
Submitted:
26 March 2025
Posted:
26 March 2025
You are already at the latest version
Abstract
Over the last 40 years several studies have provided evidence demonstrating that viral vectors can result in improved gene replacement and insertions experiments. Viruses being invisible infectious agents can severely impact on organism’s life cycle. They are obligate intracellular pathogens that have evolved efficient strategies to hijack and manipulate the host’s immune system, and this nature of viruses can be exploited for its best utilization in molecular biology. Geminiviruses, for instance, are one of the largest families of plant viruses with small circular single-stranded DNA genomes, which efficiently manipulate the host’s genetic machinery for their own replication and protein expression, entailing great potential for genetic engineering applications. In this review, we describe how geminiviruses can open up a world of opportunities for efficient genome editing, thus lessening the major technical constraints including HDR-mediated genome editing and time-inefficient tissue culture procedure. Many promoters and terminators employed in the expression cassette development are from viruses and their modifications can be utilized to enhance protein expression studies. All these insights illustrate a broader picture of plant viruses, with emphasis on engineering plant viruses to ease genome editing practices for crop improvements.
Keywords:
Geminivirus
; biotechnology
; genome editing
; vector
; replicon
1. Introduction
Global agricultural productivity faces a great threat by plant viruses, leading to significant economic loses to food security. According to the Food and Agriculture Organization (FAO), nearly 40% of global agriculture production is lost due to diseases and plant pests attack [1], with plant viruses contributing significantly. Among plant viruses, geminiviruses (family: Geminiviridae) are infamous for their wide host range, efficient vector transmission and ability to cause severe diseases in many economically important crops, including cotton, wheat, tomato, cucumber, and papaya [2]. Geminiviruses get their name from their unique twin icosahedral capsid structure. These viruses have small, circular, single-stranded (ss) DNA genome, which is around 2.5 to 5.5 kb in length, encoding proteins essential for multiplication and transmission in the host plants (reviewed in [3,4]). The destruction caused by geminiviruses is well documented. For example, the tomato yellow leaf curl virus (TYLCV), vectored by whitefly (Bemisia tabaci) has severely impacted the production of tomato across 40 countries [5]. The cassava mosaic disease (CMD), caused by cassava mosaic geminiviruses (CMGs), jeopardized food security for millions of farmers as it leads to 80% yield loses in Sub-Sahara Africa [6]. Similarly, the cotton industry of the Indian subcontinent has suffered losses of billions of dollars due to cotton leaf curl disease (CLCuD) caused by the cotton leaf curl viruses (CLCuVs) which have been responsible for multiple epidemics in the region [7,8]. One reason for the appearance of epidemics in that region is the dispersal of single species of whiteflies, B. tabaci (Asia II 1) [9]. The virus-host interaction is usually manifested by symptoms development including vein thickening, plant stunting, leaf curling and chlorosis. All these symptoms exhibit strong viral hijacking and a limited host armoury. This phenomenon provokes the idea that geminiviruses have the capacity to deliver heterologous proteins into plants and can provide help in altering plant genome for trait improvement.
Plant virus-derived vectors have been widely utilized in plant genome engineering (reviewed in [10,11]) for efficient gene targeting (GT) and precise gene insertions through homologous recombination (HR) as well as production of range of vaccines [12]. The main goal for efficient genome editing is to improve plant traits to cope with major issues arising due to climate change, increased pests’ pressure and abiotic stresses, hence there is a dire need for adopting new techniques with low risks of pleiotropic effects. Recently, the potential application of geminivirus-derived vectors has gained much attention, and this idea proved successful [13,14,15], opening the world of opportunities by expanding efficient genome editing possibilities.
The aims of this review are: 1) to provide an understanding about the biological characteristics of geminiviruses, particularly those that can use utilized as vectors for genome editing; 2) to give an overview for engineering geminivirus-based vector and their implications in plant trait improvement; 3) to provide instances of successful geminivirus-based vectors and future perspectives; 4) to compare DNA viral vectors with RNA viral vectors and 5) to highlight the regulatory frameworks and consumers concerns regarding genome editing crops.
2. Geminiviruses and Their Unique Characteristics
Geminiviruses are the second largest family of plant viruses. Several factors including their genomic organization, wide host range, insect vector, and tissue tropism make these viruses unique. Geminiviruses encompass fourteen genera: Becurtovirus, Begomovirus, Capulavirus, Citlodavirus, Curtovirus, Eragrovirus, Grablovirus, Maldovirus, Mastrevirus, Mulcrilevirus, Opunvirus, Topilevirus, Topocuvirus and Turncurtovirus [16]. All these genera are collectively containing 520 species which cause important diseases (both in monocots and dicots) in most tropical and subtropical regions around the globe. Among these genera, Begomovirus (445 species), Curtovirus and Mastrevirus (45 species) consist of most viral species. Mastreviruses and curtoviruses have monopartite (single genome component) genome, vectored by leafhoppers and causing diseases in vegetable and cereals crops [17] while begomoviruses having one (monopartite/DNA-A) and two (bipartite/DNA-A and DNA-B) genome components, are mainly transmitted by whitefly and cause diseases in vegetable, root and fiber crops [18] (Figure 1). Furthermore, begomoviruses are also reported to have associated circular, single-stranded satellite DNA molecules, termed as alphasatellites (previously known as DNA-1; ~ 1.3 kb in size), betasatellites (previously known as DNA-β; ~ 1.3 kb in size) and deltasatellites (~ 0.7 kb in size) [19]. Curtoviruses and monopartite begomoviruses have similar genomic organization comprising of one intergenic region (IR) including promoters (bidirectional) for viral genes expression and the origin of replication. They encode two/three proteins (V2 and V3; curtoviruses only) on the virion-sense strand and the capsid protein while four or five proteins (C1, C2, C3, C4 and C5) on the complementary strand. Furthermore, the bipartite begomoviruses comprises one IR and encode two proteins, one on the V-strand that encodes nuclear shuttle protein (NSP: BV1) and second on the C-strand that encodes movement protein (MP: BC1) [20]. Generally, begomoviruses can be divided into four phylogenetic groups: 1) Old World; 2) New World; 3) sweepoviruses and 4) legumoviruses. The Old World begomoviruses consist of both mono- or bipartite viruses having V2 protein. Whereas the New World begomoviruses are do not contain V2 protein and mostly bipartite [21,22]. They share a common region (CR) within the IR/long IR (LIR). On the other hand, mastreviruses have two IRs, the long IR and a short IR. These viruses utilize transcript splicing, and therefore their genomes contain introns. The C1 and C2 open-reading frames (ORFs) are involved in replication and encode replication initiator protein and have V2 and coat protein (CP) on the virion-sense strand.
3. Geminiviruses: Master of Host Genome’s Manipulator
Geminiviruses, with their small genome size, chiefly rely on host machinery for replication and symptom progression. With limited coding capacity, these viruses encode 4 to 8 multifunctional proteins including the newly identified small additional proteins necessary for infection and replication [20,23]. To redirect cellular functions for viral genome replication and spread, geminivirus proteins target a plethora of highly connected proteins (hubs) of the plant cell [24]. This virus-host interaction leads to altered gene expression, suppression of cell death mechanisms, interference with plant signaling pathways, and suppression of plant defenses, including the inhibition of antiviral silencing pathways [25,26].
3.1. Geminiviruses Lifecycle: Hijacking Host Nuclear Machinery
Geminiviruses begin their journey by entering and hijacking plant cell nuclei. The ssDNA genome of geminivirus replicates in the nucleus through rolling circle replication (RCR), followed by the conversion of ssDNA into double-stranded (ds) DNA, mediated by the plant DNA polymerase. This dsDNA acts as a template for replication and transcription of viral genes. The geminivirus-encoded (Rep/RepA/C1) protein plays a pivotal role in initiating host cell reprogramming by interacting with several host cellular proteins. Then, the Rep/RepA identifies nonanucleotide sequence within the conserved IR of the viral DNA and produces a site-specific nick on one of the two strands of dsDNA molecule. Consequently, by utilizing host DNA polymerase, Rep initiates replication, mediate circularization followed by the release of nascent virion (ssDNA) molecule. Additionally, replication enhancer protein (Ren/C3) collaborates with Rep/RepA along with other host replication-associated proteins to boost accumulation of viral genomes [27]. Eventually, newly formed viral molecules move inter-/intracellularly and invading new host cells and generating new ssDNA molecules available for acquisition by the insect vector. Viral DNA replication has also been modulated by a recombination-dependent replication process, which is based on homologous recombination [4]. However, Rep/RepA is sufficient for effective replication of viral genome.
After winning the battle in the nucleus and conquering the plant antiviral defenses, geminiviruses cross cellular boundaries, move from cell-to-cell and establish systemic infections. Within the cell, CP transfers viral DNA into the nucleus following by the uncoating and generation of new viral molecules. The newly synthesized ssDNA or dsDNA molecules are transferred from the nucleus in the cytoplasm by NSP in bipartite geminiviruses and CP in monopartite geminiviruses. Reaching the cellular periphery, plasmodesmata (PD) are reportedly involved in facilitating the cell movement of the viral genomes. Bipartite viruses MP are located at PD openings, facilitating this passage by pushing the exclusion limits of PD. In monopartite geminiviruses, similar roles have been speculated for C4 protein [28]. Understanding the roles of viral components is essential for the effective utilization of geminiviruses in plant biotechnology. The Rep and IR regions are perhaps the most vital elements required for geminivirus replication. Efficient replication of a vector system is the most desired feature in molecular biology; however, cell-to-cell movement and encapsidation—typical features of the viral life cycle—are the least desirable in viral vectors [29]. Thus, this presents an opportunity to customize geminivirus genomes for the development of geminivirus-derived vectors.
