
Submitted:
08 March 2025
Posted:
10 March 2025
You are already at the latest version
Abstract
The European Nightjar (Caprimulgus europeaus) is a Western Palearctic bird known in Europe for its distinct breeding vocalizations during summer nights. It migrates from sub-Saharan Africa, where it overwinters, to reach its breeding grounds in Europe around early to mid-April. Although nightjar ecology has been studied in some parts of its summer range (e.g. Great Britain, Belgium), the species remains unstudied in southeastern Europe. Our aim was to use passive acoustic monitoring (PAM) to study the seasonal and temporal calling activity of the species at two sites in northern Greece (Mt. Chortiatis) across an entire breeding season, examining whether variances in the daily calling frequency could be explained by environmental variables (wind, temperature, air humidity, precipitation, moon phase, cloud cover, night length). We deployed three AudioMoth acoustic sensors, scheduled to record at 48 KHz sampling rate one out of every ten minutes, at each site from 15/4/2024 to 12/10/2024. We used the BirdNet algorithm to detect the minutes with nightjar calls (recall rate 82%). The relation of environmental variables to the daily number of minutes containing nightjar calls was assessed using generalized mixed effect models (GLMMs). They are mostly consistent with previous studies on the calling activity of the species. Calling commenced in late April and continued with decreasing frequency until early September, with variations in onset and seasonal duration across sites. Most calls occurred after dusk and before dawn (crepuscular activity), with calls taking place throughout the night during peak calling period (mid-May – mid-June). Calling frequency was higher during humid, windless, moonlit nights without precipitation and shorter duration. The results provide a baseline against which to examine possible effects of climate change on the breeding behavior of Afro-Palearctic migrant bird species, and we therefore suggest that similar studies, ideally on a continental scale, should be conducted for more species, in order to detect early climate-induced ecological changes.
Keywords:
Ecoacoustics
; acoustic sensors
; Caprimulgiformes
; vocal activity
1. Introduction
A key identification characteristic of most birds at the species level is their vocalizations, which can be used to defend territories, attract females, and share information (e.g. alert about predators, parent-offspring communication) [1]. Studying the species’ vocalization is crucial in understanding species biology and ecology [2,3,4], breeding behaviors, dialects and other intraspecific interactions [1,2,3,4,5].
While direct observations remain an important method for studying bird vocalizations [2,3,5], passive acoustic monitoring (PAM), using unattended acoustic sensors deployed for days or months at a time, is increasingly recognized as a powerful alternative. This type of monitoring minimizes the level of disturbance caused by human presence, that can potentially affect observations during field surveys. By utilizing autonomous recording units (ARUs), it allows for larger scale studies, both temporally and spatially, through lowering the costs and time required for fieldwork [6,7,8,9,10,11]. Due to such characteristics, PAM has been proven to be an effective method for studying individual species, especially for cryptic [6,7], rare [8,10] or nocturnal species, as well as long term community monitoring [9,11].
The European Nightjar (Caprimulgus europaeus) is a species of the Western Palearctic well known for its distinct vocalizations during summer nights. Its breeding distribution ranges from Northwest Europe through to East Asia [12], with an overall decreasing trend in its European breeding populations [13]. Its habitat is characterized by dry heathland and woodland sites, since dry ground offering high drainage is important for the birds to lay eggs on. Some such habitats include Pine tree (Pinaceae) forests or plantations, deciduous forests with sparse tree cover, shrubland and meadows, with the species preferring mainly the edges between them, usually used for foraging on insects [14,15]. In the Mediterranean region, the species is noted to also prefer Kermes oak (Quercus coccifera) shrublands and Mediterranean maquis vegetation [15,16]. In late winter to early spring, European Nightjars migrate from their wintering grounds in sub-Saharan Africa to their breeding summer grounds in Europe, reaching the Mediterranean shores of Europe in mid to late April [17]. Males tend to arrive first [18], and after establishing their territory they start extensive vocal territorial displays to attract females [18]. The territories reach up to 32 hectares [15,19], though this number does vary between different studies and foraging range can be significantly larger [20]. Breeding occurs usually from May to September. After mating, nightjars nest on the ground where the female lays a clutch of 2 to 4 eggs. Their mating system is mostly described as monogamous and cooperative [19], although there are two cases where polygyny was detected [21]. A pair may raise two broods per season [22].
Overall, the species is understudied, with most existing studies focusing on migration patterns [23,24,25] and hunting activity patterns [20,26]. In southern Wales, the European Nightjar starts foraging after sunset following a crepuscular pattern (i.e. peaking in the night hours after dusk and before dawn) [27]. Its foraging sites may often differ from its roosting or nesting grounds, depending on food availability and ease of hunting at each area, although the distance between them varies and seems to affect its energy consumption. Its diet consists mainly of aerial insects such as Lepidoptera and Coleoptera [28].
Regarding the song, it holds two distinct syllable types, the major (series of high frequency trills) and the minor song (series of lower frequency trills) (Figure 1). It is constant, pitch is relatively stable with subtle changes and ends with a terminal phrase [29,30]. The duration of the major and minor song is thought to be able to provide information that could be used to identify individuals based on their vocal output [29], and that some males produce different song types as the night progresses [30].
It has been reported that in the western flyway European Nightjar presence, and consequently its vocal activity, is strongly related with night duration, moon phase, minimum temperature, and cloud cover [26,31,32]. Studies have shown similar effects of the moon phase or its illumination on the vocal activity of other members of the Caprimulgiformes family as well, such as the Little Nightjar (Setopagis parvula) and the Common pauraque (Nyctidromus albicollis) [33].
Studies examining species’ spatiotemporal activity can provide valuable info, since the anticipated change in climate could potentially lead into a shift of arrival dates [34], changes on onset breed [35] and increase in the danger of diseases like avian influenza which can lead to devastating results [36]. Although studies have been done in Western and Northern Europe (Western Flyway), the Eastern Flyway and more specifically the Balkan peninsula remain extremely understudied. In Greece, where the European Nightjar is common and its call typical of early summer night soundscapes in rural areas, no study has examined the species’ calling activity to date.
To address this gap in the literature, we decided to use PAM methods to study the calling activity of nightjars at two localities in northern Greece for the entire breeding season (late April – early October 2024). PAM methods have reportedly been a beneficial monitoring tool for vocal nocturnal species, mostly due to its ability to record continuously or in sampling intervals for extended periods of time [9,26]. Based on previous studies of members of the Caprimulgiformes family, our main hypothesis is that calling activity of the European nightjar, during the mating season, would decrease with low temperature, high wind speed, and high precipitation, and increase with long nights and high moon illumination [9,26,31,32,33].
2. Materials and Methods
2.1. Study Sites and Fieldwork
We recorded nightjar calling activity at two sites in northern Greece, where the presence of the species was known due to previous pilot studies, and where elevation differs from one area to the other (Figure 2). The first site (Kissos, 40.579265N, 23.090495E, 669 m) is an area announced to become a new settlement, but which remains undeveloped except for the delineation of roads (Figure 3a). The vegetation is a natural meadow interspersed with Q. coccifera thickets. The second site (Ag. Vasileios 40.645358N, 23.083563E, 420m) has similar shrubland-meadow vegetation with higher percent cover of Q. coccifera thickets (Figure 3b). The climate is typical of medium-elevation Mediterranean regions, with hot, dry summers and mild, wet winters [37]. In the studied areas for the year 2024 it was recorded 884 mm of annual precipitation and mean annual temperature of 17.45 °C.
