Submitted:
01 April 2025
Posted:
01 April 2025
You are already at the latest version
Abstract
The increasing frequency of fish escapes from aquaculture facilities poses ecological, economic, and traceability challenges. This study investigates the potential of heavy metals, fatty acid profiles, and NMR-based metabolomics to distinguish between wild, cultured, and escaped individuals of three Mediterranean fish species: Sparus aurata (gilthead seabream), Dicentrarchus labrax (European seabass), and Argyrosomus regius (meagre). Muscle tissues were analyzed using ICP-MS, GC-MS, and 1H NMR spectroscopy to quantify 15 trace metals, 35 fatty acids, and polar/apolar metabolites. Wild seabream exhibited significantly higher levels of arsenic, selenium, and mercury, and a lipid profile rich in DHA and ARA, contrasting with the linoleic acid-enriched profile of cultured fish. Metabolomic analyses revealed elevated TMAO levels in wild specimens, serving as a robust marker of environmental adaptation, while escaped fish showed intermediate metabolic signatures. The integration of multivariate statistics (MDS and PLS-LDA) enabled effective classification of fish origin, particularly in seabream. These findings highlight the utility of combining chemical and omics-based tools to enhance seafood traceability, improve aquaculture sustainability, and prevent seafood fraud.
Keywords:
Fish traceability
; Aquaculture escapes
; Heavy metals
; Fatty acid profiling
; NMR metabolomics
; Seafood authentication
; Environmental biomarkers
; Marine fish ecology
1. Introduction
The Mediterranean Sea exhibits unique environmental characteristics due to its complex topography, geographical location between the subtropics and mid-latitudes, and dynamic meteorological patterns. These factors contribute to the frequent formation of low-pressure systems, as documented in several climatological studies [1]. The increasing prevalence of extreme warm events in the Mediterranean Basin aligns with global climate change trends and rising seasonal average temperatures. Multiple simulation models have predicted that such climatic shifts may lead to an increased frequency of explosive cyclones and a higher probability of hurricane formation in the region [1,2]. These environmental changes pose a significant threat to aquaculture, a sector reliant on stable marine conditions for the cultivation and production of aquatic species [3].
Globally, fish provide approximately 17% of animal protein and 7% of total protein consumed, supplying nearly 20% of the daily protein intake for over 3.2 billion people. The contribution of aquaculture to global fisheries has increased steadily, reaching 46.8% of total seafood production by 2016 [4]. The industry continues to expand at a rate surpassing that of other food production sectors. However, as aquaculture intensifies, traditional extensive farming systems are being replaced by high-density, intensive practices that increase the risk of disease outbreaks within cultures populations [5].
Beyond economic losses, extreme weather events such as storms can severely damage aquaculture infrastructure and lead to large-scale fish escapes. Such was the case with Storm Gloria in the winter of 2020, which caused significant structural damage to fish farms along the coasts of Valencia and Murcia, allowing large numbers of cultures fish to escape into the wild [6]. Escaped fish, which are typically bred in controlled environments and fed artificial diets, differ significantly from their wild counterparts in both behavior and physiology. Their introduction into natural ecosystems poses several ecological and public health risks, including competition with native fish for food and habitat, genetic pollution through interbreedin [5,7,8].
Izquierdo-Gomez et al. [9] categorize fish escapes into three levels: routine escapes (approximately 5,000 fish per year, or 1.31 tons annually), mass escape events (up to 91 times the normal level), and catastrophic events (up to 1,800 times the normal level). The escape of cultures fish can result in the establishment of self-sustaining populations, alter genetic diversity, introduce pathogens, and disrupt local food webs [10]. In the Mediterranean Sea, cultures fish escapes have been linked to genetic hybridization and competition with native species for resources [11]. These events can also impact commercial fisheries, as escaped fish often mix with wild stocks, complicating stock assessments and reducing market value. The ability to distinguish cultures fish from wild specimens based on morphological differences—such as a more rounded and compact body shape in cultures fish—can aid in assessing the impact of these escapes [12].
One of the main challenges in managing fish escapes is accurately identifying escaped individuals once they enter natural ecosystems. The ability to distinguish between wild, cultures, and escaped fish is critical for fisheries management, conservation efforts, and ensuring the authenticity of seafood products in markets.
The analysis of heavy metals and fatty acids has emerged as a valuable tool for fish traceability. Fish accumulate metals such as arsenic (As), selenium (Se), and mercury (Hg) from their environment and diet, and the concentrations of these elements can vary depending on whether the fish were raised in aquaculture facilities or in the wild [13,14]. Cultures fish are often exposed to different dietary formulations and water conditions compared to their wild counterparts, leading to distinctive metal accumulation patterns. For instance, cultures fish tend to have lower levels of Hg and higher levels of Zn and Cu due to the composition of commercial aquafeeds. Inductively Coupled Plasma Mass Spectrometry (ICP-MS) is one of the most advanced techniques for detecting and quantifying trace metals in fish tissue. This method offers high sensitivity and precision, allowing for the identification of even trace concentrations of elements. ICP-MS analysis is particularly valuable for seafood traceability, as differences in metal content can reflect variations in feeding regimes, aquaculture water sources, and environmental exposure [14,15].
Similarly, fatty acid profiles serve as robust biochemical markers for differentiating fish from different origins. Wild fish naturally acquire a diet rich in omega-3 fatty acids (e.g., eicosapentaenoic acid [EPA] and docosahexaenoic acid [DHA]), whereas cultures fish are typically fed diets that include plant-based oils, leading to higher levels of omega-6 fatty acids, particularly linoleic acid (18:2n-6). The presence of certain fatty acids, such as linoleic acid, in fish muscle tissue can thus be indicative of aquaculture origin [16]. The ratios of omega-3 to omega-6 fatty acids, along with specific lipid biomarkers, can effectively distinguish between wild fish, escaped fish, and those reared in aquaculture facilities [6]. Gas Chromatography-Mass Spectrometry (GC-MS) is widely used to analyze fatty acid profiles in fish muscle tissue. This technique involves the conversion of fatty acids into volatile methyl esters, which are then separated and quantified. GC-MS provides detailed insights into lipid composition, allowing for the identification of key fatty acids such as EPA, DHA, and linoleic acid. The ratio of omega-3 to omega-6 fatty acids serves as a reliable indicator of dietary origin, distinguishing between wild-caught and cultures fish [6].
Beyond heavy metal and fatty acid analysis, Metabolomics has emerged as a powerful approach for studying biochemical differences between wild, escaped, and cultures fish. Metabolomics involves the comprehensive analysis of small-molecule metabolites present in biological tissues, providing insights into metabolic pathways influenced by diet, environment, and physiological status.
Nuclear Magnetic Resonance (NMR) spectroscopy is a widely used analytical tool in metabolomics, allowing for the identification and quantification of both polar and apolar metabolites [17]. By applying NMR-based metabolomics, researchers can detect metabolic differences that arise due to variations in diet, environmental exposure, and physiological adaptation [18]. Cultures fish, for example, exhibit distinct metabolic signatures characterized by higher levels of creatine, creatinine, lactate, and alanine, reflecting the impact of artificial feeding regimes and controlled rearing conditions [6]. In contrast, wild fish demonstrate higher levels of taurine, an amino acid obtained from natural prey, and an increased abundance of omega-3 fatty acids.
The application of metabolomics to escaped fish is particularly relevant in assessing how their metabolic profiles change once they enter natural ecosystems. Escaped fish often experience abrupt shifts in diet and environmental conditions, leading to metabolic adaptations that can be detected through NMR-based metabolomic profiling. This approach provides a unique means of assessing fish traceability, complementing traditional chemical analyses [6].
The metabolomic signatures of wild and cultures fish differ significantly due to variations in diet and metabolic activity. Cultures fish typically exhibit higher levels of metabolites associated with artificial feeding regimes, such as: Creatine and creatinine, indicators of higher energy reserves due to regular feeding; lactate and alanine, metabolites linked to glycolysis and anaerobic metabolism, which are more prominent in cultures fish with lower physical activity; and glycine and betaine, amino acids commonly found in aquaculture feeds. In contrast, wild fish show higher levels of metabolites that reflect a more diverse and natural diet: TMAO, a key metabolite obtained from marine prey, found at significantly higher concentrations in wild fish; or omega-3 fatty acids (EPA, DHA), essential fatty acids derived from marine food sources, crucial for cell membrane integrity and metabolic function. In this line, the Lipidomics, a subset of metabolomics, focuses on the comprehensive analysis of lipid species, providing critical insights into dietary influences and metabolic adaptations. The fatty acid composition of fish muscle tissue is a particularly valuable marker for traceability. Wild fish exhibit a lipid profile rich in omega-3 fatty acids, with dominant peaks corresponding to EPA (20:5n-3) and DHA (22:6n-3), and cultures fish have a lipid profile characterized by higher levels of omega-6 fatty acids, particularly linoleic acid (18:2n-6), due to the inclusion of plant-based oils in aquafeeds [19]. Escaped fish could be present an intermediate profile, reflecting dietary changes as they transition from aquafeed consumption to foraging in the wild.
Given the increasing incidence of fish escapes and the need for reliable traceability methods, this study aims to integrate multiple analytical approaches—metals analysis, fatty acid profiling, and NMR-based metabolomics—to differentiate between wild, cultures, and escaped fish. The specific objectives include: Characterizing heavy metal concentrations in gilthead seabream, European seabass, and meagre to assess their potential as traceability markers; analyzing fatty acid profiles using GC-MS to distinguish fish origin based on lipid composition; applying NMR-based Metabolomics and Lipidomics to identify key biochemical biomarkers for fish traceability; and developing a classification model for determining the origin of fish with high accuracy.
This comprehensive approach will contribute to improved fisheries management, aquaculture sustainability, and the prevention of seafood fraud.
2. Materials and Methods
2.1. Specimen Collection and Sample Preparation
Fish samples, 30 gilthead seabream (Sparus aurata), 30 European seabass (Dicentrarchus labrax), and 20 meagre (Argyrosomus regius), were collected from fish markets, supermarkets, and local and wholesale markets in the Valencian Community and Murcia (Spain) between 2019 and 2022. Each specimen underwent an initial external examination to assess the presence of parasites, followed by biometric measurements and photographic documentation. Muscle tissue samples were subsequently extracted and stored at -20 °C until analysis. Based on their appearance, the presence of regenerated scales [9] and traceability references (‘commercial’ labelling), the fish were classified into the following origin groups: ‘Wild’, ‘Escape’ and ‘Cultured’.
2.2. Heavy Metals Analysis by ICP-MS
Metal profiles were obtained from the digestion of muscle samples and ICP-MS analysis. These analyses were carried out at the Technical Research Services (SSTTI) of the University of Alicante. Fifteen trace elements selected on the basis of their frequency, bioaccumulation capacity and potential detrimental effect on human health (Al, Cr, Mn, Fe, Co, Ni, Cu, Zn, As, Se, Mo, Cd, Tl, Pb, Hg) were quantified. For chemical analysis, muscle samples were taken from the same individuals used in the previous biochemical analyses, i.e., 10 specimens per species (Seabream, Seabass, Meagre) and group of origin (wild, escaped, cultured), except for wild seabass, for which sufficient specimens were unavailable during the escape event period.
The frozen samples were subjected to a digestion process in a solution of 4 ml of HNO3 and 0.5 ml of H2O2. The program consisted of different phases, reaching a final temperature of 240 ºC over 45 min. Afterwards, the resulting sample was diluted to 15 ml with milli-Q water. Aliquots were taken and processed by standard ICP-MS analysis [20], extracting the concentration of the different heavy metals listed previously.
