
Submitted:
19 March 2025
Posted:
20 March 2025
You are already at the latest version
Abstract
Host genetic variability is relevant to understanding how parasites modulate natural selection in wild fish populations. Coastal lagoons are ‘transitional’ ecosystems where knowledge lacks on relationships between genotypic diversity with parasitism. The aim of this study was to assess the effect of genetic diversity on host health and parasitological traits in fish inhabiting a Mediterranean lagoon. Black-striped pipefish Syngnathus abaster were collected in August 2023–24 from the ‘Mar Menor’ (Iberian lagoon, SE Spain). Genetic diversity was measured as Internal Relatedness (IR: a homozygosity index from microsatellite markers). Population frequency was lower for the medium IR level. For this same category, both health indices (external body condition and internal organs) indicated a worse status. Parasite prevalence, abundance and an index of life-cycle complexity (heteroxenous species) were greater for the medium level of genetic diversity. Such results are explained under a scenario of parasite-mediated disruptive selection: a higher disease pressure against the phenotypically ‘intermediate’ individuals. Two contrasting strategies were detected to better control parasitism at the host genotypic level: 1) high homozygosity, 2) high het-erozygosity; which are probably reflecting better immuno-competence as a phenotypic trait. From an evolutionary perspective, parasites play a crucial role in shaping genetic diversity within host populations.
Keywords:
black-striped pipefish
; body condition
; disruptive selection
; Iberian Peninsula
; life-cycle complexity
; Mediterranean Sea
1. Introduction
Parasites can exert a strong selective pressure on host populations, from invertebrates to mammals, with profound consequences for the entire ecosystem and evolutionary processes [1,2,3]. This is because of many parasite species (diseases) can deeply impact health, disturbing a wide variety of traits related to host life-cycles, such as growth rate, reproduction, behaviour or feeding habits [1,4,5,6]. Ultimately, parasites displaying an elevated pathogenic nature may massively decrease fitness until high levels of mortality are observed within the host populations (see a dramatic case reaching up to 100% in some sites for the pen shell Pinna nobilis; [7]). In general, parasitism is an ecological relationship widely spread across the globe that usually does not lead to such extreme consequences (i.e. mass mortalities; [8]). Thus, data on physical condition, fitness or health status in hosts constitutes valuable information in order to truly assess the sub-lethal effects of this biotic interaction at the population and ecosystem levels [9,10,11].
Among vertebrate hosts, fish are pivotal communities for most aquatic ecosystems all around the world, with many stocks being of great interest for exploitation [12]. Given that wild fish are host to a wide diversity of parasite taxa (>30k helminth species; see [13]), these communities can be considered as good biological models to investigate particular ecological/evolutionary patterns from such intimate relationships [14,15,16]. As an example of a ‘simpler’ interaction, monogeneans are monoxenous gill infesting ecto-parasites through the branchial chamber [16]. More complex is the case of digeneans, which are heteroxenous parasites that typically include a variety of invertebrate and vertebrate hosts (e.g. molluscs, crustaceans, fish, birds or even aquatic mammals) in their life cycle, as well as having free-swimming stages (larvae) during their transmission between hosts [14,17]. Thus, variations in abundances of particular parasite communities may be a result of changes in structure and dynamics at the ecosystem level [15,18].
The genetic ‘aspect’ of this biotic interaction (i.e. parasitism) is of particular relevance for research on ecology and evolution, as previous studies have demonstrated genetically-based variations for key traits affecting this relationship, from both the parasite and the host perspectives (see comprehensive reviews on this topic in [19,20]). Focusing specifically on animal health, hosts have evolved/developed different traits (or strategies with their particular genetic basis) to maintain fitness when interacting with parasites. More specifically, physiological/genetic mechanisms are usually related to: 1) the defense against pathogens/parasites (i.e. immune response to avoid establishment or reduce burden), 2) the rapid body repair from the damage caused by infections (i.e. cell/tissue regeneration) [21,22]. Moreover, the link between the host genotype and the level of parasite infection appears to be highly variable. Indeed, the effect of genetic traits can be contrasting at the host population level, in terms of parasites mediating natural (either balancing, stabilizing, directional or disruptive) selection [23]. According to the ‘inbreeding depression’ hypothesis, a significant negative relationship is expected to be detected between host heterozygosity level and parasite burden [24]. This may be indicative that parasites contribute to modulate genetic diversity in their host populations through a combination of balancing and directional (positive) selection for the most heterozygous (i.e. genotypically diverse) individuals (e.g. see Zueva et al. [25] for the relationship between the Atlantic salmon Salmo salar and the ecto-parasite monogenean Gyrodactylus salaris). However, parasite-mediated disruptive selection can also occur, as it was evidenced by Blanchet et al. [26,27] in a wild population of common dace Leuciscus leuciscus and its ecto-parasite copepod Tracheliastes polycolpus. In these ‘dace’ studies, parasite burden was lower for fish individuals displaying both extremely heterozygous and homozygous levels (i.e. selective pressure against phenotypically ‘intermediate’ individuals; [23]), with global genetic variance (diversity) of this population being broadening, accordingly. Therefore, different and alternative parasite-mediated mechanisms are possible to ‘shape’ genetic diversity at the host population level. Clearly, there is a need for more research that provides insights into the relationships between genetic traits of hosts and their parasite infra-communities, particularly for fish.
In terms of applications, basic knowledge on genetic traits of wild parasitised fish could be interesting for aquaculture. Given the collapse of many extractive fisheries at the global scale [28], aquaculture is regarded as a feasible solution to provide finfish products, i.e. a source of high-quality protein (around one third of the current total consumption; see [28,29]). The world-wide development of this industry has led to an increasing awareness of the necessity to understand the genetic causes and consequences of parasite infections [30]. This is because many parasite species can result in harmful diseases and hence provoke major mortality outbreaks in farmed fish [31,32]. In fact, heritable genetic components can be identified to develop disease-resistant genetic lines tailored to particular target fish species through artificial selection [30]. Such programs of selective breeding (supported by genetic/genomic information) also contribute positively to the overall productivity and economic performance of aquaculture activities [33].
Coastal lagoons are considered as ‘transitional’ water bodies, located between flowing fresh waters (i.e. rivers, streams) and marine (salt-water) environments. These distinctive habitats have received relatively less attention than ‘pure’ freshwater or marine ecosystems [34,35]. Regarding fish parasites in transitional waters, Giari et al. [36] reviewed the relevance of such communities for shaping structure/function within these ecosystems. Almeida et al. [37] have also provided valuable information on fish parasites from a Mediterranean coastal lagoon, although this study was focused on conservation purposes for the Iberian eco-region (parasites as ‘ecological indicators’ of habitat quality). Nonetheless, genetic data are still very scarce for these aquatic environments, beyond the use for mere identification (but see Correia et al. [38] and Farjallah et al. [39] about parasite diversity). To the best of the author’s knowledge, information about diversity of fish genotypes related to their parasites has been virtually undescribed in coastal lagoons. Consequently, the assessment of these (genetic and parasitological) traits within resident populations of fish hosts, along with their health status, have a great potential to more effectively reveal ecological and evolutionary implications under these particular (transitional) environmental conditions. However, this potential is severely under-developed.
In light of this dearth of information, the aim of the present study was to assess the relationships among genetic traits, health status of fish hosts and their parasite infra-communities in a wild population inhabiting an Iberian coastal lagoon (Mediterranean Sea). Specifically, a variety of parameters were compared across host categories exhibiting different levels of genetic diversity: population frequencies and external/internal health indices (calculated from fish data); and prevalence, taxonomic composition, total abundance and an index of life-cycle complexity (calculated from parasite data).
2. Materials and Methods
2.1. Study Area
The ‘Mar Menor’ (‘smaller sea’, in English) is the largest coastal lagoon in the western Mediterranean Sea (approximate surface area = 135 km2, mean depth = 4 m, maximum depth = 6 m). This hypersaline (45 g L−1) water body is located in the south-east of the Iberian Peninsula (Murcia Region, Spain), which is separated from the Mediterranean Sea (salinity = 38 g L−1) by a sand-bar (length = 21 km, mean width = 500 m) (Figure 1). This area is characterized by a semi-arid Mediterranean climate, with intense summer drought (<5 mm) and rainfall concentrated in autumn-winter (20–30 mm). The average annual temperature typically ranges between 15–20 ºC. The lowest temperatures occur in January (3 ºC) and the highest in August (>40 ºC). Minimum water temperature is observed in January (10 ºC) and maximum in August (30 ºC). Geological substratum in the study area consists of an extensive sedimentary cover from the Quaternary, which is formed by black and red silts. Climatic and geological data were retrieved from the Ministry of Environment Spain (in Spanish: ‘Ministerio para la Transición Ecológica y el Reto Demográfico’, ‘MITECO’ hereafter; [40]). Specifically regarding aquatic vegetation, littoral shallow waters are mostly occupied by the seagrass Cymodocea nodosa, which also form mixed meadows with Ruppia cirrhosa. Fish fauna is also highly diverse and has been previously well-described by Oliva-Paterna et al. [41]. In terms of conservation, shallow habitats of the Mar Menor are extremely relevant as settlement sites and juvenile refuges for several fish species, because of its ecological role as a ‘nursery’ area. In addition, this coastal lagoon is home to important populations of highly threatened fish species, such as the European eel Anguilla anguilla and the Iberian toothcarp Aphanius iberus [42].
2.2. Field Sampling
Fish were collected in mid-August 2023 and 2024, after the breeding season (spawning until July) for the particular target species (see details below) in the study area [37]. Therefore, this sampling season avoids any effect of this physiological (i.e. reproductive) status on data. As just mentioned, this was a two-year study, which allowed the acquisition of a more representative dataset than a single-year study, according to Lanzoni et al. [43]. These authors recommend more than one year of sampling in transitional habitats (e.g. lagoons) to encompass all the variability present in fish populations from such dynamic ecosystems. The years 2023 and 2024 are considered to have been hydrologically ‘average’ in the study area [40]. As a result, the effects of particular dry or wet years on morphological and parasitological traits are avoided within the study period, which allows the data to be considered representative for this fish species and their parasites in the Mar Menor. To accomplish an appropriate spatial representativeness, six sampling sites were selected all around the lagoon, which were evenly distributed and separated >5 km (shoreline) to minimise data dependence (Figure 1.)
Sampling sites were located in the vicinity of similar environmental surroundings, with regards to vegetation, and far from influences of main tributaries, towns and fishing areas. Each site was randomly sampled on different dates during two weeks. Fish collection was always carried out at 10:00 (solar time) per site and weather conditions (e.g. temperature, humidity, visibility) were similar among sampling dates. A consistent sampling protocol was followed by wading according to the European legislation (Comité Européen de Normalisation/International Organization for Standardization: CEN/ISO; [44]). Also, sampling effort was similar per site, in terms of time (1.5 h) and research team (the same three people, see ‘Author Contributions’ below), to ensure data comparability. Similarly to Almeida et al. [37], catch method consisted of hauling a beach-seine net (10 × 2 m, 2 mm mesh size) along a 20-m section of shoreline. Five replicates (i.e. 5 × 20-m section = 100 m shoreline) were collected at each sampling site. The area covered by each haul was approximately 175 m2 (total hauled area ≈ 900 m2 per site). To encompass the existing environmental variability, fish were collected from all meso-habitats present in the study sites (e.g. silt, sandy bottoms or patches of submerged vegetation).