At each site, we deployed three autonomous recording units (ARUs; Audiomoth v1.2 with official IPX7 case). ARUs were spaced at least 900 m apart (mean distance 1,125 m), which – based on earlier on-site observations – was sufficient to ensure that same calls were not recorded in two or more sensors. According to some studies, the radius of a nightjar’s song territory reaches approximately 200 m [15]. The ARUs were scheduled to record at 48 kHz sampling rate, with medium gain, for one out of every ten minutes around the clock from April 15 to October 12, 2024. This specific scheduling was selected as a compromise between ideally continuous recording and project logistics. The SD cards (32 GB) and Lithium/Iron Disulfide AA (L91) batteries were replaced every 35 days. All ARUs were deployed on trees of the species Q. coccifera, at breast height (approx. 1.30 m), and near roads.
Since the European Nightjar is a species with territory stability [15], we assumed that the calls at each sensor represent mostly the calling activity of a single male for an entire breeding season.
2.2. Acoustic Analysis
We used the BirdNet-Analyzer v2.4 [38], accessed via Cornell University’s Raven Pro 1.6.5 acoustic analysis software (Learning Detector tool), to automatically detect European nightjar mating calls. Based on the examination of all data from five days per sensor, we selected a 0.1 threshold for our analysis as it detected at least one true-positive call segment in most 1-min audio files manually confirmed to contain a nightjar call (mean sensor “sensitivity” or “recall rate” 82%; defined as true positive / (true positive + false negative) detections [39]). All putative calls were manually reviewed to remove false positives. Since multiple segments of a single call could be tagged by the detector, we did not use the count of calls per file as a metric of nightjar calling activity. Instead, we identified 1-min files as having or not at least one true positive detection. Considering that only one minute was recorded in each 10-min interval, the maximum calling rate per hour could not exceed six one-minutes intervals.
2.2. Statistical Analyses
To assess the potential effect of environmental variables on the European Nightjar’s calling activity, we used the proportion of 1-minute acoustic files in each day (n = 144) that contained calls as a metric of calling activity. Since the proportions were not normally distributed (Shapiro-Wilk test < 0.05) and the number of days with zero detections was high, we developed Tweedie generalized linear mixed effect models in R (glmmTMB [40]) with night duration (the number of hours between sunset and sunrise), temperature (min, max, and mean (°C)), precipitation (mm), humidity (%), dew (°C), moon phase (%), moon illumination (moon phase * proportion of night with moon above the horizon [41], locality, windspeed (km/h), wind gust and cloud cover (%) as predictive variables, and site (acoustic sensor) as a nested random effect (fixed slope, random intercept). Weather data were obtained from the closest available weather station at the city of Thessaloniki (southeast foothills of Mt. Hortiates) (elevation 230 m), which was located 11 km southeast of the study area.
To decide on which variables to include in the final model, while also managing model complexity, we first examined support for inclusion of each variable in a baseline model having as fixed parameters only the intercept and night duration, which is well documented to be a good predictor of nightjar calling activity [31]. Model selection was based on the Akaike Information Criterion (AIC) [42]. Variables included in models with AIC lower than the baseline model were further considered. To exclude possible multicollinearities among variables, we kept from each pair of correlated variables (r > 0.65) the one with the lower AIC value from the previous model comparison. We used the final set of fixed variables to run all possible multivariate combinations (R package MuMIn:dredge; [43]), using AIC again for model selection. Since multiple models have ΔAIC<2, a threshold often used to identify competing best models, we calculated the model average coefficients of variables from these top models using MuMIn:mod.avg. The goodness of fit of the global model was assessed using the pseudo-R² [43]. All analyses were conducted in R software (v4.4.1) [40,44].
3. Results
All six sensors operated for 180 days (April 15 – Oct 12). In total, 39,373 sound clips were reviewed, resulting in nightjars being detected in 2,914 of the 1-min acoustic files recorded (1.9%; n = 155,520) across all sensors (range 137 – 857 files) (Table 1). The days with nightjar calling activity (i.e. ≥ 1 file with calls) ranged from 58 to 112 per sensor (mean 88 ± 22.8 SD) (Table 1). Overall calling activity across the two sites was similar (Kissos: 1,360 files; Agios Vasileios: 1,554 files), with each having a sensor with low (100 - 200), medium (500-550) and high (> 700) count of files with nightjar calls.
The recall rate of the BirdNet algorithm (0.1 threshold) of 1-min files containing nightjar calls ranged from 71 – 91.3% per sensor (mean 82%), based on the manual review of 4,320 files (five days from each sensor) containing 326 files with calls (Table 2).
Nightjar calls were first detected, sporadically, in Agios Vasileios (sensors AM16, AM17) in late April, with calling activity subsequently peaking in most sensors between mid-May and mid-June (Figure 4). Calls continued throughout July, with the calling frequency dropping rapidly after mid-August, and stopping altogether by early September.
Most calling occurs after dusk and before dawn, reaching their pick near midnight (nocturnal activity), with calls taking place throughout the night during peak calling period. Practically no calls were recorded during daytime (Figure 4). There was very little variation in the diel calling activity pattern of all sensors at seasonal level (Figure A2).
The percentage of night-time 1-min files containing nightjar calls during different quarters of the moon phase (1st, 2nd, 3rd and Full Moon) varied significantly across sensors (Figure 4). Moon phase (as percent illuminated; 0-1 range) was included in the final model of nightjar calling activity, having a significant positive effect (Table 3). In general, the model showed that nightjar calling activity was higher during humid, moonlit, shorter nights without precipitation. Temperature (minimum), when included in the multivariate model, did not have a significant effect on nightjar calling activity.
4. Discussion
Our findings describe the calling activity of the European Nightjar (Caprimulgus europaeus) throughout an entire breeding period (April – October) at two sites in northern Greece. The original hypothesis, namely that calling activity would be lower during days with high wind and precipitation and higher during days with increased proportion of the moon illuminated, is supported by the results.
Night duration was found to be the single most informative parameter affecting (negatively) nightjar calling activity, which is not surprising since night duration is shorter during late spring and summer, when breeding takes place. With the available data, we are unable to infer whether shorter nights trigger breeding behavior in the species or not. The reported effect of night duration and moon phase on the calling activity of Caprimulgiformes species has been reported before [26,31,32,33,45]. The effect of moon phase however is not universal, as it has no effect in the calling activity of the red-necked nightjar (Caprimulgus ruficollis) [46].
While a previous study by Reino et al. [31] reported lower European nightjar calling activity during cooler, cloudy nights, our results did not find such a relationship. The cloud cover was not informative enough to be included in the final set of explanatory variables (averaged model) and minimum temperature had a weak and non-significant effect.
Our finding that precipitation negatively affected nightjar calling activity is not surprising, as this relationship has been reported for birds before [47]. However, our study is the first – to our knowledge – to document this effect for the European nightjar. Unlike precipitation though, air humidity was significantly positively related to nightjar calling activity. Given that sound travels further in humid air [48], the relationship observed may reflect increased detection range by the acoustic sensors of nightjar calls and/or an actual behavioral change by the species to capitalize on this physical effect. Other species are known to benefit from this phenomenon, like those of the Apterygidae family [49].
