirović, S. The level of elements and antioxidant activity of commercial dietary supplement formulations based on edible mushrooms. Food Funct. 2014, 5, 3170–3178. [Google Scholar] [CrossRef]
- Stilinović, N.; Čapo, I.; Vukmirović, S.; Rašković, A.; Tomas, A.; Popović, M.; Sabo, A. Chemical composition, nutritional profile and in vivo antioxidant properties of the cultivated mushroom Coprinus comatus. R. Soc. Open Sci. 2020, 7, 200900. [Google Scholar] [CrossRef]
- Rašeta, M.; Rakić, M.; Čapelja, E.; Karaman, M. Chapter 2: Update on Research Data on the Nutrient Composition of Mushrooms and Their Potentials in Future Human Diets. In Food chemistry, function and analysis, Edible Fungi: Chemical Composition, Nutrition and Health Effects, edition no.1; Stojković, D., Barros, L., Eds.; Royal Society of Chemistry: Cambridge, United Kingdom, 2022; pp. 27–67. [Google Scholar]
- Gałgowska, M.; Pietrzak-Fiećko, R. Mineral composition of three popular wild mushrooms from Poland. Molecules 2020, 25, 3588. [Google Scholar] [CrossRef]
- Gnusarev, S.; Mitrofanova, N.; Churakov, B. P. The effect of mixed rot from the present tinder (Fomes fomentarius (L.:Fr.) Gill.) on the accumulation of heavy metals in the hanging birch (Betula pendula Roth.). In Materialy VIII Vserossijskoj konferencii s mezhdunarodnym uchastiem «Mediko-fiziologicheskie problemy jekologii cheloveka», Ulyanovsk, Russia, 01–04 December 2021.
- Dadáková, E.; Pelikánová, T.; Kalač, P. Content of biogenic amines and polyamines in some species of European wild-growing edible mushrooms. Eur. Food Res. Technol. 2009, 230, 163–171. [Google Scholar] [CrossRef]
- Reis, G. L.; Custódio, F. B.; Botelho, B. G.; Guidi, L. R.; Glória, M. B. Investigation of biologically active amines in some selected edible mushrooms. J. Food Compos. Anal. 2020, 86, 103375. [Google Scholar] [CrossRef]
- Kalač, P. Health effects and occurrence of dietary polyamines: a review for the period 2005-mid 2013. Food Chem. 2014, 161, 27–39. [Google Scholar] [CrossRef]
- Bal, C.; Akgul, H.; Sevindik, M.; Akata, I.; Yumrutas, O. Determination of the anti-oxidative activities of six mushrooms. Fresenius Environ. Bull. 2017, 26, 6246–6252. [Google Scholar]
- Kozarski, M.; Klaus, A.; Jakovljevic, D.; Todorović, N.; Vunduk, J.; Petrović, P. M.; Nikšić, M. P.; Vrvić, M. M.; Van Griensven, L. J. Antioxidants of edible mushrooms. Molecules 2015, 20, 19489–19525. [Google Scholar] [CrossRef] [PubMed]
- Karaman, M.; Stahl, M.; Vulić, J.; Vesić, M.; Čanadanović-Brunet, J. Wild-growing lignicolous mushroom species as sources of novel agents with antioxidative and antibacterial potentials. Int. J. Food Sci. Nutr. 2014, 65, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Tešanović, K.; Pejin, B.; Šibul, F.; Matavulj, M.; Rašeta, M.; Janjušević, L.; Karaman, M. A comparative overview of antioxidative properties and phenolic profiles of different fungal origins: fruiting bodies and submerged cultures of Coprinus comatus and Coprinellus truncorum. J. Food Sci. Technol. 2017, 54, 430–438. [Google Scholar] [CrossRef]
- Nowacka, N.; Nowak, R.; Drozd, M.; Olech, M.; Los, R.; Malm, A. Antibacterial, antiradical potential and phenolic compounds of thirty-one Polish mushrooms. PLoS ONE 2015, 10, e0140355. [Google Scholar] [CrossRef]
- Karadeniz, M.; Bakır, T. K.; Ünal, S. Investigation of the antioxidant and total phenolic substance of Fomes fomentarius and Ganoderma applanatum mushrooms showing therapeutic properties. Bilge International Journal of Science and Technology Research 2024, 8, 14–18. [Google Scholar] [CrossRef]
- Deveci, E.; Çayan, F.; Tel-Çayan, G.; Duru, M. E. Structural characterization and determination of biological activities for different polysaccharides extracted from tree mushroom species. J. Food Biochem. 2019, 43, e12965. [Google Scholar] [CrossRef]
- Saltarelli, R.; Ceccaroli, P.; Iotti, M.; Zambonelli, A.; Buffalini, M.; Casadei, L.; Vallorani, L.; Stocchi, V. Biochemical characterisation and antioxidant activity of mycelium of Ganoderma lucidum from Central Italy. Food Chem. 2009, 116, 143–151. [Google Scholar] [CrossRef]
- On-nom, N.; Suttisansanee, U.; Chathiran, W.; Charoenkiatkul, S.; Thiyajai, P.; Srichamnong, W. Nutritional security: carbohydrate profile and folk remedies of rare edible mushrooms to diversify food and diet: Thailand case study. Sustainability 2023, 15, 14034. [Google Scholar] [CrossRef]
- Ilyashenka, S. Biological activity of Fomes fomentarius. J. Pharm. Pharmacol. 2022, 10, 294–298. [Google Scholar] [CrossRef]
- Darkal, A. K.; Zuraik, M. M.; Ney, Y.; Nasim, M. J.; Jacob, C. Unleashing the biological potential of Fomes fomentarius via dry and wet milling. Antioxidants 2021, 10, 303. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H. A.; Ho, T. P.; Mangelings, D.; Van Eeckhaut, A.; Vander Heyden, Y.; Tran, H. T. Antioxidant, neuroprotective, and neuroblastoma cells (SH-SY5Y) differentiation effects of melanins and arginine-modified melanins from Daedaleopsis tricolor and Fomes fomentarius. BMC Biotechnol. 2024, 24, 89. [Google Scholar] [CrossRef] [PubMed]
- Pero, R. W.; Lund, H.; Leanderson, T. Antioxidant metabolism induced by quinic acid. increased urinary excretion of tryptophan and nicotinamide. Phytother. Res. 2009, 23, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.H.; Kang, H.K.; Hosseindoust, A.; Mun, J.Y.; Moturi, J.; Tajudeen, H.; Lee, H.; Cheong, E.J.; Kim, J.S. Effects of scopoletin supplementation and stocking density on growth performance, antioxidant activity, and meat quality of Korean native broiler chickens. Foods 2021, 10, 1505. [Google Scholar] [CrossRef]
