
Submitted:
11 March 2025
Posted:
12 March 2025
You are already at the latest version
Abstract
Sanderia malayensis is a species from the phylum Cnidaria, class Scyphozoa, and order Semaeostomeae, found in tropical waters across the Indian Ocean, Pacific Ocean, Red Sea, and Malaysia, with its range extending to Australian and Japanese waters. This study aimed to evaluate the effects of salinity on the growth and survival of S. malayensis ephyrae and to identify its optimal salinity range. The experimental design involved two temperature conditions (20°C and 24°C) and three salinity levels (21 PSU, 24 PSU, 27 PSU). The results indicated that growth and feeding abilities of S. malayensis ephyrae were significantly higher in 24 PSU and 27 PSU environments compared to 21 PSU at both temperatures. Survival rates were also higher in 24 PSU (20°C: 90%, 24°C: 79%) and 27 PSU (20°C: 87%, 24°C: 86%) compared to 21 PSU (20°C: 70%, 24°C: 55%). Despite lower survival in 21 PSU, the species exhibited notable environmental adaptability. These findings suggest that S. malayensis ephyrae are highly adaptable to varying salinity conditions, indicating the potential for expansion into South Korean waters affected by climate change, with possible implications for local marine ecosystems.
Keywords:
Sanderia malayensis
; Salinity
; Growth
; Survival Rate
; Feeding Rate
; Ephyrae
; Koreans waters
1. Introduction
Jellyfish outbreaks have significant impacts on marine ecosystems and human activities [1,2,3,4]. In particular, they are known to have highly negative social and economic effects on human activities, especially in sectors such as fisheries and tourism [5,6]. These impacts include damage to aquaculture species, loss of fishing nets, a decrease in fishery resources due to food competition, disruptions to coastal power plant systems, and human sting incidents at beaches [7,8,9,10,11].
Jellyfish outbreaks occurring worldwide are primarily attributed to Scyphozoan species from the phylum Cnidaria [12,13], and species such as Aurelia coerulea and Nemopilema nomurai frequently experience annual outbreaks in East Asian waters, including those of Korea, Japan, and China [14,15,16,17].
The life cycle of most Scyphozoans is primarily carried out through sexual reproduction [18]. Eggs fertilized through mating develop into planula larvae, which then attach to the substrate and transform into polyps. These polyps undergo asexual reproduction to form numerous colonies. Subsequently, polyps release ephyrae through strobilation, and the ephyrae grow and develop into adult medusae.
Research on Scyphozoans primarily focuses on various ecological studies of Aurelia spp. (A. coerulea, Aurelia solida, Aurelia labiata) [19,20,21,22,23,24]. Aurelia spp. have the widest distribution range globally and are not only easy to sample for experimental research but also exhibit exceptional survival in diverse environmental conditions. As a result, they have long been used as standardized model organisms for experimental studies on Scyphozoan ecology. [23,25]. However, outside of Aurelia spp. ecological research on Scyphozoans is highly limited, and there is an increasing need for broader ecological studies on Scyphozoans. [3,26]. This is particularly important as recent phenomena have shown a rise in the abundance of various Scyphozoan populations [27], and climate change-driven alterations in marine environments may lead to the expansion of their distribution ranges [28]. Such changes could potentially destabilize marine ecosystems, posing a risk of significant ecological impacts [4].
Sanderia malayensis (Hereafter S. malayensis) is a species belonging to the phylum Cnidaria, class Scyphozoa [29], and is reported to inhabit tropical waters including the Indian Ocean, Pacific Ocean, Red Sea, and Malaysian waters [30,31]. However, the distribution of this species has expanded to include the northern Australian waters and East Asian waters, including those of Japan [32,33]. The first discovery of S. malayensis in Japan was made in 1938 in the Amakusa region of Kyushu [34]. Later, in 2004, the discovery of natural polyps of S. malayensis forming colonies and inhabiting deep-sea areas of the Kagoshima region was reported [35].
Ecological studies on S. malayensis have primarily focused on various asexual reproductive strategies and habitat conditions during the polyp stage [29,30,36,37]. However, ecological research on the ephyra stage, the free-swimming phase produced through the strobilation process, remains highly limited [38].
S. malayensis has continued to be observed in the East Asian waters of Japan since 2010, with reports indicating that its distribution range is gradually expanding in the Japanese waters due to the influence of marine environmental changes [39,40,41]. This suggests that S. malayensis could potentially become an invasive species with the possibility of expanding its distribution into South Korean waters, which are geographically the closest to Japan. Shin and Choi [38] conducted an experiment that simulated the summer temperature conditions of the southern coast of Korea and confirmed the stable growth and high survival rates of S. malayensis ephyrae, suggesting that the environmental factors affecting the growth and survival of this species are more strongly influenced by temperature than by salinity. Additionally, they reported that no significant effects on growth and survival were observed when exposed to salinity levels close to those found in natural environments [38]. The ephyra stage of Scyphozoans is known to be significantly influenced by salinity conditions, which are one of the key factors affecting growth and survival [42,43], suggesting the need for further research to specifically evaluate the impact of salinity on the growth and survival of S. malayensis ephyrae.
Therefore, this study aimed to predict the potential changes in growth and survival of S. malayensis ephyrae when exposed to coastal marine environments with frequent salinity fluctuations, by creating salinity conditions lower than the natural salinity environment used in the experiment by Shin and Choi [38], and to identify the optimal salinity range for this species. Through this, the study aims to understand the impact of salinity environmental factors on the growth and survival of this species, providing fundamental insights, and contributing to the use of this data as an ecological indicator to assess the potential for the expansion of its distribution into South Korean waters due to changes in marine environments caused by future climate change.
2. Materials and Methods
2.1. Acquisition of S. malayensis Polyps and Ephyrae
For the acquisition of S. malayensis ephyrae in this experiment, S. malayensis polyps were provided by researchers at the Shinagawa Aquarium, located in Minato, Tokyo, Japan (provided in May 2015), and the original source of the polyps could not be verified. S. malayensis polyps were cultured and maintained in an incubator at the Aqua Planet Gwangyo Jellyfish Laboratory in Suwon, South Korea. The method for inducing strobilation to obtain ephyrae was based on the temperature stimulation method used by Fuchs et al. [44] and the temperature conditions under which ephyrae were released from S. malayensis polyps as reported by Avian et al. [29]. The ephyrae, released in large quantities from the polyps, were stored following the method of Shin and Choi [38]. They were maintained in a transparent acrylic cylindrical tank (Diameter 35 cm, Height 40 cm, Thickness 5 mm), where water flow was generated through an air stick to prevent the ephyrae from sinking to the bottom, until the experiment commenced. The seawater used for storage and cultivation was prepared using Red Sea Salt (Public Aquarium Part A) artificial seawater product from Red Sea, which underwent three stages of filtration: stage 1: sand filter, stage 2: biological filter, and stage 3: housing filter. No additional feeding was provided to the ephyrae to limit their growth until the commencement of the experiment.
2.2. Evaluation of Growth, Survival, and Feeding of S. malayensis Ephyrae According to the Effect of Salinity
The setup of the experimental treatments (Temperature and Salinity conditions) and the rearing information are shown in Figure 1. The tanks used in the experiment were Breeding Air Kreisel tanks designed and manufactured by Schuran Seawater Equipment BV (Netherlands). The temperature settings for the experimental treatments were designed based on the natural temperature range of 20–22°C reported by Uchida and Sugiura [45-46] and the temperature condition of 24°C, which showed the highest survival rate (98%) in the study by Shin and Choi [38]. The two temperature conditions, 20°C and 24°C, were chosen as the optimal temperature environments for evaluating the effects of salinity. The salinity settings for the experimental treatments were designed to evaluate the effects of low salinity conditions, with three low salinity treatments at 21 PSU, 24 PSU, and 27 PSU, which were established to be lower than the salinity range found in natural environments, and these treatments were applied equally in the two temperature environments of 20°C and 24°C. All experimental treatments contained 50 ephyrae, which were cultured as described in section 2-1, with each treatment set up in replicates. The seawater used in this experiment was filtered artificial seawater, the same product as used in section 2-1. The experiment lasted for a total of 20 days. During the experimental period, feeding was provided according to the method proposed by Purcell et al. [2] and the feeding method used in the study by Riisgård [47], where a sufficient amount of Artemia sp. was supplied once a day to ensure that all ephyrae in each treatment received enough food. This was to ensure that the feeding behavior of the cultured ephyrae was not restricted by providing a sufficient amount of Artemia sp. to each treatment. The water quality maintenance for ephyrae health was managed through 100% water exchange every other day with pre-filtered clean artificial seawater following the feeding of Artemia sp.
The growth, feeding, and survival records of the ephyrae were measured a total of 10 times at 2-day intervals throughout the 20-day experimental period. Growth and feeding were recorded by randomly extracting 10 ephyrae from each treatment, which were set up in replicates, and observing them under an optical microscope (Olympus model SZX2-ILLK, Tokyo, Japan). The growth size increase (mm) was measured according to the method devised by Straehler-Pohl et al. [48], using the total diameter of the ephyrae (Central disk diameter + Total marginal lappet length) as the measurement standard (Figure 2). On day 1, 10, and 20 of the experiment, the largest individual in terms of diameter was visually selected from each treatment to observe the growth changes of the ephyrae, and images were captured using an optical microscope.
The growth rate of the ephyrae (% d-1) was calculated according to the method for measuring ephyra growth rate devised by Widmer [43], as follows. This method was developed through Båmstedt et al. [49].
% Growth day-1 = ln [(D2 / D1)3] / (t2 – t1) × 100
Here, D1 represents the average diameter of the ephyrae on the first measurement day (Day 1), and D2 represents the average diameter two days later (Day 2). Since measurements were recorded at 2-day intervals for a total of 10 times during the 20-day experimental period, the average diameter at each measurement point was represented as D1 and D2 and calculated accordingly. The feeding records of the ephyrae were measured by randomly extracting ephyrae from each treatment 1 hour after feeding Artemia sp. and visually confirming the gut content under an optical microscope. The number of Artemia sp. consumed, which was considered completed feeding, was counted and recorded. The survival records of the ephyrae were measured by counting all surviving ephyrae from each treatment and selecting individuals that showed good swimming and feeding behavior, which were then recorded.
3. Statistical Analysis
To evaluate the effects of salinity environmental factors on the growth and feeding of S. malayensis ephyrae, we applied the Aligned Rank Transform ANOVA (ART-ANOVA) method. The dependent variables were growth (mm) and feeding (number of individuals), while the independent variables were the three salinity levels (21 PSU, 24 PSU, 27 PSU) and the number of days measured during the experimental period (a total of 10 measurements: 2-day intervals over the 20-day experimental period). All analyses were performed independently under the experimental temperature conditions of 20°C and 24°C. To assess the effects of salinity over the course of the experimental period, the interaction effect of salinity × experimental days was included for each temperature condition.
As a result of preliminary testing for homogeneity of variance using Levene’s test, it was found that the assumption of homogeneity was not met for the growth changes measured at the three salinity levels at 20°C and the feeding changes measured at the three salinity levels at 24°C. Therefore, to avoid the potential occurrence of statistical errors when applying traditional ANOVA, we employed a non-parametric approach based on ART-ANOVA analysis. ART-ANOVA is a validated non-parametric alternative known to effectively adjust for errors due to violations of homogeneity of variance and can also assess interaction effects between factors [50,51].
