
Submitted:
02 April 2025
Posted:
04 April 2025
You are already at the latest version
Abstract
Microplastics (MPs) are emerging pollutants that are ubiquitous in aquatic ecosystems and can affect the stability of aquatic food webs. They are intentionally produced in a size of less than 5 mm for specific purposes or are the result of fragmentation of larger plastic debris. Zooplankton can be affected directly by ingestion of MPs or indirectly by interference caused by suspended plastic particles. Various environmental agencies recommend the genus Moina for assessing risk from water pollutants. However, this genus has received less attention in research compared to non-indigenous cladocerans commonly used as test organisms. We evaluated the effects of artificially fragmented Acrylonitrile Butadiene Styrene microplastics (ABS-MPs) on key demographic parameters such as survival, mortality, life expectancy, fecundity, and feeding rates of Moina macrocopa americana. We exposed M. macrocopa neonates to a diet consisting of the green microalgae Chlorella vulgaris and ABS-MPs particles. Four treatments were set with different concentrations of ABS-MPs particles (5, 10, and 20 mg L-1). Survivorship, mortality, and reproduction were recorded daily until the last individual from the original cohort died. ABS-MPs significantly reduced M. macrocopa consumption rates of C. vulgaris, with an 85% decrease compared to the control. Although no statistically significant differences were found in life expectancy, net reproduction, or generation time among the toxic treatments, these parameters were drastically reduced compared to the control, even at the lowest concentration (5 mg L⁻¹); this resulted in a 34% reduction in average lifespan. The ABS-MPs interfere with the long-term population dynamics of M. macrocopa and change their consumption rates, potentially decreasing their fitness.
Keywords:
Microplastics
; Moina macrocopa
; ABS-MPs
1. Introduction
Since their first production in the 1950s, plastic materials have been considered an innovative product utilized in daily life. However, their excessive consumption, inadequate recycling, and difficulty in removal have led to environmental problems due to the high concentrations recorded [1,2]. Plastic particles smaller than 5 mm are classified as microplastics (MPs) and are almost imperceptible due to their size [3,4]. Primary MPs are manufactured industrially, such as microspheres for cosmetics, toothpaste, and facial scrubs. On the other hand, secondary MPs are formed by fragmentation or degradation of larger plastic items [5]. Nowadays, MPs are considered an emerging pollutant found in terrestrial and aquatic ecosystems, including coral reefs, Antarctic deep waters, and Arctic glaciers [6,7,8]. Research on microplastic pollution in aquatic environments is focused on marine habitats. However, freshwater ecosystems are also vulnerable due to direct anthropogenic influence, with plastic concentrations that could be comparable to, or even exceed, those observed in marine systems [9,10,11].
MPs access freshwater environments through different pathways, including treated water effluent, wind, stormwater runoff, surface runoff, and activities such as tourism [12,13]. Once MPs reach waterbodies, they negatively impact aquatic organisms when ingested voluntarily or involuntarily, disrupting the digestive tract and causing damage at all levels of biological organization [14,15]. The risk associated with MPs is also related to additives present in plastic materials, such as plasticizers, antioxidants, heat stabilizers, or slip agents. Additionally, MPs act as vectors for pollutants through adsorption and can host pathogens that adhere to the plastisphere [16,17].
Zooplanktonic organisms have an essential function in aquatic food webs as primary consumers [18]. Despite their importance and vulnerability, studies on their interaction with plastic particles are less common than those focused on mollusks and fish [19,20]. Zooplankton consume MPs through non-selective filtration or by mistaking them for prey, resulting in false satiation. This reduces their survival by interfering with the energetic budget associated with the presence of plastic particles and decreasing their natural ingestion rates, as observed in rotifers, copepods, and cladocerans [21,22,23]. The damage caused by MPs is related to factors such as size, shape, type, and concentration [24,25].
Most studies examining the impact of MPs on zooplankton have focused on standard polystyrene (PS) and polyethylene (PE) materials with spherical shapes and uniform sizes. However, in freshwater environments, most microplastics consist of irregular fragments and fibers of varying sizes [19]. In addition, other plastics commonly found in aquatic ecosystems include polyvinyl chloride (PVC), polyethylene terephthalate (PET), and acrylonitrile butadiene styrene (ABS), which are often overlooked [19,26]. ABS plastics are produced in large quantities, are highly toxic, and difficult to biodegrade. Despite their persistence, they have not been widely utilized in aquatic toxicity studies [27,28].
The genus Moina is a common cladoceran in tropical freshwater bodies, particularly in polluted waters. It exhibits high densities throughout the year making it an ideal organism to examine sensitivity to environmental stressors [29]. Their transparent body allows direct observation of contaminants through their gut, and their short lifespan and parthenogenetic reproduction make them suitable for both short-term (acute) and long-term (chronic) assessments. In addition, they are recognized as bioindicators, which increases their importance in assessing the effects of toxic substances, including microplastics [25]. This research aims to evaluate the effects of secondary ABS-MPs with irregular shapes and several sizes on demographic indicators such as survival, mortality, life expectancy, and fecundity in Moina macrocopa.
2. Materials and Methods
2.1. Moina macrocopa and Chlorella vulgaris Maintenance
The parthenogenetic populations of the cladoceran Moina macrocopa americana used to establish the stock cultures were obtained from the culture collection of the Aquatic Zoology Laboratory of UNAM, Mexico. These populations were isolated from the Ramsar site Lake Xochimilco and have been maintained for more than five years under controlled laboratory conditions. Cultures were established in EPA medium (Environmental Protection Agency medium, moderately hard water), consisting of g 1.9 g NaHCO3, 1.2 g CaSO4, 1.2 g MgSO4, and 0.04 g KCl dissolved in 20 L distilled water [30]. Stocks were maintained in 1 L glass beakers at room temperature with 12:12 h photoperiods. Their medium was renewed every three days, and the microcrustaceans were fed with the green microalga Chlorella vulgaris as an exclusive diet.
Axenic cultures of the microalga C. vulgaris (strain CL-V-3 CICESE, MX) were grown using a standardized Bold medium [30], supplemented with 0.5 g NaHCO3 every three days as an additional carbon source. Cultures were established in 2 L graduated Erlenmeyer flasks with 1.5 L of sterile medium Initial microalgal inoculation was initiated with 1 x 105 cells mL-1. The algae were grown in a batch regime at a temperature of 25 ± 2 °C, under a 16:8 h photoperiodic cycle, and with an irradiance of 150 μmol m-2 s-1. Moderate bubbling aeration was used to avoid cell sedimentation. C. vulgaris cells were decanted, the supernatant was removed, and the cells were rinsed and subsequently resuspended in an EPA medium. The algal cell density was quantified as number of cells per mL, using a hemocytometer (Neubauer chamber).