- Chen, N.; Jiang, T.; Xu, J.; Xi, W.; Shang, E.; Xiao, P.; Duan, J. The relationship between polysaccharide structure and its antioxidant activity needs to be systematically elucidated. Int. J. Biol. Macromol. 2024, 270. [Google Scholar] [CrossRef]
- Murray Stewart, T.; Dunston, T. T.; Woster, P. M.; Casero, R. A. Jr. Polyamine catabolism and oxidative damage. J. Biol. Chem. 2018, 293, 18736–18745. [Google Scholar] [CrossRef]
- Casero, R. A. Jr.; Murray Stewart, T.; Pegg, A. E. Polyamine metabolism and cancer: treatments, challenges and opportunities. Nat. Rev. Cancer. 2018, 18, 681–695. [Google Scholar] [CrossRef]
- Moss, D. E. Improving anti-neurodegenerative benefits of acetylcholinesterase inhibitors in Alzheimer's disease: are irreversible inhibitors the future? Int. J. Mol. Sci. 2020, 21, 3438. [Google Scholar] [CrossRef]
- Murray, A. P.; Faraoni, M. B.; Castro, M. J.; Alza, N. P.; Cavallaro, V. Natural AChE inhibitors from plants and their contribution to Alzheimer's disease therapy. Curr. Neuropharmacol. 2013, 11, 388–413. [Google Scholar] [CrossRef]
- Borai, I. H.; Ezz, M. K.; Rizk, M. Z.; Aly, H. F.; El-Sherbiny, M.; Matloub, A. A.; Fouad, G. I. Therapeutic impact of grape leaves polyphenols on certain biochemical and neurological markers in AlCl3-induced Alzheimer’s disease. Biomed. Pharmacother. 2017, 93, 837–851. [Google Scholar] [CrossRef]
- Ahmed, S.; Khan, S.; Zargaham, M. K.; Khan, A.; Khan, S.; Hussain, A.; Uddin, J.; Khan, A.; Al-Harrasi, A. Potential therapeutic natural products against Alzheimer's disease with reference of acetylcholinesterase. Biomed. Pharmacother. 2021, 139, 111609. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, I.; Sankhe, R.; Mudgal, J.; Arora, D.; Nampoothiri, M. Spermidine, an autophagy inducer, as a therapeutic strategy in neurological disorders. Neuropeptides 2020, 83, 102083. [Google Scholar] [CrossRef] [PubMed]
- Arthur, R.; Jamwal, S.; Kumar, P. A review on polyamines as promising next-generation neuroprotective and anti-aging therapy. Eur. J. Pharmacol. 2024, 978, 176804. [Google Scholar] [CrossRef] [PubMed]
- Cör, D.; Botić, T.; Gregori, A.; Pohleven, F.; Knez, Ž. (2017). The effects of different solvents on bioactive metabolites and “in vitro” antioxidant and anti-acetylcholinesterase activity of Ganoderma lucidum fruiting body and primordia extracts. Maced. J. Chem. Chem. Eng. 2017, 36, 129–141. [Google Scholar] [CrossRef]
- Jabir, N. R.; Khan, F. R.; Tabrez, S. Cholinesterase targeting by polyphenols: a therapeutic approach for the treatment of Alzheimer's disease. CNS Neurosci. Ther. 2018, 24, 753–762. [Google Scholar] [CrossRef]
- Budryn, G.; Majak, I.; Grzelczyk, J.; Szwajgier, D.; Rodríguez-Martínez, A.; Pérez-Sánchez, H. Hydroxybenzoic acids as acetylcholinesterase inhibitors: calorimetric and docking simulation studies. Nutrients 2022, 14, 2476. [Google Scholar] [CrossRef]
- Barbosa, J. R.; de Carvalho Júnior, R. N. Polysaccharides obtained from natural edible sources and their role in modulating the immune system: biologically active potential that can be exploited against COVID-19. Trends Food Sci. Technol. 2021, 108, 223–235. [Google Scholar] [CrossRef]
- Sułkowska-Ziaja, K.; Balik, M.; Szczepkowski, A.; Trepa, M.; Zengin, G.; Kała, K.; Muszyńska, B. A review of chemical composition and bioactivity studies of the most promising species of Ganoderma spp. Diversity 2023, 15, 882. [Google Scholar] [CrossRef]
- Guerra, G. P.; Rubin, M. A.; Mello, C. F. Modulation of learning and memory by natural polyamines. Pharmacol. Res. 2016, 112, 99–118. [Google Scholar] [CrossRef]
- Toro-Funes, N.; Bosch-Fusté, J.; Veciana-Nogués, M. T.; Izquierdo-Pulido, M.; Vidal-Carou, M. D. In vitro antioxidant activity of dietary polyamines. Food Res. Int. 2013, 51, 141–147. [Google Scholar] [CrossRef]
- Makletsova, M. G.; Rikhireva, G. T.; Kirichenko, E. Y.; Trinitatsky, I. Y.; Vakulenko, M. Y.; Ermakov, A. M. The role of polyamines in the mechanisms of cognitive impairment. J. Neurochem. 2022, 16, 283–294. [Google Scholar] [CrossRef]
- Zhang, L.; Gu, C.; Liu, J. Nature spermidine and spermine alkaloids: occurrence and pharmacological effects. Arab. J. Chem. 2022, 15, 104367. [Google Scholar] [CrossRef]
- Shnyreva, A. V.; Shnyreva, A. A.; Espinoza, C.; Padrón, J. M.; Trigos, Á. Antiproliferative activity and cytotoxicity of some medicinal wood-destroying mushrooms from Russia. Int. J. Med. Mushrooms 2018, 20, 1–11. [Google Scholar] [CrossRef]
- Rosendahl, A. H.; Perks, C. M.; Zeng, L.; Markkula, A.; Simonsson, M.; Rose, C.; Ingvar, C.; Holly, J. M. P.; Jernstrom, H. Caffeine and caffeic acid inhibit growth and modify estrogen receptor and insulin-like growth factor i receptor levels in human breast cancer. Clin. Cancer Res. 2015, 21, 1877–1887. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Geographical distribution of sampled Fomes fomentarius specimens across the Balkans. The map shows sampling localities in Croatia (FC), Bosnia and Herzegovina (FB), and Serbia (FS). The designated markers indicate specific collection sites: National Park “Štrbački Buk” (FC), Vrelo Bosne (FB), and National Park “Fruška Gora” (FS) (Map image sourced from Google Maps).
Figure 1.
Geographical distribution of sampled Fomes fomentarius specimens across the Balkans. The map shows sampling localities in Croatia (FC), Bosnia and Herzegovina (FB), and Serbia (FS). The designated markers indicate specific collection sites: National Park “Štrbački Buk” (FC), Vrelo Bosne (FB), and National Park “Fruška Gora” (FS) (Map image sourced from Google Maps).
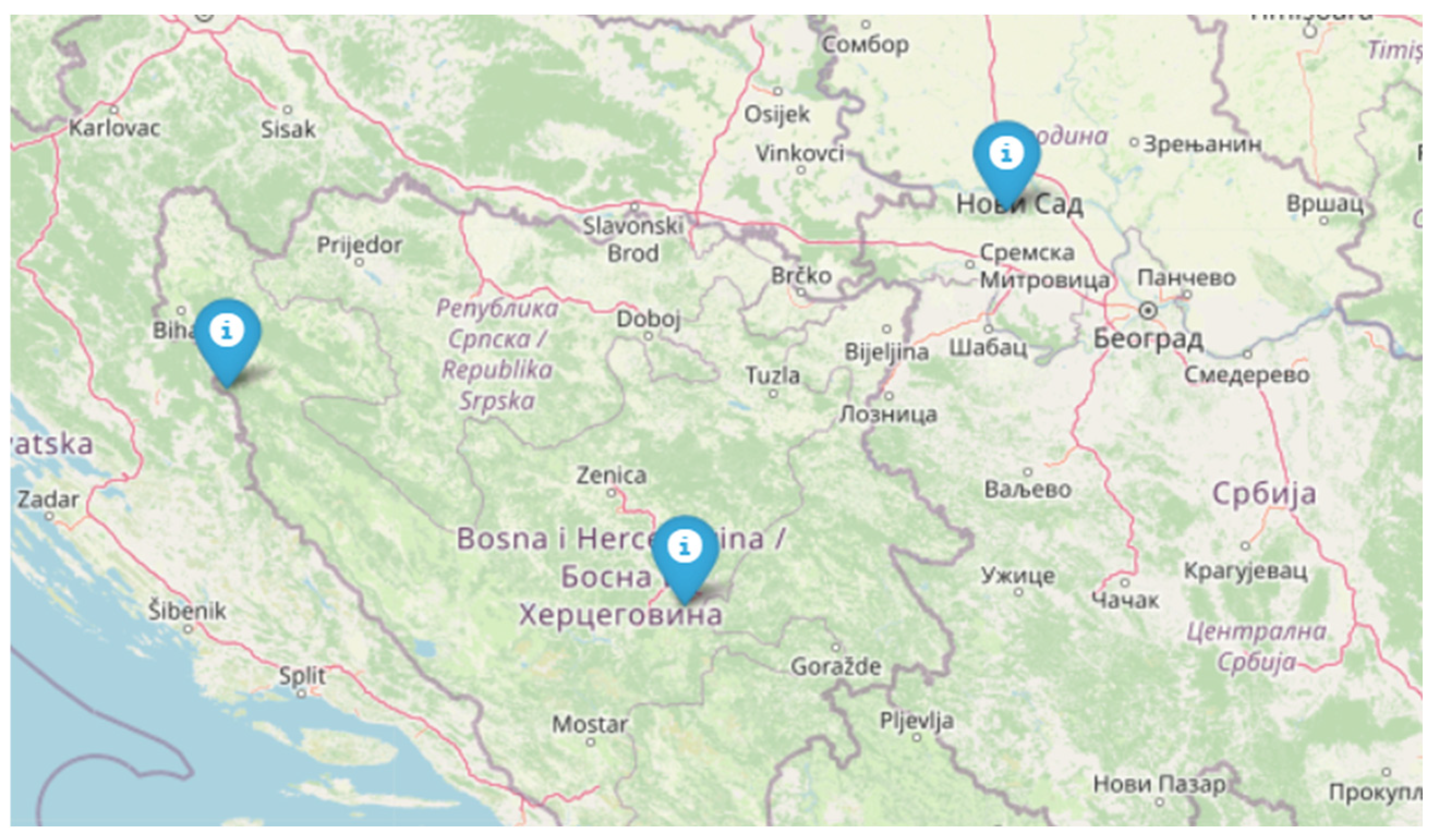
Figure 2.