Since the assumption of homogeneity of variance was met for the feeding changes measured at the three salinity levels at 20°C according to Levene’s test, ANOVA analysis was applied. This was done to ensure high statistical power through the parametric method of ANOVA and to estimate clear statistical results and reliability (ART-ANOVA analysis results for the three salinity levels of feeding changes measured at 20°C, Supplementary Table S1, S2). The Holm adjustment was applied as a post-hoc test (For the growth changes at three salinity levels at 20°C and 24°C, and for the feeding changes at three salinity levels at 24°C). For the feeding changes measured at three salinity levels at 20°C, Tukey's Honest Significant Difference (HSD) test was used to compare the differences between groups.
To evaluate the effect of salinity environmental factors on the survival rate of S. malayensis ephyrae, we independently compared survival curves for each of the three salinity conditions at 20°C and 24°C using Kaplan-Meier survival analysis. The hazard ratio based on the survival analysis was assessed using the Cox Proportional Hazards Model to evaluate the impact of salinity differences. All statistical analyses were performed using R version 4.4.2 (R Core Team, 2024). The ARTool package [52] was applied for growth and feeding changes analysis (ART-ANOVA), and the Survival package [51] was used for survival analysis.
4. Results
4.1. Effect of Salinity Environmental Factors on the Growth of S. malayensis Ephyrae
During the 20-day experimental period, the growth changes of S. malayensis ephyrae cultured under three different salinity conditions (21 PSU, 24 PSU, 27 PSU) at 20°C and 24°C were captured on Day 1, Day 10, and Day 20. The images of these growth changes for each treatment are shown in Figure 3 and Figure 4. At the end of the 20-day experimental period, the growth changes (size increase) of S. malayensis ephyrae were measured as follows: at 20°C, the size increased to 6.34 ± 0.14 mm (Mean ± SD) at 21 PSU, 8.47 ± 0.15 mm (Mean ± SD) at 24 PSU, and 8.51 ± 0.13 mm (Mean ± SD) at 27 PSU (Figure 5, Table 1). At 24°C, the size increased to 6.35 ± 0.14 mm (Mean ± SD) at 21 PSU and 8.50 ± 0.13 mm (Mean ± SD) at both 24 PSU and 27 PSU (Figure 5, Table 1). The growth rate (% d-1) of S. malayensis ephyrae during the 20-day experimental period was as follows: at 20°C, the growth rate was 10.31 % d-1 at 21 PSU, 14.88 % d-1 at 24 PSU, and 14.86 % d-1 at 27 PSU. At 24°C, the growth rate was 10.24 % d-1 at 21 PSU, 14.80 % d-1 at 24 PSU, and 14.94 % d-1 at 27 PSU (Table 2). The results of the ART-ANOVA analysis for growth changes under three different salinity conditions at 20°C and 24°C showed statistically significant main effects for time (20°C: F = 633.97, 24°C: F = 633.98), salinity (20°C: F = 6023.03, 24°C: F = 6284.18), and the interaction effect between time and salinity (20°C: F = 234.34, 24°C: F = 207.09). These results are presented in Table 3 (p <0.001). To further examine the interaction effect between time and the three different salinity environments, the Holm post-hoc test results showed that the growth at 21 PSU was significantly lower than at 24 PSU and 27 PSU at both 20°C and 24°C (Table 1, p < 0.001). However, the difference between 24 PSU and 27 PSU was not statistically significant (Table 1, 20°C: p = 0.461, 24°C: p = 0.840).
4.2. Effect of Salinity Environmental Factors on the Feeding of S. malayensis Ephyrae
During the 20-day experimental period, the average feeding rate (Preys ind·hr⁻¹) of S. malayensis ephyrae was as follows: at 20°C, 2.10 ± 0.908 Preys ind·hr⁻¹ (Mean ± SD) at 21 PSU, 7.58 ± 1.138 Preys ind·hr⁻¹ (Mean ± SD) at 24 PSU, and 7.69 ± 1.165 Preys ind·hr⁻¹ (Mean ± SD) at 27 PSU (Figure 6, Table 4). At 24°C, the average feeding rate was 2.16 ± 0.869 Preys ind·hr⁻¹ (Mean ± SD) at 21 PSU, 7.75 ± 1.118 Preys ind·hr⁻¹ (Mean ± SD) at 24 PSU, and 7.64 ± 1.136 Preys ind·hr⁻¹ (Mean ± SD) at 27 PSU (Figure 6, Table 4). Levene’s test was used for preliminary testing of homogeneity of variance, and based on the results, the analysis method was chosen. The average feeding rate at 20°C was analyzed using ANOVA, while the average feeding rate at 24°C was analyzed using ART-ANOVA (Section 3. Statistical Analysis). The results of the ANOVA analysis for the average feeding rate (Preys ind·hr⁻¹) under three different salinity conditions (21 PSU, 24 PSU, 27 PSU) at 20°C showed statistically significant main effects for time (F = 2029.04) and salinity (F = 3.16; Table 5, p < 0.001). However, the interaction effect between time and salinity (F = 1.47) was not statistically significant (Table 5, p = 0.085). To further examine the differences in the average feeding rate (Preys ind·hr⁻¹) between the three salinity conditions, Tukey post-hoc analysis revealed significant differences between 21 PSU and 24 PSU, as well as between 21 PSU and 27 PSU (Table 4, p < 0.001). However, no significant difference was found between 24 PSU and 27 PSU, indicating that the average feeding rate was similar in the relatively higher salinity environments of 24 PSU and 27 PSU compared to 21 PSU (Table 4, p = 0.522).
The results of the ART-ANOVA analysis for the average feeding rate (Preys ind·hr⁻¹) under three different salinity conditions at 24°C showed statistically significant main effects for time (F = 633.18), salinity (F = 3.27), and the interaction effect between time and salinity (F = 2.75) (Table 6, p <0.001). To further examine the interaction effects between time and the three salinity conditions, the Holm post-hoc test revealed significant differences between 21 PSU and 24 PSU, as well as between 21 PSU and 27 PSU (Table 4, p < 0.05). However, no significant difference was found between 24 PSU and 27 PSU, indicating that the average feeding rate was similar in the relatively higher salinity environments of 24 PSU and 27 PSU compared to 21 PSU (Table 4, p = 0.100).
4.3. Effect of Salinity Environmental Factors on the Survival of S. malayensis Ephyrae
The Kaplan–Meier survival analysis results for S. malayensis ephyrae cultured under three different salinity conditions (21 PSU, 24 PSU, 27 PSU) at 20°C and 24°C during the 20-day experimental period are shown in Figure 7. The survival rates at 20°C were 70% (95% CI: 62–80%) at 21 PSU, 90% (95% CI: 84–96%) at 24 PSU, and 87% (95% CI: 81–94%) at 27 PSU (Figure 7, Table 7). The results of the Log-rank test showed a significant difference in the survival curves between the three different salinity conditions (χ² = 17.746, df = 2, Figure 7, p < 0.001). The survival rates at 24°C were 55% (95% CI: 46–66%) at 21 PSU, 79% (95% CI: 69–86%) at 24 PSU, and 86% (95% CI: 79–93%) at 27 PSU (Figure 7, Table 7). The Log-rank test results also showed a significant difference in the survival curves between the three salinity conditions (χ² = 28.008, df = 2, Figure 7, p < 0.001). In the Cox Hazard Ratio analysis to assess the effects of the three different salinity conditions (21 PSU, 24 PSU, 27 PSU), with 21 PSU set as the reference, the survival rates at both 20°C and 24°C were found to significantly increase at 24 PSU and 27 PSU (p < 0.001, Supplementary Figure S1, S2). At 20°C, the hazard ratios for 24 PSU and 27 PSU were 0.29 (95% CI: 0.14–0.59, p < 0.001) and 0.38 (95% CI: 0.20–0.72, p = 0.003), respectively. At 24°C, the hazard ratios for 24 PSU and 27 PSU were 0.42 (95% CI: 0.25–0.69, p < 0.001) and 0.25 (95% CI: 0.14–0.46, p < 0.001), respectively. The p-value for the Global Log-Rank Test was calculated to be <0.001, indicating that there was a statistically significant difference in survival rates between the three different salinity conditions (p <0.001).
5. Discussion
This study aimed to evaluate the effects of salinity on the growth and survival of S. malayensis ephyrae and to determine its optimal salinity range. S. malayensis ephyrae showed stable growth changes in environments with salinities of 24 PSU and 27 PSU (Figure 5, Table 1), which was also confirmed by the survival rate results (Figure 7, Table 7). The environment with 21 PSU showed significantly lower growth changes (Figure 5, Table 1, p < 0.001) and survival rates (Figure 7, Table 7, p < 0.001) compared to 24 PSU and 27 PSU. However, a certain level of environmental adaptability was observed. These findings demonstrate that this species has the ability to adapt very well to a wide range of salinity conditions. Previous studies on S. malayensis ephyrae have primarily focused on morphological characteristics [36,48,53], as well as ecological studies during the polyp stage [29-30,37,46]. However, biological and ecological research on the free-swimming phase, which follows the polyp stage, remains largely unexplored [38]. This may be due to the technical challenges involved in studying the free-swimming stage, including the need for advanced techniques and the establishment of life-support systems [54]. S. malayensis is known to be a representative species of Scyphozoans commonly exhibited in aquariums worldwide [29], and aquariums possess the necessary technologies and manuals for year-round display of various Scyphozoans [55]. This aspect may present one of the significant challenges faced by researchers in studying the free-swimming phase of Scyphozoan species, including S. malayensis. The effects of salinity environmental factors on the survival and growth of Scyphozoan ephyrae during the free-swimming stage have been reported in various studies [23,42-43,56-58]. Båmstedt et al. [42] found no significant impact on growth rate and growth efficiency between 17.5 PSU and 35 PSU in their experiment on A. aurita, but they reported very high feeding rates when exposed to 35 PSU, suggesting that the stable feeding rate of A. aurita is attributed to higher salinity environments. Purcell et al. [56] suggested that C. quinquecirrha prefers survival in environments with salinities below 20 PSU, and Windmer [43] confirmed a high growth rate of C. capillata in the low salinity environment of 21 PSU, while C. hyoscella showed maximum growth rate at 34 PSU. Additionally, in studies on A. coerulea, Fu et al. [57] reported that A. coerulea increased its growth rate when exposed to lower salinity, and research by Schäfer et al. [23] and Yu et al. [58] also suggested that salinity levels of 20 PSU and 25 PSU resulted in higher growth rates of A. coerulea compared to environments with salinities above 30 PSU, indicating that lower salinity environments had a significant effect on growth. In this study, S. malayensis ephyrae showed a tendency for both growth (Figure 5, Table 1) and survival (Figure 7, Table 7) to increase with higher salinity, while feeding rates were found to be lower in the 21 PSU environment compared to 24 PSU and 27 PSU (Figure 6, Table 4, p < 0.001). These results suggest that this species has the ability to adapt to a wide range of salinity conditions, but it prefers higher salinity environments for stable growth and survival. In the study by Shin and Choi [38], which simulated the summer temperature conditions of the southern coast of Korea, no significant differences were observed in the growth and survival of S. malayensis ephyrae under salinity conditions of 30–33 PSU at 24°C, with a very high survival rate of 98%. In this study, the survival rates at 24 PSU and 27 PSU were 90% and 87% (Figure 7, Table 7) at 20°C, and 79% and 86% (Figure 7, Table 7) at 24°C. However, the survival rate at 21 PSU was significantly lower than at 24 PSU and 27 PSU (20°C: 70%, 24°C: 55%, P < 0.001). Therefore, based on the results of both the Shin and Choi [38] study and this study, the ideal salinity range for the growth and survival of S. malayensis ephyrae is estimated to be between 24 PSU and 33 PSU. In the study by Shin and Choi [38], the salinity range of 30 PSU to 33 PSU may represent the stable optimal salinity range for this species. Chrysaora species such as C. capillata and C. lamarckii have a medusivores ecology, feeding on gelatinous plankton [43], and Avian et al. [29] reported in a laboratory study that S. malayensis ephyrae were observed to unilaterally prey on A. solida ephyrae. In natural environments, the abundance of zooplankton is a contributing factor to Scyphozoan outbreaks [1] and feeding on prey with appropriate nutrients has a significant impact on the growth and survival of Scyphozoans [59]. The most commonly used zooplankton in global Scyphozoan research are Artemia sp. and Brachiounus spp. [8,60]. In this study, Artemia sp. was provided as the prey for S. malayensis ephyrae, with no additional nutritional supplementation. Considering that S. malayensis has a medusivores ecology feeding on gelatinous plankton [29], it is possible that rapid growth could be induced when S. malayensis ephyrae prey on Aurelia spp. (A. solida, A. labiata, A. coerulea). Additionally, if S. malayensis expands its distribution into South Korean waters in the future, it may compete for food in the marine ecosystem with A. coerulea, which is known for its outbreaks during the summer and may prey on A. coerulea [4]. This possibility is supported by the findings of this study, which demonstrate that S. malayensis is highly adaptable to a wide range of salinity conditions, similar to the results found in studies on Aurelia spp. [23,42,57-58,61]. Shin and Choi [38] suggested that the optimal temperature range for the growth and survival of S. malayensis is between 20°C and 24°C, while Avian et al. [29] reported that the polyps of this species have the ability to reproduce even at 20°C. Furthermore, this species has a wide distribution range, not only in the Indian Ocean, Pacific Ocean, and Suez Canal but also in East Asian waters, including Japan [31]. Notably, the species has been reported to appear from spring to summer May to August in the waters of Japan, which are geographically close to the waters of South Korea [39-41]. This study confirmed that S. malayensis ephyrae possess a high level of adaptability, allowing for stable growth and survival across a range of salinity environments. These findings suggest that S. malayensis could potentially become an invasive species capable of expanding its distribution into South Korean waters, which are affected by climate change. Furthermore, there is a pressing need for additional biological and ecological research to understand the potential impacts of this species' presence on marine ecosystems.