2.2. Preparation of ABS Microplastics
The small polymeric pieces were collected from an industrial ABS plastic manufacturing plant located in Mexico City. The synthetic material was mechanically shredded several times to create secondary ABS-MPs of less than 5 mm. The shredded plastics were then sieved to obtain ABS-MPs with sizes between 10 and 150 µm. The size of the ABS-MPs was verified microscopically. Dust and other contaminants were removed from the surface of the ABS-MPs by washing them with ethanol, rinsing with distilled water, and drying at 45 °C. The ABS-MPs were then resuspended in distilled water and the number of plastic particles was obtained using a Sedgewick-Rafter chamber. Resuspended ABS-MPs were diluted in an EPA medium to prepare a stock solution from which working concentrations used in the bioassays were obtained. As MPs tend to adhere to the forming plastic beds, the stock solution was sonicated for 3 min at 20 KHz before use.
2.3. Ingestion of Moina macrocopa to ABS-MPs
To analyze the ingestion of M. macrocopa, neonates of the third brood that were less than 6 h old were used. The microcrustaceans were fed ad libitum with C. vulgaris for 24 h prior to the experiments. Subsequently, ten neontaes were randomly transferred to 50 mL borosilicate vials containing 40 mL of EPA medium and subjected to a one-hour starvation period. Neonates were fed for 30 min with an initial concentration of 0.5 x 106 cells of C. vulgaris mL-1. Ingestion rates were evaluated under two conditions: C. vulgaris alone and combined with ABS-MPs at concentrations of 5, 10, and 20 mg L-1. Each test was performed in quadruplicate at room temperature. At the end of the filtration period, neonates were removed, and unconsumed cells were immediately fixed with 3% formaldehyde to ensure accurate quantification. The concentration of the cells was measured using a Neubauer chamber, and the ingestion rate was calculated by finding the difference between the initial and final algal densities.
2.4. Exposure of Moina macrocopa to ABS-MPs
The chronic experiments were performed as follows: 10 M. macrocopa neonates (less than 24 h born) were placed in 50 mL transparent containers with 40 mL of EPA medium (filtered with a cellulose filter (Millipore®) to avoid contamination). Each group of neonates was exposed to one of three concentrations of ABS-MPs (5, 10, and 20 mg L-1). All experiments were performed in quadruplicate, with each replicate containing C. vulgaris at a concentration of 0.5 x 10⁶ cells mL-1 as the sole food source. A control group without plastic particles subjected to the same diet as the toxic treatments was included (Figure 1). The vessels were covered with aluminum foil to prevent external contamination by airborne particles. The experiment was maintained under a 12:12 h light/dark cycle at room temperature (22 to 24 °C, confirmed with data recording equipment). Every 24 h, the EPA medium was replaced with C. vulgaris and ABS-MPs. Using a stereo microscope (Nikon SMZ800), the number of live and dead cladocerans was determined on a daily basis. Neonates were removed to exclusively monitor the initial cohort. With the data obtained, the demographic curves of survival, mortality and fecundity were plotted for each treatment.
In addition, the demographic variables of average life span, net reproduction, generation time and population growth were calculated [31]. In this instance, x represents age; lx is the proportion of individuals surviving to age x; mx denotes the proportion of females born during time interval x; ex corresponds to the average life expectancy of organisms alive at the beginning of the lifespan; Tx is the total number of individuals alive on average at age 0; nx indicates the initial population size at age 0. The constant e in the population rate equation is equal to 2.71828; r represents the intrinsic rate of population increase.
Average life span:
Net reproductive rate:
Generation time:
Rate of population increase:
2.5. Statistical Analysis
Statistical analyses were performed by analysis of variance (ANOVA) followed by Tukey's test to identify significant effects. All analyses were performed using SigmaPlot® 11 software (Systat Software, Inc., San Jose, CA, USA, https://systatsoftware.com).
3. Results
3.1. Cells Ingestion of Moina macrocopa
The presence of microplastics altered the number of cells ingested during the filtration process of M. macrocopa. The general trend showed that as the concentration of ABS-MPs increased, the number of food cells ingested decreased significantly (Figure 2). In the control treatment, the cladocerans consumed 625 cells of C. vulgaris ind-1 min-1. In contrast, at 20 mg L-1, an 85% reduction (94 cells ind-1 min-1) was observed. Neonates exposed to 5 mg L-1 ABS-MPs consumed 341 cells ind-1 min-1, which corresponded to approximately half the amount observed in the treatment without ABS-MPs. This indicates that the presence of ABS-MP influences filtration, even at low concentrations. The treatments at 10 mg and 20 mg L-1 did not show significant differences from each other (Table 1, P > 0.05).
3.2. Survivorship
Survival of cladocerans exposed to microparticles decreased compared to the control treatment, which showed a mean survival of 16 days. In contrast, the ABS-MPs treatments resulted in survivorship ranging from 11 to 13 days (Figure 3). The Kaplan-Meier survival analysis indicated a significant difference in survival curves (P = 0.038). However, the Holm-Sidak post hoc test (P < 0.05) suggested that, although there was an overall trend towards reduced survival, the variations between groups were not large enough to be statistically significant. Survival curves for controls indicated that cladocerans died mainly in the last days of their life cycle, corresponding to a type I curve. In treatments with the presence of ABS-MPs, M. macrocopa showed a decrease in survival, with a reduction of ~39% in the first four days. By day 10, when the control population reached a mean survival rate of 0.5 (lx), toxic treatments showed a 70% decrease in survivorship.
Figure 3.
Survival of the cladoceran Moina macrocopa exposed to different concentrations of ABS secondary microplastics (5, 10 and 20 mg L⁻¹) and control. Values are expressed as means ± standard error based on four replicates.
Figure 3.
Survival of the cladoceran Moina macrocopa exposed to different concentrations of ABS secondary microplastics (5, 10 and 20 mg L⁻¹) and control. Values are expressed as means ± standard error based on four replicates.

Figure 3.
Mortality rate of the cladoceran Moina macrocopa exposed to different concentrations of ABS secondary microplastics (5, 10 and 20 mg L⁻¹) and control. Values are expressed as means ± standard error based on four replicates.
Figure 3.
Mortality rate of the cladoceran Moina macrocopa exposed to different concentrations of ABS secondary microplastics (5, 10 and 20 mg L⁻¹) and control. Values are expressed as means ± standard error based on four replicates.
