
Submitted:
05 March 2025
Posted:
07 March 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
An increase in IgG4 levels is typically associated with immunological tolerance states and develops after prolonged exposure to antigens. Accordingly, IgG4 is considered an anti-inflammatory antibody with a limited ability to trigger efficient immune responses. Additionally, IgG4 reduces allergic reactions by blocking immunoglobulin E (IgE) activity. In the case of COVID-19, it has been reported that the repeated administration of some vaccines induces high IgG4 levels. The latest research data has revealed a surprising IgE anti-receptor binding domain response after both natural infection and several SARS-CoV-2 vaccines. The presence of IgG4 and IgE in COVID-19 disease suggests that the virus may induce an “allergic-like” response to evade immune surveillance, leading to a shift from Th1 to Th2, which promotes tolerance to the virus and potentially contributes to chronic infection. The spike protein from vaccines could also induce such a response. Interestingly, "allergen-like" epitopes and IgE responses have been reported for other viruses, such as HIV and respiratory syncytial virus (RSV). The impact of this viral-induced tolerance will be discussed, concerning the protective efficacy of vaccines.
Keywords:
SARS-CoV-2
; HIV
; RSV
; IgG4
; IgE
; COVID-19 vaccines
; immune evasion
1. Introduction
Attempts to rapidly contain the COVID-19 pandemic were limited by a lack of precise understanding of SARS-CoV-2-host interactions, particularly regarding viral biology and host immune response. The overall body of evidence points to the importance of the COVID-19 vaccines in decreasing the severity of sickness, hospitalization, and mortality in virus-infected individuals [1,2,3,4,5]. However, emerging data suggest that repeated vaccination may have unexpected consequences. One of them is the increased production of IgG4 antibodies, a subclass of immunoglobulins traditionally associated with immune regulation and tolerance. IgG4 antibodies are unique among the IgG subclasses due to their ability to undergo Fab-arm exchange (FAE), resulting in bispecific antibodies that are functionally monovalent, and it has lower affinity for antigens when compared with other IgG subclasses [6,7,8,9]. This property allows IgG4 to act as an anti-inflammatory agent by blocking immune complex formation and reducing Fc-mediated effector functions, such as antibody-dependent cellular cytotoxicity (ADCC), antibody-dependent cellular phagocytosis (ADCP), and complement activation [6,10,11]. For example, IgG4 is sometimes referred to as a protective blocking antibody because it can inhibit or stop inflammation in allergy by competing with inflammatory IgE for antigen binding [12,13,14,15]. It is also implicated in certain autoimmune diseases including IgG4-related disease (IgG4-RD), which is characterized by inflammation and fibrosis in various organs [16,17,18].
An increase in IgE production is often associated with natural exposure to a relevant allergen. IgG4 appears to inhibit IgE via various routes. First, through direct competition, IgG4 can prevent IgE from binding to particular allergens in serum. Direct suppression of IgE-mediated mast cell activation is the second and most important mechanism (Figure 1). IgE causes mast cells to degranulate when it interacts with the high-affinity receptor FcϵRI. On the other hand, IgG4 inhibits the aforementioned effects during both the initiation and effector phases of allergic immune responses by binding to the inhibitory receptor FcγRIIb [19].
Clinical improvement has been associated with increases in IgG4 levels, according to data from several studies [20,21]. The protection provided by treatment correlates with increased anti-venom IgG levels in venom allergy, at least at the onset of desensitization [22,23]. IgE-facilitated allergen presentation to T cells, allergen-induced increases in memory IgE synthesis following high allergen exposure during the pollen season, and allergen-induced release of inflammatory mediators from mast cells and basophils appears to be inhibited by IgG4 blocking antibodies. Grass pollen immunotherapy leads to a significant increase in allergen-specific IgG4 antibodies, which are crucial for modulating allergic responses by blocking IgE-facilitated allergen binding to B cells, T cells, mast cells, and basophils [24,25]. IgE, both total and allergen-specific is strongly suppressed by IL-10, which also causes an increase in IgG4 synthesis [26]. Therefore, in addition to producing tolerance in T cells, IL-10 also controls the generation of specific isotypes and skews the specific response from an IgE- to an IgG4-dominated phenotype. During allergen-specific immunotherapy (AIT), there is an initial rise in IgE levels, which then decline, followed by a sustained increase in specific IgG4 levels [27].
While this regulatory role is beneficial in conditions like allergies, it may be less desirable in the context of viral infections, where robust and sustained immune responses are required for protection and viral clearance [28]. In the context of the vaccination campaign against SARS-CoV-2, scientists have demonstrated that the antibody subclasses IgG1 and IgG3 immediately dominated the IgG response following the first two mRNA injections [29]. Nonetheless, an unanticipated long-term side effect has been reported following the second mRNA vaccine injection: a change in the isotype of IgG antibodies towards IgG4 [29]. Several human investigations [29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45] and at least one animal study [46] have now reported the phenomenon of increased IgG4 antibodies following mRNA vaccination. In comparison to all spike-specific IgG antibodies, IgG4 antibodies increased, on average, from 0.04% soon after the second vaccination to 19.27% late after the third [29]. A further study confirmed this, showing that the median level of IgG4 antibodies against the spike protein was 21.2% of total IgG antibodies [34]. On the other hand, this phenomenon has not been documented in unvaccinated individuals [31,32,38,40,43].
Numerous studies have now shown that mRNA-based vaccines (Pfizer/BioNTech or Moderna) and inactivated vaccines such as CoronaVac and BBIBP-CorV cause this response [29,30,31,33,35,38,47,48]; while recipients of adenoviral vector-based or protein-based vaccines did not produce this increase in IgG4 concentrations [30,42]. IgG4 levels slightly increased [30,33,39] or not at all [29,35] when heterologous SARS-CoV-2 vaccines were used (mixing of mRNA vs. adenoviral vector vaccines). It has been proposed that an increase in IgG4 levels may protect against immunological over-activation, similar to what occurs after successful AIT by suppressing IgE-induced effects [29]. In contrast, other authors proposed that such an increase may not be a mechanism of protection, but rather an immunological tolerance mechanism to the spike protein that, by inhibiting natural antiviral responses, may encourage unopposed SARS-CoV2 infection and replication [28]. In this work, we expand on the previous hypothesis and argue that SARS-CoV-2 presents "allergen-like" epitopes, which induce the synthesis of IgE to evade the immune system and generate a chronic infection. Experimental evidence shows that other viruses may also use IgE to suppress immune responses, such as influenza A [49] and RSV [50]. In response, the immune system produces IgG4 antibodies to neutralize the negative effects of IgE. It is also proposed that repeated vaccination with an allergen-like antigen may not be the best option if it results in tolerance rather than protective immunity.
2. Evidence of the Involvement of IgE Antibodies in COVID-19 and in Vaccination
Only a limited number of studies have investigated the role of IgE antibodies and their association with disease severity. One study assessed how changes in inflammatory markers and basal serum total immunoglobulin E (IgE) levels affected the clinical development of COVID-19 patients who were admitted to the hospital. Clinically deteriorated COVID-19 patients had considerably lower serum eosinophil levels (0.015 [0-1.200] vs 0.040 [0-1.360], p=0.002) and significantly higher serum total IgE concentrations (172.90 [0-2124] vs 38.70 [0-912], p<0.001). This study found that high IgE levels could be used as a negative predictor of clinical development in severe COVID-19 [51]. Regarding the role of eosinophils in COVID-19, it has been reported that the disease was less severe in allergic individuals with high blood eosinophil numbers [52]. Low eosinophil counts, on the other hand, are thought to be a predictor of the course of the disease. , Such a reduction was often observed in individuals who passed away from COVID-19 [53]. A study analyzed the medical records of 85 fatal cases of COVID-19 and observed that 81% of the patients had eosinophil numbers below the normal level (absolute eosinophil counts <0.02 × 109 cells/L) at the time of admission [54]. During respiratory viral infections, eosinophils are known to have a role in the immune response that facilitates cytokine release and the recruitment of CD8 + T cells. Additionally, the virus can be neutralized by enzymes produced by eosinophils [55].
Another study found that IgE antibodies produced against the SARS-CoV-2 N protein were detected in 100 % of patients with severe disease, 92.6 % with moderate disease, and 66.7% with mild disease [56]. Furthermore, a recent study discovered an unusual IgE anti-receptor binding domain response after spontaneous infection and after receiving different SARS-CoV-2 vaccines [45]. Following COVID-19 infection, samples from 59 subjects were evaluated after primary vaccination with vectored (ChAdOx1), inactivated (CoronaVac) vaccines, and booster vaccination with mRNA (BNT162b2) vaccine. IgE was naturally induced by COVID-19, but immunization raised its levels. All groups showed comparable IgE levels following boosting with BNT162b2, but those who received two doses of ChAdOx1 showed a more robust response than those who received two doses of CoronaVac. Particularly following the booster vaccination, IgE exhibited moderate to high avidity. Additionally, they detected IgG4 antibodies, primarily following the booster, and these showed a moderate correlation with IgE [45]. Th2 cytokines facilitate class-switching to IgG4 and IgE antibodies, which are found in the upstream area of the Ig locus, and B cells require sustained antigenic stimulation for this to occur [57,58].
Although it is well known that aluminum hydroxide creates a potent Th2 milieu that facilitates this response [59], ChAdOx1, which does not contain aluminum, produced significantly greater IgE and IgG4 levels than CoronaVac, an alum-adjuvanted vaccine. The researchers suggested that the active immunogen ( the spike protein) triggers such IgE and IgG4 responses [45]. When IgE-coated antigen binds to mast cell FcεRI receptors, the IgE-FcεRI complex is cross-linked, which activates mast cells and releases histamine along with several pro-inflammatory chemokines and cytokines, such as IL-6, IL-1β, and TNF-α [60,61]. Mast cell activation syndrome may be connected to the emergence of the cytokine storm and hyper-inflammation, according to several investigations that have demonstrated that mast cells are activated in patients with severe COVID-19 [61,62,63]. Another study supported the use of IgE-blocking drugs such as omalizumab for the treatment of COVID-19 by indicating a link between mast cell activation and high antibody levels in severe COVID-19 cases [64].
An inevitable question arises: what purpose would it be for a virus to have an allergenic epitope? This is highly unusual in a viral infection. It is supposed that, upon contact with a virus, the immune system would produce only IgG1 and IgG3, which are the main antiviral antibodies [65,66,67,68]. A plausible explanation is that the virus possesses an epitope that triggers an allergic response to evade the immune system and establish a chronic infection. Though infrequent, viral protein antigens can elicit IgE responses, as seen following HIV [69,70,71,72,73,74,75,76,77,78], respiratory syncytial virus [79,80,81], and varicella zoster infections [82], as well as with hepatitis B [83] and influenza vaccinations [84]. In COVID-19, two investigations have described the presence of IgE antibodies after a spontaneous infection with SARS-CoV-2. Seric IgE was reported to correlate with the severity of COVID-19 infection, which likely contributed to hyper-inflammation [51,56]. Interestingly, other studies discovered that lethal COVID-19 was associated with high IgG4 levels [41,85,86,87]. It is hypothesized that elevated IgG4 levels were a consequence, rather than a cause, of death. The etiology of death (among other factors) may be attributable to a severe IgE-mediated allergic reaction directed against the N and spike proteins. It is conceivable that the immune system generated the production of IgG4s in an attempt to minimize the cytokine storm. Indeed, it has been reported that IgG4 competes with specific IgE for allergen binding as a "blocking antibody," which prevents mast cell and basophil degranulation [88,89]. Another possibility is that in severe COVID-19 pneumonia, SARS-CoV-2 induces IgG4 synthesis in an IgE-independent manner, inhibiting IgG3 binding to its Fc receptor, thus impairing viral phagocytosis [90]. According to a recent study, the fatal outcome of severe COVID-19 was correlated to significantly increased IgG4 levels in the lung tissue. IgG4 expression peaked at 80-90% (Figure 2) in acute stages, whereas in the organization and fibrosis stages decreased to 5-10% [87]. It would be interesting to investigate the presence of IgE antibodies in lung samples from patients who died of severe pneumonia and who also showed high levels of IgG4. This would help determine whether death resulted from elevated IgE levels and whether the increase in IgG4 was an attempt by the immune system to minimize IgE effects or if IgG4 independently contributed to inhibiting the phagocytosis of virus-infected cells.