All these sampling conditions allowed the collection of a representative and comparable sample of the target fish species, the black-striped pipefish Syngnathus abaster (‘pipefish’, hereafter), at each site. Target species selection was a key point of the present research design for several reasons: 1) This small-bodied fish is easy to be transported and maintained in aquaria (see transportation and laboratory procedures below); 2) It is not under any threatened category (in terms of ‘IUCN Red List’) at the regional level in Murcia (and listed as ‘Least Concern’ in the Mediterranean area; see [45]); 3) It is among the most abundant and common fishes in the Mar Menor coastal lagoon [41,42]. Consequently, the extraction of a relatively large fish sample (see n below in the ‘Morphological Examination’ section) had virtually no effect on this population, nor on subsequent data acquisition of the present (two-year) study; and 4) Pipefish belongs to the taxonomic Order Syngnathiformes, being a zooplanktivorous species, swimming through aquatic vegetation at a certain distance from the bottom (i.e. limnetic habits) and inhabiting a coastal lagoon [46]. Such phylogenetic ‘position’ and environmental requirements of the study species are very contrasting with respect to previous studies analysing the ‘connections’ between genetic diversity of hosts and fish parasites (e.g. see the case of a river leuciscid species in Blanchet et al. [26,27]; or the Atlantic salmon in Zueva et al. [25]). Thus, the present study would contribute towards providing a more complete ‘picture’ of this biotic relationship affecting host population traits (e.g. parasites mediating directional, balancing or disruptive selection; see the ‘Discussion’ section).
After each survey was concluded per site, captured fish were identified and the target species (i.e. pipefish) was separately placed in tanks with cooled water (by using cooling accumulators) and supplied oxygen (two battery-operated aerators with portable pumps). Within the complete fish sample per site, the largest individuals were visually selected for laboratory examinations. To ensure collection of the ‘largest size’ sub-sample, fish were measured for Standard Length (SL, ±1 mm). Such selection was to minimise the effect of fish size on data. In fact, no significant differences were found between sites and genetic categories (see ‘Data Analyses’ and ‘Results’ sections). Also, the use of this sub-sample ensures sufficient maturity to more effectively reveal health and parasitological patterns for small-bodied fishes (see a similar procedure in Almeida et al. [37,47]). In particular for the Mar Menor, pipefish reach an age of up to 4 years [37]. Thus, the capture of larger/older specimens allows that those individuals had completed 2–3 reproductive cycles, minimising the effect of extraction on this wild population, according to the license for scientific fish sampling (see details below). Fish were then transported to a laboratory facility at the University of Murcia, which is located <1 h from the study area. Thus, all fish individuals collected at one particular site (see sampling size below in the ‘Morphological Examination’ section) were processed on the date of capture, in the afternoon. This overall field procedure helped to: 1) minimise the effect of stress during handling/transportation, and 2) reduce disturbance of their parasite infra-communities (e.g. specimen loss). This is a more appropriate alternative to maintaining fish in aquaria for more than 24 h (see a comprehensive review on fish captivity in Näslund and Johnsson [48]).
All field procedures complied with animal use and care regulations of Europe and Spain (a specific license was granted for scientific field research in Murcia: AUF/2023-24/0007). Fish were collected by trained personnel (i.e. the holder of the license, F.J. Oliva-Paterna). Thus, no adverse effects were caused on the wildlife in the study area and all native fish fully recovered from the collection methods. Except for the study species, fish were released after recovery at the same sites of the lagoon where they were captured.
2.3. Morphological Examination
On arrival at the laboratory, one 50-L aquarium was filled with water collected from the same site of the Mar Menor lagoon. This aquarium was equipped with constant oxygen supply, water filter, temperature/pH controller and environmental enrichment (e.g. similar substrate and submerged vegetation as per each site), according to Näslund and Johnsson [48] recommendations. Fish were allowed to acclimatise for a 30-min period by swapping water between the laboratory aquarium and the transportation tank, before being completely released into aquarium. Fish were individually euthanised by immersion in an overdose solution of anaesthetic (dropwise addition of clove oil) for 5 min (adapted from Chapman et al. [49]), followed by severance of the spinal cord. Immediately after euthanasia, each fish specimen was measured again for SL (±1 mm). As per Latorre et al. [50], this particular fish length was selected because it avoids potential ‘noise’ given by variation of caudal fin length not related to body size (e.g. wounds and cuts in the fish skin and rays). In total, n = 360 individuals (30 ind. × 6 sites × 2 years) were examined. The size range was 80–125 mm SL. After size measurements, fish were dissected and sex determined.
Body Condition (‘BC’, hereafter) is basically any index calculated by using the body mass relative to size and then compared to a reference value. Therefore, the BC concept emerges as the plumpness, fatness or well-being of the animal [51,52]. Indeed, a BC index can be also used as an external morphological approach to measure fish health [37,53]. In this study, eviscerated Body Mass (eBM) was measured (electronic balance, ±1 mg) to provide an integrated quantification of BC, avoiding bias from gonad mass and gut content. The eBM range was 192–799 mg. Then, data on fish size (SL) were used for the calculation of BC (see details in the ‘Data Analyses’ section).
Adapted from Adams et al. [39], a Health Assessment Index (‘HAI’, hereafter) was computed as another morphological approach to estimate fish fitness. Unlike other ‘external’ condition indices (e.g. BC), a variety of internal organs are usually examined to compute the HAI (see details below). Thus, the simultaneous use of both BC and HAI (i.e. on the same fish individuals) provides a better understanding on the environmental and physiological processes related to fish health status. Moreover, HAI helps to complement data on parasite infra-communities, with these (health and parasitological) traits being related to the resistance/tolerance capacity of fish hosts (see the ‘Discussion’ section). Specifically, gills and skin (fins, scales) were externally examined for abnormalities. Then, a variety of organs were internally examined to assess the colour, size and texture: eyes, alimentary tract, heart, liver, spleen and gonads. Special care was taken to avoid damaging parasite fauna during this health assessment (see details below in the ‘Parasitological Examination’ section). Scores for each examined category ranges from 0 to 30 points and they were combined to compute a particular HAI value per individual. An HAI score of zero indicates a fish that did not have any appreciable abnormalities, whereas a high score reflects an individual perceived to be unhealthy. This index was calculated according to the following formula:
where HSi is the ‘Health Score’ of the anatomical region/organ i (range: 0–30 points) and n is the total number of examined anatomical regions/organs (n = 8) on a particular fish individual.
n
HAI = ∑ HSi,
i
2.4. Parasitological Examination
Ecto- and endo-parasites were identified and counted per fish individual during the HAI examination (i.e. the same anatomical regions and organs, see above). In this study, the number of fish examined for parasites was much higher than in other studies, where species accumulation curves were applied to calculate a representative sample size (e.g. see Figure 1 showing <20 fish individuals per site in Chapman et al. [49]). The sample size in the present work (n = 30 fish individuals per site) achieved a good representativeness of parasite infra-communities for pipefish (see other examples of this procedure in Cruz et al. [37] and Almeida et al. [47,53]). Fish were examined under a dissecting microscope (40×) for the presence of parasites according to standard protocols [54,55]. Additionally, parasites were identified using a light microscope (1000×) equipped with phase contrast and differential interference contrast. Parasites were excysted, when necessary, by using fine forceps. Where necessary, parasites were fixed in 70% ethanol for later staining (e.g. Iron-Aceto-Carmine, ‘IAC’) and identification. Parasites were identified (and counted) using the keys/databases in Brewster [56], Bruno et al. [57], Falaise [58] and Stoyanov et al. [59]. For particular parasite taxa, Family or Genus were the lowest practical taxonomic levels, since these parasite categories provide information enough to assess their ecological role at the host population level [49,60]. All scientific names for parasite taxa were checked in the Global Biodiversity Information Facility (‘GBIF’), appearing as ‘accepted’ [61]. After examinations, specimens were deposited in the Parasitological Collection of the Department of Biological Sciences at the USP-CEU University (Madrid, Spain).
2.5. Genetic Analyses
Microsatellite markers are often used as a surrogate for genome-wide diversity to infer associations between parasite load and host genetic traits [24], including in fish [26,27,62]. Adapted from Aljanabi and Martinez [63], a sample (≈25 mg) of muscle tissue was taken from each pipefish individual (right flank) to extract total DNA by using the DNeasy® Blood & Tissue Kit (Qiagen, Valencia, CA, USA). DNA was quantified in a Nanodrop® Lite Spectrophotometer (ThermoScientific, Wilmington, DE, USA). Based on previous genetic analyses in Syngnathus species [64,65], n = 10 polymorphic microsatellite loci were selected for this study (Table 1) due to their high variability and similar conditions for amplification. For each pipefish specimen, loci markers were co-amplified by using the QIAGEN Multiplex PCR Kit (Qiagen, Valencia, CA, USA). Polymerase Chain Reactions (PCR) were carried out in a 16 μL final volume containing 20–25 ng of genomic DNA, 8 μL of 2 × QIAGEN Multiplex PCR Master Mix and locus-specific optimized combination of primers (detailed protocols are available from the authors upon reasonable request). PCR amplifications were performed in a Sure Cycler 8800 machine (Agilent Technologies, Santa Clara, CA, USA) under the following program: 15 min at 95 ºC, followed by 30 cycles × [30 s at 94 ºC, 90 s at 56 ºC and 60 s at 72 ºC] and finally a 45-min final elongation step at 72 ºC. Amplified fragments were separated on an ABI PRISM 3730 automated capillary sequencer (Applied Biosystems, Foster City, CA, USA) in the Department of Genomics/Proteomics at the Complutense University (Madrid, Spain). Allelic sizes were scored using the MW marker GeneScan-500 LIZ® (Applied Biosystems, Foster City, CA, USA). Genotypes were analysed by using the software Peak Scanner v.2.0 (Applied Biosystems, Foster City, CA, USA) to generate individual profiles with the 10 microsatellites.
2.6. Data Analyses
General Linear Models (‘GLMs’) were used to test for differences between males and females on the examined traits, as per Almeida et al. [47]. Given that no difference was found (overall p-values > 0.05), this categorical factor (i.e. gender) was not included in subsequent models. This allowed the simplification of data analyses and thus increased the statistical power of the remaining sources of variation, which would otherwise be seriously compromised [37,47]. Data were pooled because no spatial/temporal effect was found to be significant, after previous Generalised Linear Mixed Models (‘GLMMs’) using ‘site’ and ‘year’ as the random factors (a comprehensive review of this statistical technique is given in Johnson et al. [66]).
With respect to genetic traits, no null alleles or linkage disequilibrium were detected in the selected set of microsatellites. For the latter data analysis, the adjusted threshold of statistical significance was set at α = 0.0005 (see a similar procedure in Blanchet et al. [26]). Two genetic indices of homozygosity were calculated: ‘Internal Relatedness’ (IR, see specific algorithm in Amos et al. [67]) and ‘Homozygosity by locus’ (HL, see specific algorithm in Aparicio et al. [68]). As per Blanchet et al. [26], both indices were highly correlated (r = 0.986, p < 0.001), with only IR (range: from −0.125 to 0.470) being finally reported in the present study (see a similar genetic approach in Cruz et al. [53]). This genetic measure (i.e. IR) attempts to estimate the relatedness of an individual’s parents using the extent of allele sharing relative to random expectations [67]. Therefore, IR was used as a measure of ‘Genetic Diversity’ in the present study (similar to Blanchet et al. [26]). Additionally, IR often outperforms other indices when predicting individual fitness [69]. Individuals with low IR values are more heterozygous within the population, whereas high IR levels correspond to more homozygous specimens. Cruz et al. [53] and Blanchet et al. [26] revealed simple-linear and curvi-linear (i.e. quadratic) regressions (respectively) between IR and parasite burden of leuciscid fishes. However, no continuous association was found to be statistically significant (overall p-values > 0.05) for the present data set, after using Generalised Additive Models (‘GAMs’, as per Cruz et al. [53]). Alternatively and according to the IR range (total amplitude ≈ 0.6, see above), three categories of genetic diversity were arbitrarily established (interval ≈ 0.2 per category): ‘Low’ (IR > 0.3), ‘Medium’ (0.1 < IR < 0.3) and ‘High’ (IR < 0.1) levels. Then, all fish data were distributed into each corresponding category to test for genetically based differences in population, morphological and parasitological traits (see the ‘Results’ section). This analytical technique (i.e. splitting IR into categories) was used to more clearly detect and visualise variation patterns (see a similar procedure in Blanchet et al. [27]), as well as ease biological interpretations.
