
Submitted:
11 May 2025
Posted:
12 May 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Background/Objectives: Cases of the new COVID-19 coronavirus infection, which manifested in 2019 and caused a global socioeconomic crisis, still continue to be registered worldwide. The high mutational activity of SARS-CoV-2 leads to the emergence of new antigenic variants of the virus, which significantly reduces the effectiveness of COVID-19 vaccines, as well as the sensitivity of diagnostic test systems based on variable viral antigens. These problems may be solved by the focusing on highly conserved coronavirus antigens, for example nucleocapsid (N) protein, which is actively expressed by coronavirus-infected cells and serves as a target for the production of virus-specific antibodies and T-cell responses. It is known that anti-N antibodies are non-neutralizing, but their protective potential and functional activity are not sufficiently studied. Methods: Protective effect of anti-N antibodies was studied in Syrian hamsters passively immunized with polyclonal sera raised to N(B.1) recombinant protein. Animals were infected with 105 or 104 TCID50 of SARS-CoV-2 (B.1, Wuhan or BA.2.86.1.1.18, Omicron) 6 hours after serum passive transfer, and protection was assessed by weight loss, clinical manifestation of disease, viral titers in respiratory tract, as well as by histopathological evaluation of lung tissues. Functional activity of anti-N (B.1) antibodies was evaluated by complement-dependent cytotoxicity (CDC) and antibody-dependent cytotoxicity (ADCC) assays. Results: Protection of anti-N antibodies was evident only against lower dose of SARS-CoV-2 (B.1) challenge, whereas almost no protection was revealed against BA.2.86.1.1.18 variant. Anti-N (B.1) monoclonal antibodies were able to stimulate CDC and ADCC. Conclusions: Anti-N (B.1) antibodies possess protective activity against homologous challenge infection, which is possibly mediated by innate Fc-mediated immune reactions. These data may be informative for the development of N-based broadly-protective COVID-19 vaccines.
Keywords:
COVID-19 vaccine
; SARS-CoV-2
; Syrian hamster
; nucleocapsid protein
; antiviral antibodies
1. Introduction
A novel COVID-19 coronavirus infection caused by SARS-CoV-2 continues to pose severe socioeconomic burden in all countries of the world. Prevention of infection with new antigenic variants of SARS-CoV-2 using licensed vaccines and antiviral drugs based on monoclonal antibodies turned to be ineffective due to the constant evolutionary variability of the virus [1]. In this regard, an urgent task of public health care today is the development of new cross-protective vaccines based on highly conserved SARS-CoV-2 proteins, the protective effects of which does not depend on the variability of antigenic properties of the virus. One of such promising antigens is the nucleocapsid protein (N), which is significantly less variable than the surface-exposed spike protein (S) [2,3]. According to the literature data, N protein is actively produced in infected cells, including being detected on the cell surface during the progression of natural infection [4] and is highly immunogenic, inducing the formation of specific T-cells and antibodies [5]. The vast majority of studies consider N protein as a target for the induction of T-cell immune responses [6,7,8], whereas no convincing data on the ability of anti-N antibodies to protect against natural infection have been presented so far.
Although antibodies against N protein lack neutralizing activity [9], they are intensively produced [10], persist in the organism significantly longer than S-specific antibodies [11] and hypothetically may provide antiviral protection by their functional activity acting as activators of complement cascade reactions [12] or antibody-mediated cellular phagocytosis/cytotoxicity (ADCP/ADCC) [13]. In addition to natural infection, the production of N-specific antibodies can also be stimulated by immunization with whole-virion inactivated [14,15] or live attenuated [16] vaccines, but the mechanism of participation of these immunoglobulins in protection against COVID-19 remains largely unexplored. Moreover, there is evidence that antibodies to N protein may have autoimmune properties, provoking the development of various immunopathologic conditions [17].
Studying the protective role of anti-N antibodies is necessary both in light of prediction of infection course/outcomes and for the development of universal N-based vaccines against COVID-19. Here, we evaluated the antiviral potential of anti-N immunoglobulins against homologous and heterologous infections and studied a possible mechanism of this protection.
2. Materials and Methods
2.1. Cells, Virus and Sera
African green monkey kidney Vero CCL81 cells were purchased from the American Type Culture Collection (ATCC) and maintained in DMEM supplemented with 10% fetal bovine serum (FBS) and 1 × antibiotic–antimycotic (AA) (all from Capricorn Scientific, Ebsdorfergrund, Germany). SARS-CoV-2 viruses HCoV-19/Russia/StPetersburg-3524/2020 (B.1 Lineage, Wuhan) and hCoV-19/Russia/SPE-RII-9293S/2023 (BA.2.86.1.1.18 Lineage, Omicron) were obtained from the Smorodintsev Research Institute of Influenza (Saint Petersburg, Russia). They were propagated and titrated as previously described [18] in Vero CCL81 or Vero E6 cells, respectively, at MOI 0.01 using DMEM supplemented with 2% FBS, 10 mM of HEPES and 1 × AA (all from Capricorn Scientific, Ebsdorfergrund, Germany) at 37 °C and 5% CO2. As full cytopathic effect was achieved, the virus-containing media was harvested, clarified via centrifugation at 3000 rpm and stored at −70 °C in aliquots. All experiments with live SARS-CoV-2 were performed in a biosafety-level-3 (BSL3) laboratory.
2.2. Hamster Study Design
Golden Syrian hamsters (Mesocricetus auratus) were purchased from Stezar cattery (Vladimir region, Russia). The experimental design was approved by the local ethics committee of the Institute of Experimental Medicine (protocol no 4/24, dated 24 October 2024). The animals were maintained in standard laboratory vivarium conditions with free access to food and water. The study was conducted in accordance with Directive 2010/63/EU [19].
Animals aged 8 weeks and body weight 80-100 g (five animals per group) were pre-screened in ELISA on 2 µg/mL immobilized recombinant N (B.1) protein, then intraperitoneally immunized with 2 mL of anti-N(B.1) or control hamster serum previously inactivated for 1 hour at 56 °C and 1:1 diluted with PBS. Then, 6 hours later hamsters were infected intranasally under light ether anesthesia with HCoV-19/Russia/StPetersburg-3524/2020 (B.1, Wuhan) or hCoV-19/Russia/SPE-RII-9293S/2023 (BA.2.86.1.1.18, Omicron) viruses, at a dose of 104 or 105 TCID50. Clinical symptoms of the disease and body weight dynamics were monitored for 6 days after the challenge. The clinical features of the disease course were evaluated according to the following criteria: coat condition: 0 - normal, 1 - lack of care; interaction with other animals: 0 - normal, 1 - reduced; feed consumption: 0 - normal, 1 - reduced; behavior in the open area: 0 - active, 1 - reduced; reaction to being taken: 0 - normal, 1 - reduced.