Regarding the timing of the European Nightjar’s spring migrationour findings are in agreement with previous studies which reported mid to late April arrival of the species in the Mediterranean regions of southern Europe [17,18,19].
One potential limitation of the study is its non-continuous recording schedule, dictated by technical limitations of the acoustic sensor used – namely the maximum recording duration possible with the provided battery pack. Ideally, a continuous recording would provide higher temporal resolution on nightjar calling activity variations. The fact that the species calls often and for extended periods of time during the night makes us believe that the resolution lost is not such that would affect our conclusions. However, future studies could – at least at some recording stations – compare the impact of our recording schedule against that of a continuous one. In general, given the increased analytical effort required for continuous recordings, the recommendation of PAM protocols for birds is often to record around the clock with a given interval (similar to what we have done) [50]. Although there are studies supporting PAM as a good practice for monitoring Nightjars, there’s criticism by others. According to the findings of Zwart et al. [51], ARUs were found to be better at detecting nightjar calls than traditional human surveys. But in study of Eisenring et al. [52] hey were found to perform insufficiently. Another limitation of our study is the distance between the study sites and the meteorological station. As mentioned in the “materials and methods” chapter, data were obtained from the closest available weather station with continuous recordings. Although these measurements might not capture microclimatic differences between the exact locations, we believe that they adequately represent the regional weather patterns in which this study focuses, as is done in other similar studies [9].
5. Conclusions
The findings of the study are mostly consistent with previous studies on the calling activity of the European Nightjar during summer breeding season. While small in spatial scale, the study is the first of its kind for the species in the Balkan peninsula, included data from an entire breeding season, and showcases the value of passive acoustic monitoring (PAM) as a tool for effectively studying the behavior of cryptic/nocturnal species such as the European Nightjar. The results also serve as a baseline against which to examine possible effects of climate change on the breeding behavior of Afro-Palearctic migrant bird species (e.g. onset, duration, response to changes in environmental variables) in the future. We propose that similar studies, across a larger – ideally continental - scale, are conducted across a range of species to provide early warning of climate-induced ecological changes. To achieve this, we propose that national level PAM grids are developed across Europe, following the example of the Australian Acoustic Observatory [53], with recording and analysis protocols being coordinated at EU level. Given the high spatiotemporal resolution provided by PAM, the decreasing costs of deploying and operating PAM grids, and the increasingly automated analysis of the datasets (e.g. using algorithms such as BirdNet), we believe that developing an EU-wide acoustic observatory is primarily an issue of coordination, collaboration and political priority settings, and less of funding limitations.
Author Contributions
Conceptualization, D.T., C.A. and E.N.; methodology, D.T. and C.A.; software, D.T. and A.R.; validation, D.T., A.R., E.N., A.K., T.P. and C.A.; formal analysis, D.T. and A.R.; investigation, D.T., C.A. and T.P.; resources, C.A.; data curation, D.T. and A.R.; writing—original draft preparation, D.T. and C.A.; writing—review and editing, D.T., A.R., E.N., A.K., T.P. and C.A.; visualization, D.T.; supervision, C.A. and TP; project administration, D.T. and C.A.; funding acquisition, C.A.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data may be available from the authors upon request.
Acknowledgments
We would like to thank Panagiotis Papadopoulos for his help and assistance in the field. This study is dedicated in the memory of Stefanos Pantelatos who played a key role in the implementation of a pilot study in 2023 that eventually led to the inception of the current study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Figure A1.
Representative landscape photographs of the two studied areas: (a) Kissos; (b) Agios Vasileios.
Figure A1.
Representative landscape photographs of the two studied areas: (a) Kissos; (b) Agios Vasileios.

Figure A2.
Hourly distribution of 1-min files containing European Nightjar calls per sensor, for the duration of the study period.
Figure A2.
Hourly distribution of 1-min files containing European Nightjar calls per sensor, for the duration of the study period.
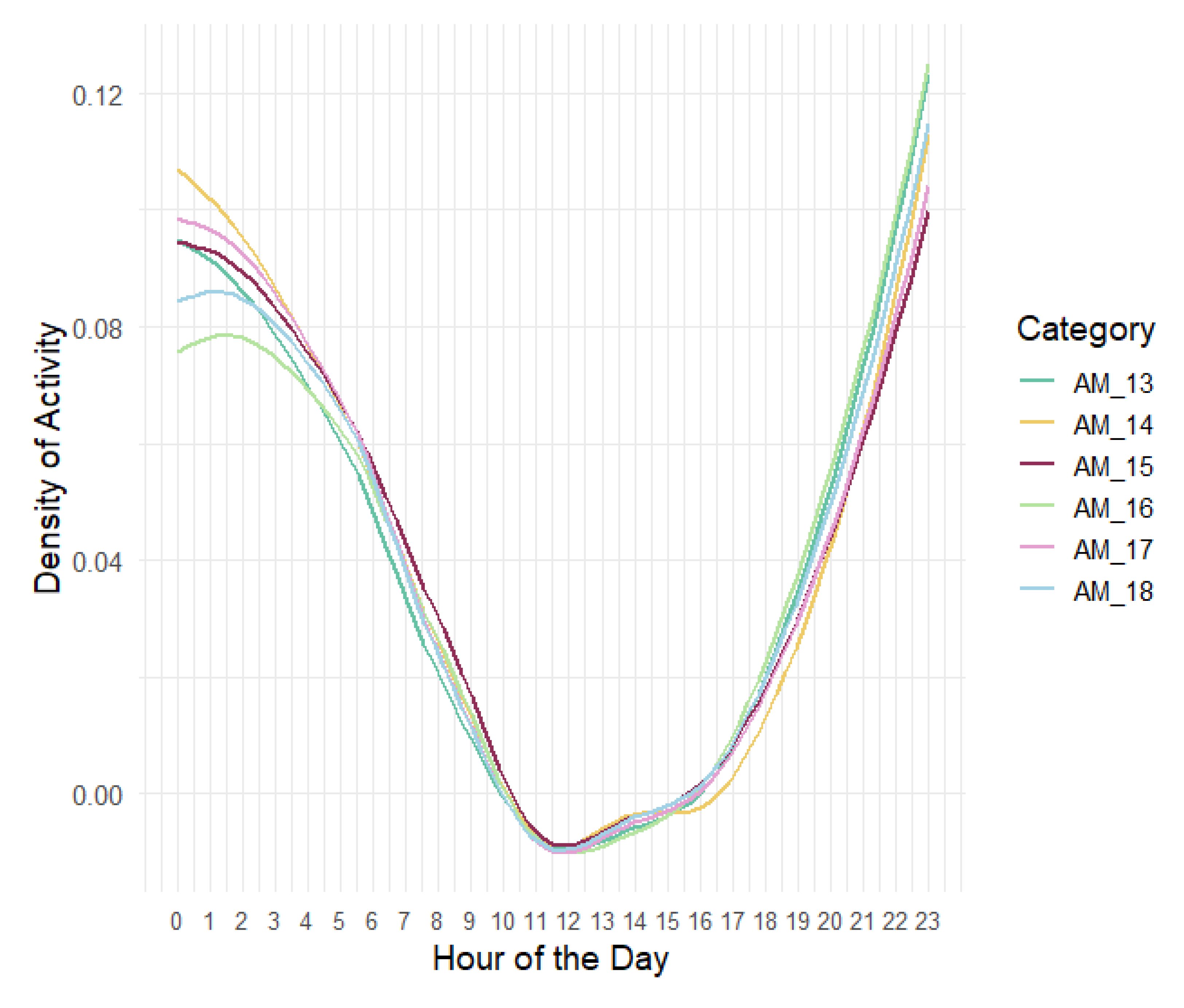

Table A1.