6. Conclusions
This study confirmed that S. malayensis ephyrae exhibit stable growth (Figure 5, Table 1) and high survival rates (Figure 7, Table 7) across a range of salinity environments, demonstrating their high adaptability to varying salinity conditions. Considering that S. malayensis is distributed in the East Asian waters, including Japan, these findings suggest the potential for this species to expand its distribution into South Korean waters, which are influenced by climate change. If the distribution of this species expands, there is a pressing need for additional research to understand the potential impacts on marine ecosystems. Future studies should focus on thoroughly analyzing the potential ecological consequences of this species' distribution expansion on local ecosystems.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1, S2: ART-ANOVA analysis results for feeding rate changes at three salinity levels (21 PSU, 24 PSU, 27 PSU) measured at 20°C. Figure S1, S2: Cox Hazard Ratio analysis results for survival rates at salinities of 21 PSU, 24 PSU, and 27 PSU under two different temperatures (20°C and 24°C) for evaluation.
Author Contributions
Conceptualization, K.H.S. and K.H.C.; Methodology, K.H.S. and K.H.C.; Formal Analysis and Investigation, K.H.S.; Supervision, K.H.C.; Writing—Original Draft Preparation, K.H.S.; Writing—Review and Editing, K.H.S. and K.H.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was conducted with guidance from Chungnam National University, but without external funding or institutional support.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are available upon reasonable request. The data are either included in the main body of the article or provided as supplementary materials.
Acknowledgements
We would like to express our sincere gratitude to Aqua Planet Aquarium for providing and supporting the Sanderia malayensis ephyrae for this study. We also extend our heartfelt thanks to Keun-Hyung Choi from the Department of Earth, Environmental and Space Sciences at Chungnam National University for his extensive guidance and invaluable advice throughout the course of this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Purcell, J.E.; Uye, S.; Lo, W.-T. Anthropogenic causes of jellyfish blooms their direct consequences for humans: A review. Mar. Ecol. Prog. Ser. 2007, 350, 153-174. https://doi.org/10.3354/meps07093. [CrossRef]
- Purcell, J.E.; Atienza, D.; Fuentes, V.; Olariaga, A.; Tilves, U.; Colahan, C.; Gili, J.M. Temperature effects on asexual reproduction rates of scyphozoan species from the northwest Mediterranean Sea. Hydrobiologia 2012, 690, 169-180. http://doi.org/10.1007s10750-012-1047-7. [CrossRef]
- Pitt, K.A.; Lucas, C.H.; Condon, R.H.; Duarte, C.M.; Stewart-Koster, B. Claims That Anthropogenic Stressors Facilitate Jellyfish Blooms Have Been Amplified Beyond the Available Evidence: A Systematic Review. Front. Mar. Sci. 2018, 5, 451. http://doi.org/10.3389/fmars.2018.00451. [CrossRef]
- Lee, S.H.; Scotti, M.; Jung, S.J.; Hwang, J.S. Jellyfish blooms challenge the provisioning of ecosystem services in the Korean coastal waters. Hydrobiologia 2023, 850, 2855-2870. http://doi.org/10.1007/s10750-022-05076-4. [CrossRef]
- Bosch-Belmar, M.; Milisenda, G.; Basso, L.; Doyle, T.K. Jellyfish Impacts on Marine Aquaculture and Fisheries. Rev. Fish. Sci. Aquac. 2023, 29, 242-259. http://doi.org/10.1080/23308249.2020.1806201. [CrossRef]
- Fernández-Alías, A.; Marcos, C.; Ismael, J.; Sabah, S.; Pérez-Ruzafa, A. Population dynamics and growth in three scyphozoan jellyfishes, and their relationship with environmental conditions in a coastal lagoon. Estuar. Coast. Shelf. Sci. 2020, 243, 106901. https://doi.org/10.1016/j.ecss.2020.106901. [CrossRef]
- Quiñones, J.; Monroy, A.; Acha, E. M.; Mianzan, H. Jellyfish bycatch diminishes profit in an anchovy fishery off Peru. Fish. Res. 2013, 139, 47-50. http://doi.org/10.1016/j.fishres.2012.04.014. [CrossRef]
- Purcell, J.E.; Baxter, E.J.; Fuentes, V. 13-Jellyfish as products and problems of aquaculture. In Advances in Aquaculture Hatchery Technology, 1st ed.; Allan, G., Burnell, G., Eds.; Food Science, Technology and Nutrition; Woodhead Publishing: Cambridge, UK, 2013; pp. 404-430. http://doi.org/10.1533/9780857097460.2.404. [CrossRef]
- De Donno, A.; Idolo, A.; Bagordo, F.; Grassi, T.; Leomanni, A.; Serio, F.; Guido, M.; Canitano, M.; Zampardi, S.; Boero, F.; Piraino, S. Impact of stinging jellyfish proliferations along south Italian coasts: Human health hazards, treatment and social costs. Int. J. Environ. Res. Public Health 2014, 1, 2488-2503. http://doi.org/10.3390/ijerph110302488. [CrossRef]
- Milisenda, G.; Martinez-Quintana, A.; Fuentes, V.L.; Bosch-Belmar, M.; Aglieri, G.; Boero, F.; Piraino, S. Reproductive and bloom patterns of Pelagia noctiluca in the Strait of Messina, Italy. Estuar. Coast. Shelf Sci. 2018, 201, 29-39. http://doi.org/10.1016/j.ecss.2016.01.002. [CrossRef]
- Tilves, U.; Fuentes, V.L.; Milisenda, G.; Parrish, C.C.; Vizzini, S.; Sabatés, A. Trophic interactions of the jellyfish Pelagia noctiluca in the NW Mediterranean: Evidence from stable isotope signatures and fatty acid composition. Mar. Ecol. Prog. Ser. 2018, 591, 101-116. http://doi.org/10.3354/meps12332. [CrossRef]
- Frolova, A.; Miglietta, M.P. Insights on Bloom Forming Jellyfish (Class: Scyphozoa) in the Gulf of Mexico: Environmental Tolerance Ranges and Limits Suggest Differences in Habitat Preference and Resistance to Climate Change Among Congeners. Front. Mar. Sci. 2020, 7, 93. http://doi.org/10.3389/fmars.2020.00093. [CrossRef]
- Tang, C.; Sun, S.; Zhang, F. Natural predators of polyps of three scyphozoans: Nemopilema nomurai, Aurelia coerulea, and Rhopilema esculentum. J.Oceanol. Limnol. 2021, 39, 598-608. http://doi.org/10.1007/s00343-020-0284-2. [CrossRef]
- Ishii, H. The influence of environmental changes upon the coastal plankton ecosystems, with special reference to mass occurrence of jellyfish. Bull. Plankt. Soc. Jpn. 2001, 48, 55-61.
- Wang, S.; Zhang, G.; Sun, S.; Wang, Y.; Zhao, Z. Population dynamics of three Scyphozoan Jellyfish species during summer of 2011 in Jiaozhou Bay. Oceanol. Limnol. Sin. 2012, 43, 471-479.
- Sun, M.; Dong, J.; Purcell, J.E.; Li, Y.; Duan, Y.; Wang, A.; Wang, B. Testing the influence of previous–year temperature and food supply on development of Nemopilema nomurai blooms. Hydrobiologia 2015, 754, 85–86. http://doi.org/10.1007/s10750-014-2046-7. [CrossRef]
- Lee, S.H.; Hwang, J.S.; Kim, K.Y.; Molinero, J.C. Contrasting Effects of Regional and Local Climate on the Interannual Variability and Phenology of the Scyphozoan, Aurelia coerulea and Nemopilema nomurai in the Korean Peninsula. Diversity 2021, 13, 214. http://doi.org/10.3390/d13050214. [CrossRef]
- Helm, R.R. Evolution and development of scyphozoan jellyfish. Biol. Rev. Camb. Philos. Soc. 2018, 93, 1228-1250. http://doi.org/10.1111/brv.12393. [CrossRef]
- Feng, S.; Wang, S.; Sun, S.; Zhang, F.; Zhang, G.; Mengtan, L.; Uye, S-I. Strobilation of three scyphozoans (Aurelia coelurea, Nemopilema nomurai and Rhopilema esculentum) in the field at Jiaozhou Bay, China. Mar. Ecol. Prog. Ser. 2018, 591, 141-153. http://doi.org/10.3354/meps12276. [CrossRef]
- Xing, Y.; Liu, Q.; Zhang, M.; Zhen, Y.; Mi, T.; Yu, Z. Effects of temperature and salinity on the asexual reproduction of Aurelia coerulea polyps. J. Oceanol. Limnol. 2019, 38, 133-142. http://doi.org/10.1007/s00343-019-8337-0. [CrossRef]
- Loveridge, A.; Lucas, C.H.; Pitt, K.A. Shorter, warmer winters may inhibit production of ephyrae in a population of the moon jellyfish Aurelia Aurita. Hydrobiologia 2021, 848, 739-749. https://doi.org/10.1007/s10750-020-04483-9. [CrossRef]
- Takauchi, S.; Miyake, H.; Hirata, N.; Nagai, M.; Suzuki, N.; Ogiso, S.; Ikeguchi, S. Morphological characteristics of ephyrae of Aurelia coerulea derived from planula strobilation. Fish. Sci. 2021, 87, 617- 679. http://doi.org/10.1007/s12562-021-01541-6. [CrossRef]
- Schäfer, S.; Gueroun, S.K.M.; Andrade, C.; Canning-Clode, J. Combined Effects of Temperature and Salinity on Polyps and Ephyrae of Aurelia solida (Cnidaria: Scyphozoa). Diversity 2021, 13, 573. http://doi.org/10.3390/d13110573. [CrossRef]
- Fernández-Alías, A.; Marcos, C.; Perez-Ruzafa, A. Reconstructing the Biogeographic History of the Genus Aurelia Lamarck, 1816 (Cnidaria, Scyphozoa), and Reassessing the Nonindigenous Status of A. solida and A. coerulea in the Mediterranean Sea. Diversity 2023, 15, 1181. http://doi.org/10.3390/d15121181. [CrossRef]
- Scorrano, S.; Aglieri, G.; Boero, F.; Dawson, M.N.; Piraino, S. Unmasking Aurelia species in the Mediterranean Sea: An integrative morphometric and molecular approach. Zool. J. Linn. Soc. 2017, 180, 243-267. http://doi.org/10.1111/zoj.12494. [CrossRef]
- Fernández-Alías, A.; Marcos, C.; Pérez-Ruzafa, A. The unpredictability of scyphozoan jellyfish blooms. Front, Mar. Sci 2024, 11. http://doi.org/10.3389/fmars.2024.1349956. [CrossRef]
- Fernández-Alías, A.; Marcos, C.; Perez-Ruzafa, A. Larger scyphozoan species dwelling in temperate, shallow waters show higher blooming potential. Mar. pollut. Bull. 2021, 173, 113100. http://doi.org/10.1016/j.marpolbul.2021.113100. [CrossRef]
- Boero, F.; Brotz, L.; Gibbons, M.J.; Piraino, S.; Zampardi, S. 3.10 Impacts and Effects of Ocean Warming on Jellyfish. In Explaining Ocean Warming: Causes, Scale, Effects and Consequences; IUCN: Gland, Switzerland, 2016; pp. 213-237. Available online: http://www.researchgate.net/publication/309209466.