On the 6th day, to assess the tissue levels of virus replication, animals were removed from the experiment by ether overdose and nasal passages and lungs were aseptically isolated. The tissues were weighed and homogenized with the steel beads in the Qiagen TissueLyser LT, followed by titration on Vero CCL81 (for B.1 virus) or Vero E6 cells (for BA.2.86.1.1.18 virus) with FFU counting by the previously described approach [20]. For this, 10−1 to 10−3 dilutions of clarified homogenates were used to infect 96-well cell monolayers in triplicates. After 20 h of incubation, virus-infected cells were formalin-fixed overnight (ON), then permeabilized and consistently treated with anti-N(B.1) biotynilated antibody and eGFP-streptavidin fusion protein. The FFU counting was performed using AID vSpot Spectrum (Autoimmun Diagnostika GmbH, Germany).
One lung lobe was intended for histological studies and fixed in 10% buffered formalin solution (pH = 7.4) for 48 hours. Then, routine histologic wiring was applied on a histoprocessor Histo-Tek VP1 (Sakura, Japan), followed by embedding of specimens in paraffin blocks. The 3 μm slices were stained either with hematoxylin and eosin solution according to a standard protocol. In addition, MSB-stain was used for elective staining for blood clots and vascular condition. With this staining, fibrin blood clots and red blood cells turn red, collagen adventitia becomes blue, whicle lymphocytes turn purple. These slides were analyzed using LEICA DM1000 microscope, while capturing and measurements were performed using ADF Image Capture 4.17 software package. Morphometric analyses were performed as earlier described [21], using alveolar wall thickness as the primary parameter for the assessment of damage to lung tissues.
2.3. Assessment of Functional Activity of Anti-N Antibodies
The ability of anti-N (B.1) antibodies to promote Fc-mediated innate immune reactions was assessed both by antibody-dependent natural killer (NK) cell degranulation assay, which is a surrogate assay for the assessment of antibody-dependent cellular cytotoxicity (ADCC) responses [22], and by complement-dependent cytotoxicity assay (CDC). In the first case, 2 µg/mL of recombinant N (B.1) protein in carbonate-bicarbonate buffer were used to coat high-sorbent 96-well plates (Corning, USA) at 4 °C overnight. Then, anti-N (B.1) mAbs (5 ug/mL) or serum samples (1:50) were added to the washed wells and incubated at 37 °C for 1 h. Then, 2 × 106 splenocytes collected from naïve C57BL/6J mice were added in 100 µL of CR-10 to each well and incubated at 37 °C and 5% CO2 for 24 h. On the next day, the cell suspensions were collected and treated with ZombieAqua fixable viability dye, anti-CD3 (clone 2E7), anti-CD49b (clone DX5), anti-CD45.2 (clone 104) and anti-CD107a (clone 1D4B) conjugated antibodies (Biolegend, San Diego, CA, USA) diluted in staining buffer (SB) (PBS supplemented with 0.2% BSA and 0.05% sodium azide), for 20 min at RT in the darkness. In this staining protocol, a transmembrane protein CD107a of cytolytic granules was targeted on the cell surface as a degranulation marker [23]. The staining procedure was followed by washing with 200 µL SB twice, fixation of the cells in 1% formaldehyde and flow cytometry analysis. At least 30,000 events were measured using a CytoFlex cytometer equipped with CytExpert software (Beckman Coulter, USA).
To perform the CDC assay, Vero CCL81 monolayers seeded the day before on 24-well plates were infected with a HCoV-19/Russia/StPetersburg-3524/2020 (B.1 Lineage, Wuhan) virus at MOI 0.01 and incubated in DMEM supplemented with 2% FBS and 1 x AA at 37 °C in 5% CO2 overnight. Then, the medium was removed and cells were washed with 300 µL PBS, followed by treatment with 10 µg of anti-N (B.1) mAbs or 1:2 sera diluted in DMEM at 37 °C and 5% CO2 for 15 min. Then, 50 µL of naïve guinea pig sera diluted 1:10 in DMEM were added as a complement source, and plates were further incubated for 3 h at 37 °C and 5% CO2. The well supernatants were transferred to the 2 mL tubes, and the remaining cell monolayers were washed twice with PBS prior to the accutase treatment for 10 min at 37 °C and 5% CO2. After the accutase inactivation by addition of 1 mL DMEM supplemented with 10% FBS, the cells were thoroughly resuspended and collected to the same tubes with corresponding supernatants. The tubes were centrifuged for 7 min, 300× g, washed with PBS twice, resuspended in 300 µL PBS, stained with propidium iodide and YO-PRO iodide (Thermo Fisher Scientific, USA) and analyzed using CytoFlex Cytometer. CDC induction was determined as an increase in the percent of cells in the late apoptotic phase.
2.4. Statistical Analysis
Data were processed using the statistical tool of GraphPad Prism 6.0 Software (GraphPad Software, San Diego, CA, USA). Compliance with the normal distribution was checked by the Shapiro-Wilk test. Differences between several test groups were analyzed by two-way ANOVA with Sidak’s multiple comparisons test. The significance level was set at p < 0.05.
3. Results
3.1. Protection Against SARS-CoV-2 Infection
For this study, we generated hyperimmune hamster sera against recombinant N(B.1) protein for subsequent use for serum passive transfer experiments. To collect it, we expressed the antigen in bacterial system (E. coli BL21(DE3) cells) using previously developed protocol [5] and obtained sufficient quantities for triple immunization of 5 hamsters with 100 µg of recombinant N (B.1) protein with a 14-day interval. The resulting titer of pooled anti-N(B.1) sera assessed in ELISA on N(B.1) was 1:7290. At the same time, the titer of naïve sera was also checked and appeared to be lower than ELISA sensitivity threshold.
The pre-screened naïve hamsters were immunized intraperitoneally with 2 mL of 0.22 µm filtered naïve or anti-N(B.1) sera diluted 1:1 in PBS and 6 h later challenged with 104 or 105 TCID50 of SARS-CoV-2 B.1 (Wuhan) or BA.2.86.1.1.18 (Omicron). The animals were weighted and monitored prior to the infection and then daily. The nasal washes were also collected just before the challenge (day 0) and then on day 2 and 4.