Akaike Information Criterion based model selection of univariable models and baseline model (daily proportion of activity ~ Night duration + (1|Sensor)) used to identify informative variables which were subsequently considered in multivariable models.
Table A1.
Akaike Information Criterion based model selection of univariable models and baseline model (daily proportion of activity ~ Night duration + (1|Sensor)) used to identify informative variables which were subsequently considered in multivariable models.
| Model | AIC score | ΔAIC score |
|---|---|---|
| Humidity | - 1635.716 | 0 |
| Temperature Min | - 1622.213 | 13.503 |
| Temperature Mean | - 1620.230 | 15.486 |
| Wind gust | - 1616.328 | 19.388 |
| Moon Phase | - 1614.372 | 21.345 |
| Temperature Max | - 1614.227 | 21.489 |
| Precipitation | - 1609.066 | 26.650 |
| Illuminocity index | - 1608.762 | 26.954 |
| Cloud cover | - 1608.300 | 27.416 |
| Baseline model | - 1607.721 | 27.995 |
| Windspeed | - 1606.082 | 29.634 |
| Locality | - 1605.890 | 29.826 |
| Dew | - 1605.741 | 29.975 |
Table A2.
Monthly detections per locality and sensor.
| Sensor | April | May | June | July | August | September | October |
|---|---|---|---|---|---|---|---|
| Kissos: | |||||||
| AM13 | 0 | 56 | 232 | 181 | 49 | 5 | 0 |
| AM14 | 0 | 215 | 314 | 105 | 64 | 2 | 0 |
| AM15 | 0 | 23 | 88 | 22 | 4 | 0 | 0 |
| Ag. Vasileios: | |||||||
| AM16 | 2 | 179 | 110 | 149 | 59 | 6 | 0 |
| AM17 | 1 | 552 | 152 | 90 | 62 | 0 | 0 |
| AM18 | 0 | 56 | 89 | 37 | 9 | 1 | 0 |
References
- Catchpole, C. K.; Slater, P. J. B. Bird Song: Biological Themes and Variations, 2nd ed; Cambridge university press: NY, USA, 2008; 93-158.
- Oñate-Casado, J.; Porteš, M.; Beran, V.; Petrusek, A.; Petrusková, T. Guess Who? Evaluating Individual Acoustic Monitoring for Males and Females of the Tawny Pipit, a Migratory Passerine Bird with a Simple Song. J Ornithol 2023, 164, 845-858. [CrossRef]
- Petrusková, T.; Pišvejcová, I.; Kinštová, A.; Brinke, T.; Petrusek, A. Repertoire-Based Individual Acoustic Monitoring of a Migratory Passerine Bird with Complex Song as an Efficient Tool for Tracking Territorial Dynamics and Annual Return Rates. Methods Ecol Evol 2016, 7, 274-284. [CrossRef]
- Gallego-Abenza, M.; Kraft, F.-L.H.; Ma, L.; Rajan, S.; Wheatcroft, D. Responses in Adult Pied Flycatcher Males Depend on Playback Song Similarity to Local Population. Behavioral Ecology 2024, 36, arae090. [CrossRef]
- Pipek, P.; Petrusková, T.; Petrusek, A.; Diblíková, L.; Eaton, M.A.; Pyšek, P. Dialects of an Invasive Songbird Are Preserved in Its Invaded but Not Native Source Range. Ecography 2018, 41, 245-254. [CrossRef]
- Matsubayashi, S.; Nakadai, K.; Suzuki, R.; Ura, T.; Hasebe, M.; Okuno, H.G. Auditory Survey of Endangered Eurasian Bittern Using Microphone Arrays and Robot Audition. Front Robot AI 2022, 9, 854572. [CrossRef]
- Znidersic, E.; Watson, D.M.; Towsey, M.W. A New Method to Estimate Abundance of Australasian Bittern (Botaurus poiciloptilus) from Acoustic Recordings. Avian Conserv Ecol 2024, 19, 16. [CrossRef]
- De Araújo, C.B.; Zurano, J.P.; Torres, I.M.D.; Simões, C.R.M.A.; Rosa, G.L.M.; Aguiar, A.G.; Nogueira, W.; Vilela, H.A.L.S.; Magnago, G.; Phalan, B.T.; et al. The Sound of Hope: Searching for Critically Endangered Species Using Acoustic Template Matching. Bioacoustics 2023, 32, 708-723. [CrossRef]
- Astaras, C.; Valeta, C.; Vasileiadis, I. Acoustic Ecology of Tawny Owl (Strix aluco) in the Greek Rhodope Mountains Using Passive Acoustic Monitoring Methods. Folia Oecologica 2022, 49, 110-116. [CrossRef]
- Wang, M.; Mei, J.; Liu, F. Temporal Acoustic Patterns of the Oriental Turtle Dove in a Subtropical Forest in China. Diversity (Basel) 2022, 14, 1043. [CrossRef]
- Pérez-Granados, C.; Schuchmann, K.L. Passive Acoustic Monitoring of Chaco Chachalaca (Ortalis canicollis) Over a Year: Vocal Activity Pattern and Monitoring Recommendations. Trop Conserv Sci 2021, 14, 19400829211058295. [CrossRef]
- Day, G.; Fox, G.; Hipperson, H; Maher, K. H.; Tucker, R.; Horsburgh, G. J.; Waters, D.; Durant, K. L.; Burke, T.; Slate, J.; Arnold, K. E. Revealing the Demographic History of the European Nightjar (Caprimulgus europaeus). Ecol Evol 2024, 14, e70460. [CrossRef]
- Data Zone by BirdLife. Available online: https://datazone.birdlife.org/species/factsheet/european-nightjar-caprimulgus-europaeus (accessed on 01/03/2025).
- Mitchell, L. J.; Kohler, T.; White, P. C. L.; Arnold K. E. High interindividual variability in habitat selection and functional habitat relationships in European nightjars over a period of habitat change. Ecol Evol 2020, 10, 5932-5945. [CrossRef]
- Sharps, K.; Henderson, I.; Conway, G.; Armour-Chelu, N.; Dolman, P.M. Home-Range Size and Habitat Use of European Nightjars Caprimulgus europaeus Nesting in a Complex Plantation-Forest Landscape. Ibis 2015, 157, 260-272. [CrossRef]
- Warman, S.R. Handbook of the Birds of Europe, the Middle East and North Africa, Volume V: Tyrant Flycatchers to Thrushes (Birds of the Western Palearctic). Trends Ecol Evol 1988, 3, 282-283. [CrossRef]
- Norevik, G.; Åkesson, S.; Hedenström, A. Migration Strategies and Annual Space-Use in an Afro-Palearctic Aerial Insectivore – the European Nightjar Caprimulgus europaeus. J Avian Biol 2017, 48, 738-747. [CrossRef]
- Cleere, N.; Christie, D.A.; Rasmussen, P.C. Eurasian Nightjar (Caprimulgus europaeus), Version 1.1., Rasmussen, P.C., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2021. [CrossRef]
- Cleere, N. Nightjars: A Guide to the Nightjars, Nighthawks, and Their Relatives, 1st ed.; Yale University Press: New Haven, Connecticut, USA, 1998.
