
Submitted:
17 March 2025
Posted:
18 March 2025
You are already at the latest version
Abstract
The overwhelming amount of cassava residues and okara is a foremost challenge for the food processing industry environmental loading. The purpose of this article is to utilize Pleurotus ostreatus mycelium to ferment solid cassava and soybean residue, resulting in mycelial biomass with nutritional values and promising prebiotic activities from fermented waste-sources. By a blending ratio of 80% cassava and 20% soybean residues, the mycelium spread rapidly after 3 days of culture, from 1.73 mm on the first day to 13.32 mm on the third day, and completely covered the surface after 9 days of culture (120 mm). Using the SSF method allowed us to improve the content of substances isolated from mycelium biomass, where polysaccharide content rose by 2.1 times to 3.44 mg/g, and the protein content increased by 1.84 times over the initial substrate. The prebiotic activity of isolated-PS was greatest in P. acidilactici NBD8 (1.58); for L. pentosus NH1, L. argentoraten NH15, and L. plantarum WCFS1 strains, the indices were 0.11, 0.17, and 0.3, respectively. SSF process with P. ostreatus mycelium has the potential to be an effective method for improving the nutrition and digestibility of soybean and cassava residues for application in the production of nature-derived animal feed, as well as contributing to fully utilize agricultural residue, agriculture's circular economy, reducing environmental issues, and achieve net-zero carbon emissions target by 2050, as Vietnam government committed during the COP26 World Leaders' Summit in 2021.
Keywords:
Oyster mushroom
; Pleurotus ostreatus
; polysaccharide
; prebiotic activity
; soybean residue
; cassava residue
; solid-state fermentation
1. Introduction
Solid state fermentation (SSF) refers to a fermentation process in which microorganisms grow on a solid substrate that lacks or limited free water, and complex materials are transformed into simpler ones [1,2,3]. It is considered one of the technologies for conversion of agricultural by-products into valuable secondary products for nutrition and human health [4,5], animal feed [6,7,8], biofuel [9,10,11,12], and bio-protection for grain crops [13,14,15]. SSF regularly employs agricultural and agro-industrial by-products as low-economic-value raw materials rather than discarding them as an environmental concern; these materials are used as substrates in the fermentation process [16,17]. Various agricultural substrates have been successfully used in SSF to produce ligninolytic enzymes, which decompose lignin, a major contributor to the total carbon of agro-industrial wastes, and produce polycyclic aromatic hydrocarbon compounds that can inhibit DNA synthesis and induce cancerous tumors in the liver, lung, larynx, and cervix in animals and humans [18,19]. Pleurotus ostreatus (P. ostreatus) is one of significant edible mushrooms, capable of decomposing lignocellulose without the need for chemical or biological preparation due to its enzymatic complex system, which contains phenol oxidases and peroxidases [1,20].
In Vietnam, the cassava starch and tofu processing, and soybean milk production industries are large-scale sectors that generate significant amounts of residue. This source is often only partially used in its fresh form as feed for livestock or solid compost, with the majority not being utilized, leading to environmental pollution and the wastage of this recyclable resource. Soybean residue (also called okara) and cassava residues retain a substantial amount of fiber and nutrients, making them suitable substrates for the growth of edible and medicinal mushrooms. Wet okara contains about 6% fat, 8.08% protein, 12.01% carbohydrates, 5% dietary fiber, 1% ash, and 32.05% total solid, meanwhile, these compositions in dried okara were 12.06, 34.15, 48.9, 33, 2.05, and 95.04%, respectively [21,22,23]. In other studies, it was reported that, okara had an amount of 10% lipid, 25% protein, and 50% fiber [24,25,26]. As for cassava residue, Yimin (2015) announced that, crude protein and neutral detergent fiber content were 1.30 – 16.41% and 25.4 – 52.9% on a dry matter basis, respectively [27]. Parts of cassava plant contain different concentrations of lysine, ranged from 3.9 g/100g in leaves to 7.2 g/100g dry weight in roots, soluble and insoluble dietary fiber contents in cassava pulp by-products reached 2.29% and 15.07%, respectively [28,29]. In another study, it was reported that cassava residue had a high organic content, including approximately 2.19% crude protein, 60.37% starch, 0.52% crude lipid, total minerals at 2.01%, and 21.01% fiber [30].
As for recent achievements around the world, Zhu (2020) designed, synthesized, and characterized superabsorbent hydrogels that had high capacities of water absorbency, holding, and retention through graft copolymerization of soybean residue with acrylic acid and acrylamide [31]. Sabater (2020) made a minireview to introduce new ways of valorization of vegetable food waste and by-products through fermentation processes to improve nutritional value, or to produce biologically active compounds from those waste and by-products [32]. Oktaviani (2021) applied bioconversion of cassava peel residue into yeasts to produce cell wall mannoprotein as an antioxidant [33]. Suriyapha (2022) delved into how to make bioconversion of agro-industrial residues as a protein source supplementation for cows. In that study, authors tried to compare the effects of Holstein Thai crossbreed cows with citric waste fermented yeast waste on absorption, digestibility, fermentation activity in the stomach of ruminants, blood metabolites, cognation of purine, milk production, and economical efficiency of tropical lactating cows [34]. Verardi (2023) reviewed research on agricultural residue recovery through fermentation technology, analyzed the key steps in the agro-residue bioconversion process, and the most common microorganisms employed in this procedure [35]. Bala (2023) addressed pathways to convert agro-residues, into valuable bioproducts and bioactive compounds, as well as their applications [36]. Blasi (2023) reviewed the valorization methods employed for biotransformation of lignocellulosic agricultural waste into economically and environmentally valuable products, such as the production of biofuels, the synthesis of platform chemicals; the creation of materials that is composed entirely of matter taken from living microorganisms; the production of extracellular fungal enzymes, organic acids, and compounds that has a biological activity [37]. Cruz (2021) and Adnane (2024) provided a thorough examination or evaluation of anaerobic co-digestion technology as a biochemical recovery pathway of cassava residue and other agricultural residues for the production biogas that fulfill the global target of renewable energy [38,39]. In this study, we proposed a method for bioconversion of cassava and soybean residues to derive valuable bio-substances, such as polysaccharide (PS) and protein by the purple P. ostreatus mycelium through SSF, moreover the prebiotic activity of PS isolated from fermented substrate was also evaluated.
P. ostreatus is one of the most popular mushrooms cultivated in Vietnam due to eating culture and humid tropical monsoon conditions. It contains a variety of nutrients and bioactive substances such as proteins, fibers, lipids, carbohydrates, minerals, vitamins, and bioactive compounds like PS, phenols, and flavonoids [40]. The applications of PS obtained from mushroom mycelium mainly focus on antioxidant activities, anti-tumor effects, immune modulation, stimulation of macrophage functions, and lung cell protection [41]. The biologically active PS in purple oyster mushroom was reported to occur from the mycelial growth stage without the need for the fruiting body to fully develop [42]. Thus, SSF to harvest mycelial biomass may shorten the cultivation cycle, easily control the cultivation conditions on an industrial scale to obtain a large amount of mycelial biomass, opening prospects for the development of diverse substrate sources for producing mushroom fiber rich in polysaccharides, both of high medicinal value and nutritional value for use in the livestock industry, and reduce environmental issues caused by agricultural residue.
2. Materials and Methods
2.1. Cassava and Soybean Residue Sources
Fresh cassava residue was collected from a tapioca starch processing factory in Quang Nam province (Q86F+498, ĐT611, Que Cuong commune, Que Son district, Quang Nam, Vietnam). Fresh soybean residue was sampled from Hoa Khanh market (Dong Ke Street, Hoa Khanh Bac town, Lien Chieu district, Danang city, Vietnam). These residues then were sun-dried and ground into fine powder. The purple oyster mushroom (P. ostreatus) was provided by the Laboratory of Biology – Environment, University of Science and Education – The University of Danang (459 Ton Duc Thang Street, Lien Chieu district, Danang, Vietnam). Probiotic strains, Lactiplantibacillus plantarum WCFS1, Lactiplantibacillus pentosus NH1, Lactiplantibacillus argentoraten NH15, Pediococcus acidilactici NBD8, and the pathogenic strain Escherichia coli ATCC 85922 were used to assess the prebiotic activity of polysaccharides fermented from mushroom mycelia. Commercial prebiotics (FOS, inulin, and GOS) were distributed by Southeast Asia Pharmaceutical and Trading company (46 Lot 5, Den Lu 2 Urban Area, Hoang Van Thu Ward, Ha Noi, Ha Noi Vietnam).