2.1. Does SARS-CoV-2 Possess “Allergen-Like” Epitopes?
To date, several works have investigated this interesting topic. Heterologous immunity mediated by virus-induced T cells has been shown to either promote immunopathology or provide protection against different antigens [91,92,93,94]. The shared characteristics between the SARS-CoV-2 proteome and some aeroallergens may offer a protective benefit to individuals sensitized to such allergens, potentially mitigating the risk of an excessive Th1 response and the subsequent cytokine storm. In contrast, these heterologous immune responses may have a negative impact by weakening the antiviral response. Due to reduced CD8+ responses, a Th2 immune skew may result in insufficient viral clearance [91].
This can be exemplified by findings from a large population-based cohort investigation which discovered that adults with non-allergic asthma had a higher risk of severe COVID-19. In contrast, the risk of severe COVID-19 was not found to be significantly increased in patients with allergic asthma [95]. From the immunological perspective, the differential risk of severe COVID-19 in individuals with non-allergic versus allergic asthma can be attributed to the distinct immune pathways underlying these conditions [95,96,97].
Other “allergen-like” epitopes have been reported to be present in SARS-CoV-2. In 2020, researchers discovered four insertions in the spike glycoprotein (S) that are unique to SARS-CoV-2 and do not occur in other coronaviruses. Crucially, all four inserts included amino acid residues similar to those found in HIV-1 gp120 or Gag proteins. The scientists hypothesized that it was improbable that these four different inserts in SARS-CoV-2 were accidental, suggesting an artificial origin [98]. This conclusion led to the retraction of the paper. Alternatively, these four inserts could be acquired naturally. It is widely recognized that viruses can acquire new genes through genetic recombination [99,100,101]. It is proposed that the alleged presence of such inserts should be rigorously investigated. In the event of confirmation, this could provide a rationale for the presence of IgE and IgG4 antibodies in response to SARS-CoV-2 infection.
Many viruses have developed sophisticated immune evasion techniques, and SARS-CoV-2 is no exception [102]. In this work, we hypothesize that during its evolution, this virus has acquired two epitopes (the N and the Spike protein) that, acting as allergens, induce the production of IgE antibodies to impair antiviral defenses. In respect of the Spike protein, a galectin-3 (Gal-3)-like fold has been identified within the N-terminal domain of SARS-CoV-2[103]. Gal-3 is a β-galactoside-binding lectin which is associated with fibrosis and conditions such as idiopathic pulmonary fibrosis (IPF)[104]. Gal-3 is a low-affinity receptor for IgE, and it has been theorized that both Gal-3 and the Gal-3-like fold within the NTD act as allergens by binding to IgE on the FcεRI or other related mast cell receptors [105,106]. In response, the immune system promotes IgG4 synthesis, trying to block the negative effects of high IgE levels in a manner similar to AIT. A significant correlation between IgG4 anti-Gal-3 autoantibodies and serum Gal-3 levels has been found in patients with IgG4-RD [107]. This, and other research, supports the hypothesis that multiple allergen-like epitopes on SARS-CoV-2 are implicated [108,109]. Since allergies are associated with a switch from Th1 to Th2 response, and Th2 induces immune tolerance [110], it is proposed that SARS-CoV-2 induces a Th2 response to promote tolerance. Such a response would be evolutionarily advantageous for the virus since the immune system would ignore it instead of attacking it, thus allowing the establishment of chronic infection. Considering there is not much information about the mechanism that mediates the induction of IgE antibodies by HIV and RSV viruses, the following section will describe how they have evolved an immune evasion mechanism through the presence of allergen-like epitopes.
3. Allergen-Like Epitopes in HIV Envelope Proteins
It is well known that no effective vaccine has been produced against HIV. In 2004, it was suggested that the steady rise in interleukin 4 (IL-4) and IgE levels during HIV-1 infection, as well as the stimulation of T helper 2 (Th2) cytokine generation, could be an allergic reaction to HIV-1 proteins [111]. For example, one study found that the HIV-1 gp41 heptad repeat 2 (HR2) protein contains an amino acid domain similar to the allergen domain in the fungus Aspergillus fumigatus Asp f1 protein [112]. The same author expanded on this theory in a later review, citing experimental evidence that show gp120 proteins released by HIV-1 resemble allergens, which bind to IgE molecules attached to Fc epsilon receptor I (FcεRI) cells (mast cells, basophils, monocytes, and dendritic cells (DCs)) and cause them to produce and release IL-4. This cytokine was shown to induce the following responses: the inactivation of T helper 1 (Th1) cells, the inhibition of the antiviral cytotoxic T cell (CTL) response, the synthesis of IgE by B cells and the inhibition of antiviral IgG3 synthesis [111]. It was discovered that whereas CD4+ Th1 cells did not effectively support HIV replication, CD4+ Th2 and the majority of Th0 cells did. These findings indicated that HIV replication preferentially occurs in T cells that produce Th2-type cytokines [113], given that interferon-gamma (IFN-γ) has been demonstrated to inhibit HIV infection of Th1 cells [114].
This is highly relevant since the Th1 and Th2 pathways are mutually inhibitory: IFN-γ restricts TH2 cell growth, while IL-4 and IL-10 preferentially promote TH2 cell growth and inactivate Th1 cells [115,116]. These results imply that HIV-1 attacks the human immune system in two ways: first, it infects Th2 cells, macrophages, and DCs, where it then replicates, and second, the gp120 proteins it sheds trigger an allergic reaction. As a result, this “allergic-like” context inhibits Th1 cell differentiation, cytokine release, and antiviral cytotoxic T cells that are essential to destroy virus-infected cells [117]. The theory that HIV-induced AIDS is an allergy and that gp120 is the allergen was proposed. Thus, it can be concluded that the ongoing replication of HIV-1 in infected individuals continuously exposes the immune system to an increase in allergen concentration until immune tolerance is achieved, ultimately compromising host immunity [117]. At the time of its formulation, this theory was probably considered to have no logical basis. However, considering the recent discoveries referenced in this review, there is a strong probability that the hypothesis is accurate, although the statement that AIDS is an allergy is an oversimplification. In our opinion, such a hypothesis can be reformulated as follows: the induction of a Th1 to Th2 switch during HIV infection mimics the human immune system´s response to environmental allergens. When the immune system determines that an allergen is not a pathogen, but is regarded as an innocuous particle, it develops a tolerance response to ignore it. Such a response includes IgG4 antibody synthesis to block IgE proinflammatory effects. It is probable that HIV, through convergent evolution, has developed a similar mechanism. Via the gp120 protein (which has allergen-like epitopes), HIV tricks the immune system into inducing a tolerance response (Th2) instead of an antiviral response (Th1).
Several works have evaluated IgE levels and their association with disease severity. A study revealed that in the initial phases of HIV-1 infection, there is an increase in plasma IgE levels before a decrease in CD4+ cell levels [73]. Furthermore, it is noteworthy that the degree of immunodeficiency (as determined by the CD4+ cell counts) was significantly correlated with the IgE concentration, that is, the higher the concentration of IgE antibodies, the lower the CD4+ count [76]. It was suggested that IgE overproduction during HIV infection may be associated with CD4+ cell depletion and their decreased ability to produce IFN [76].
In a particular study, the IgE levels of 315 HIV-seropositive patients, with or without AIDS, were measured, and the results were then compared to those of 100 HIV-seronegative participants. The research found that HIV-infected patients had higher IgE levels compared to the seronegative control subjects (p < 0.05). This difference was particularly pronounced between the AIDS patients and the control volunteers (p < 0.005). Furthermore, survival analysis revealed a 24-month incidence rate of AIDS of 83% in subjects with IgE>150 KIU/L versus 44% in those with IgE<150 (p=0.016). Researchers also evaluated the prognostic value of IgE elevation during the progression of the illness. The analysis revealed that individuals with IgE levels greater than 150 had a 100% risk of developing AIDS, while those with IgE levels less than 150 had a 9% risk (p=0.003). These findings suggest that IgE levels may serve as a highly selective indicator among patients in the later stages of HIV infection [76]. In another work, the total blood IgE levels of 18 uninfected homosexual male participants, 27 uninfected heterosexual male subjects, and 67 infected male subjects were examined. The analysis revealed that participants with a helper T-lymphocyte number ≤200/mm3 exhibited a mean IgE level of 132 IU/ml, which was significantly higher than the mean IgE levels observed in the uninfected heterosexual (38 IU/ml) and homosexual (35 IU/ml) groups. Furthermore, the study identified a negative correlation between IgE levels, helper T cells, and suppressor/cytotoxic T cell counts. However, these correlations were not observed for IgG or IgA levels [69]. In research involving children, persistent elevation of IgE was found to be significantly associated with a substantial decrease in CD4+ counts (≥30% over one year) and an increased susceptibility to bacterial infections. A considerable decrease in CD4+ cell count appears to be associated with protracted IgE hyperproduction, suggesting that this clinical test (as in adults) serves as a valuable indicator of the progression of the disease [118].
4. The Role of IgE in RSV Infection
It is well known that reinfection is common and immunity to the respiratory syncytial virus is not optimal. Relative protection is associated with RSV-specific IgG, IgM, and IgA antibodies. One study showed an association between wheezing and hypoxia in RSV infection and histamine and RSV-specific IgE levels in nasopharyngeal secretions [81]. There is experimental evidence that allergic inflammation can interfere with antiviral responses [119,120,121,122], and observational investigations have shown strong correlations between allergy sensitization and the incidence of acute virus-induced wheezing [123,124,125]. The mechanisms by which IgE antibodies against RSV modulate the inhibition of the immune response have been described. Uncommon blood immune cells known as plasmacytoid dendritic cells (pDCs) have been shown to release significant amounts of type 1 IFN in response to exposure to virus-derived nucleic acids that are identified by toll-like receptors (TLRs) 7 and 9 [126].
pDCs generate significant levels of type I IFN, primarily IFN-α and IFN-β. They are essential for producing antiviral molecules, up to 1,000 times more than any other white blood cell type [127]. Purified pDCs were exposed to influenza viruses, and IFN-α production was measured to assess their function in asthma patients and controls. Flow cytometry was used to assess the expression of FcεRIα on pDCs in blood samples from both controls and patients with allergic asthma. When exposed to the influenza A virus, pDCs from asthmatic patients secreted significantly less IFN-α (572 versus 2815; p=0.03), and secretion was inversely associated with blood IgE levels. These findings demonstrated that IgE binding reduces pDC antiviral responses [119]. Notably, anti-IgE therapy reduces viral exacerbations in asthma by downregulating the high-affinity IgE receptor (FcεRI) on pDCs, thus improving antiviral immunity [128,129,130].