2.3. Fatty Acid Analysis by GC-MS
The fatty acid profiles were constructed by extracting a battery of 35 of the most common fatty acids (Annex) by direct extraction of fatty acid methyl esters (FAME) [21]. The analysis performed by the external Technical Services of the Institute of Animal Science and Technology - UPV, provided a quantitative description of the different fatty acids, as well as a chromatogram corresponding to each individual.
In the experimental process, muscle samples from the specimens used for metabolomic analysis were used. The experimental process consisted of direct FAME synthesis [21]. In detail, the frozen samples were divided into 0.5 g subsamples which were ground at room temperature between 10 and 15 seconds. The derivative was placed in Pyrex tubes with: 1 ml of C13:0 standard (0.5 mg C13:0/ml MeOH); 0.7 ml of 10 N KOH in water; and 5.3 ml of MeOH. The tubes were incubated in a water bath at 55 °C for 1.5 h with 5 sec shaking every 20 min to dissolve and hydrolyze the sample. Subsequently, the samples were brought to room temperature by applying a cold bath and 0.58 ml of 24 N H2SO4 in water was added. The tubes were mixed by inversion and incubated in water at 55 °C for an additional 1.5 h, shaking them gently for 5 s every 20 min. After a second cooling, 3 ml of hexane were added and homogenized in vortex for 5 min. Subsequently, they were centrifuged for an additional 5 min. The hexane layers, containing the FAMEs, were transferred to gas chromatography vials, stored at -20 °C. Gas chromatographic (GC) analysis followed a standard protocol [21]. The GC-MS results were the concentrations of each fatty acid in mg per 100 g of sample.
2.4. 1H NMR Acquisition and Data Processing Parameters
A 500 µL sample was place in a 5 mm NMR tube, and spectra were referenced to TSP at 0.00 ppm (polar samples) or to chloroform at 7.26 ppm. All 1H NMR experiments were performed on a Bruker Avance 400 MHz equipped with a 5 mm HBB13C TBI probe with an actively shielded Z-gradient. The 1D solution state 1H NMR experiments had a 2 s recycle delay, 32,768 time-domain points and 2.556 s acquisition time. In total, 1024 scans were performed, and the experiment was conducted at 298ºK. Spectra were apodised through multiplication with an exponential decay, producing a 0.3 Hz line broadening in the transformed spectrum. The 1H NMR spectra were normalised and reduced to ASCII files using TopSpin (Bruker) and aligned using icoshift (version 1.0; available at www.models.kvl.dk) [6]. Processing of 1H NMR spectra was performed in MATLAB (MathWorks, Natick, MA, USA). The region of water (4.60–4.95 ppm) and extreme high and low fields (<0.5 ppm and 10 ppm, respectively) were removed. Metabolites were identified in one-dimensional spectra using The Human Metabolome Database (HMDB, https://hmdb.ca/) and the literature cited in this study [6].
2.5. Statistical Analysis
Data from metal and fatty acids quantification were analyzed using Nonclassical Multidimensional Scaling (nMDS), specifically employing the Sammon mapping algorithm to visualize sample relationships based on pairwise similarity matrices. This method reduces dimensionality while preserving topological distances, emphasizing subtle group differences.
Metabolomic data (polar and apolar fractions) were organized into feature matrices and analyzed using Partial Least Squares–Linear Discriminant Analysis (PLS-LDA) in MATLAB [22]. Pareto scaling was applied, and three components were used to build classification models. Model performance was evaluated through standard statistical parameters including R²X (explained variance in predictors), R²Y (explained variance in response), sensitivity, specificity, and area under the ROC curve (AUC).
3. Results and Discussion
3.1. Heavy Metals Analysis in Seabream, Seabass, and Meagre
The present study evaluated the fatty acid profiles and heavy metal concentrations as potential tracers for distinguishing escaped fish from wild and cultured counterparts. The findings reveal species-specific variations in metal accumulation and fatty acid composition, highlighting their potential use in traceability and food safety considerations.
The analysis of heavy metals in fish muscle tissue revealed significant differences between wild, escaped, and cultures fish (Figure 1, Figure 2 and Figure 3). Using Inductively Coupled Plasma Mass Spectrometry (ICP-MS), we quantified the concentrations of 15 trace elements, including Al, Cr, Mn, Fe, Co, Ni, Cu, Zn, As, Se, Mo, Cd, Tl, Pb, and Hg. Wild seabream exhibited higher levels of arsenic (As), selenium (Se), and mercury (Hg) compared to farm-raised fish. This is attributed to their natural diet, which includes benthic organisms and filter-feeding mollusks known for bioaccumulating arsenic. Notably, arsenic in wild seabream is primarily present in the form of arsenobetaine, which is considered non-toxic to humans [23]. The presence of arsenic in seafood has been widely studied due to its potential health risks, particularly when present in inorganic forms, which are highly toxic. However, marine organisms tend to accumulate organic arsenic forms, which have lower toxicity. Despite this, continuous monitoring is required to assess the potential long-term effects of arsenic accumulation in marine food chains.
The metabolism of arsenic in aquatic organisms is a complex process influenced by environmental and biological factors. Arsenic exists in both inorganic (iAs) and organic (oAs) forms, with arsenobetaine (AsB) being the predominant non-toxic form in marine fish [23]. The biotransformation of arsenic involves redox reactions and methylation processes, which are mediated by microbial and enzymatic activity [24]. In aquatic environments, arsenic can induce oxidative stress by generating reactive oxygen species (ROS), which can damage cellular components such as lipids, proteins, and DNA [25]. Antioxidant enzymes, including superoxide dismutase (SOD) and glutathione peroxidase (GPx), play a crucial role in mitigating oxidative stress, highlighting the intricate balance between arsenic toxicity and cellular defense mechanisms [26].
Selenium is an essential micronutrient that plays a crucial role in antioxidant defense, immune function, and thyroid hormone metabolism. Marine fish generally contain moderate to high selenium levels, which contribute to their nutritional value. Interestingly, selenium has been shown to counteract mercury toxicity by forming biologically inactive Se–Hg complexes, which reduce mercury’s bioavailability and toxicity [27]. The protective role of selenium is particularly relevant in wild seabream, where higher Se levels may mitigate the adverse effects of elevated mercury concentrations. However, excessive selenium intake can also be detrimental, leading to toxicity symptoms such as oxidative stress and metabolic disturbances. The balance between selenium and mercury is, therefore, an important factor in seafood safety and should be further investigated in future studies.
Mercury levels were also elevated in wild seabream, likely due to bioaccumulation through the food chain, given that seabream is a carnivorous species. The trophic transfer of mercury from prey to predator results in higher concentrations in top consumers, which is a concern for both ecosystem health and human consumption. The observed differences in metal concentrations suggest that metal profiles can serve as biomarkers to differentiate wild seabream from cultured counterparts.
In contrast, seabass did not show significant differences in heavy metal content between wild and cultured groups. The similarity in metal accumulation suggests that cultured seabass diets may closely resemble those of wild seabass, leading to a homogenization of metal profiles. This observation limits the utility of heavy metals as a distinguishing factor for seabass traceability. Additionally, seabass are more pelagic feeders compared to seabream, potentially leading to a different bioaccumulation dynamic that reduces metal differentiation.
For meagre, which included only cultured and escaped groups, escaped specimens exhibited higher concentrations of As, Se, and Hg. These findings suggest that escaped fish consume wild prey, leading to a bioaccumulative effect similar to that observed in wild seabream. The absence of wild meagre in this region of the Mediterranean prevents direct comparisons, but the observed differences highlight the potential of heavy metal analysis in tracing fish escapes.
3.2. Fatty Acid Profiles in Seabream, Seabass, and Meagre
The fatty acid analysis yielded patterns consistent with heavy metal findings. In seabream, distinct lipid biomarkers enabled differentiation between wild, cultured, and escaped groups. Wild seabream displayed higher levels of arachidonic acid (C20:4n6) and docosapentaenoic acid (DHA; C22:5n-3) compared to cultured individuals. These differences are linked to dietary variations, as wild seabream consume marine-derived lipids rich in long-chain polyunsaturated fatty acids (PUFAs), while cultured fish are fed diets containing vegetable oils [28,29].
Fatty acids play fundamental roles in fish metabolism, influencing membrane fluidity, signaling pathways, and energy storage. DHA, for instance, is critical for neural development, vision, and immune function in fish. It is also a key component of phospholipids in cell membranes, enhancing membrane stability and function. Arachidonic acid (ARA) serves as a precursor for eicosanoids, which regulate inflammatory responses, immunity, and stress adaptation. Wild seabream’s elevated DHA and ARA levels reflect their natural diet, which provides a more balanced and diverse lipid profile compared to cultured fish [30,31,32].
Conversely, cultured seabream exhibited elevated levels of linoleic acid (C18:2n6c) and linolenic acid (C18:3n3), consistent with the use of terrestrial plant-derived feed ingredients [30]. While these fatty acids can be elongated and desaturated to form long-chain PUFAs, their conversion efficiency in marine fish is relatively low. The higher prevalence of plant-derived fatty acids in cultured fish has implications for nutritional quality, as these lipids may alter the functional properties of cellular membranes and reduce the availability of essential fatty acids for physiological processes [33].
Seabass and meagre did not show significant differences in fatty acid profiles between wild and cultured individuals, aligning with the findings from heavy metal analysis. The lack of differentiation suggests that fatty acid composition alone may not serve as a reliable biomarker for traceability in these species. However, in seabream, normalization of fatty acid values improved classification, supporting the use of lipidomics as a tool for distinguishing fish origin (Figure 4, Figure 5 and Figure 6).
3.3. Implications for Food Safety and Traceability
The findings of this study have important implications for seafood safety and traceability. The concentrations of heavy metals in all fish samples remained below regulatory safety limits, ensuring that consumption poses no immediate health risks. However, the bioaccumulation of arsenic and mercury in wild and escaped fish raises concerns about long-term exposure effects, particularly for consumers with high seafood intake. The selenium-to-mercury molar ratio observed in wild seabream suggests a mitigating effect on mercury toxicity, but further research is needed to determine the threshold for safe consumption [34].
From a traceability perspective, seabream demonstrated the greatest potential for differentiation based on both heavy metal and fatty acid profiles. The clear distinction between wild and cultured individuals highlights the feasibility of using these biomarkers in monitoring programs for escaped fish. In contrast, seabass and meagre require additional molecular or isotopic analyses to improve classification accuracy. The integration of fatty acid profiling with stable isotope analysis may enhance differentiation by providing insights into trophic interactions and feeding histories.
The data for metals and AAGGS analyses were analyzed using the MDS method. Variations in the concentrations of metals and AAGGs in the different types of fish (cultured, escaped or wild) can be visualized by Sammon mapping (Figure 7). Sammon mapping was used as this method diminishes the influence of large distances, which can completely dominate the map. The negative part of the x-axis in the Sammon mapping for metals was dominated by cultured and escaped fish samples, with wild fish samples dominating the positive part, due to the higher concentration in As, Se and Hg (Figure 1, Figure 2 and Figure 3). These samples are from sea bream. Sea bream are fish that consume algae that accumulate these metals. The wild sea bass samples, since they do not have these differences in these metals, are placed next to the cultured and escaped fish. For Sammon's map for AAGGs, we have a similar distribution, but here, on the negative side we do not have the wild sea bass samples, which are grouped with the wild sea bream samples. The AAGGs profiles are very similar in wild fish for both sea bream and sea bass (Figure 4, Figure 5 and Figure 6).