2.2. Cultivation of Pleurotus ostreatus Mycelium in Liquid Medium
The PDB+ medium was prepared by adding 200 g of potatoes, 20 g of D-glucose, 2 g of peptone, and 2 g of yeast extract into 1 L of distilled water, sterilized twice. 100 mL of PDB+ medium was dispensed into 250 mL Erlenmeyer flasks and then sterilized at 121°C for 30 minutes. After cooling, a uniform piece of mycelium (approximately 1 cm²) was inoculated into each flask from a first-generation culture tube to proceed with the shake culture at a speed of 150 rpm at a temperature range of 25 to 28°C. After 7 days, the mycelium was harvested.
2.3. Solid-State Fermentation of Cassava and Soybean Residue with Pleurotus ostreatus Mycelium
The experiment was conducted to evaluate the growth of the Pleurotus ostreatus mycelium on various experimental treatments containing a mixture of soybean and cassava residues as detailed in Table 1.
100 g of the substrate in each formula was placed into plastic boxes (with an upper-diameter of 12 cm and bottom-diameter of 10 cm) to sterilize at 121°C, 1 atm, for 20 minutes. After cooling down, 5 mL of the liquid mycelium solution was inoculated into each box. The cultivation phase of the mushroom mycelium was maintained at a temperature range of 25 – 28°C, and subsequently, the spread rate and morphological characteristics of the mycelium were evaluated after 15 days of cultivation.
2.4. Extraction and Quantification of PS
Taking 25 g of dry, finely ground mycelial biomass obtained after the SSF and mixing it with 375 mL of distilled water, followed by incubation at 70°C for 3 hours. The mixture was then vigorously shaken and filtered through Whatman filter paper, and the filtrate was precipitated using 96% alcohol in a 1:4 ratio for 12 – 24 hours at 4°C. Subsequently, the mixture was centrifuged at 10,000 rpm for 10 minutes. The supernatant was discarded, and the sediment was dissolved in 1M NaOH at 60°C for one hour. The quantification of PS was conducted using the phenol-sulfuric acid method [43].
2.5. Assessment of the Impact of PS on the Growth of Gut Probiotics
The experimental medium used to evaluate the impact of PS on the growth of beneficial gut bacteria were prepared as follows: GF: MRS medium with glucose removed; PS: GF medium supplemented with 1 g/L PS; Pre: GF medium supplemented with 1 g/L commercial prebiotic (used as a control); Glc: GF medium supplemented with 1 g/L glucose; GlPs: Glc medium supplemented with 1 g/L PS; and GlPre: Glc medium supplemented with 1 g/L commercial prebiotic.
The probiotic strains were inoculated into the above mediums and then cultivated at 37°C for 48 hours. Subsequently, the cell density of the experimental treatments was determined by measuring the optical density at a wavelength of 600 nm at time points of 0 hours, 24 hours, and 48 hours [44].
2.6. Calculation of Prebiotic Index
The prebiotic activity of PS extracted from the mycelium after the SSF of cassava and soybean residues was evaluated through the prebiotic index (PI). The beneficial bacterial strains were cultured in GF, PS, Pre, and Glc mediums. The strain E. coli was cultured in M9 medium with glucose eliminated, supplemented with 1 g/L PS, 1 g/L glucose, and 1g/L commercial prebiotic. The bacterial strains were incubated at 37°C for 48 hours. The optical density of the liquid samples at a wavelength of 600 nm (OD600) was measured to determine the biomass accumulation capacity. The PI was calculated using the following formula [45].
where, , , , and were OD600 values of beneficial gut bacteria cultured in Pre; GF; and Glc medium at 0 hours and 24 hours. , , and were OD600 values of E. coli cultured in Pre; Glc; and GF medium at 0 hours and 24 hours.
2.7. Assessment of Mycelial Biomass Quality
The protein content was determined according to ISO 5983-2: 2009 for livestock feed - this standard specifies the method for determining nitrogen content and calculating crude protein content using the Kjeldahl method [46]. Lipid content was analyzed in accordance with ISO 6492:1999 for livestock feed - this specifies the determination of fat content by hexane extraction [47]. The crude ash content was analyzed according to ISO 5984:2022 for livestock feed [48].
2.8. Data Analysis
Each experimental formula was replicated three times, with each replication consisting of five identical treatments. Descriptive statistics were used to determine mean values and standard deviations. The treatments were compared using one-way analysis of variance (ANOVA) and the Tukey HSD Test for a significance level of 95%.
3. Results and Discussion
3.1. Impact of the Cultivation Substrate on the Growth of Pleurotus ostreatus Mycelium
Agricultural residue typically contains a variety of substances and numerous nutrients remaining after the processing phase. These provide a rich source of raw materials for SSF to produce mycelial biomass from various types of mushrooms. The nutrient content within the substrate significantly influences the growth and development of the mushroom mycelium during SSF. Changes in the diameter of the Pleurotus ostreatus mycelium on the surface of the substrate in the treatments during the SSF process are depicted in Table 2 and Figure 1.
The findings indicated that the mixing ratio between cassava and soybean residues significantly affects the spread rate and morphology of the mushroom mycelium. Treatments with a higher ratio of cassava residue showed rapid mycelium spread and completed occupying of the whole substrate surface quicker than those with a higher ratio of soybean residue. In treatments CT1, CT2, and CT3, the mycelium spread rapidly and covered the surface of the medium after 9 days of cultivation (120 mm). In CT4 and CT5, the spread rate was slower, the mycelium covering approximately 90% and 70.24% of the substrate surface after 9 days (108.07 mm and 84.29 mm, respectively). CT1 (100% cassava residue) showed the fastest spread rate, with the mycelium covering 92% of the surface by the 7th day (110.62 mm), followed by CT2 and CT3 with about 70% (83.18 mm and 82.51 mm, respectively) (Table 2).
According to Nair (2012), the high fiber and carbohydrates content (14.88 g/100g and 63.85 g/100g dry weight), good thermal stability, and high crystallinity in cassava residue might make the substrate more porous, allowing the mycelium to develop and spread more quickly [49]. This can explain the early surface coverage of mycelium observed in CT1, CT2, and CT3, because soybean residue contains mostly crude fiber composed of cellulose, hemicellulose, and lignin, about 25% protein, 10-15% oil, but little starch or simple carbohydrates [50]. Hence, a high proportion of soybean residue in treatments will result in increasing compaction and reduce the porosity of the substrate, while the diversity and richness in nutrients remaining in the soybean residue often led to slower mycelium development due to a lack of oxygen for respiration and decomposition of complex compounds. This demonstrated the slower mycelium spread on the substrate in treatments CT4 and CT5 where soybean residue occupied 30% and 40%, respectively.
Sensory evaluation of the mycelium thickness on the substrate surface revealed distinct differences among the treatments. Pleurotus ostreatus mycelium in CT3, CT4, and CT5 had the highest density, was fluffy, evenly white, while CT1 and CT2 had a lower density. This might be due to the rich and higher nutrients content in soybean residue compared to those in cassava residue [28,50] with high levels of nitrogen and vitamins, facilitating the growth and biomass accumulation of the mycelium during fermentation. The productivity and quality of the mycelium depend on the nutritional status from the substrate source, such as the C/N ratio, vitamins, plant hormones, and trace and macro minerals [51].
After 30 days of cultivation, the mycelium had covered the entire substrate in all treatments (Figure 1). In CT4 and CT5, although the mycelium had covered the surface of the substrate, the bottom of the boxes still had sparse mycelium, and some boxes showed no further mycelium spread upon continued cultivation. In CT1 and CT2, the low nutrient content might have led to low mycelial biomass and the mycelium drying out and dying over time. In contrast, for CT3, the mycelium developed fully and appeared fluffy and white across the box when the SSF period was extended, showing signs of fruiting body development on the substrate surface. Therefore, the 80% cassava + 20% soybean residue mix in CT3 was selected for further experiments.