Another study found that, in contact with an allergen, IL-33 reduced the expression of the genes for viperin and interferon regulatory factor 7 (IRF7) and quickly reduced the expression of IL-1 receptor-associated kinase 1 in pDCs both in vivo and in vitro. This resulted in reduced IFN-α production and hypo-responsiveness of TLR7 [121]. All these data suggest that by shifting the balance of Th1/Th2 cytokines towards elevated levels of Th2 cytokines and IgE (which are indicators of allergy), RSV avoids the human adaptive immune system [80].
5. Repeated Vaccination with Allergen-Like Epitopes
The presence of “allergen-like” epitopes in some viruses raises an intriguing possibility: they may use such epitopes to induce immune tolerance, allowing them to persist in the host for a long time. The immunological effects of repeated immunization with viral antigens that share structural similarities to allergens require thorough investigation. In certain situations, vaccine-induced immunity may shift from a strong protective immunity to a state resembling immunological tolerance. Allergenic viral epitopes may preferentially stimulate a Th2-skewed immune response, which results in the generation of IgE and IgG4 antibodies. For strong antiviral immunity, a Th1 response—which is defined by the generation of IFN-γ and the activation of cytotoxic T cells—is usually necessary.
A shift toward a Th2 response, promoting immune tolerance, could explain why HIV and RSV vaccines have failed to prevent infection and disease progression. It is imperative to note that the consequences of immune tolerance induction exhibit variations depending on the specific pathogen. In the context of SARS-CoV-2, it has been documented that emerging variants exhibit reduced pathogenicity compared to the original lineage [131,132,133,134]. However, in the case of HIV, no comparable reduction in pathogenicity has been observed, as the virus continues to evade immune control and drives disease progression [135].
According to a recent study, delayed disease progression was attributed to low IgG2 and IgG4 levels and high levels of HIV-1-specific IgG1 three years after seroconversion. In particular, there was a significant correlation between delayed disease progression and the ability to interact with all Fcγ receptors (FcγRs) and complement component C1q, specifically with FcγRIIa [136]. Since AIDS progression is also associated with high IgE levels [76,118], the combined elevated presence of IgE and IgG4 antibodies supports the hypothesis that “allergen-like” proteins in the HIV envelope induce a state of tolerance to establish chronic infection. A study was conducted to compare the efficacy of repeated immunization with similar HIV vaccines in a trial scenario. The protection (31.2%) provided by one vaccine (RV144) was attributed to the production of IgG1 and IgG3 antibodies, whereas the protection achieved by the other vaccine (VAX003) was found to be negligible and was linked to the production of IgG4 antibodies after multiple vaccinations [137]. Because VAX003 was associated with higher levels of IgG4, which are typically associated with lower functionality, scientists sought to determine whether the induction of these antibodies was simply associated with a poorly coordinated functional response or if it contributed directly to it. To determine that, IgG4 antibodies were selectively removed from 16 samples from the two vaccine trials (VAX003 and RV144) to assess their impact on effector functions [137].
In VAX003, the removal of IgG4 antibodies resulted in a significant increase in antibody-dependent cell phagocytosis (ADCP), suggesting that IgG4 may competitively inhibit Fcγ receptor-mediated phagocytosis. Additionally, a tendency toward increased antibody-dependent cell cytotoxicity (ADCC) was discovered, indicating that IgG4 may dampen natural killer cell-mediated cytotoxicity. In RV144, no significant difference was observed between total and IgG4-depleted samples. This is likely due to the inherently low levels of IgG4 in RV144-induced immune responses [137]. Regarding the relevance of the elevated levels of IgG4 antibodies detected in the failed HIV trials, it has been suggested that the shift in the IgG subclass from fully efficacious antibodies (IgG3) to IgG4 would be a significant barrier to the effectiveness of HIV vaccines [138]. Indeed, in the RV144 trial, high IgG3 levels were associated with a lower risk of infection [139], while IgG4 production was directly associated with reduced efficacy in protecting people from getting HIV infection. Moreover, it favored breakthrough infections [137].
The induction of tolerance to a virus and enhancement by repeated vaccination is not recommended, as this may result in the immune system ignoring the pathogen, thereby facilitating reinfection and unopposed replication [28]. Indeed, in COVID-19, several studies demonstrated that the more vaccines a person receives, the greater the likelihood of acquiring SARS-CoV-2 [140,141,142,143,144,145]. Depletion assays were used in a study with convalescent participants and results showed that IgG4 had little or no impact on SARS-CoV-2 neutralization [146], thus confirming similar results from other studies [67,147]. Other research evaluated IgG4 levels in healthy and immunocompromised patients who received three mRNA vaccines, showing they peaked suddenly, remained elevated for a long time, and had little effect on neutralizing capacity [40]. Furthermore, a decline in the capacity of spike-specific antibodies to stimulate ADCP and complement deposition was also observed, coinciding with a class switch to IgG4 antibodies [29]. Although the virus is now considered less dangerous, it can still lead to chronic inflammation and severe damage or death in adults with comorbidities and immunosuppressed individuals [148].
6. Implications for Vaccine Design
A new pandemic virus is likely to emerge in the future. In such a scenario, it is suggested that virologists should analyze its genetic sequence to find allergen-like epitopes. Such identification requires a combination of bioinformatics, structural analysis, and immunological validation. Some useful tools are:
- AlgPred 2.0 (https://webs.iiitd.edu.in/raghava/algpred2/), which uses multiple machine learning approaches to predict allergen-like sequences.
- AllergenFP (http://ddg-pharmfac.net/AllergenFP/), uses a descriptor-based fingerprint to detect potential allergens.
- BLASTp (https://blast.ncbi.nlm.nih.gov/Blast.cgi), finds regions of similarity between biological sequences. The program compares nucleotide or protein sequences to the sequence database and calculates the statistical significance.
- AlphaFold (https://swissmodel.expasy.org/), predicts protein structures for docking studies.
- Molecular docking (Autodock, HADDOCK) (https://rascar.science.uu.nl/haddock2.4/),
If such epitopes are identified using these tools, they should be excluded from vaccines. A gene-editing technique known as CRISPR-Cas9 [149] could be used to delete or modify an “allergen-like” epitope on a virus or bacterium to prevent the shift to a Th2 response and redirect toward a Th1 response.
7. Conclusion
A comprehensive review of the existing literature suggests that certain viruses, such as HIV and RSV, possess “allergen-like” epitopes. These viruses induce an allergic response that counteracts antiviral defenses and promotes immunological tolerance, allowing them to persist in the host. Such tolerance may be a key factor contributing to the failure of vaccines targeting these viruses, as it leads to the induction of a Th2 rather than a Th1 response. Several HIV vaccine trials showed no protection against infection, and that was associated with high IgG4 levels. In addition, vaccines against pertussis and malaria have also been shown to increase IgG4 production beyond normal levels (for review, see ref. [28]). For instance, research demonstrated that a decreased risk of malaria in the second year was associated with higher concentrations of IgG1 and IgG3 antibodies. The probability of contracting malaria was reduced by almost 50% when IgG1 levels doubled and by roughly 60% when IgG3 levels doubled. In stark contrast, when levels of non-neutralizing IgG4 doubled, the likelihood of getting malaria increased by about three times [150]. Interestingly, a different study also discovered a connection between elevated IgG4 levels and an increased risk of infection and malaria exacerbations [151]. These findings strongly suggest that pertussis and malaria pathogens also possess “allergen-like” epitopes.
Additionally, it has been shown that IgG4 inhibits IgG1 and IgG3 from opsonizing malaria-infected erythrocytes in vitro [152]. Opsonization is an immunological process in which invading pathogens are tagged with opsonins (antibodies) so that phagocytes can eliminate them. The negatively charged cell walls of the pathogen and the phagocyte repel each other in the absence of antibodies. Consequently, the infection can continue to grow in the human body without being destroyed. Opsonins help the macrophage to ingest the pathogen by overcoming the repulsive barrier between negatively charged cell walls. An antimicrobial method to eradicate and stop the transmission of the disease is opsonization [153].
The presence of “allergen-like “epitopes in SARS-CoV-2 is suggested, as evidenced by the induction of IgE and IgG4 responses by the N and spike proteins. Consequently, it is imperative to verify or refute their existence. If this hypothesis is confirmed, it would suggest that the virus has developed an additional immune evasion mechanism by mimicking an allergen. Therefore, repeated vaccination could unintentionally promote unopposed reinfection and replication, allowing chronic infection [28]. In the context of SARS-CoV-2 vaccines, repeated mRNA vaccination also leads to the production of high levels of IgG4 antibodies, which are likely to be generated to counteract the effects of IgE antibodies induced by the N or the spike protein. However, as noted above, the induction of tolerance to an allergen is beneficial for the host, whereas, in a viral infection, tolerance induction will allow the persistence of the pathogen rather than effective clearance.
Author Contributions
Conceptualization, A.R.-C., V.N.U., and E.M.R.; formal analysis, V.N.U., A.R.-C., and E.M.R.; investigation, E.M.R., D.C., V.N.U., M.P., C.B., and A.R.-C.; data curation, E.M.R., D.C., V.N.U., C.B., and A.R.-C.; writing—original draft preparation, V.N.U., A.R.-C., and E.M.R.; writing—review and editing, E.M.R., D.C., V.N.U., M.P., C.B. and A.R.-C.; visualization, A.R.-C., V.N.U., supervision, V.N.U. and E.M.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- DeSilva, M.B.; Mitchell, P.K.; Klein, N.P.; Dixon, B.E.; Tenforde, M.W.; Thompson, M.G.; Naleway, A.L.; Grannis, S.J.; Ong, T.C.; Natarajan, K. Protection of two and three mRNA vaccine doses against severe outcomes among adults hospitalized with COVID-19—Vision Network, August 2021 to March 2022. The Journal of infectious diseases 2023, 227, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Tenforde, M.W.; Self, W.H.; Adams, K.; Gaglani, M.; Ginde, A.A.; McNeal, T.; Ghamande, S.; Douin, D.J.; Talbot, H.K.; Casey, J.D. Association between mRNA vaccination and COVID-19 hospitalization and disease severity. Jama 2021, 326, 2043–2054. [Google Scholar] [CrossRef] [PubMed]
- Lauring, A.S.; Tenforde, M.W.; Chappell, J.D.; Gaglani, M.; Ginde, A.A.; McNeal, T.; Ghamande, S.; Douin, D.J.; Talbot, H.K.; Casey, J.D. Clinical severity of, and effectiveness of mRNA vaccines against, covid-19 from omicron, delta, and alpha SARS-CoV-2 variants in the United States: prospective observational study. bmj 2022, 376. [Google Scholar] [CrossRef] [PubMed]
- Tenforde, M.W.; Self, W.H.; Zhu, Y.; Naioti, E.A.; Gaglani, M.; Ginde, A.A.; Jensen, K.; Talbot, H.K.; Casey, J.D.; Mohr, N.M. Protection of mRNA vaccines against hospitalized COVID-19 in adults over the first year following authorization in the United States. Clinical Infectious Diseases: An Official Publication of the Infectious Diseases Society of America 2022, ciac381.