In order to reveal the genetic pattern at the population level, the numbers of collected fish individuals were used to calculate percentages per IR category. Parasite prevalence (the proportion of examined hosts infected with parasites) was also calculated as a percentage across the three categories. Then, chi-square tests were used to detect significant differences for these two response variables (i.e. population frequencies and prevalences) between genetic categories, along with a post-hoc Marascuilo tests for pair-wise comparisons.
One-way ANOVA was used to ensure the existence of highly significant differences (p < 0.001) in the IR means between the three genetic categories. Then, a post-hoc Tukey-Kramer Honestly Significant Difference (HSD) test was used to perform pair-wise comparisons across genetic categories. ANOVA was also used to confirm that similar fish sizes (SL) were selected between genetic categories (see the ‘Results’ section below). Despite no differences being found in fish length, the effect of body size was controlled for subsequent comparisons by using Analysis of Covariance (ANCOVA) to avoid any subtle interaction. In particular for BC, ANCOVA allowed the comparison of eBM between genetic categories, after controlling for fish size (covariate: SL). This analytical procedure, i.e. the use of body length as the covariate, is statistically preferable to control for size effects, because it avoids the bias of using the residuals from linear regressions between the study parameters and the size [52] or computing indices/ratios (e.g. Fulton’s condition factor) [52,70].
As per Almeida et al. [47], quantitative descriptors were used to properly analyse the variation in parasitological traits between categories of genetic diversity. Two parameters of parasite infra-communities were calculated for each fish individual (adapted from Bush et al. [71]): ‘Total Abundance’ (TA, total number of parasite specimens detected from the examined anatomical regions/organs) and a ‘Life-cycle Complexity Index’ (LCI). The latter descriptor was calculated according to the following formula:
where ai is the abundance of a parasite taxon i, hi is the number of different hosts in the life cycle of i and n is the total number of parasite taxa on a particular fish individual. The number of hosts was allocated to each parasite taxon according to available literature (see identification keys above). This number ranged from 1 to 3 hosts for the parasite taxa found in the present study and consequently, the descriptor LCI (i.e. number of hosts weighed by the abundance of each parasite taxon) also varied within this range. As with the procedure for BC, ANCOVA (covariate: eBM) was used to test for significant differences between genetic categories for the response variables HAI and the two parasite descriptors (TA, LCI).
n n
LCI = ∑ (ai × hi) / ∑ ai,
i i
Data were transformed by using log10 (x + 1). Assumptions of normality of distributions and homogeneity of variances were verified through Shapiro-Wilks and Levene’s tests, respectively. Statistical analyses were performed with R v.4.1.0 [72]. The significance level was set at α = 0.05. Values reported in results are arithmetic Means ± Standard Deviations (SD).
3. Results
Genetic diversity (measured as IR, in an inverted order) significantly varied across the three categories (ANOVA: F2,357 = 106.22, p < 0.001), with all the groups being different (Tukey’s HSD test: p < 0.05). Specific results were: Low = 0.369 ± 0.0712, Medium = 0.189 ± 0.0758 and High = 0.005 ± 0.0684 levels. The calculation of population frequencies showed a lower percentage of fish individuals displaying a medium level of genetic diversity (26.7%), although this difference was marginally non-significant (χ2 = 5.65, p = 0.059). After pair-wise comparisons, the difference between the low (35.8%) and medium (26.7%) levels was close to the significance threshold (post-hoc Marascuilo: p = 0.070), whereas the medium (26.7%) and high (37.5%) levels was significantly different (post-hoc Marascuilo: p < 0.05) (Figure 2).
Fish sizes were similar across the three genetic categories (ANOVA: F2,357 = 0.66, p = 0.518), with SL results (mm) as follows: Low = 93.9 ± 15.32, Medium = 89.9 ± 12.85 and High = 91.4 ± 14.56 levels. After controlling for fish size (SL), adjusted eBM values (i.e. BC) were lower (mean < 315 mg) for the medium level in comparison with the low and high levels (>400 mg), although this difference did not reach the statistical significance (but p < 0.1 from ANCOVA; see Table 2). After controlling for fish mass (eBM), adjusted HAI means were very different among genetic groups (ANCOVA: p < 0.01; see Table 2). In particular, this health index was significantly higher (>35 points) for the medium level, whereas the low and high levels were similar (≈25 points) (Table 2).
Pipefish showed a higher percentage of parasite prevalence for the medium level of genetic diversity (74.0%) relative to the low (65.9%) and high (62.2%) levels, although this difference was not statistically significant (χ2 = 3.53, p = 0.171). After pair-wise comparisons, the difference between the high (62.2%) and medium (74.0%) levels was close to the significance threshold (post-hoc Marascuilo: p = 0.061) (Figure 3).
A total of 13 parasite taxa (Family, Genus or Species level) were identified for pipefish (Table 3). As higher hierarchical taxonomic groups, parasites ranged from single-cell organisms to metazoan: Ciliophora, Cestoda, Monogenea, Digenea, Nematoda, Acanthocephala and Crustacea. Digenean species were the most diverse and abundant parasites (six taxa accounting for >65% of total parasite abundance from the three genetic categories). In particular, the most abundant digenean parasites were the taxonomic Family Cyathocotylidae, followed by Podocotyle atherinae, Cryptocotyle concava and the Genus Diplostomum. For all these parasites (as well as the rest of digeneans, the nematode Contracaecum microcephalum and the cestode Proteocephalus sp.), the highest abundances were always found in the medium level of genetic diversity (Table 3). Other common parasites for pipefish were the copepod Ergasilus ponticus, the monogenean Genus Gyrodactylus, the acanthocephalan Pomphorhynchus laevis and the ciliate Trichodina partidisci, with these four parasite taxa being less abundant in the medium category. The digenean Timoniella imbutiformis was not observed in more heterozygous pipefish, whereas Echinochasmus perfoliatus (digenean) and Paracanthocephaloides incrassatus (acanthocephalan) were not detected in more homozygous hosts (Table 3).
4. Discussion
The observed population pattern (i.e. frequencies of individuals) in the present study could be in accordance with a scenario of disruptive selection, i.e. leading to a bimodal trait distribution [23,73]. In particular, a negative selection appeared to mainly focus on individuals displaying ‘intermediate’ genetic diversity (i.e. the medium level being less common), whereas the most homozygous and heterozygous pipefish would be positively selected (i.e. ‘extremes’ being more frequent). Mechanistically, the selective pressure promoting such ‘disruption’ within this pipefish population may be related to a differential effect of parasitism, depending on host genetic traits, as suggested per Blanchet et al. [26,27]. However, such differences among population percentages were not (statistically) clear in the present study, according to this particular type of natural selection (only for the ‘high’ category in comparison with the ‘medium’). A potential explanation for this partial lack of statistical significance may be because of the examined specimens belonging to a particular fraction of the entire population (i.e. the oldest adults, as the largest specimens were analysed and fish typically exhibit continuous growth throughout their full life-span). In the Iberian Peninsula, clear ecological patterns relating a variety of parasitological traits were observed after using larger/older specimens to assess colonisation capacity (see the case of a minnow population in Cruz et al. [53] and Almeida et al. [47]) and as ecological indicators (see the case of pipefish and goby species in Almeida et al. [37]). Moreover, Blanchet et al. [26] found no effect (i.e. statistical interaction) of host body size on any mathematical model including IR and parasite load for a French dace population. These previous studies provided the rationale to select the largest individuals of pipefish in the Mar Menor. However, juvenile fish may be more vulnerable to parasitism and consequently, better reflect this effect on populations under certain environmental circumstances, such as heterogeneity of habitat features [49,74]. Thus, future research should focus on a broader size range to overcome this potential limitation. Overall, this more comprehensive analysis would truly reveal the differential effect of parasites depending on genetic diversity at the entire population level (i.e. juvenile, sub-adult and adult individuals). Additionally, the assessment of genetic markers in juvenile pipefish could also shed light on the effect of parasites on population recruitment, a key trait to assess viability of fish communities on a long-term basis [4,75].
Parasite prevalence and abundance usually follow the same variation trends, as observed by Chapman et al. [49] in a freshwater fish (pumpkinseed sunfish Lepomis gibbosus). Similarly, these two parasitological parameters showed the same pattern across the genetic categorisation, with both more homozygous and heterozygous individuals harbouring, on average, fewer parasites than pipefish from the intermediate level of genetic diversity. Despite significant differences not being clearly detected, such results do merit some consideration, as again they could be understood through a mechanism of disruptive selection. Thus, this was contrary to the expectation on directional selection (i.e. lower parasite burden) only for the most heterozygous individuals [24,76], which has also been observed in different fishes (e.g. see Zueva et al. [25] for Atlantic salmon or Fraser and Neff [77] for guppy Poecilia reticulata). In this genetic context where heterozygosity is highly advantageous, balancing selection (i.e. overdominance of heterozygous specimens) could also occur at a certain degree of magnitude (e.g. see Mäkinen et al. [78] for a study on three-spined stickleback Gasterosteus aculeatus), which would contribute to the maintenance of the same level of genetic diversity over time [79]. Alternatively, Blanchet et al. [26] found that disruptive selection was acting on a dace population infected by a fin-feeder parasite, by using both continuous data and categories of genetic diversity [27]. As per these authors, a similar genetic/parasitological pattern was observed by using IR categories, prevalence percentages and TA values from pipefish from the Mar Menor, i.e. explained as disruptive selection. These overall findings are very relevant, as this same evolutionary trend has been observed on very contrasting species, in terms of phylogenetic ‘distance’ and ecological requirements, such as a leuciscid fish in fresh waters and a syngnathid species in transitional waters. Indeed, parasitological examinations were much less complete in the mentioned studies (e.g. one helminth species in Zueva et al. [25], two species from the same Genus in Fraser and Neff [77] and one copepod species in Blanchet et al. [26,27]), whereas a higher variety of parasites were analysed in the present work: protozoans, cestodes, monogeneans, digeneans, nematodes, acanthocephalans and crustaceans. Such assessment of several parasite taxa at the same time is obviously more representative, but it poses a risk for biological interpretation, as they can display a wide range of traits (e.g. virulence, transmission) that may differentially affect interactions with host individuals. Therefore, this elevated variability could be the reason why statistical differences did not reach the significance level for prevalence and parasite abundance in pipefish, potentially masking the underlaying ecological and evolutionary processes.