4. Geminiviruses: How They Can Help in Plant Genome Editing
4.1. Overview of CRISPR-Cas System
In the field of plant genetic engineering, manipulation of the plant’s genome to improve its production or resistance to abiotic and biotic stresses is fundamental. DNA manipulation is achieved through genome editing technologies, which can be divided into two categories. 1) Conventional genome editing technologies which includes, meganucleases, zinc-finger nucleases (ZFNs), and transcription activator like effector nucleases (TALENs); and 2) clustered, regularly interspaced short palindromic repeats-CRISPR/CRISPR-associated proteins (CRISPR-Cas) which is more efficient and precise than conventional ones. These techniques are based on the site-directed nucleases (SDNs), which introduce specific and targeted modifications at the DNA double-stranded breaks (DSBs), generated by the DNA cutting nucleases (for instance, Cas9) [30]. To preserve the genome integrity, plant cellular defense machinery repairs the DSBs either with nonhomologous end joining (NHEJ) pathway or with homology-directed repair (HDR) pathway [31]. Based on the editing outcomes, there are three types of SDNs: 1) SDN-1, which occurs without any DNA template and results in small indels (additions/deletions). The indels occur through NHEJ repair mechanism, thus has the potential to knock out corresponding gene [32]; 2) SDN-2, which uses a short DNA template (donor DNA; homologous to the target region) and generates a small indel or single-nucleotide variant (SNV) through homology-directed repair (HDR); 3) SDN-3, which involves the insertion of large DNA template (kilobase-scale) into a desired locus via HDR to the cleavage site. As the latter two types use HDR cellular repair mechanism, thereby allowing precise allele or gene replacement or insertions (Figure 2). In general, CRISPR-based genome editing involves different approaches: nuclease editing, which can create small indels, base and prime editing, which enable base substitutions and small indels; and large DNA fragment editing, which employs strategies similar to prime editing, CRISPR-associated recombinases or transposases [33,34,35]. The HDR-driven knock-in (KI) remains very challenging due to the delivery of repair template (i.e., donor DNA). However, several approaches have been employed to enhance HDR frequency including, the expression of genes involved in HR, use of single-stranded donor DNA and by using viral vectors such as DNA or RNA viruses [36]. One of the main challenges for optimal editing outcomes is the delivery of CRISPR reagents (Cas and gRNAs) and repair template into the plant cell. To date, Agrobacterium-, biolistic-, particle bombardment-, floral dip-, poly (ethylene glycol) (PEG)- and protoplast-mediated transformation are extensively used as delivery methods. However, these methods have several limitations, including species dependency, embryonic mortality, high cost, time inefficient and low transformation efficiency. As an alternative, plant viruses including RNA and DNA viruses-mediated vectors for CRISPR reagents delivery, offering great potential solution (reviewed in [10,11,37]. The virus’s ability to autonomously replicate within plant cells and produces high copy number of donor DNA leads to enhance protein expression, thus making viruses well-suited for cargo delivery.
4.2. Geminivirus-Based Vectors: A Stalwart Approach for Genome Editing
Over the last 40 years, geminiviruses have been exploited to deliver CRISPR reagents and repair template of varying length into plant cells. Geminiviral DNA replication mainly depends on two viral elements i.e., Rep/RepA and the IR. These two elements act as a building block in the replication of replicons in plant cell. The sequence of interest is flanked between two IR (or LIR in case of Mastreviruses) (Figure 3a). Upon Rep/RepA protein expression, DNA molecules comprising gene of interest will be produced. Note that, replicons production is independent of whether the Rep/RepA protein is expressed from the viral genome or transgenically. Furthermore, these molecules replicate due to the action of Rep/RepA, and it accumulates in abundance in the cell, making geminiviruses well suited for the templated HDR-based genome editing, as this method contains donor DNA and requires in high copy number. By following this ability of geminivirus-derived replicons, Gil-Humanes et. al., and Wang et al., did successful insertions in wheat and rice genomes [13,38] which is discussed later in this review.
The short and long intergenic regions (SIR and LIR, respectively) on geminiviral genome allow a linear construct to become circular by joining of two LIR sequencing with the help of viral Rep/RepA proteins (Figure 3b). This allows employing ‘LSL’ (LIR-SIR-LIR) vectors, by which linear construct (e.g., BeYDV (deconstructed virus; (Figure 3c) detailed mentioned in next paragraph), consisting of a SIR and flanking by two LIR can be transferred to the Agrobacterium strain, followed by the Agrobacterium-mediated plant transformation (Figure 4) [39]. Broadly, there are two methods by which geminiviruses can be modified and utilized as a nucleic acid delivery: 1) full virus vector strategy; and 2) deconstructed virus vector strategy [40].
4.3. Full- and Deconstructed- Virus Vector Strategy
In full virus vector strategy, geminiviruses keep most of all the genes necessary for its replication and infection of the host. The gene of interest (heterologous sequence) can be introduced into the virus genome by replacing CP gene. In some bipartite viruses, the CP is not required for cell-to-cell movement (Figure 3c) [41]. In a study by Hyes et al., a bipartite geminivirus (tomato golden mosaic virus; TGMV) has been used to express neomycin phosphotransferase (neo) in N. tabacum cv. Samsun [42]. The authors developed a chimeric DNA by replacing viral CP with the neo gene and showed that it has successfully replicates and spread systemically through the plant. Note that, CP is not required for successful viral infection or systemic spread throughout the plant [43]. Another example of DNA virus deployed as a gene replacement vector is cassava latent virus (CLV). The CLV CP has been replaced with the bacterial chloramphenicol acetyl transferase (CAT) gene. The CAT expression was mediated under the control of CP promoter and they successfully exhibited that the CAT expression of 80 U/mg occurred in systemic infected N. benthamiana leaves [44]. Furthermore, Baltes et al., utilized a recombinant cabbage leaf curl virus (CaLCuV) to deliver repair template. They target Arabidopsis thaliana alcohol dehydrogenase (ADH1) gene. The authors modified DNA A genome of CaLCuV by incorporating 18-bp sequence in the repair template (total size of the donor DNA was 600 bp). After infecting the Arabidopsis plants with CaLCuV, they identified one plant with the desired insertion. Thus, suggesting that full viruses vector can be deployed for precise gene insertion [15].
The disadvantage of this strategy is that it may not be possible to integrate large heterologous sequences of up to 800 bp [45]. Virus genome size limitations are due to the cell-to-cell movement via plasmodesmata and not by replication [46]. However, this capacity is not enough for expressing TALEN and Cas9, but it is suitable to express gRNAs. Another approach, researchers utilized is to generate Cas9 stable transgenic plant and they express gRNAs in viral replicons [47]. Thus, eliminating the size constraints and make it possible for targeted genome engineering.
In deconstructed virus vector strategy, the undesired components (e.g., MP and CP) are removed and only useful components [ e.g., cis-(IR) and trans-(Rep/RepA)], important for replication, are retained to overcome the genome size constraints (Figure 3c). This movement requirement is fulfilled by using Agrobacterium-mediated viral vector transformation to the plant cells.
4.3. Geminivirus-Based Homology-Directed Repair: Case Studies
Two geminiviruses, wheat dwarf virus (WDV) and tomato leaf curl virus (ToLCV) were used for gene targeting in wheat, corn and rice [13,38]. In a study by Gil-Humanes et. al., the authors generated a deconstructed version of WDV and expressed reporter gene in the wheat cells in a 110-fold increase compared to the control. They also checked gene targeting efficiency by delivering Cas9 and repair template in a ubiquitin locus that resulted in 12-fold higher efficiency than non-viral methods. They further highlighted that nuclease (Cas9) promoter played a significant role in efficient GT by comparing maize ubiquitin promoter (ZmUbi) with viral LIR promoter and suggested that ZmUbi have significantly enhance the GT. The findings of targeted integration by HDR in all three wheat homoeoalleles (A, b and D) [13], providing evidence that geminiviral-based replicons have the potential for precise and efficient HR-based genome editing in cereals.
In another study by Wang et al., authors performed geminiviral-based knock-in (KI) in rice. They selected two loci (ACT1 and GST) for targeted KI and incorporated GFP fused cassettes (ACT1-GFP and GST-GFP). For this, first they inserted these cassettes into WDV replicon to make WDV expression cassettes (WDV1-ACT1 and WDV2-GST, respectively). Next, they incorporated U6-driven gRNA. One barrier in the study was the larger Cas9 size that they remained unable to incorporate into the replicon. To overcome this, they generated transgenic rice calli expressing Cas9 and utilized to inoculate WDV-based generated vectors. They found that the targeted KI frequency for ACT1 and GST were 8.5% and 4.7%, respectively, which was lower than using Cas9 expressing rice calli [38].
Another geminivirus-based vector, Beet curly top virus (BCTV) was deployed to deliver Cas12a in GFP-transgenic N. benthamiana. The authors removed the virion-sense genes (V1, V2 and V3) and inserted constitutive cauliflower mosaic virus (CaMV) 35S promoter upstream of the Cas12a. They found higher indel rate of approximately 40 % in the GFP target region compared to Agrobacterium-mediated T-DNA delivery. Furthermore, they compared three different Cas nucleases (LbCas12a, AsCas9 and SpCas9) and found that LbCas12a nuclease showed higher efficiency with no evidence of off-target mutations [48]. Moreover, BCTV represented higher rate mutagenesis than previously deployed geminiviruses including BeYDV and Sweet potato leaf curl virus (SPLCV) [15,49].
Recently, in a preprint posted last year, the team performed experiments to determine optimal composition of BCTV-based viral vector to deliver donor DNA in gene targeting experiment and to produce high levels of expression of a desire gene. They utilized GoldenBraid (GB) modular cloning system that is based on Type IIS restriction enzymes and DNA parts that consist of standardized ‘grammar’ of 4-nt overhangs [50]. Furthermore, they also found that native BCTV promoter inclusion upstream to GOI performed better than previously used constitutive CaMV 35S promoter. Moreover, BCTV-replicons was able to accommodate cargo of up to 4 kb long without impacting replicon accumulation and removing virion-sense genes further enhance cargo expression [51].
Over the past decade, several advancements in the field of genome editing and omics provide much evidence for utilizing geminiviral vector technology [34,52,53]. Furthermore, HDR-based genome editing which is quite challenging in the past have now possible by utilizing the power of geminivirus vectors. Although, just like every past technology, virus-induced genome editing (VIGE) is also having few drawbacks including off-targets, limited cargo capacity, elimination of viruses in systemically infected plant, for heritable editing, germline cells must be infected, etc.