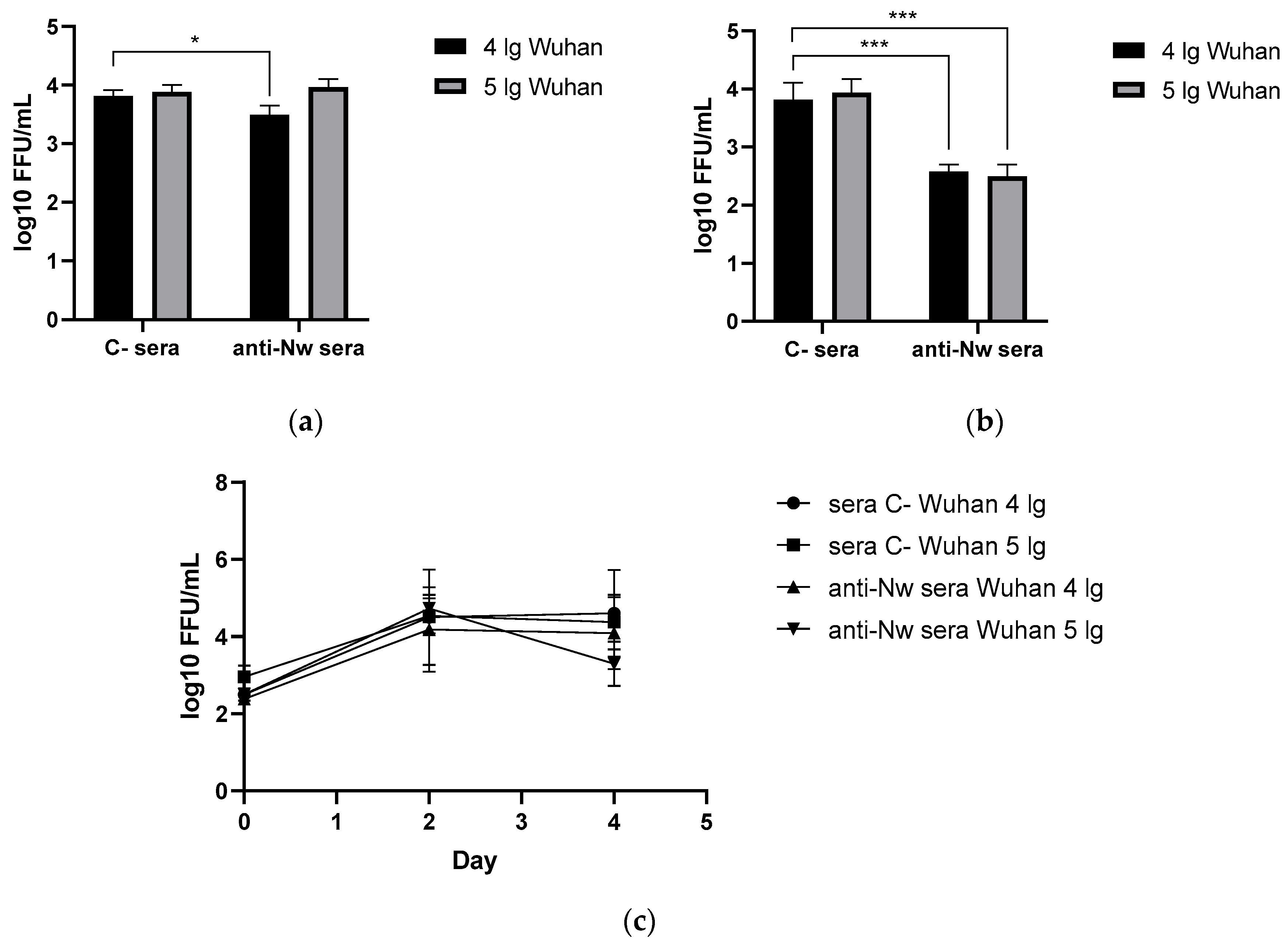
On the 6th day after challenge with Wuhan (B.1) variant, lungs and nasal turbinates were harvested from immunized and control hamsters, where SARS-CoV-2 (B.1) titers were determined by FFU counting in Vero CCL81. Surprisingly, anti-N (B.1) sera significantly reduced virus replication both in the upper and the lower respiratory tract if the challenge dose was 4 lg, while in case of 5 lg reduction in nasal and pulmonary viral titers compared to the mock group did not reach statistical significance. At the same time, no significant reduction of viral titers in the nasal washes of the immunized animals was revealed, which may be associated with the non-mucosal nature and intraperitoneal administration way of serum anti-N(B.1) immunoglobulins. Taken together, the data obtained confirm the ability of anti-N antibodies to form protective immunity against the homologous challenge virus at a low dose in the lower respiratory tract.
Figure 1.
Viral replication rates in animals immunized with control or anti-N (B.1) sera and challenged with Wuhan (B.1) virus : (a) in nasal turbinates; (b) in lung tissue; (c) in nasal washes. * - p < 0.05; *** - p < 0.001.
Figure 1.
Viral replication rates in animals immunized with control or anti-N (B.1) sera and challenged with Wuhan (B.1) virus : (a) in nasal turbinates; (b) in lung tissue; (c) in nasal washes. * - p < 0.05; *** - p < 0.001.
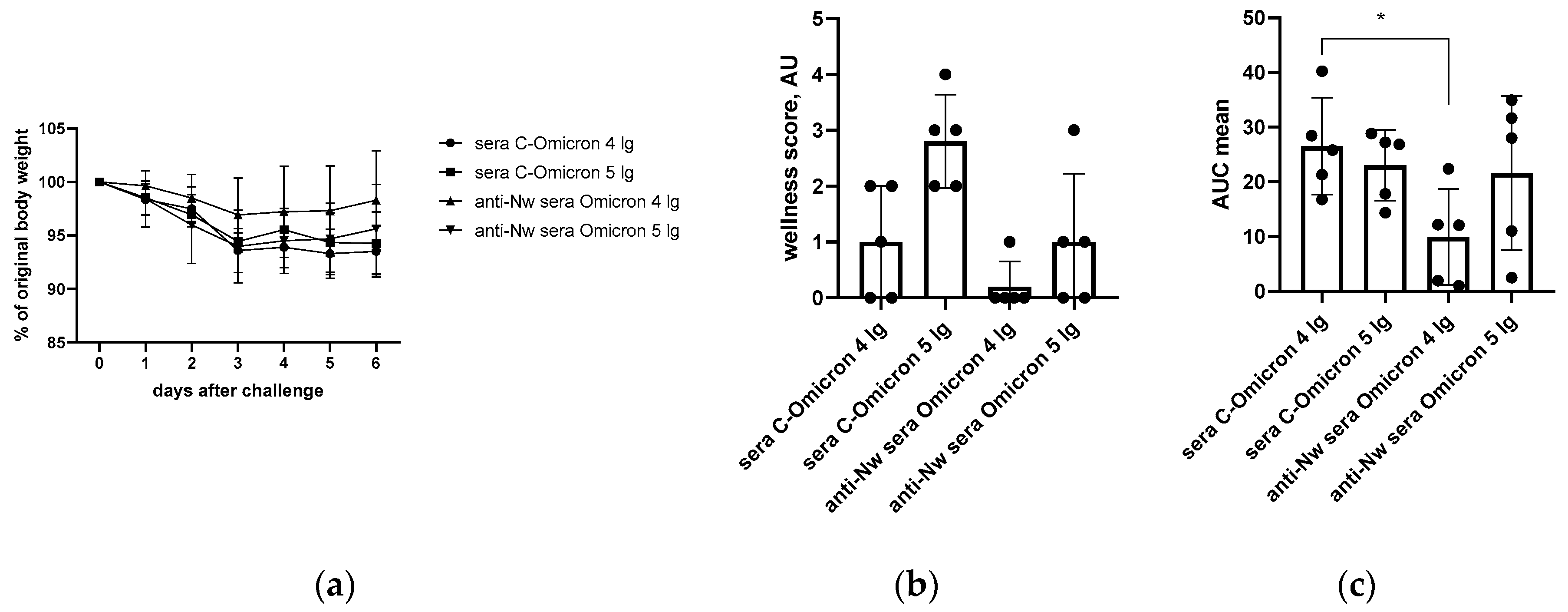
Body weight and wellness score monitoring conducted within 6 days after challenge revealed that anti-N (B.1)-immunized hamsters were less affected by the infection than animals that received the control sera (Figure 2b). As expected, only vaccinated hamsters challenged with 4 log of B.1 (Wuhan) were protected from the weight loss, while this effect was not observed at a higher challenge dose (Figure 2a).