- Avian, M.; Motta, G.; Prodan, M.; Tordoni, E.; Macaluso, V.; Beran, A.; Goruppi, A.; Bacaro, G.; Tirelli, V. Asexual reproduction and strobilation of S. malayensis (Scyphozoa, Pelagiidae) in relation to temperature: Experimental evidence and implications. Diversity 2021, 13, 37. https://doi.org/10.3390/d13020037. [CrossRef]
- Adler, L.; Jarms, G. New insights into reproductive traits of scyphozoans: Special methods of propagation in S. malayensis Goette, 1886 (Pelagiidae, Semaeostomeae) enable establishing a new classification of asexual reproduction in the class Scyphozoa. Mar Biol 2009, 156, 1411-1420. https://doi.org/10.1007/s00227-009-1181-6. [CrossRef]
- Morandini, A.C.; Gul, S. Rediscovery of S. malayensis and remarks on Rhopilema nomadica record in Pakistan (Cnidaria: Scyphozoa) Pap. Avulsos Zool 2016, 56, 171-175. https://doi.org/10.1590/0031-1049.2016.56.15. [CrossRef]
- Kramp, P.L. Synopsis of the medusae of the world. J. Mar. Biol. Assoc. UK 1961, 40, 7-382.
- Gershwin, L.A.; Zeidler, W. Two new jellyfishes (Cnidaria: Scyphozoa) from tropical Australian waters. Zootaxa 2008, 1764, 41-52. https://doi.org/10.5281/zenodo.181987. [CrossRef]
- Uchida, T. Medusae in the vicinity of the Amakusa Marine Biological Station. Bull. Biogeogr. Soc. Japan 1938, 8, 143–149.
- Miyake, H.; Hashimoto, J.; Chikuchishin, M.; Miura, T. Scyphopolyps of Sanderia malayensis and Aurelia aurita attached to tubes of vestimentiferan tubeworm (Lamellibrachia satsuma) at submarine fumaroles in Kagoshima Bay. Mar. Biotechnol. 2004, 6, S174-S178. Straehler-Pohl, I.; Widmer, C.L.; Morandini, A.C. Characterizations of juvenile stages of some semaeostome Scyphozoa (Cnidaria), with recognition of a new family (Phacellophoridae). Zootaxa 2011, 2741, 1-37. https://doi.org/10.11646/zootaxa.2741.1.1. [CrossRef]
- Schiariti, A.; Morandini, A.C.; Jarms, G.; Paes, R.V.G.; Franke, S.; Mianzan, H. Asexual reproduction strategies and blooming potential in Scyphozoa. Mar. Ecol. Prog. Ser. 2014, 510, 241-253. http://doi.org/10.3354/meps10798. [CrossRef]
- Shin, K.H.; Choi, K.H. Effects of the seawater temperature and salinity on the survival and growth of the Sanderia malayensis (Cnidaria: Scyphozoa) ephyrae. Mar. Biol. Res. 2025, in press (accepted). https://doi.org/10.1080/17451000.2025.2462828. [CrossRef]
- Kubota, S. Appearance of Sanderia malayensis at Shirahama, Wakayama Prefecture, Japan after half a century absence. Nanki Biol. Soc. 2012, 54, 147-148.
- Minemizu, R.; Kobota, S.; Hirano, Y.; Lindsay, D.J. A photographic guide to the jellyfishes of Japan [Japanese]; Heibonsha: Tokyo, Japan, 2015; pp. 268.
- Toshino, S.; Yamashiro, H.; Tanimoto, M. New record of Sanderia malayensis from the Ryukyu Archipelago, southern Japan. Fauna Ryukyuana 2018, 43, 19-25.
- Båmstedt, U.; Lane, J.; Martinussen, M.B. Bioenergetics of ephyra larvae of the scyphozoan jellyfish Aurelia aurita in relation to temperature and salinity. Mar. Biol. 1999, 135, 89-98. http://doi.org/10.1007/s002270050605. [CrossRef]
- Widmer, C.L. Influences of temperature and salinity on asexual reproduction and development of scyphozoan jellyfish from the British Isles. Dissertation, University of St Andrews, 2014; pp. 72-85.
- Fuchs, B.; Wang, W.; Graspeuntner, S.; Li, Y.; Insua, S.; Herbst, E-M.; Dirksen, P.; Boehm, A-M.; Hemmrich, G.; Sommer, F.; et al. Regulation of polyp-to-jellyfish transition in Aurelia aurita. Curr. Biol. 2014, 24, 263-273. http://doi.org/10.1016/j.cub.2013.12.003. [CrossRef]
- Uchida, T.; Sugiura, Y. On the ephyra and postephyra of Semaeostome medusa, Sanderia malayensis Goette, 1886. J. Fac. Sci. Hokkaido Univ. Ser. VI Zool. 1975, 19, 879-881. Uchida, T.; Sugiura, Y. On the polyp of the Scyphomedusa, Sanderia malayensis and its reproduction. J. Fac. Sci. Hokkaido Univ. Ser. VI Zool. 1978, 21, 279-286.
- Riisgård, H.U. Superfluous Feeding and Growth of Jellyfish Aurelia aurita. J Mar Sci Eng 2022, 10, 1368. https://doi.org/10.3390/jmse10101368. [CrossRef]
- Straehler-Pohl, I.; Jarms, G. Identification key for young ephyrae: A first step for early detection of jellyfish blooms. Hydrobiologia 2010, 645, 3-21. http://doi.org/10.1007/s10750-010-0226-7. [CrossRef]
- Båmstedt, U.; Ishii, H.; Martinussen, M.B. Is the scyphomedusa Cyanea capillata (L.) dependent on gelatinous prey for its early development? Sarsia 1997, 82, 269-273.
- Wobbrock, J.O.; Findlater, L.; Gergle, D.; Higgins, J.J. The Aligned Rank Transform for Nonparametric Factorial Analyses Using Only Anova Procedures. In Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, Vancouver, BC, Canada, 7–12 May 2011; pp. 143-146. http://doi.org/10.1145/1978942.1978963. [CrossRef]
- Elkin, L.A.; Kay, M.; Higgins, J.J.; Wobbrock, J.O. An Aligned Rank Transform procedure for multifactor contrast tests. In Proceedings of the 34th Annual ACM Symposium on User Interface Software and Technology, 10-14 October 2021. http://doi.org/10.1145/3472749.3474784. [CrossRef]
- Therneau, T.M.; Grambsch, P.M. The cox model. In Modeling Survival Data: Extending the Cox Model; Springer: Berlin/Heidelberg, Germany, 2000; pp. 39-77. http://doi.org/10.1007/978-1-4757-3294-8. [CrossRef]
- Heins, A.; Sötje, I.; Holst, S. Assessment of investigation techniques for scyphozoan statoliths, with focus on early development of the jellyfish Sanderia malayensis. Mar. Ecol. Prog. Ser. 2018, 591, 37-56.
- Ballesteros, A.; Siles, P.; Jourdan, E.; Gili, J-M. Versatile aquarium for jellyfish: A rearing system for the biomass production of early life stages in flow-through or closed systems. Front Mar Sci 2022, 9, 942094. https://doi.org/10.3389/fmars.2022.942094. [CrossRef]
- Crow, J.; Howard, M.; Lévesque, V.; Matsushige, L.; Spina, S.; Schaadt, M.; Sowinski, N, Widmer, C.L et al. AZA Aquatic Invertebrate TAG. In Jellyfish (Cnidaria/Ctenophora) Care Manual; Schaadt, M., Ed.; Association of Zoos and Aquariums (AZA): Silver Spring, MD, USA, 2013; pp. 5-79.
- Purcell, J.E.; White, J.R.; Nemazie, D.A.; Wright, D.A. Temperature, salinity and food effects on asexual reproduction and abundance of the scyphozoan Chrysaora quinquecirrha. Mar. Ecol. Prog. Ser. 1999, 180, 187-196. http://doi.org/10.3354/meps180187. [CrossRef]
- Fu, Z.; Li, J.; Wang, J.; Lai, J.; Liu, Y.; Sun, M. Combined effects of temperature and salinity on the growth and pulsation of moon jellyfish (Aurelia coerulea) ephyrae. Am. J. Life Sci. 2020, 8, 144-151. https://doi.org/10.11648/j.ajls.20200805.19. [CrossRef]
- Yu, S.; Song, D.; Fan, M.; Xie, C. Effects of temperature and salinity on growth of Aurelia aurita. Ecol. Model. 2023, 476, 110229. http://doi.org/10.1016/j.ecolmodel.2022.110229. [CrossRef]
- Duarte, I.M.; Marques, S.C.; Leandro, S.M.; Calado, R. An overview of jellyfish aquaculture: For food, feed, pharma and fun. Rev. Aquac. 2022, 14, 265–287. http://doi.org/10.1111/raq.12597. [CrossRef]
- Miranda, F.A.S.; Chambel, J.; Almeida C.; Pires, D.; Dauarte I.; Esteves, L.; Maranhão, P. Effect of different diets on growth and survival of the white-spotted jellyfish, Phyllorhiza punctata. Front. Mar. Sci. 2016, 3. http://doi.org/10.3389/conf.FMARS.2016.04.00042. [CrossRef]
- Widmer, C.L. Effects of temperature on growth of north-east Pacific moon jellyfish ephyrae, Aurelia labiata (Cnidaria: Scyphozoa). J. Mar. Biol. Assoc. United Kingd. 2005, 85, 569-573. https://doi.org/10.1017/S0025315405011495. [CrossRef]
- Experimental design: Breeding Air Kreisel tank setup for S.malayensis ephyrae.