3.4. Metabolomic and Lipidomic Profiling Using NMR
This study aimed to evaluate the utility of metabolomic and lipidomic profiling in distinguishing between cultured, escaped, and wild fish specimens, specifically focusing on seabass (Dicentrarchus labrax) and meagre (Argyrosomus regius). The metabolomic and lipidomic analyses revealed significant differences between groups, with particular emphasis on the role of trimethylamine N-oxide (TMAO) as a key biomarker distinguishing wild fish from cultured and escaped fish [6,19,35,36,37].
The NMR-based metabolomic analysis of the polar fraction from seabass muscle revealed a clear separation between cultured and wild specimens, while escaped fish exhibited intermediate characteristics (Figure 8). The multivariate PLS-LDA analysis effectively grouped wild fish apart from the cultured and escaped specimens were made for seabream [6]. We have used the build model for sea bream and tried to classify sea bass and sea bass specimens. We obtained an excellent classification of the samples (Figure 9), suggesting a good tool for traceability.
A key finding was the significantly higher levels of TMAO in wild seabass compared to cultured and escaped specimens. TMAO plays a critical role in osmoregulation in marine fish, stabilizing proteins against denaturation and oxidative stress in varying salinity conditions [38,39]. Wild fish, exposed to fluctuating salinities, accumulate higher levels of TMAO, whereas cultured fish, kept under stable conditions, exhibit lower levels. Escaped fish had intermediate levels, suggesting partial metabolic adaptation but not full alignment with wild conditions. These findings are consistent with recent studies on gilthead seabream (Sparus aurata), which also reported significantly elevated TMAO in wild fish, indicating its role as a robust metabolic marker of environmental adaptation [37,40,41,42,43].
TMAO serves multiple physiological functions beyond osmoregulation. It has been identified as a key metabolite for stabilizing proteins and membranes against environmental stressors such as temperature fluctuations and oxidative stress. Studies indicate that TMAO enhances mitochondrial efficiency, contributing to improved energy metabolism in wild fish [41]. Additionally, wild fish tend to have a more diverse gut microbiota, which enhances TMAO biosynthesis. In contrast, cultured fish, often exposed to antibiotic treatments, exhibit reduced microbial diversity, limiting their ability to produce TMAO [38].
In aquaculture, lower TMAO levels might indicate reduced adaptation to natural environmental conditions and could be linked to inferior muscle quality. Furthermore, TMAO levels in wild fish have been correlated with increased muscle firmness, a key quality attribute for seafood consumers [40].
For meagre, a similar pattern emerged, with TMAO serving as the primary distinguishing metabolite. The intermediate TMAO levels in escaped meagre reinforce the idea that metabolic adaptation to the wild is an ongoing, time-dependent process [40].
Furthermore, cultured fish showed elevated levels of betaine, creatine, and glucose, reflecting metabolic adaptations associated with captivity. These metabolites, involved in energy storage and anaerobic metabolism, highlight the restricted mobility and high-energy diets typical of aquaculture environments [44,45,46]. The higher lactate levels in escaped fish suggest increased muscular activity, supporting the hypothesis of a metabolic shift following escape.
Lipid profiling using ¹H NMR on the apolar fraction further confirmed dietary influences on metabolic composition. Cultured fish exhibited significantly higher levels of linoleic acid (C18:2n6) and α-linolenic acid (C18:3n3), characteristic of vegetable oils used in aquafeeds. These fatty acids were absent in wild fish, underscoring diet as a key determinant of lipid composition (Figure 10). Wild fish showed higher concentrations of arachidonic acid (C20:4n6) and docosahexaenoic acid (DHA, C22:6n-3), derived from marine prey. These long-chain polyunsaturated fatty acids (LC-PUFAs) are essential for cell membrane function, neural development, and immune responses [47]. Their enrichment in wild fish reflects their natural diet, rich in marine-derived lipids, in contrast to cultured fish, which receive diets supplemented with plant oils.
The distinct triplet signal at 2.79 ppm, corresponding to di-unsaturated fatty acids (DUFAs), was exclusively found in cultured and escaped fish (Figure 10). This signal was linked to linoleic acid, confirming its role as a dietary biomarker for aquaculture feeds. Adjusting the classification model to include this biomarker improved the differentiation of wild fish, validating its potential use in traceability applications [6].
The fatty acid composition in fish muscle reflects not only dietary influences but also physiological adaptations to energy metabolism, immune function, and stress resilience. DHA, for instance, is a crucial component of neuronal and retinal membranes, playing a fundamental role in cognitive function and vision in fish [48,49]. Meanwhile, arachidonic acid (ARA) acts as a precursor for eicosanoids, which modulate inflammatory responses and immune defense.
Fish raised in aquaculture systems often have lower DHA and ARA levels due to their plant-based diets, which lack these essential lipids. This may have implications for immune competence, as studies have shown that fish with higher dietary ARA intake exhibit enhanced stress tolerance and disease resistance.
The PLS-LDA model created to classify the seabream specimens into farmed, escaped and wild according to the 1H NMR spectra of the apolar metabolites [6], was used to classify the spectra obtained from the seabass and meagre samples (Figure 9). In this case, as with the spectra obtained from the 1H NMR analysis of the polar metabolites, we also obtained an excellent separation and classification of the samples (Figure 11).
4. Conclusions
This study presents a multidisciplinary approach to trace the origin of fish through the integration of heavy metal analysis, fatty acid profiling, and 1H NMR-based metabolomics and lipidomics. By analyzing three economically relevant Mediterranean species—Sparus aurata (gilthead seabream), Dicentrarchus labrax (European seabass), and Argyrosomus regius (meagre)—we demonstrate that biochemical signatures can effectively distinguish between wild, farmed, and escaped individuals.
Our findings highlight the following key insights:
Heavy metals, particularly arsenic (As), selenium (Se), and mercury (Hg), were significantly elevated in wild and escaped gilthead seabream, supporting their potential as traceability biomarkers. These differences are linked to natural feeding habits and environmental exposure.
Fatty acid composition revealed species-specific patterns, with wild seabream showing higher levels of DHA and ARA, and cultured fish presenting elevated linoleic acid. These differences reflect dietary sources and feeding regimes, though they were less pronounced in seabass and meagre.
Metabolomic analysis, especially the quantification of TMAO, creatine, and betaine, provided robust discrimination between groups. TMAO emerged as a key marker of environmental adaptation and was consistently higher in wild fish. Escaped specimens displayed intermediate metabolic profiles, reflecting partial adaptation to natural conditions.
Lipidomic profiling confirmed the influence of aquaculture diets on muscle lipid composition. The presence of linoleic acid and other plant-derived fatty acids in cultured fish, and their absence in wild individuals, further reinforced dietary traceability.
The application of multivariate statistical models (MDS, PLS-LDA) enabled accurate classification of fish origin, particularly in gilthead seabream. These models may be adapted for broader use in seafood authentication programs.
Overall, this work demonstrates that combining metal, lipid, and metabolite analyses offers a reliable and integrative strategy for fish traceability. Such tools are essential for strengthening seafood authentication, detecting aquaculture escape events, supporting sustainable fisheries management, and protecting consumer trust.
Future perspectives should explore the integration of compound-specific isotope analysis, microbiota profiling, and long-term studies to enhance traceability systems. The approach presented here lays the groundwork for the development of molecular traceability platforms applicable to both regulatory and commercial contexts.
These findings contribute to improving seafood traceability, ensuring food safety, and supporting sustainable fisheries management.
Author Contributions
Funding obtained: KT-G. Conceived and designed the experiments: FCM-E and KT-G. Collected fish samples: KT-G, JMV-R, and AV-M Performed the experiments: WB, KT-G, JMV-R, AV-M, and FCM-E. Analyzed the data: WB, KT-G, JMV-R, AV-M, and FCM-E. Wrote the manuscript: FCM-E, WB and KT-G. Reviewed the manuscript: FCM-E, WB, KT-G, JMV-R, and AV-M. All authors have read and agreed to the published version of the manuscript.
Funding
KT-G was funded by a postdoctoral tenure program Juan de la Cierva Formación (FJCI-2014-20100) and Juan de la Cierva Incorporación (IJCI-2017-34174) funded by the Spanish National Research Agency. This study was funded by the Project ‘GLObal change Resilience in Aquaculture-2 (GLORiA2)’, supported by the Biodiversity Foundation of the Spanish Ministry for the Ecological Transition and Demographic Challenge, through the Pleamar Program and co-financed by the European Maritime and Fisheries Fund. The research is also part of the LIFE IP INTEMARES Project ‘Integrated, innovative and participatory management of the Natura 2000 Network in the Spanish marine environment’. This study forms part of the ThinkInAzul programme and was supported by MCIN with funding from European Union NextGenerationEU (PRTR-C17.I1) and by Generalitat Valenciana (THINKINAZUL/2021/044-TOWARDS). WB thanks the Government of Argelia for the award of a doctoral fellowship to support her studies at the University of Alicante.
Acknowledgments
The authors wish to thank Emilio Lorenzo, PhD (SSTTI of University of Alicante) for technical assistance with NMR analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three letter acronym |
| LD | Linear dichroism |
References
- Makris, C. V., K. Tolika, V. N. Baltikas, K. Velikou and Y. N. Krestenitis. "The impact of climate change on the storm surges of the mediterranean sea: Coastal sea level responses to deep depression atmospheric systems." Ocean Modelling 181 (2023): 102149. https://www.sciencedirect.com/science/article/pii/S1463500322001639. [CrossRef]
- Lionello, P., I. F. Trigo, V. Gil, M. L. R. Liberato, K. M. Nissen, J. G. Pinto, C. C. Raible, M. Reale, A. Tanzarella, R. M. Trigo, et al. "Objective climatology of cyclones in the mediterranean region: A consensus view among methods with different system identification and tracking criteria." Tellus A: Dynamic Meteorology and Oceanography (2016). [CrossRef]
- Toledo-Guedes, K., J. Atalah, D. Izquierdo-Gomez, D. Fernandez-Jover, I. Uglem, P. Sanchez-Jerez, P. Arechavala-Lopez and T. Dempster. "Domesticating the wild through escapees of two iconic mediterranean farmed fish species." Scientific Reports 14 (2024): 23772. [CrossRef]
- FAO. The state of world fisheries and aquaculture (sofia). Rome, Italy FAO, 2022, #266 p.