In contrast to the results of B.1 challenge study, anti-N (B.1) sera appeared to be completely ineffective against the infection caused by BA.2.86.1.1.18 strain (Figure 3). These data are in line with the histopathology assessment of the H&E histological assessment of lung tissues collected from the SARS-CoV-2 (B.1)-challenged hamsters revealed that approximately 60% of the histological slides in the control group were characterized by more than 50% damage to lung and vascular tissues. Furthermore, this group had most severe alveolar inflammation with loss of normal septal histoarchitectonics (Figure 5, left panels). In addition, there was a more frequent desquamation of the necrotic bronchiolar epithelium into the bronchi lumen compared with the other control groups. In group immunized with anti-N (B.1) sera and challenged with 4 lg of B.1 virus, lung pathology was less pronounced compared to the control animals. In particular, alveoli were affected in a smaller percentage and at a lower severity. Bronchioles as well as vessels were affected mainly in the apical segments of the lungs. In animals which received anti-N (B.1) sera and 5 lg of SARS-CoV-2 (B.1), the lung histoarchitectonics were not comparable to that of the intact hamsters’ lungs, but this group was the least affected by B.1 (Wuhan) challenge.
Figure 3.
Viral replication rates in animals immunized with control or anti-N (B.1) sera and challenged with Omicron (BA.2.86.1.1.18) virus: (a) in nasal turbinates; (b) in lung tissue; (c) in nasal washes. * - p < 0.05.
Figure 3.
Viral replication rates in animals immunized with control or anti-N (B.1) sera and challenged with Omicron (BA.2.86.1.1.18) virus: (a) in nasal turbinates; (b) in lung tissue; (c) in nasal washes. * - p < 0.05.
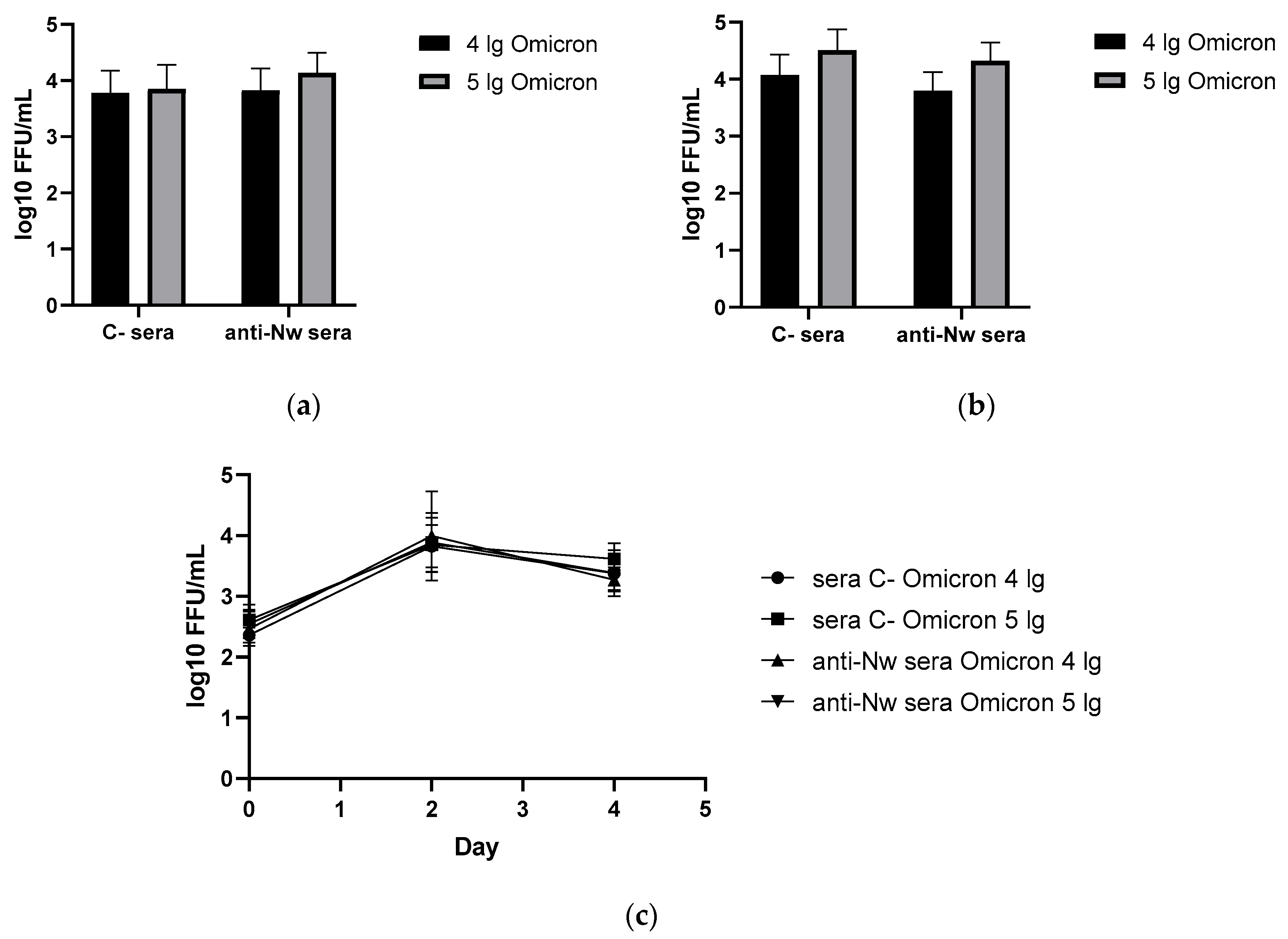
Figure 4.
The monitoring of performance of hamsters immunized with anti-N (B.1) or control sera after Omicron (BA.2.86.1.1.18) challenge: (a) individual body weight dynamics; (b) grouped wellness score; (c) AUCs of lost body weight.