- Evens, R.; Beenaerts, N.; Witters, N.; Artois, T. Study on the Foraging Behaviour of the European Nightjar Caprimulgus europaeus Reveals the Need for a Change in Conservation Strategy in Belgium. J Avian Biol 2017, 48, 1238-1245. [CrossRef]
- Evens, R.; Lathouwers, M.; Creemers, J.; Ulenaers, E.; Eens, M.; Kempenaers, B. A case of facultative polygyny in an enigmatic monogamous species, the European Nightjar (Caprimulgus europaeus). Ecol evol 2024, 14, e70366. [CrossRef]
- Langston, R.H.W.; Liley, D.; Murison, G.; Woodfield, E.; Clarke, R.T. What Effects Do Walkers and Dogs Have on the Distribution and Productivity of Breeding European Nightjar Caprimulgus europaeus? Ibis 2007, 149, 27-36. [CrossRef]
- Jacobsen, L.B.; Jensen, N.O.; Willemoes, M.; Hansen, L.; Desholm, M.; Fox, A.D.; Tøttrup, A.P.; Thorup, K. Annual Spatiotemporal Migration Schedules in Three Larger Insectivorous Birds: European Nightjar, Common Swift and Common Cuckoo. Animal Biotelemetry 2017, 5, 4. [CrossRef]
- Norevik, G.; Åkesson, S.; Andersson, A.; Bäckman, J.; Hedenström, A. Flight Altitude Dynamics of Migrating European Nightjars across Regions and Seasons. Jour Exper Biology 2021, 224, jeb242836. [CrossRef]
- Cresswell, B.; Edwards, D. Geolocators Reveal Wintering Areas of European Nightjar (Caprimulgus europaeus). Bird Study 2013, 60, 77-86. [CrossRef]
- Norevik, G.; Åkesson, S.; Andersson, A.; Bäckman, J.; Hedenström, A. The Lunar Cycle Drives Migration of a Nocturnal Bird. PLoS Biol 2019, 17, e3000456. [CrossRef]
- Jenks, P.; Green, M.; Cross, T. Foraging Activity and Habitat Use by European Nightjars in South Wales. British Birds 2014, 107.
- Mitchell, L. The influence of environmental variation on individual foraging and habitat selection behaviour of the European nightjar. Doctoral dissertation, University of York, Heslington, York, UK, 2019.
- Rebbeck, M.; Corrick, R.; Eaglestone, B.; Stainton, C. Recognition of Individual European Nightjars Caprimulgus europaeus from Their Song. Ibis 2001, 143, 468-475. [CrossRef]
- Docker, S.; Lowe, A.; Abrahams, C. Identification of Different Song Types in the European Nightjar Caprimulgus europaeus. Bird Study 2020, 67, 119-127. [CrossRef]
- Reino, L.; Porto, M.; Santana, J.; Osiejuk, T.S. Influence of Moonlight on Nightjars’ Vocal Activity: A Guideline for Nightjar Surveys in Europe. Biologia (Poland) 2015, 70, 968-973. [CrossRef]
- Evens, R.; Kowalczyk, C.; Norevik, G.; Ulenaers, E.; Davaasuren, B.; Bayargur, S.; Artois, T.; Åkesson, S.; Hedenström, A.; Liechti, F.; et al. Lunar Synchronization of Daily Activity Patterns in a Crepuscular Avian Insectivore. Ecol Evol 2020, 10, 7106-7116. [CrossRef]
- Pérez-Granados, C.; Schuchmann, K.L.; Marques, M.I. Addicted to the Moon: Vocal Output and Diel Pattern of Vocal Activity in Two Neotropical Nightjars Is Related to Moon Phase. Ethol Ecol Evol 2022, 34, 66-81. [CrossRef]
- Gordo, O. Why Are Bird Migration Dates Shifting? A Review of Weather and Climate Effects on Avian Migratory Phenology. Clim Res 2007, 35, 37-58. [CrossRef]
- Cuchot, P.; Bonnet, T.; Dehorter, O.; Henry, P.Y.; Teplitsky, C. How Interacting Anthropogenic Pressures Alter the Plasticity of Breeding Time in Two Common Songbirds. Jour Animal Ecol 2024, 93, 918–931. [CrossRef]
- Prosser, D.J.; Teitelbaum, C.S.; Yin, S.; Hill, N.J.; Xiao, X. Climate Change Impacts on Bird Migration and Highly Pathogenic Avian Influenza. Nat Microbiol 2023, 8, 2223-2225. [CrossRef]
- Lionello, P.; Malanotte-Rizzoli, P.; Boscolo, R.; Alpert, P.; Artale, V.; Li, L.; Luterbacher, J.; May, W.; Trigo, R.; Tsimplis, M.; et al. The Mediterranean Climate: An Overview of the Main Characteristics and Issues. Develop Earth Envi Sci 2006, 4, 1-26. [CrossRef]
- Kahl, S.; Wood, C.M.; Eibl, M.; Klinck, H. BirdNET: A Deep Learning Solution for Avian Diversity Monitoring. Ecol Info 2021, 61, 101236. [CrossRef]
- Knight, E.C.; Hannah, K.C.; Foley, G.J.; Scott, C.D.; Brigham, R.M.; Bayne, E. Recommendations for acoustic recognizer performance assessment with application to five common automated signal recognition programs. Avian Conserv Ecol 2017, 12, 14. [CrossRef]
- Kuznetsova, A., Brockhoff, P.B., & Christensen, R.H.B. (2024). GlmmTMB: Generalized Linear Mixed Models Using Template Model Builder. R Package Version 1.1.2. https://CRAN.R-Project.Org/Package=glmmTMB.
- Astaras, C.; Linder, J.M.; Wrege, P.; Orume, R.; Johnson, P.J.; MacDonald, D.W. Boots on the Ground: The Role of Passive Acoustic Monitoring in Evaluating Anti-Poaching Patrols. Environ Conserv 2020, 47, 213-216. [CrossRef]
- Burnham, K.P.; Anderson, D.R. Statistical Theory and Numerical Results. In Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed; Springer: NY, USA, 2002; pp. 352–436. ISBN 978-0-387-22456-5.
- Bartoń, Kamil. (2024). MuMIn: Multi-Model Inference. R Package Version 1.48.4. https://CRAN.R-Project.Org/Package=MuMIn1.