- Arechavala-Lopez, P., K. Toledo-Guedes, D. Izquierdo-Gomez, T. Segvic-Bubic and P. Sanchez-Jerez. "Implications of sea bream and sea bass escapes for sustainable aquaculture management: A review of interactions, risks and consequences." Reviews in Fisheries Science & Aquaculture 26 (2018): 214-34. <Go to ISI>://WOS:000428059800006. [CrossRef]
- Badaoui, W., F. C. Marhuenda-Egea, J. M. Valero-Rodriguez, P. Sanchez-Jerez, P. Arechavala-Lopez and K. Toledo-Guedes. "Metabolomic and lipidomic tools for tracing fish escapes from aquaculture facilities." ACS Food Science & Technology 4 (2024): 871-79. [CrossRef]
- Arechavala-Lopez, P., J. M. Valero-Rodriguez, J. Penalver-Garcia, D. Izquierdo-Gomez and P. Sanchez-Jerez. "Linking coastal aquaculture of meagre argyrosomus regius and western mediterranean coastal fisheries through escapes incidents." Fisheries Management and Ecology 22 (2015): 317-25<Go to ISI>://WOS:000358453400006. [CrossRef]
- Jackson, D., A. Drumm, S. McEvoy, O. Jensen, D. Mendiola, G. Gabina, J. A. Borg, N. Papageorgiou, I. Karakassis and K. D. Black. "A pan-european valuation of the extent, causes and cost of escape events from sea cage fish farming." Aquaculture 436 (2015): 21-26. <Go to ISI>://WOS:000346088200005. [CrossRef]
- Izquierdo-Gomez, D., P. Arechavala-Lopez, J. T. Bayle-Sempere and P. Sanchez-Jerez. "Assessing the influence of gilthead sea bream escapees in landings of mediterranean fisheries through a scale-based methodology." Fisheries Management and Ecology 24 (2017): 62-72. <Go to ISI>://WOS:000394962400006. [CrossRef]
- Naylor, R., S. Fang and J. Fanzo. "A global view of aquaculture policy." Food Policy 116 (2023): 102422. https://www.sciencedirect.com/science/article/pii/S0306919223000209. [CrossRef]
- Šegvić-Bubić, T., I. Lepen, Ž. Trumbić, J. Ljubković, D. Sutlović, S. Matić-Skoko, L. Grubišić, B. Glamuzina and I. Mladineo. "Population genetic structure of reared and wild gilthead sea bream (sparus aurata) in the adriatic sea inferred with microsatellite loci." Aquaculture 318 (2011): 309-15. https://www.sciencedirect.com/science/article/pii/S004484861100487X. [CrossRef]
- Jiménez-Segura, L. F., D. Restrepo-Santamaria, J. G. Ospina-Pabón, M. C. Castellanos-Mejía, D. Valencia-Rodríguez, A. F. Galeano-Moreno, J. L. Londoño-López, J. Herrera-Pérez, V. M. Medina-Ríos, J. Álvarez-Bustamante, et al. "Fish databases for improving their conservation in colombia." Scientific Data 12 (2025): 262. [CrossRef]
- Di Bella, C., A. Calagna, G. Cammilleri, P. Schembri, D. Lo Monaco, V. Ciprì, L. Battaglia, G. Barbera, V. Ferrantelli, S. Sadok, et al. "Risk assessment of cadmium, lead, and mercury on human health in relation to the consumption of farmed sea bass in italy: A meta-analytical approach." Frontiers in Marine Science 8 (2021): https://www.frontiersin.org/journals/marine-science/articles/10.3389/fmars.2021.616488.
- Lounas, R., H. Kasmi, S. Chernai, N. Amarni, L. Ghebriout and B. Hamdi. "Heavy metal concentrations in wild and farmed gilthead sea bream from southern mediterranean sea-human health risk assessment." Environ Sci Pollut Res Int 28 (2021): 30732-42. [CrossRef]
- Salem, A. M. and M. E. A. El-Metwally. "Bioaccumulation and health risk potential of heavy metals including cancer in wild and farmed meagre argyrosomus regius (asso, 1801), mediterreanean sea, egypt." Egyptian Journal of Aquatic Biology and Fisheries 28 (2024): 1277-301. https://ejabf.journals.ekb.eg/article_374235.html. [CrossRef]
- Lenas, D., D. Triantafillou, S. Chatziantoniou and C. Nathanailides. "Fatty acid profile of wild and farmed gilthead sea bream ( sparus aurata )." Journal für Verbraucherschutz und Lebensmittelsicherheit 6 (2011). [CrossRef]
- Lindon, J. C., J. K. Nicholson and E. Holmes. The handbook of metabonomics and metabolomics. Elsevier Science, 2011.
- Southam, A., A. Lange, R. Al-Salhi, E. Hill, C. Tyler and M. Viant. "Distinguishing between the metabolome and xenobiotic exposome in environmental field samples analysed by direct-infusion mass spectrometry based metabolomics and lipidomics." Metabolomics 10 (2014): 1050-58. [CrossRef]
- Marhuenda-Egea, F. C. and P. Sanchez-Jerez. Metabolomic insights into wild and farmed gilthead seabream (sparus aurata): Lipid composition, freshness indicators, and environmental adaptations. 30. 2025.
- Griboff, J., D. A. Wunderlin and M. V. Monferran. "Metals, as and se determination by inductively coupled plasma-mass spectrometry (icp-ms) in edible fish collected from three eutrophic reservoirs. Their consumption represents a risk for human health?" Microchemical Journal 130 (2017): 236-44. https://www.sciencedirect.com/science/article/pii/S0026265X16302016. [CrossRef]
- O'Fallon, J. V., J. R. Busboom, M. L. Nelson and C. T. Gaskins. "A direct method for fatty acid methyl ester synthesis: Application to wet meat tissues, oils, and feedstuffs." J Anim Sci 85 (2007): 1511-21. [CrossRef]
- Li, H. D., Q. S. Xu and Y. Z. Liang. "Libpls: An integrated library for partial least squares regression and linear discriminant analysis." Chemometrics and Intelligent Laboratory Systems 176 (2018): 34-43<Go to ISI>://WOS:000431936700004. [CrossRef]
- Zhang, W., A.-J. Miao, N.-X. Wang, C. Li, J. Sha, J. Jia, D. S. Alessi, B. Yan and Y. S. Ok. "Arsenic bioaccumulation and biotransformation in aquatic organisms." Environment International 163 (2022): 107221. https://www.sciencedirect.com/science/article/pii/S0160412022001477. [CrossRef]
- Sharma, V. K. and M. Sohn. "Aquatic arsenic: Toxicity, speciation, transformations, and remediation." Environment International 35 (2009): 743-59. https://www.sciencedirect.com/science/article/pii/S0160412009000051. [CrossRef]
- Valko, M., H. Morris and M. T. D. Cronin. "Metals, toxicity and oxidative stress." Current Medicinal Chemistry 12 (2005): 1161-208. http://www.eurekaselect.com/article/5248. [CrossRef]
- Byeon, E., H.-M. Kang, C. Yoon and J.-S. Lee. "Toxicity mechanisms of arsenic compounds in aquatic organisms." Aquatic Toxicology 237 (2021): 105901. https://www.sciencedirect.com/science/article/pii/S0166445X21001600. [CrossRef]
- Burger, J. and M. Gochfeld. "Selenium and mercury molar ratios in saltwater fish from new jersey: Individual and species variability complicate use in human health fish consumption advisories." Environmental Research 114 (2012): 12-23. https://www.sciencedirect.com/science/article/pii/S0013935112000679. [CrossRef]
- Benedito-Palos, L., J. C. Navarro, A. Sitjà-Bobadilla, J. Gordon Bell, S. Kaushik and J. Pérez-Sánchez. "High levels of vegetable oils in plant protein-rich diets fed to gilthead sea bream (sparus aurata l.): Growth performance, muscle fatty acid profiles and histological alterations of target tissues." British Journal of Nutrition 100 (2008): 992-1003. https://www.cambridge.org/core/product/AFBDB03ACA2AD6AA3B66016BD4BE5651. [CrossRef]
- Robin, J. H. and B. Vincent. "Microparticulate diets as first food for gilthead sea bream larva (sparus aurata): Study of fatty acid incorporation." Aquaculture 225 (2003): 463-74. https://www.sciencedirect.com/science/article/pii/S0044848603003107. [CrossRef]
- Xu, H., Q. Ai, K. Mai, W. Xu, J. Wang, H. Ma, W. Zhang, X. Wang and Z. Liufu. "Effects of dietary arachidonic acid on growth performance, survival, immune response and tissue fatty acid composition of juvenile japanese seabass, lateolabrax japonicus." Aquaculture 307 (2010): 75-82. https://www.sciencedirect.com/science/article/pii/S0044848610003960. [CrossRef]
- Calder, P. C. "The relationship between the fatty acid composition of immune cells and their function." Prostaglandins, Leukotrienes and Essential Fatty Acids 79 (2008): 101-08. https://www.sciencedirect.com/science/article/pii/S0952327808001361. [CrossRef]
- Furne, M., E. Holen, P. Araujo, K. K. Lie and M. Moren. "Cytokine gene expression and prostaglandin production in head kidney leukocytes isolated from atlantic cod (gadus morhua) added different levels of arachidonic acid and eicosapentaenoic acid." Fish & Shellfish Immunology 34 (2013): 770-77. https://www.sciencedirect.com/science/article/pii/S1050464812004470. [CrossRef]
- Hardy, R. W. and C.-s. Lee. "Aquaculture feed and seafood quality." Bulletin of Fisheries Research and Development Agency 31 (2010): 43-50.
- Ralston, N. V. C. "Selenium health benefit values as seafood safety criteria." EcoHealth 5 (2008): 442-55. [CrossRef]
- Melis, R., R. Sanna, A. Braca, E. Bonaglini, R. Cappuccinelli, H. Slawski, T. Roggio, S. Uzzau and R. Anedda. "Molecular details on gilthead sea bream (sparus aurata) sensitivity to low water temperatures from 1h nmr metabolomics." Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 204 (2017): 129-36. https://www.sciencedirect.com/science/article/pii/S1095643316302574. [CrossRef]
- Martinez, I., T. Bathen, I. Standal, J. Halvorsen, M. Aursand, I. Gribbestad and D. Axelson. "Bioactive compounds in cod (gadus morhua) products and suitability of h-1 nmr metabolite profiling for classification of the products using multivariate data analyses." Journal of Agricultural and Food Chemistry 53 (2005): 6889-95. [CrossRef]
- Yancey, P. H., A. L. Samerotte, G. L. Brand and J. C. Drazen. "Increasing contents of tmao with depth within species as well as among species of teleost fish." Integrative and Comparative Biology 44 (2004): 669-69<Go to ISI>://WOS:000226721401179.
- Solé-Jiménez, P., F. Naya-Català, M. C. Piazzon, I. Estensoro, J. À. Calduch-Giner, A. Sitjà-Bobadilla, D. Van Mullem and J. Pérez-Sánchez. "Reshaping of gut microbiota in gilthead sea bream fed microbial and processed animal proteins as the main dietary protein source." Frontiers in Marine Science 8 (2021): https://www.frontiersin.org/articles/10.3389/fmars.2021.705041. [CrossRef]
- Quero, G. M., R. Piredda, M. Basili, G. Maricchiolo, S. Mirto, E. Manini, A. M. Seyfarth, M. Candela and G. M. Luna. "Host-associated and environmental microbiomes in an open-sea mediterranean gilthead sea bream fish farm." Microbial Ecology 86 (2023): 1319-30. [CrossRef]
- Kunkel-Patterson, A., J. C. Fiess, L. Mathias, A. Samerotte, T. Hirano and P. H. Yancey. "Effects of salinity and temperature on osmolyte composition in tilapia." Integrative and Comparative Biology 45 (2005): 1156-56<Go to ISI>://WOS:000235337601284.
- Samerotte, A. L., J. C. Drazen, G. L. Brand, B. A. Seibel and P. H. Yancey. "Correlation of trimethylamine oxide and habitat depth within and among species of teleost fish: An analysis of causation." Physiological and Biochemical Zoology 80 (2007): 197-208<Go to ISI>://WOS:000243966400004. [CrossRef]
- Treberg, J. R. and W. R. Driedzic. "Maintenance and accumulation of trimethylamine oxide by winter skate (leucoraja ocellata):: Reliance on low whole animal losses rather than synthesis." American Journal of Physiology-Regulatory Integrative and Comparative Physiology 291 (2006): R1790-R98<Go to ISI>://WOS:000241768400027. [CrossRef]
- Yancey, P. H. "Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses." Journal of Experimental Biology 208 (2005): 2819-30. 10.1242/jeb.01730. [CrossRef]
- Schrama, D., M. Cerqueira, C. S. Raposo, A. M. Rosa da Costa, T. Wulff, A. Gonçalves, C. Camacho, R. Colen, F. Fonseca and P. M. Rodrigues. "Dietary creatine supplementation in gilthead seabream (sparus aurata): Comparative proteomics analysis on fish allergens, muscle quality, and liver." Frontiers in Physiology 9 (2018): https://www.frontiersin.org/journals/physiology/articles/10.3389/fphys.2018.01844. [CrossRef]
- Teles, A., L. Guzmán-Villanueva, M. A. Hernández-de Dios, M. Maldonado-García and D. Tovar-Ramírez. "Taurine enhances antioxidant enzyme activity and immune response in seriola rivoliana juveniles after lipopolysaccharide injection." (2024):.