Recently, to reduce CRISPR-mediated off-targets, Vu et al., utilized a novel combination of prime editing (PE) components which consists of RNA chaperone and modified prime editing guide RNAs along with a geminiviral (BeYDV)-based replicon system and obtained 9.7% desired PE efficiency at the callus stage. They carried out this experiment in tomatoes and Arabidopsis and obtained 38.2% positive transformants contained desired PE alleles [54], proving a potential heritable PE transmission and unlocking the practical applications of geminiviral replicons in dicots. Another strategy to obtain genome edited progeny is to deliver only gRNA with geminiviral replicon into stable transgenic plant, expressing Cas9. This method has emerged as a powerful tool in plant genome engineering. In a study by Yin et al., the authors engineered a cabbage leaf curl virus (CaLCuV) to deliver gRNA by replacing its CP with gRNA [47]. Compared to the conventional virus-induced gene silencing (VIGS), geminiviral replicon-based genome editing (VIGE) have two advantages which are; 1) VIGS can cause additional non-specific silencing especially for homologous genes while CRISPR-based VIGE can only target specific gene and cause knock-out after repairing DSB by NHEJ, providing optimal tool to study specific gene function; 2) In VIGS, cloning is required to synthesis a fragment of a targeted gene by PCR while in VIGE, only small fragment of about 20 bp in length target sequence into gRNA is required, providing a better way to establish a high-throughput genome wide function analysis.
4.4. BeYDV: A Successful and Pioneer Geminivirus-Based Vector
Several successful reports of utilizing deconstructed viral replicons are available. For instance, one of the pioneer geminivirus-based vector, bean yellow dwarf virus (BeYDV) was deployed for the expression of vaccine proteins, reporter protein and monoclonal antibodies [39]. Deconstructed virus vector strategy has several advantages over the use of full virus vector strategy: 1) no cargo size limit reported with this strategy. However, replication efficiency may be affected with the increased replicon size.; 2) lethal outcomes driven from the other viral proteins are ignored; 3) less restrictions imposed by the host range, however, needs to be considered. A comprehensive list of deconstructed geminiviruses-based replicons has been provided in supplementary file 1.
Geminiviruses-derived deconstructed vectors including BeYDV was used to efficiently deliver large sequences including repair templates and ZFNs and TALEN to induce mutations in potato [55], tomato [14] and tobacco [15]. This replicon has 10- to 100-times higher efficiency than traditional Agrobacterium T-DNA delivery. The BeYDV replicons system were also utilized to express Streptococcus pyogenes Cas9 (SpCas9). Furthermore, BeYDV replicons was also utilized for targeted mutations in potato and cassava for enhance herbicide tolerance [55,56]. Besides gene targeting, geminiviral vectors can replicates inside the host’s nucleus with high copy numbers. Thus, unveiling the utilization of geminiviral replicons as a repair template for HDR. By merging this replicon strategy with Agrobacterium-mediated T-DNA, it would be possible to get high editing outcomes in a more efficiency manner [57]. One way is to utilize this strategy is loading repair template is on geminiviral replicon and nucleases can be expressed using either viral replicon or traditional T-DNA. An example of employing BeYDV as an HDR-based genome editing is in tobacco and tomato in which the replicons successfully recover the precise wild-type sequence provided by a donor template and incorporate strong 35S promoter upstream of the tomato endogenous ANT1 gene. The efficiency was ~12 times higher than standard Agrobacterium T-DNA delivery [14]. BeYDV-derived vectors have effectively investigated the effects of phytohormones on plant biomass using transient expression system and efficiency of expression was recorded 19-25% more than controls for different hormones [58]. Abovementioned example suggests that geminiviral replicons can be modified and be merged with other delivery methods T-DNA and nanoparticles, etc. We have provided a list geminiviruses that have been deployed as vectors for genome editing, along with detailed descriptions in Table 1. Additionally, an extensive list of geminiviral vectors employed in plant genome engineering is provided in supplementary file 1.
5. Comparison of Geminivirus-Based Vectors with Other Plant Viral Vectors
The circular, ssDNA genome of geminiviruses have certain advantages and disadvantages over linear RNA viruses. For instance, cargo capacity, genomic size and organization and ability of hijacking host genetic material. We are listing few points based on the research that has been made so far and provide few potential strategies which help in the utilization of plant viral vectors. The points include 1) the ability of geminiviruses to autonomously replicate within the host nucleus at high levels has been the key to their success as vector. To date, HDR-based genome editing remains highly challenging. However, recent studies have demonstrated the feasibility of using geminivirus-based vectors to deliver donor DNA in plants [15,38,48]. Thus, geminiviruses can easily integrate heterologous proteins in their circular genome and can offer a potential to stably deliver nucleases, gRNAs and donor DNA than linear genome of RNA viruses; 2) the size constraints of geminiviruses can be compromised by removing unnecessary viral genes including coat and movement proteins, strategy known as deconstructed viral vector (Figure 3c) [13,14]. Thus, providing a huge potential to facilitate larger proteins in geminivirus-based vectors. In addition to removing virion-sense genes, various heterologous promoters have also been utilized. Eini et al., successfully delivered Cas12a (Cpf1; ~1300 amino acids) into plants by removing the virion-sense genes of the Beet curly top virus (BCTV). To express Cas12a and other cargoes, they employed CaMV 35S promoter [48]. Studies have shown that a BCTV-based vector can accommodate cargo up to 4 kb in size without affecting viral replication [29]; 3) newly discovered smaller size Cas nucleases can provide a way forward to precisely and efficiently screening the gene function of plants. The smaller Cas nucleases include Campylobacter jejuni Cas9 (CjCas9; 984 amino acids), CasΦ (Cas12j; 800 amino acids) and more recently discovered NanoCas (425 amino acids) [61,62,63]. Note that NanoCas has not been tested in plants yet. However, if it demonstrates equal or higher efficiency than traditional Cas9, it could be a game-changer for plant biotechnology, as it would allow the use of a single viral vector instead of generating two viral cassettes to express Cas and gRNAs, simultaneously; 4) RNA-viruses like SNYV, lack a DNA phase during replication and due to this ability RNA-virus-based vectors have the advantage to not integrate into the host genome. Thus, mutant plants regenerated from infected tissues are non-transgenic. On contrary, it is extremely difficult to eliminate geminivirus-based vectors by regeneration as geminiviral protein interact with various host cellular proteins for viral replication. A plausible solution to this would be modification of strictly inducible geminivirus vectors that can only express when plant reaches a specific biomass; 5) one of the major disadvantages of using geminiviral vectors is their perturbation into plant cell cycle during viral replication process [64]. Whereas RNA viruses do not have this drawback as these viruses’ genomes packaged within their capsids. The extensive list of RNA viruses-based vectors is provided in Table 2.
6. Outlook to Viral Vector-Based Crop Design
The deployment of plant viruses-based vectors to deliver heterologous proteins to plants, offers effective and time-efficient method. To date, number of RNA viruses including TRV, bamboo mosaic virus (BMV), BYSMV, BSMV, beet necrotic yellow vein virus (BNYVV), pea early browning virus (PEBV), PVX, and SYNV have successfully demonstrated efficient, targeted and heritable mutations [66,68,69,77,82] in dicot plants whereas, a number of viral vectors including WDV, BSMV and foxtail mosaic virus (FoMV) have been reported in monocots plants [13,71,72,74]. Most of these viruses were reported to induce targeted, stable and heritable mutations. Apart from this, a number of RNA viruses including BaMV, BSMV, BYSMV, brome mosaic virus (BMV), cymbidium mosaic virus (CymMV), cucumber mosaic virus (CMV), FoMV, rice tungro bacilliform virus (RTBV) and wheat streak mosaic virus (WSMV) are deployed for gene suppressing and systemic protein expression analysis [72,81,83,84,85,86,87,88]. Once the editing efficiency and heritable mutations of modified alleles are optimized, VIGE would bypass the need of plant regeneration and transformation, thus boosting the breeding of new climate-friendly crop varieties. Moreover, absence of exogenous sequences (in case of RNA viruses-derived vectors) would make genome editing indistinguishable from the traditional mutagenesis approaches and also benefitting for regulatory approval [89].
Plant viral vectors can be utilized to rewire different biosynthetic pathways by transferring of transcription factors, precise and targeted knock-down or overexpression of metabolic genes or heterologous enzymes. Similar methods might be applied for field-grown crops biofortification as an alternative or in conjunction to traditional breeding and transgenic approaches. Furthermore, viral vectors can be used to express positive regulators ectopically as discussed before, in an example of FT gene [82]. Virus-induced flowering enhance FT accumulation, early flowering and has further been applied to promote breeding programmes and genetic studies of cotton, citrus and apple [90]. In addition, virus-induced flowering is also helpful for tree species with long juvenile phase that can take years or even decades.
Geminiviruses have a wide host range that make them unprecedented in the development of viral vector for delivering genome editing reagents. With the discovery of miniature Cas (NanoCas), it would be much easier to manage multiple gRNAs along with nuclease for precise and efficient gene targeting. Multipartite viruses including geminiviruses (helper viruses/satellites) can also be engineered for multiple gene silencing and protein co-expression assays as suggested by Liou et al. [91]. Moreover, geminiviral vectors have potentially useful for efficient genome editing, especially in transient assays in model plants such as tobacco, as well as in the regeneration of transgenic plants harbouring the desire mutations. However, it can take several months in regeneration and is difficult to scale up to a high-throughput level. As geminiviruses are not seed-transmissible, thus deconstructed geminiviral strategies could potentially be utilized for genome editing in plants, offering a novel system to obtain stable mutant progeny in a single generation, for example, by modifying reproductive tissues.
Lastly, regulatory frameworks and consumers concerns regarding genome edited crops and the use of viral vectors cannot be overlooked. Globally, consumers are showing misconceptions, inadequate understanding and a general unfamiliarity with genome editing technologies in plants [92]. Such consumer behavior is influenced by cultural, social, personal, psychological and economic factors including lifestyle, income, personal welfare, scientific knowledge, religion and beliefs [93]. Nonetheless, without embracing these ground-breaking technologies, the issues of genome editing and, hence, global food security will be a monumental challenge.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
MAM provided the outlines of the review and contributed the key ideas. MAM, MWS, II and MQS wrote the manuscript. MAM prepared the figures. RZN, SM and MAM reviewed and improved the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were generated.