With respect to health status, BC and HAI have previously been used to detect variations in physical fitness of fishes inhabiting transitional waters [37]. Both morphological parameters exhibited the same pattern, with pipefish individuals that are either more homozygous or more heterozygous displaying a better condition/health. These findings were in accordance with the parasitological pattern, likely related to the effect of host genotypes on susceptibility to infection and parasite transmission (see a detailed explanation below). However, a statistical significant difference across genetic categories was only found for HAI. In the specific case of BC, this index did not reach the significance level because other factors not related to parasites may be also modulating fish condition. In particular, indices calculated from data on body mass are very sensitive to variations in food supply [51] and this may be affecting results on BC. The pipefish species selected for this study chiefly feeds on copepods and cladocerans (two important biological communities within zooplankton) in Iberian waters [46,80]. These crustacean ‘food items’ are not usually a limiting factor in the Mar Menor because environmental conditions can periodically change from an oligotrophic status to an eutrophication level. Such episodes are unfortunately common within this coastal lagoon, which is of conservation concern [81]. Consequently, this ‘bottom-up’ effect promotes an increase in phytoplankton, with the corresponding biomass of zooplankton ‘grazers’ [82]. Statistically speaking (i.e. showing significant differences), HAI was a good indicator of fitness status for pipefish in relation to genetic diversity, with higher HAI values being found in the medium category (i.e. more parasites and worse health). In this respect, Chapman et al. [49] observed that this same index could reflect physiological/histological effects of infectious diseases (e.g. parasitism on gills and guts by ecto-/endo-helminths), with positive correlations being found between HAI scores and parasite loads. Parasitism can directly impact fish organs while feeding, which promotes a variety of histo-pathological effects, such as tissue inflammation, hyperplasia, hypertrophy and necrosis [83]. The closer ‘connection’ with these tissue damages was likely the reason why this health index was able to provide clearer differences between levels of genetic diversity. Such findings demonstrate the importance of using both ‘external’ and ‘internal’ indices of fish health status, with different levels of sensitivity, to better detect variations across genetic, morphological and parasitological traits [53].
According to Råberg et al. [21], hosts have evolved two different strategies to maintain fitness when interacting with parasites and pathogens: 1) ‘resistance’ (the ability to actively limit and reduce parasite burden before or after infection has occurred), and 2) ‘tolerance’ (the ability to actively limit and reduce the damage caused by a given parasite burden). These authors also demonstrated how both strategies deeply depended on genetic variation, i.e. displaying different reaction norms across host genotypes. It follows then, that TA can be considered as a proxy for ‘resistance’, whereas HAI can be considered as a proxy for ‘tolerance’ in the present study. Overall, both mechanisms (i.e. resistance and tolerance) may act in a synergistic manner, decreasing prevalence and abundance of diseases, as well as maintaining health, for the highest levels of homozygosity and heterozygosity within this pipefish population. Contrary to this interpretation, Råberg et al. [21] and Blanchet et al. [27] provided evidence that a negative genetic correlation existed between resistance and tolerance, such that a highly tolerant individual is necessarily weakly resistant (and vice versa). Focusing particularly on the ‘fish’ study (i.e. the most equivalent to the present work: [27]), the level of fin degradation was the only trait used in common dace as an inverse descriptor of host health for a given number of parasites (i.e. tolerance). Nevertheless, the assessment of health status was more comprehensive in pipefish from the Mar Menor, with each individual being examined for up to eight anatomical regions/organs (see the ‘Morphological Examination’ section). This could be a potential explanation to have found a positive association between TA and HAI values (i.e. inverses for resistance and tolerance, respectively) across IR intervals in this study.
Regarding the genetic basis of these two physiological processes, specific genes and genomic regions have been related to the immunological capability for resisting parasites at the molecular (e.g. immuno-globulins) and cellular (e.g. lymphocytes, macrophages) levels [25,84,85]. In relation to tolerance, particular host genotypes are better able to control an exacerbated immune (inflammatory) response, by acting on cytokines or mast cells [84,86], as well as reducing tissue degradation through investment in tissue regeneration [22]. Following this genetic perspective and according to the ‘immunological hypothesis’, research on polymorphic loci coding for the Major Histocompatibility Complex (‘MHC’, hereafter) is promising, given their important role in the adaptive immune response of vertebrates (presenting pathogen-derived antigenic peptides to lymphocytes) [3]. Thus, an elevated diversity of these genes (via heterozygosity) is considered a good indicator of resistance in host fishes [77,87]. Within the study pipefish population, the present results appear to reveal two well-defined and efficient contrasting strategies against parasites at the host genotypic level: 1) more heterozygosity, and 2) more homozygosity. The first one is easy to understand, according to previous observations [24,87], where an elevated genetic diversity is surely reflected in a wider ‘repertoire’ of immune response. This promotes a rapid detection on a great variety of parasites and pathogens (antigens), and hence hosts can readily eliminate them [25,87]. The second strategy is less intuitive, but a potential explanation could be based on the polygenic nature of the immune response [25,30,88]. A large number of allelic copies encoding for particular products (e.g. antibodies, cytokines, MHC molecules), although less diverse due to homozygosity, could reach a higher synthesis rate (i.e. a ‘gene-dose’ effect; [89]), with this also being an effective mechanism to control parasites/pathogens (see Spiering and de Vriesh [90] for an example on a higher bi-allelic expression/synthesis against COVID-19). For the medium category, these pipefish individuals would be neither ‘diverse’ nor ‘productive’ enough to display a suitable level of immuno-competence, facilitating penetration/migration of parasites through host tissues and consequently, being more susceptible to infections. Such (worse) genotypic context was also reflected in the highest HAI mean (i.e. poor health status).
Parasite taxa recovered from pipefish were similar to those found in previous research on the same fish species, although richness was lower [60,91]. This finding (i.e. a lower richness) may be due to the particular hypersaline conditions within the Mar Menor lagoon (see description of the ‘Study Area’ above). In terms of poor water quality, elevated salinity commonly affects fish parasites via direct (e.g. ciliated and free-swimming larvae, such as miracidium searching for mollusc hosts) and indirect effects (e.g. host tolerance) [92]. In the present study, pipefish belonging to the low/high levels of genetic diversity were able to avoid infections from three parasite species (two digeneans and one acanthocephalan), as well as maintaining lower TA (marginally non-significant) and LCI (clearly significant) values, with respect to the medium category. For the low/high categories, monoxenous ecto-parasites (skin and gills) were more abundant, such as Ergasilus ponticus (crustacean), Gyrodactylus (monogenean) and Trichodina partidisci (ciliate), which display direct cycles, i.e. they are transferred from fish to fish. The acanthocephalan Pomphorhynchus laevis was also more frequent in pipefish intestines from those two categories. This helminthiasis needs two species to complete its heteroxenous life cycle: amphipods (intermediate crustacean host) and fish. Therefore, all these parasite taxa established a common link with pipefish, which always played a role as the unique or definitive host (i.e. where sexual reproduction takes place by the adult stage). Thus, fish usually remained alive after this parasitic interaction [8]. Yet, parasitism was very different for pipefish from the medium category, where all the observed digeneans (including the most abundant parasites: Cyathocotylidae, Podocotyle, Cryptocotyle and Diplostomum), as well as the nematode Contracaecum microcephalum and the cestode Proteocephalus, were more frequent. All these taxa display heteroxenous cycles. Except for Podocotyle atherinae and Proteocephalus sp. (with small zooplanktivorous fish being the definitive host), the rest of those helminths ‘used’ pipefish as an intermediate host that must be preyed upon by larger predatory fish (e.g. sea bass Dicentrarchus labrax) or (mainly) aquatic birds (e.g. cormorants, gulls, herons and egrets) as the definitive hosts [93]. These piscivorous species are common in the study area (see Oliva-Paterna et al. [41] for fishes and Farinós-Celdrán et al. [94] for birds in the Mar Menor) and they usually prey on pipefish species [95,96]. In fish hosts, encysted metacercaria larvae can shelter in the muscle/skin (Cryptocotyle concavum), eye lens (Diplostomum sp., causing impaired vision) and other vital organs (Cyathocotylidae). This parasite stage is aimed at changing fish behaviour, to become more vulnerable to predation by piscivorous definitive hosts (e.g. swimming next to the surface; see Barber [97] for a comprehensive review about the role of parasites in fish-bird interactions). In the case of Contracaecum microcephalum, definitive hosts are marine mammals, such as seals and cetaceans. For this nematode species, fish play a role as an intermediate or even more specifically, as a paratenic host, which is a ‘non-essential’ species for the parasite cycle, although it can increase the transmission rate (similarly to the phylogenetically related and well-known Genus Anisakis; [98]). These kinds of parasite species (digeneans and nematodes) were more common in the medium category, where such infestations of those anatomical systems/organs may cause severe tissue damage. Thus, a stronger pathological effect would be necessary for these parasites ultimately to provoke the fish death via predation by definitive hosts. Such negative pressure on pipefish from the medium level of genetic diversity was likely reflected in higher HAI values (see above), promoting clear statistical differences. These findings on parasite composition were directly related to the LCI results, with an average significantly higher for the medium category. This indicated an elevated abundance of heteroxenous parasites that need several ecologically diverse hosts (e.g. zooplankton, snails, fish, birds or mammals) to complete their life cycles. Apparently, pipefish from the low/high categories were capable of better resisting (i.e. stronger immune response) those more ‘noxious’ parasites, i.e. heteroxenous cycles that necessarily implies fish mortality [93]).
5. Conclusions
This study is underpinned by the use of particular traits to evaluate the intimate ecological and evolutionary relationships between parasitism and host genotypes in animal populations. Thus, all the analysed parameters in the present work (i.e. frequencies of host population, health indices and parasitological traits) always provided patterns that could be easily explained under a scenario of disruptive selection. Overall results suggests that this type of natural selection has been occurring in pipefish inhabiting the Mar Menor coastal lagoon. However, it is also necessary to highlight that the clearest differences (statistically speaking) were revealed for HAI (a more ‘internal’ index) and LCI (an indicator of more ‘noxious’ parasites), probably because these two parameters are reflecting deeper effects on hosts (see comments above). From a strict (micro-)evolutionary perspective, parasites appear to play a crucial role for shaping genetic diversity within local communities of animal hosts. In particular for the study pipefish population, genetic variability (or variance, in mathematical terms) was broadened through disruptive selection, which had already been observed by Blanchet et al. [26,27] in a very different fish species: the leuciscid common dace inhabiting fresh waters. Therefore, this study demonstrates that parasite-mediated disruptive selection is a widespread phenomenon across finfish populations, as well as other wild aquatic communities (see Duffy et al. [99] for an example on a crustacean species from zooplankton). Also, the present results complement the most intuitive view that a high heterozygosity is the only possible means to achieve a fitter status [24,25,76], with two main strategies being displayed at the host genotypic level to better deal with parasites: 1) high homozygosity (‘gene-dose’ effect on immune synthesis), and 2) high heterozygosity (wider ‘repertoire’ of immune response). Mechanistically, homozygosity level (i.e. genotypic variability measured by the IR) may reflect immuno-competence as a phenotypic trait [100], with parasites exerting a higher negative (purifying) selection on host individuals from the ‘intermediate’ range (displaying a poorer immune response). These findings open new perspectives for future research lines on the genetic interaction between hosts and their parasites, as well as the underlying processes. Consequently, researchers from different academic disciplines, such as animal health, ecology and evolution, may find the present study a valuable contribution to understand the genetic mechanisms by which hosts ‘adapt’ to pathogen/parasite infections.
Author Contributions
M.T., F.J.O.-P. and D.A. contributed to the study design. M.T., F.J.O.-P. and D.A. carried out the field sampling. A.C., E.L., C.L., L.O.-D., G.C.-G., M.T. and D.A. carried out the laboratory work. A.C., E.L., C.L., L.O.-D., G.C.-G., D.A. and D.H.F. performed the statistical analyses. M.T. and F.J.O.-P. were responsible for the acquisition of funds. D.A. and D.H.F. led the writing of the manuscript. M.T. and F.J.O.-P. contributed substantially to revisions of the manuscript. All authors gave final approval for submission.