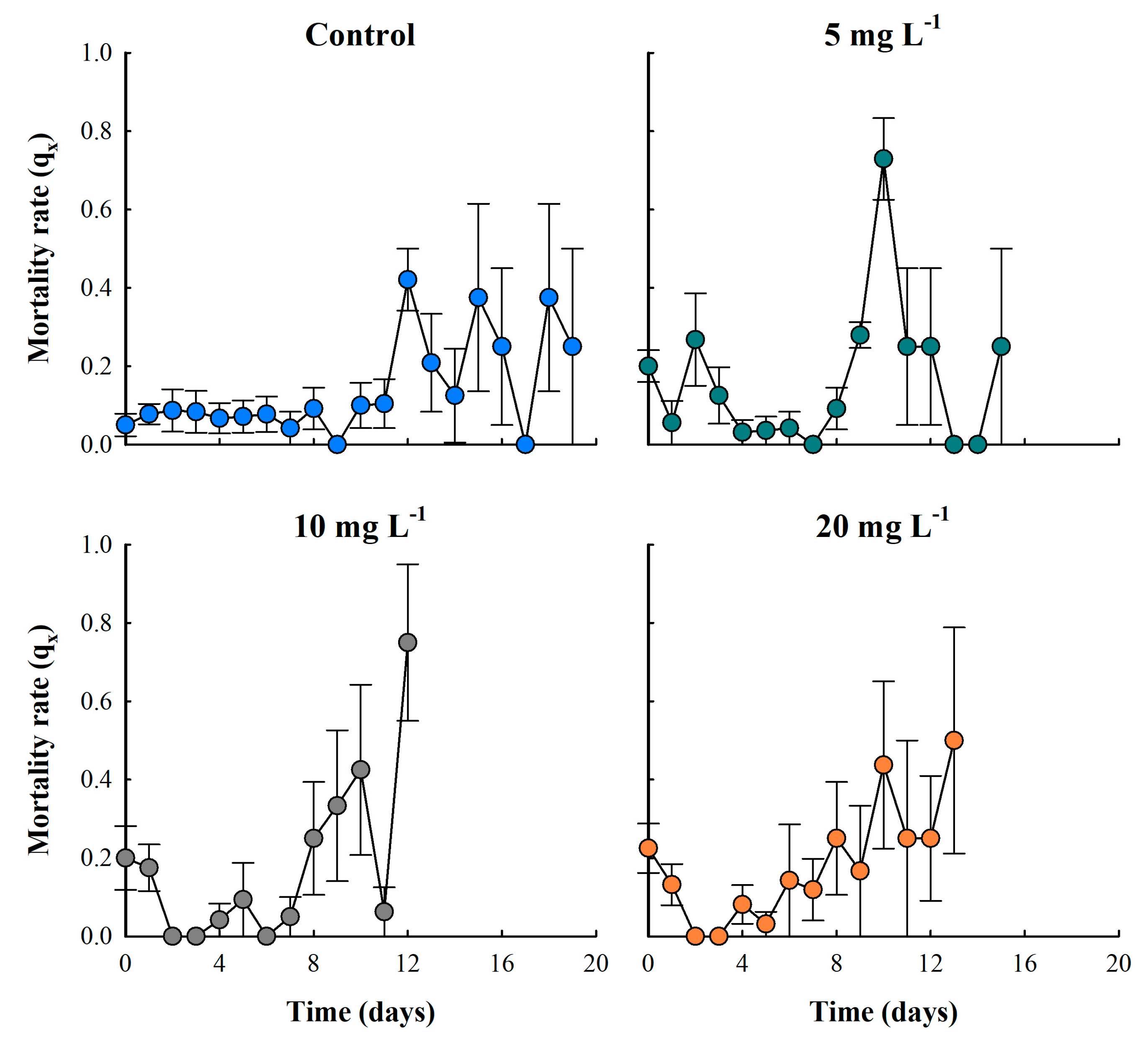
3.3. Mortality Rate
The particulate-free treatment (control) showed low mortality rates during the first 11 days. After this period, the population began experiencing senescence, resulting in a continuous increase in mortality. In the particle concentration treatment of 5 mg L-1, the lowest mortality rate (qx) was observed from days four to eight. In the 10 mg L-1 treatment, the lowest mortality rate occurred between days two and seven, while in the highest concentration treatment of 20 mg L-1, mortality rates remained low between days two and six of life (Figure 4).
Figure 4.
Fecundity (number of neonates per female) of the cladoceran Moina macrocopa exposed to different concentrations of ABS secondary microplastics (5, 10 and 20 mg L-1). Values are expressed as means ± standard error based on four replicates.
Figure 4.
Fecundity (number of neonates per female) of the cladoceran Moina macrocopa exposed to different concentrations of ABS secondary microplastics (5, 10 and 20 mg L-1). Values are expressed as means ± standard error based on four replicates.
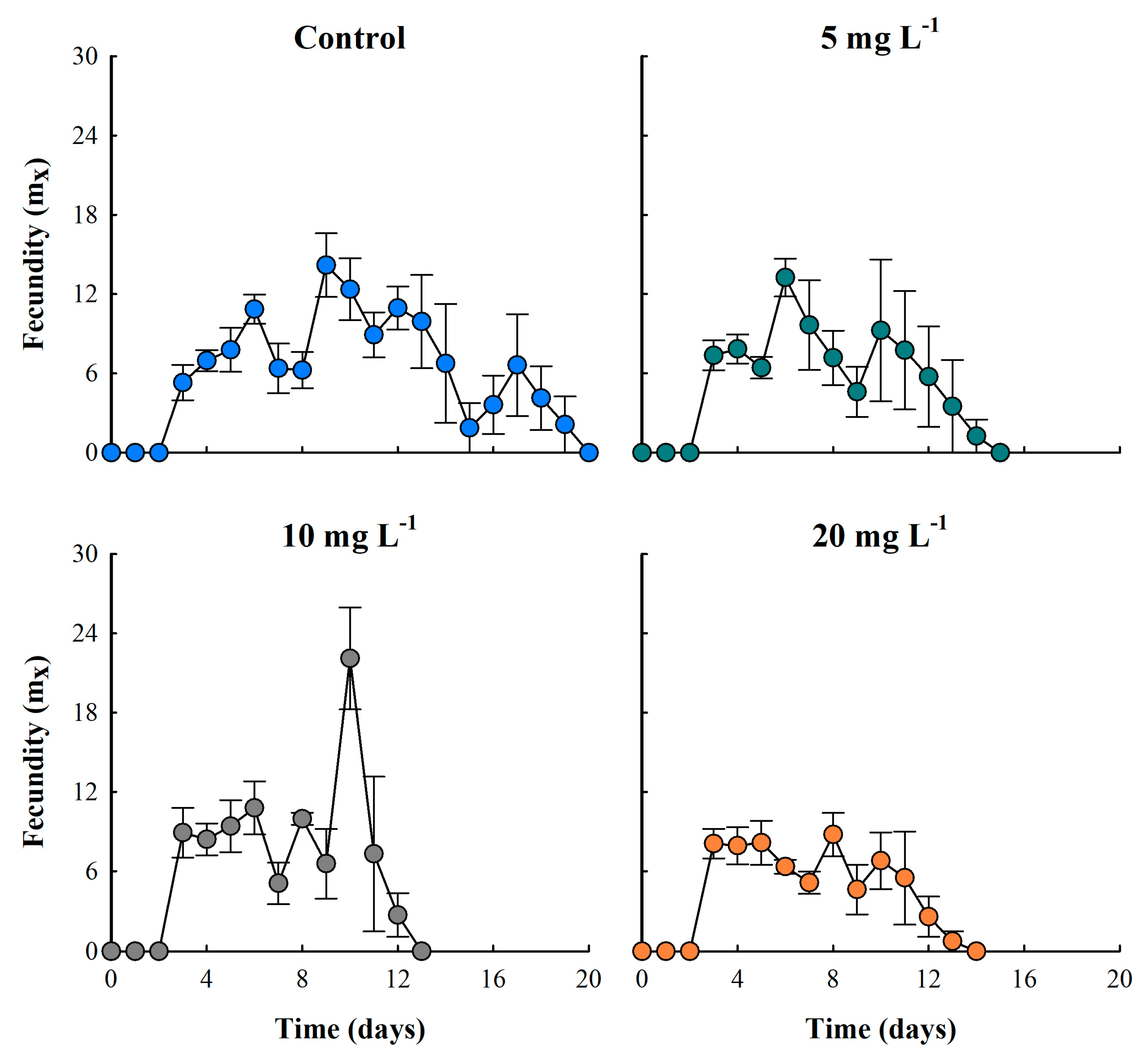
3.4. Fecundity
Fecundity curves of M. macrocopa, evaluated by age-specific analysis, showed that all treatments reached reproductive maturity at 72 hours (day three). In the control treatment, this variable remained constant for 17 days. On the contrary, in the ABS-MPs treatments, particularly in the concentrations of 10 and 20 mg L-1, the fecundity was inversely proportional to concentration, showing a 37.7% reduction compared to the control (Figure 5). Nevertheless, the 10 mg L-1 treatment exhibited the highest reproductive rates, with a peak production of 22.1 neonates per female per day, representing a 35.8% increase in reproductive effort compared to the control. Fluctuations in fecundity seen in the ABS-MPs treatments were most likely due to the toxic effects of MPs or changes in energy cost. Throughout the experiment, MPs were observed stored in the thoracic appendages of cladocerans, in some cases blocking the intestine and in others adhering to the carapace (Figure 5).