Acknowledgments
We are grateful to reviewers for all the suggestions.
Conflicts of Interest
The authors declare no competing interests.
References
- FAO. FAO’s Plant Production and Protection Division. 2022. [CrossRef]
- Mansoor, S.; Zafar, Y.; Briddon, R.W. Geminivirus disease complexes: the threat is spreading. Trends Plant Sci 2006, 11, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Mansoor, S.; Briddon, R.W.; Zafar, Y.; Stanley, J. Geminivirus disease complexes: an emerging threat. Trends Plant Sci 2003, 8, 128–134. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: masters at redirecting and reprogramming plant processes. Nat Rev Microbiol 2013, 11, 777–788. [Google Scholar] [CrossRef]
- Wan, H.J.; Yuan, W.; Wang, R.Q.; Ye, Q.J.; Ruan, M.Y.; Li, Z.M.; Zhou, G.Z.; Yao, Z.P.; Yang, Y.J. Assessment of the genetic diversity of tomato yellow leaf curl virus. Genet Mol Res 2015, 14, 529–537. [Google Scholar] [CrossRef]
- Uke, A.; Tokunaga, H.; Utsumi, Y.; Vu, N.A.; Nhan, P.T.; Srean, P.; Hy, N.H.; Ham, L.H.; Lopez-Lavalle, L.A.B.; Ishitani, M.; et al. Cassava mosaic disease and its management in Southeast Asia. Plant Mol Biol 2022, 109, 301–311. [Google Scholar] [CrossRef]
- Mahmood, M.A.; Ahmed, N.; Hussain, A.; Naqvi, R.Z.; Amin, I.; Mansoor, S. Dominance of Cotton leaf curl Multan virus-Rajasthan strain associated with third epidemic of cotton leaf curl disease in Pakistan. Sci Rep 2024, 14, 13532. [Google Scholar] [CrossRef]
- Mansoor, S.; Amin, I.; Iram, S.; Hussain, M.; Zafar, Y.; Malik, K.A.; Briddon, R.W. Breakdown of resistance in cotton to cotton leaf curl disease in Pakistan. Plant Pathol 2003, 52, 784–784. [Google Scholar] [CrossRef]
- Mahmood, M.A.; Ahmed, N.; Hussain, S.; Muntaha, S.T.; Amin, I.; Mansoor, S. Dominance of Asia II 1 species of Bemisia tabaci in Pakistan and beyond. Sci Rep 2022, 12, 1528. [Google Scholar] [CrossRef]
- Mahmood, M.A.; Naqvi, R.Z.; Rahman, S.U.; Amin, I.; Mansoor, S. Plant Virus-Derived Vectors for Plant Genome Engineering. Viruses 2023, 15. [Google Scholar] [CrossRef]
- Abrahamian, P.; Hammond, R.W.; Hammond, J. Plant Virus-Derived Vectors: Applications in Agricultural and Medical Biotechnology. Annu Rev Virol 2020, 7, 513–535. [Google Scholar] [CrossRef]
- Hefferon, K. Plant Virus Expression Vectors: A Powerhouse for Global Health. Biomedicines 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Gil-Humanes, J.; Wang, Y.; Liang, Z.; Shan, Q.; Ozuna, C.V.; Sánchez-León, S.; Baltes, N.J.; Starker, C.; Barro, F.; Gao, C.; et al. High-efficiency gene targeting in hexaploid wheat using DNA replicons and CRISPR/Cas9. Plant J 2017, 89, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Čermák, T.; Baltes, N.J.; Čegan, R.; Zhang, Y.; Voytas, D.F. High-frequency, precise modification of the tomato genome. Genome Biol 2015, 16, 232. [Google Scholar] [CrossRef]
- Baltes, N.J.; Gil-Humanes, J.; Cermak, T.; Atkins, P.A.; Voytas, D.F. DNA replicons for plant genome engineering. Plant Cell 2014, 26, 151–163. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Lett, J.M.; Martin, D.P.; Roumagnac, P.; Varsani, A.; Zerbini, F.M.; Navas-Castillo, J. ICTV Virus Taxonomy Profile: Geminiviridae 2021. J Gen Virol 2021, 102. [Google Scholar] [CrossRef]
- Shepherd, D.N.; Martin, D.P.; Van Der Walt, E.; Dent, K.; Varsani, A.; Rybicki, E.P. Maize streak virus: an old and complex 'emerging' pathogen. Mol Plant Pathol 2010, 11, 1–12. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu Rev Phytopathol 2011, 49, 219–248. [Google Scholar] [CrossRef]
- Lozano, G.; Trenado, H.P.; Fiallo-Olivé, E.; Chirinos, D.; Geraud-Pouey, F.; Briddon, R.W.; Navas-Castillo, J. Characterization of Non-coding DNA Satellites Associated with Sweepoviruses (Genus Begomovirus, Geminiviridae) - Definition of a Distinct Class of Begomovirus-Associated Satellites. Front Microbiol 2016, 7, 162. [Google Scholar] [CrossRef]
- Ahmed, N.; Mahmood, M.A.; Amin, I.; Mansoor, S. Geminiviruses also encode small proteins with specific functions. Trends Microbiol 2021, 29, 1052–1054. [Google Scholar] [CrossRef]
- Trenado, H.P.; Orílio, A.F.; Márquez-Martín, B.; Moriones, E.; Navas-Castillo, J. Sweepoviruses cause disease in sweet potato and related Ipomoea spp.: fulfilling Koch's postulates for a divergent group in the genus begomovirus. PLoS One 2011, 6, e27329. [Google Scholar] [CrossRef]
- Ilyas, M.; Qazi, J.; Mansoor, S.; Briddon, R.W. Molecular characterisation and infectivity of a "Legumovirus" (genus Begomovirus: family Geminiviridae) infecting the leguminous weed Rhynchosia minima in Pakistan. Virus Res 2009, 145, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Tan, H.; Zhao, S.; Li, H.; Liu, H.; Ma, Y.; Zhang, X.; Rong, J.; Fu, X.; Lozano-Durán, R.; et al. Geminiviruses encode additional small proteins with specific subcellular localizations and virulence function. Nat Commun 2021, 12, 4278. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Durán, R.; Rosas-Díaz, T.; Luna, A.P.; Bejarano, E.R. Identification of host genes involved in geminivirus infection using a reverse genetics approach. PLoS One 2011, 6, e22383. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lozano-Durán, R. Manipulation of plant RNA biology by geminiviruses. J Exp Bot 2023, 74, 2311–2322. [Google Scholar] [CrossRef]
- Mahmood, M.A.; Naqvi, R.Z.; Amin, I.; Mansoor, S. Salicylic acid-driven innate antiviral immunity in plants. Trends Plant Sci 2024, 29, 715–717. [Google Scholar] [CrossRef]
- Settlage, S.B.; See, R.G.; Hanley-Bowdoin, L. Geminivirus C3 protein: replication enhancement and protein interactions. J Virol 2005, 79, 9885–9895. [Google Scholar] [CrossRef]
- Gafni, Y.; Epel, B.L. The role of host and viral proteins in intra-and inter-cellular trafficking of geminiviruses. Physiol Mol Plant Pathol 2002, 60, 231–241. [Google Scholar] [CrossRef]
- Neubauer, M.; Vollen, K.; Ascencio-Ibanez, J.T.; Hanley-Bowdoin, L.; Stepanova, A.N.; Alonso, J.M. Development of modular geminivirus-based vectors for high cargo expression and gene targeting in plants. BioRxiv, 2024. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Symington, L.S.; Gautier, J. Double-strand break end resection and repair pathway choice. Annu Rev Genet 2011, 45, 247–271. [Google Scholar] [CrossRef]
- Mahmood, M.A.; Naqvi, R.Z.; Mansoor, S. Engineering crop resistance by manipulating disease susceptibility genes. Mol Plant 2022, 15, 1511–1513. [Google Scholar] [CrossRef]
- Mahmood, M.A.; Greenwood, J.R. A prime example of precisely delivered DNA. Trends Genet 2023, 39, 717–718. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, M.A.; Mansoor, S. PASTE: The Way Forward for Large DNA Insertions. Crispr j 2023, 6, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Awan, M.J.A.; Mahmood, M.A.; Naqvi, R.Z.; Mansoor, S. PASTE: a high-throughput method for large DNA insertions. Trends Plant Sci 2023, 28, 509–511. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Du, J.; Yun, S.; Xue, C.; Yao, Y.; Rao, S. Recent advances in CRISPR-Cas9-based genome insertion technologies. Mol Ther Nucleic Acids 2024, 35, 102138. [Google Scholar] [CrossRef]