- Sibanda, B.; Haryanto, B. Assessing the Impact of COVID-19 Vaccination Programs on the Reduction of COVID-19 Cases: A Systematic Literature Review. Annals of Global Health 2024, 90, 45. [Google Scholar] [CrossRef] [PubMed]
- Rispens, T.; Huijbers, M.G. The unique properties of IgG4 and its roles in health and disease. Nature Reviews Immunology 2023, 23, 763–778. [Google Scholar] [CrossRef] [PubMed]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG subclasses and allotypes: from structure to effector functions. Frontiers in immunology 2014, 5, 520. [Google Scholar] [CrossRef]
- Rispens, T.; Ooijevaar-de Heer, P.; Bende, O.; Aalberse, R.C. Mechanism of immunoglobulin G4 Fab-arm exchange. Journal of the American Chemical Society 2011, 133, 10302–10311. [Google Scholar] [CrossRef]
- Aalberse, R.; Stapel, S.; Schuurman, J.; Rispens, T. Immunoglobulin G4: an odd antibody. Clinical & Experimental Allergy 2009, 39, 469–477. [Google Scholar]
- Schuurman, J.; Van Ree, R.; Perdok, G.a.; Van Doorn, H.; Tan, K.; Aalberse, R. Normal human immunoglobulin G4 is bispecific: it has two different antigen-combining sites. Immunology 1999, 97, 693–698. [Google Scholar] [CrossRef]
- Van Der Neut Kolfschoten, M.; Schuurman, J.; Losen, M.; Bleeker, W.K.; Martínez-Martínez, P.; Vermeulen, E.; Den Bleker, T.H.; Wiegman, L.; Vink, T.; Aarden, L.A. Anti-inflammatory activity of human IgG4 antibodies by dynamic Fab arm exchange. Science 2007, 317, 1554–1557. [Google Scholar]
- Akdis, C.; Blaser, K. Mechanisms of allergen-specific immunotherapy. Allergy 2000, 55. [Google Scholar] [CrossRef] [PubMed]
- Akdis, M. Healthy immune response to allergens: T regulatory cells and more. Current opinion in immunology 2006, 18, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Larché, M.; Akdis, C.A.; Valenta, R. Immunological mechanisms of allergen-specific immunotherapy. Nature Reviews Immunology 2006, 6, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Durham, S.R.; Walker, S.M.; Varga, E.-M.; Jacobson, M.R.; O'Brien, F.; Noble, W.; Till, S.J.; Hamid, Q.A.; Nouri-Aria, K.T. Long-term clinical efficacy of grass-pollen immunotherapy. New England Journal of Medicine 1999, 341, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Khosroshahi, A.; Stone, J.H. A clinical overview of IgG4-related systemic disease. Current opinion in rheumatology 2011, 23, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Della-Torre, E.; Lanzillotta, M.a.; Doglioni, C. Immunology of IgG4-related disease. Clinical & Experimental Immunology 2015, 181, 191–206. [Google Scholar]
- Shiokawa, M.; Kodama, Y.; Kuriyama, K.; Yoshimura, K.; Tomono, T.; Morita, T.; Kakiuchi, N.; Matsumori, T.; Mima, A.; Nishikawa, Y. Pathogenicity of IgG in patients with IgG4-related disease. Gut 2016, 65, 1322–1332. [Google Scholar] [CrossRef]
- Qin, L.; Tang, L.-F.; Cheng, L.; Wang, H.-Y. The clinical significance of allergen-specific IgG4 in allergic diseases. Frontiers in Immunology 2022, 13, 1032909. [Google Scholar] [CrossRef]
- Flicker, S.; Valenta, R. Renaissance of the blocking antibody concept in type I allergy. International archives of allergy and immunology 2003, 132, 13–24. [Google Scholar] [CrossRef]
- Wachholz, P.A.; Durham, S.R. Mechanisms of immunotherapy: IgG revisited. Current opinion in allergy and clinical immunology 2004, 4, 313–318. [Google Scholar] [CrossRef]
- Golden, D.B.; Meyers, D.A.; Kagey-Sobotka, A.; Valentine, M.D.; Lichtenstein, L.M. Clinical relevance of the venom-specific immunoglobulin G antibody level during immunotherapy. Journal of Allergy and Clinical Immunology 1982, 69, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Müller, U.; Helbling, A.; Bischof, M. Predictive value of venom-specific IgE, IgG and IgG subclass antibodies in patients on immunotherapy with honey bee venom. Allergy 1989, 44, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Nouri-Aria, K.T.; Wachholz, P.A.; Francis, J.N.; Jacobson, M.R.; Walker, S.M.; Wilcock, L.K.; Staple, S.Q.; Aalberse, R.C.; Till, S.J.; Durham, S.R. Grass pollen immunotherapy induces mucosal and peripheral IL-10 responses and blocking IgG activity. The Journal of Immunology 2004, 172, 3252–3259. [Google Scholar] [CrossRef] [PubMed]
- Shamji, M.; Ljørring, C.; Francis, J.; A Calderon, M.; Larche, M.; Kimber, I.; Frew, A.; Ipsen, H.; Lund, K.; Würtzen, P. Functional rather than immunoreactive levels of IgG4 correlate closely with clinical response to grass pollen immunotherapy. Allergy 2012, 67, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Meiler, F.; Klunker, S.; Zimmermann, M.; Akdis, C.A.; Akdis, M. Distinct regulation of IgE, IgG4 and IgA by T regulatory cells and toll-like receptors. Allergy 2008, 63, 1455–1463. [Google Scholar] [CrossRef]
- Akdis, M.; Akdis, C.A. Mechanisms of allergen-specific immunotherapy: multiple suppressor factors at work in immune tolerance to allergens. Journal of Allergy and Clinical Immunology 2014, 133, 621–631. [Google Scholar] [CrossRef]
- Uversky, V.N.; Redwan, E.M.; Makis, W.; Rubio-Casillas, A. IgG4 Antibodies Induced by Repeated Vaccination May Generate Immune Tolerance to the SARS-CoV-2 Spike Protein. Vaccines 2023, 11, 991. [Google Scholar] [CrossRef]
- Irrgang, P.; Gerling, J.; Kocher, K.; Lapuente, D.; Steininger, P.; Habenicht, K.; Wytopil, M.; Beileke, S.; Schäfer, S.; Zhong, J. Class switch toward noninflammatory, spike-specific IgG4 antibodies after repeated SARS-CoV-2 mRNA vaccination. Science immunology 2022, 8, eade2798. [Google Scholar] [CrossRef]
- Buhre, J.S.; Pongracz, T.; Künsting, I.; Lixenfeld, A.S.; Wang, W.; Nouta, J.; Lehrian, S.; Schmelter, F.; Lunding, H.B.; Dühring, L. mRNA vaccines against SARS-CoV-2 induce comparably low long-term IgG Fc galactosylation and sialylation levels but increasing long-term IgG4 responses compared to an adenovirus-based vaccine. Frontiers in immunology 2023, 13. [Google Scholar] [CrossRef]
- Kiszel, P.; Sík, P.; Miklós, J.; Kajdácsi, E.; Sinkovits, G.; Cervenak, L.; Prohászka, Z. Class switch towards spike protein-specific IgG4 antibodies after SARS-CoV-2 mRNA vaccination depends on prior infection history. Scientific Reports 2023, 13, 13166. [Google Scholar] [CrossRef]
- Emmenegger, M.; Fiedler, S.; Brugger, S.D.; Devenish, S.R.; Morgunov, A.S.; Ilsley, A.; Ricci, F.; Malik, A.Y.; Scheier, T.; Batkitar, L. Both COVID-19 infection and vaccination induce high-affinity cross-clade responses to SARS-CoV-2 variants. Iscience 2022, 25. [Google Scholar] [CrossRef]
- Selva, K.J.; Ramanathan, P.; Haycroft, E.R.; Reynaldi, A.; Cromer, D.; Tan, C.W.; Wang, L.-F.; Wines, B.D.; Hogarth, P.M.; Downie, L.E. Preexisting immunity restricts mucosal antibody recognition of SARS-CoV-2 and Fc profiles during breakthrough infections. JCI insight 2023, 8. [Google Scholar] [CrossRef] [PubMed]
- Valk, A.M.; Keijser, J.B.; van Dam, K.P.; Stalman, E.W.; Wieske, L.; Steenhuis, M.; Kummer, L.Y.; Spuls, P.I.; Bekkenk, M.W.; Musters, A.H. Suppressed IgG4 class switching in dupilumab-and TNF inhibitor-treated patients after mRNA vaccination. Allergy 2024. [Google Scholar] [CrossRef]
- Hartley, G.E.; Fryer, H.A.; Gill, P.A.; Boo, I.; Bornheimer, S.J.; Hogarth, P.M.; Drummer, H.E.; O'Hehir, R.E.; Edwards, E.S.; van Zelm, M.C. Third dose COVID-19 mRNA vaccine enhances IgG4 isotype switching and recognition of Omicron subvariants by memory B cells after mRNA but not adenovirus priming. bioRxiv 2023, 2023.2009. 2015.557929.
- Farkash, I.; Feferman, T.; Cohen-Saban, N.; Avraham, Y.; Morgenstern, D.; Mayuni, G.; Barth, N.; Lustig, Y.; Miller, L.; Shouval, D.S. Anti-SARS-CoV-2 antibodies elicited by COVID-19 mRNA vaccine exhibit a unique glycosylation pattern. Cell Reports 2021, 37. [Google Scholar] [CrossRef]
- Sheehan, J.; Ardizzone, C.M.; Khanna, M.; Trauth, A.J.; Hagensee, M.E.; Ramsay, A.J. Dynamics of Serum-Neutralizing Antibody Responses in Vaccinees through Multiple Doses of the BNT162b2 Vaccine. Vaccines 2023, 11, 1720. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, M.; Sakamoto, A.; Ozuru, R.; Kurihara, Y.; Itoh, R.; Ishii, K.; Shimizu, A.; Chou, B.; Nabeshima, S.; Hiromatsu, K. The appearance of anti-spike receptor binding domain immunoglobulin G4 responses after repetitive immunization with messenger RNA-based COVID-19 vaccines. International Journal of Infectious Diseases 2024, 139, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.; Islam, M.R.; Khaton, F.; Soltana, U.H.; Jafrin, S.A.; Rahman, S.I.A.; Tauheed, I.; Ahmed, T.; Khan, I.I.; Akter, A. Appearance of tolerance-induction and non-inflammatory SARS-CoV-2 spike-specific IgG4 antibodies after COVID-19 booster vaccinations. Frontiers in Immunology 2023, 14. [Google Scholar] [CrossRef]
- Espino, A.M.; Armina-Rodriguez, A.; Alvarez, L.; Ocasio-Malavé, C.; Ramos-Nieves, R.; Rodriguez Martinó, E.I.; López-Marte, P.; Torres, E.A.; Sariol, C.A. The Anti-SARS-CoV-2 IgG1 and IgG3 Antibody Isotypes with Limited Neutralizing Capacity against Omicron Elicited in a Latin Population a Switch toward IgG4 after Multiple Doses with the mRNA Pfizer–BioNTech Vaccine. Viruses 2024, 16, 187. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.; Oltz, E.; Bednash, J.; Horowitz, J.; Rubinstein, M.; Vlasova, A.N. Brief Research Report: Impact of vaccination on antibody responses and mortality from severe COVID-19. Frontiers in Immunology 15, 1325243.