Funding
This research was partly funded by the Environmental Service and the Mar Menor Service of the Government of the Autonomous Community of Murcia, Spain (Project: “Estudio del estado de la ictiofauna indicadora de zonas someras, mejora de la información y aplicación en la redacción de proyectos en zona sumergida del Mar Menor”-TRAGSATEC [in Spanish]).
Data Availability Statement
Data are available from the authors upon reasonable request.
Acknowledgments
The authors are grateful to Antonio Zamora-López, Adrián Guerrero-Gómez and Antonio Guillén (Department of Zoology and Physical Anthropology, University of Murcia, Spain), who helped with the field and laboratory works.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Coen, L.D.; Melanie, J.B. The ecology, evolution, impacts and management of host-parasite interactions of marine molluscs. J. Invertebr. Pathol. 2015, 131, 177–211. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.J.F. Complex networks of parasites and pollinators: Moving towards a healthy balance. Phil. Trans. R. Soc. B 2022, 377, 20210161. [Google Scholar] [CrossRef]
- Fleischer, R.; Eibner, G.J.; Schwensow, N.I.; Pirzer, F.; Paraskevopoulou, S.; Mayer, G.; Corman, V.M.; Drosten, C.; Wilhelm, K.; Heni, A.C.; et al. Immunogenetic-pathogen networks shrink in Tome’s spiny rat, a generalist rodent inhabiting disturbed landscapes. Commun. Biol. 2024, 7, 169. [Google Scholar] [CrossRef]
- Longshaw, M.; Frear, P.A.; Nunn, A.D.; Cowx, I.G.; Feist, S.W. The influence of parasitism on fish population success. Fish. Manag. Ecol. 2010, 17, 426–434. [Google Scholar] [CrossRef]
- Lutermann, H.; Butler, K.B.; Bennett, N.C. Parasite-mediated mate preferences in a cooperatively breeding rodent. Front. Ecol. Evol. 2022, 10, 838076. [Google Scholar] [CrossRef]
- Andriolli, F.S.; Cardoso Neto, J.A; Fine, P.V.A.; Salazar, D.; Figueroa, G.; Torres, D.V.; de Morais, J.W.; Baccaro, F.B. When zombies go vegan: Ophiocordyceps unilateralis hosts are selecting to bite palm leaves before dying? Acta Oecol. 2025, 126, 104055. [Google Scholar] [CrossRef]
- Cabanellas-Reboredo, M.; Vázquez-Luis, M.; Mourre, B. Tracking a mass mortality outbreak of pen shell Pinna nobilis populations: A collaborative effort of scientists and citizens. Sci. Rep. 2019, 9, 13355. [Google Scholar] [CrossRef]
- Bush, A.O.; Fernández, J.C. ; Esch, G; Seed, R. Parasitism: The Diversity and Ecology of Animal Parasites, 1st ed.; Cambridge University Press: Cambridge, UK, 2001; p. 576. [Google Scholar]
- Adams, S.M.; Brown, A.M.; Goede, R.W. A quantitative health assessment index for rapid evaluation of fish conditions in the field. Trans. Am. Fish. Soc. 1993, 122, 63–73. [Google Scholar] [CrossRef]
- Krist, A.C.; Jokela, J.; Wiehn, J.; Lively, C.M. Effects of host condition on susceptibility to infection, parasite developmental rate, and parasite transmission in a snail-trematode interaction. J. Evol. Biol. 2004, 17, 33–40. [Google Scholar] [CrossRef]
- Tschirren, B.; Bischoff, L.L.; Saladin, V.; Richner, H. Host condition and host immunity affect parasite fitness in a bird-ectoparasite system. Funct. Ecol. 2007, 21, 372–378. [Google Scholar] [CrossRef]
- Holmlund, C.M; Hammer, M. Ecosystem services generated by fish populations. Ecol. Econ. 1999, 29, 253–268. [Google Scholar] [CrossRef]
- Williams, H.H.; Jones, A. Parasitic Worms of Fish, 1st ed.; Taylor & Francis Ltd: London, UK, 1994; p. 610. [Google Scholar]
- Cribb, T.H.; Bray, R.A.; Littlewood, D.T.J. The nature and evolution of the association among digeneans, molluscs and fishes. Int. J. Parasitol. 2001, 31, 997–1011. [Google Scholar] [CrossRef] [PubMed]
- Marcogliese, D.J. Parasites of the superorganism: Are they indicators of ecosystem health? Int. J. Parasitol. 2005, 35, 705–706. [Google Scholar] [CrossRef]
- Šimková, A. Host-specific monogeneans parasitizing freshwater fish: The ecology and evolution of host-parasite associations. Parasite 2024, 31, 61. [Google Scholar] [CrossRef]
- Pietrock, M.; Marcogliese, D.J. Free-living endohelminth stages: At the mercy of environmental conditions. Trends Parasitol. 2003, 19, 293–299. [Google Scholar] [CrossRef]
- Lafferty, K.D. Ecosystem consequences of fish parasites. J. Fish Biol. 2008, 73, 2083–2093. [Google Scholar] [CrossRef]
- Valenzuela-Sánchez, A.; Wilber, M.Q.; Canessa, S.; Bacigalupe, L.D.; Muths, E.; Schmidt, B.R.; Cunningham, A.A.; Ozgul, A.; Johnson, P.T.J.; Cayuela, H. Why disease ecology needs life-history theory: A host perspective. Ecol. Lett. 2021, 24, 876–890. [Google Scholar] [CrossRef]
- Buckingham, L.J.; Ashby, B. Coevolutionary theory of hosts and parasites. J. Evol. Biol. 2002, 35, 205–224. [Google Scholar] [CrossRef]
- Råberg, L.; Sim, D.; Read, A.F. Disentangling genetic variation for resistance and tolerance to infectious diseases in animals. Science 2007, 318, 812–814. [Google Scholar] [CrossRef]
- Barrett, D.E.; Estensoro, I.; Sitjà-Bobadilla, A.; Bartholomew, J.L. Intestinal transcriptomic and histologic profiling reveals tissue repair mechanisms underlying resistance to the parasite Ceratonova shasta. Pathogens 2021, 10, 1179. [Google Scholar] [CrossRef]
- Endler, J.A. Natural Selection in the Wild, 1st ed.; Princeton University Press: Princeton, NJ, USA, 1986; p. 354. [Google Scholar]
- Coltman, D.W.; Pilkington, J.G.; Smith, J.A.; Pemberton, J.M. Parasite-mediated selection against inbred soay sheep in a free-living island population. Evolution 1999, 53, 1259–1267. [Google Scholar] [CrossRef] [PubMed]
- Zueva, K.J.; Lumme, J.; Veselov, A.E.; Kent, M.P.; Lien, S. Footprints of directional selection in wild Atlantic salmon populations: Evidence for parasite-driven evolution? PLoS ONE 2014, 9, e91672. [Google Scholar] [CrossRef]
- Blanchet, S.; Rey, O.; Berthier, P.; Lek, S.; Loot, G. Evidence of parasite-mediated disruptive selection on genetic diversity in a wild fish population. Mol. Ecol. 2009, 18, 1112–23. [Google Scholar] [CrossRef]
- Blanchet, S.; Rey, O.; Loot, G. Evidence for host variation in parasite tolerance in a wild fish population. Evol. Ecol. 2010, 24, 1129–1139. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture: Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Froehlich, H.E.; Couture, J.; Falconer, L.; Krause, G.; Morris, J.A.; Perez, M.; Stentiford, G.D.; Vehviläinen, H.; Halpern, B.S. Mind the gap between ICES nations’ future seafood consumption and aquaculture production. ICES J. Mar. Sci. 2001, 78, 468–477. [Google Scholar] [CrossRef]
- Nguyen, N.H. Genetics and genomics of infectious diseases in key aquaculture species. Biology 2024, 13, 29. [Google Scholar] [CrossRef]
- Timi, J.T.; Buchmann, K. A century of parasitology in fisheries and aquaculture. J. Helminthol. 2023, 97, 1–18. [Google Scholar] [CrossRef]
- Madsen, H.; Stauffer, J.R. Aquaculture of animal species: Their eukaryotic parasites and the control of parasitic infections. Biology 2024, 13, 41. [Google Scholar] [CrossRef]
- Gjedrem, T.; Rye, M. Selection response in fish and shellfish: A review. Rev. Aquac. 2018, 10, 168–179. [Google Scholar] [CrossRef]
- Kennish, M.J.; Paerl, H.W. Coastal Lagoons. Critical Habitats of Environmental Change, 1st ed.; CRC Press: Boca Raton, FL, USA, 2010; p. 568. [Google Scholar]
- Elliott, M.; Whitfield, A.K. Challenging paradigms in estuarine ecology and management. Estuar. Coast. Shelf Sci. 2011, 94, 306–314. [Google Scholar] [CrossRef]
- Giari, L.; Castaldelli, G.; Timi, J.T. Ecology and effects of metazoan parasites of fish in transitional waters. Parasitology 2022, 149, 1829–1841. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.; Cruz, A.; Llinares, C.; Torralva, M.; Lantero, E.; Fletcher, D.H.; Oliva-Paterna, F.J. Fish morphological and parasitological traits as ecological indicator of habitat quality in a Mediterranean coastal lagoon. Aquat. Conserv. Mar. Freshw. Ecosyst. 2023, 33, 1229–1244. [Google Scholar] [CrossRef]
- Correia, S.; Fernández-Boo, S.; Magalhães, L.; de Montaudouin, X.; Daffe, G.; Poulin, R.; Vera, M. Trematode genetic patterns at host individual and population scales provide insights about infection mechanisms. Parasitology 2023, 150, 1207–1220. [Google Scholar] [CrossRef]
- Farjallah, S.; Amor, N.; Montero, F.E.; Repullés-Albelda, A.; Villar-Torres, M.; Nasser Alagaili, A.; Merella, P. Assessment of the genetic diversity of the monogenean gill parasite Lamellodiscus echeneis (Monogenea) infecting wild and cage-reared populations of Sparus aurata (Teleostei) from the Mediterranean Sea. Animals 2024, 14, 2653. [Google Scholar] [CrossRef]
- MITECO. Available online: https://www.aemet.es/es/serviciosclimaticos (accessed on 15 March 2025).
- Oliva-Paterna, F.J.; Andreu, A. , Miñano, P.A.; Verdiell, D.; Egea, A.; De Maya, J.A.; Ruiz-Navarro, A.; García-Alonso, J.; Fernández-Delgado, C.; Torralva, M. Y-O-Y fish species richness in the littoral shallows of the meso-saline coastal lagoon (Mar Menor, Mediterranean coast of the Iberian Peninsula). J. Appl. Ichthyol. 2006, 22, 235–237. [Google Scholar] [CrossRef]
- Guerrero-Gómez, A.; Zamora-López, A.; Guillén-Beltrán, A.; Zamora-Marín, J.M.; Sánchez-Pérez, A.; Torralva, M.; Oliva-Paterna, F.J. An updated checklist of YOY fish occurrence in the shallow perimetral areas of the Mar Menor (Western Mediterranean Sea). Limnetica 2022, 41, 101–106. [Google Scholar] [CrossRef]
- Lanzoni, M.; Gaglio, M.; Gavioli, A.; Fano, E.A.; Castaldelli, G. Seasonal variation of functional traits in the fish community in a brackish lagoon of the Po River Delta (Northern Italy). Water 2021, 13, 679. [Google Scholar] [CrossRef]
- European Commission. Commission Directive 2014/101/EU of 30 October 2014 amending Directive 2000/60/EC of the European Parliament and of the Council establishing a framework for Community action in the field of water policy. Official Journal of the European Union-Legislation 2014, 311, 32–35. Available online: http://data.europa.eu/eli/dir/2014/101/oj (accessed on 15 March 2025).