Figure 4.
The monitoring of performance of hamsters immunized with anti-N (B.1) or control sera after Omicron (BA.2.86.1.1.18) challenge: (a) individual body weight dynamics; (b) grouped wellness score; (c) AUCs of lost body weight.
MSB staining was alternatively used for elective staining for blood clots and assessment of vascular condition (Figure 5, right panels). In this case, the fibrin blood clots and red blood cells dyed in red, while collagen adventitia is in blue and lymphocytes are in purple. This staining approach allowed to reveal that the condition of the lung parenchyma in naïve animals had no features, corresponded to the species norm of hamsters. There were no blood clots in the lumens of large vessels, and the wall thickness of the vessels was normal. At the same time, in the C-sera-immunized groups infected with 4 or 5 lg of B.1 (Wuhan) strain, we detected both a decrease in the airiness of the pulmonary parenchyma due to thickening of the alveolar walls and vascular damage - the presence of intraluminal thrombi, edema and thickening of the media, edema of the adventitia with progressed infiltration of lymphocytes (Figure 5a). The administration of anti-N(B.1) sera, especially in the group infected with 4 lg of the B.1 (Wuhan) virus, resulted in a protective effects such as the increase in the airiness of the lung parenchyma (by 40%) due to the lower thickness of the alveolar walls, as well as protection against vascular damage. In this group, the magnitude of perivasculitis was moderate compared to that of infection control. In case of challenge with 5 lg of the B.1 (Wuhan) virus, the effect of immunization was not so significant.
Figure 5.
Histopathological assessment of protective effect of anti-N (B.1) sera against Wuhan (B.1) (a) and Omicron (BA.2.86.1.1.18) (b) viruses in a hamster model on day 6 after challenge. Representative micrographs of hematoxylin–eosin or MSB-stained lung sections are shown using 200x and 400x magnifications, respectively.
Figure 5.
Histopathological assessment of protective effect of anti-N (B.1) sera against Wuhan (B.1) (a) and Omicron (BA.2.86.1.1.18) (b) viruses in a hamster model on day 6 after challenge. Representative micrographs of hematoxylin–eosin or MSB-stained lung sections are shown using 200x and 400x magnifications, respectively.
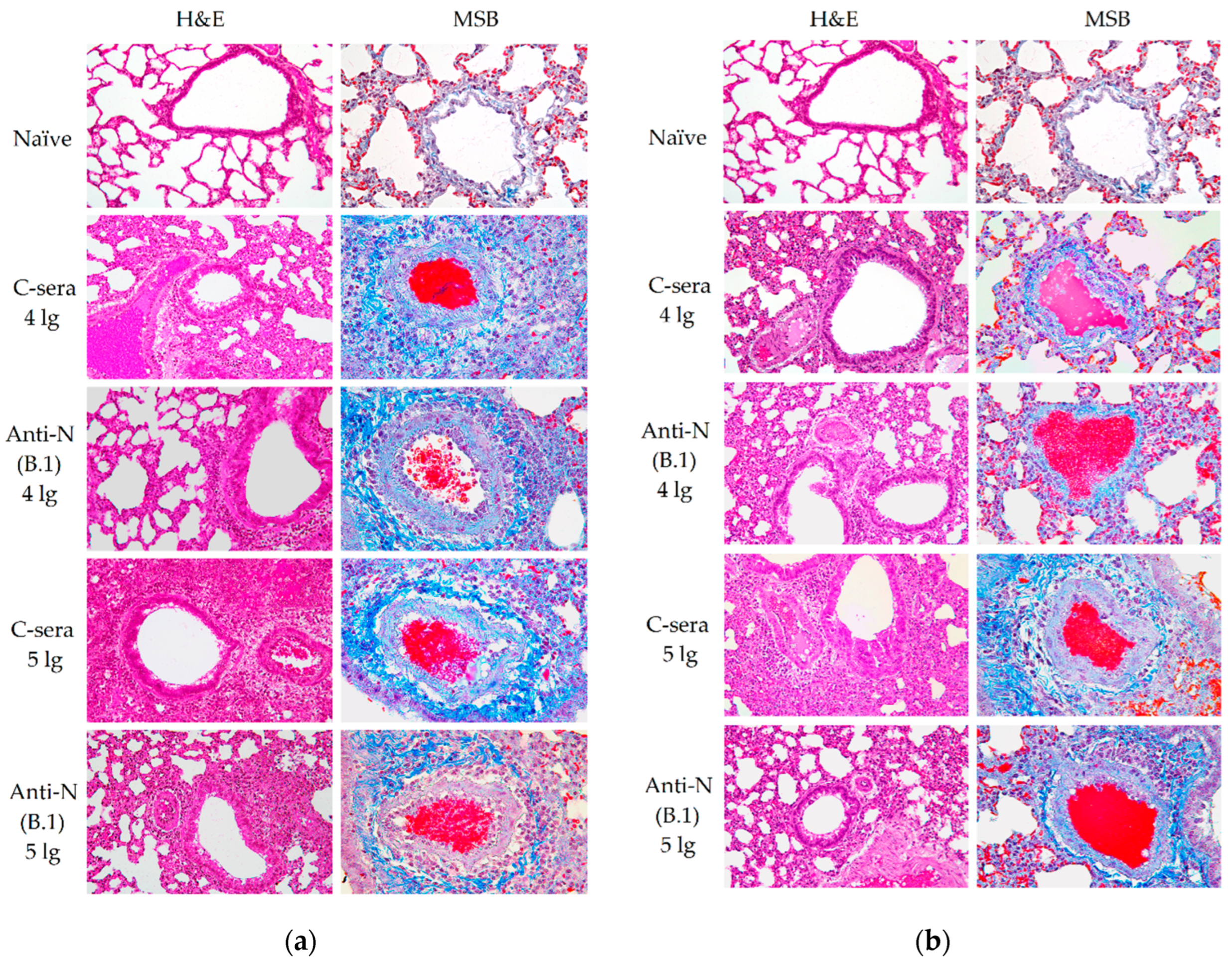
In a series of experiments with infection with the BA.2.86.1.1.18 (Omicron) strain (Figure 5b), the alveolar walls were less damaged in the hamsters immunized with C-sera, but there was a greater lesion of the perivascular space due to lymphocytic infiltration. It should be noted that the use of anti-N(B.1) sera led to an apparent increase in vascular damage and the development of perivasculitis.
We further calculated and compared the thickness of the alveolar septa in hamsters immunized with C- or anti-N(B.1) sera and challenged with Wuhan (B.1) or Omicron (BA.2.86.1.1.18) viruses (Figure 6). It was found that regardless of the infecting strain the values were significantly closer to those in the naïve animals in hamsters vaccinated with anti-N(B.1) sera, and thickness differences between animals which received C- or anti-N(B.1) sera were more pronounced in Wuhan (B.1) challenge study (Figure 6a).