- Zaidi, S.S.; Mansoor, S. Viral Vectors for Plant Genome Engineering. Front Plant Sci 2017, 8, 539. [Google Scholar] [CrossRef]
- Wang, M.; Lu, Y.; Botella, J.R.; Mao, Y.; Hua, K.; Zhu, J.K. Gene Targeting by Homology-Directed Repair in Rice Using a Geminivirus-Based CRISPR/Cas9 System. Mol Plant 2017, 10, 1007–1010. [Google Scholar] [CrossRef]
- Mor, T.S.; Moon, Y.S.; Palmer, K.E.; Mason, H.S. Geminivirus vectors for high-level expression of foreign proteins in plant cells. Biotechnol Bioeng 2003, 81, 430–437. [Google Scholar] [CrossRef]
- Gleba, Y.; Marillonnet, S.; Klimyuk, V. Engineering viral expression vectors for plants: the 'full virus' and the 'deconstructed virus' strategies. Curr Opin Plant Biol 2004, 7, 182–188. [Google Scholar] [CrossRef]
- Gardiner, W.E.; Sunter, G.; Brand, L.; Elmer, J.S.; Rogers, S.G.; Bisaro, D.M. Genetic analysis of tomato golden mosaic virus: the coat protein is not required for systemic spread or symptom development. Embo j 1988, 7, 899–904. [Google Scholar] [CrossRef]
- Hayes, R.J.; Petty, I.T.D.; Coutts, R.H.A.; Buck, K.W. Gene amplification and expression in plants by a replicating geminivirus vector. Nature 1988, 334, 179–182. [Google Scholar] [CrossRef]
- Pooma, W.; Gillette, W.K.; Jeffrey, J.L.; Petty, I.T. Host and viral factors determine the dispensability of coat protein for bipartite geminivirus systemic movement. Virology 1996, 218, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.; Etessami, P.; Stanley, J. Expression of a bacterial gene in plants mediated by infectious geminivirus DNA. Embo j 1988, 7, 1583–1587. [Google Scholar] [CrossRef]
- Muangsan, N.; Robertson, D. Geminivirus vectors for transient gene silencing in plants. Methods Mol Biol 2004, 265, 101–115. [Google Scholar] [CrossRef]
- Gilbertson, R.L.; Sudarshana, M.; Jiang, H.; Rojas, M.R.; Lucas, W.J. Limitations on geminivirus genome size imposed by plasmodesmata and virus-encoded movement protein: insights into DNA trafficking. Plant Cell 2003, 15, 2578–2591. [Google Scholar] [CrossRef]
- Yin, K.; Han, T.; Liu, G.; Chen, T.; Wang, Y.; Yu, A.Y.; Liu, Y. A geminivirus-based guide RNA delivery system for CRISPR/Cas9 mediated plant genome editing. Sci Rep 2015, 5, 14926. [Google Scholar] [CrossRef]
- Eini, O.; Schumann, N.; Niessen, M.; Varrelmann, M. Targeted mutagenesis in plants using Beet curly top virus for efficient delivery of CRISPR/Cas12a components. N Biotechnol 2022, 67, 1–11. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, X.; Sun, H.; Liang, Q.; Wang, W.; Zhang, C.; Bian, X.; Cao, Q.; Li, Q.; Xie, Y.; et al. Improving CRISPR-Cas-mediated RNA targeting and gene editing using SPLCV replicon-based expression vectors in Nicotiana benthamiana. Plant Biotechnol J 2020, 18, 1993–1995. [Google Scholar] [CrossRef]
- Sarrion-Perdigones, A.; Falconi, E.E.; Zandalinas, S.I.; Juárez, P.; Fernández-del-Carmen, A.; Granell, A.; Orzaez, D. GoldenBraid: an iterative cloning system for standardized assembly of reusable genetic modules. PLoS One 2011, 6, e21622. [Google Scholar] [CrossRef]
- Neubauer, M.; Vollen, K.; Ascencio-Ibanez, J.T.; Hanley-Bowdoin, L.; Stepanova, A.N.; Alonso, J.M. Development of modular geminivirus-based vectors for high cargo expression and gene targeting in plants. bioRxiv, 2024. [Google Scholar] [CrossRef]
- Naqvi, R.Z.; Siddiqui, H.A.; Mahmood, M.A.; Najeebullah, S.; Ehsan, A.; Azhar, M.; Farooq, M.; Amin, I.; Asad, S.; Mukhtar, Z.; et al. Smart breeding approaches in post-genomics era for developing climate-resilient food crops. Front Plant Sci 2022, 13, 972164. [Google Scholar] [CrossRef]
- Naqvi, R.Z.; Mahmood, M.A.; Mansoor, S.; Amin, I.; Asif, M. Omics-driven exploration and mining of key functional genes for the improvement of food and fiber crops. Front Plant Sci 2023, 14, 1273859. [Google Scholar] [CrossRef]
- Vu, T.V.; Nguyen, N.T.; Kim, J.; Song, Y.J.; Nguyen, T.H.; Kim, J.Y. Optimized dicot prime editing enables heritable desired edits in tomato and Arabidopsis. Nat Plants 2024, 10, 1502–1513. [Google Scholar] [CrossRef] [PubMed]
- Butler, N.M.; Baltes, N.J.; Voytas, D.F.; Douches, D.S. Geminivirus-Mediated Genome Editing in Potato (Solanum tuberosum L.) Using Sequence-Specific Nucleases. Front Plant Sci 2016, 7, 1045. [Google Scholar] [CrossRef] [PubMed]
- Hummel, A.W.; Chauhan, R.D.; Cermak, T.; Mutka, A.M.; Vijayaraghavan, A.; Boyher, A.; Starker, C.G.; Bart, R.; Voytas, D.F.; Taylor, N.J. Allele exchange at the EPSPS locus confers glyphosate tolerance in cassava. Plant Biotechnol J 2018, 16, 1275–1282. [Google Scholar] [CrossRef]
- Peyret, H.; Lomonossoff, G.P. When plant virology met Agrobacterium: the rise of the deconstructed clones. Plant Biotechnol J 2015, 13, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, M.; Wang, X.; Zhang, Y.J.; Da, X.W.; Jia, L.Y.; Pang, H.L.; Feng, H.Q. Effects of plant growth regulators on transient expression of foreign gene in Nicotiana benthamiana L. leaves. Bioresour Bioprocess 2021, 8, 124. [Google Scholar] [CrossRef]
- Butler, N.M.; Atkins, P.A.; Voytas, D.F.; Douches, D.S. Generation and Inheritance of Targeted Mutations in Potato (Solanum tuberosum L.) Using the CRISPR/Cas System. PLoS One 2015, 10, e0144591. [Google Scholar] [CrossRef]
- Dahan-Meir, T.; Filler-Hayut, S.; Melamed-Bessudo, C.; Bocobza, S.; Czosnek, H.; Aharoni, A.; Levy, A.A. Efficient in planta gene targeting in tomato using geminiviral replicons and the CRISPR/Cas9 system. Plant J 2018, 95, 5–16. [Google Scholar] [CrossRef]
- Kim, E.; Koo, T.; Park, S.W.; Kim, D.; Kim, K.; Cho, H.-Y.; Song, D.W.; Lee, K.J.; Jung, M.H.; Kim, S. In vivo genome editing with a small Cas9 orthologue derived from Campylobacter jejuni. Nat Commun 2017, 8, 14500. [Google Scholar] [CrossRef]
- Pausch, P.; Al-Shayeb, B.; Bisom-Rapp, E.; Tsuchida, C.A.; Li, Z.; Cress, B.F.; Knott, G.J.; Jacobsen, S.E.; Banfield, J.F.; Doudna, J.A. CRISPR-CasΦ from huge phages is a hypercompact genome editor. Science 2020, 369, 333–337. [Google Scholar] [CrossRef]
- Rauch, B.J.; DeLoughery, A.; Sper, R.; Chen, S.; Yunanda, S.; Masnaghetti, M.; Chai, N.; Lin, J.C.; Neckelmann, A.; Bjornson, Y.; et al. Single-AAV CRISPR editing of skeletal muscle in non-human primates with NanoCas, an ultracompact nuclease. bioRxiv, 2025. [Google Scholar] [CrossRef]
- Ruhel, R.; Chakraborty, S. Multifunctional roles of geminivirus encoded replication initiator protein. Virusdisease 2019, 30, 66–73. [Google Scholar] [CrossRef]
- Ellison, E.E.; Nagalakshmi, U.; Gamo, M.E.; Huang, P.J.; Dinesh-Kumar, S.; Voytas, D.F. Multiplexed heritable gene editing using RNA viruses and mobile single guide RNAs. Nat Plants 2020, 6, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Uranga, M.; Aragonés, V.; Selma, S.; Vázquez-Vilar, M.; Orzáez, D.; Daròs, J.A. Efficient Cas9 multiplex editing using unspaced sgRNA arrays engineering in a Potato virus X vector. Plant J 2021, 106, 555–565. [Google Scholar] [CrossRef]
- Uranga, M.; Aragonés, V.; García, A.; Mirabel, S.; Gianoglio, S.; Presa, S.; Granell, A.; Pasin, F.; Daròs, J.A. RNA virus-mediated gene editing for tomato trait breeding. Hortic Res 2024, 11, uhad279. [Google Scholar] [CrossRef] [PubMed]