- Wickham, M.H. Package “ggplot2” Type Package Title An Implementation of the Grammar of Graphics; 2014;
- Ashdown, R.A.M.; McKechnie, A.E. Environmental Correlates of Freckled Nightjar (Caprimulgus Tristigma) Activity in a Seasonal, Subtropical Habitat. J Ornithol 2008, 149, 615-619. [CrossRef]
- De Felipe, M.; Sáez-Gómez, P.; Camacho, C. Environmental Factors Influencing Road Use in a Nocturnal Insectivorous Bird. Eur J Wildl Res 2019, 65, 1-7. [CrossRef]
- Alonso, J.C.; Abril-Colón, I.; Ucero, A.; Palacín, C. Precipitation and Female Experience Are Major Determinants of the Breeding Performance of Canarian Houbara Bustards. Wildlife Biol 2024, e01345. [CrossRef]
- Motegi, T.; Mizutani, K.; Wakatsuki, N. Simultaneous Measurement of Air Temperature and Humidity Based on Sound Velocity and Attenuation Using Ultrasonic Probe. Jpn J Appl Phys 2013, 52, 07HC05. [CrossRef]
- Digby, A.; Towsey, M.; Bell, B.D.; Teal, P.D. Temporal and Environmental Influences on the Vocal Behaviour of a Nocturnal Bird. J Avian Biol 2014, 45, 591-599. [CrossRef]
- Metcalf, O.; Abrahams, C.; Ashington, B.; Baker, E.; Bradfer-Lawrence, T.; Browning, E.; Carruthers-Jones, J.; Darby, J.; Dick, J.; Eldridge, A.; Elliott, D.; Heath, B.; Howden-Leach, P.; Johnston, A.; Lees, A.; Meyer, C.; Ruiz Arana, U.; Smyth, S. Good Practice Guidelines for Long-Term Ecoacoustic Monitoring in the UK. The UK Acoustics Network 2023.
- Zwart, M.C.; Baker, A.; McGowan, P.J.K.; Whittingham, M.J. The Use of Automated Bioacoustic Recorders to Replace Human Wildlife Surveys: An Example Using Nightjars. PLoS ONE 2014, 9, e102770. [CrossRef]
- Eisenring, E.; Eens, M.; Pradervand, J. N.; Jacot, A.; Baert, J.; Ulenaers, E.; Evens, R. Quantifying Song Behavior in a Free-Living, Light-Weight, Mobile Bird Using Accelerometers. Ecol. Evol. 2022, 12(1), e8446. [CrossRef]
- Roe, P.; Eichinski, P.; Fuller, R.A.; McDonald, P.G.; Schwarzkopf, L.; Towsey, M.; Truskinger, A.; Tucker, D.; Watson, D.M. The Australian Acoustic Observatory. Methods Ecol Evol 2021, 12, 1802-1808. [CrossRef]
Figure 1.
Part of a typical European Nightjar call containing both major and minor songs.
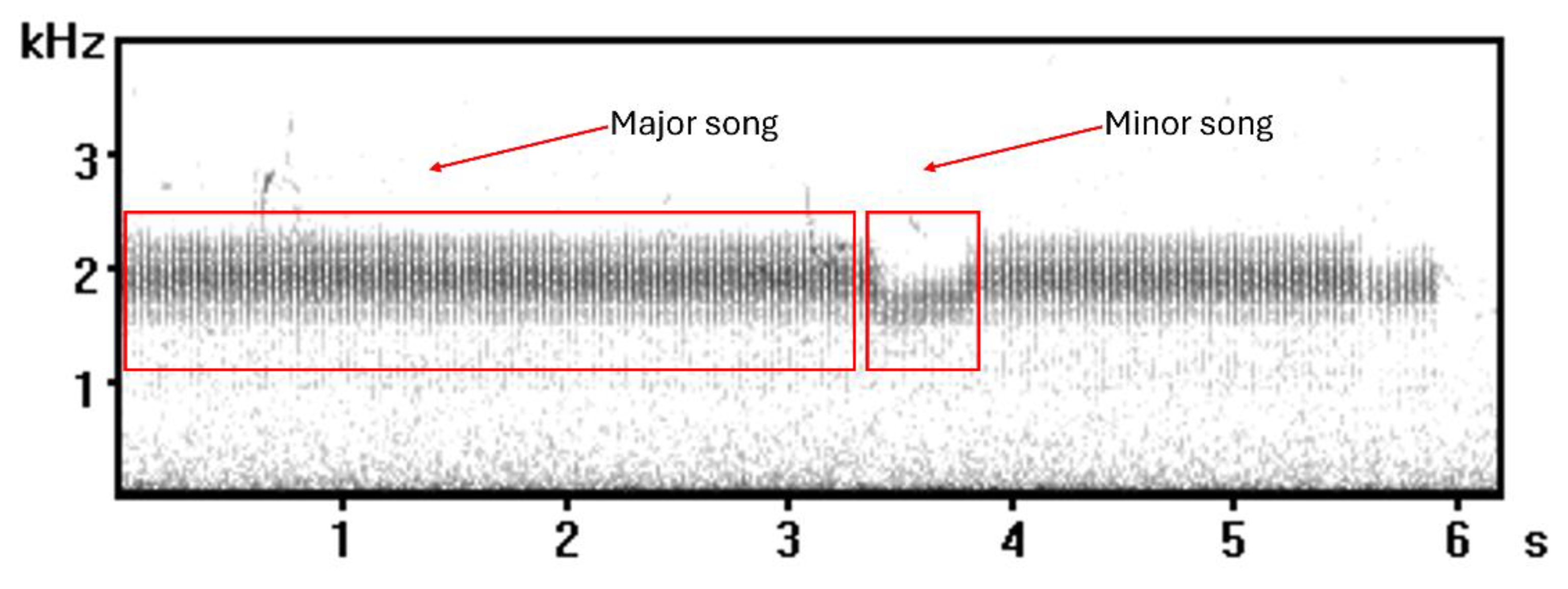
Figure 2.
Map of the study area.