Figure 4.
Life expectancy of the cladoceran Moina macrocopa; Generation time; Net reproduction and Finite rate of increase, exposed to different concentrations of ABS secondary microplastics (5, 10 and 20 mg L-1) and control. Values are expressed as means ± standard error based on four replicates.
Figure 4.
Life expectancy of the cladoceran Moina macrocopa; Generation time; Net reproduction and Finite rate of increase, exposed to different concentrations of ABS secondary microplastics (5, 10 and 20 mg L-1) and control. Values are expressed as means ± standard error based on four replicates.
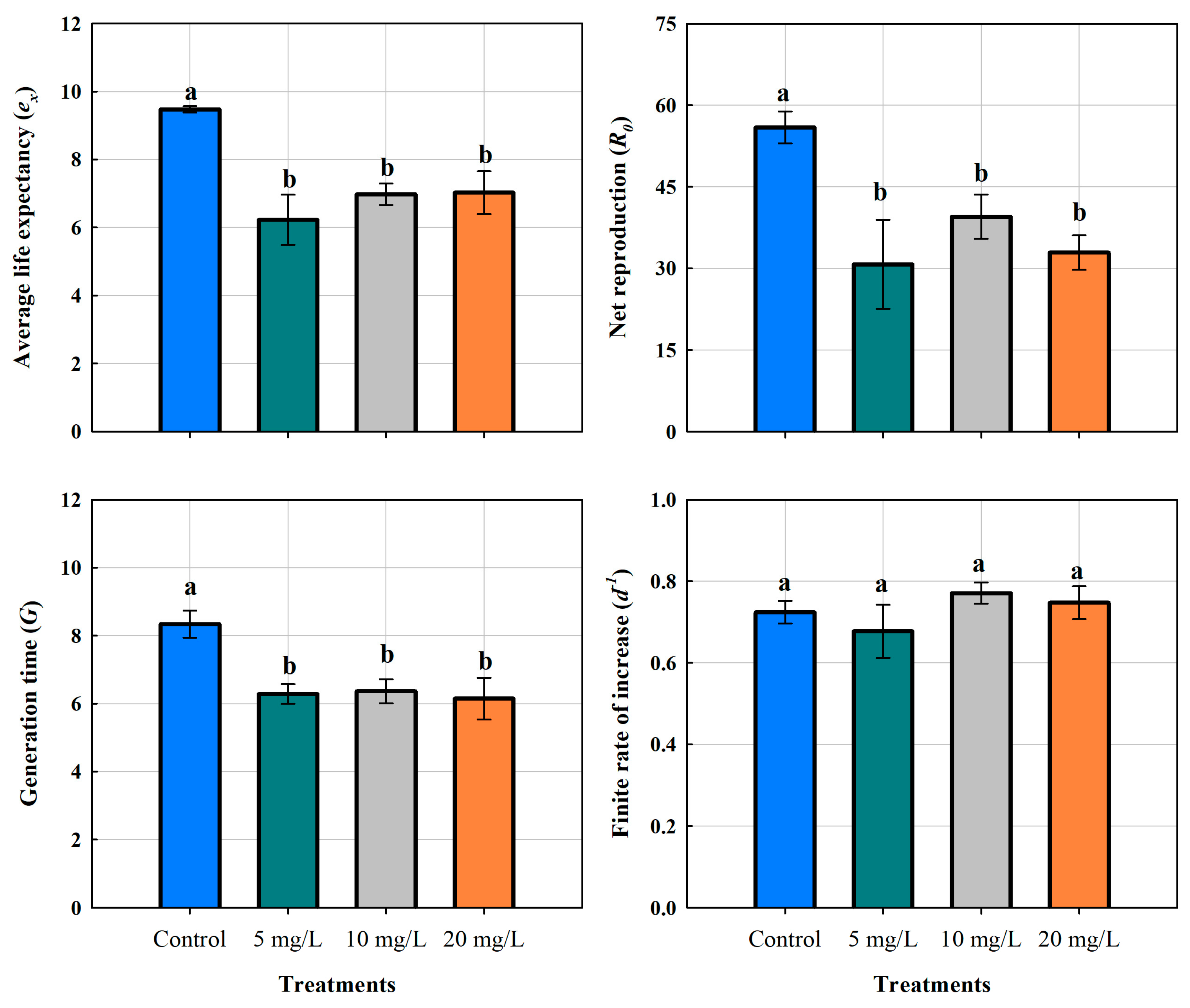
3.5. Life Table Parameters
The life expectancy (eₓ) of M. macrocopa in the control group was 9.48 ± 0.09 days. On the other hand, in the ABS-MPs treatments it decreased significantly to 6.23 ± 0.74, 6.98 ± 0.31, and 7.03 ± 0.63 days for 5, 10 and 20 mg L-1, respectively. A one-way ANOVA confirmed significant differences (P < 0.01), indicating that the presence of MPs had a negative effect on this demographic variable (Table 2). However, MPs concentration did not result in statistically significant differences. Tukey's post hoc test (P < 0.05) further confirmed that all ABS-MP treatments resulted in a comparable reduction in life expectancy, suggesting that even the lowest concentration (5 mg L-1) had a negative impact on the survival of M. macrocopa, reducing it by 34.3%.