Mineral composition in dried samples of F. fomentarius from three different localities in the Balkan region: a) Macroelement content (mg/g d.w.) and b) Microelement content (mg/kg d.w.). The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; d.w.–dry weight. Different small letters indicate significant differences among different species; Tukey`s significant difference (HSD) post hoc test (p < 0.05). Data represent the mean ± standard deviation (SD).
Figure 2.
Mineral composition in dried samples of F. fomentarius from three different localities in the Balkan region: a) Macroelement content (mg/g d.w.) and b) Microelement content (mg/kg d.w.). The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; d.w.–dry weight. Different small letters indicate significant differences among different species; Tukey`s significant difference (HSD) post hoc test (p < 0.05). Data represent the mean ± standard deviation (SD).
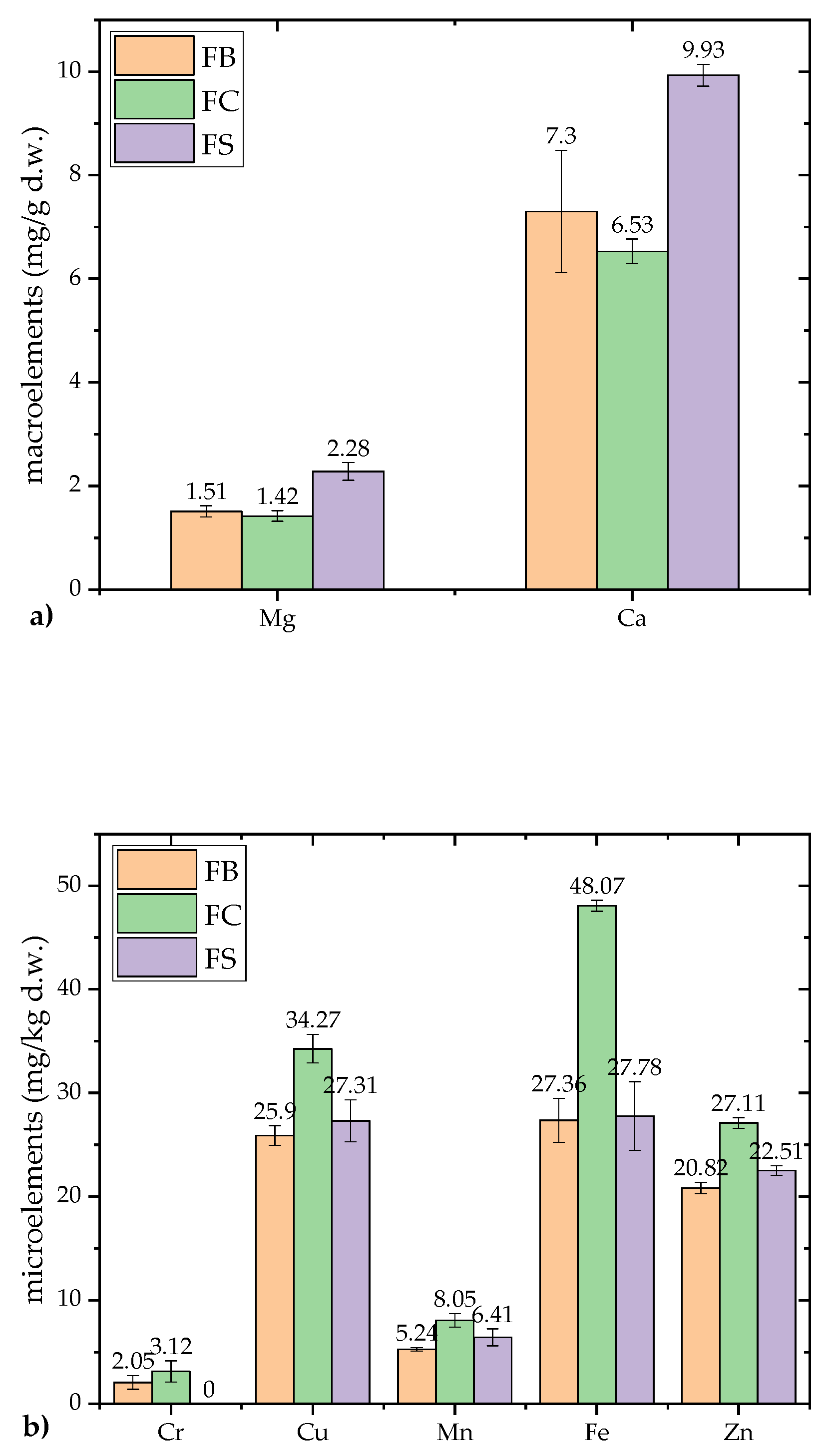
Figure 3.
Polyamine content in dried samples of F. fomentarius from three different localities in the Balkan region, expressed as putrescine, spermidine and spermine levels. The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; d.w.–dry weight. Different small letters indicate significant differences among different species; Tukey`s significant difference (HSD) post hoc test (p < 0.05). Data represent the mean ± standard deviation (SD).
Figure 3.
Polyamine content in dried samples of F. fomentarius from three different localities in the Balkan region, expressed as putrescine, spermidine and spermine levels. The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; d.w.–dry weight. Different small letters indicate significant differences among different species; Tukey`s significant difference (HSD) post hoc test (p < 0.05). Data represent the mean ± standard deviation (SD).