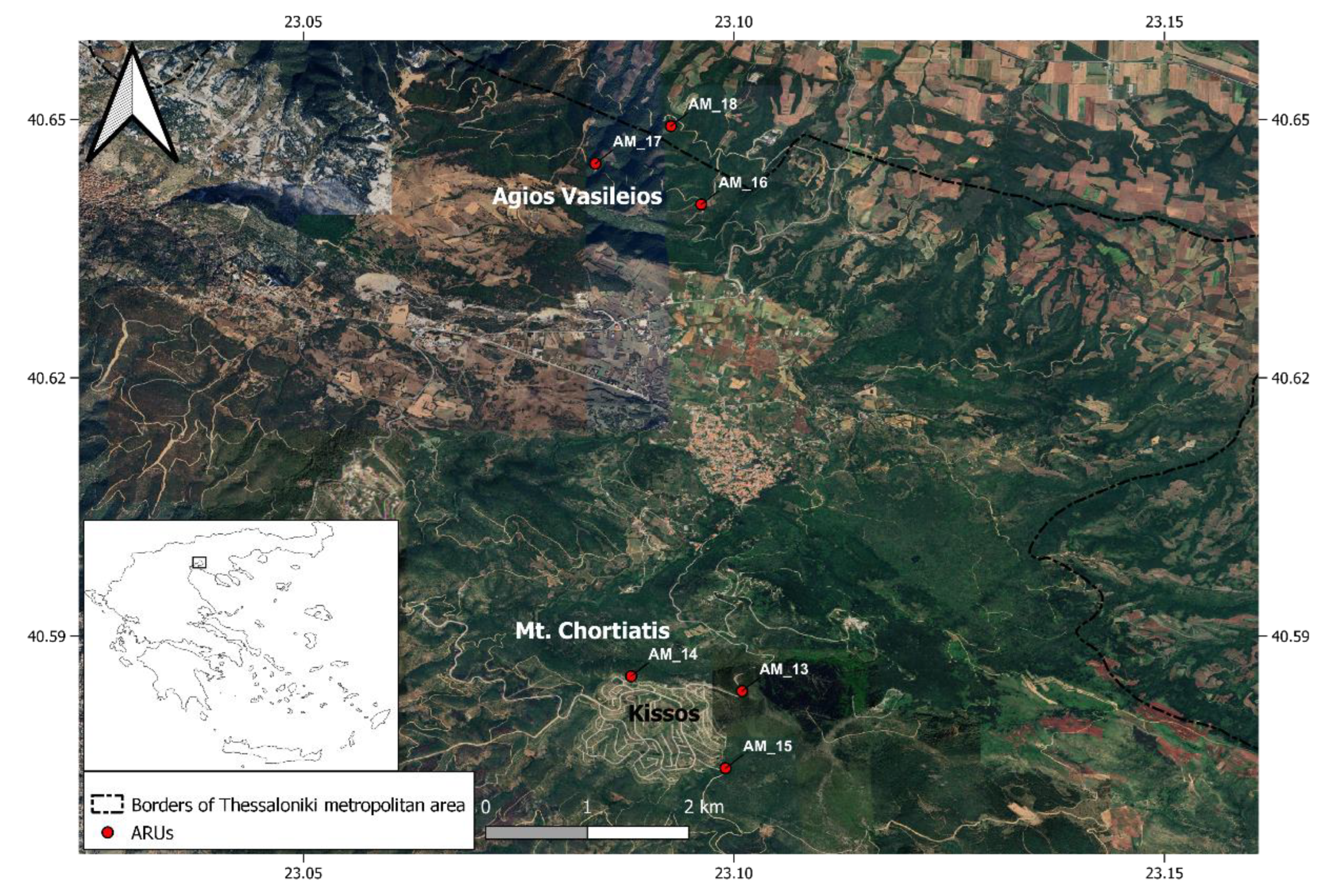
Figure 3.
Distribution and count of 1-min audio files containing European Nightjar (Caprimulgus Europaeus) mating calls per hour, day, month, and acoustic sensor. Since the sensors only recorded 1 min out of each 10-min period, the maximum count of files with calls per hour is six. October is omitted from the graph since no calls were recorded then.
Figure 3.
Distribution and count of 1-min audio files containing European Nightjar (Caprimulgus Europaeus) mating calls per hour, day, month, and acoustic sensor. Since the sensors only recorded 1 min out of each 10-min period, the maximum count of files with calls per hour is six. October is omitted from the graph since no calls were recorded then.
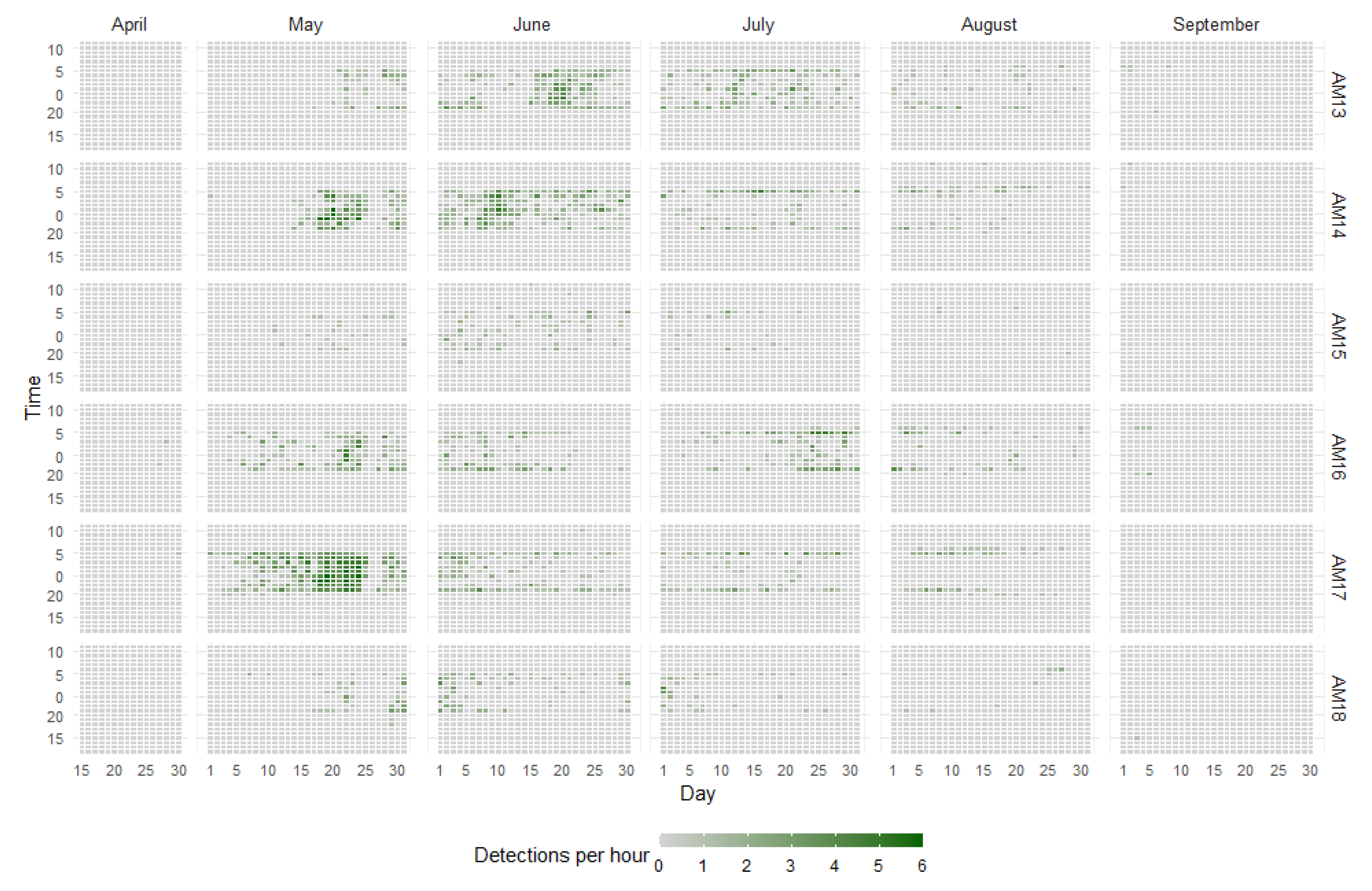
Figure 4.
Box plots of proportion of night 1-min files containing at least one nightjar mating call, (May – August). Moon phase: (1) First quarter of the moon, (2) Second quarter of the moon, (3) Third quarter of the moon, (4) Full moon.
Figure 4.
Box plots of proportion of night 1-min files containing at least one nightjar mating call, (May – August). Moon phase: (1) First quarter of the moon, (2) Second quarter of the moon, (3) Third quarter of the moon, (4) Full moon.