- Ali, Z.; Eid, A.; Ali, S.; Mahfouz, M.M. Pea early-browning virus-mediated genome editing via the CRISPR/Cas9 system in Nicotiana benthamiana and Arabidopsis. Virus Res 2018, 244, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Li, S.; Li, Z.; Li, H.; Song, W.; Zhao, H.; Lai, J.; Xia, L.; Li, D.; Zhang, Y. A barley stripe mosaic virus-based guide RNA delivery system for targeted mutagenesis in wheat and maize. Mol Plant Pathol 2019, 20, 1463–1474. [Google Scholar] [CrossRef]
- Li, T.; Hu, J.; Sun, Y.; Li, B.; Zhang, D.; Li, W.; Liu, J.; Li, D.; Gao, C.; Zhang, Y.; et al. Highly efficient heritable genome editing in wheat using an RNA virus and bypassing tissue culture. Mol Plant 2021, 14, 1787–1798. [Google Scholar] [CrossRef]
- Wang, W.; Yu, Z.; He, F.; Bai, G.; Trick, H.N.; Akhunova, A.; Akhunov, E. Multiplexed promoter and gene editing in wheat using a virus-based guide RNA delivery system. Plant Biotechnol J 2022, 20, 2332–2341. [Google Scholar] [CrossRef]
- Tamilselvan-Nattar-Amutha, S.; Hiekel, S.; Hartmann, F.; Lorenz, J.; Dabhi, R.V.; Dreissig, S.; Hensel, G.; Kumlehn, J.; Heckmann, S. Barley stripe mosaic virus-mediated somatic and heritable gene editing in barley (Hordeum vulgare L.). Front Plant Sci 2023, 14, 1201446. [Google Scholar] [CrossRef]
- Zhang, X.; Kang, L.; Zhang, Q.; Meng, Q.; Pan, Y.; Yu, Z.; Shi, N.; Jackson, S.; Zhang, X.; Wang, H.; et al. An RNAi suppressor activates in planta virus-mediated gene editing. Funct Integr Genomics 2020, 20, 471–477. [Google Scholar] [CrossRef]
- Baysal, C.; Kausch, A.P.; Cody, J.P.; Altpeter, F.; Voytas, D.F. Rapid and efficient in planta genome editing in sorghum using foxtail mosaic virus-mediated sgRNA delivery. Plant J 2025, 121, e17196. [Google Scholar] [CrossRef]
- Cody, W.B.; Scholthof, H.B.; Mirkov, T.E. Multiplexed Gene Editing and Protein Overexpression Using a Tobacco mosaic virus Viral Vector. Plant Physiol 2017, 175, 23–35. [Google Scholar] [CrossRef]
- Jiang, N.; Zhang, C.; Liu, J.Y.; Guo, Z.H.; Zhang, Z.Y.; Han, C.G.; Wang, Y. Development of Beet necrotic yellow vein virus-based vectors for multiple-gene expression and guide RNA delivery in plant genome editing. Plant Biotechnol J 2019, 17, 1302–1315. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Yang, J.; Gu, Y.; Wang, Q.; Zhang, Z.; Guo, H.; Zhao, L.; Zhang, H.; Gu, L. Bamboo mosaic virus-mediated transgene-free genome editing in bamboo. New Phytol 2025, 245, 1810–1816. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Wang, D.; Wang, H.; Sun, Y.; Yin, C.; Li, J.; Ye, J. Development and application of sugarcane streak mosaic virus vectors. Virology 2024, 593, 110028. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, X.; Liu, H.; Li, Z. Highly efficient DNA-free plant genome editing using virally delivered CRISPR-Cas9. Nat Plants 2020, 6, 773–779. [Google Scholar] [CrossRef]
- Zhao, C.; Lou, H.; Liu, Q.; Pei, S.; Liao, Q.; Li, Z. Efficient and transformation-free genome editing in pepper enabled by RNA virus-mediated delivery of CRISPR/Cas9. J Integr Plant Biol 2024, 66, 2079–2082. [Google Scholar] [CrossRef]
- Gao, Q.; Xu, W.Y.; Yan, T.; Fang, X.D.; Cao, Q.; Zhang, Z.J.; Ding, Z.H.; Wang, Y.; Wang, X.B. Rescue of a plant cytorhabdovirus as versatile expression platforms for planthopper and cereal genomic studies. New Phytol 2019, 223, 2120–2133. [Google Scholar] [CrossRef]
- Liu, D.; Ellison, E.E.; Myers, E.A.; Donahue, L.I.; Xuan, S.; Swanson, R.; Qi, S.; Prichard, L.E.; Starker, C.G.; Voytas, D.F. Heritable gene editing in tomato through viral delivery of isopentenyl transferase and single-guide RNAs to latent axillary meristematic cells. Proc Natl Acad Sci U S A 2024, 121, e2406486121. [Google Scholar] [CrossRef]
- Bouton, C.; King, R.C.; Chen, H.; Azhakanandam, K.; Bieri, S.; Hammond-Kosack, K.E.; Kanyuka, K. Foxtail mosaic virus: A Viral Vector for Protein Expression in Cereals. Plant Physiol 2018, 177, 1352–1367. [Google Scholar] [CrossRef]
- Bredow, M.; Natukunda, M.I.; Beernink, B.M.; Chicowski, A.S.; Salas-Fernandez, M.G.; Whitham, S.A. Characterization of a foxtail mosaic virus vector for gene silencing and analysis of innate immune responses in Sorghum bicolor. Mol Plant Pathol 2023, 24, 71–79. [Google Scholar] [CrossRef]
- Chen, W.; Huang, C.; Luo, C.; Zhang, Y.; Zhang, B.; Xie, Z.; Hao, M.; Ling, H.; Cao, G.; Tian, B.; et al. A New Method for Rapid Subcellular Localization and Gene Function Analysis in Cotton Based on Barley Stripe Mosaic Virus. Plants (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.S.; Schneider, W.L.; Chaluvadi, S.R.; Mian, M.A.; Nelson, R.S. Characterization of a Brome mosaic virus strain and its use as a vector for gene silencing in monocotyledonous hosts. Mol Plant Microbe Interact 2006, 19, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.H.; Lu, H.C.; Pan, Z.J.; Yeh, H.H.; Wang, S.S.; Chen, W.H.; Chen, H.H. Optimizing virus-induced gene silencing efficiency with Cymbidium mosaic virus in Phalaenopsis flower. Plant Sci 2013, 201-202, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Liou, M.R.; Huang, Y.W.; Hu, C.C.; Lin, N.S.; Hsu, Y.H. A dual gene-silencing vector system for monocot and dicot plants. Plant Biotechnol J 2014, 12, 330–343. [Google Scholar] [CrossRef]
- Globus, R.; Qimron, U. A technological and regulatory outlook on CRISPR crop editing. J Cell Biochem 2018, 119, 1291–1298. [Google Scholar] [CrossRef]
- McGarry, R.C.; Klocko, A.L.; Pang, M.; Strauss, S.H.; Ayre, B.G. Virus-Induced Flowering: An Application of Reproductive Biology to Benefit Plant Research and Breeding. Plant Physiol 2017, 173, 47–55. [Google Scholar] [CrossRef]
- Liou, M.-R.; Hu, C.-C.; Lin, N.-S.; Hsu, Y.-H. Development and application of satellite-based vectors. In Viroids and Satellites; Elsevier: 2017; pp. 597-604.
- Wunderlich, S.; Gatto, K.A. Consumer perception of genetically modified organisms and sources of information. Adv Nutr 2015, 6, 842–851. [Google Scholar] [CrossRef]
- Ewa, W.G.; Agata, T.; Milica, P.; Anna, B.; Dennis, E.; Nick, V.; Godelieve, G.; Selim, C.; Naghmeh, A.; Tomasz, T. Public perception of plant gene technologies worldwide in the light of food security. GM Crops Food 2022, 13, 218–241. [Google Scholar] [CrossRef]
Figure 1.
Genomic organization of Begomoviruses, Curtoviruses and Mastreviruses. Created in https://BioRender.com. .
Figure 1.
Genomic organization of Begomoviruses, Curtoviruses and Mastreviruses. Created in https://BioRender.com. .
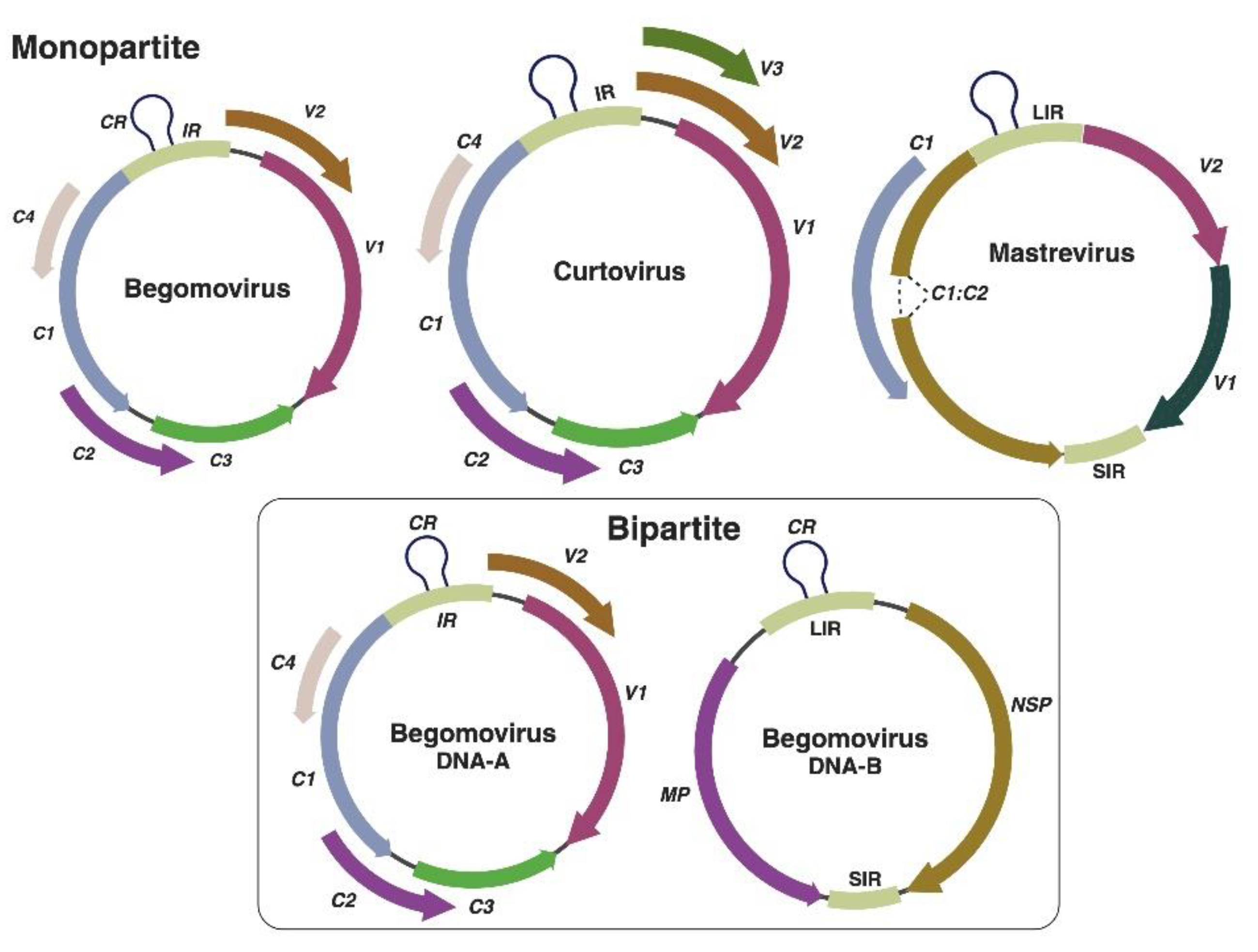
Figure 2.
CRISPR-Cas9 system and their types. The basic working principle of CRISPR-Cas9 system. This system works on a simple DNA-RNA base pairing, the PAM sequence and target specific gene precisely. Upon targeting, it results in DNA DSBs, which will be repaired either with NHEJ which is an error-prone process and results in random indels and gene disruption at the cleavage site (termed as SDN-1 process) or with HDR which precisely repair the DSBs. HDR process can be harnessed to integrate specific DNA template at the cleavage site (termed as SDN-2 and 3). The donor template could be several hundreds to thousands base pair long. Created in https://BioRender.com.
Figure 2.