3.2. Functional Activity of the N(B.1)-Specific Antibodies
In our previous experiments, no direct inhibition of SARS-CoV-2 propagation in Vero CCL81 by anti-N sera or monoclonal antibodies was revealed (Figure 7). This findings are in line with the well-known data on non-neutralizing properties of these antibodies [9].
As the direct neutralization of virus appeared to be ineffective, we further attempted to elucidate the mechanisms by which the defense by anti-N (B.1) antibodies is mediated. Due to the non-neutralizing nature and the obvious impact of the anti-N antibodies on the SARS-CoV-2-induced cross-protective potential, it is important to assess functional activity of these antibodies, i.e., Fc-driven cytotoxicity effects. Functional activity of the anti-N(B.1) sera and monoclonal antibodies was assessed in two assays—complement dependent cytotoxicity and an antibody-dependent natural killer (NK) degranulation activity, which served as a surrogate assay for evaluation of antibody-dependent cellular cytotoxicity (ADCC).
3.2.1. Complement Dependent Cytotoxicity (CDC)
The antiviral defense mechanisms that N-protein-based vaccines are aimed to trigger have not yet been definitively established. One of the actual questions is whether the complement system plays an important role in this protection. Previously obtained serum sample from COVID-19 convalescent and mice three times immunized with recombinant N (B.1) protein as well as anti-N monoclonal antibodies [24] were added to Vero CCL81 cells infected with HCoV-19/Russia/StPetersburg-3524/2020 (B.1 Lineage, Wuhan) virus, followed by incubation with guinea pig naïve sera (the complement source). This resulted in the increased late apoptosis of the infected cells in case of treatment with convalescent serum and NCL10 antibody (Figure 8a). The assay served as an indicator which is considered to correlate well with the CDC-mediating antibody activity [25].
Interestingly, serum anti-N (B.1) antibodies from immunized mice demonstrated no CDC activity, compared to monoclonal antibodies NCL10, and in the last case, the percent of induced late apoptotic cells was comparable to those increased by the addition of convalescent sera. Since the hamsters passively immunized with anti-N (B.1) sera had significantly reduced viral pulmonary titers six days after challenge compared to the control group, the promotion of CDC response by the convalescent sera and anti-N monoclonal antibodies suggests that complement system may play an important role in reducing SARS-CoV-2 propagation in the lungs.
3.2.2. Antibody-Dependent Cellular Cytotoxicity (ADCC)
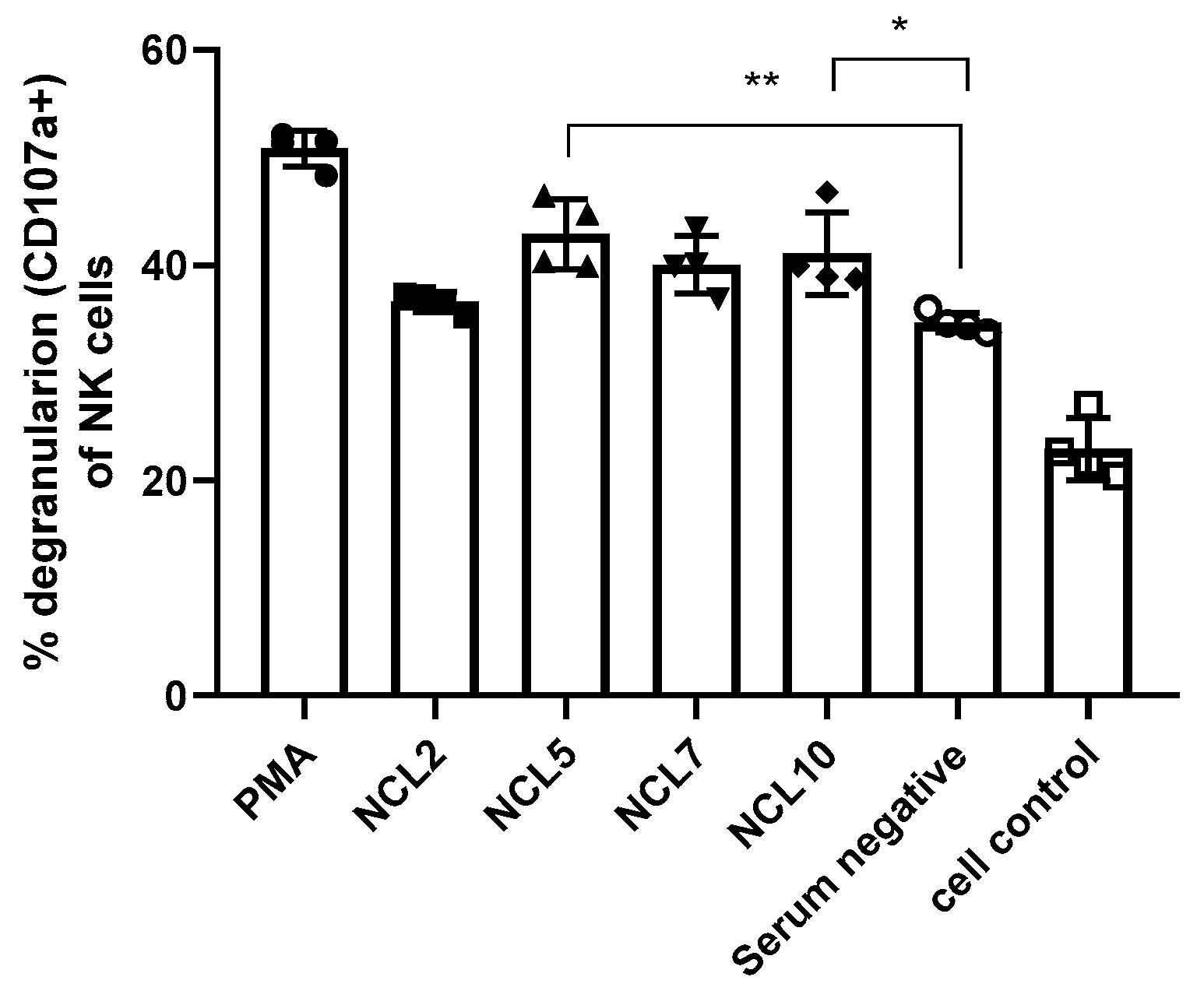
Due to the lack of fluorescently labeled antibodies to the surface markers of Syrian hamsters, possible ADCC activity of N-specific antibodies was assessed for the previously generated mouse monoclonal antibodies NCL2, NCL5, NCL7 and NCL10. A stimulation of antibody-dependent cellular cytotoxicity was revealed for two of the four mAbs, NCL5 and NCL10, as evidenced by the level of NK cell degranulation induced by incubation of these antibodies with splenocytes of naïve C57BL/6J mice as a source of NK cells (Figure 9). These results suggest that the ADCC mechanism may play some role in the protection from SARS-CoV-2 infection provided by anti-N antibodies, although these findings are not directly transferable to the results of evaluating the protective activity of antibodies in hamster experiments.