Figure 1.
The experimental design to assess the effects of low salinity on the survival and growth of S. malayensis ephyrae is as follows. The experiment was conducted under two temperature conditions, 20°C and 24°C, with three different salinity levels. Each experimental group was replicated twice, with 50 ephyrae cultured in each Breeding Air Kreisel tank. As a result, 100 ephyrae were cultured per salinity level under each temperature condition, totaling 600 ephyrae across both temperature conditions.
Figure 1.
The experimental design to assess the effects of low salinity on the survival and growth of S. malayensis ephyrae is as follows. The experiment was conducted under two temperature conditions, 20°C and 24°C, with three different salinity levels. Each experimental group was replicated twice, with 50 ephyrae cultured in each Breeding Air Kreisel tank. As a result, 100 ephyrae were cultured per salinity level under each temperature condition, totaling 600 ephyrae across both temperature conditions.
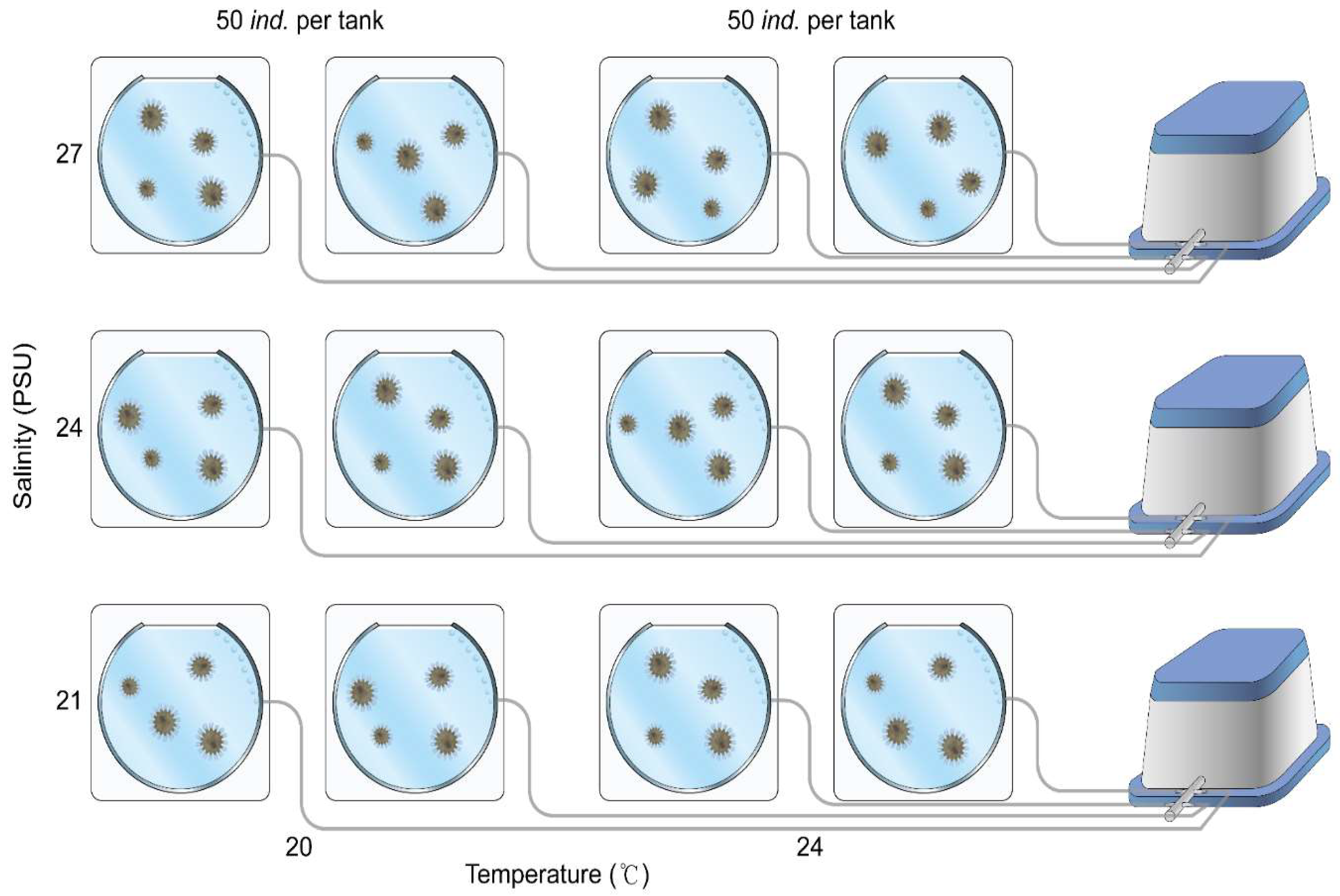
Figure 2.
In this experiment, the measurement points and range criteria for assessing the growth changes of S. malayensis ephyrae were adapted from Straehler-Pohl et al. (2010). Measurements were taken based on the central disc diameter (mm) and total marginal lappet length (mm) of the ephyrae.
Figure 2.
In this experiment, the measurement points and range criteria for assessing the growth changes of S. malayensis ephyrae were adapted from Straehler-Pohl et al. (2010). Measurements were taken based on the central disc diameter (mm) and total marginal lappet length (mm) of the ephyrae.
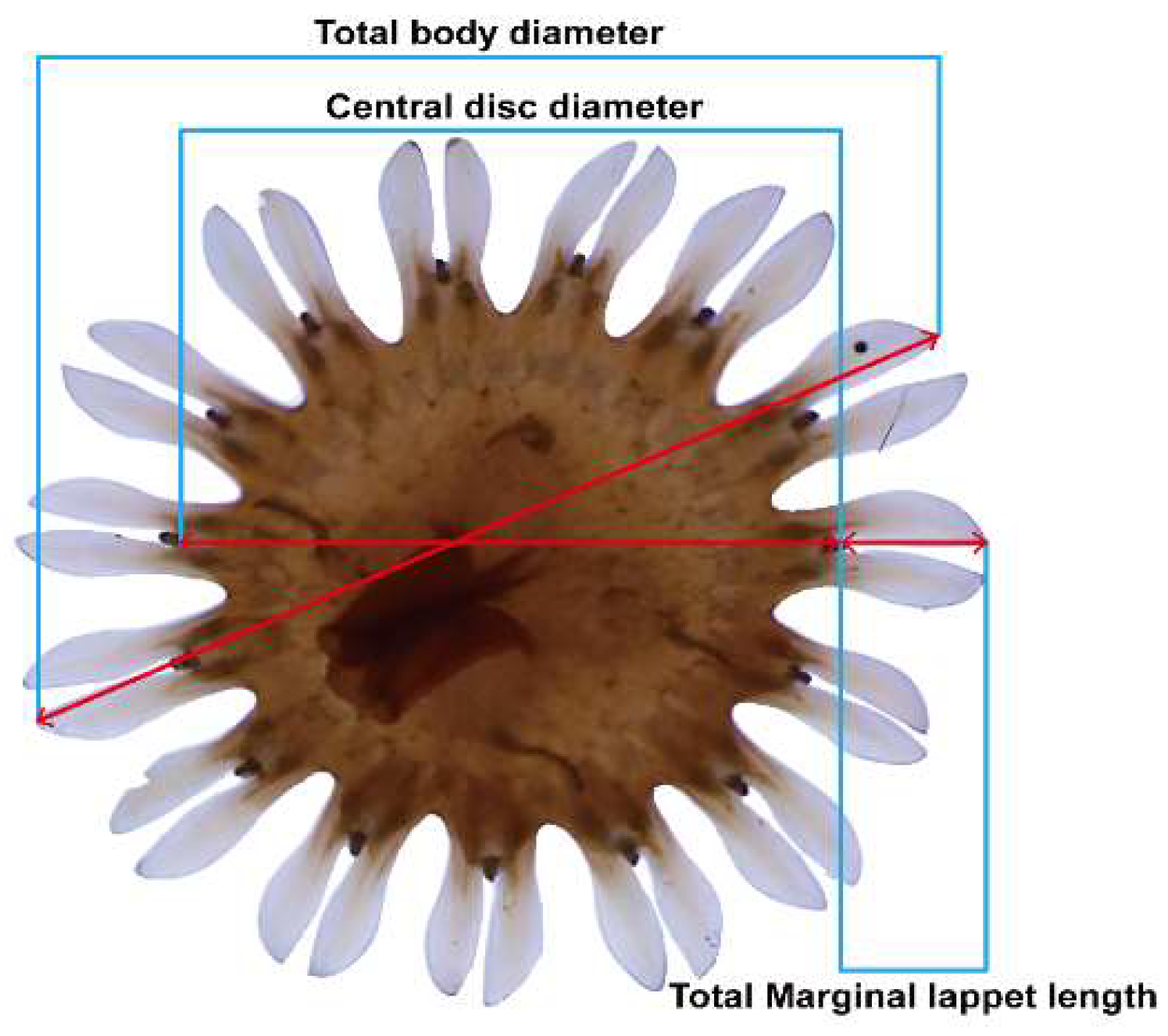
Figure 3.
The growth changes of S. malayensis ephyrae were recorded three times during the 20-day experimental period (On days 1, 10, and 20). The experiment was conducted at a temperature of 20°C with three different salinity conditions: 21PSU, 24PSU, and 27PSU.
Figure 3.
The growth changes of S. malayensis ephyrae were recorded three times during the 20-day experimental period (On days 1, 10, and 20). The experiment was conducted at a temperature of 20°C with three different salinity conditions: 21PSU, 24PSU, and 27PSU.

Figure 4.
The growth changes of S. malayensis ephyrae were recorded three times during the 20-day experimental period (On days 1, 10, and 20). The experiment was conducted at a temperature of 24°C with three different salinity conditions: 21PSU, 24PSU, and 27PSU.
Figure 4.
The growth changes of S. malayensis ephyrae were recorded three times during the 20-day experimental period (On days 1, 10, and 20). The experiment was conducted at a temperature of 24°C with three different salinity conditions: 21PSU, 24PSU, and 27PSU.