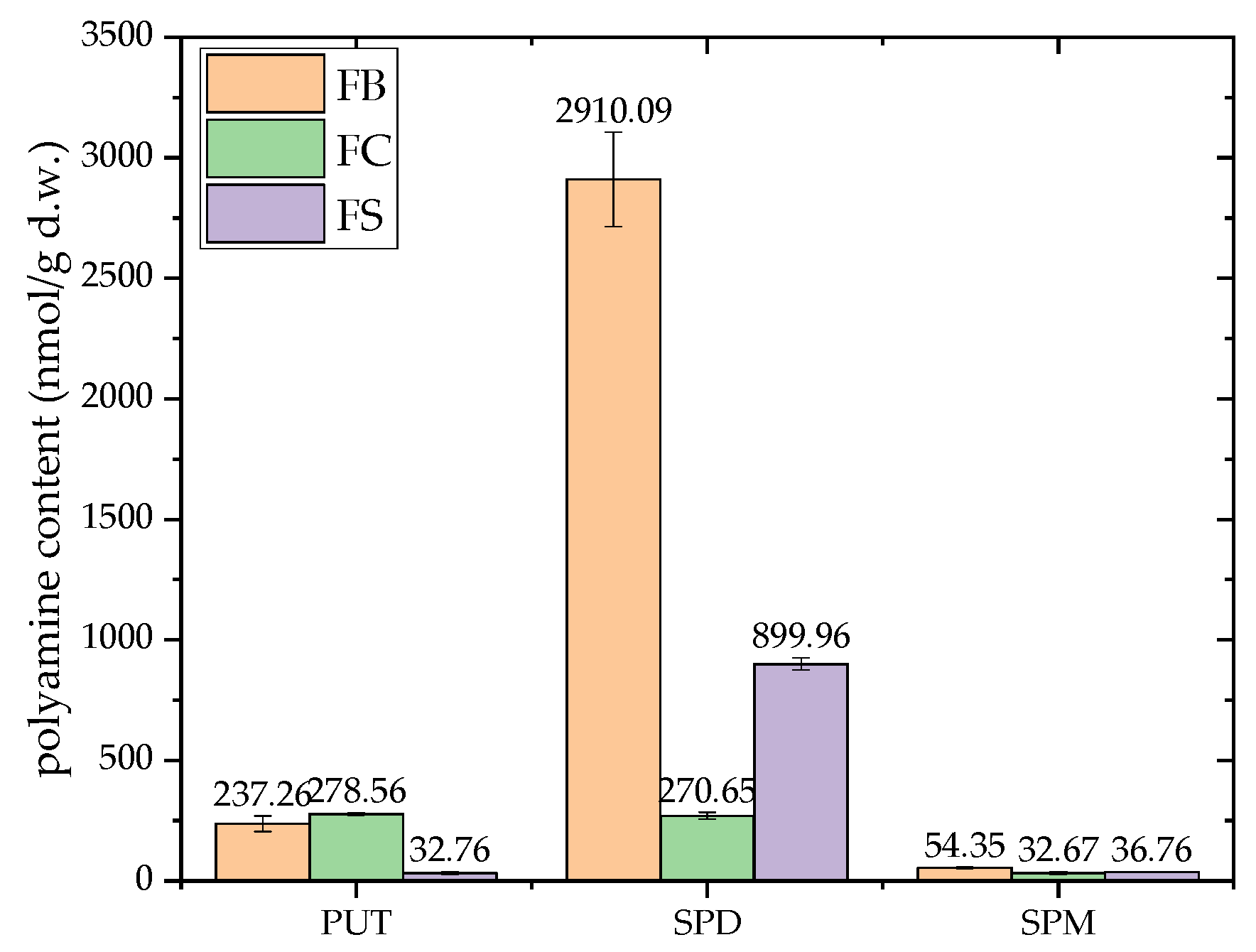
Figure 4.
PCA analysis of Fomes fomentarius extracts and identified polyphenolics using LC-MS/MS analysis. Distribution of variables in: (above) the score plot and (below) the loading plot of the first two principal components. The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; 70% EtOH–hydroethanolic extract prepared with 70% EtOH; H2O–hot water extract; 80% MeOH–hydromethanolic extract prepared with 80% MeOH; CHCl3–chloroform extract.
Figure 4.
PCA analysis of Fomes fomentarius extracts and identified polyphenolics using LC-MS/MS analysis. Distribution of variables in: (above) the score plot and (below) the loading plot of the first two principal components. The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; 70% EtOH–hydroethanolic extract prepared with 70% EtOH; H2O–hot water extract; 80% MeOH–hydromethanolic extract prepared with 80% MeOH; CHCl3–chloroform extract.
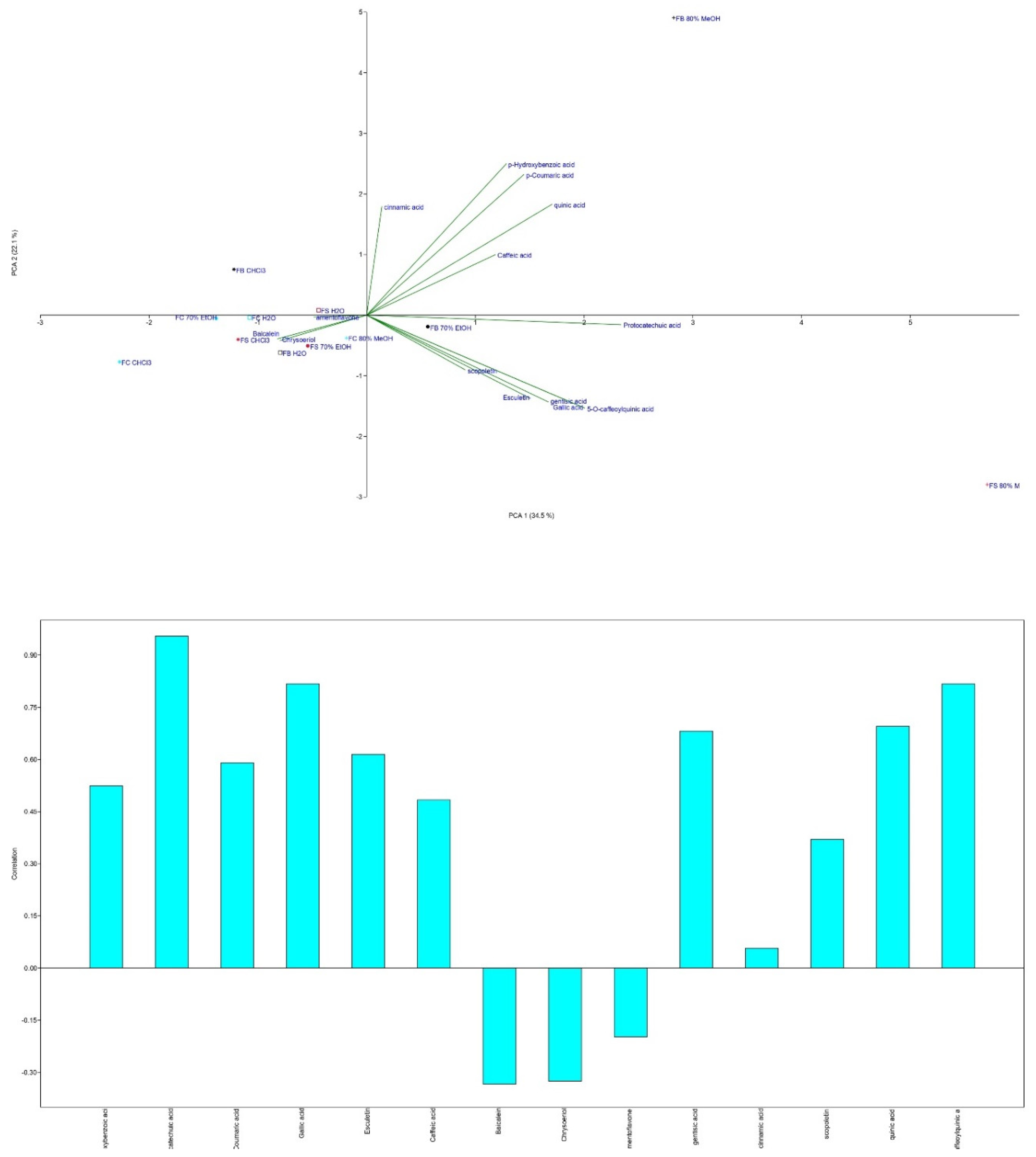
Figure 5.
Total phenolic content (TPC) and total carbohydrate content (TCC) of F. fomentarius extracts from three different localities in the Balkan region: a) Total phenolic content (TPC); b) Total carbohydrate content (TCC). The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; EtOH–hydroethanolic extract prepared with 70% EtOH; MeOH–hydromethanolic extract prepared with 80% MeOH; H2O–hot water extract; CHCl3–chloroform extract; GAE–gallic acid equivalents; d.w.–dry weight; GluE–Glucose equivalents; Different small letters indicate significant differences among different species; Tukey`s significant difference (HSD) post hoc test (p < 0.05). Data represent the mean ± standard deviation (SD).
Figure 5.
Total phenolic content (TPC) and total carbohydrate content (TCC) of F. fomentarius extracts from three different localities in the Balkan region: a) Total phenolic content (TPC); b) Total carbohydrate content (TCC). The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; EtOH–hydroethanolic extract prepared with 70% EtOH; MeOH–hydromethanolic extract prepared with 80% MeOH; H2O–hot water extract; CHCl3–chloroform extract; GAE–gallic acid equivalents; d.w.–dry weight; GluE–Glucose equivalents; Different small letters indicate significant differences among different species; Tukey`s significant difference (HSD) post hoc test (p < 0.05). Data represent the mean ± standard deviation (SD).