The net reproduction rate (R₀) showed a significant decrease in response to MPs exposure. In the control group, the mean value was 55.93 ± 5.89, while all treated groups showed lower means: 30.75 ± 13.39 (5 mg L-1), 39.50 ± 8.12 (10 mg L-1), and 32.90 ± 6.35 (20 mg L-1). The greatest variability in responses was observed in the 5 mg L-1 treatment, suggesting inconsistent responses amongst individuals. Remarkably, the 10 mg L-1 group presented an intermediate effect, while the 20 mg L-1 treatment did not show a clear dose-dependent trend, as its mean value was comparable to that of 5 mg L-1. The 5 mg L-1 concentration caused a reduction of 25 offspring per female, representing approximately 45% fewer compared to the control group (P < 0.05, Tukey's test). Nevertheless, the same was observed at the medium and high concentrations, which were significantly different from the control but not from the 5 mg L-1 treatment (P > 0.05, Tukey's test). These results indicate that ABS-MPs affect reproductivity, but the response does not follow a strictly linear pattern with increasing concentrations.
Cladocerans exposed to MPs showed a generation time of 6.28 ± 0.68 days for 5 mg L-1, 6.36 ± 0.70 days for 10 mg L-1, and 6.15 ± 1.16 days for 20 mg L-1 (Figure 6). The significant decrease in generation time was due to fragmented plastics exposure (P < 0.05). In the control group, the mean was 8.33 ± 0.77 days, which is 24-26% longer. Although all MPs treatments resulted in a significantly shorter generation time, no significant differences were detected among the three concentrations (P > 0.05). Furthermore, individuals from the 20 mg L-1 treatment showed the lowest values (4.69 days), which could indicate a stress-induced acceleration of reproduction.
Growth rates of M. macrocopa ranged from 0.55 to 0.87 d-1. Statistical analysis revealed no significant differences between treatments (P > 0.05). The mean growth rates were 0.72 ± 0.05 d-1 in the control group, 0.68 ± 0.11 d-1 at 5 mg L-1, 0.77 ± 0.05 d-1 at 10 mg L-1, and 0.75 ± 0.07 d-1 at 20 mg L-1 (Figure 6). These results suggest that increase rates remained stable over time, implying a combination of reproductive strategies, at least in the F0 generation. Further studies are required to investigate these mechanisms and their possible impact on long-term population dynamics.
4. Discussion
Some authors have reported that microplastics in freshwater environments commonly range between 20 and 100 μm [32] and that cladocerans can ingest particles between 23 and 264 μm [33]. Similarly, it has been shown that M. macrocopa can grow and reproduce when feeding on organic particles between 35-40 μm [34], which supports the findings presented here.
Our results indicate that the effects of ABS secondary microplastics (10-150 µm) on the demographic variables of Moina macrocopa are not necessarily concentration-dependent. Even at the lowest concentration tested, we observed detrimental effects on cladocerans comparable to those at medium and high concentrations (10 and 20 mg L-1). We suggest that the observed decrease in fitness may be due to intestinal obstruction and reduced efficiency of food intake. Both digestive obstruction and external adhesion impair cladoceran fitness, leading to abnormal movement and reduced feeding efficiency [35,36]. Consistent with this, our study found an 85% reduction in green algal consumption. Similarly, Reyes-Santillán et al. [37] reported that Daphnia pulex exhibited a 50% decrease in ingestion rate and altered heart rate when exposed to 30 µm plastic microspheres at 40 mg L-1. Even when plastic particles are not ingested due to their size, their mere presence can increase energy expenditure by interfering with algal consumption and requiring additional energy for the removal of clogged plastic particles [23]. De Felice et al. [38] reported that microplastics induced behavioral changes in terms of swimming activity, phototactic behavior and reproduction.
Ingestion has been shown to be a critical factor in determining the negative effects of microplastics and represents the first indication of their mechanistic impact. Moina macrocopa has a non-selective filter-feeding behavior, which favors the ingestion of microplastics along with organic particles [39]. Consequently, microplastics lodge between the thoracic appendages and clog the gut. Although not measured, a personal observation we noted is that cladocerans frequently used their postadomen, whose function is cleaning, suggesting stress caused by the presence of particles. This is in agreement with the findings of Rehse et al. [39] and Frydkjær et al. [24], who reported that secondary microplastics, due to their irregular morphology, caused erratic movements.
Mortality in M. macrocopa increased even at the lowest microplastic concentration, consistent with findings by Castro et al. [40] for D. similis. The authors attributed this to appendage damage and reduced feeding. Similarly, Ogonowski et al. [41] reported increased mortality in D. magna due to lower algal consumption. Diet appears to play a critical role, as Tang et al. [42] found increased expression of arginine kinase, an enzyme involved in energy production, in D. magna exposed to microplastics, suggesting an attempt to compensate for food limitation.
Fecundity and net reproduction were significantly reduced until 45% in the presence of ABS-MPs versus control, contrary to what De Felice et al. [35] reported. They showed an increase of offspring production in D. magna exposed to primary microplastics. This discrepancy may be due to differences in microplastic type. Castro et al. [40] and Ziajahromi et al. [43] observed reduced fecundity in the cladoceran Ceriodaphnia dubia and D. similis cladocerans exposed to secondary microplastics, associated to the expended energy during swminming. Given the high energy demands of reproduction in cladocerans [44], gut blockage likely impaired nutrition, further reducing fecundity.
Mortality of Moina macrocopa increased even at the lowest concentration of microplastics, consistent with the findings of Castro et al. [40] for Daphnia similis. The authors attributed this to appendage damage and reduced feeding. Similarly, Ogonowski et al. [41] reported increased mortality in Daphnia magna due to reduced algal consumption. Diet plays a critical role, Tang et al. [42] found increased expression of arginine kinase, an enzyme involved in energy production in D. magna exposed to microplastics, suggesting an attempt to compensate for food limitation.
Fecundity and net reproduction decreased significantly, by up to 45%, in the presence of ABS-MPs compared to the control, contrary to the findings of De Felice et al. [38], who reported increased offspring production in D. magna exposed to primary microplastics. This dissimilarity may be due to the type of microplastic. Castro et al. [40] and Ziajahromi et al. [43] observed a decrease in fecundity variables in Ceriodaphnia dubia and D. similis exposed to secondary microplastics due to higher energy expenditure.
Interestingly, microplastics did not alter the timing of the first reproductive event in M. macrocopa, as both control and exposed individuals released their first offspring on the fourth day. This contrasts with De Felice et al. [38], who reported earlier reproduction in D. magna exposed to microplastics, possibly as a survival strategy. In our study, both fecundity and survival declined at the lowest microplastic concentration, suggesting that M. macrocopa did not prioritize reproduction over survival.