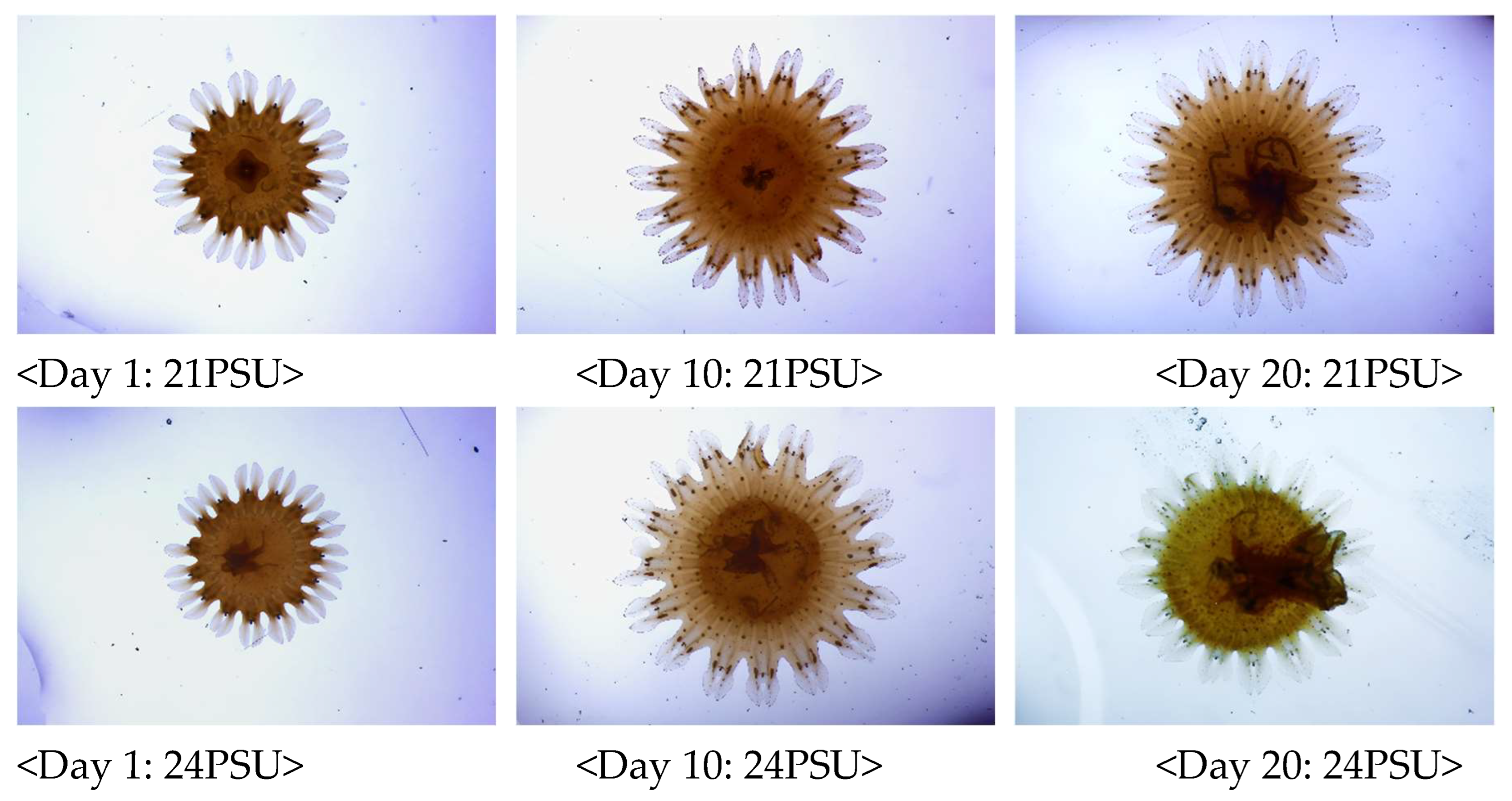
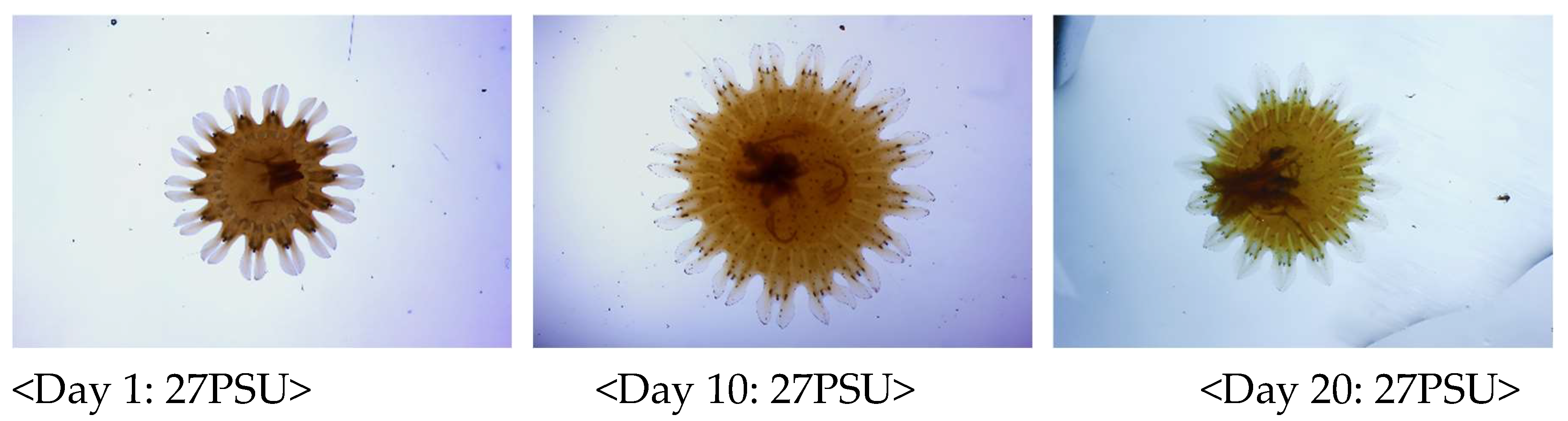
Figure 5.
The growth changes (size increase: mm) of S. malayensis ephyrae over 20 days: At temperatures of 20°C and 24°C, the growth in the 21PSU salinity environment was significantly lower than in the 24PSU and 27PSU environments (A, B: p < 0.001). However, no significant differences were observed between the 24PSU and 27PSU salinity environments (A: p = 0.461, B: p = 0.840).
Figure 5.
The growth changes (size increase: mm) of S. malayensis ephyrae over 20 days: At temperatures of 20°C and 24°C, the growth in the 21PSU salinity environment was significantly lower than in the 24PSU and 27PSU environments (A, B: p < 0.001). However, no significant differences were observed between the 24PSU and 27PSU salinity environments (A: p = 0.461, B: p = 0.840).
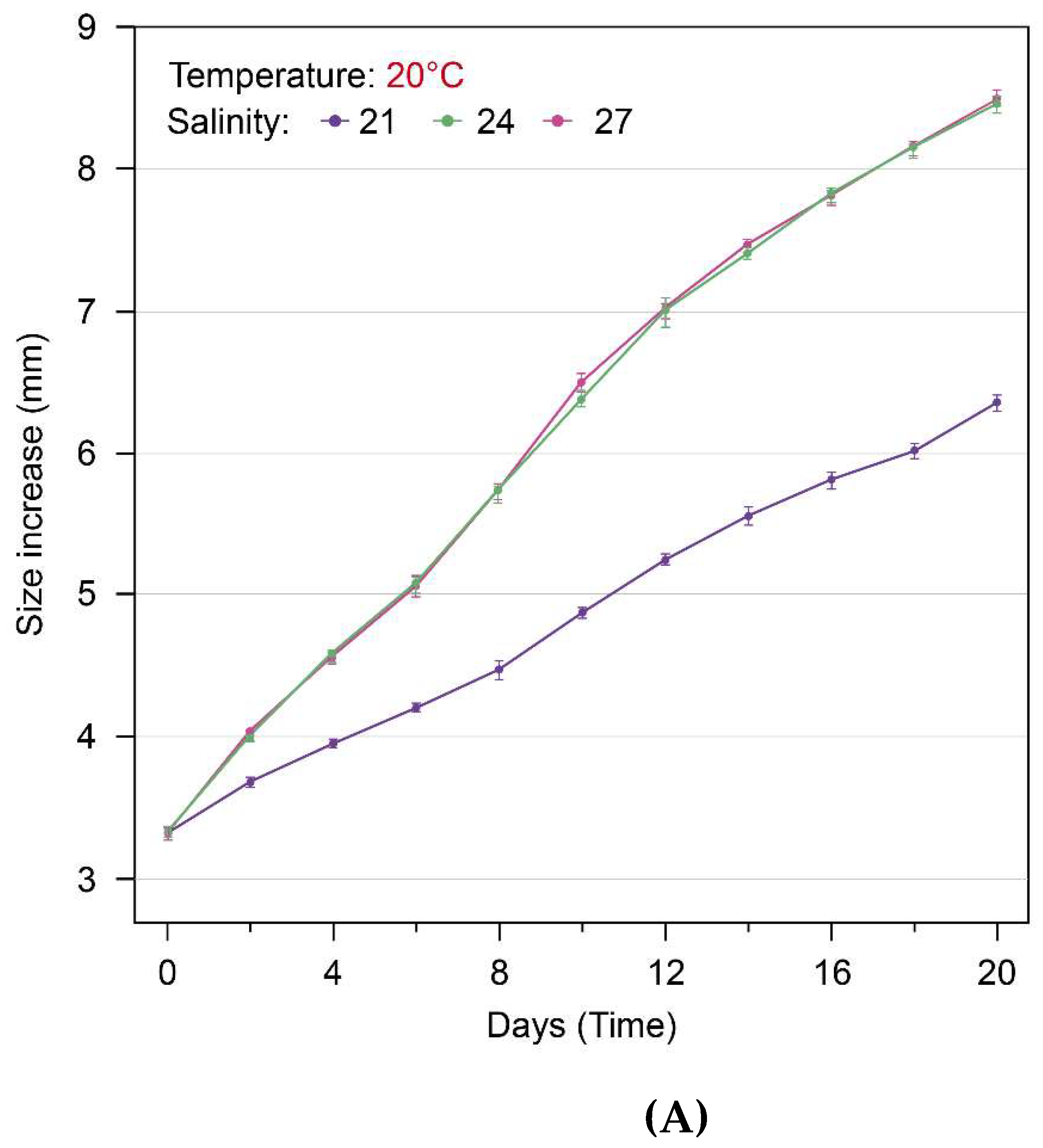
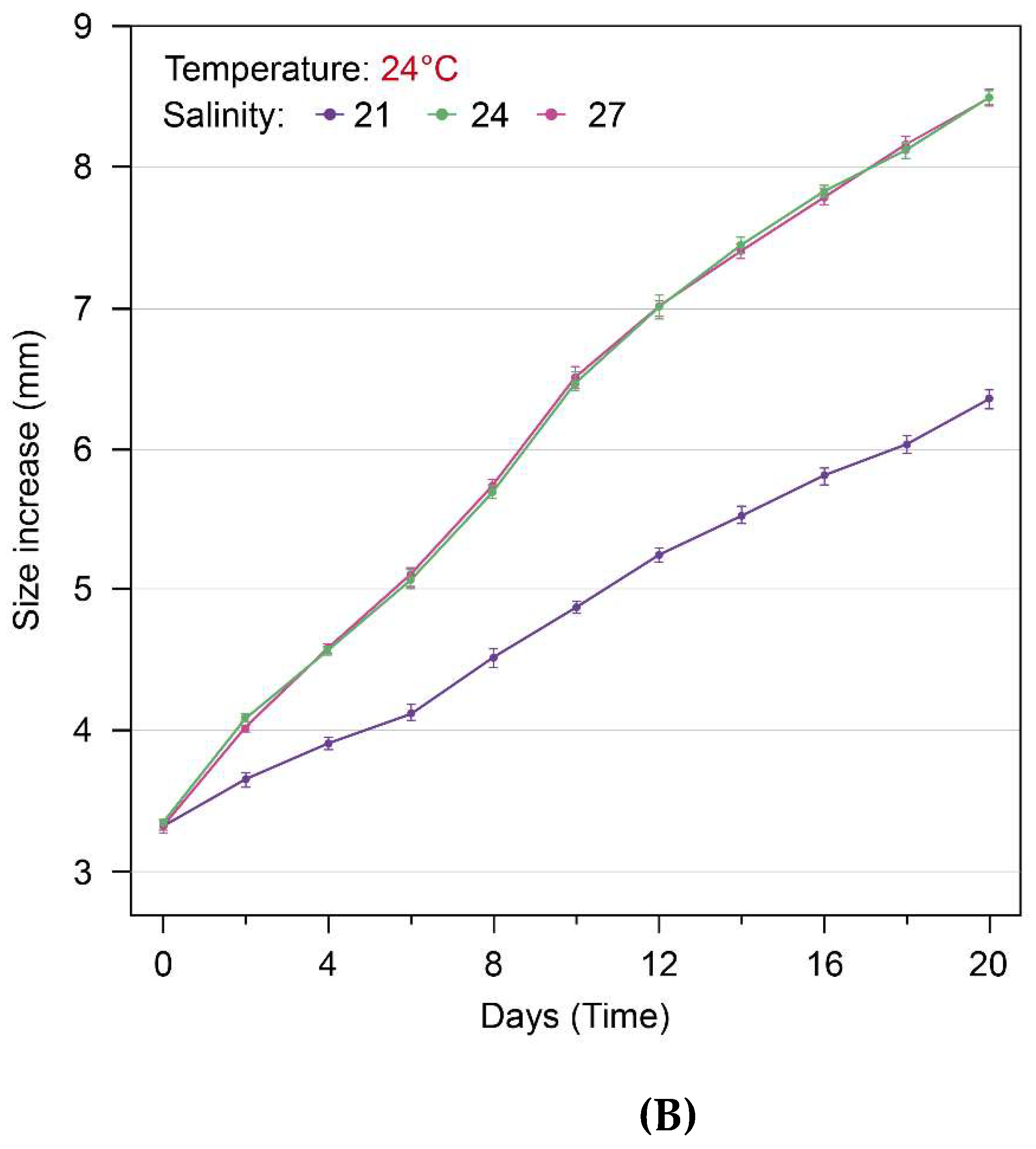
Figure 6.