- Ford, M. 2024. Syngnathus abaster. The IUCN Red List of Threatened Species 2024: e.T21257A135092980. https://dx.doi.org/10.2305/IUCN.UK.2024-2.RLTS.T21257A135092980.en (accessed on 15 March 2025).
- Doadrio, I.; Perea, S.; Garzón-Heydt, P.; González, J.L. Ictiofauna Continental Española. Bases para su Seguimiento (in Spanish); DG Medio Natural y Política Forestal, Ministry of Environment: Madrid, Spain, 2011; p. 612. [Google Scholar]
- Almeida, D.; Alcaraz-Hernández, J.D.; Cruz, A.; Lantero, E.; Fletcher, D.H.; García-Berthou, E. Seasonal effects on health status and parasitological traits of an invasive minnow in Iberian Waters. Animals 2024, 14, 1502. [Google Scholar] [CrossRef]
- Näslund, J.; Johnsson, J.I. Environmental enrichment for fish in captive environments: Effects of physical structures and substrates. Fish. Fish. 2016, 17, 1–30. [Google Scholar] [CrossRef]
- Chapman, J.M.; Marcogliese, D.J.; Suski, C.D.; Cooke, S.J. Variation in parasite communities and health indices of juvenile Lepomis gibbosus across a gradient of watershed land-use and habitat quality. Ecol. Indic. 2015, 57, 564–572. [Google Scholar] [CrossRef]
- Latorre, D.; Masó, G.; Hinckley, A.; Verdiell-Cubedo, D.; Castillo-García, G.; González-Rojas, A.G.; Black-Barbour, E.N.; Vila-Gispert, A.; García-Berthou, E.; Miranda, R.; et al. Interpopulation variability in dietary traits of invasive bleak Alburnus alburnus (Actinopterygii, Cyprinidae) across the Iberian Peninsula. Water 2020, 12, 2200. [Google Scholar] [CrossRef]
- Jakob, E.M.; Marshall, S.D.; Uetz, G.W. Estimating fitness: A comparison of body condition indices. Oikos 1996, 77, 61–67. [Google Scholar] [CrossRef]
- García-Berthou, E. On the misuse of residuals in ecology: Testing regression residuals vs. the analysis of covariance. J. Anim. Ecol. 2001, 70, 708–711. [Google Scholar] [CrossRef]
- Cruz, A.; Llinares, C.; Martín-Barrio, I.; Castillo-García, G.; Arana, P.; García-Berthou, E.; Fletcher, D.H.; Almeida, D. Comparing morphological, parasitological and genetic traits of an invasive minnow between intermittent and perennial stream reaches. Freshw. Biol. 2022, 67, 2035–2049. [Google Scholar] [CrossRef]
- Weber, E.P.; Govett, P. Parasitology and necropsy of fish. Compend. Contin. Educ. Vet. 2009, 31, E12. [Google Scholar]
- Stoskopf, M.K. Fish Medicine, 2nd ed.; Saunders Ltd.: Philadelphia, PA, USA, 2011; p. 882. [Google Scholar]
- Brewster, B. Aquatic Parasite Information - A Database on Parasites of Freshwater and Brackish Fish in the United Kingdom. PhD Thesis, Kingston University, London, 2016. [Google Scholar]
- Bruno, D.; Nowak, B.F.; Elliot, D. Guide to the identification of fish protozoan and metazoan parasites in stained tissue sections. Dis. Aquat. Organ. 2006, 70, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Falaise, P. Les Parasites de Poisson: Agents de Zoonoses (in French). PhD Thesis, University of Toulouse, Toulouse, 2017. [Google Scholar]
- Stoyanov, B.; Yasen, M.; Boyko, B. Helminth parasites of black-striped pipefish Syngnathus abaster Risso, 1827 (Actinopterygii: Syngnathidae) from a Black Sea coastal wetland, Bulgaria. Acta Zool. Bulg. 2024, 76, 115–128. [Google Scholar]
- Ondračková, M.; Slováčková, I.; Trichkova, T.; Polačik, M.; Jurajda, P. Shoreline distribution and parasite infection of black-striped pipefish Syngnathus abaster Risso, 1827 in the lower River Danube. J. Appl. Ichthyol. 2012, 28, 590–596. [Google Scholar] [CrossRef]
- GBIF. Available online: https://www.gbif.org/species/search (accessed on 15 March 2025).
- Ondračková, M.; Bartáková, V.; Kvach, Y.; Bryjová, A.; Trichkova, T.; Ribeiro, F.; Carassou, L.; Martens, A.; Masson, G.; Zechmeister, T.; Jurajda, P. Parasite infection reflects host genetic diversity among non-native populations of pumpkinseed sunfish in Europe. Hydrobiologia 2021, 848, 2169–2187. [Google Scholar] [CrossRef]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high-quality genomic DNA for PCR-based techniques. Nucl. Acids. Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef] [PubMed]
- Diekmann, O.E.; Gouveia, L.; Serrão, E.A.; Van de Vilet, M.S. Highly polymorphic microsatellite markers for the black striped pipefish, Syngnathus abaster. Mol. Ecol. Resour. 2009, 9, 1460–1466. [Google Scholar] [CrossRef]
- Jones, A.G.; Rosenqvist, G.; Berglund, A.; Avise, J.C. The genetic mating system of a sex-role-reversed pipefish (Syngnathus typhle): A molecular inquiry. Behav. Ecol. Sociobiol. 1999, 46, 357–365. [Google Scholar] [CrossRef]
- Johnson, P.C.D.; Barry, S.J.E.; Ferguson, H.M.; Müller, P. Power analysis for generalized linear mixed models in ecology and evolution. Methods Ecol. Evol. 2015, 6, 133–142. [Google Scholar] [CrossRef]
- Amos, W.; Wilmer, J.W.; Fullard, K.; Burg, T.M.; Croxall, J.P.; Bloch, D.; Coulson, T. The influence of parental relatedness on reproductive success. Proc. Biol. Sci. 2001, 268, 2021–2027. [Google Scholar] [CrossRef]
- Aparicio, J.M.; Ortego, J.; Cordero, P.J. What should we weigh to estimate heterozygosity, alleles or loci? Mol. Ecol. 2006, 15, 4659–4665. [Google Scholar] [CrossRef]
- Chapman, J.R.; Nakagawa, S.; Coltman, D.W.; Slate, J.; Sheldon, B.C. A quantitative review of heterozygosity-fitness correlations in animal populations. Mol. Ecol. 2009, 18, 2746–2765. [Google Scholar] [CrossRef]
- Packard, G.C.; Boardman, T.J. The use of percentages and size-specific indices to normalize physiological data for variation in body size: Wasted time, wasted effort? Comp. Biochem. Physiol. A 1999, 122, 37–44. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- R Core Team. R: A language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.R-project.org/. (accessed on 15 February 2025).
- Rueffler, C.; Van Dooren, T.J.M.; Leimar, O.; Abrams, P.A. Disruptive selection and then what? Trends Ecol. Evol. 2006, 21, 238–245. [Google Scholar] [CrossRef]
- Khan, R.A. Host-parasite interactions in some fish species. J. Parasitol. Res. 2012, 237280. [Google Scholar] [CrossRef] [PubMed]
- Krkosek, M.; Revie, C.W.; Gargan, P.G.; Skilbrei, O.T.; Finstad, B.; Todd, C.D. Impact of parasites on salmon recruitment in the Northeast Atlantic Ocean. Proc. Biol. Sci. 2013, 280, 20122359. [Google Scholar] [CrossRef] [PubMed]
- Ortego, J.; Aparicio, J.M.; Calabuig, G.; Cordero, P.J. Risk of ectoparasitism and genetic diversity in a wild lesser kestrel population. Mol Ecol. 2007, 16, 3712–3720. [Google Scholar] [CrossRef]
- Fraser, B.A.; Neff, B.D. Parasite mediated homogenizing selection at the MHC in guppies. Genetica 2010, 138, 273–278. [Google Scholar] [CrossRef]
- Mäkinen, H.S.; Cano, J.M.; Merilä, J. Identifying footprints of directional and balancing selection in marine and freshwater three-spined stickleback (Gasterosteus aculeatus) populations. Mol Ecol. 2008, 17, 3565–3582. [Google Scholar] [CrossRef]
- Hedrick, P.W. Balancing selection. Curr. Biol. 2007, 17, 230–231. [Google Scholar] [CrossRef]
- Castro-Fernández, J.; Castejón-Silvo, I.; Arechavala-Lopez, P.; Terrados, J.; Morales-Nin, B. Feeding ecology of pipefish species inhabiting Mediterranean seagrasses. Mediterr. Mar. Sci. 2020, 21, 705–718. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Morkune, R.; Marcos, C.; Pérez-Ruzafa, I.; Razinkovas-Baziukas, A. Can an oligotrophic coastal lagoon support high biological productivity? Sources and pathways of primary production. Mar. Environ. Res. 2020, 153, 104824. [Google Scholar] [CrossRef]
- Gilabert, J. Seasonal plankton dynamics in a Mediterranean hypersaline coastal lagoon: The Mar Menor. J. Plankton Res. 2001, 23, 207–218. [Google Scholar] [CrossRef]
- Feist, S.W.; Longshaw, M. Histopathology of fish parasite infections – importance for populations. J. Fish Biol. 2008, 73, 2143–2160. [Google Scholar] [CrossRef]
- Alvarez-Pellitero, P. Fish immunity and parasite infections: From innate immunity to immunoprophylactic prospects. Vet. Immunol. Immunopathol. 2008, 126, 171–198. [Google Scholar] [CrossRef] [PubMed]
- Piazzon, M.C.; Galindo-Villegas, J.; Pereiro, P.; Estensoro, I.; Calduch-Giner, J.A.; Gómez-Casado, E.; Novoa, B.; Mulero, V.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Differential modulation of IgT and IgM upon parasitic, bacterial, viral, and dietary challenges in a perciform fish. Front. Immunol. 2016, 7, 637. [Google Scholar] [CrossRef]
- Holzer, A.S.; Piazzon, M.C.; Barrett, D.; Bartholomew, J.L.; Sitjà-Bobadilla, A. To react or not to react: The dilemma of fish immune systems facing Myxozoan Infections. Front. Immunol. 2021, 12, 734238. [Google Scholar] [CrossRef]
- Eizaguirre, C.; Lenz, T.L. Major histocompatibility complex polymorphism: Dynamics and consequences of parasite-mediated local adaptation in fishes. J. Fish Biol. 2010, 77, 2023–2047. [Google Scholar] [CrossRef]
- Daub, J.T.; Hofer, T.; Cutivet, E.; Dupanloup, I.; Quintana-Murci, L.; Robinson-Rechavi, M.; Excoffier, L. Evidence for polygenic adaptation to pathogens in the human genome. Mol. Biol. Evol. 2013, 30, 1544–1558. [Google Scholar] [CrossRef]
- Hartwell, L.H; Goldberg, M.L.; Fischer, J.A.; Hood, L.; Aquadro, C.F. Genetics: From genes to genomes, 5th ed.; McGraw-Hill Education: New York, NY, USA, 2014; p. 816. [Google Scholar]
- Spiering, A.E.; de Vries, T.J. Why females do better: The X chromosomal TLR7 gene-dose effect in COVID-19. Front. Immunol. 2021, 12, 756262. [Google Scholar] [CrossRef]
- Polyakova, T.A.; Kornyychuk, Y.M.; Pronkina, N.V. Checklist of Syngnathidae parasites in the Black Sea and the Sea of Azov. Inland Water Biol. 2023, 16, 1141–1158. [Google Scholar] [CrossRef]
- Kornyychuk, Y.; Anufriieva, E.; Shadrin, N. Diversity of parasitic animals in hypersaline waters: A review. Diversity 2023, 15, 409. [Google Scholar] [CrossRef]
- Bullard, S.A.; Overstreet, R.M. Digeneans as enemies of fishes. In Fish Diseases, 1st ed.; Eiras, J., Segner, H., Wahli, T., Kapoor, B.G., Eds.; CRC Press: Boca Raton, FL, USA, 2008; Volume 2, pp. 817–976. [Google Scholar]
- Farinós-Celdrán, P.; Robledano-Aymerich, F.; Carreño, M.F.; Martínez-López, J. Spatiotemporal assessment of littoral waterbirds for establishing ecological indicators of Mediterranean coastal lagoons. ISPRS Int. J. Geo-Inf. 2017, 6, 256. [Google Scholar] [CrossRef]
- Franco, A.; Franzoi, P.; Malavasi, S.; Riccato, F.; Torricelli, P.; Mainardi, D. Use of shallow water habitats by fish assemblages in a Mediterranean coastal lagoon. Estuar. Coas. Shelf Sci. 2006, 66, 67–83. [Google Scholar] [CrossRef]
- Kleiber, D.; Blight, L.K.; Caldwell, I.R.; Vincent, A.C.J. The importance of seahorses and pipefishes in the diet of marine animals. Rev. Fish Biol. Fisheries 2011, 21, 205–223. [Google Scholar] [CrossRef]
- Barber, I. The role of parasites in fish-bird interactions: A behavioural ecological perspective. In Interactions between Fish and Birds: Implications for Management, 1st ed.; Cowx, I.G., Ed.; Blackwell Publishing Ltd: Hoboken, NJ, USA, 2003; Volume 1, pp. 221–243. [Google Scholar] [CrossRef]
- Shamsi, S. Parasite loss or parasite gain? Story of Contracaecum nematodes in antipodean waters. Parasite Epidemiol. Control 2019, 4, e00087. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.A.; Brassil, C.E.; Hall, S.R.; Tessier, A.J.; Cáceres, C.E.; Conner, J.K. Parasite-mediated disruptive selection in a natural Daphnia population. BMC Evol. Biol. 2008, 8, 80. [Google Scholar] [CrossRef]
- Wegner, K.M.; Kalbe, M.; Kurtz, J.; Reusch, T.B.H.; Milinski, M. Parasite selection for immunogenetic optimality. Science 2003, 301, 1343. [Google Scholar] [CrossRef]
Figure 1.