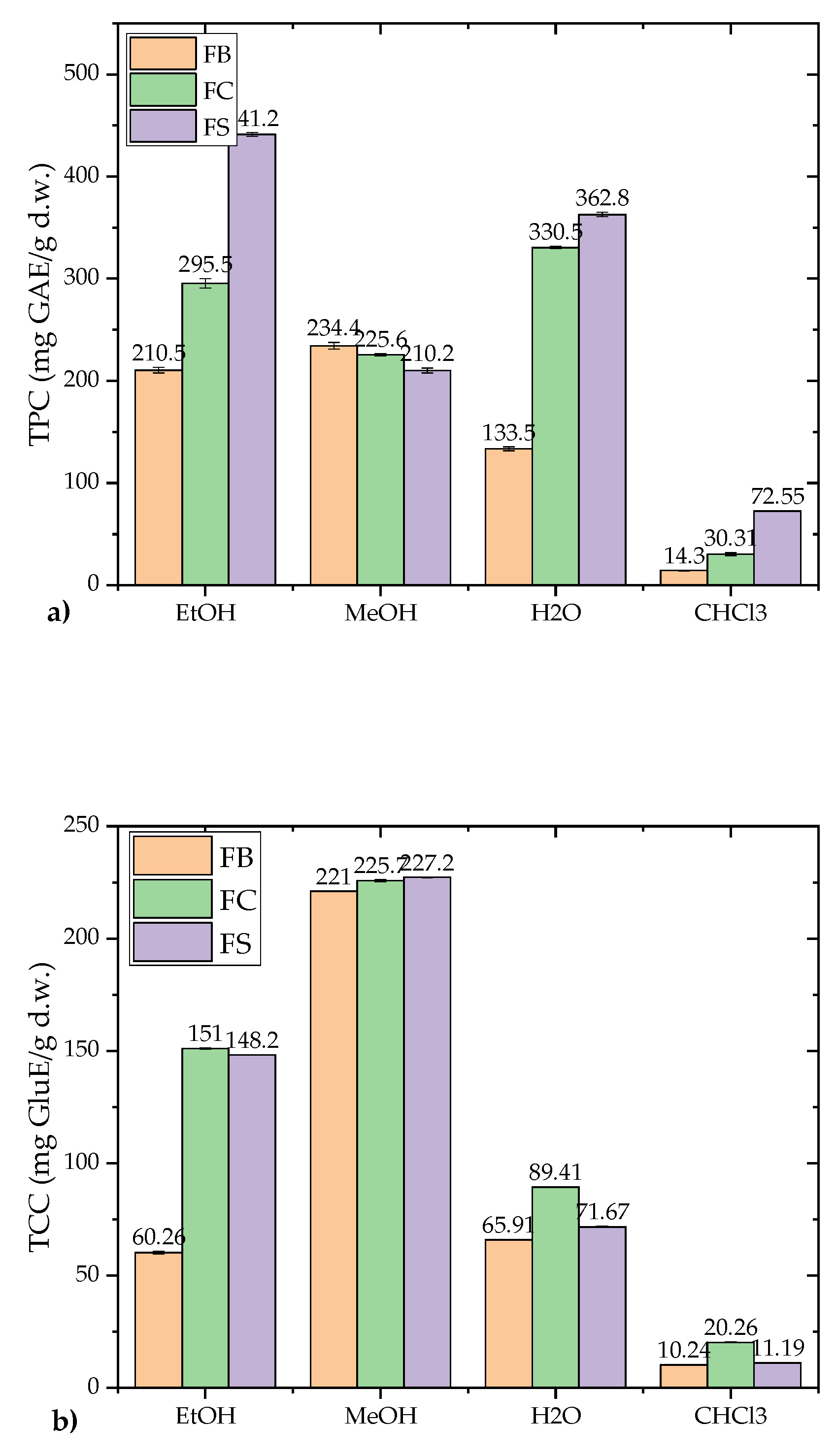
Figure 6.
Antioxidant and anti-acetycholinesterase activity of Fomes fomentarius extracts from three different localities in the Balkan region: a) DPPH scavenging activity; b) ABTS scavenging activity; c) NO scavenging activity; d) Reduction potential (FRAP assay); e) Fe²⁺/ascorbate-induced lipid peroxidation (LP, tested only in FB strain extracts); f) Anti-AChE activity (tested only in FB strain extracts, MeOH extracts of FC and FS). The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; EtOH–hydroethanolic extract prepared with 70% EtOH; MeOH–hydromethanolic extract prepared with 80% MeOH; H2O–hot water extract; CHCl3–chloroform extract; PG–Propyl gallate; d.w.–dry weight; DPPH–radical scavenger capacity against 2,2-diphenyl-1-picrylhydrazyl radical, DPPH•; ABTS–radical scavenger capacity against 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid, ABTS•+; NO–radical scavenging capacity against NO•; FRAP– ferric reducing antioxidant power; AAE–ascorbic acid equivalents; TE–Trolox equivalents; anti-AChE–anti-acetylcholinesterase. Different small letters indicate significant differences among different species; Tukey`s significant difference (HSD) post hoc test (p < 0.05). Data represent the mean ± standard deviation (SD).
Figure 6.
Antioxidant and anti-acetycholinesterase activity of Fomes fomentarius extracts from three different localities in the Balkan region: a) DPPH scavenging activity; b) ABTS scavenging activity; c) NO scavenging activity; d) Reduction potential (FRAP assay); e) Fe²⁺/ascorbate-induced lipid peroxidation (LP, tested only in FB strain extracts); f) Anti-AChE activity (tested only in FB strain extracts, MeOH extracts of FC and FS). The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; EtOH–hydroethanolic extract prepared with 70% EtOH; MeOH–hydromethanolic extract prepared with 80% MeOH; H2O–hot water extract; CHCl3–chloroform extract; PG–Propyl gallate; d.w.–dry weight; DPPH–radical scavenger capacity against 2,2-diphenyl-1-picrylhydrazyl radical, DPPH•; ABTS–radical scavenger capacity against 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid, ABTS•+; NO–radical scavenging capacity against NO•; FRAP– ferric reducing antioxidant power; AAE–ascorbic acid equivalents; TE–Trolox equivalents; anti-AChE–anti-acetylcholinesterase. Different small letters indicate significant differences among different species; Tukey`s significant difference (HSD) post hoc test (p < 0.05). Data represent the mean ± standard deviation (SD).