The feeding rate changes (preys ind-1 h-1) of S. malayensis ephyrae over 20 days: At temperatures of 20°C (Tukey post-hoc test) and 24°C (Holm post-hoc test), the feeding rate in the 21PSU salinity environment was significantly lower compared to the 24PSU and 27PSU environments (A, B: p < 0.001). However, no significant differences were observed between the 24PSU and 27PSU salinity environments (A: p = 0.522, B: p = 0.100).
Figure 6.
The feeding rate changes (preys ind-1 h-1) of S. malayensis ephyrae over 20 days: At temperatures of 20°C (Tukey post-hoc test) and 24°C (Holm post-hoc test), the feeding rate in the 21PSU salinity environment was significantly lower compared to the 24PSU and 27PSU environments (A, B: p < 0.001). However, no significant differences were observed between the 24PSU and 27PSU salinity environments (A: p = 0.522, B: p = 0.100).
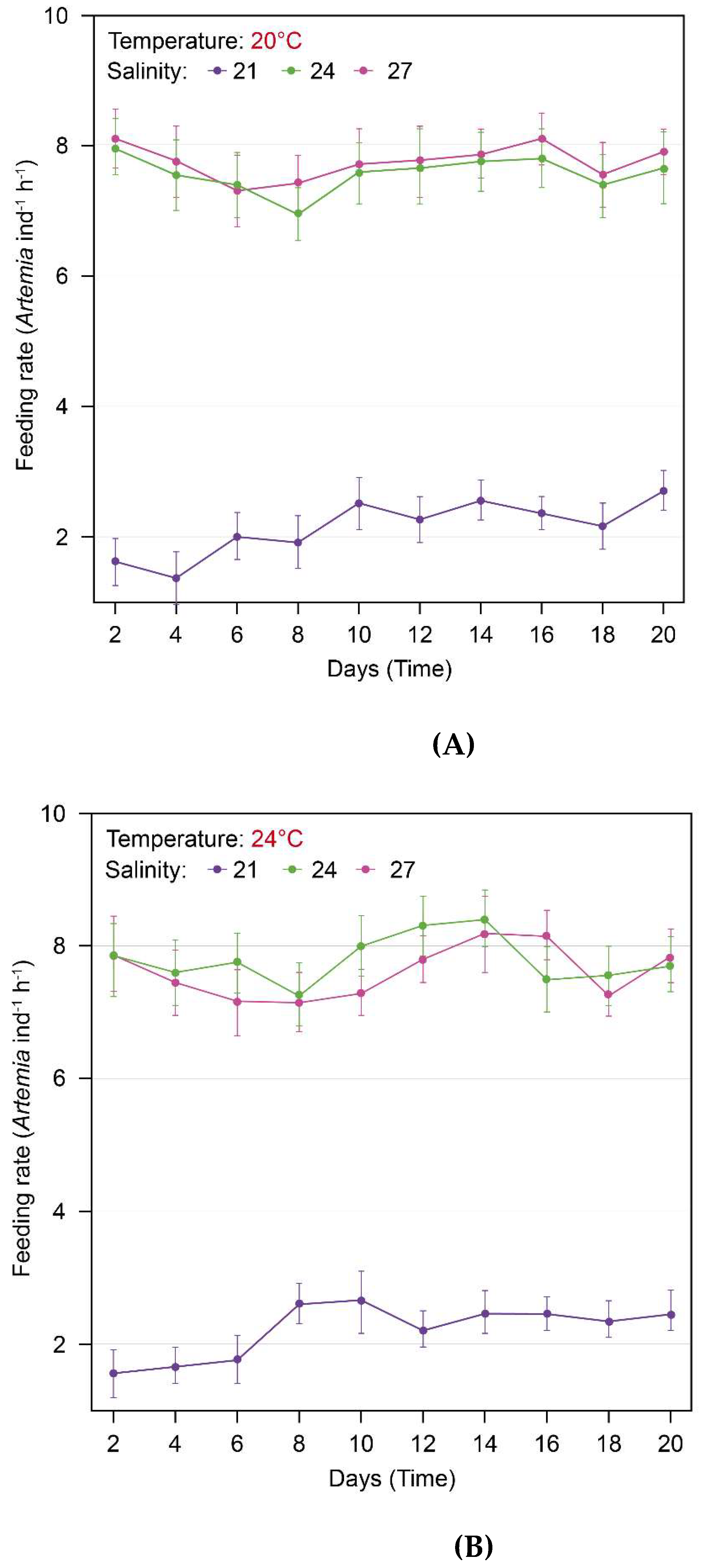
Figure 7.
Kaplan-Meier survival analysis of S. malayensis ephyrae over 20 days: Significant differences in survival curves were observed between the salinity conditions (21PSU, 24PSU, 27PSU) at 20°C and 24°C. (20°C: χ² = 17.746, DF = 2; 24°C: χ² = 28.008, DF = 2, p < 0.001). The effects of the three different salinity conditions were further evaluated using the Cox Hazard Ratio analysis, as shown in Figure S1 and S2.
Figure 7.
Kaplan-Meier survival analysis of S. malayensis ephyrae over 20 days: Significant differences in survival curves were observed between the salinity conditions (21PSU, 24PSU, 27PSU) at 20°C and 24°C. (20°C: χ² = 17.746, DF = 2; 24°C: χ² = 28.008, DF = 2, p < 0.001). The effects of the three different salinity conditions were further evaluated using the Cox Hazard Ratio analysis, as shown in Figure S1 and S2.
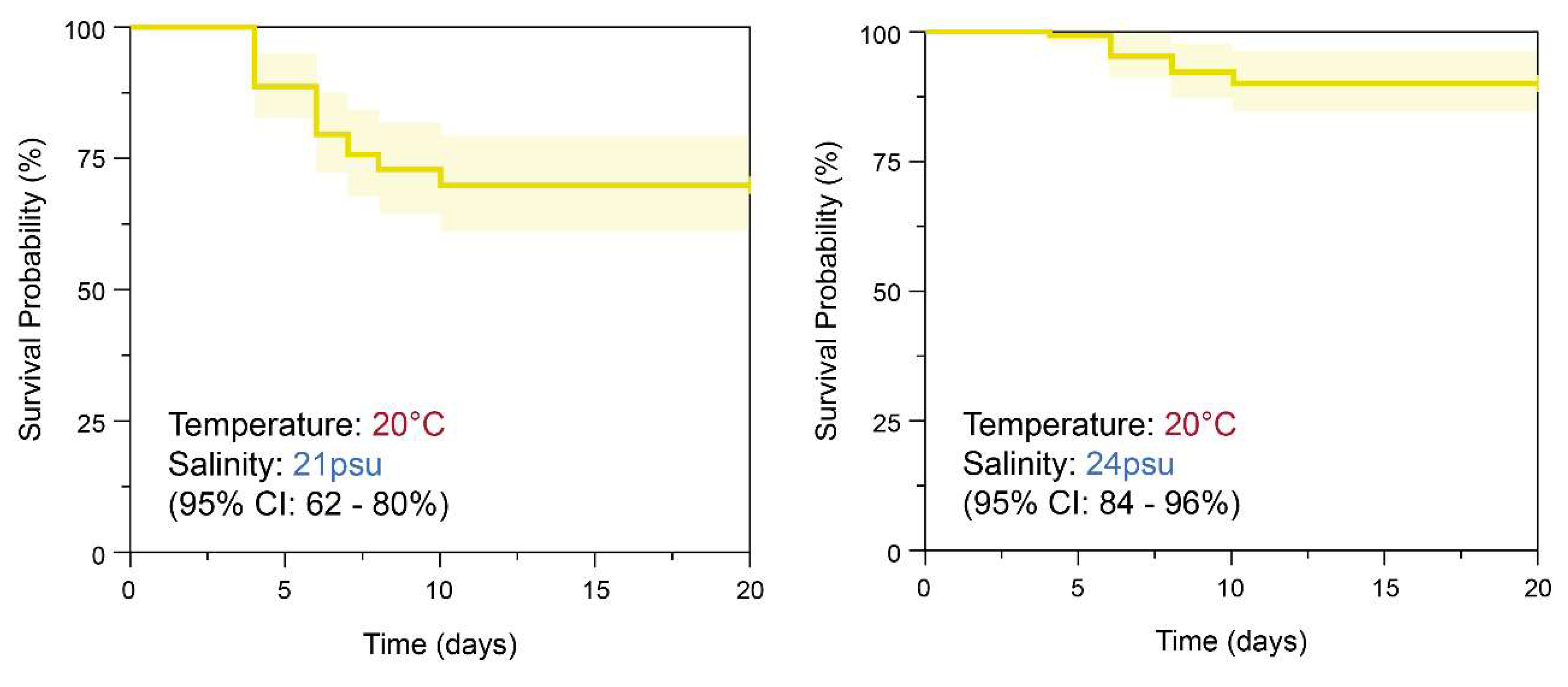
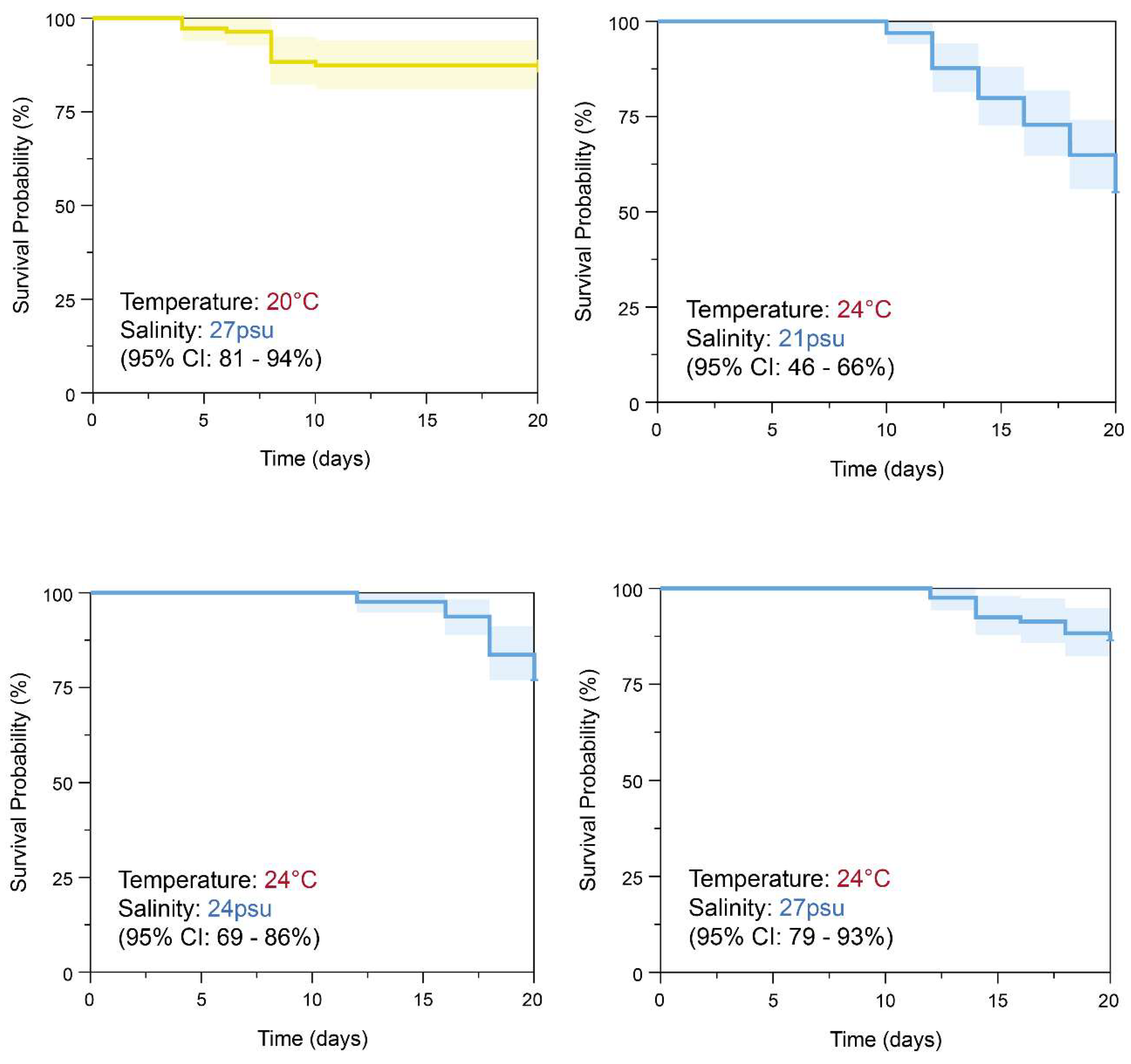

Table 1.