3.2. Impact of the Cultivation Substrate on the Growth of Pleurotus ostreatus Mycelium
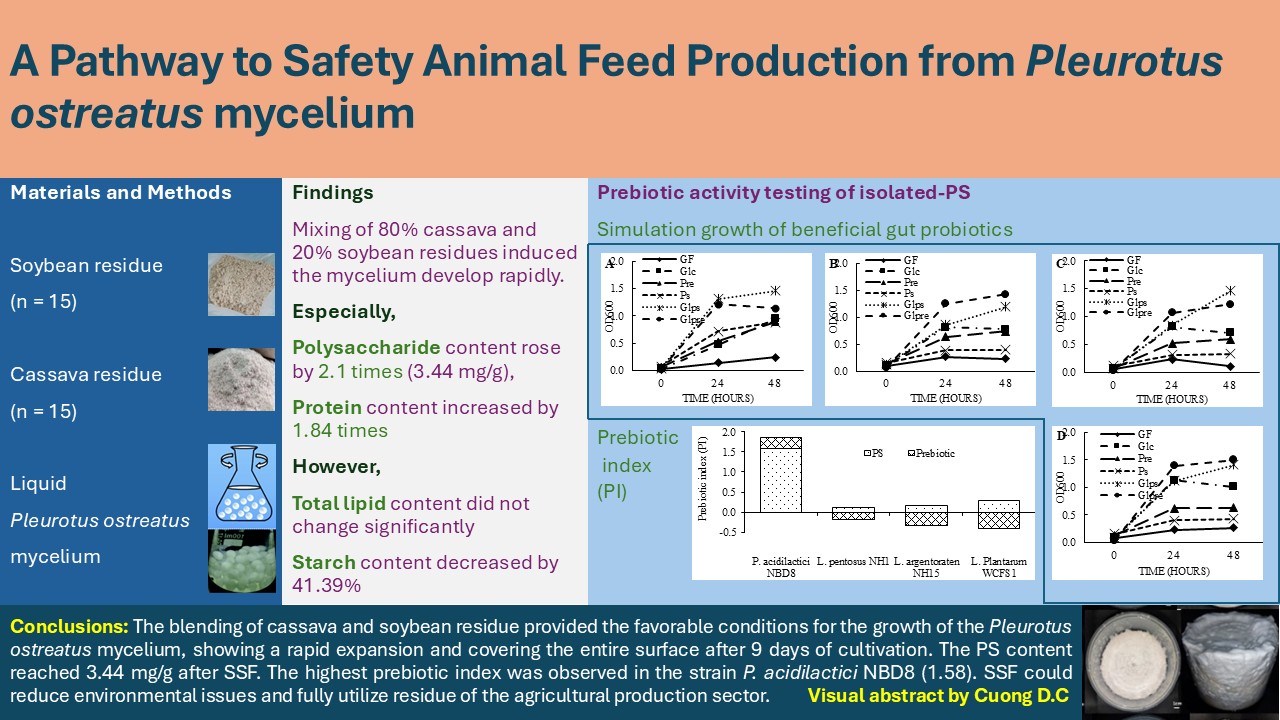
As presented in Figure 2, the PS content in the substrate after SSF increased approximately 2.12 times compared to the unfermented raw substrate (from 1.62 mg/100g to 3.44 mg/100g). The increase in PS content could be attributed to the mycelium utilizing a rich carbon source, including compounds such as starch, cellulose, and lignocellulose, for growth and accumulating intracellular polysaccharides from the fungal fibers and unutilized crude polysaccharides following lignocellulose hydrolysis. The rise in PS content after SSF by the mushroom mycelium has also been demonstrated in the study by Lu (2023) when conducted on substrates of corn stalks and xyloma sawdust (a plant belonging to the Willow family, Salicaceae) using Shiitake (Lentinula edodes) and oyster mushroom (Pleurotus ostreatus) mycelium [52].
Protein, lipid, carbohydrate, and total ash content are also important quality indicators of the product after SSF. Substrates from agricultural by-products typically contain relatively low nutrient levels. However, after fermentation with mushroom mycelium, the nutritional composition was significantly improved, especially in terms of protein content [53]. In this study, there was a significant increase in protein content in the substrate after SSF, about 1.84 times (from 4.29 mg/100g to 7.91 mg/100g). This increase might be due to the high protein content (28.85%) of the oyster mushroom [54,55], thus the increase in mycelium biomass during growth led to an increase in accumulated protein content. The total lipid content in the substrate before and after fermentation did not change significantly (0.82 – 0.83 mg/100g). Meanwhile, the starch content after SSF decreased by about 41.39% (from 55.66 mg/100g to 32.62 mg/100g). Starch usually constitutes a high proportion in the raw substrate composition. During fermentation, the mushroom mycelium used starch as a carbon source for growth and transformation into other compounds, leading to a decrease in starch content in the substrate after SSF [56]. The total ash content in the substrate after fermentation increased by 1.86 times (from 1.69 mg/100g to 3.14 mg/100g). The increase in total ash content could be due to the presence of minerals in the second-stage fermentation broth of the oyster mushroom when inoculating and some minerals from the water added to the substrate to maintain moisture.
3.3. Prebiotic Activity of PS after Extraction
Prebiotics, as defined by the International Scientific Association for Probiotics and Prebiotics (ISAPP), are non-digestible food components that selectively feed beneficial microorganisms [57]. Prebiotics added to the diet will not be digested and absorbed by poultry. However, they can play a crucial role in selectively promoting the growth of certain beneficial bacteria, improving the gut microbiome, and enhancing the absorption and utilization of nutrients [58]. In the present study, we evaluated the prebiotic activity of PS extracted from the substrate after SSF with purple oyster mushroom mycelium by assessing the ability of probiotic strains to utilize PS as a nutritional source and evaluating through the prebiotic activity index.
3.3.1. The Impact of PS on the Growth of Beneficial Gut Probiotics
The prebiotic activity of PS is due to the selective metabolic capability of the probiotics (beneficial bacteria), thereby stimulating their growth [59]. The ability of probiotic strains to metabolize carbohydrates similarly to glucose is a prerequisite for assessing the prebiotic activity of a carbohydrate. In this study, four bacterial strains, P. acidilactici NBD8, L. pentosus NH1, L. argentoraten NH15, and L. plantarum WCFS1 were used to evaluate the ability to use PS extracted from the mixture of substrate and mycelium after SSF.
The three Lactiplantibacillus strains showed the highest growth rate in the medium with glucose as the carbon source (Figure 3). The control glucose medium (Glc) allowed mycelium development easily because glucose is a common metabolized sugar for living cells. However, P. acidilactici NBD8 grew better in the PS and Pre medium.
In medium supplemented with PS, all four bacterial strains showed higher growth than in those without added carbon source. Thus, P. acidilactici NBD8, L. pentosus NH1, L. argentoraten NH15, and L. plantarum WCFS1 could use PS extracted from the substrate after SSF as a carbon source. However, in the PS medium, the accumulation of mycelial biomass was lower than in the Pre medium; this could be because the PS extracted from the substrate after SSF is crude PS not yet purified and contains some insoluble impurities, whereas commercial prebiotics are purified PS proven to selectively stimulate the growth of beneficial bacteria species. Yet, P. acidilactici NBD8 showed significant growth within 24 hours. This may be due to each probiotic strain producing different amounts of hydrolyzing enzymes and having varying rates of carbon source degradation for nutrition; hence, their growth rates differ. This finding aligns with the study conducted by Phirom-On (2021), where L. plantarum SKKL1, L. plantarum TISTR 2075, and L. casei TISTR 1463 could use cello-dextrin extracted from banana peels as a carbon source alternative to glucose for growth [44].
Both GlPs and GlPre mediums strongly stimulated the growth of probiotics. For P. acidilactici NBD8 cultured on GlPs medium, growth was better than on GlPre medium. After 48 hours of cultivation in GlPre medium, bacteria entered a degradation phase, cells were decomposed, so the OD600 value decreased, while in GlPs medium, the OD600 value continued to increase.
For the three beneficial probiotic strains L. pentosus NH1, L. argentoraten NH15, and L. plantarum WCFS1, after 24 hours of cultivation, the OD600 value in GlPs medium was lower than those in GlPre medium. And after 48 hours of cultivation, the OD600 value in both mediums continued to increase, especially L. argentoraten cultured in GlPs medium had a higher OD600 value than in GlPre medium. This may be because PS extracted from mycelium contains more complex structural components than commercial prebiotics, so the probiotic strains in the adaptation phase could not immediately use them for growth. They had to accumulate a certain biomass and produce enough extracellular enzymes to degrade and utilize these prebiotics, leading to a longer time required to achieve equivalent biomass levels. This result was similar to the findings of Nguyen Thi Bich Hang (2023), when supplemented 10 mg/mL PS extracted from Cordyceps militaris mycelium to evaluate the stimulatory effect on the growth of L. plantarum strain [60].