- Kalkeri, R.; Zhu, M.; Cloney-Clark, S.; Plested, J.S.; Parekh, A.; Gorinson, D.; Cai, R.; Mahato, S.; Ramanathan, P.; Aurelia, L.C. Altered IgG4 Antibody Response to Repeated mRNA versus Protein COVID Vaccines. medRxiv 2024, 2024.2001. 2017.24301374.
- Nziza, N.; Deng, Y.; Wood, L.; Dhanoa, N.; Dulit-Greenberg, N.; Chen, T.; Kane, A.S.; Swank, Z.; Davis, J.P.; Demokritou, M. Humoral profiles of toddlers and young children following SARS-CoV-2 mRNA vaccination. Nature Communications 2024, 15, 905. [Google Scholar] [CrossRef]
- Jain, S.; Kumar, S.; Lai, L.; Linderman, S.; Malik, A.A.; Ellis, M.L.; Godbole, S.; Solis, D.; Sahoo, M.K.; Bechnak, K. XBB. 1.5 monovalent booster improves antibody binding and neutralization against emerging SARS-CoV-2 Omicron variants. bioRxiv 2024, 2024.2002. 2003.578771.
- Portilho, A.I.; Silva, V.O.; Da Costa, H.H.M.; Yamashiro, R.; de Oliveira, I.P.; de Campos, I.B.; Prudencio, C.R.; Matsuda, E.M.; de Macedo Brígido, L.F.; De Gaspari, E. An unexpected IgE anti-receptor binding domain response following natural infection and different types of SARS-CoV-2 vaccines. Scientific Reports 2024, 14, 20003. [Google Scholar] [CrossRef]
- Routhu, N.K.; Stampfer, S.D.; Lai, L.; Akhtar, A.; Tong, X.; Yuan, D.; Chicz, T.M.; McNamara, R.P.; Jakkala, K.; Davis-Gardner, M.E. Efficacy of mRNA-1273 and Novavax ancestral or BA. 1 spike booster vaccines against SARS-CoV-2 BA. 5 infection in non-human primates. Science Immunology 2023, eadg7015.
- Valk, A.M.; Keijser, J.B.; van Dam, K.P.; Stalman, E.W.; Wieske, L.; Steenhuis, M.; Kummer, L.Y.; Spuls, P.I.; Bekkenk, M.W.; Musters, A.H. Suppressed IgG4 class switching in dupilumab-and TNF inhibitor-treated patients after repeated SARS-CoV-2 mRNA vaccination. medRxiv 2023, 2023.2009. 2029.23296354.
- Liu, Z.; Cai, L.; Xing, M.; Qiao, N.; Liu, J.; Li, X.; Zhang, C.; Tang, N.; Xu, Z.; Guo, Y. Evaluation of antibody responses in healthy individuals receiving SARS-CoV-2 inactivated vaccines. Biosafety and Health 2024, 6, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Suzuki, Y.; Yamamoto, N.; Matsumoto, Y.; Shirai, A.; Okubo, T. Influenza A virus infection increases IgE production and airway responsiveness in aerosolized antigen-exposed mice. Journal of allergy and clinical immunology 1998, 102, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Dakhama, A.; Lee, Y.-M.; Ohnishi, H.; Jing, X.; Balhorn, A.; Takeda, K.; Gelfand, E.W. Virus-specific IgE enhances airway responsiveness on reinfection with respiratory syncytial virus in newborn mice. Journal of Allergy and Clinical Immunology 2009, 123, 138–145. e135. [Google Scholar] [CrossRef] [PubMed]
- Guclu, O.A.; Goktas, S.S.; Dilektasli, A.G.; Ozturk, N.A.A.; Demirdogen, E.; Coskun, F.; Ediger, D.; Ursavas, A.; Uzaslan, E.; Erol, H.A. A pilot study for IgE as a prognostic biomarker in COVID-19. Internal Medicine Journal 2022, 10.1111/imj. 15728.
- Ferastraoaru, D.; Hudes, G.; Jerschow, E.; Jariwala, S.; Karagic, M.; de Vos, G.; Rosenstreich, D.; Ramesh, M. Eosinophilia in asthma patients is protective against severe COVID-19 illness. The Journal of Allergy and Clinical Immunology: In Practice 2021, 9, 1152–1162. e1153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-j.; Dong, X.; Cao, Y.-y.; Yuan, Y.-d.; Yang, Y.-b.; Yan, Y.-q.; Akdis, C.A.; Gao, Y.-d. Clinical characteristics of 140 patients infected with SARS-CoV-2 in Wuhan, China. Allergy 2020, 75, 1730–1741. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Tu, L.; Zhu, P.; Mu, M.; Wang, R.; Yang, P.; Wang, X.; Hu, C.; Ping, R.; Hu, P. Clinical features of 85 fatal cases of COVID-19 from Wuhan. A retrospective observational study. American journal of respiratory and critical care medicine 2020, 201, 1372–1379. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo-Muñoz, J.; Sastre, B.; Cañas, J.; Gil-Martínez, M.; Redondo, N.; Del Pozo, V. Eosinophil response against classical and emerging respiratory viruses: COVID-19. J Investig Allergol Clin Immunol 2021, 31, 94–107. [Google Scholar] [CrossRef]
- Plūme, J.; Galvanovskis, A.; Šmite, S.; Romanchikova, N.; Zayakin, P.; Linē, A. Early and strong antibody responses to SARS-CoV-2 predict disease severity in COVID-19 patients. Journal of Translational Medicine 2022, 20, 176. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.M.; Jackson, K.J. A temporal model of human IgE and IgG antibody function. Frontiers in immunology 2013, 4, 235. [Google Scholar] [CrossRef] [PubMed]
- Jeannin, P.; Delneste, Y.; Lecoanet-Henchoz, S.; Gretener, D.; Bonnefoy, J.-Y. Interleukin-7 (IL-7) enhances class switching to IgE and IgG4 in the presence of T cells via IL-9 and sCD23. Blood, The Journal of the American Society of Hematology 1998, 91, 1355–1361. [Google Scholar] [CrossRef]
- He, P.; Zou, Y.; Hu, Z. Advances in aluminum hydroxide-based adjuvant research and its mechanism. 2015.
- Kinet, J.-P. The high-affinity IgE receptor (FcϵRI): from physiology to pathology. Annual review of immunology 1999, 17, 931–972. [Google Scholar] [CrossRef] [PubMed]
- Kempuraj, D.; Selvakumar, G.P.; Ahmed, M.E.; Raikwar, S.P.; Thangavel, R.; Khan, A.; Zaheer, S.A.; Iyer, S.S.; Burton, C.; James, D. COVID-19, mast cells, cytokine storm, psychological stress, and neuroinflammation. The Neuroscientist 2020, 26, 402–414. [Google Scholar] [PubMed]
- Afrin, L.B.; Weinstock, L.B.; Molderings, G.J. Covid-19 hyperinflammation and post-Covid-19 illness may be rooted in mast cell activation syndrome. International journal of infectious diseases 2020, 100, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Motta Junior, J.d.S.; Miggiolaro, A.F.R.d.S.; Nagashima, S.; De Paula, C.B.V.; Baena, C.P.; Scharfstein, J.; De Noronha, L. Mast cells in alveolar septa of COVID-19 patients: a pathogenic pathway that may link interstitial edema to immunothrombosis. Frontiers in Immunology 2020, 11, 574862. [Google Scholar] [CrossRef] [PubMed]
- Farmani, A.R.; Mahdavinezhad, F.; Moslemi, R.; Mehrabi, Z.; Noori, A.; Kouhestani, M.; Noroozi, Z.; Ai, J.; Rezaei, N. Anti-IgE monoclonal antibodies as potential treatment in COVID-19. Immunopharmacology and immunotoxicology 2021, 43, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Diaz, A.; Romero, M.; Mendez, N.V.; Landin, A.M.; Blomberg, B.B. Effects of age on H1N1-specific serum IgG1 and IgG3 levels evaluated during the 2011–2012 influenza vaccine season. Immunity & ageing 2013, 10, 1–9. [Google Scholar]
- Cavacini, L.A.; Kuhrt, D.; Duval, M.; Mayer, K.; Posner, M.R. Binding and neutralization activity of human IgG1 and IgG3 from serum of HIV-infected individuals. AIDS research and human retroviruses 2003, 19, 785–792. [Google Scholar] [PubMed]
- Suthar, M.S.; Zimmerman, M.G.; Kauffman, R.C.; Mantus, G.; Linderman, S.L.; Hudson, W.H.; Vanderheiden, A.; Nyhoff, L.; Davis, C.W.; Adekunle, O. Rapid generation of neutralizing antibody responses in COVID-19 patients. Cell Reports Medicine 2020, 1. [Google Scholar] [CrossRef]
- Mazzini, L.; Martinuzzi, D.; Hyseni, I.; Benincasa, L.; Molesti, E.; Casa, E.; Lapini, G.; Piu, P.; Trombetta, C.M.; Marchi, S. Comparative analyses of SARS-CoV-2 binding (IgG, IgM, IgA) and neutralizing antibodies from human serum samples. Journal of Immunological Methods 2021, 489, 112937. [Google Scholar] [CrossRef]
- Wright, D.N.; Nelson Jr, R.P.; Ledford, D.K.; Fernandez-Caldas, E.; Trudeau, W.L.; Lockey, R.F. Serum IgE and human immunodeficiency virus (HIV) infection. Journal of allergy and clinical immunology 1990, 85, 445–452. [Google Scholar] [CrossRef]
- Ammann, A.J.; Abrams, D.; Conant, M.; Chudwin, D.; Cowan, M.; Volberding, P.; Lewis, B.; Casavant, C. Acquired immune dysfunction in homosexual men: immunologic profiles. Clinical immunology and immunopathology 1983, 27, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Ring, J.; Fröschl, M.; Brunner, R.; Braun-Falco, O. LAV/HTLV-III infection and atopy: serum IgE and specific IgE antibodies to environmental allergens. Acta dermato-venereologica 1986, 66, 530–532. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.Y. Chronic diffuse dermatitis and hyper-IgE in HIV infection. Acta dermato-venereologica 1988, 68, 486–491. [Google Scholar] [PubMed]