CRISPR-Cas9 system and their types. The basic working principle of CRISPR-Cas9 system. This system works on a simple DNA-RNA base pairing, the PAM sequence and target specific gene precisely. Upon targeting, it results in DNA DSBs, which will be repaired either with NHEJ which is an error-prone process and results in random indels and gene disruption at the cleavage site (termed as SDN-1 process) or with HDR which precisely repair the DSBs. HDR process can be harnessed to integrate specific DNA template at the cleavage site (termed as SDN-2 and 3). The donor template could be several hundreds to thousands base pair long. Created in https://BioRender.com.
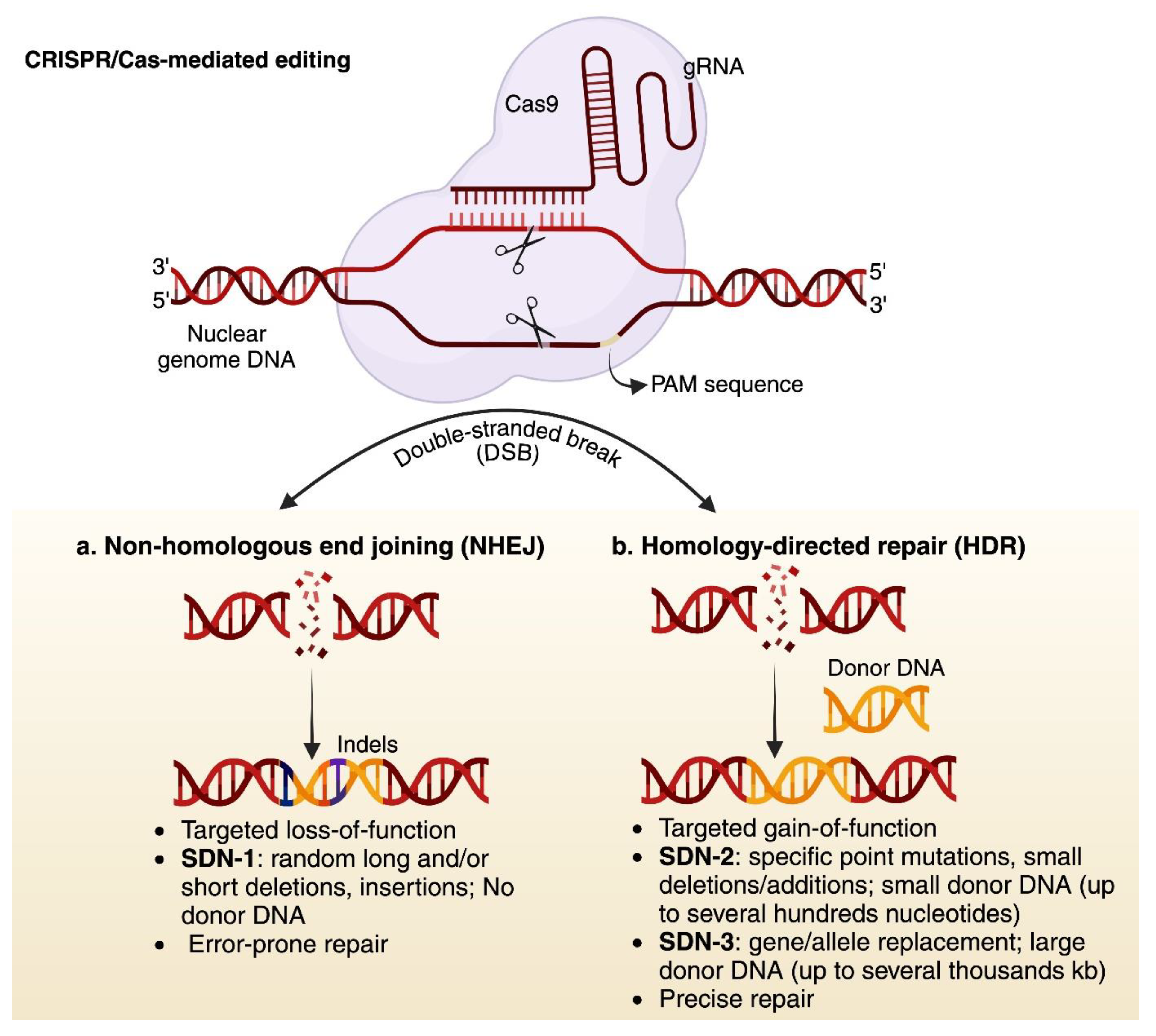
Figure 3.
Geminivirus-based replicons structure. a) Basic geminiviral-based replicon mechanism. The gene of interest is flanked by two direct IRs. Upon expression of Rep, episomal replicons will be generated. Thus, accumulating to high levels in the cell. b) Geminiviral replicons structures for genome editing. Three version of modified cassettes have been shown here. In A), the Cas9, donor DNA and Rep protein are included as part of the replicon; in B), the donor DNA and Rep protein are part of the replicon while Cas9 is outside the replicon and will only be expressed from T-DNA; and in C), only the donor DNA is part of the replicon while the Cas9 and Rep protein are outside the replicon. Note that the Rep protein does not require to be the part of same T-DNA but can also be provided independently. Replicons such as A) and B) have been successfully utilized in genome editing [15]. Created in https://BioRender.com. .
Figure 3.
Geminivirus-based replicons structure. a) Basic geminiviral-based replicon mechanism. The gene of interest is flanked by two direct IRs. Upon expression of Rep, episomal replicons will be generated. Thus, accumulating to high levels in the cell. b) Geminiviral replicons structures for genome editing. Three version of modified cassettes have been shown here. In A), the Cas9, donor DNA and Rep protein are included as part of the replicon; in B), the donor DNA and Rep protein are part of the replicon while Cas9 is outside the replicon and will only be expressed from T-DNA; and in C), only the donor DNA is part of the replicon while the Cas9 and Rep protein are outside the replicon. Note that the Rep protein does not require to be the part of same T-DNA but can also be provided independently. Replicons such as A) and B) have been successfully utilized in genome editing [15]. Created in https://BioRender.com. .
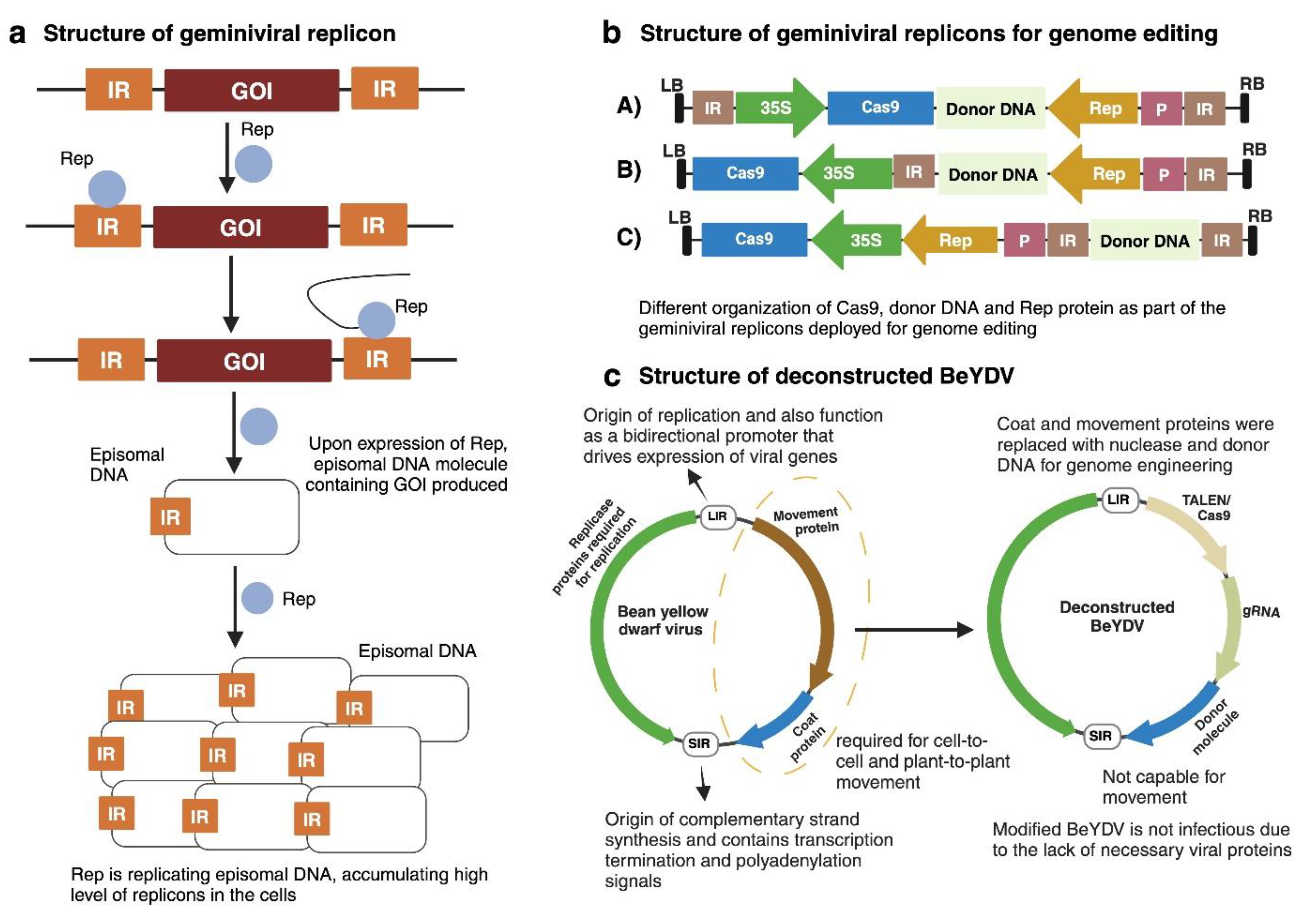
Figure 4.
Illustration of geminivirus-mediated HDR genome editing. The geminiviral genome containing cas9, gRNA and donor DNA is cloned into the transfer DNA (T-DNA) vector. To ensure the release of T-DNA in the plant cell, one LIR is integrated on each side of the viral genome. During Agrobacterium infection, the T-DNA molecules are transferred to the plant cell nucleus where the viral genome (circular) amplified into thousands of copies via rolling circle replication driven by replicase proteins. On the other side, the complex of gRNA and Cas9 induces DSBs at the target locus and the donor DNA is integrated into the target site via homology-directed repair (HDR). The high copy number of donor DNA increases the gene targeting frequency. Created in https://BioRender.com. .