Aida (2009) also emphasized that the mycelium of many mushroom species contains PS with prebiotic activity (such as chitin, hemicelluloses, β and α–glucan, mannan, xylan, and galactan) [61]. α– (1–3)–glucan, one of the PS extracted from the cell walls of two species P. ostreatus and P. eryngii, has been shown to stimulate the growth of bifidobacterium and Lactobacillus [62].
3.3.2. Prebiotic Index (PI)
According to Huebner (2007), the activity of a prebiotic is defined as the ability of a specific substrate, typically a prebiotic substance, to support the growth of beneficial bacteria in the gut compared to other bacteria. To evaluate the activity of a prebiotic, it is often compared to the growth on a non-prebiotic substrate, usually glucose. Therefore, a carbohydrate exhibits a positive prebiotic activity index if it is metabolized like glucose by probiotic strains; and is selectively metabolized by probiotics rather than by other gut bacteria [63].
The OD600 value measured after 24 hours of culture clearly demonstrated that the culture medium influenced the growth of both beneficial and harmful bacteria. All strains grew slowly in MRS medium without carbon (GF) with OD600 values ranged from 0.06 – 0.26 but increased faster in medium supplemented with glucose and commercial prebiotics with OD600 values ranged from 0.29 to 1.13 for the Glc medium and 0.25 to 0.64 for the Pre medium. Notably, on medium supplemented with PS, growth stimulation was greater for probiotic strains, particularly P. acidilactici NBD8 (OD600 = 0.71) than for E. coli ATCC 85922 (OD600 = 0.09) (Figure 4).
In treatments using PS as a carbon source, all probiotic strains showed positive prebiotic indices (ranging from 0.11 to 1.58). The highest prebiotic index was observed in the strain P. acidilactici NBD8 (1.58); the PI values of other the strains L. pentosus NH1, L. argentoraten NH15, and L. plantarum WCFS1 were 0.11, 0.17, and 0.30, respectively (Figure 5). The lower PI in the three Lactiplantibacillus strains could be due to their poorer growth in the medium supplemented with PS compared to the one supplemented with glucose. In the Pre medium, although the growth stimulation effect on the probiotic strains was higher than in the PS medium, the three Lactiplantibacillus strains showed negative prebiotic indices (–0.18; –0.32; and –0.41), and only the strain P. acidilactici NBD8 showed a positive PI (0.28). The reason for this negative value could be that the commercial prebiotic stimulated the growth of harmful microorganisms (E. coli ATCC 85922) as much as glucose did. According to Huebner (2007), strains L. plantarum 12006 and L. acidophilus 33200 showed negative PI when cultured on all prebiotics, except for purified GOS [63].
This study also showed that the same type of PS can have different prebiotic indices in different bacterial strains. This can be explained by the fact that different microbial strains have different metabolic capabilities, leading to variations in prebiotic indices. Probiotic strains need specific hydrolytic enzyme systems to utilize prebiotics. Therefore, the genes encoding these metabolic systems may or may not be present in different strains, leading to different prebiotic activity indices. The hydrolytic enzyme systems play a crucial role in the ability of probiotic strains to use prebiotics. Each probiotic strain may have genetic variations and permutations in these systems, which can create diversity in their ability to utilize and metabolize prebiotics. This causes variations in prebiotic indices [64].
The positive prebiotic indices indicate that the level of PS extracted from the substrate after SSF with mycelium selectively promoted the growth of the strains P. acidilactici NBD8, L. pentosus NH1, L. argentoraten NH15, and L. plantarum WCFS1. Additionally, the strain E. coli ATCC 85922 was unable to use this PS as a carbon source for growth.
4. Conclusions
The blending ratio in the experimental formula containing 80% cassava and 20% soybean residues provided the most favorable conditions for the growth of the P. ostreatus mycelium, showing a rapid expansion after 3 days of cultivation (from 1.73 mm to 13.32 mm) and covering the entire plate surface after 9 days of cultivation (120 mm). The polysaccharide content extracted from the substrate after SSF reached 3.44 mg/g, increased by 2.1 times, with protein and mineral content increasing about 1.84 times compared to the initial substrate. The highest prebiotic index was observed in the strain P. acidilactici NBD8 (1.58); whereas, in the strains L. pentosus NH1, L. argentoraten NH15, and L. plantarum WCFS1, the indices were 0.11, 0.17, and 0.30, respectively. The SSF process using P. ostreatus mycelium holds potential as an effective method to enhance the nutritional and digestibility of soybean and cassava residues for application in the production of natural-origin animal feeds, as well as contributing to the circular economy in agriculture by fully utilizing by-products of the agricultural production industry.
Author Contributions
Chi Cuong Doan: Conceptualization, software, validation, formal analysis, investigation, writing—review and editing. Nguyen Thi Bich Hang: methodology, visualization, project administration, writing—original draft preparation. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by VIETNAM MINISTRY OF EDUCATION AND TRAINING, grant number B2024.DNA.11.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data and material of this study are available on request.
Acknowledgments
The authors thank Bui Duc Thang and Truong Cong Phat for technical support at the Mushroom Laboratory, Faculty of Biology and Environmental Sciences – University of Science and Education – The University of Danang.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Arvanitoyannis, I.S.; Kassaveti, A.; Ladas, D. 6 - Food Waste Treatment Methodologies. In Food Science and Technology; Arvanitoyannis, I.S.B.T.-W.M. for the F.I., Ed.; Academic Press: Amsterdam, 2008; pp. 345–410 ISBN 978-0-12-373654-3.
- Parmar, A.B.; Patel, V.; Usadadia, S.; Rathwa, S.; Prajapati, D. A Solid State Fermentation, Its Role in Animal Nutrition: A Review. 2019, 4626–4633.
- Mitchell, D.A.; de Lima Luz, L.F.; Krieger, N.; Berovič, M. 2.25 - Bioreactors for Solid-State Fermentation. In; Moo-Young, M.B.T.-C.B. (Second E., Ed.; Academic Press: Burlington, 2011; pp. 347–360 ISBN 978-0-08-088504-9.
- Feng, X.; Ng, K.; Ajlouni, S.; Zhang, P.; Fang, Z. Effect of Solid-State Fermentation on Plant- Sourced Proteins: A Review Effect of Solid-State Fermentation on Plant-Sourced Proteins: A Review. Food Reviews International 2023. [CrossRef]
- Cano y Postigo, L.O.; Jacobo-Velázquez, D.A.; Guajardo-Flores, D.; Garcia Amezquita, L.E.; García-Cayuela, T. Solid-State Fermentation for Enhancing the Nutraceutical Content of Agrifood by-Products: Recent Advances and Its Industrial Feasibility. Food Biosci 2021, 41, 100926. [CrossRef]
- Yang, L.; Zeng, X.; Qiao, S. Advances in Research on Solid-State Fermented Feed and Its Utilization: The Pioneer of Private Customization for Intestinal Microorganisms. Animal Nutrition 2021, 7, 905–916. [CrossRef]
- Vandenberghe, L.P.S.; Pandey, A.; Carvalho, J.C.; Letti, L.A.J.; Woiciechowski, A.L.; Karp, S.G.; Thomaz-Soccol, V.; Martínez-Burgos, W.J.; Penha, R.O.; Herrmann, L.W.; et al. Solid-State Fermentation Technology and Innovation for the Production of Agricultural and Animal Feed Bioproducts. Systems Microbiology and Biomanufacturing 2021, 1, 142–165. [CrossRef]
- Rothmann, C.; Rothmann, L.; Viljoen, B.; Cason, E.D. Application of Solid-State Fermentation Using Mushrooms for the Production of Animal Feed. J Basic Microbiol 2023, 63, 1153–1164. [CrossRef]
- Farinas, C.S. Developments in Solid-State Fermentation for the Production of Biomass-Degrading Enzymes for the Bioenergy Sector. Renewable and Sustainable Energy Reviews 2015, 52, 179–188. [CrossRef]
- Onu Olughu, O.; Tabil, L.G.; Dumonceaux, T.; Mupondwa, E.; Cree, D. Optimization of Solid-State Fermentation of Switchgrass Using White-Rot Fungi for Biofuel Production. Fuels 2022, 3, 730–752.
- Khot, M.B. Solid-State Fermentation: An Alternative Approach to Produce Fungal Lipids as Biodiesel Feedstock BT - Status and Future Challenges for Non-Conventional Energy Sources Volume 2. In; Joshi, .Sanket J, Sen, R., Sharma, A., Salam, P.A., Eds.; Springer Nature Singapore: Singapore, 2022; pp. 123–137 ISBN 978-981-16-4509-9.