Cladoceran survival depends on whether they are exposed to primary or secondary microplastics. While smooth microbeads accumulate in the digestive tract without causing significant mortality [22,41], secondary microplastics with irregular morphology cause internal and external injury. Frydkjær et al. [24] found that cladocerans could rapidly remove smooth microspheres but retained those with irregular shapes, which compromised their survival. A similar pattern was observed in our experiments, where the life expectancy was reduced by approximately 35% in the presence of plastic materials.
5. Conclusions
In this study the effects of three different concentrations of ABS-MPs on the life table of the freshwater cladoceran M. macrocopa were evaluated. It was observed that even at the lowest concentration tested had significantly negative consequences on the life table of M. macrocopa, decreasing survival, life expectancy and fecundity, and increasing mortality. The interaction of zooplankton with the ABS particles obstructed their digestive system, causing immobilization and impaired swimming, which led to deficient feeding and declining nutrition.
Author Contributions
Work design and provision of background theory and intellectual property was developed by CAZB, DJCH and PRG. Data collection (experiments) and analysis were performed by DLM, DJCH and CAZB. Writing was developed by DLM and CAZB. All authors revised subsequent versions of the manuscript.
Funding
This research was funded by Posgrado en Ciencias Biológicas (PCBiol), Universidad Nacional Autónoma de México (UNAM); El Consejo Nacional de Humanidades, Ciencias y Tecnología (CONAHCyT) (1145780) and DGAPA-PAPIIT (IN229523).
Institutional Review Board Statement
Not applicable for studies not involving humans or animals.
Data Availability Statement
All the data produced or examined in this study are provided within this published article.
Acknowledgments
MGDL thanks Posgrado en Ciencias Biológicas (PCBiol) by Universidad Nacional Autónoma de México (UNAM) for its support in carrying out this project and CONAHCyT for a master scholarship granted (1145780). All authors thank Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (DGAPA-PAPIIT IN229523) for financial assistance. CAZB thanks Dr. Nandini Sarma for supplying Moina macrocopa americana cultures for the experiments and CONAHCyT Mexico (492558). All the authors thank the discount granted.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MPs | Microplastics |
| ABS-MPs | Acrylonitrile Butadiene Styrene microplastics |
| PS | polystyrene |
| PE | polyethylene |
| PVC | polyvinyl chloride |
| PET | polyethylene terephthalate |
References
- Barnes, D.K.; Galgani, F.; Thompson, R.C.; Barlaz, M. Accumulation and fragmentation of plastic debris in global environments. Philosophical Transactions of the Royal Society B: Biological Sciences 1985, 364, 1985–1998. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, T.; Sakane, F.; Kinoshita, C.; Sato, K.; Mizukawa, K.; Takada, H. Covid-19-derived plastic debris contaminating marine ecosystem: alert from a sea turtle. Marine Pollution Bulletin 2022, 175, 113389. [Google Scholar] [CrossRef] [PubMed]
- Frias, J.P.; Nash, R. Microplastics: Finding a consensus on the definition. Marine Pollution Bulletin 2019, 138, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.; Hamid, A.K.; Krebsbach, S.A.; He, J.; Wang, D. Critical review of microplastics removal from the environment. Chemosphere 2022, 293, 133557. [Google Scholar] [CrossRef]
- Andrady, A.L. Microplastics in the marine environment. Marine Pollution Bulletin 2011, 62, 1596–1605. [Google Scholar] [CrossRef]
- Ramos, L.; Berenstein, G.; Hughes, E.A.; Zalts, A.; Montserrat, J.M. Polyethylene film incorporation into the horticultural soil of small periurban production units in Argentina. Science of the Total Environment 2015, 523, 74–81. [Google Scholar] [CrossRef]
- Hall, N.M.; Berry, K.L.E.; Rintoul, L.; Hoogenboom, M.O. Microplastic ingestion by scleractinian corals. Marine Biology 2015, 162, 725–732. [Google Scholar] [CrossRef]
- Cunningham, E.M.; Ehlers, S.M.; Dick, J.T.; Sigwart, J.D.; Linse, K.; Dick, J.J.; Kiriakoulakis, K. High abundances of microplastic pollution in deep-sea sediments: evidence from Antarctica and the Southern Ocean. Environmental Science & Technology 2020, 54, 13661–13671. [Google Scholar] [CrossRef]
- McCormick, A.R.; Hoellein, T.J.; London, M.G.; Hittie, J.; Scott, J.W.; Kelly, J.J. Microplastic in surface waters of urban rivers: concentration, sources, and associated bacterial assemblages. Ecosphere 2016, 7, e01556. [Google Scholar] [CrossRef]
- D’Avignon, G.; Gregory-Eaves, I.; Ricciardi, A. Microplastics in lakes and rivers: an issue of emerging significance to limnology. Environmental Reviews 2022, 30, 228–244. [Google Scholar] [CrossRef]
- Nava, V. , Chandra, S., Aherne, J., Alfonso, M. B., Antão-Geraldes, A. M., Attermeyer, K., & Leoni, B. (2023). Plastic debris in lakes and reservoirs. Nature 2023, 619, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Dris, R.; Gasperi, J.; Tassin, B. Sources and fate of microplastics in urban areas: a focus on Paris Megacity. In Freshwater microplastics: emerging environmental contaminants?. M. Wagner; S. Lambert Eds.; Springer, Heidelberg, 2017; pp. 69-84. [CrossRef]
- Lambert, S.; Wagner, M. Microplastics are contaminants of emerging concern in freshwater environments: an overview. In Freshwater microplastics: emerging environmental contaminants? M. Wagner & S. Lambert Eds.; Springer, Heidelberg, 2018; pp. 1-23. [CrossRef]
- Hurley, R.R.; Woodward, J.C.; Rothwell, J.J. Ingestion of microplastics by freshwater tubifex worms. Environmental Science & Technology 2017, 51, 12844–12851. [Google Scholar] [CrossRef]
- Galloway, T.S.; Cole, M.; Lewis, C. Interactions of microplastic debris throughout the marine ecosystem. Nature Ecology & Evolution 2017, 1, 0116. [Google Scholar] [CrossRef]
- Wang, T.; Yu, C.; Chu, Q.; Wang, F.; Lan, T.; Wang, J. Adsorption behavior and mechanism of five pesticides on microplastics from agricultural polyethylene films. Chemosphere 2020, 244, 12549. [Google Scholar] [CrossRef]