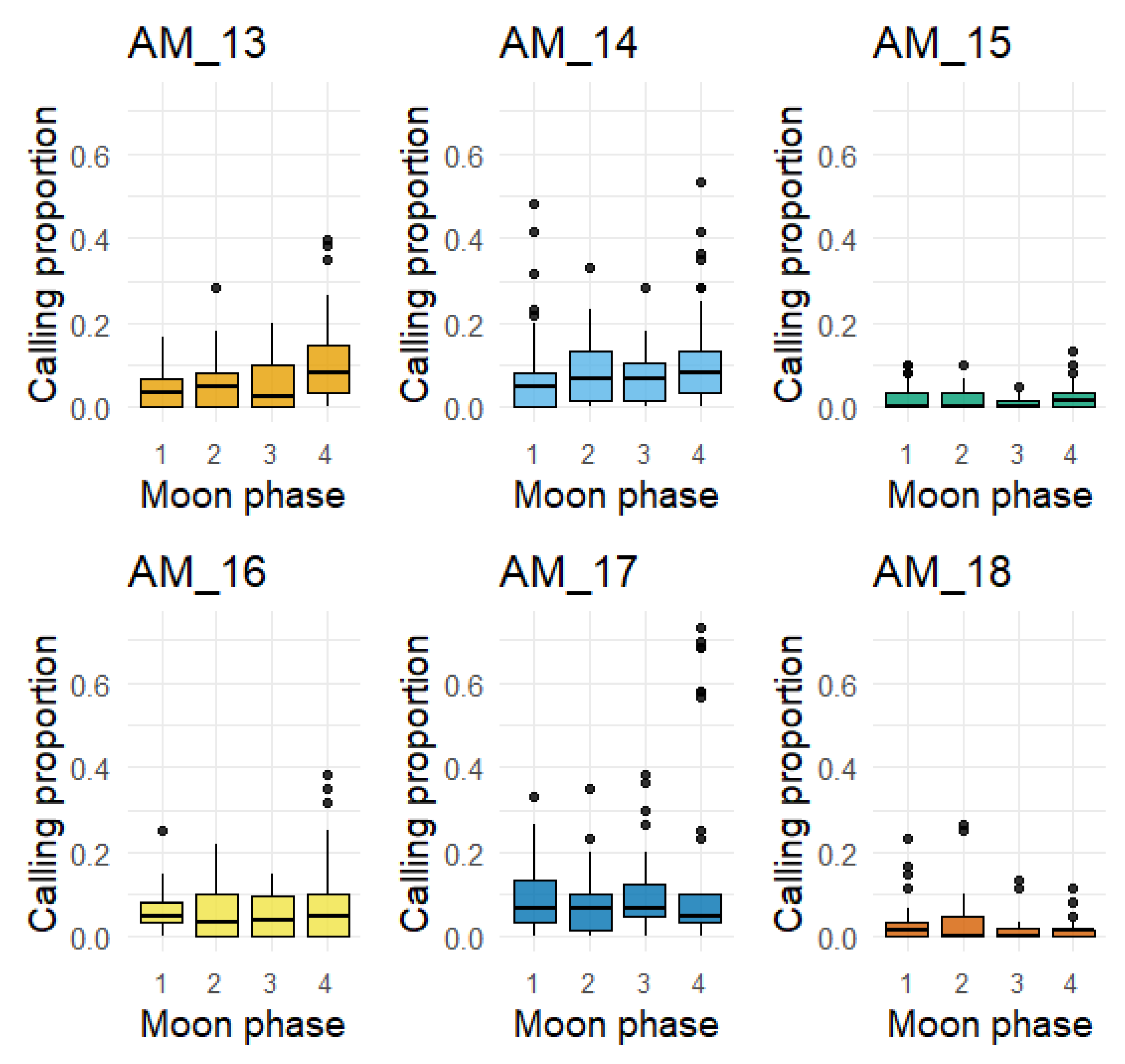
Table 1.
Nightjar detection frequency per sensor during the study period (15/4 – 12/10/2024). All sensors recorded all days (n = 180). Number of 1-min files per day is 144 (one every ten minutes).
Table 1.
Nightjar detection frequency per sensor during the study period (15/4 – 12/10/2024). All sensors recorded all days (n = 180). Number of 1-min files per day is 144 (one every ten minutes).
| Site | Sensor | Algorithm detections | 1-min files with nightjar calls | Days with calls | Mean number 1-min files with calls per day (excl. inactive days) |
|---|---|---|---|---|---|
| Kissos | AΜ13 | 6,527 | 523 | 95 | 5.50 |
| AΜ14 | 3,939 | 700 | 104 | 6.73 | |
| AΜ15 | 2,888 | 137 | 58 | 2.36 | |
| Ag. Vasileios | AΜ16 | 6,064 | 505 | 98 | 5.15 |
| AΜ17 | 15,173 | 857 | 112 | 7.65 | |
| AΜ18 | 4,782 | 192 | 61 | 3.14 |
Table 2.
Detections with Raven pro learning detector vs hand browsed and the recall rate of the algorithm, calculated at five (different) days for each sensor.
Table 2.
Detections with Raven pro learning detector vs hand browsed and the recall rate of the algorithm, calculated at five (different) days for each sensor.
| Sensor | Date | Learning Detector | visual browsing | Recall Rate % |
|---|---|---|---|---|
| AΜ13 | 1 - 5/7/2024 | 17 | 19 | 89.5 |
| AΜ14 | 12 - 16/6/2024 | 46 | 62 | 74.2 |
| AΜ15 | 3 - 7/6/2024 | 21 | 23 | 91.3 |
| AΜ16 | 20 - 24/5/2024 | 76 | 107 | 71.0 |
| AΜ17 | 7 - 11/5/2024 | 69 | 77 | 89.6 |
| AΜ18 | 29/6- 3/7/2024 | 29 | 38 | 76.3 |
Table 3.
Estimates and significance of the fixed effect variables predicting European Nightjar mating calling activity, as measured in number of 1-min files with calls per day per sensor. [AIC: - 1665.5, pseudo-R2 (conditional) = 0.817 (marginal) = 0.711; generalized linear mixed effect model with sensor as a random effect (intercept only)].
Table 3.
Estimates and significance of the fixed effect variables predicting European Nightjar mating calling activity, as measured in number of 1-min files with calls per day per sensor. [AIC: - 1665.5, pseudo-R2 (conditional) = 0.817 (marginal) = 0.711; generalized linear mixed effect model with sensor as a random effect (intercept only)].
| Variable | Estimate | Std. Error | Adjusted SE | Z-value | Lower 95 % CI | Lower 95 % CI | Pr(>|z|) |
|---|---|---|---|---|---|---|---|
| (Intercept) | 10.293 | 0.782 | 0.783 | 13.141 | 8.760 | 11.825 | < 0.001 *** |
| Humidity | 3.133 | 0.511 | 0.512 | 6.122 | 2.131 | 4.134 | < 0.001 *** |
| Moon Phase | 0.265 | 0.107 | 0.107 | 2.474 | 0.055 | 0.474 | 0.013 * |
| Precipitation | - 1.761 | 0.400 | 0.400 | 4.401 | - 2.545 | - 0.977 | < 0.001 *** |
| Wind gust | - 0.342 | 0.312 | 0.313 | 1.095 | - 0.953 | 0.270 | 0.274 |
| Night duration | - 40.935 | 1.837 | 1.839 | 22.254 | - 44.535 | - 37.334 | < 0.001 *** |
| Temperature (Min) | - 0.021 | 0.114 | 0.114 | 0.185 | - 0.244 | 0.202 | 0.853 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