- Karimi, F.; Mazaheri, D.; Saei Moghaddam, M.; Mataei Moghaddam, A.; Sanati, A.L.; Orooji, Y. Solid-State Fermentation as an Alternative Technology for Cost-Effective Production of Bioethanol as Useful Renewable Energy: A Review. Biomass Convers Biorefin 2021. [CrossRef]
- Banat, I.M.; Carboué, Q.; Saucedo-Castañeda, G.; De Jesús Cázares-Marinero, J. Biosurfactants: The Green Generation of Speciality Chemicals and Potential Production Using Solid-State Fermentation (SSF) Technology. Bioresour Technol 2021, 320, 124222. [CrossRef]
- Faria, D.J.; Carvalho, A.P.; Conte-Junior, C.A. Valorization of Fermented Food Wastes and Byproducts: Bioactive and Valuable Compounds, Bioproduct Synthesis, and Applications. Fermentation 2023, 9.
- Mattedi, A.; Sabbi, E.; Farda, B.; Djebaili, R.; Mitra, D.; Ercole, C.; Cacchio, P.; Del Gallo, M.; Pellegrini, M. Solid-State Fermentation: Applications and Future Perspectives for Biostimulant and Biopesticides Production. Microorganisms 2023, 11.
- Dattatraya Saratale, G.; Bhosale, R.; Shobana, S.; Banu, J.R.; Pugazhendhi, A.; Mahmoud, E.; Sirohi, R.; Kant Bhatia, S.; Atabani, A.E.; Mulone, V.; et al. A Review on Valorization of Spent Coffee Grounds (SCG) towards Biopolymers and Biocatalysts Production. Bioresour Technol 2020, 314, 123800. [CrossRef]
- Lizardi-Jiménez, M.A.; Hernández-Martínez, R. Solid State Fermentation (SSF): Diversity of Applications to Valorize Waste and Biomass. 3 Biotech 2017, 7, 44. [CrossRef]
- Couto, S.R.; Gundı́n, M.; Lorenzo, M.; Sanromán, M.Á. Screening of Supports and Inducers for Laccase Production by Trametes Versicolor in Semi-Solid-State Conditions. Process Biochemistry 2002, 38, 249–255. [CrossRef]
- Tsutsui, T.; Hayashi, N.; Maizumi, H.; Huff, J.; Barrett, J.C. Benzene-, Catechol-, Hydroquinone- and Phenol-Induced Cell Transformation, Gene Mutations, Chromosome Aberrations, Aneuploidy, Sister Chromatid Exchanges and Unscheduled DNA Synthesis in Syrian Hamster Embryo Cells. Mutat Res 1997, 373, 113–123. [CrossRef]
- Leonowicz, A.; Matuszewska, A.; Luterek, J.; Ziegenhagen, D.; Wojtaś-Wasilewska, M.; Cho, N.S.; Hofrichter, M.; Rogalski, J. Biodegradation of Lignin by White Rot Fungi. Fungal Genet Biol 1999, 27, 175–185. [CrossRef]
- Razavizadeh, S.; Alencikiene, G.; Salaseviciene, A.; Vaiciulyte-Funk, L.; Ertbjerg, P.; Zabulione, A. Impact of Fermentation of Okara on Physicochemical, Techno-Functional, and Sensory Properties of Meat Analogues. European Food Research and Technology 2021, 247, 2379–2389. [CrossRef]
- Sengupta, S.; Chakraborty, M.; Bhowal, J.; Bhattacharya, D. Study on the Effects of Drying Process on the Composition and Quality of Wet Okara. Int J Sci Environ Technol 2012.
- Toda, K.; Chiba, K.; Ono, T. Effect of Components Extracted from Okara on the Physicochemical Properties of Soymilk and Tofu Texture. J Food Sci 2007, 72, C108-13. [CrossRef]
- Li, B.; Qiao, M.; Lu, F. Composition, Nutrition, and Utilization of Okara (Soybean Residue). Food Reviews International 2012, 28, 231–252. [CrossRef]
- Kim, H.-S.; Yu, O.-K.; Byun, M.-S.; Cha, Y.-S. Okara, a Soybean by-Product, Prevents High Fat Diet-Induced Obesity and Improves Serum Lipid Profiles in C57BL/6J Mice. Food Sci Biotechnol 2016, 25, 607–613. [CrossRef]
- Asghar, A.; Afzaal, M.; Saeed, F.; Ahmed, A.; Ateeq, H.; Shah, Y.A.; Islam, F.; Hussain, M.; Akram, N.; Shah, M.A. Valorization and Food Applications of Okara (Soybean Residue): A Concurrent Review. Food Sci Nutr 2023, 11, 3631–3640. [CrossRef]
- Yimin, C.; Napasirth, V.; Napasirth, P.; Sulinthone, T.; Phommachanh, K. Microbial Population, Chemical Composition and Silage Fermentation of Cassava Residues. Animal Science Journal 2015, 86, 842–848. [CrossRef]
- Fanelli, N.S.; Torres-Mendoza, L.J.; Abelilla, J.J.; Stein, H.H. Chemical Composition of Cassava-Based Feed Ingredients from South-East Asia. Anim Biosci 2023, 36, 908–919. [CrossRef]
- Okrathok, S.; Thumanu, K.; Pukkung, C.; Molee, W.; Khempaka, S. Extraction of Dietary Fibers from Cassava Pulp and Cassava Distiller’s Dried Grains and Assessment of Their Components Using Fourier Transform Infrared Spectroscopy to Determine Their Further Use as a Functional Feed in Animal Diets. Anim Biosci 2022, 35, 1048–1058. [CrossRef]
- Pham Kim Dang; Duong Thu Huong Protein Enrichment of Cassava Residues by Simultaneous Saccharification and Fermentation. Vietnam J. Agri. Sci. 2018, 16, 207–214.
- Zhu, J.; Tan, W.K.; Song, X.; Gao, Z.; Wen, Y.; Ong, C.N.; Loh, C.S.; Swarup, S.; Li, J. Converting Okara to Superabsorbent Hydrogels as Soil Supplements for Enhancing the Growth of Choy Sum (Brassica Sp.) under Water-Limited Conditions. ACS Sustain Chem Eng 2020, 8, 9425–9433. [CrossRef]
- Sabater, C.; Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Margolles, A. Valorization of Vegetable Food Waste and By-Products through Fermentation Processes. Front Microbiol 2020, 11. [CrossRef]
- Oktaviani, N.; Sarwono, K.A.; Utama, G. lara Bioconversion Rice Bran and Cassava Peel into Yeasts Cell Walls Mannoprotein as Environmental Friendly Antioxidant. E3S Web Conf. 2021, 249, 3004. [CrossRef]
- Suriyapha, C.; Supapong, C.; So, S.; Wanapat, M.; Cherdthong, A. Bioconversion of Agro-Industrial Residues as a Protein Source Supplementation for Multiparous Holstein Thai Crossbreed Cows. PLoS One 2022, 17, e0273916. [CrossRef]
- Verardi, A.; Sangiorgio, P.; Blasi, A.; Lopresto, C.G.; Calabrò, V. Bioconversion of Crop Residues Using Alternative Fermentation-Based Approaches. Frontiers in Bioscience 2023, 15, 17-null.
- Bala, S.; Garg, D.; Sridhar, K.; Inbaraj, B.S.; Singh, R.; Kamma, S.; Tripathi, M.; Sharma, M. Transformation of Agro-Waste into Value-Added Bioproducts and Bioactive Compounds: Micro/Nano Formulations and Application in the Agri-Food-Pharma Sector. Bioengineering (Basel) 2023, 10. [CrossRef]
- Blasi, A.; Verardi, A.; Lopresto, C.G.; Siciliano, S.; Sangiorgio, P. Lignocellulosic Agricultural Waste Valorization to Obtain Valuable Products: An Overview. Recycling 2023, 8. [CrossRef]
- Adnane, I.; Taoumi, H.; Elouahabi, K.; Lahrech, K.; Oulmekki, A. Valorization of Crop Residues and Animal Wastes: Anaerobic Co-Digestion Technology. Heliyon 2024, 10, e26440. [CrossRef]
- Cruz, I.A.; Santos Andrade, L.R.; Bharagava, R.N.; Nadda, A.K.; Bilal, M.; Figueiredo, R.T.; Romanholo Ferreira, L.F. Valorization of Cassava Residues for Biogas Production in Brazil Based on the Circular Economy: An Updated and Comprehensive Review. Clean Eng Technol 2021, 4, 100196. [CrossRef]
- Deepalakshmi, K.; Sankaran, M. Pleurotus Ostreatus: An Oyster Mushroom with Nutritional and Medicinal Properties. J Biochem Technol 2014, 5, 718–726.