Map of the study area showing the geographic location (coordinates: 37º45’N–0º47’W) for the Mar Menor coastal lagoon (SE Spain). Black dots: sampling sites.
Figure 1.
Map of the study area showing the geographic location (coordinates: 37º45’N–0º47’W) for the Mar Menor coastal lagoon (SE Spain). Black dots: sampling sites.
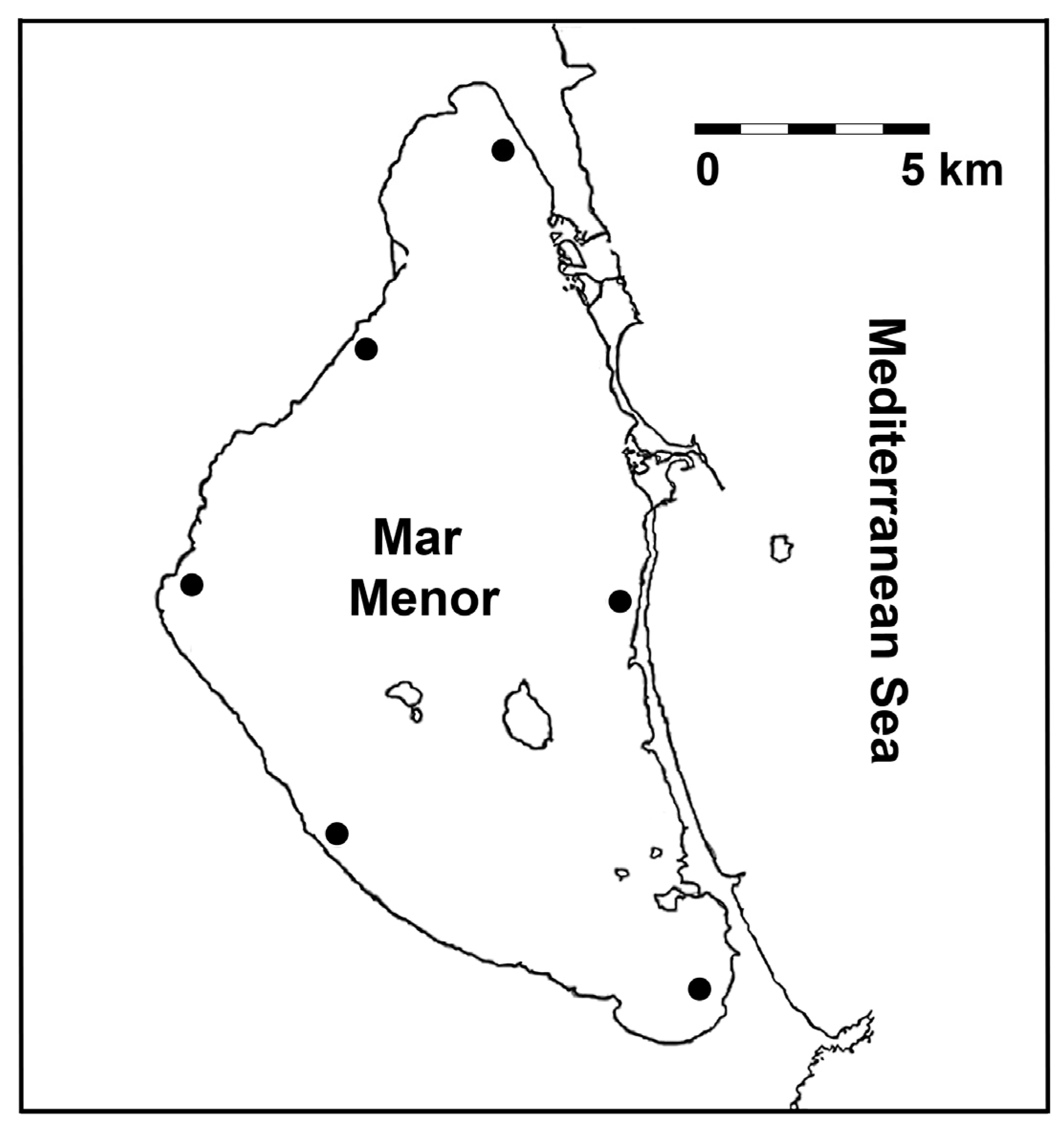
Figure 2.
Population frequencies for black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. Results are percentages of fish individuals, reported per category (‘level’) of genetic diversity.
Figure 2.
Population frequencies for black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. Results are percentages of fish individuals, reported per category (‘level’) of genetic diversity.
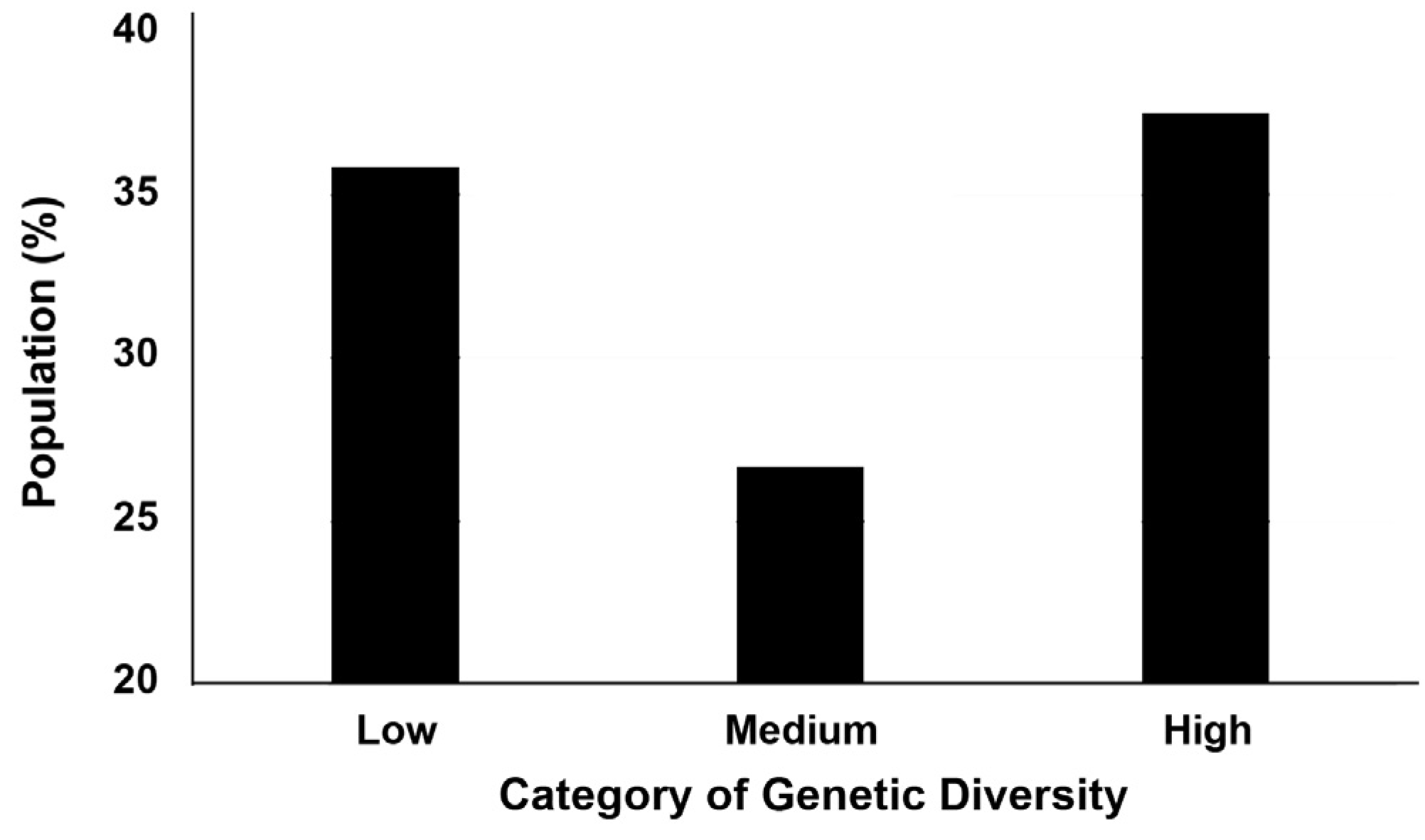
Figure 3.
Parasite prevalences for black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. Results are percentages of parasitised fish individuals, reported per category (‘level’) of genetic diversity.
Figure 3.
Parasite prevalences for black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. Results are percentages of parasitised fish individuals, reported per category (‘level’) of genetic diversity.