- Sampath, W. W. H. A., R. M. D. S. Rathnayake, M. Yang, W. Zhang and K. Mai. "Roles of dietary taurine in fish nutrition." Marine Life Science & Technology 2 (2020): 360-75<Go to ISI>://WOS:000649454300005. [CrossRef]
- Houston, S. J. S., V. Karalazos, J. Tinsley, M. B. Betancor, S. A. M. Martin, D. R. Tocher and O. Monroig. "The compositional and metabolic responses of gilthead seabream (sparus aurata) to a gradient of dietary fish oil and associated n-3 long-chain pufa content." British Journal of Nutrition 118 (2017): https://www.cambridge.org/core/product/84301CB4F616131CACB2C2E07141C1C1. [CrossRef]
- Stubhaug, I., D. R. Tocher, J. G. Bell, J. R. Dick and B. E. Torstensen. "Fatty acid metabolism in atlantic salmon (salmo salar l.) hepatocytes and influence of dietary vegetable oil." Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 1734 (2005): 277-88. https://www.sciencedirect.com/science/article/pii/S1388198105000880. [CrossRef]
- Izquierdo, M. S., A. Obach, L. Arantzamendi, D. Montero, L. Robaina and G. Rosenlund. "Dietary lipid sources for seabream and seabass: Growth performance, tissue composition and flesh quality." Aquaculture Nutrition 9 (2003): 397-407. [CrossRef]
Figure 1.
Boxplots of the concentrations of different metals (As, Se and Hg) determined in meagre.
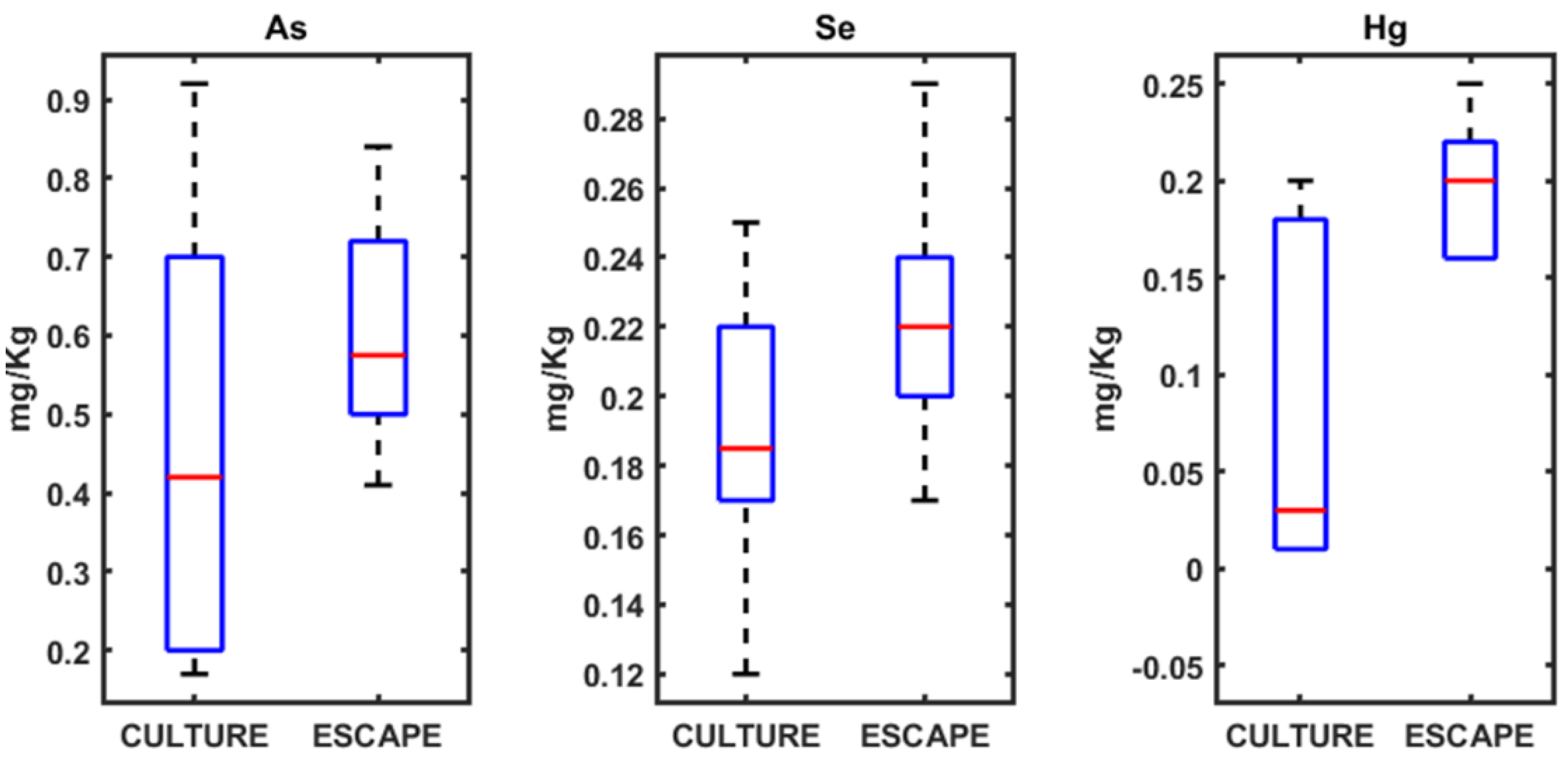
Figure 2.
Boxplots of the concentrations of different metals (As, Se and Hg) determined in seabream.
Figure 2.
Boxplots of the concentrations of different metals (As, Se and Hg) determined in seabream.
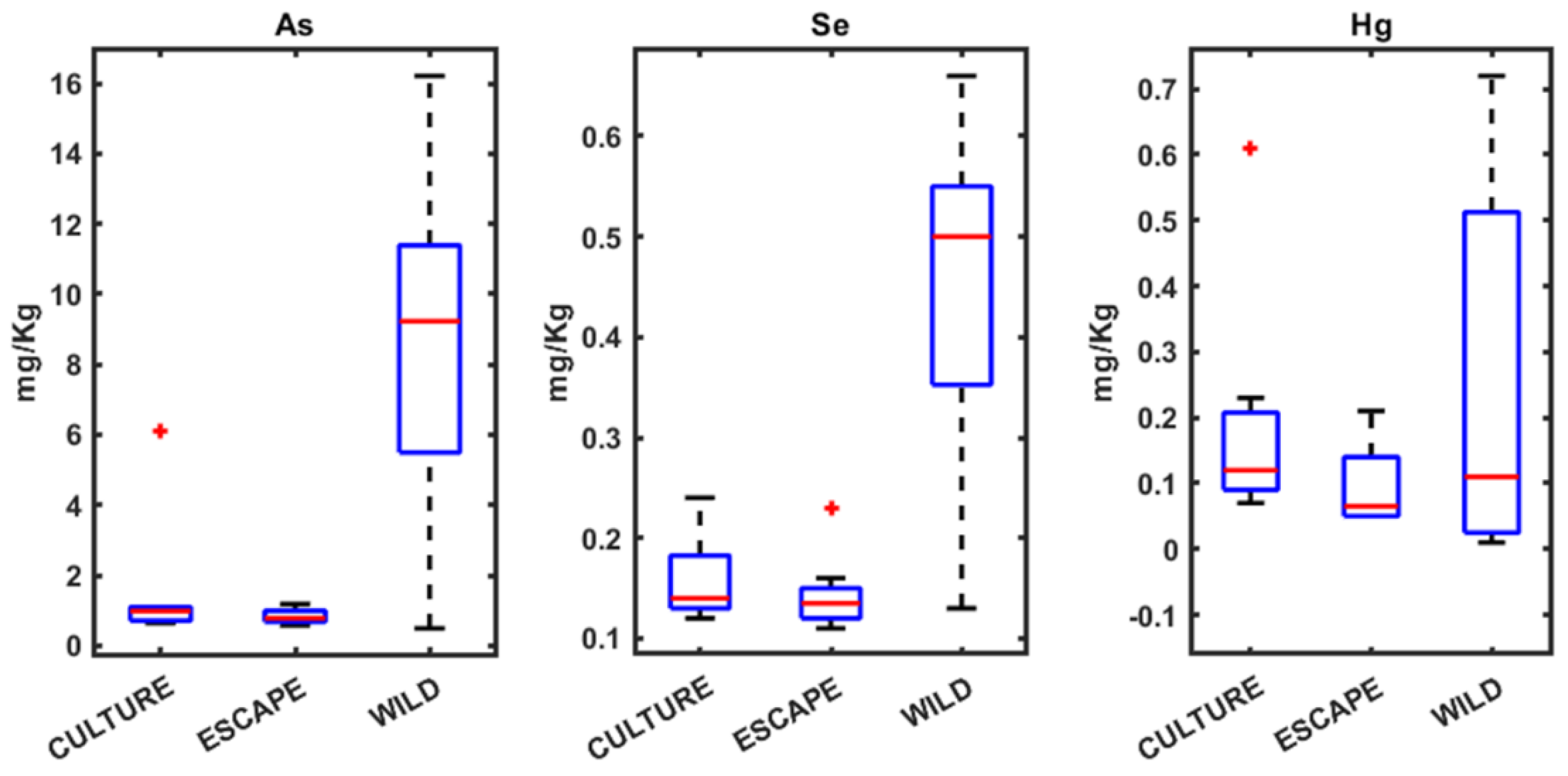
Figure 3.
Boxplots of the concentrations of different metals (As, Se and Hg) determined in seabass.