- Murphy, E.J.; Rezoagli, E.; Pogue, R.; Simonassi-Paiva, B.; Abidin, I.I.Z.; Fehrenbach, G.W.; O’neil, E.; Major, I.; Laffey, J.G.; Rowan, N. Immunomodulatory Activity of β-Glucan Polysaccharides Isolated from Different Species of Mushroom - A Potential Treatment for Inflammatory Lung Conditions. Sci Total Environ 2022, 809, 152177. [CrossRef]
- Yang, B.-K.; Gu, Y.-A.; Jeong, Y.-T.; Song, C.-H. Anti-Complementary Activities of Exo- and Endo-Biopolymer Produced by Submerged Mycelial Culture of Eight Different Mushrooms. Mycobiology 2007, 35, 145–149. [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal Chem 1956, 28, 350–356. [CrossRef]
- Phirom-On, K.; Apiraksakorn, J. Development of Cellulose-Based Prebiotic Fiber from Banana Peel by Enzymatic Hydrolysis. Food Biosci 2021, 41, 101083. [CrossRef]
- Huebner, J.; Wehling, R.L.; Parkhurst, A.; Hutkins, R.W. Effect of Processing Conditions on the Prebiotic Activity of Commercial Prebiotics. Int Dairy J 2008, 18, 287–293. [CrossRef]
- International Standard Organization ISO 5983-2:2009. Animal Feeding Stuffs — Determination of Nitrogen Content and Calculation of Crude Protein Content 2009.
- International Standard Organization ISO 6492:1999. Animal Feeding Stuffs — Determination of Fat Content 1999.
- International Standard Organization ISO 5984:2022. Animal Feeding Stuffs — Determination of Crude Ash 2022.
- Nair, M.P.; Padmaja, G.; Sankarakutty, S.; Sheriff, T. Bioconversion of Cellulo-Starch Waste from Cassava Starch Industries for Ethanol Production: Pretreatment Techniques and Improved Enzyme Systems. Industrial Biotechnology 2012, 8, 300–308. [CrossRef]
- O’Toole, D.K. Characteristics and Use of Okara, the Soybean Residue from Soy Milk Production-a Review. J Agric Food Chem 1999, 47, 363–371. [CrossRef]
- Adenipekun, C.O.; Jonathan, G. Nutritional Requirements of Pleurotus Florida (Mont.) Singer, A Nigerian Mushroom. Pakistan Journal of Nutrition 2006, 5, 597–600. [CrossRef]
- Lu, X.; Zhao, Y.; Li, F.; Liu, P. Active Polysaccharides from Lentinula Edodes and Pleurotus Ostreatus by Addition of Corn Straw and Xylosma Sawdust through Solid-State Fermentation. Int J Biol Macromol 2023, 228, 647–658. [CrossRef]
- Heidari, F.; Øverland, M.; Hansen, J.Ø.; Mydland, L.T.; Urriola, P.E.; Chen, C.; Shurson, G.C.; Hu, B. Solid-State Fermentation of Pleurotus Ostreatus to Improve the Nutritional Profile of Mechanically-Fractionated Canola Meal. Biochem Eng J 2022, 187, 108591. [CrossRef]
- Tolera, K.D.; Abera, S. Nutritional Quality of Oyster Mushroom (Pleurotus Ostreatus) as Affected by Osmotic Pretreatments and Drying Methods. Food Sci Nutr 2017, 5, 989–996. [CrossRef]
- Ayimbila, F.; Keawsompong, S. Nutritional Quality and Biological Application of Mushroom Protein as a Novel Protein Alternative. Curr Nutr Rep 2023, 12, 290–307. [CrossRef]
- Tu, J.; Brennan, M.; Brennan, C. An Insight into the Mechanism of Interactions between Mushroom Polysaccharides and Starch. Curr Opin Food Sci 2021, 37, 17–25. [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat Rev Gastroenterol Hepatol 2017, 14, 491–502. [CrossRef]
- Peng, M.; Tabashsum, Z.; Anderson, M.; Truong, A.; Houser, A.K.; Padilla, J.; Akmel, A.; Bhatti, J.; Rahaman, S.O.; Biswas, D. Effectiveness of Probiotics, Prebiotics, and Prebiotic-like Components in Common Functional Foods. Compr Rev Food Sci Food Saf 2020, 19, 1908–1933. [CrossRef]
- De Figueiredo, F.C.; De Barros Ranke, F.F.; De Oliva-Neto, P. Evaluation of Xylooligosaccharides and Fructooligosaccharides on Digestive Enzymes Hydrolysis and as a Nutrient for Different Probiotics and Salmonella Typhimurium. LWT 2020, 118, 108761. [CrossRef]
- Nguyen Thi Bich Hang; Doan Chi Cuong; Dang Minh Nhat; Bui Duc Thang Prebiotic Properties of Polysaccharides Isolated from Cordyceps Militaris Mycelia. Vietnam Trade and Industry Review 2023, 4, 413–420.
- Aida, F.M.N.A.; Shuhaimi, M.; Yazid, M.; Maaruf, A.G. Mushroom as a Potential Source of Prebiotics: A Review. Trends Food Sci Technol 2009, 20, 567–575. [CrossRef]
- Synytsya, A.; Míčková, K.; Synytsya, A.; Jablonský, I.; Spěváček, J.; Erban, V.; Kováříková, E.; Čopíková, J. Glucans from Fruit Bodies of Cultivated Mushrooms Pleurotus Ostreatus and Pleurotus Eryngii: Structure and Potential Prebiotic Activity. Carbohydr Polym 2009, 76, 548–556. [CrossRef]
- Huebner, J.; Wehling, R.L.; Hutkins, R.W. Functional Activity of Commercial Prebiotics. Int Dairy J 2007, 17, 770–775. [CrossRef]
- Barrangou, R.; Altermann, E.; Hutkins, R.; Cano, R.; Klaenhammer, T. Functional and Comparative Genomic Analyses of an Operon Involved in Fructooligosaccharide Utilization by Lactobacillus Acidophilus. Proc Natl Acad Sci U S A 2003, 100, 8957–8962. [CrossRef]
Figure 1.
The development of the Pleurotus ostreatus mycelium during SSF. Which, CT1: 100% cassava residue; CT2: 90% cassava + 10% soybean residue; CT3: 80% cassava + 20% soybean residue; CT4: 70% cassava + 30% soybean residue; CT5: 60% cassava + 40% soybean residue.
Figure 1.
The development of the Pleurotus ostreatus mycelium during SSF. Which, CT1: 100% cassava residue; CT2: 90% cassava + 10% soybean residue; CT3: 80% cassava + 20% soybean residue; CT4: 70% cassava + 30% soybean residue; CT5: 60% cassava + 40% soybean residue.

Figure 2.
Content of PS, protein, starch, total lipid, and total ash (mg/100g) in the substrate before and after SSF.
Figure 2.
Content of PS, protein, starch, total lipid, and total ash (mg/100g) in the substrate before and after SSF.