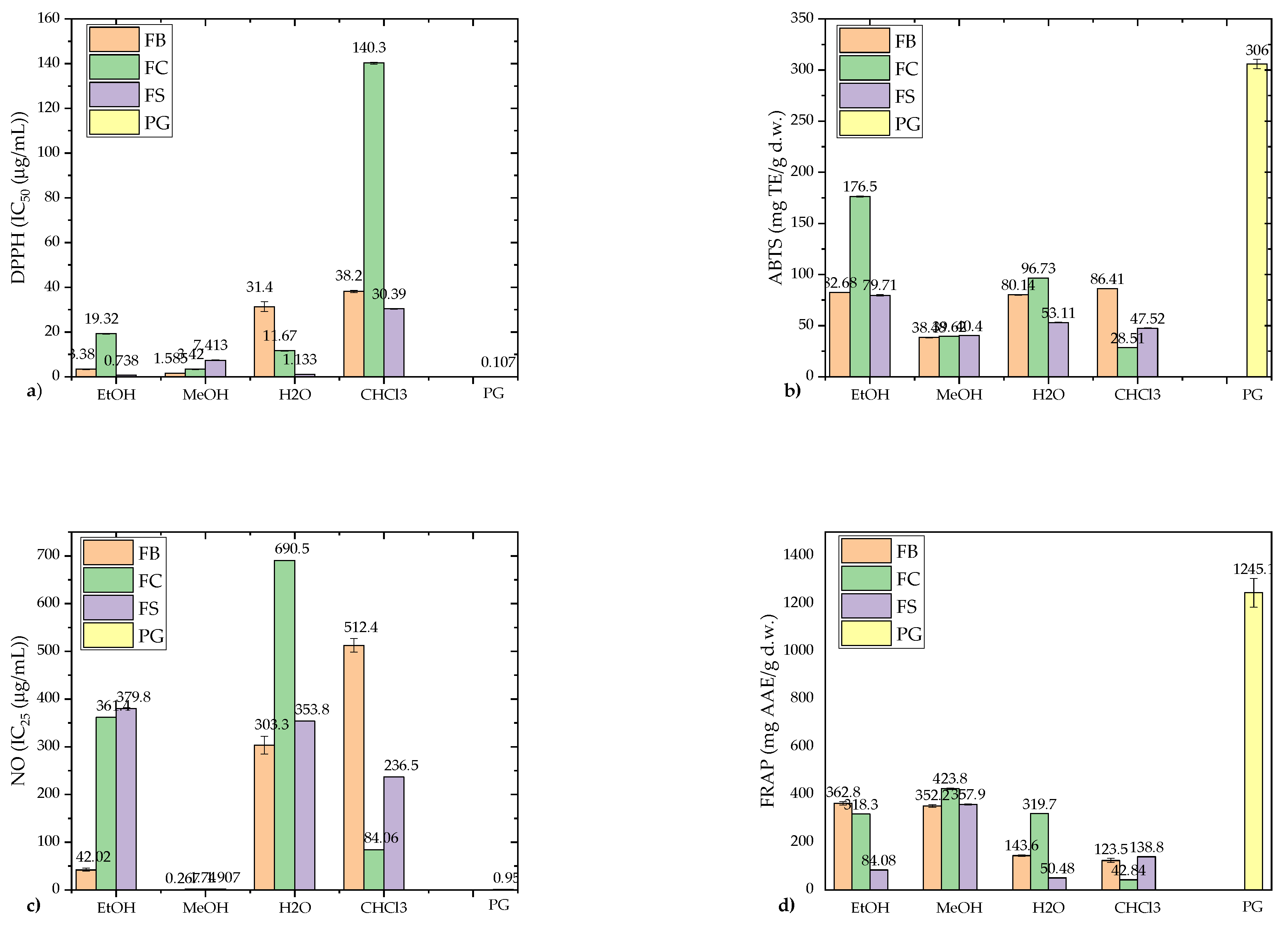
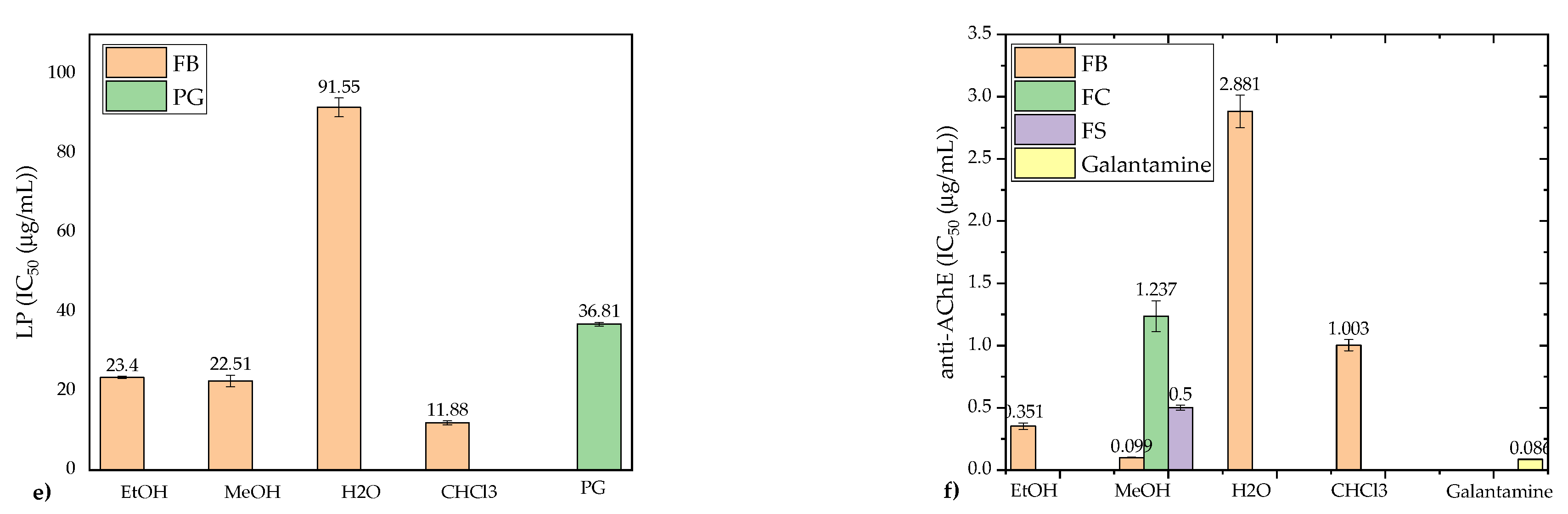
Figure 7.
PCA analysis of analyzed Fomes fomentarius extracts and conducted antioxidant assays. Distribution of variables in: (above) the score plot and (below) the loading plot of the first two principal components. The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; 70% EtOH–hydroethanolic extract prepared with 70% EtOH; H2O–hot water extract; 80% MeOH–hydromethanolic extract prepared with 80% MeOH; CHCl3–chloroform extract; DPPH–radical scavenger capacity against 2,2-diphenyl-1-picrylhydrazyl radical, DPPH•; ABTS–radical scavenger capacity against 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid, ABTS•+; NO–radical scavenging capacity against NO•; LP–lipid peroxidation assay; FRAP– ferric reducing antioxidant power.
Figure 7.
PCA analysis of analyzed Fomes fomentarius extracts and conducted antioxidant assays. Distribution of variables in: (above) the score plot and (below) the loading plot of the first two principal components. The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; 70% EtOH–hydroethanolic extract prepared with 70% EtOH; H2O–hot water extract; 80% MeOH–hydromethanolic extract prepared with 80% MeOH; CHCl3–chloroform extract; DPPH–radical scavenger capacity against 2,2-diphenyl-1-picrylhydrazyl radical, DPPH•; ABTS–radical scavenger capacity against 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid, ABTS•+; NO–radical scavenging capacity against NO•; LP–lipid peroxidation assay; FRAP– ferric reducing antioxidant power.