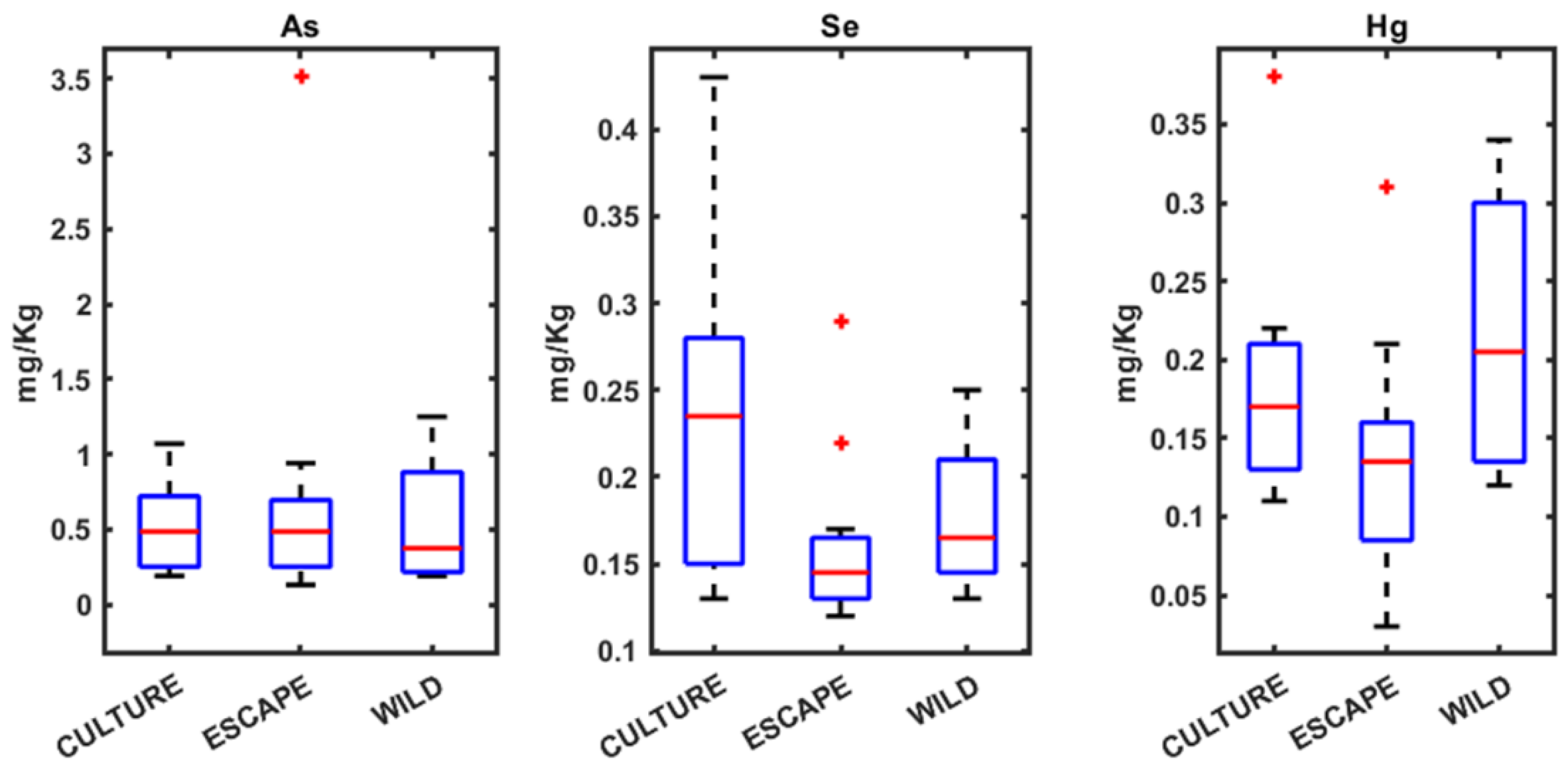
Figure 4.
Boxplots of the concentrations of different AAGGs determined in meagre.
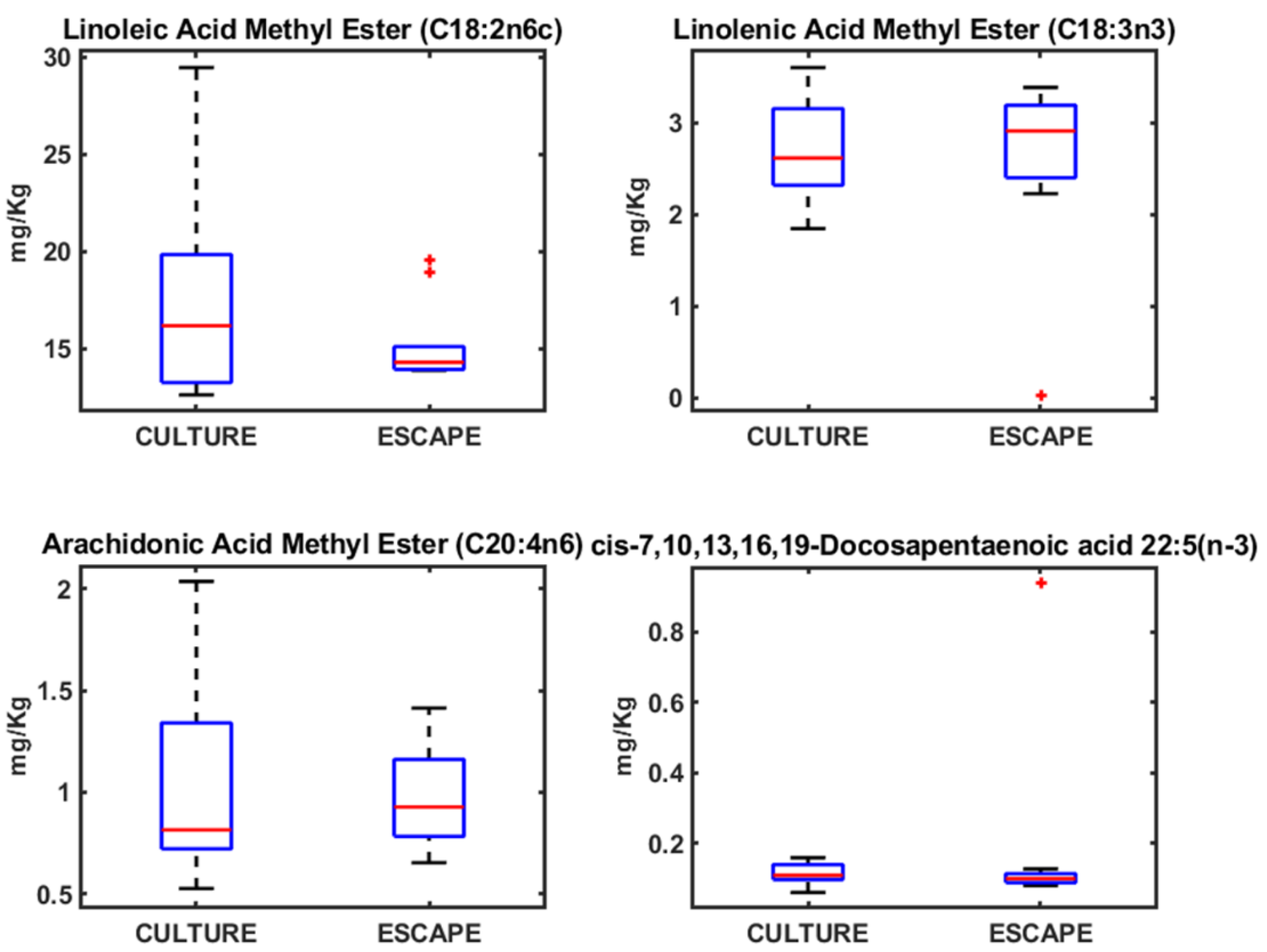
Figure 5.
Boxplots of the concentrations of different AAGGs determined in seabream.
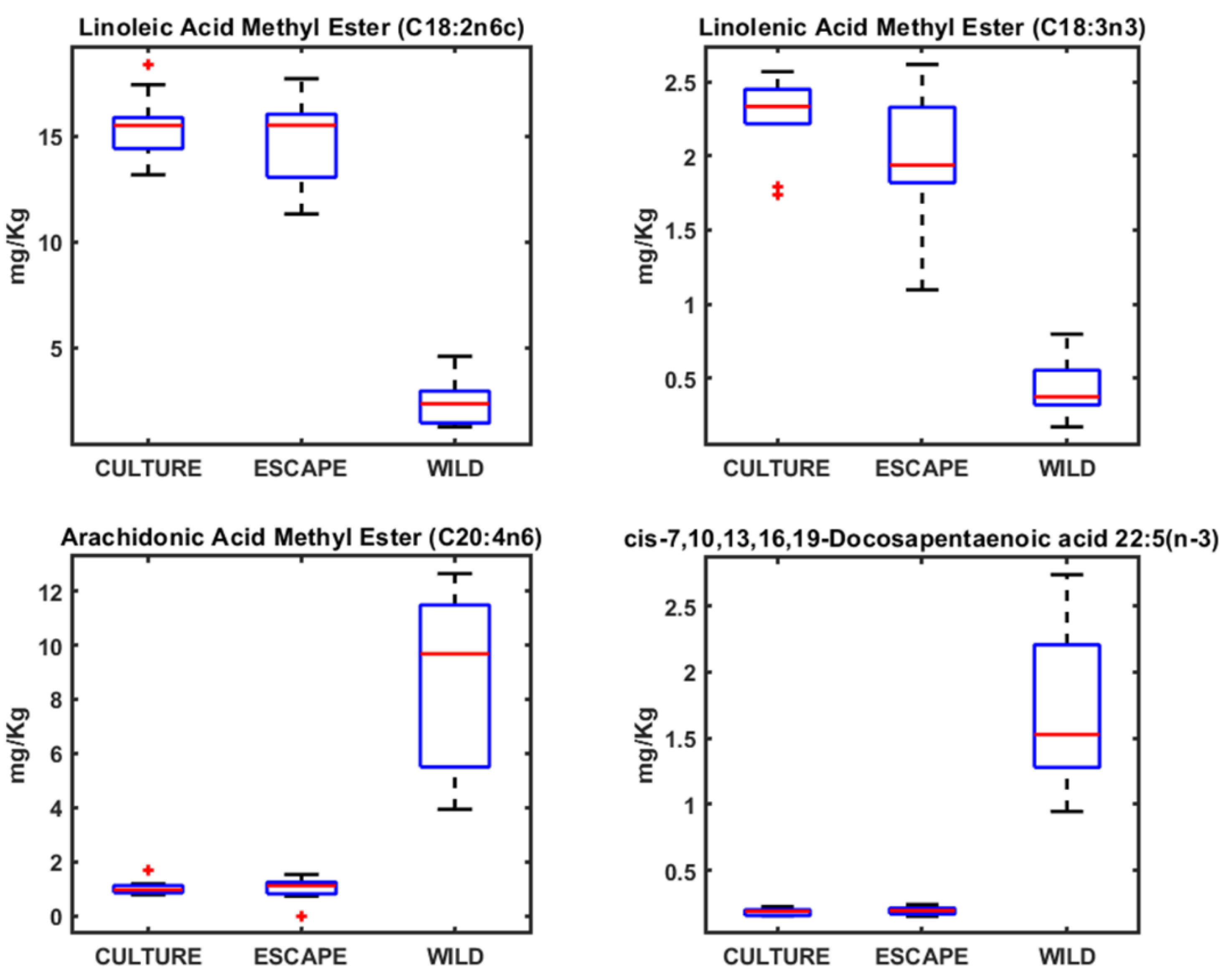
Figure 6.
Boxplots of the concentrations of different AAGGs determined in seabass.