4. Discussion
Despite the official end of the COVID-19 pandemic declared by WHO in May 2023 [26], this disease continues to cause significant socio-economic damage to the global community. Effective control of the spread of this infection is impossible without understanding the molecular mechanisms of its progression, and in the absence of highly specific etiotropic therapeutics and vaccine prophylaxis for COVID-19. Although the use of first-generation vaccines was able to contain the spread of the original SARS-CoV-2 strain and minimize the impact of the early phase of COVID-19 pandemic [27,28], they targeted predominantly the coronavirus spike protein, which has led to the global spread of SARS-CoV-2 antigenic variants and a significant reduction in vaccine efficacy [29,30].
Due to the considerable variability of coronavirus surface antigens, the efforts of researchers have been focused on creating a universal vaccines against COVID-19 based on the most conserved components of SARS-CoV-2 [31]. One of them is the N protein of coronavirus, the main component of virions actively synthesizing by infected cells and causing intensive production of antiviral antibodies [32]. The N antigen also serves as one of the main targets of antiviral T-cell immune responses [8], which makes it a promising basis for the development of universal recombinant vaccines against COVID-19 [2,3].
Despite the general characterization of the properties of anti-N antibodies produced in response to immunization or during coronavirus infection, their protective activity still remains poorly understood. Some early studies described the ability of monoclonal antibodies specific to the N-protein of murine hepatitis coronavirus to protect animals from wild-type challenge [33]. In addition, the efficacy of prior passive immunization with monoclonal anti-N antibodies or serum from animals immunized with the N protein of SARS-CoV-2 to mitigate the effects of infection was previously demonstrated by Dangi et al. [34]. N-specific antibodies are known to be non-neutralizing [9], and the ability to trigger ADCC has been shown including for antibodies against SARS-CoV-2 that do not have neutralizing activity [35,36,37]. Moreover, there is evidence for preferential activation of NK cells by antibodies against SARS-CoV-2 antigens other than the Spike protein [38]. Antibodies generated in response to COVID-19 infection were found to be able to induce ADCC significantly more actively compared to antibodies generated in response to vaccination [39], and this fact also indicates a possible role for anti-N antibodies in mediating the innate cell protective responses. The direct possibility of induction of ADCC and ADCP reactions by anti-N antibodies from plasma of COVID-19 convalescents was recently shown by Díez et al. [40] and Hagemann et al. [41]. In addition, the ability of some monoclonal N-specific antibodies against SARS-CoV-2 [12] and anti-N antibodies generated in COVID-19 convalescents [42] to induce the CDC initiation has been demonstrated. Here, we also found that anti-N(B.1) monoclonal antibodies were able to provoke CDC, and some of them had the ADCC-stimulating ability.
Moreover, our results clearly indicate the protective properties of anti-N (B.1) antibodies against homologous challenge, and the inability of sera against N (B.1) to inhibit the infection caused by evolutionary distant BA.2.86.1.1.18 variant, which is in line with the previously obtained data on the restricted specificity of anti-N (B.1) sera to the recombinant N protein of more recent variants of the coronavirus [24]. The combination of these facts suggests that antiviral antibody generation alone is insufficient for protection and that induction of T-cell immune responses is necessary for elimination of infection.
The fact of involvement of N-specific antibodies in the realization of innate immunity reactions makes the development of N-based vaccines very promising, but requiring a careful study in light of the reported autoreactivity of anti-N immunoglobulins. The evaluation of this phenomenon is mandatory to assess the risk of autoimmune pathologies, which may be due to partial specificity of anti-N antibodies to antigens of the human and should be considered as an undesirable effect of their generation caused by vaccination or natural infection [17,43]. In light of innate protection mechanisms triggered by N protein, further studies should be focused on the evaluating the feasibility of vaccination that induces the production of anti-N antibodies versus the risk of developing autoimmune pathologies.
5. Conclusions
Our findings suggest that anti-N antibodies, while not possessing direct neutralizing activity, can provide protection against homologous SARS-CoV-2 infection, presumably by activating innate immunity responses such as CDC and ADCC reactions. The obtained results can be used to predict possible consequences of coronaviral infection, as well as for the further development of N-based vaccines against COVID-19. Further studies are needed to clarify the type of triggered defense reactions and to elucidate the mechanisms of Fc-effector activation.
Author Contributions
Conceptualization, A.R. and I.I.-S.; methodology, A.R., V.M. and I.I.-S.; software, A.R. and I.I.-S.; validation, A.R. and E.B.; formal analysis, A.R.; investigation, A.R., E.B., V.M., A.K. and Y.O.; resources, A.R. and K.S.; data curation, A.R., V.M. and Y.O.; writing—original draft preparation, A.R. and K.S.; writing—review and editing, A.R. and I.I.-S.; visualization, A.R., Y.O.; supervision, I.I.-S.; project administration, I.I.-S.; funding acquisition, A.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the RSCF grant number 24-75-00010.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of the Institute of Experimental Medicine (protocol № 4/24, dated 24 October 2024).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Senevirathne, T.H.; Wekking, D.; Swain, J.W.R.; Solinas, C.; De Silva, P. COVID-19: From emerging variants to vaccination. Cytokine & growth factor reviews 2024, 76, 127–141. [Google Scholar] [CrossRef]
- Oronsky, B.; Larson, C.; Caroen, S.; Hedjran, F.; Sanchez, A.; Prokopenko, E.; Reid, T. Nucleocapsid as a next-generation COVID-19 vaccine candidate. International journal of infectious diseases : IJID : official publication of the International Society for Infectious Diseases 2022, 122, 529–530. [Google Scholar] [CrossRef]