The growth changes (size increase) of S. malayensis ephyrae were measured under two temperature conditions (20°C, 24°C) across three salinity levels (21PSU, 24PSU, 27PSU). Statistical significance was determined using the Holm post-hoc test. Growth at 21PSU was significantly lower than at 24PSU and 27PSU (p < 0.001), while no significant difference in growth was observed between 24PSU and 27PSU (p > 0.05).
Table 1.
The growth changes (size increase) of S. malayensis ephyrae were measured under two temperature conditions (20°C, 24°C) across three salinity levels (21PSU, 24PSU, 27PSU). Statistical significance was determined using the Holm post-hoc test. Growth at 21PSU was significantly lower than at 24PSU and 27PSU (p < 0.001), while no significant difference in growth was observed between 24PSU and 27PSU (p > 0.05).
| Temperature (°C) | Salinity (PSU) |
Size Increase (mm, n = 10) | 95% Confidence interval | |||
|---|---|---|---|---|---|---|
| Mean SD | Lower Limit (CI) |
Upper Limit (CI) |
||||
| 20 | 21 | 6.34a | 0.14 | 6.10 | 6.70 | |
| 24 | 8.47b | 0.15 | 8.20 | 8.70 | ||
| 24 | 27 | 8.51b | 0.13 | 8.30 | 8.70 | |
| 21 24 27 |
6.35a 8.50b 8.50b |
0.14 0.13 0.13 |
6.108.30 8.30 |
6.608.70 8.70 |
||
Table 2.
The growth rates (% d-1) of S. malayensis ephyrae at three salinity levels (21PSU, 24PSU, 27PSU) under two temperature conditions (20°C, 24°C) were measured following the method outlined by Bamstedt et al. (1997). The sample size for each treatment group was n = 10.
Table 2.
The growth rates (% d-1) of S. malayensis ephyrae at three salinity levels (21PSU, 24PSU, 27PSU) under two temperature conditions (20°C, 24°C) were measured following the method outlined by Bamstedt et al. (1997). The sample size for each treatment group was n = 10.
| Temperature (°C) | Salinity (PSU) |
95% Confidence interval | ||||
|---|---|---|---|---|---|---|
| Growth rate (% d-1, n = 10) |
Lower Limit (CI) | Upper Limit (CI) | ||||
| 20 | 21 | 10.31 | 10.19 | 10.71 | ||
| 24 | 14.88 | 14.83 | 14.86 | |||
| 24 | 27 | 14.86 | 14.83 | 15.05 | ||
| 21 24 27 |
10.24 14.80 14.94 |
10.19 14.38 14.38 |
10.47 15.05 15.05 |
|||
Table 3.
The statistical significance of growth changes (size increase) in S. malayensis ephyrae under two temperature conditions (20°C, 24°C) and three salinity levels (21PSU, 24PSU, 27PSU) was assessed using ART-ANOVA. A significant interaction effect between time and salinity was observed (p < 0.001), and the results of the Holm post-hoc analysis are provided in the main text.
Table 3.
The statistical significance of growth changes (size increase) in S. malayensis ephyrae under two temperature conditions (20°C, 24°C) and three salinity levels (21PSU, 24PSU, 27PSU) was assessed using ART-ANOVA. A significant interaction effect between time and salinity was observed (p < 0.001), and the results of the Holm post-hoc analysis are provided in the main text.
| Temperature(°C) Source | Size Increase (mm, n = 10) | |||||
|---|---|---|---|---|---|---|
| Degrees of freedom |
Sum of Squares F-value | p-Value | ||||
| Time | 2 | 15988153.54 633.97 | <0.001 | |||
| 20 Salinity | 10 | 23703043.10 6023.03 | <0.001 | |||
| Salinity × Days | 20 | 20791713.35 234.03 | <0.001 | 9 | <0.001 | |
| Time | 2 | 15974650.91 633.98 | <0.001 | 2 | <0.001 | |
| 24 Salinity | 10 | 23715381.33 6284.18 | <0.001 | 18 | <0.001 | |
| Salinity × Days | 20 | 20437093.72 207.09 | <0.001 | |||
Table 4.
The feeding rate (Preys ind-1 hr-1) results of S. malayensis ephyrae under two temperature conditions (20°C, 24°C) and three salinity levels (21PSU, 24PSU, 27PSU) are presented. Statistical significance was determined using the Tukey post-hoc test at 20°C and the Holm post-hoc test at 24°C. Growth at 21PSU was significantly lower compared to 24PSU and 27PSU (p < 0.001), while no significant difference in growth was observed between 24PSU and 27PSU (p > 0.05).
Table 4.
The feeding rate (Preys ind-1 hr-1) results of S. malayensis ephyrae under two temperature conditions (20°C, 24°C) and three salinity levels (21PSU, 24PSU, 27PSU) are presented. Statistical significance was determined using the Tukey post-hoc test at 20°C and the Holm post-hoc test at 24°C. Growth at 21PSU was significantly lower compared to 24PSU and 27PSU (p < 0.001), while no significant difference in growth was observed between 24PSU and 27PSU (p > 0.05).
| Temperature (°C) | Salinity (PSU) |
Feeding rate (Artemia ind-1 h-1, n = 10) |
95% Confidence interval | |||
|---|---|---|---|---|---|---|
| Mean SD | Lower Limit (CI) |
Upper Limit (CI) |
||||
| 20 | 21 | 2.10a | 0.908 | 0.00 | 4.00 | |
| 24 | 7.58b | 1.138 | 5.00 | 10.00 | ||
| 24 | 27 | 7.69b | 1.165 | 4.00 | 10.00 | |
| 21 24 27 |
2.16a 7.75b 7.64b |
0.869 1.118 1.136 |
0.00 5.00 5.00 |
5.00 10.00 11.00 |
||
Table 5.
The statistical significance of feeding rates of S. malayensis ephyrae at three salinity levels (21PSU, 24PSU, 27PSU) under a temperature condition of 20°C was assessed using ANOVA. No significant interaction effect between time and salinity was observed (p = 0.085), and the results of the Tukey post-hoc analysis are provided in the main text.
Table 5.
The statistical significance of feeding rates of S. malayensis ephyrae at three salinity levels (21PSU, 24PSU, 27PSU) under a temperature condition of 20°C was assessed using ANOVA. No significant interaction effect between time and salinity was observed (p = 0.085), and the results of the Tukey post-hoc analysis are provided in the main text.
| Source | Feeding rate (Artemia ind-1 h-1, n = 10) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sum of squares | Degrees offreedom | Mean ofsquares | F-value | p-Value | ||||||
| Time | 4489 | 2 | 2245 | 2029.04 | <0.001 | |||||
| Salinity | 35 | 10 | 3 | 3.16 | <0.001 | |||||
| Salinity × Days | 33 | 20 | 2 | 1.47 | 0.085 | 9 | <0.001 | |||
Table 6.
The statistical significance of feeding rates of S. malayensis ephyrae at three salinity levels (21PSU, 24PSU, 27PSU) under a temperature condition of 24°C was assessed using ART-ANOVA. A significant interaction effect between time and salinity was observed (p = 0.001), and the results of the Holm post-hoc analysis are provided in the main text.
Table 6.
The statistical significance of feeding rates of S. malayensis ephyrae at three salinity levels (21PSU, 24PSU, 27PSU) under a temperature condition of 24°C was assessed using ART-ANOVA. A significant interaction effect between time and salinity was observed (p = 0.001), and the results of the Holm post-hoc analysis are provided in the main text.
| Source | Feeding rate (Artemia ind-1 h-1, n = 10) | |||||||||
| Degress of freedom | Sum of squares |
F-value | p-Value | |||||||
| Time | 2 | 15983505.68 | 633.18 | <0.001 | ||||||
| Salinity | 10 | 1186144.58 | 3.27 | <0.001 | ||||||
| Salinity × Days | 20 | 1927731.32 | 2.75 | <0.001 | 9 | <0.001 | ||||
Table 7.
The Kaplan-Meier survival analysis results of S. malayensis ephyrae under two temperature conditions (20°C, 24°C) and three salinity levels (21PSU, 24PSU, 27PSU) are presented. The Log-rank test revealed a significant difference in survival curves among the three salinity levels (p < 0.001).
Table 7.
The Kaplan-Meier survival analysis results of S. malayensis ephyrae under two temperature conditions (20°C, 24°C) and three salinity levels (21PSU, 24PSU, 27PSU) are presented. The Log-rank test revealed a significant difference in survival curves among the three salinity levels (p < 0.001).
| Temperature (°C) | Salinity(PSU) | 95% Confidence interval | ||||
|---|---|---|---|---|---|---|
| Survival rate (%) | ObservedEvent | Lower CI | Upper CI | |||
| 20 | 21a | 70 30 | 0.62 | 0.80 | ||
| 24b | 90 10 | 0.84 | 0.96 | |||
| 24 | 27c | 87 13 | 0.81 | 0.94 | ||
| 21a 24b 27c |
55 45 79 21 86 14 |
0.46 0.69 0.79 |
0.66 0.86 0.93 |
|||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