- Cao, Y.; Zhao, M.; Ma, X.; Song, Y.; Zuo, S.; Li, H.; Deng, W. A critical review on the interactions of microplastics with heavy metals: mechanism and their combined effect on organisms and humans. Science of the Total Environment 2021, 788, 147620. [Google Scholar] [CrossRef]
- Turner, J.T. Zooplankton fecal pellets, marine snow, phytodetritus, and the ocean’s biological pump. Progress in Oceanography, 2015, 130, 205–248. [Google Scholar] [CrossRef]
- Rodrigues, S.M.; Elliott, M.; Almeida, C.M.R.; Ramos, S. Microplastics and plankton: Knowledge from laboratory and field studies to distinguish contamination from pollution. Journal of Hazardous Materials 2021, 417, 126057. [Google Scholar] [CrossRef]
- Cera, A.; Scalici, M. Freshwater wild biota exposure to microplastics: A global perspective. Ecology and Evolution 2021, 11, 9904–9916. [Google Scholar] [CrossRef]
- Lan, R.; Wei, Y. ,Xue, R. Uptake of polystyrene microplastics by marine rotifers under different experimental conditions. Environmental Earth Sciences 2021, 687, 012071. [Google Scholar] [CrossRef]
- Huang, C.H. , Chu, T.W., Kuo, C.H., Hong, M.C., Chen, Y.Y., & Chen, B. Effects of microplastics on reproduction and growth of freshwater live feeds Daphnia magna. Fishes 2022, 7, 181. [Google Scholar] [CrossRef]
- Zamora-Barrios, C. A.; Nandini, S.; Sarma, S. S. S. Effect of microplastics on the demography of Brachionus calyciflorus Pallas (Rotifera) over successive generations. Aquatic Toxicology 2024, 275, 107061. [Google Scholar] [CrossRef] [PubMed]
- Frydkjær, C. K.; Iversen, N.; Roslev, P. Ingestion and egestion of microplastics by the cladoceran Daphnia magna: Effects of regular and irregular shaped plastic and sorbed phenanthrene. Bulletin of Environmental Contamination and Toxicology 2017, 99, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Jemec, A.; Horvat, P.; Kunej, U.; Bele, M.; Kržan, A. Uptake and effects of microplastic textile fibers on freshwater crustacean Daphnia magna. Environmental Pollution 2016, 219, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Sighicelli, M.; Pietrelli, L.; Lecce, F.; Iannilli, V.; Falconieri, M.; Coscia, L.; Di Vito, S.; Nuglio, S.; Zampetti, G. Microplastic pollution in the surface waters of Italian Subalpine Lakes. Environmental Pollution 2018, 236, 645–651. [Google Scholar] [CrossRef]
- Lithner, D.; Larsson, Å.; Dave, G. Environmental and health hazard ranking and assessment of plastic polymers based on chemical composition. Science of the Total Environment 2011, 409, 3309–3324. [Google Scholar] [CrossRef]
- Yuan, Z.; Nag, R. ; Cummins, E Ranking of potential hazards from microplastic polymers in the marine environment. Journal of Hazardous Materials 2022, 429, 128399. [Google Scholar] [CrossRef]
- Vignatti, A. M.; Cabrera, G. C.; Echaniz, S. A. Distribution and biological aspects of the introduced species Moina macrocopa (Straus, 1820) (Crustacea, Cladocera) in the semi-arid central region of Argentina. Biota Neotropica 2013, 13, 86–92. [Google Scholar] [CrossRef]
- Weber, C. I. Methods for measuring the acute toxicity of effluents and receiving waters to freshwater and marine organisms (4th ed.). United States Environmental Protection Agency, Cincinnati, Ohio, 1993, EPA/600/4-90/027F, xv.
- KREBS.
- Uurasjärvi, E.; Hartikainen, S.; Setälä, O. , Lehtiniemi, M.; Koistinen, A. Microplastic concentrations, size distribution, and polymer types in the surface waters of a northern European lake. Water Environment Research 2020, 92, 149–156. [Google Scholar] [CrossRef]
- Kokalj, A. J.; Kunej, U.; Skalar, T. Screening study of four environmentally relevant microplastic pollutants: Uptake and effects on Daphnia magna and Artemia franciscana. Chemosphere 2018, 208, 522–529. [Google Scholar] [CrossRef]
- Kumar, R. , Hwang, J. S. Ontogenetic shifts in the ability of the cladoceran, Moina macrocopa Straus and Ceriodaphnia cornuta Sars to utilize ciliated protists as food source. International Review of Hydrobiology, 2008, 93, 284–296. [Google Scholar] [CrossRef]
- Cole, M0.; Lindeque, P. K.; Fileman, E.; Halsband, C.; Goodhead, R.; Moger, J., Galloway, T. S. Microplastic ingestion by zooplankton. Environmental Science & Technology 2013, 47, 6646–6655. [CrossRef]
- Cole, M.; Lindeque, P. K.; Fileman, E.; Halsband, C.; Galloway, T. S. The impact of polystyrene microplastics on feeding, function, and fecundity in the marine copepod Calanus helgolandicus. Environmental Science & Technology 2015, 49, 1130–1137. [Google Scholar] [CrossRef]
- Reyes-Santillán, M. C., Nandini, S., Sarma, S. S. S. The combined effect of temperature and microplastics on Daphnia pulex Leydig, 1860 (Cladocera). Inland Waters, 2025 (just-accepted), 1.
- De Felice, B.; Sabatini, V.; Antenucci, S.; Gattoni, G.; Santo, N.; Bacchetta, R.; Ortenzi, M. A. , Parolini, M. Polystyrene microplastics ingestion induced behavioral effects to the cladoceran Daphnia magna. Chemosphere 2019, 231, 423–431. [Google Scholar] [CrossRef]
- Rehse, S.; Kloas, W.; Zarfl, C. Short-term exposure with high concentrations of pristine microplastic particles leads to immobilization of Daphnia magna. Chemosphere 2016, 153, 91–99. [Google Scholar] [CrossRef]
- Castro, D. G. D.; Destro, A. L. F.; Coimbra, E. C. L.; Silva, A. L. L. D.; Mounteer, A. H. Effects of PET microplastics on the freshwater crustacean Daphnia similis Claus, 1976. Acta Limnologica Brasiliensis 2023, 35, e6. [Google Scholar] [CrossRef]
- Ogonowski, M.; Schür, C.; Jarsén, Å.; Gorokhova, E. The effects of natural and anthropogenic microparticles on individual fitness in Daphnia magna. PLOS ONE, 2016, 11, e0155063. [Google Scholar] [CrossRef]
- Tang, J.; Wang, X.; Yin, J.; Han, Y.; Yang, J.; Lu, X.; Xie, T.; Akbar, S.; Lyu, K.; Yang, Z. Molecular characterization of thioredoxin reductase in waterflea Daphnia magna and its expression regulation by polystyrene microplastics. Aquatic Toxicology 2019, 208, 90–97. [Google Scholar] [CrossRef]
- Ziajahromi, S.; Kumar, A.; Neale, P. A. , Leusch, F. D. Impact of microplastic beads and fibers on waterflea (Ceriodaphnia dubia) survival, growth, and reproduction: Implications of single and mixture exposures. Environmental Science & Technology 2017, 51, 13397–13406. [Google Scholar] [CrossRef]
- Enserink, L.; de la Haye, M.; Maas, H. Reproductive strategy of Daphnia magna: Implications for chronic toxicity tests. Aquatic Toxicology 1993, 25, 111–123. [Google Scholar] [CrossRef]
Figure 1.