- Miguez-Burbano, M.; Shor-Posner, G.; Fletcher, M.A.; Lu, Y.; Moreno, J.; Carcamo, C.; Page, B.; Quesada, J.; Sauberlich, H.; Baum, M. Immunoglobulin E levels in relationship to HIV-1 disease, route of infection, and vitamin E status. Allergy 1995, 50, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Ouaaz, F.; Ruscetti, F.W.; Dugas, B.; Mikovits, J.; Agut, H.; Debr, P.; Mossalayi, M.D. Role of IgE Immune Complexes in the Regulation of HIV-1 Replication and Increased Cell Death of Infected U1 Monocytes: Involvement of CD23/Fc ε RII-Mediated Nitric Oxide and Cyclic AMP Pathways. Molecular Medicine 1996, 2, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Ellaurie, M.; Rubinstein, A.; Rosenstreich, D.L. IgE levels in pediatric HIV-1 infection. Annals of allergy, asthma & immunology: official publication of the American College of Allergy, Asthma, & Immunology 1995, 75, 332–336. [Google Scholar]
- Israël-Biet, D.; Labrousse, F.; Tourani, J.-M.; Sors, H.; Andrieu, J.-M.; Even, P. Elevation of IgE in HIV-infected subjects: a marker of poor prognosis. Journal of allergy and clinical immunology 1992, 89, 68–75. [Google Scholar] [CrossRef] [PubMed]
- LUCEY, D.R.; ZAJAC, R.A.; MELCHER, G.P.; BUTZIN, C.A.; BOSWELL, R.N. Serum IgE levels in 622 persons with human immunodeficiency virus infection: IgE elevation with marked depletion of CD4+ T-cells. AIDS research and human retroviruses 1990, 6, 427–429. [Google Scholar] [CrossRef]
- Shor-Posner, G.; Miguez-Burbano, M.J.; Lu, Y.; Feaster, D.; Fletcher, M.; Sauberlich, H.; Baum, M.K. Elevated IgE level in relationship to nutritional status and immune parameters in early human immunodeficiency virus–1 disease. Journal of allergy and clinical immunology 1995, 95, 886–892. [Google Scholar] [CrossRef]
- Bul, R.H.D.; Molinaro, G.A.; Kettering, J.D.; Heiner, D.C.; Imagawa, D.T.; Geme Jr, J.W.S. Virus-specific IgE and IgG4 antibodies in serum of children infected with respiratory syncytial virus. The Journal of pediatrics 1987, 110, 87–90. [Google Scholar]
- Becker, Y. Respiratory syncytial virus (RSV) evades the human adaptive immune system by skewing the Th1/Th2 cytokine balance toward increased levels of Th2 cytokines and IgE, markers of allergy—a review. Virus genes 2006, 33, 235–252. [Google Scholar] [PubMed]
- Welliver, R.C.; Sun, M.; Rinaldo, D.; Ogra, P.L. Respiratory syncytial virus-specific IgE responses following infection: evidence for a predominantly mucosal response. Pediatric research 1985, 19, 420–424. [Google Scholar] [PubMed]
- Smith-Norowitz, T.A.; Josekutty, J.; Silverberg, J.I.; Lev-Tov, H.; Norowitz, Y.M.; Kohlhoff, S.; Nowakowski, M.; Durkin, H.G.; Bluth, M.H. Long term persistence of IgE anti-Varicella Zoster Virus in pediatric and adult serum post chicken pox infection and after vaccination with Varicella Virus vaccine. International Journal of Biomedical Science: IJBS 2009, 5, 353. [Google Scholar]
- Smith-Norowitz, T.A.; Tam, E.; Norowitz, K.B.; Chotikanatis, K.; Weaver, D.; Durkin, H.G.; Bluth, M.H.; Kohlhoff, S. IgE anti Hepatitis B virus surface antigen antibodies detected in serum from inner city asthmatic and non asthmatic children. Human Immunology 2014, 75, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Smith-Norowitz, T.A.; Wong, D.; Kusonruksa, M.; Norowitz, K.B.; Joks, R.; Durkin, H.G.; Bluth, M.H. Long term persistence of IgE anti-influenza virus antibodies in pediatric and adult serum post vaccination with influenza virus vaccine. International journal of medical sciences 2011, 8, 239. [Google Scholar] [CrossRef] [PubMed]
- Della-Torre, E.; Lanzillotta, M.; Strollo, M.; Ramirez, G.A.; Dagna, L.; Tresoldi, M. Serum IgG4 level predicts COVID-19 related mortality. European Journal of Internal Medicine 2021, 93, 107–109. [Google Scholar] [PubMed]
- Moura, A.D.; da Costa, H.H.; Correa, V.A.; de, S. Lima, A.K.; Lindoso, J.A.; De Gaspari, E.; Hong, M.A.; Cunha-Junior, J.P.; Prudencio, C.R. Assessment of avidity related to IgG subclasses in SARS-CoV-2 Brazilian infected patients. Scientific Reports 2021, 11, 17642. [Google Scholar] [CrossRef]
- Genova, S.N.; Pencheva, M.M.; Abadjieva, T.I.; Atanasov, N.G. Cellular and immune response in fatal COVID-19 pneumonia. The Pan African Medical Journal 2024, 49. [Google Scholar]
- Santos, A.F.; James, L.K.; Bahnson, H.T.; Shamji, M.H.; Couto-Francisco, N.C.; Islam, S.; Houghton, S.; Clark, A.T.; Stephens, A.; Turcanu, V. IgG4 inhibits peanut-induced basophil and mast cell activation in peanut-tolerant children sensitized to peanut major allergens. Journal of Allergy and Clinical Immunology 2015, 135, 1249–1256. [Google Scholar]
- Lambin, P.; Bouzoumou, A.; Murrieta, M.; Debbia, M.; Rouger, P.; Leynadier, F.; Levy, D. Purification of human IgG4 subclass with allergen-specific blocking activity. Journal of immunological methods 1993, 165, 99–111. [Google Scholar] [CrossRef]
- Rubio-Casillas, A.; Redwan, E.M.; Uversky, V.N. Does SARS-CoV-2 induce IgG4 synthesis to evade the immune system? Biomolecules 2023, 13, 1338. [Google Scholar] [CrossRef] [PubMed]
- Balz, K.; Kaushik, A.; Chen, M.; Cemic, F.; Heger, V.; Renz, H.; Nadeau, K.; Skevaki, C. Homologies between SARS-CoV-2 and allergen proteins may direct T cell-mediated heterologous immune responses. Scientific reports 2021, 11, 4792. [Google Scholar] [CrossRef] [PubMed]
- Balz, K.; Trassl, L.; Härtel, V.; Nelson, P.P.; Skevaki, C. Virus-induced T cell-mediated heterologous immunity and vaccine development. Frontiers in immunology 2020, 11, 513. [Google Scholar] [CrossRef]
- Pusch, E.; Renz, H.; Skevaki, C. Respiratory virus-induced heterologous immunity: Part of the problem or part of the solution? Allergo Journal 2018, 27, 28–45. [Google Scholar] [CrossRef]
- Karagöz, I.K.; Kaya, M.; Rückert, R.; Bozman, N.; Kaya, V.; Bayram, H.; Yıldırım, M. A bioinformatic analysis: Previous allergen exposure may support anti-SARS-CoV-2 immune response. Computational Biology and Chemistry 2023, 107, 107961. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Hasegawa, K.; Ma, B.; Fujiogi, M.; Camargo, C.A.; Liang, L. Association of asthma and its genetic predisposition with the risk of severe COVID-19. Journal of Allergy and Clinical Immunology 2020, 146, 327–329. e324. [Google Scholar] [CrossRef] [PubMed]
- Skevaki, C.; Karsonova, A.; Karaulov, A.; Fomina, D.; Xie, M.; Chinthrajah, S.; Nadeau, K.C.; Renz, H. SARS-CoV-2 infection and COVID-19 in asthmatics: a complex relationship. Nature Reviews Immunology 2021, 21, 202–203. [Google Scholar] [CrossRef] [PubMed]
- Skevaki, C.; Karsonova, A.; Karaulov, A.; Xie, M.; Renz, H. Asthma-associated risk for COVID-19 development. Journal of allergy and clinical immunology 2020, 146, 1295–1301. [Google Scholar] [CrossRef]
- Pradhan, P.; Pandey, A.K.; Mishra, A.; Gupta, P.; Tripathi, P.K.; Menon, M.B.; Gomes, J.; Vivekanandan, P.; Kundu, B. Uncanny similarity of unique inserts in the 2019-nCoV spike protein to HIV-1 gp120 and Gag. BioRxiv 2020. [Google Scholar]
- Worobey, M.; Holmes, E.C. Evolutionary aspects of recombination in RNA viruses. Journal of General Virology 1999, 80, 2535–2543. [Google Scholar] [CrossRef]
- Lai, M. RNA recombination in animal and plant viruses. Microbiological reviews 1992, 56, 61–79. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.L.; Hahn, B.H.; Sharp, P.M. Recombination in AIDS viruses. Journal of molecular evolution 1995, 40, 249–259. [Google Scholar] [CrossRef]
- Rubio-Casillas, A.; Redwan, E.M.; Uversky, V.N. SARS-CoV-2: a master of immune evasion. Biomedicines 2022, 10, 1339. [Google Scholar] [CrossRef]
- Caetano-Anollés, K.; Hernandez, N.; Mughal, F.; Tomaszewski, T.; Caetano-Anollés, G. The seasonal behaviour of COVID-19 and its galectin-like culprit of the viral spike. In Methods in Microbiology; Elsevier: 2022; Volume 50, pp. 27-81.