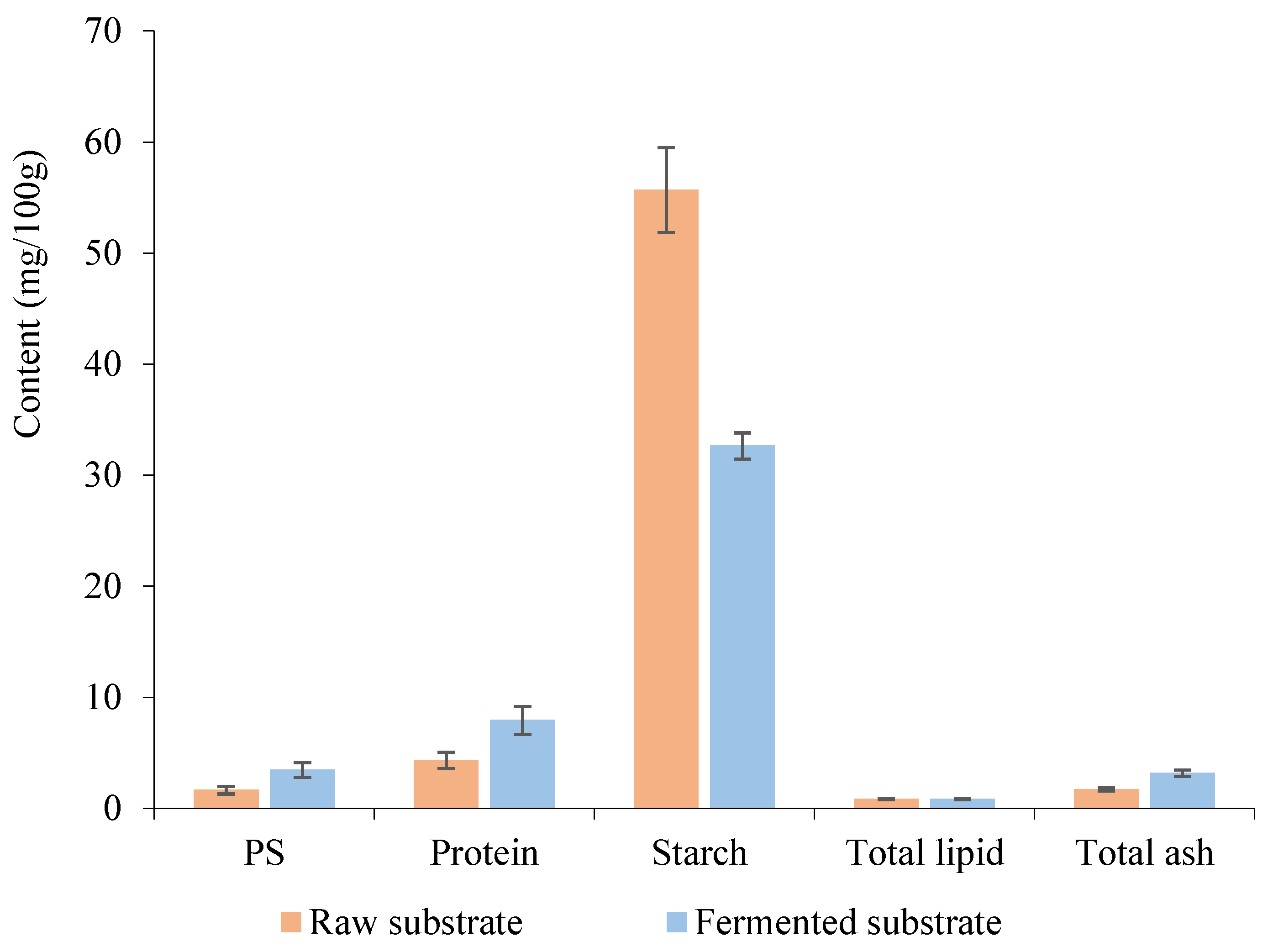
Figure 3.
Impact of polysaccharides extracted from fermented substrate on probiotic growth. (a): P. acidilactici NBD8, (b): L. pentosus NH1, (c): L. argentoraten NH15, (d): L. plantarum WCFS1. Where: GF: MRS medium without glucose, Glc: GF medium supplemented with 1 g/L glucose, PS: GF medium supplemented with 1 g/L PS, Pre: GF medium supplemented with 1g/L commercial prebiotic, GlPs: Glc medium supplemented with 1g/L PS, and GlPre: Glc medium supplemented with 1g/L commercial prebiotic.
Figure 3.
Impact of polysaccharides extracted from fermented substrate on probiotic growth. (a): P. acidilactici NBD8, (b): L. pentosus NH1, (c): L. argentoraten NH15, (d): L. plantarum WCFS1. Where: GF: MRS medium without glucose, Glc: GF medium supplemented with 1 g/L glucose, PS: GF medium supplemented with 1 g/L PS, Pre: GF medium supplemented with 1g/L commercial prebiotic, GlPs: Glc medium supplemented with 1g/L PS, and GlPre: Glc medium supplemented with 1g/L commercial prebiotic.
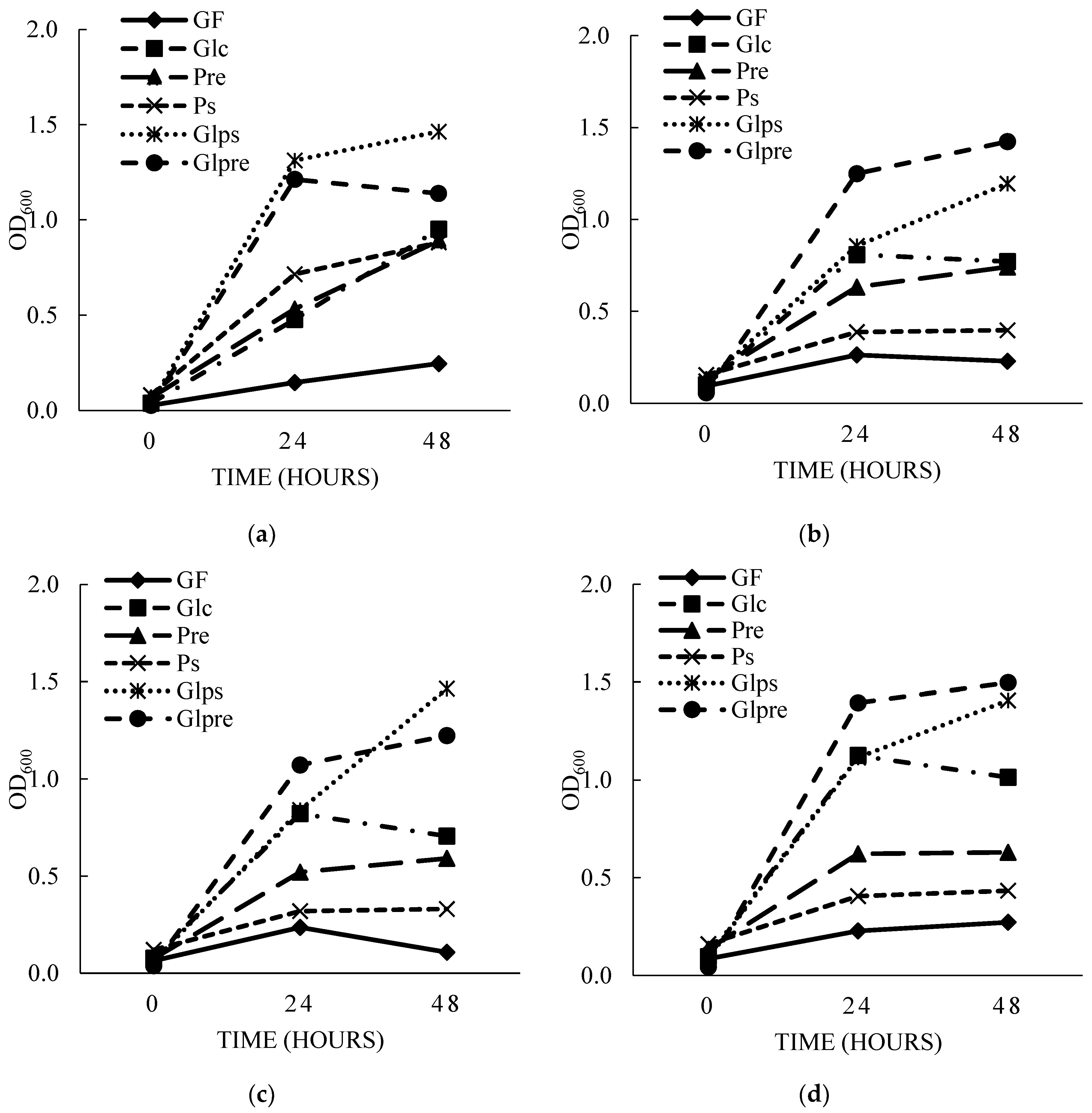
Figure 4.
The optical density values of bacteria when cultivated in medium with different carbon sources at 37°C over 24 hours. Where GF: MRS medium without glucose, Glc: GF medium supplemented with 1 g/L glucose, Pre: GF medium supplemented with 1g/L commercial prebiotic; PS: GF medium supplemented with 1 g/L PS.
Figure 4.
The optical density values of bacteria when cultivated in medium with different carbon sources at 37°C over 24 hours. Where GF: MRS medium without glucose, Glc: GF medium supplemented with 1 g/L glucose, Pre: GF medium supplemented with 1g/L commercial prebiotic; PS: GF medium supplemented with 1 g/L PS.