Experimental design.
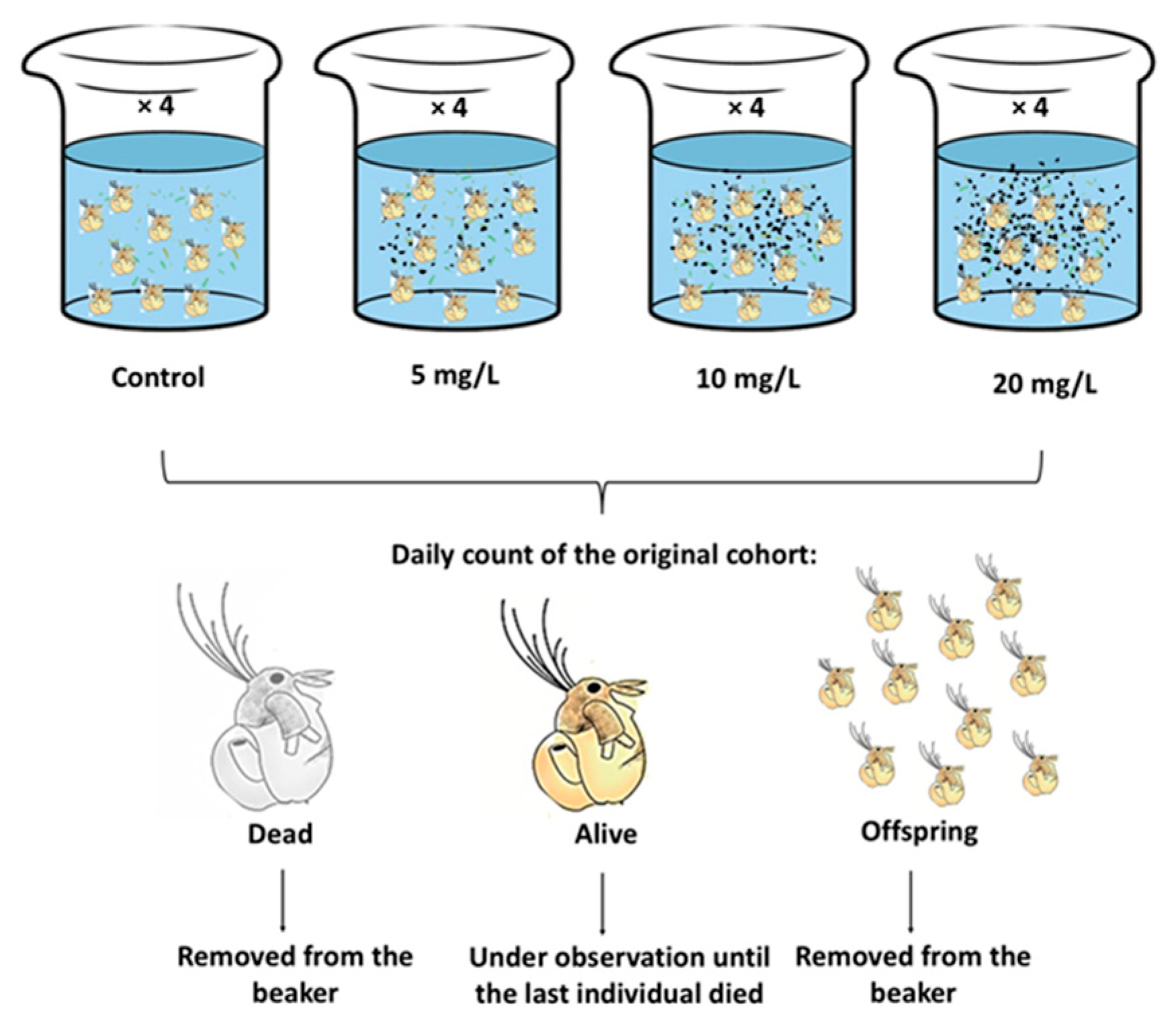
Figure 2.
Ingestion of C. vulgaris by the cladoceran Moina macrocopa americana to ABS-MPs.

Figure 5.
Secondary microplastics clogging appendages and gut, with others attached to the carapace of the cladoceran Moina macrocopa.
Figure 5.
Secondary microplastics clogging appendages and gut, with others attached to the carapace of the cladoceran Moina macrocopa.


Table 1.
One-way analysis of variance applied to ingestion rates of Moina macrocopa exposed to ABS-MPs. The table provides information on degrees of freedom (DF), the sum of squares (SS), mean squared (MS), Fisher’s test statistic (F), and the corresponding P- values.
Table 1.
One-way analysis of variance applied to ingestion rates of Moina macrocopa exposed to ABS-MPs. The table provides information on degrees of freedom (DF), the sum of squares (SS), mean squared (MS), Fisher’s test statistic (F), and the corresponding P- values.
| Source of Variation | DF | SS | MS | F | P |
|---|---|---|---|---|---|
| Between Groups | 3 | 179587.383 | 59862.461 | 22.398 | <0.001 |
| Residual | 12 | 32071.468 | 2672.622 | ||
| Total | 15 | 211658.851 |
Table 2.
One-way analysis of variance (ANOVA) applied to the demographic variables of Moina macrocopa exposed to three concentrations of ABS microplastics (5, 10, and 20 mg L⁻¹). The reported statistical parameters include degrees of freedom (DF), sum of squares (SS), mean square (MS), Fisher's test statistic (F), and the corresponding P-values.
Table 2.
One-way analysis of variance (ANOVA) applied to the demographic variables of Moina macrocopa exposed to three concentrations of ABS microplastics (5, 10, and 20 mg L⁻¹). The reported statistical parameters include degrees of freedom (DF), sum of squares (SS), mean square (MS), Fisher's test statistic (F), and the corresponding P-values.
| Source of Variation | DF | SS | MS | F | P |
|---|---|---|---|---|---|
| Average life span | |||||
| Between Groups | 3 | 25.007 | 8.336 | 10.474 | 0.001 |
| Residual | 12 | 9.550 | 0.796 | ||
| Total | 15 | 34.557 | |||
| Net reproductive rate | |||||
| Between Groups | 3 | 2114.262 | 704.754 | 10.429 | 0.001 |
| Residual | 12 | 810.915 | 67.576 | ||
| Total | 15 | 2925.177 | |||
| Generation time | |||||
| Between Groups | 3 | 12.974 | 4.325 | 5.826 | 0.011 |
| Residual | 12 | 8.908 | 0.742 | ||
| Total | 15 | 21.882 | |||
| Population growth rate | |||||
| Between Groups | 3 | 0.0191 | 0.00638 | 0.872 | 0.483 |
| Residual | 12 | 0.0879 | 0.00732 | ||
| Total | 15 | 0.107 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