- Hirani, N.; MacKinnon, A.C.; Nicol, L.; Ford, P.; Schambye, H.; Pedersen, A.; Nilsson, U.J.; Leffler, H.; Sethi, T.; Tantawi, S. Target inhibition of galectin-3 by inhaled TD139 in patients with idiopathic pulmonary fibrosis. European Respiratory Journal 2021, 57. [Google Scholar] [CrossRef] [PubMed]
- Bambouskova, M.; Polakovicova, I.; Halova, I.; Goel, G.; Draberova, L.; Bugajev, V.; Doan, A.; Utekal, P.; Gardet, A.; Xavier, R.J. New regulatory roles of galectin-3 in high-affinity IgE receptor signaling. Molecular and cellular biology 2016, 36, 1366–1382. [Google Scholar] [CrossRef] [PubMed]
- Tsilioni, I.; Theoharides, T.C. Recombinant SARS-CoV-2 spike protein stimulates secretion of chymase, tryptase, and IL-1β from human mast cells, augmented by IL-33. International Journal of Molecular Sciences 2023, 24, 9487. [Google Scholar] [PubMed]
- Perugino, C.A.; AlSalem, S.B.; Mattoo, H.; Della-Torre, E.; Mahajan, V.; Ganesh, G.; Allard-Chamard, H.; Wallace, Z.; Montesi, S.B.; Kreuzer, J. Identification of galectin-3 as an autoantigen in patients with IgG4-related disease. Journal of Allergy and Clinical Immunology 2019, 143, 736–745. e736. [Google Scholar]
- Motta, R.V.; Culver, E.L. IgG4 autoantibodies and autoantigens in the context of IgG4-autoimmune disease and IgG4-related disease. Frontiers in Immunology 2024, 15, 1272084. [Google Scholar]
- Raszek, M.; Cowley, D.; Redwan, E.M.; Uversky, V.N.; Rubio-Casillas, A. Exploring the possible link between the spike protein immunoglobulin G4 antibodies and cancer progression. Exploration of immunology 2024, 4, 267–284. [Google Scholar]
- Umetsu, D.T.; DeKruyff, R.H. TH1 and TH2 CD4+ cells in human allergic diseases. Journal of allergy and Clinical Immunology 1997, 100, 1–6. [Google Scholar]
- Becker, Y. The changes in the T helper 1 (Th1) and T helper 2 (Th2) cytokine balance during HIV-1 infection are indicative of an allergic response to viral proteins that may be reversed by Th2 cytokine inhibitors and immune response modifiers–a review and hypothesis. Virus genes 2004, 28, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Becker, Y. HIV-1 gp41 heptad repeat 2 (HR2) possesses an amino acid domain that resembles the allergen domain in Aspergillus fumigatus Asp f1 protein: review, hypothesis and implications. Virus Genes 2007, 34, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, S.; Del Prete, G.; Manetti, R.; Ravina, A.; Annunziato, F.; De Carli, M.; Mazzetti, M.; Piccinni, M.-P.; D'Elios, M.M.; Parronchi, P. Role of TH1/TH2 cytokines in HIV infection. Immunological reviews 1994, 140, 73–92. [Google Scholar] [CrossRef]
- Wong, G.H.; Goeddel, D.V. Tumour necrosis factors α and β inhibit virus replication and synergize with interferons. Nature 1986, 323, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Gajewski, T.F.; Fitch, F.W. Anti-proliferative effect of IFN-gamma in immune regulation. I. IFN-gamma inhibits the proliferation of Th2 but not Th1 murine helper T lymphocyte clones. Journal of immunology (Baltimore, Md.: 1950) 1988, 140, 4245–4252. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T.R.; Moore, K.W. The role of IL-10 in crossregulation of TH1 and TH2 responses. Immunology today 1991, 12, A49–A53. [Google Scholar] [CrossRef] [PubMed]
- Becker, Y. HIV-1 induced AIDS is an allergy and the allergen is the Shed gp120–a review, hypothesis, and implications. Virus Genes 2004, 28, 319–331. [Google Scholar] [CrossRef]
- Da Silva, L.; Da Silva, J. Relationship between atopy, allergic diseases and total serum IgE levels among HIV-infected children. European annals of allergy and clinical immunology 2013, 45, 155–159. [Google Scholar]
- Gill, M.A.; Bajwa, G.; George, T.A.; Dong, C.C.; Dougherty, I.I.; Jiang, N.; Gan, V.N.; Gruchalla, R.S. Counterregulation between the FcεRI pathway and antiviral responses in human plasmacytoid dendritic cells. The Journal of Immunology 2010, 184, 5999–6006. [Google Scholar] [CrossRef]
- Gielen, V.; Sykes, A.; Zhu, J.; Chan, B.; Macintyre, J.; Regamey, N.; Kieninger, E.; Gupta, A.; Shoemark, A.; Bossley, C. Increased nuclear suppressor of cytokine signaling 1 in asthmatic bronchial epithelium suppresses rhinovirus induction of innate interferons. Journal of Allergy and Clinical Immunology 2015, 136, 177–188. e111. [Google Scholar] [CrossRef]
- Lynch, J.P.; Werder, R.B.; Simpson, J.; Loh, Z.; Zhang, V.; Haque, A.; Spann, K.; Sly, P.D.; Mazzone, S.B.; Upham, J.W. Aeroallergen-induced IL-33 predisposes to respiratory virus–induced asthma by dampening antiviral immunity. Journal of Allergy and Clinical Immunology 2016, 138, 1326–1337. [Google Scholar] [CrossRef] [PubMed]
- Durrani, S.R.; Montville, D.J.; Pratt, A.S.; Sahu, S.; DeVries, M.K.; Rajamanickam, V.; Gangnon, R.E.; Gill, M.A.; Gern, J.E.; Lemanske Jr, R.F. Innate immune responses to rhinovirus are reduced by the high-affinity IgE receptor in allergic asthmatic children. Journal of Allergy and Clinical Immunology 2012, 130, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Duff, A.L.; Pomeranz, E.S.; Gelber, L.E.; Price, G.W.; Farris, H.; Hayden, F.G.; Platts-Mills, T.A.; Heymann, P.W. Risk factors for acute wheezing in infants and children: viruses, passive smoke, and IgE antibodies to inhalant allergens. Pediatrics 1993, 92, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Teach, S.J.; Gergen, P.J.; Szefler, S.J.; Mitchell, H.E.; Calatroni, A.; Wildfire, J.; Bloomberg, G.R.; Kercsmar, C.M.; Liu, A.H.; Makhija, M.M. Seasonal risk factors for asthma exacerbations among inner-city children. Journal of Allergy and Clinical Immunology 2015, 135, 1465–1473. e1465. [Google Scholar] [CrossRef] [PubMed]
- Soto-Quiros, M.; Avila, L.; Platts-Mills, T.A.; Hunt, J.F.; Erdman, D.D.; Carper, H.; Murphy, D.D.; Odio, S.; James, H.R.; Patrie, J.T. High titers of IgE antibody to dust mite allergen and risk for wheezing among asthmatic children infected with rhinovirus. Journal of Allergy and Clinical Immunology 2012, 129, 1499–1505. e1495. [Google Scholar] [CrossRef] [PubMed]
- Swiecki, M.; Colonna, M. The multifaceted biology of plasmacytoid dendritic cells. Nature Reviews Immunology 2015, 15, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Bencze, D.; Fekete, T.; Pázmándi, K. Type I interferon production of plasmacytoid dendritic cells under control. International journal of molecular sciences 2021, 22, 4190. [Google Scholar] [CrossRef] [PubMed]
- Gill, M.A.; Liu, A.H.; Calatroni, A.; Krouse, R.Z.; Shao, B.; Schiltz, A.; Gern, J.E.; Togias, A.; Busse, W.W. Enhanced plasmacytoid dendritic cell antiviral responses after omalizumab. Journal of Allergy and Clinical Immunology 2018, 141, 1735–1743. e1739. [Google Scholar] [CrossRef] [PubMed]
- Teach, S.J.; Gill, M.A.; Togias, A.; Sorkness, C.A.; Arbes Jr, S.J.; Calatroni, A.; Wildfire, J.J.; Gergen, P.J.; Cohen, R.T.; Pongracic, J.A. Preseasonal treatment with either omalizumab or an inhaled corticosteroid boost to prevent fall asthma exacerbations. Journal of Allergy and Clinical Immunology 2015, 136, 1476–1485. [Google Scholar] [CrossRef]
- Esquivel, A.; Busse, W.W.; Calatroni, A.; Togias, A.G.; Grindle, K.G.; Bochkov, Y.A.; Gruchalla, R.S.; Kattan, M.; Kercsmar, C.M.; Khurana Hershey, G. Effects of omalizumab on rhinovirus infections, illnesses, and exacerbations of asthma. American journal of respiratory and critical care medicine 2017, 196, 985–992. [Google Scholar] [CrossRef]
- Meng, B.; Abdullahi, A.; Ferreira, I.A.; Goonawardane, N.; Saito, A.; Kimura, I.; Yamasoba, D.; Gerber, P.P.; Fatihi, S.; Rathore, S. Altered TMPRSS2 usage by SARS-CoV-2 Omicron impacts infectivity and fusogenicity. Nature 2022, 603, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Peacock, T.P.; Brown, J.C.; Zhou, J.; Thakur, N.; Sukhova, K.; Newman, J.; Kugathasan, R.; Yan, A.W.; Furnon, W.; De Lorenzo, G. The altered entry pathway and antigenic distance of the SARS-CoV-2 Omicron variant map to separate domains of spike protein. Biorxiv 2022, 2021.2012. 2031.474653.
- McMahan, K.; Giffin, V.; Tostanoski, L.H.; Chung, B.; Siamatu, M.; Suthar, M.S.; Halfmann, P.; Kawaoka, Y.; Piedra-Mora, C.; Jain, N. Reduced pathogenicity of the SARS-CoV-2 omicron variant in hamsters. Med 2022, 3, 262–268. e264. [Google Scholar] [CrossRef] [PubMed]
- Willett, B.J.; Grove, J.; MacLean, O.A.; Wilkie, C.; De Lorenzo, G.; Furnon, W.; Cantoni, D.; Scott, S.; Logan, N.; Ashraf, S. SARS-CoV-2 Omicron is an immune escape variant with an altered cell entry pathway. Nature microbiology 2022, 7, 1161–1179. [Google Scholar] [CrossRef] [PubMed]
- Ariën, K.K.; Vanham, G.; Arts, E.J. Is HIV-1 evolving to a less virulent form in humans? Nature Reviews Microbiology 2007, 5, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Grobben, M.; Bakker, M.; Schriek, A.I.; Levels, L.J.; Umotoy, J.C.; Tejjani, K.; van Breemen, M.J.; Lin, R.N.; de Taeye, S.W.; Ozorowski, G. Polyfunctionality and breadth of HIV-1 antibodies are associated with delayed disease progression. PLoS pathogens 2024, 20, e1012739. [Google Scholar] [CrossRef]
- Chung, A.W.; Ghebremichael, M.; Robinson, H.; Brown, E.; Choi, I.; Lane, S.; Dugast, A.-S.; Schoen, M.K.; Rolland, M.; Suscovich, T.J. Polyfunctional Fc-effector profiles mediated by IgG subclass selection distinguish RV144 and VAX003 vaccines. Science translational medicine 2014, 6, 228ra238–228ra238. [Google Scholar] [CrossRef] [PubMed]
- Karnasuta, C.; Akapirat, S.; Madnote, S.; Savadsuk, H.; Puangkaew, J.; Rittiroongrad, S.; Rerks-Ngarm, S.; Nitayaphan, S.; Pitisuttithum, P.; Kaewkungwal, J. Comparison of antibody responses induced by RV144, VAX003, and VAX004 vaccination regimens. AIDS research and human retroviruses 2017, 33, 410–423. [Google Scholar] [CrossRef]
- Yates, N.L.; Liao, H.-X.; Fong, Y.; DeCamp, A.; Vandergrift, N.A.; Williams, W.T.; Alam, S.M.; Ferrari, G.; Yang, Z.-y.; Seaton, K.E. Vaccine-induced Env V1-V2 IgG3 correlates with lower HIV-1 infection risk and declines soon after vaccination. Science translational medicine 2014, 6, 228ra239–228ra239. [Google Scholar]
- Ioannou, G.N.; Locke, E.R.; O’Hare, A.M.; Bohnert, A.S.; Boyko, E.J.; Hynes, D.M.; Berry, K. COVID-19 vaccination effectiveness against infection or death in a national US health care system: a target trial emulation study. Annals of Internal Medicine 2022, 175, 352–361. [Google Scholar] [CrossRef]
- Nakatani, E.; Morioka, H.; Kikuchi, T.; Fukushima, M. Behavioral and Health Outcomes of mRNA COVID-19 Vaccination: A Case-Control Study in Japanese Small and Medium-Sized Enterprises. Cureus 2024, 16. [Google Scholar] [CrossRef]
- Eythorsson, E.; Runolfsdottir, H.L.; Ingvarsson, R.F.; Sigurdsson, M.I.; Palsson, R. Rate of SARS-CoV-2 reinfection during an omicron wave in Iceland. JAMA network open 2022, 5, e2225320–e2225320. [Google Scholar] [CrossRef] [PubMed]
- Chemaitelly, H.; Tang, P.; Hasan, M.R.; AlMukdad, S.; Yassine, H.M.; Benslimane, F.M.; Al Khatib, H.A.; Coyle, P.; Ayoub, H.H.; Al Kanaani, Z. Waning of BNT162b2 vaccine protection against SARS-CoV-2 infection in Qatar. New England Journal of Medicine 2021, 385, e83. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.K.; Burke, P.C.; Nowacki, A.S.; Gordon, S.M. Risk of coronavirus disease 2019 (COVID-19) among those up-to-date and not up-to-date on COVID-19 vaccination by US CDC criteria. PLoS One 2023, 18, e0293449. [Google Scholar] [CrossRef] [PubMed]
- Feldstein, L.R.; Ruffin, J.; Wiegand, R.; Grant, L.; Babu, T.M.; Briggs-Hagen, M.; Burgess, J.L.; Caban-Martinez, A.J.; Chu, H.Y.; Ellingson, K.D. Protection from covid-19 vaccination and prior sars-cov-2 infection among children aged 6 months–4 years, united states, september 2022–april 2023. Journal of the Pediatric Infectious Diseases Society 2025, 14, piae121. [Google Scholar] [CrossRef]
- Kober, C.; Manni, S.; Wolff, S.; Barnes, T.; Mukherjee, S.; Vogel, T.; Hoenig, L.; Vogel, P.; Hahn, A.; Gerlach, M. IgG3 and IgM identified as key to SARS-CoV-2 neutralization in convalescent plasma pools. PLoS One 2022, 17, e0262162. [Google Scholar] [CrossRef] [PubMed]
- Whitcombe, A.L.; McGregor, R.; Craigie, A.; James, A.; Charlewood, R.; Lorenz, N.; Dickson, J.M.; Sheen, C.R.; Koch, B.; Fox-Lewis, S. Comprehensive analysis of SARS-CoV-2 antibody dynamics in New Zealand. Clinical & translational immunology 2021, 10, e1261. [Google Scholar]
- Gazit, S.; Shlezinger, R.; Perez, G.; Lotan, R.; Peretz, A.; Ben-Tov, A.; Cohen, D.; Muhsen, K.; Chodick, G.; Patalon, T. Comparing SARS-CoV-2 natural immunity to vaccine-induced immunity: reinfections versus breakthrough infections. MedRxiv 2021. [Google Scholar]
- Redman, M.; King, A.; Watson, C.; King, D. What is CRISPR/Cas9? Archives of Disease in Childhood-Education and Practice 2016, 101, 213–215. [Google Scholar] [CrossRef]
- Dobaño, C.; Quelhas, D.; Quintó, L.; Puyol, L.; Serra-Casas, E.; Mayor, A.; Nhampossa, T.; Macete, E.; Aide, P.; Mandomando, I. Age-dependent IgG subclass responses to Plasmodium falciparum EBA-175 are differentially associated with incidence of malaria in Mozambican children. Clinical and Vaccine Immunology 2012, 19, 157–166. [Google Scholar] [CrossRef]
- Aucan, C.; Traoré, Y.; Tall, F.o.; Nacro, B.; Traoré-Leroux, T.r.s.; Fumoux, F.; Rihet, P. High immunoglobulin G2 (IgG2) and low IgG4 levels are associated with human resistance to Plasmodium falciparum malaria. Infection and immunity 2000, 68, 1252–1258. [Google Scholar] [CrossRef]
- Groux, H.; Gysin, J. Opsonization as an effector mechanism in human protection against asexual blood stages of Plasmodium falciparum: functional role of IgG subclasses. Research in immunology 1990, 141, 529–542. [Google Scholar] [PubMed]
- Thau, L.; Asuka, E.; Mahajan, K. Physiology, opsonization. StatPearls [Internet] 2023.