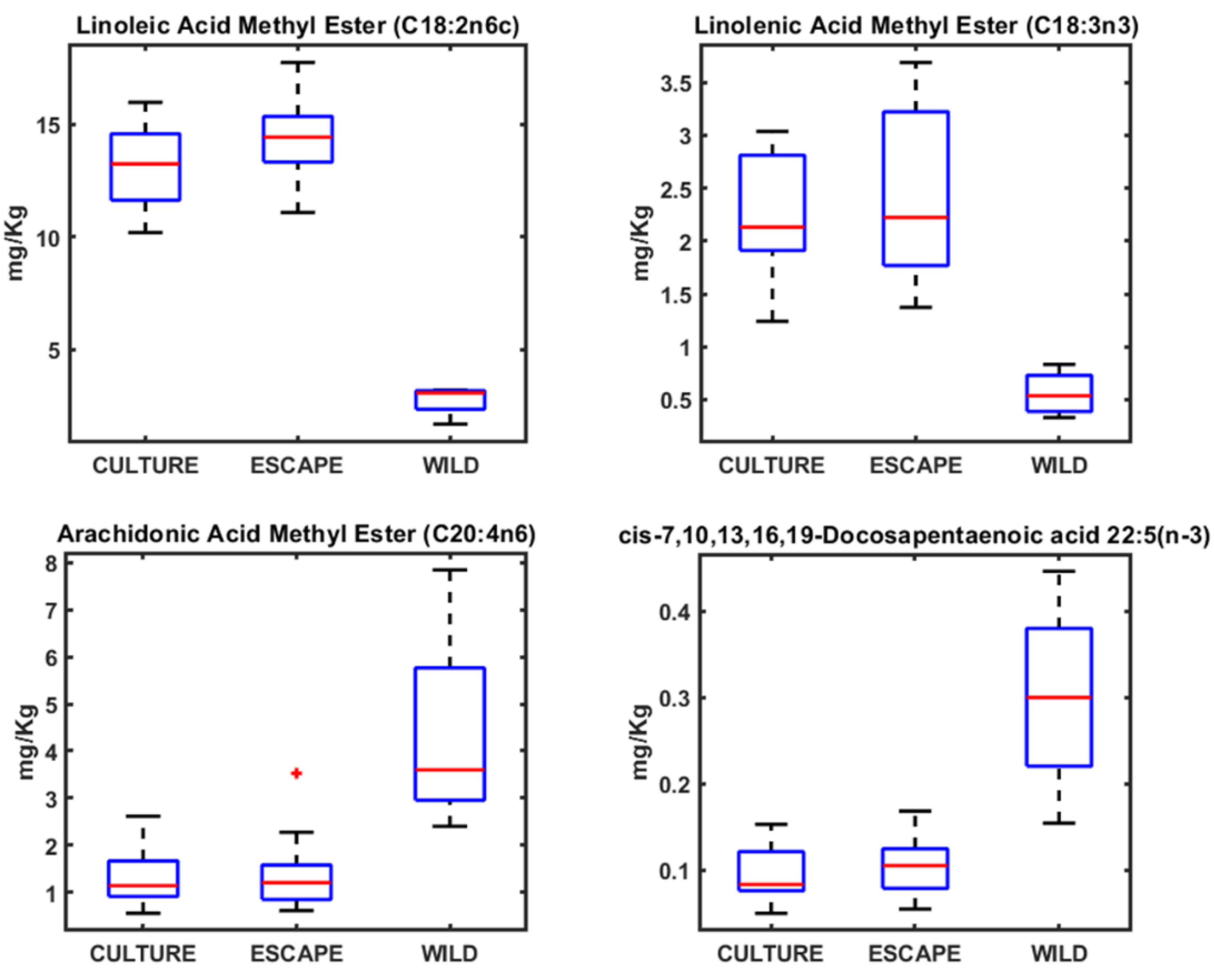
Figure 7.
Multidimensional scaling approaches on the metals and the AAGGs obtained from the muscle of meagre, seabream and seabass. The legend of the symbols is in the figure: AR (meagre), SA (seabream), DL (seabass), C (culture), E (escaped) and W (wild).
Figure 7.
Multidimensional scaling approaches on the metals and the AAGGs obtained from the muscle of meagre, seabream and seabass. The legend of the symbols is in the figure: AR (meagre), SA (seabream), DL (seabass), C (culture), E (escaped) and W (wild).
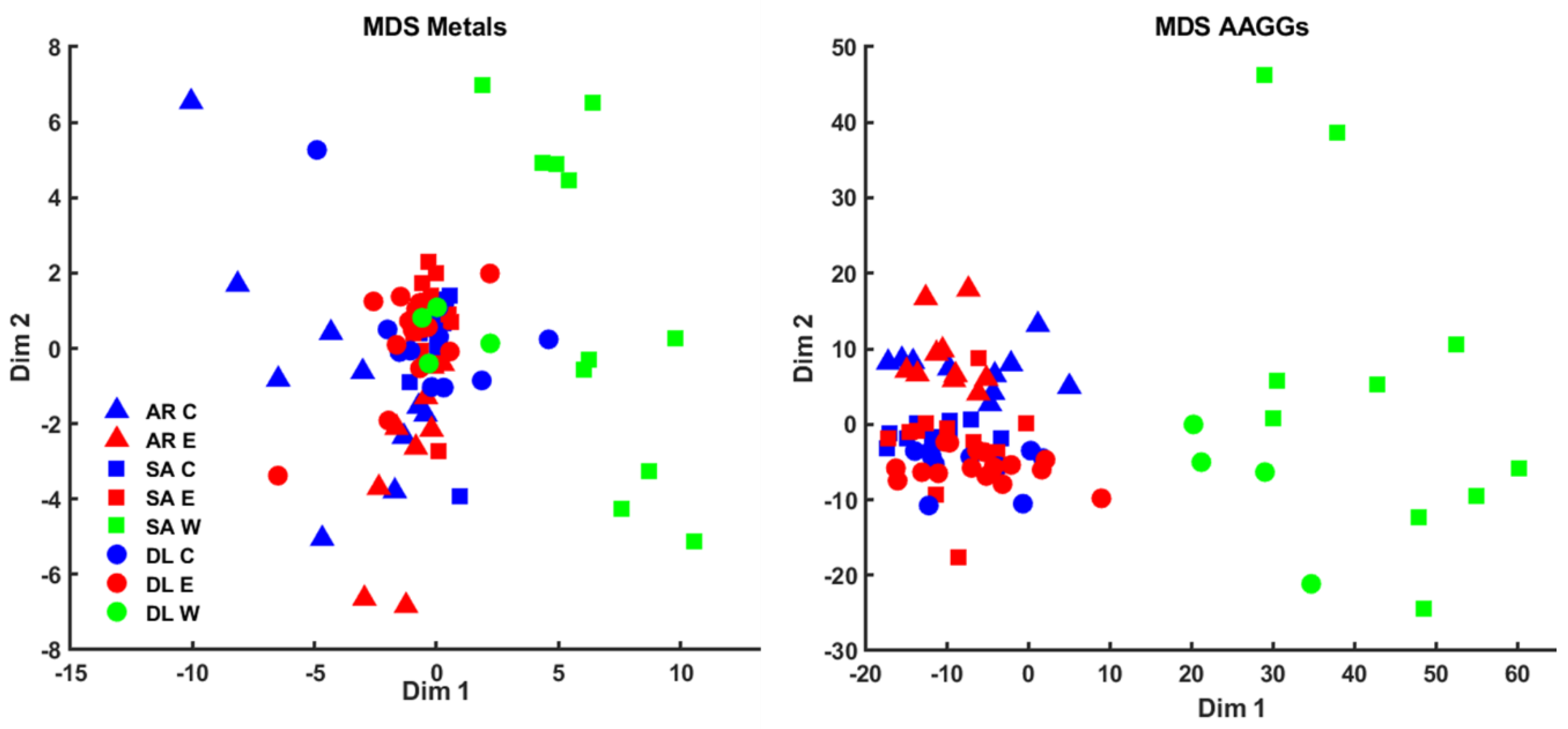
Figure 8.
Magnified representation of the region of the 1D 1H-HRMAS NMR spectra of muscle tissue from wild seabass (red line) and cultured and escaped seabass (blue line), and cultured and escaped meagre (green line) with the signals of the, taurine (Tau) (S-CH2, 3.26), (TMAO (N-CH3, 3.28 ppm), taurine (Tau) (N-CH2, 3.42), and glicine (Gly) (CH, 3.56 ppm). The signals of TMAO and Gly are singlets, and the signals of Tau are triplets.
Figure 8.
Magnified representation of the region of the 1D 1H-HRMAS NMR spectra of muscle tissue from wild seabass (red line) and cultured and escaped seabass (blue line), and cultured and escaped meagre (green line) with the signals of the, taurine (Tau) (S-CH2, 3.26), (TMAO (N-CH3, 3.28 ppm), taurine (Tau) (N-CH2, 3.42), and glicine (Gly) (CH, 3.56 ppm). The signals of TMAO and Gly are singlets, and the signals of Tau are triplets.
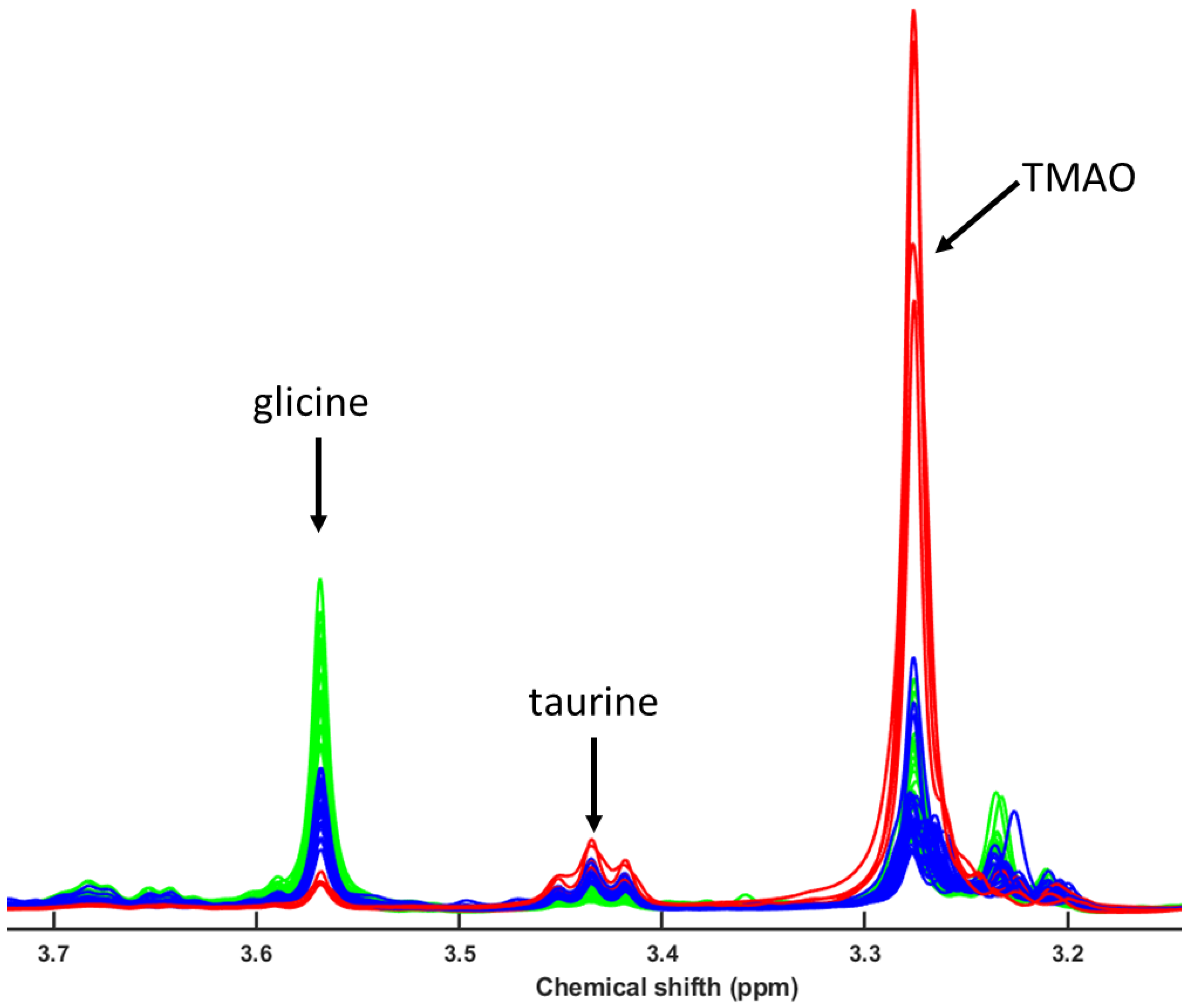
Figure 9.