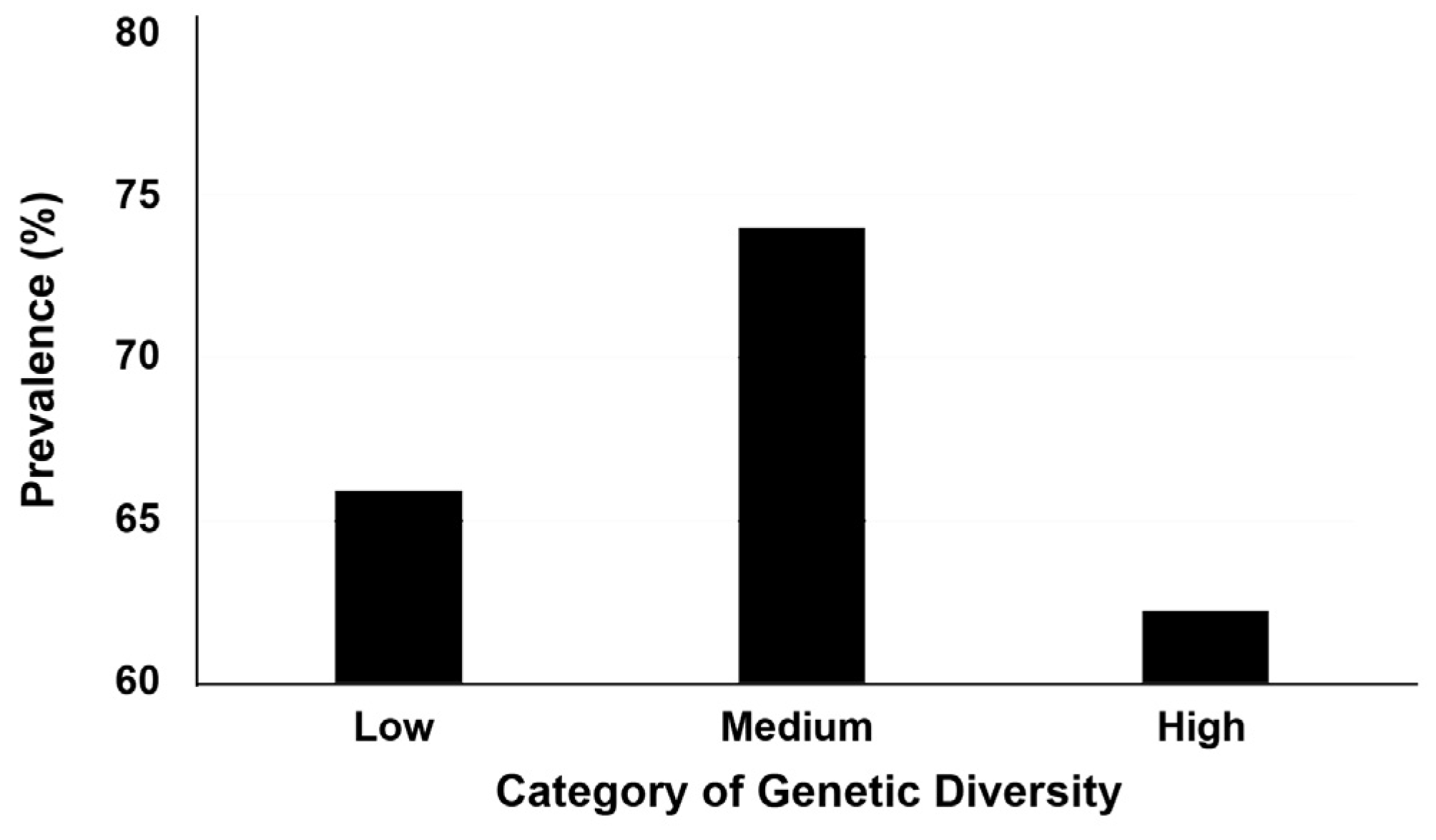

Table 1.
Description of the microsatellites (n = 10) used in this study to estimate genetic diversity in black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. For each pair of primers (F-Forward, R-Reverse), the following information is provided: locus name and GenBank ID, specific sequences, fluorophore name (F/R-attached), allelic size range (Size), allelic richness (NA), core motif and bibliographic reference.
Table 1.
Description of the microsatellites (n = 10) used in this study to estimate genetic diversity in black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. For each pair of primers (F-Forward, R-Reverse), the following information is provided: locus name and GenBank ID, specific sequences, fluorophore name (F/R-attached), allelic size range (Size), allelic richness (NA), core motif and bibliographic reference.
| locus (GenBank ID) | Primers 5’–3’ | Size | NA | Core Motif | References |
|---|---|---|---|---|---|
| Sabas1 (GQ168557) | F-FAM: GGCATGTATCAAACGAATGTCCTC | 147–451 | 28 | (ATCT)32 | [64] |
| R: TTTGCGAAACAGTGTATGACTATACTT | |||||
| Sabas2 (GQ168558) | F-ATTO565: CAGGTTGTTGACCATTTGAGTGT | 170–262 | 11 | (TC)33 | [64] |
| R: CGAATATGATTATGTGAGTCCTAAGGC | |||||
| Sabas3 (GQ168559) | F: TTCCCCCTAGGACCAATAAAGTATCT | 149–279 | 21 | (ATCT)37 | [64] |
| R-Bodipy530: TGAGAGTGGTTGCCTCCAGC | |||||
| Sabas4 (GQ168560) | F: CAAAATGCAAGTGATCCTGTGTAGG | 207–298 | 27 | (TCTA) 38 | [64] |
| R-ATTO550: TGGTGTGGTGGAACTGAATGACG | |||||
| Sabas5 (GQ168561) | F: CATTGAAACTGCATTGATTTTATGATT | 249–360 | 29 | (ATCT)44 | [64] |
| R-FAM: AGGGGGTTGTAAGTCTTTGTG | |||||
| Sabas6 (GQ168562) | F-ATTO565: TCGTGTTCCGGGACGCACATGG | 244–451 | 35 | (AGAT)4CTAT(ATCT)31 | [64] |
| R: ATGTCCGAGGTCAAACACGGCGA | |||||
| Sabas7 (GQ168563) | F-Bodipy530: CGATGTGCGAGACCTGTTGCG | 184–322 | 24 | (GATA)33 | [64] |
| R: AAAGAGGCGGAGCTTGTGTAAGGA | |||||
| Sabas8 (GQ168564) | F: FAM-TATGTGTGCCCTGCGACTGGTTGR: CAGGAGATAAGGGAGCGTTTATAGCGG | 187–522 | 31 | (TAGA)42 | [64] |
| Sabas9 (GQ168565) | F: TGATTTGGAATGACACGGGTGGTTTG | 200–276 | 17 | (ATAG)33 | [64] |
| R-FAM: TCGTTTTGTGTGCACCGAGTGTT | |||||
| typh16 (–) | F-ATTO565: CAGGACACGCTGGAAAGAC | 223–307 | 20 | (GATG)15 | [65] |
| R: GCAACACCTTGAAGAGGAAAGT |
Table 2.
Morphological traits of health status (BC and HAI) for black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. Results are (adjusted) means ± SD, reported per category (‘level’) of genetic diversity. F-ratios, degrees of freedom and significance levels (p-values) are presented, after ANCOVA (covariates: SL/eBM, see ‘Data Analyses’ for details). Significant results are highlighted in bold. Lower-case superscript letters indicate significant differences between categories (Tukey’s HSD test: p < 0.05).
Table 2.
Morphological traits of health status (BC and HAI) for black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. Results are (adjusted) means ± SD, reported per category (‘level’) of genetic diversity. F-ratios, degrees of freedom and significance levels (p-values) are presented, after ANCOVA (covariates: SL/eBM, see ‘Data Analyses’ for details). Significant results are highlighted in bold. Lower-case superscript letters indicate significant differences between categories (Tukey’s HSD test: p < 0.05).
| S. abaster | Low | Medium | High | F2,356 | p-value |
|---|---|---|---|---|---|
| BC (mg) | 411.5 ± 200.90 | 314.1 ± 190.85 | 402.5 ± 211.62 | 2.36 | 0.095 |
| HAI (points) | 24.2 ± 13.16 | a 35.3 ± 14.07 | 25.4 ± 15.55 | 4.95 | 0.008 |
Table 3.
Parasite list found in black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. Mean values of parasite abundances are shown per Family, Genus or Species, and category (‘level’) of genetic diversity. Dashes indicate parasites were undetected on examined pipefish individuals (n = 85, 71 and 84 for the low, medium and high levels of genetic diversity; respectively).
Table 3.
Parasite list found in black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. Mean values of parasite abundances are shown per Family, Genus or Species, and category (‘level’) of genetic diversity. Dashes indicate parasites were undetected on examined pipefish individuals (n = 85, 71 and 84 for the low, medium and high levels of genetic diversity; respectively).
| Parasite (Taxon) | Parasite (Family/Genus/Species) | Low | Medium | High |
|---|---|---|---|---|
| Ciliophora | Trichodina partidisci | 0.7 | 0.3 | 0.7 |
| Cestoda | Proteocephalus sp. | 0.3 | 0.5 | 0.4 |
| Monogenea | Gyrodactylus sp. | 2.2 | 0.6 | 2.9 |
| Digenea | Cryptocotyle concava | 1.5 | 2.8 | 1.2 |
| Cyathocotylidae | 3.8 | 7.3 | 4.4 | |
| Diplostomum sp. | 1.3 | 2.2 | 1.5 | |
| Echinochasmus perfoliatus | – | 1 | 0.2 | |
| Podocotyle atherinae | 2.7 | 3.9 | 2.5 | |
| Timoniella imbutiformis | 0.9 | 1.7 | – | |
| Nematoda | Contracaecum microcephalum | 0.1 | 0.8 | 0.3 |
| Acanthocephala | Paracanthocephaloides incrassatus | – | 0.3 | 0.6 |
| Pomphorhynchus laevis | 1.3 | 0.2 | 1.1 | |
| Crustacea | Ergasilus ponticus | 2.9 | 1.3 | 2.7 |
Regarding the two parameters of parasite infra-communities (TA and LCI), adjusted values (covariate: eBM) were always higher (>20 parasite specimens and ≈2 host species, respectively per parameter) for the medium group relative to the low and high levels (≈18 and ≈1.6, respectively per parameter) of genetic diversity. Such differences were close to the statistical significance (ANCOVA: p < 0.08; see Table 4) for TA and were clearly significant (ANCOVA: p ≈ 0.02; see Table 4) for LCI. In particular for this parasite index of complexity, the medium group was significantly different with respect to the low and high levels (Tukey’s HSD test: p < 0.05; see Table 4).
Table 4.
Parasitological traits (TA and LCI) for black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. Results are (adjusted) means ± SD, reported per category (‘level’) of genetic diversity. F-ratios, degrees of freedom and significance levels (p-values) are presented, after ANCOVA (covariate: eBM, see ‘Data Analyses’ for details). Significant results are highlighted in bold. Lower-case superscript letters indicate significant differences between categories (Tukey’s HSD test: p < 0.05).
Table 4.
Parasitological traits (TA and LCI) for black-striped pipefish Syngnathus abaster from the Mar Menor coastal lagoon. Results are (adjusted) means ± SD, reported per category (‘level’) of genetic diversity. F-ratios, degrees of freedom and significance levels (p-values) are presented, after ANCOVA (covariate: eBM, see ‘Data Analyses’ for details). Significant results are highlighted in bold. Lower-case superscript letters indicate significant differences between categories (Tukey’s HSD test: p < 0.05).
| S. abaster | Low | Medium | High | F2,236 | p-value |
|---|---|---|---|---|---|
| TA (number of specimens) | 17.7 ± 8.68 | 22.9 ± 9.37 | 18.5 ± 8.36 | 2.59 | 0.077 |
| LCI (range: 1–3 host species) | 1.57 ± 0.685 | a 1.99 ± 0.701 | 1.56 ± 0.806 | 3.92 | 0.021 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