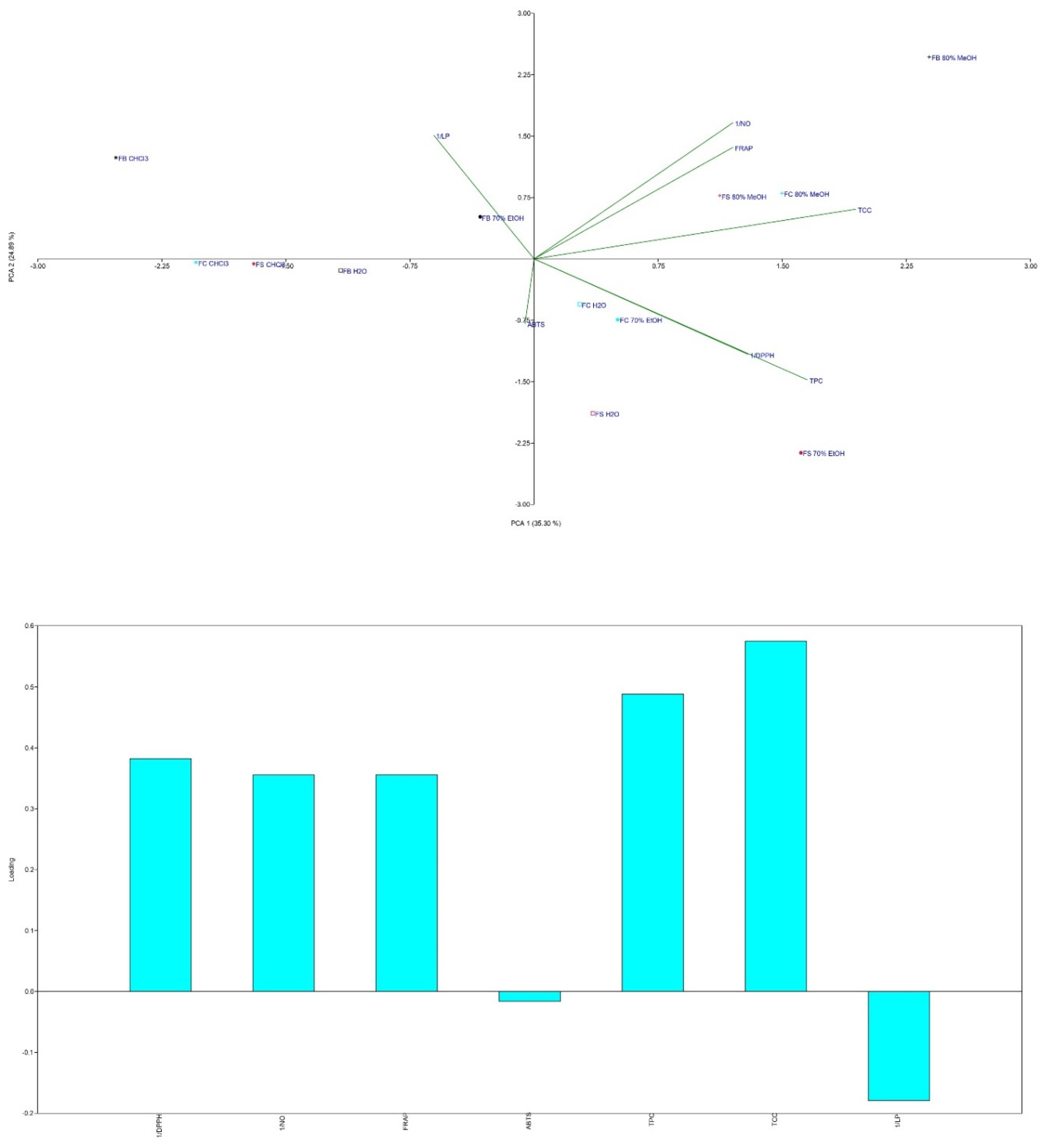
Figure 8.
PCA analysis of analyzed Fomes fomentarius extracts, mycochemical profile including TPC, TCC, and PAs and conducted antioxidant assays and measured anti-AChE properties. Distribution of variables in: (above) the score plot and (below) the loading plot of the first two principal components. The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; 70% EtOH–hydroethanolic extract prepared with 70% EtOH; H2O–hot water extract; 80% MeOH–hydromethanolic extract prepared with 80% MeOH; CHCl3–chloroform extract; anti-AChE–anti-acetylcholinesterase; DPPH–radical scavenger capacity against 2,2-diphenyl-1-picrylhydrazyl radical, DPPH•; ABTS–radical scavenger capacity against 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid, ABTS•+; NO–radical scavenging capacity against NO•; LP–lipid peroxidantion assay; FRAP– ferric reducing antioxidant power; TPC–total phenolic content; TCC–total carbohydrate content; PAs–polyamines; PUT–putrescine; SPD–spermidine; SPM–spermine.
Figure 8.
PCA analysis of analyzed Fomes fomentarius extracts, mycochemical profile including TPC, TCC, and PAs and conducted antioxidant assays and measured anti-AChE properties. Distribution of variables in: (above) the score plot and (below) the loading plot of the first two principal components. The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; 70% EtOH–hydroethanolic extract prepared with 70% EtOH; H2O–hot water extract; 80% MeOH–hydromethanolic extract prepared with 80% MeOH; CHCl3–chloroform extract; anti-AChE–anti-acetylcholinesterase; DPPH–radical scavenger capacity against 2,2-diphenyl-1-picrylhydrazyl radical, DPPH•; ABTS–radical scavenger capacity against 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid, ABTS•+; NO–radical scavenging capacity against NO•; LP–lipid peroxidantion assay; FRAP– ferric reducing antioxidant power; TPC–total phenolic content; TCC–total carbohydrate content; PAs–polyamines; PUT–putrescine; SPD–spermidine; SPM–spermine.
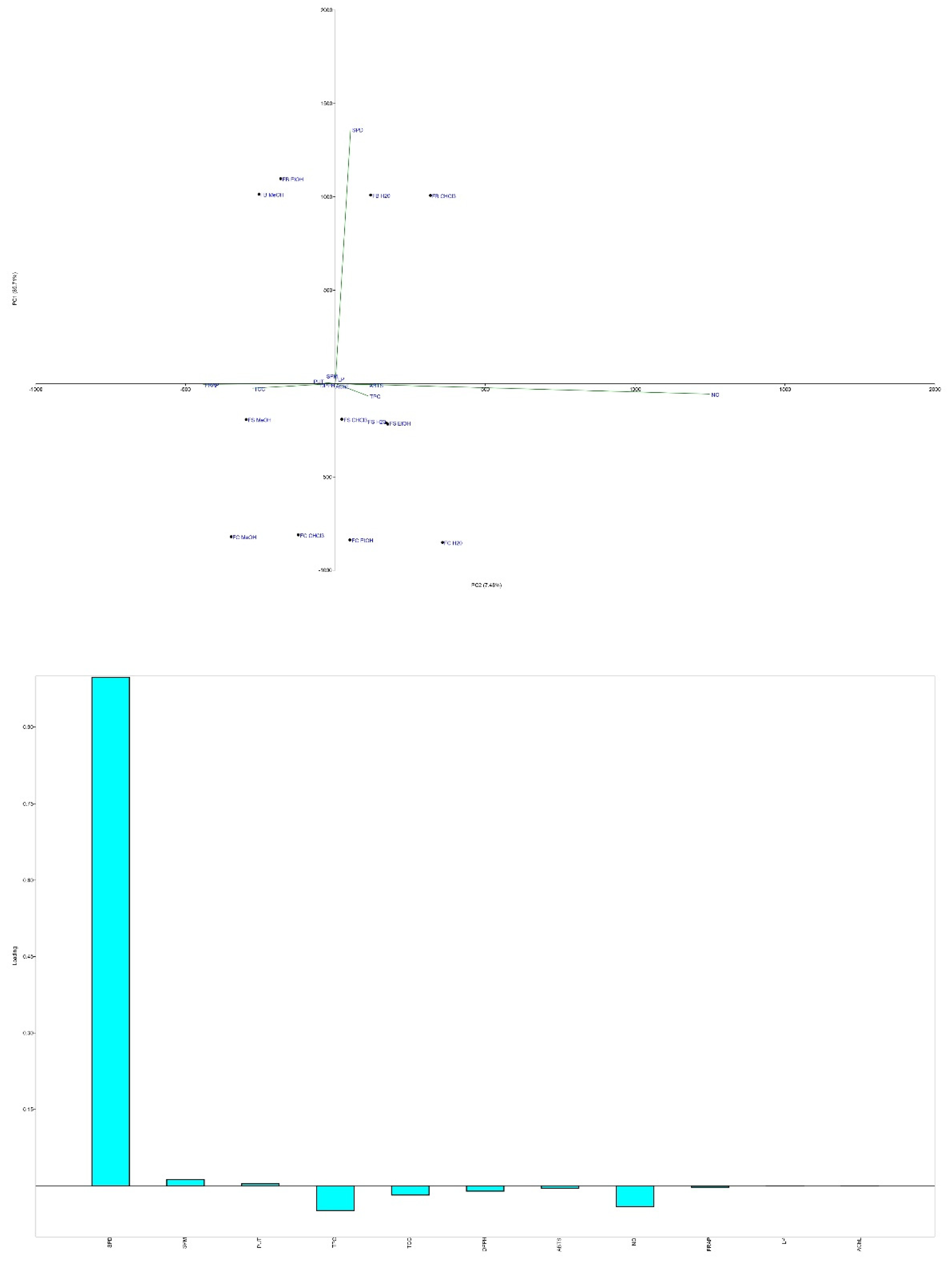

Table 1.
Concentrations of selected phenolic compounds determined using the LC-MS/MS method in the analyzed extracts of three different Fomes fomentarius strains.
Table 1.