The PLS-LDA model score plots of 1H NMR spectra for the polar fraction of gilthead sea bream muscle samples (blue circles for escaped and cultured seabream, and red circles for wild seabream) was used for classification the spectra for the polar fraction for wild seabass (red diamond) and escaped and cultures seabass and meagre (purple diamond. The VIP scores and the pseudospectrum format PLS-LDA tpLoading was included [6].
Figure 9.
The PLS-LDA model score plots of 1H NMR spectra for the polar fraction of gilthead sea bream muscle samples (blue circles for escaped and cultured seabream, and red circles for wild seabream) was used for classification the spectra for the polar fraction for wild seabass (red diamond) and escaped and cultures seabass and meagre (purple diamond. The VIP scores and the pseudospectrum format PLS-LDA tpLoading was included [6].
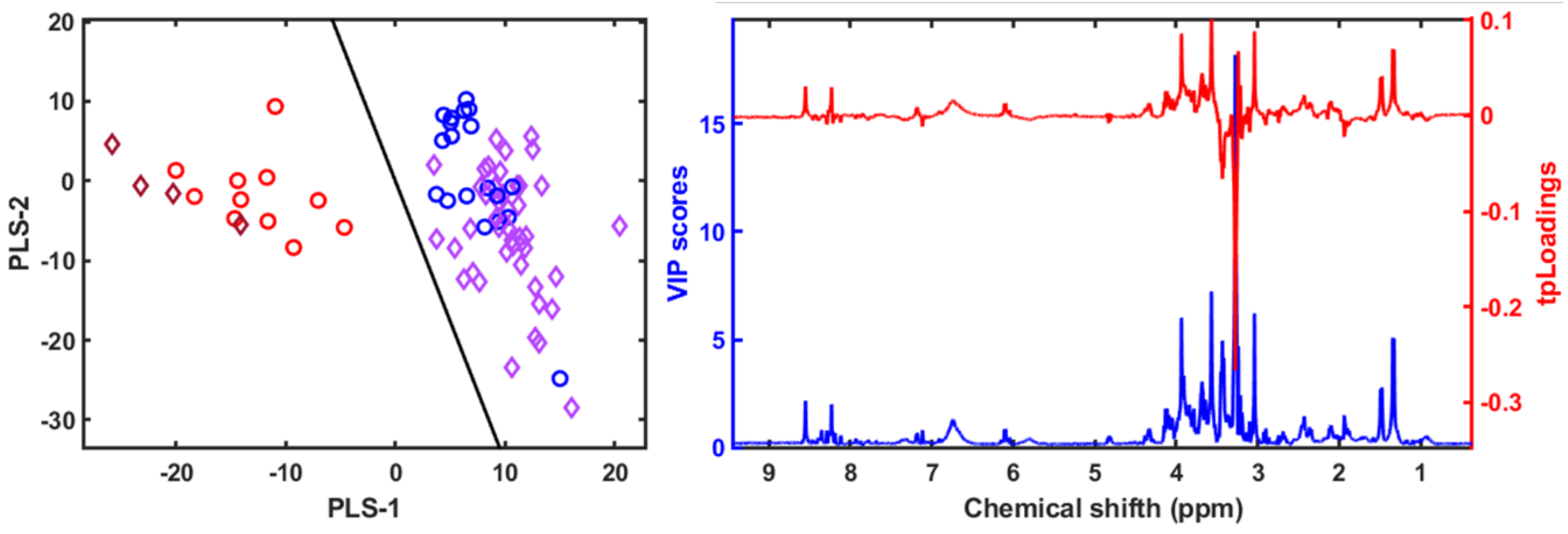
Figure 10.
1H NMR spectra of the nonpolar fractions of wild seabass (red line), cultured and escaped seabass muscle samples (green line), and cultured and escaped meagre (blue line). The insert shows the enhanced linoleic acid region.
Figure 10.
1H NMR spectra of the nonpolar fractions of wild seabass (red line), cultured and escaped seabass muscle samples (green line), and cultured and escaped meagre (blue line). The insert shows the enhanced linoleic acid region.
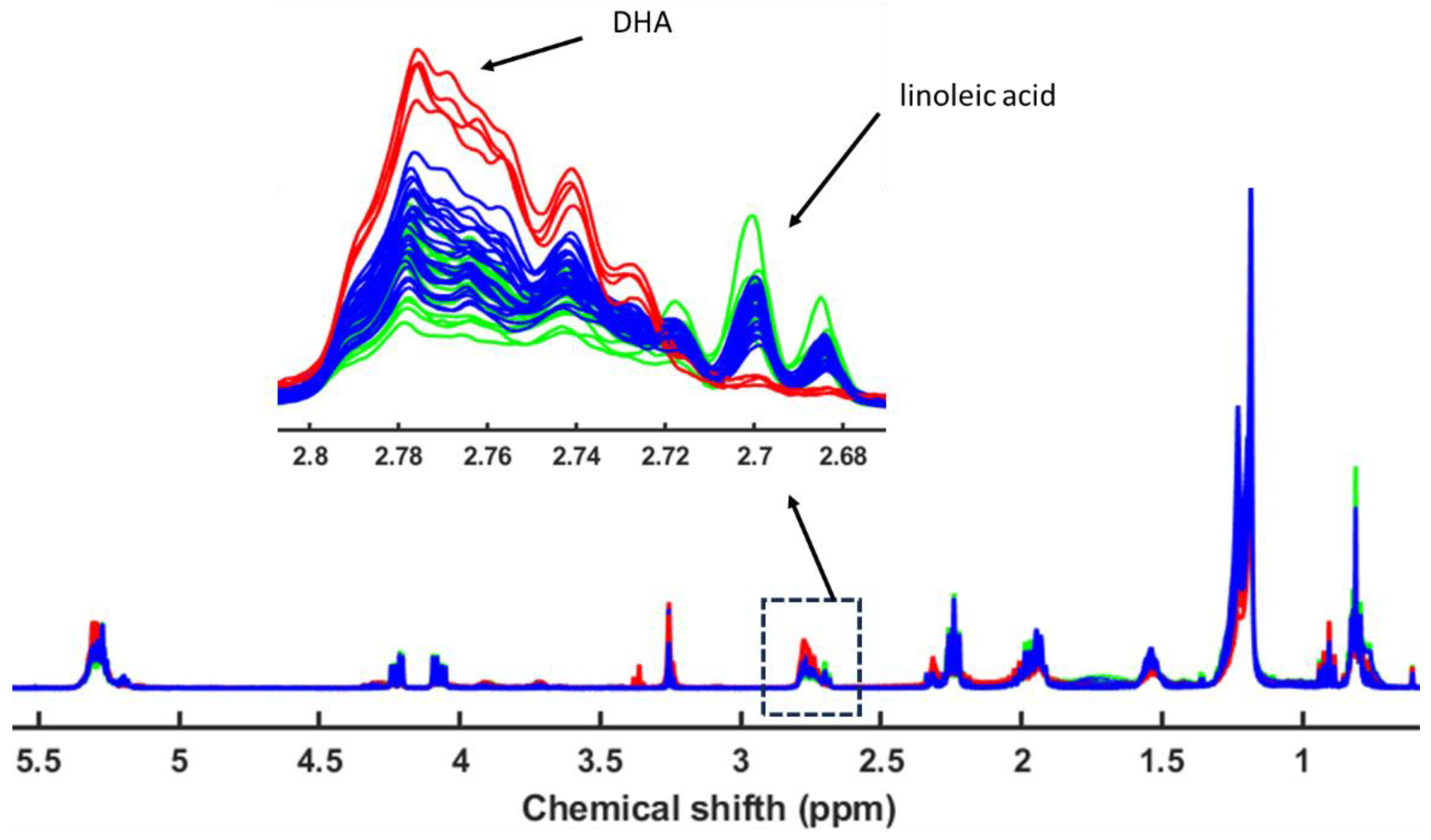
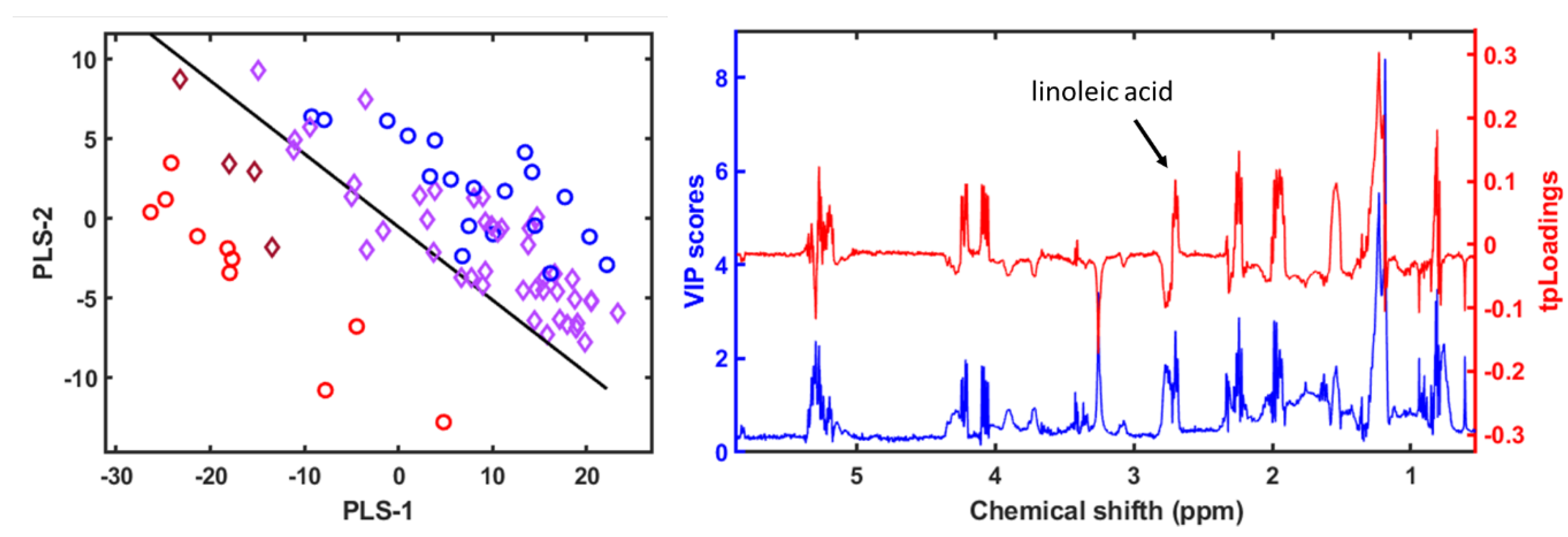
Figure 11.
The PLS-LDA model score plots of 1H NMR spectra for the apolar fraction of gilthead sea bream muscle samples (blue circles for escaped and cultured seabream, and red circles for wild seabream) was used for classification the spectra for the polar fraction for wild seabass (red diamond) and escaped and cultures seabass and meagre (purple diamond). The VIP scores and the pseudospectrum format PLS-LDA tpLoading was included [6].
Figure 11.
The PLS-LDA model score plots of 1H NMR spectra for the apolar fraction of gilthead sea bream muscle samples (blue circles for escaped and cultured seabream, and red circles for wild seabream) was used for classification the spectra for the polar fraction for wild seabass (red diamond) and escaped and cultures seabass and meagre (purple diamond). The VIP scores and the pseudospectrum format PLS-LDA tpLoading was included [6].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