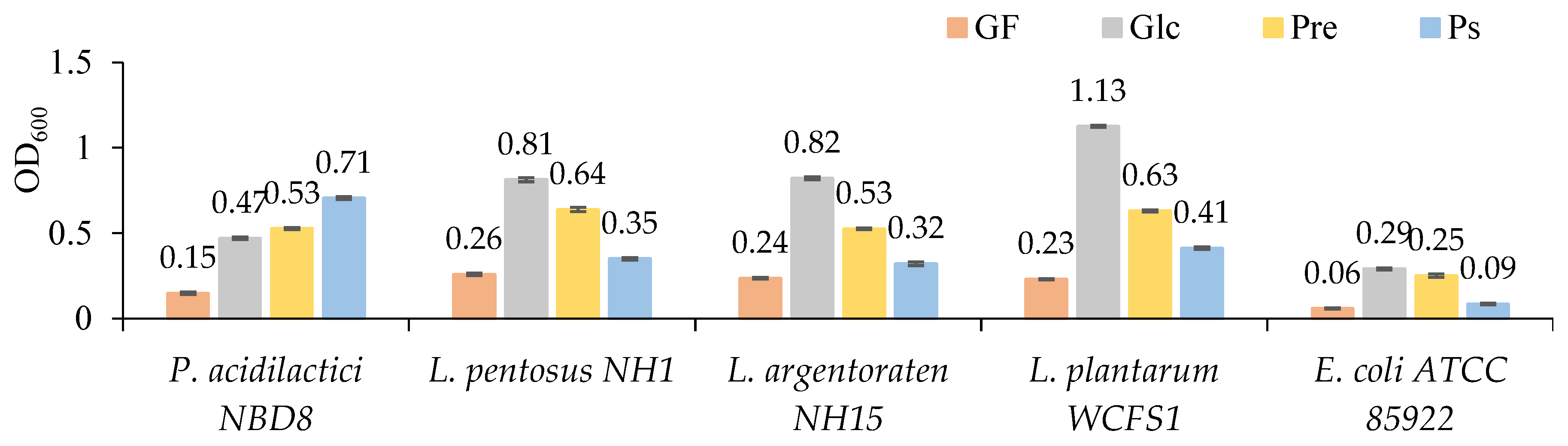
Figure 5.
Selective prebiotic stimulation characteristics of bacterial strains through the prebiotic index.
Figure 5.
Selective prebiotic stimulation characteristics of bacterial strains through the prebiotic index.
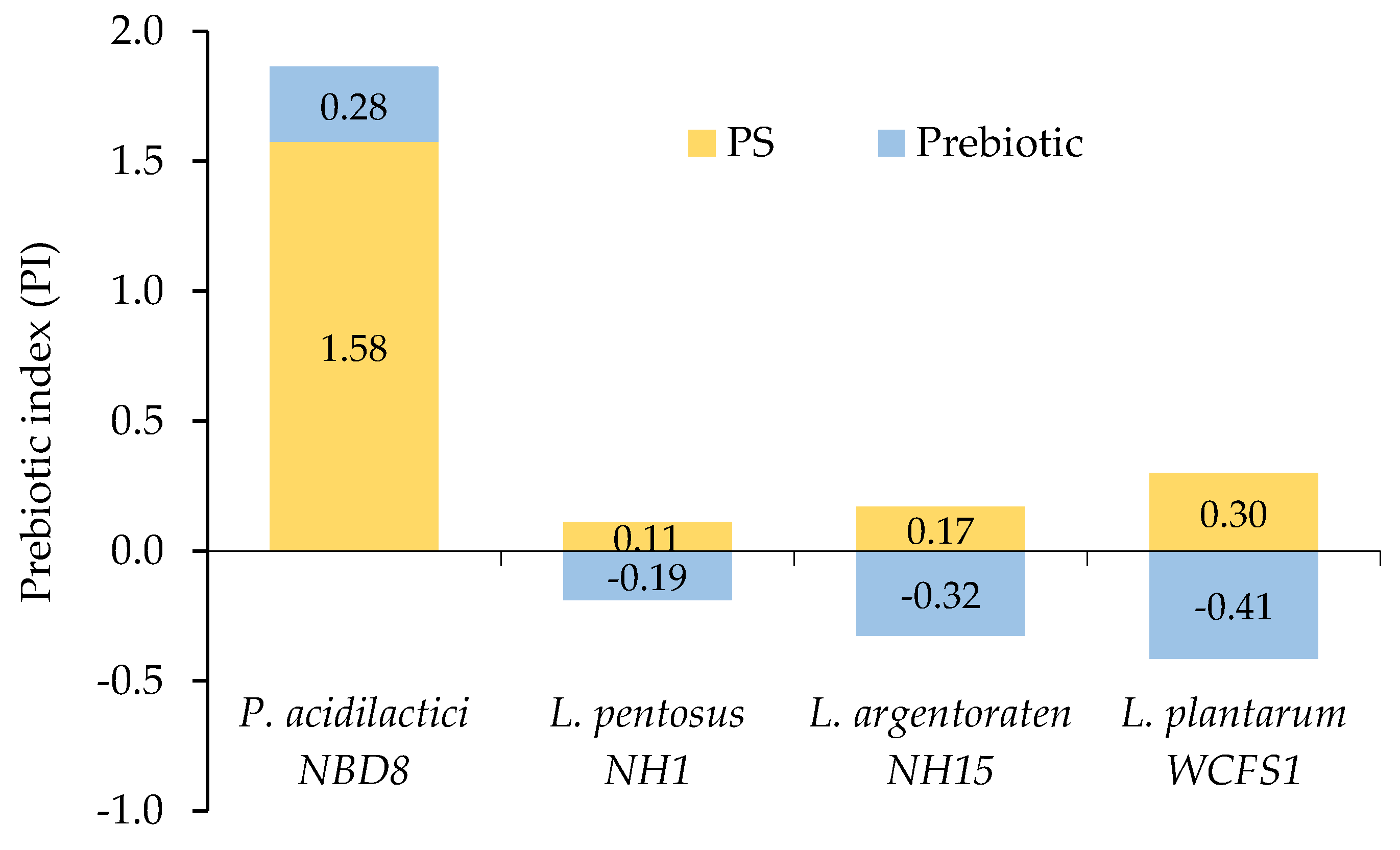

Table 1.
Blending treatments for cassava and soybean residues.
| Treatments 1 | Cassava residue (%) | Soybean residue (%) |
|---|---|---|
| CT1 (n = 15) | 100 | 0 |
| CT2 (n = 15) | 90 | 10 |
| CT3 (n = 15) | 80 | 20 |
| CT4 (n = 15) | 70 | 30 |
| CT5 (n = 15) | 60 | 40 |
1Each formula was prepared in 5 samples and repeated three times. Both cassava and soybean residues were in dry substrate.
Table 2.
Changes in the diameter (mm) of Pleurotus ostreatus mycelium on various substrate during cultivation.
Table 2.
Changes in the diameter (mm) of Pleurotus ostreatus mycelium on various substrate during cultivation.
| treatments | Culture time (days) | Characteristics of mycelium | ||||
|---|---|---|---|---|---|---|
| 1st | 3rd | 5th | 7th | 9th | ||
| CT1 (n = 15) | 1.77 ± 0.15a | 19.37 ± 1.32a | 51.8 ± 3.93a | 110.62 ± 3.45a | 120 ± 0a | Fine, low density |
| CT2 (n = 15) | 0.91 ± 0.05b | 10.26 ± 1.42c | 33.99 ± 2.04b | 83.18 ± 3.59b | 120 ± 0a | Fine, evenly white, low density |
| CT3 (n = 15) | 1.73 ± 0.17a | 13.32 ± 1.04b | 34.56 ± 2.41b | 82.51 ± 3.66b | 120 ± 0a | Thick, evenly white, high density |
| CT4 (n = 15) | 0.97 ± 0.03b | 6.47 ± 0.81d | 24.01 ± 1.68c | 60.06 ± 2.99c | 108.07 ± 3.85b | Thick, fluffy, evenly white, high density |
| CT5 (n = 15) | 0.91 ± 0.03b | 2.69 ± 0.53e | 11.56 ± 1.29d | 41.61 ± 2.35d | 84.29 ± 3.52c | Thick, fluffy, high density |
Note: Values are mean ± standard error of the mean (n = 15). Means followed by the same letter within a column are not significantly different according to the Tukey HSD Test for a significance level of 95%.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