Concentrations of selected phenolic compounds determined using the LC-MS/MS method in the analyzed extracts of three different Fomes fomentarius strains.
| Class | Compound | Amount of selected compound (μg/g d.w. ± SD) | |||||||||||
| F. fomentariusand extract type | |||||||||||||
| FB | FC | FS | |||||||||||
| CHCl3 | H2O | EtOH | MeOH | CHCl3 | H2O | EtOH | MeOH | CHCl3 | H2O | EtOH | MeOH | ||
| Flavones | Baicalein | <12.2 | <12.2 | <12.2 | <12.2 | 213.9 ± 42.78a | <12.2 | <12.2 | <12.2 | <12.2 | 147.5 ± 29.50b | 44.63 ± 8.926c | <12.2 |
| Chrysoeriol | <3.05 | <3.05 | <3.05 | <3.05 | 3.410 ± 0.102a | <3.05 | <3.05 | <3.05 | <3.05 | <3.05 | <3.05 | <3.05 | |
| Biflavonoid | Amentoflavone | <3.05 | <3.05 | <3.05 | <3.05 | <3.05 | <3.05 | 10.97 ± 0.329a | <3.05 | <3.05 | <3.05 | <3.05 | <3.05 |
| Hydroxybenzoic acids | p-Hydroxybenzoic acid | <6.1 | <6.1 | 32.79 ± 1.967c | 420.0 ± 25.20a | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | 56.03 ± 3.362b |
| Protocatechuic acid | <6.1 | <6.1 | 10.90 ± 0.872c | 13.28 ± 1.062b | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | 25.48 ± 2.038a | |
| Gentisic acid | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | 15.91 ± 1.273b | <12.2 | <12.2 | <12.2 | 22.46 ± 1.797a | |
| Gallic acid | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | 15.30 ± 1.377a | |
| Cinnamic acid | 179.9 ± 35.98a | <97.5 | <97.5 | 106.1 ± 21.22b | <97.5 | <97.5 | <97.5 | <97.5 | <97.5 | <97.5 | <97.5 | <97.5 | |
| Hydroxycinnamic acids | p-Coumaric acid | <12.2 | <12.2 | <12.2 | 83.12 ± 7.481a | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | <12.2 | 19.23 ± 1.731b |
| Caffeic acid | <6.1 | <6.1 | 22.18 ± 1.553e | 83.36 ± 5.835b | <6.1 | <6.1 | <6.1 | 6.95 ± 0.438f | <6.1 | 105.95 ± 7.416a | 63.32 ± 4.432c | 49.23 ± 3.446d | |
| Coumarins | Esculetin | <6.1 | 51.36 ± 3.082b | 73.66 ± 4.420a | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | 84.03 ± 5.042a |
| Scopoletin | <6.1 |
<6.1 |
9.770 ± 0.782g | 88.02 ± 7.042e | <6.1 | <6.1 | <6.1 | 23.63 ± 1.890f | 209.1 ± 16.73d | 511.7 ± 40.94c | 853.1 ± 68.25a | 580.8 ± 46.46b | |
| Cyclohexanecarboxylic acid | Quinic acid | <48.85 |
58.99 ± 5.589g | 414.2 ± 41.42c | 996.8 ± 99.68a | <48.85 | 207.5 ± 20.75f | 211.7 ± 21.17f | 379.3 ± 37.93d | <48.85 | 247.8 ± 24.78e | <48.85 | 454.2 ± 45.42b |
| Chlorogenic acid | 5-O-caffeoylquinic acid | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | <6.1 | 6.740 ± 0.337a |
| Total | 179.88 | 110.35 | 563.48 | 1790.69 | 217.28 | 207.49 | 222.70 | 425.77 | 209.05 | 1012.91 | 961.02 | 1313.49 | |
Significant differences were determined by one-way ANOVA using the Tukey's HSD test. Data represent the mean ± standard deviation (SD), and differences were considered statistically significant at p < 0.05. Bold number: amount of quantified phenolic compounds in examined extracts. Values marked with the symbol “<” indicate concentrations below the LoQ (limit of quantification) but above the LoD (limit of detection), where the compound was detected but not quantifiable. The following abbreviations are used for the analyzed parameters: FB—strain sampled from Bosnia and Herzegovina; FC—strain sampled from Croatia; FS—strain sampled from Serbia; EtOH—hydroethanolic extract prepared with 70% EtOH; H2O—hot water extract; MeOH— hydromethanolic extract prepared with 80% MeOH CHCl3—chloroform extract; nd—not detected (no peak observed).
Table 2.
The antiproliferative activity of the selected Fomes fomentarius extracts.
| Cell line | Extract concentration (μg/mL) | Growth inhibition (% ± SEM) | |||||
| FB | FC | FS | |||||
| H2O | EtOH | MeOH | CHCl3 | MeOH | MeOH | ||
| MDA-MB-231 | 50 | 12.77±3.30d | 29.15±1.58c | 74.17±0.34a | < 10* | 12.83±0.59d | 46.55±1.79b |
| 100 | 13.05±2.57d | 89.06±0.87b | 94.98±0.78a | < 10* | 51.39±0.77c | 91.39±0.83b | |
| MCF-7 | 50 | 29.17±3.44e | 65.84±2.35c | 85.23±0.50a | 38.14±2.50d | 60.99±1.30c | 75.88±0.78b |
| 100 | 70.97±3.10e | 91.34±0.86c | 95.96±0.36a | 49.22±1.52f | 80.84±0.72d | 93.42±0.57b | |
| T47D | 50 | 17.45±2.08e | 55.38±0.72b | 51.51±1.50cd | 49.19±1.33d | 53.06±0.72bc | 68.55±0.94a |
| 100 | 20.72±1.80e | 78.62±0.83b | 82.80±1.58a | 68.40±1.19c | 63.59±1.12d | 80.79±0.90ab | |
| A2780 | 50 | < 10 | 91.12±0.43c | 100.30±0.36a | 25.07±3.29e | 69.20±1.66d | 96.35±0.72b |
| 100 | 36.95±3.79c | 100.50±0.33a | 99.94±0.34a | 52.43±0.99b | 100.20±0.31a | 99.21±0.47a | |
| SiHa | 50 | 16.44±2.02e | 66.97±0.33b | 93.13±0.92a | 44.56±1.62d | 51.43±2.86c | 68.77±0.31b |
| 100 | 20.09±1.58d | 95.42±0.76a | 95.75±0.48a | 43.25±1.82c | 73.51±1.26b | 93.13±1.02a | |
| HeLa | 50 | 15.46±2.47e | 50.25±2.04c | 60.86±3.10b | 36.87±0.54d | 62.75±2.12b | 68.69±1.35a |
| 100 | 27.81±1.07e | 92.80±3.38a | 96.31±0.50a | 50.79±1.50d | 70.31±2.96c | 82.82±2.48b | |
*: < 10: extracts exhibiting growth inhibition lower than 10% are considered ineffective and their data are not given numerically for clarity. Significant differences were determined by one-way ANOVA using the Tukey's HSD test. Data represent the mean ± SEM of two independent measurements, and differences were considered statistically significant at p < 0.05. Means within each row with different letters (a–f) are statistically significant. The following abbreviations are used for the examined parameters: FB–strain sampled from Bosnia and Herzegovina; FC–strain sampled from Croatia; FS–strain sampled from Serbia; d.w.–dry weight; CHCl3–chloroform extract; H2O–hot water extract; EtOH–hydroethanolic extract prepared with 70% EtOH; MeOH– hydromethanolic extract prepared with 80% MeOH; SEM– standard error of the mean; MDA-MB-231–Human breast cancer cell line; MCF7–Human breast cancer cell line; T47D–Human breast cancer cell line; A2780–Ovarian cancer cell line; SiHa–cervical cancer cell line; HeLa–Cervical cancer cell line.The H₂O extract had the mildest effect, with the most notable impact on the MCF-7 breast cancer cell line. Conversely, the CHCl₃ extract showed a more potent inhibition of cancer cell viability, especially affecting the T47D breast cancer line, while the MeOH and EtOH extracts, particularly from the FB strain, were identified as the most effective in inhibiting cancer cell growth.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