Figure 4.
Illustration of geminivirus-mediated HDR genome editing. The geminiviral genome containing cas9, gRNA and donor DNA is cloned into the transfer DNA (T-DNA) vector. To ensure the release of T-DNA in the plant cell, one LIR is integrated on each side of the viral genome. During Agrobacterium infection, the T-DNA molecules are transferred to the plant cell nucleus where the viral genome (circular) amplified into thousands of copies via rolling circle replication driven by replicase proteins. On the other side, the complex of gRNA and Cas9 induces DSBs at the target locus and the donor DNA is integrated into the target site via homology-directed repair (HDR). The high copy number of donor DNA increases the gene targeting frequency. Created in https://BioRender.com. .
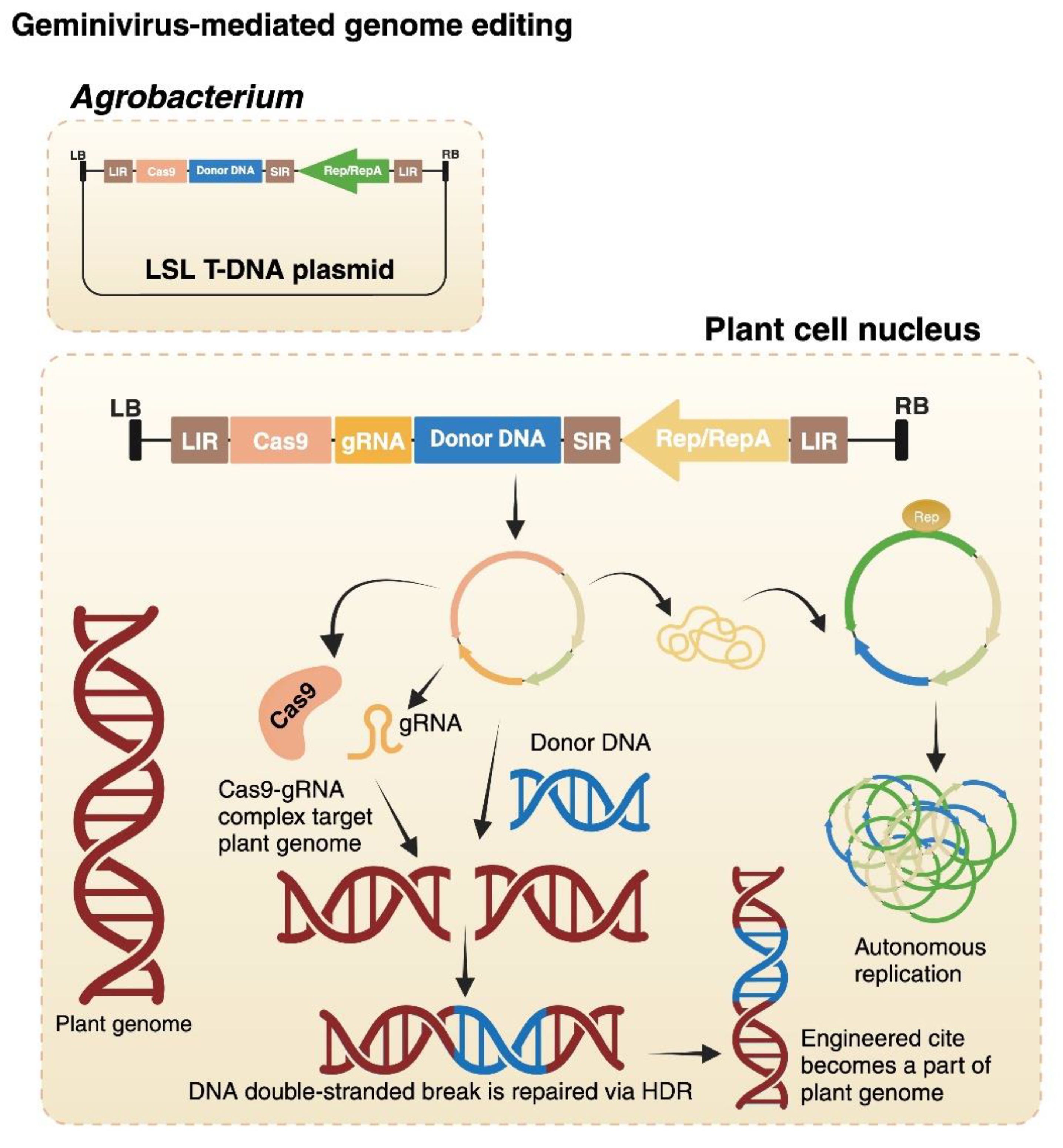

Table 1.
List of geminivirus-based vectors used in genome editing.
| Viruses | Genus | GE platform | gRNA type | Plant species | Targeted gene* | Heritability | Reference |
|---|---|---|---|---|---|---|---|
| BeYDV | Mastrevirus | CRISPR-Cas9 | AtU6-gRNA | Solanum tuberosum | StALS1 | Yes | [59] |
| TALEN and CRISPR-Cas9 | AtU6-gRNA | S. lycopersicum | ALS1 | No | [55] | ||
| ZFN, TALEN and CRISPR-Cas9 | AtU6-gRNA | N. tabacum | ALS and P-GUS: NPTII | No | [15] | ||
| TALEN and CRISPR-Cas9 | AtU6-gRNAs | Tomato cv. MicroTom | ANT1 | Yes | [14] | ||
| CRISPR-Cas9 | AtU6-gRNAs | Lycopersicon esculentum | CRTISO and PSY1 | Yes | [60] | ||
| WDV | CRISPR-Cas9 | TaU6-gRNA | Triticum aestivum | Ubi, MLO, and GFP | No | [13] | |
| Cas9 expressing rice | OsU6-gRNAs | Oryza sativa | ACT1 and GST | No | [38] | ||
| CaLCuV | Begomovirus | Cas9 expressing tobacco | AtU6-gRNA | N. benthamiana | NbPDS3 and NbIspH | No | [47] |
| SPLCV | LwaCas13, LbCas12a and Cas9 | AtU6-gRNA | N. benthamiana | NbPDS1 and mGFP5 | No | [49] | |
| BCTV | Curtovirus | CRISPR/Cas12a | - | N. benthamiana | NbGFP | Yes | [48] |
*Solanum tuberosum Acetolactate synthase 1 (StALS1), Acetolactate synthase 1 (ALS1), promoter of GUS and neomycin phosphotransferase II (P-GUS: NPTII), Anthocyanin mutant 1 (ANT1), Carotenoid isomerase (CRITSO), Phytoene synthase 1 (PSY1) ubiquitin (Ubi), Mildew Locus O (MLO), Green fluorescent protein (GFP), Actin-1 (ACT1), Glutathione S-transferase (GST), Nicotiana benthamiana Phytoene desaturase 3 (NbPDS3), Nicotiana benthamiana isopentenyl/dimethylallyl diphosphate synthase (IDDS). .
Table 2.
List of RNA viruses-based vectors used in genome editing.
| Virus | Plant species | Targeted gene | Component delivered | Heritability | Reference |
|---|---|---|---|---|---|
| Positive-sense RNA virus | |||||
| TRV | N. benthamiana |
PDS, AG (gRNA fusion with FT/tRNA) |
gRNA | Yes | [65] |
| PVX | N. benthamiana | NbXT2B | sgRNA | Yes | [66] |
| Solanum lycopersicum | PDS and SGR | sgRNA | Yes | [67] | |
| TRV and PEBV | N. benthamiana and A. thaliana | GL1, TT4 and NbPDS | sgRNAs | No | [68] |
| BSMV | N. benthamiana, Triticum aestivum and Zea mays | NbPDS, mGFP5, TaGASR7 and ZmTMS5 | gRNA | - | [69] |
| Triticum aestivum | TaPDS, TaGW2 and TaGASR7 | sgRNAs | Yes | [70,71] | |
| Hordeum vulgare | ALBOSTRIANS, ASY1, MUS81 and ZYP1 | sgRNA | Yes | [72] | |
| FoMV | N. benthamiana | NbPDS | Cas9, sgRNA, RNAi suppressor p19 | No | [73] |
| Sorghum bicolor | MgCh, Lw1 and PDS | sgRNAs and AmCyan fluorescent protein | Yes | [74] | |
| TMV | N. benthamiana | NbAGO1 | sgRNAs | No | [75] |
| BNYVV | N. benthamiana and Beta vulgaris | NbPDS3 | gRNAs, mCherry, eCFP-HA, sGFP, GUS | No | [76] |
| BaMV | N. benthamiana, Dendrocalamus latiflorus Munro and phyllostachys edulis | NbPDS6, NbPDS, PheRDR6 and DlmRDR6 | Ascas12f1-YHAM, Ascas12f1-HKRA Cas9 and sgRNA | No | [77] |
| SCSMV | N. benthamiana and Saccharum spp. | - | EGFP | - | [78] |
| Negative-strand RNA virus | |||||
| SYNV | N. benthamiana | GFP, PDS, RDR6 and SG3 | SpCas9 and gRNA | Yes | [79] |
| TSWV | N. benthamiana, C. annuum | PDS3 and PDS4 | Cas9, Cas12a, adenine and cytosine base editors | Yes | [80] |
| BYSMV | N. benthamiana, Laodelphax striatellus, Barley, wheat, maize and foxtail millet | GFP | GUS, RFP and SpCas9 and gRNA | No | [81] |
* PDS and AG: Phytoene Desaturase and AGAMOUS; GL1 and TT4: Arabidopsis GLABRA1 and TRANSPARENT TESTA GLABRA4; NbPDS: N. benthamiana Phytoene Desaturase; RDR6: RNA-dependent RNA polymerase 6; SG3: Suppressor Gene Silencing 3; ZmTM5: Thermosensitive genic male sterile 5; GFP: Green fluorescent protein; SGR: STAYGREEN; MgCh: Magnesium chelatase; Lw1: Lemon white 1; GUS and RFP: β-glucuronidase and red fluorescent protein.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