Figure 1.
The function that allergen-specific IgG4 plays in exposure to allergens. (A) Acute allergen exposure may cause IgE-mediated histamine-induced hypersensitivity reactions through the high-affinity receptor FcϵRI. (B) Long-term exposure to allergens may cause the development of IgG4, which then binds to the allergen. IgG4 then binds to the inhibitory receptor FcγRIIb located on mast cells and basophils, thus limiting hypersensitive reactions and promoting allergy tolerance. Source: [19]. This is an open-access article and distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Figure 1.
The function that allergen-specific IgG4 plays in exposure to allergens. (A) Acute allergen exposure may cause IgE-mediated histamine-induced hypersensitivity reactions through the high-affinity receptor FcϵRI. (B) Long-term exposure to allergens may cause the development of IgG4, which then binds to the allergen. IgG4 then binds to the inhibitory receptor FcγRIIb located on mast cells and basophils, thus limiting hypersensitive reactions and promoting allergy tolerance. Source: [19]. This is an open-access article and distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
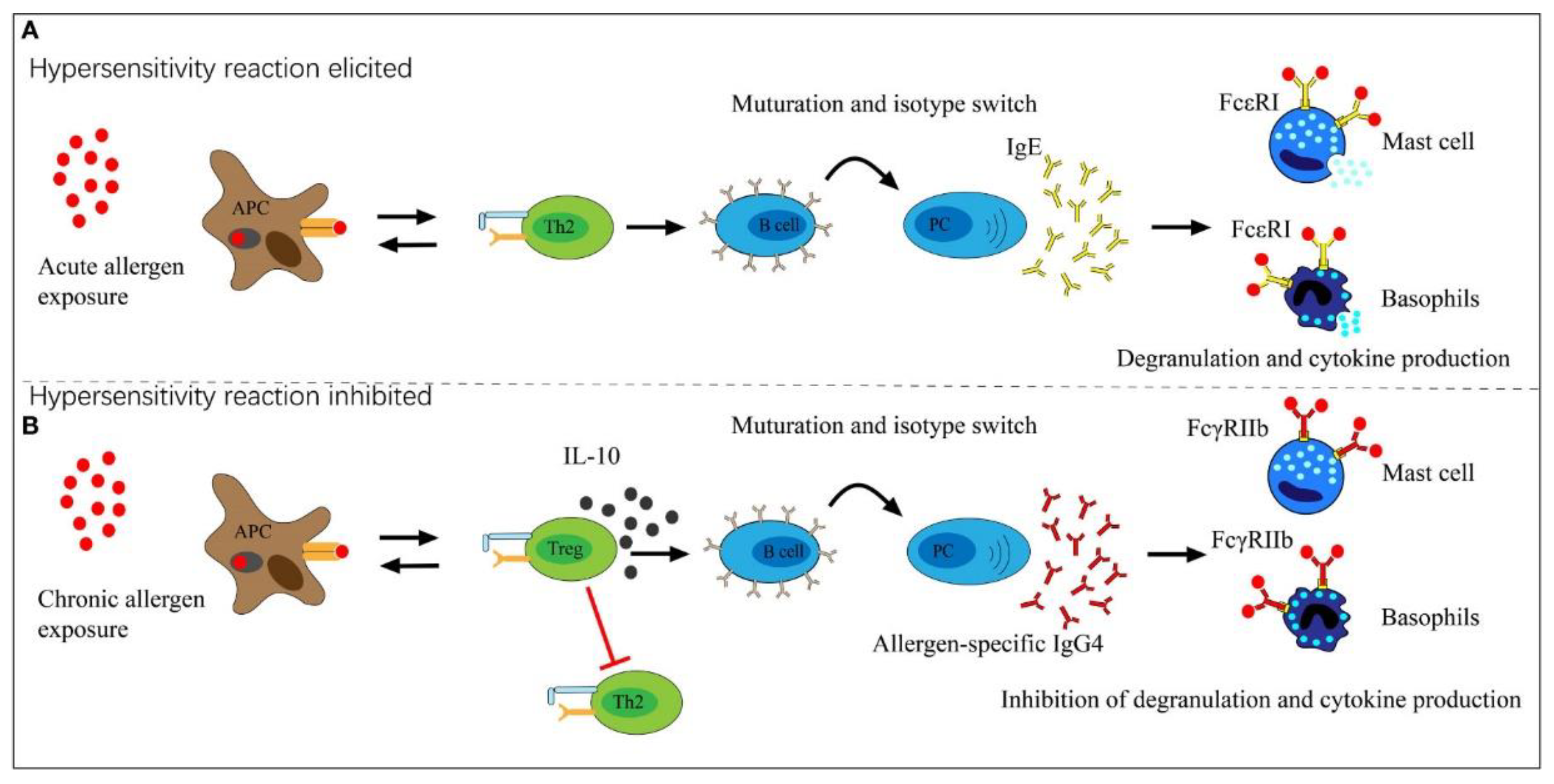
Figure 2.
Immunochemical staining of lung tissues. In the acute phase (ARDS and DAD) IgG4 was intensely expressed in lung tissue, reaching 80-90%, but was essentially absent during the stages of organization and fibrosis. ARDS (acute respiratory distress syndrome), DAD (diffuse alveolar damage). Modified from [87]. This is an open-access article and distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Figure 2.
Immunochemical staining of lung tissues. In the acute phase (ARDS and DAD) IgG4 was intensely expressed in lung tissue, reaching 80-90%, but was essentially absent during the stages of organization and fibrosis. ARDS (acute respiratory distress syndrome), DAD (diffuse alveolar damage). Modified from [87]. This is an open-access article and distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
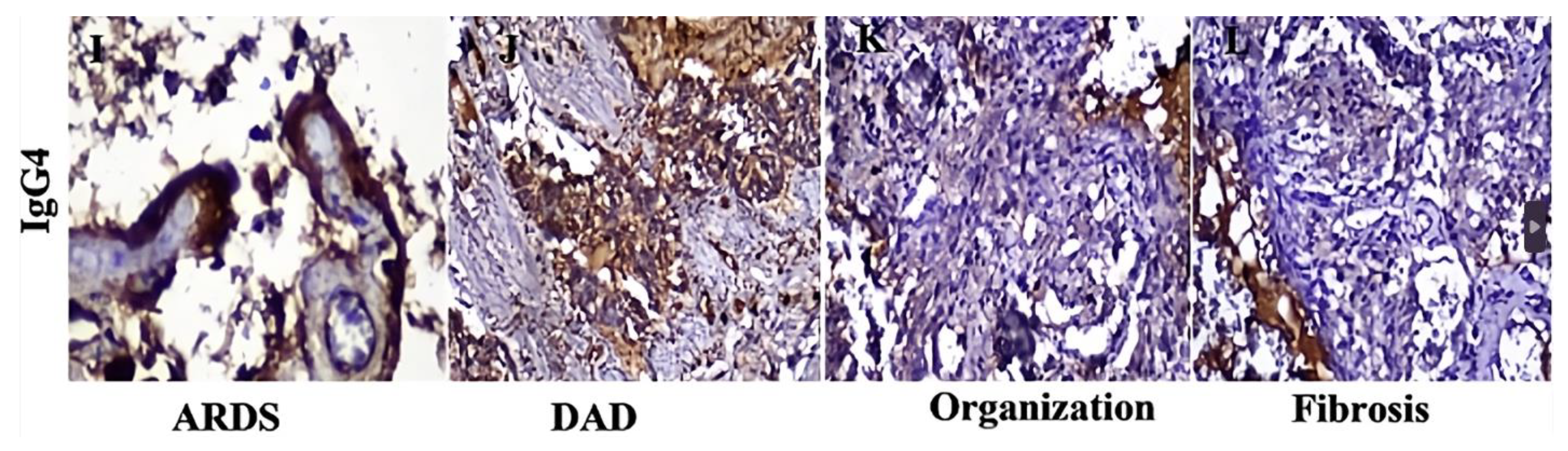
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