- Dutta, N.K.; Mazumdar, K.; Gordy, J.T. The Nucleocapsid Protein of SARS-CoV-2: a Target for Vaccine Development. Journal of virology 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- López-Muñoz, A.D.; Yewdell, J.W. Cell surface RNA virus nucleocapsid proteins: a viral strategy for immunosuppression? npj Viruses 2024, 2, 41. [Google Scholar] [CrossRef] [PubMed]
- Rak, A.; Donina, S.; Zabrodskaya, Y.; Rudenko, L.; Isakova-Sivak, I. Cross-Reactivity of SARS-CoV-2 Nucleocapsid-Binding Antibodies and Its Implication for COVID-19 Serology Tests. Viruses 2022, 14, 2041. [Google Scholar] [CrossRef]
- Grifoni, A.; Sidney, J.; Vita, R.; Peters, B.; Crotty, S.; Weiskopf, D.; Sette, A. SARS-CoV-2 human T cell epitopes: Adaptive immune response against COVID-19. Cell host & microbe 2021, 29, 1076–1092. [Google Scholar] [CrossRef]
- Sette, A.; Sidney, J.; Crotty, S. T Cell Responses to SARS-CoV-2. Annual review of immunology 2023, 41, 343–373. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef]
- Wajnberg, A.; Amanat, F.; Firpo, A.; Altman, D.R.; Bailey, M.J.; Mansour, M.; McMahon, M.; Meade, P.; Mendu, D.R.; Muellers, K.; et al. Robust neutralizing antibodies to SARS-CoV-2 infection persist for months. Science 2020, 370, 1227–1230. [Google Scholar] [CrossRef]
- Movsisyan, M.; Truzyan, N.; Kasparova, I.; Chopikyan, A.; Sawaqed, R.; Bedross, A.; Sukiasyan, M.; Dilbaryan, K.; Shariff, S.; Kantawala, B.; et al. Tracking the evolution of anti-SARS-CoV-2 antibodies and long-term humoral immunity within 2 years after COVID-19 infection. Scientific reports 2024, 14, 13417. [Google Scholar] [CrossRef]
- Koerber, N.; Priller, A.; Yazici, S.; Bauer, T.; Cheng, C.C.; Mijocevic, H.; Wintersteller, H.; Jeske, S.; Vogel, E.; Feuerherd, M.; et al. Dynamics of spike-and nucleocapsid specific immunity during long-term follow-up and vaccination of SARS-CoV-2 convalescents. Nature communications 2022, 13, 153. [Google Scholar] [CrossRef]
- Kang, S.; Yang, M.; He, S.; Wang, Y.; Chen, X.; Chen, Y.Q.; Hong, Z.; Liu, J.; Jiang, G.; Chen, Q.; et al. A SARS-CoV-2 antibody curbs viral nucleocapsid protein-induced complement hyperactivation. Nature communications 2021, 12, 2697. [Google Scholar] [CrossRef] [PubMed]
- Caddy, S.L.; Vaysburd, M.; Papa, G.; Wing, M.; O'Connell, K.; Stoycheva, D.; Foss, S.; Terje Andersen, J.; Oxenius, A.; James, L.C. Viral nucleoprotein antibodies activate TRIM21 and induce T cell immunity. The EMBO journal 2021, 40, e106228. [Google Scholar] [CrossRef]
- Dinc, H.O.; Saltoglu, N.; Can, G.; Balkan, II; Budak, B.; Ozbey, D.; Caglar, B.; Karaali, R.; Mete, B.; Tuyji Tok, Y.; et al. Inactive SARS-CoV-2 vaccine generates high antibody responses in healthcare workers with and without prior infection. Vaccine 2022, 40, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ning, J.; Chen, Y.; Li, B.; Shi, L.; He, T.; Zhang, F.; Chen, X.; Zhai, A.; Wu, C. The BBIBP-CorV inactivated COVID-19 vaccine induces robust and persistent humoral responses to SARS-CoV-2 nucleocapsid, besides spike protein in healthy adults. Frontiers in microbiology 2022, 13, 1008420. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Liu, J.; Xia, H.; Zou, J.; Muruato, A.E.; Periasamy, S.; Kurhade, C.; Plante, J.A.; Bopp, N.E.; et al. A live-attenuated SARS-CoV-2 vaccine candidate with accessory protein deletions. Nature communications 2022, 13, 4337. [Google Scholar] [CrossRef]
- Matyushkina, D.; Shokina, V.; Tikhonova, P.; Manuvera, V.; Shirokov, D.; Kharlampieva, D.; Lazarev, V.; Varizhuk, A.; Vedekhina, T.; Pavlenko, A.; et al. Autoimmune Effect of Antibodies against the SARS-CoV-2 Nucleoprotein. Viruses 2022, 14, 1141. [Google Scholar] [CrossRef]
- Matyushenko, V.; Isakova-Sivak, I.; Kudryavtsev, I.; Goshina, A.; Chistyakova, A.; Stepanova, E.; Prokopenko, P.; Sychev, I.; Rudenko, L. Detection of IFNgamma-Secreting CD4(+) and CD8(+) Memory T Cells in COVID-19 Convalescents after Stimulation of Peripheral Blood Mononuclear Cells with Live SARS-CoV-2. Viruses 2021, 13, 1490. [Google Scholar] [CrossRef]
- Anonimous. Directive 2010/63/EU of the European parliament and of the council of September 22, 2010, on the protection of animals used for scientific purposes. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32010L0063.
- Rak, A.; Matyushenko, V.; Prokopenko, P.; Kostromitina, A.; Polyakov, D.; Sokolov, A.; Rudenko, L.; Isakova-Sivak, I. A novel immunofluorescent test system for SARS-CoV-2 detection in infected cells. PloS one 2024, 19, e0304534. [Google Scholar] [CrossRef]
- Yakovlev, K.S.; Mezhenskaya, D.А.; Sivak, K.V.; Rudenko, L.G.; Isakova-Sivak, I.N. Comparative study of the pathogenicity of SARS-CoV-2 B.1 AND B.1.617.2 lineages for syrian hamsters. Medical academic journal 2022, 22, 125–136. [Google Scholar] [CrossRef]
- Morrison, B.J.; Roman, J.A.; Luke, T.C.; Nagabhushana, N.; Raviprakash, K.; Williams, M.; Sun, P. Antibody-dependent NK cell degranulation as a marker for assessing antibody-dependent cytotoxicity against pandemic 2009 influenza A(H1N1) infection in human plasma and influenza-vaccinated transchromosomic bovine intravenous immunoglobulin therapy. Journal of virological methods 2017, 248, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Betts, M.R.; Price, D.A.; Brenchley, J.M.; Lore, K.; Guenaga, F.J.; Smed-Sorensen, A.; Ambrozak, D.R.; Migueles, S.A.; Connors, M.; Roederer, M.; et al. The functional profile of primary human antiviral CD8+ T cell effector activity is dictated by cognate peptide concentration. Journal of immunology 2004, 172, 6407–6417. [Google Scholar] [CrossRef] [PubMed]
- Rak, A.; Gorbunov, N.; Kostevich, V.; Sokolov, A.; Prokopenko, P.; Rudenko, L.; Isakova-Sivak, I. Assessment of Immunogenic and Antigenic Properties of Recombinant Nucleocapsid Proteins of Five SARS-CoV-2 Variants in a Mouse Model. Viruses 2023, 15, 230. [Google Scholar] [CrossRef]
- Duensing, T.D.; Watson, S.R. Complement-Dependent Cytotoxicity Assay. Cold Spring Harbor protocols 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Sarker, R.; Roknuzzaman, A.S.M.; Nazmunnahar; Shahriar, M.; Hossain, M.J.; Islam, M.R. The WHO has declared the end of pandemic phase of COVID-19: Way to come back in the normal life. Health science reports 2023, 6, e1544. [Google Scholar] [CrossRef]
- Das, K.; Behera, R.L.; Paital, B. CHAPTER 8 - Socio-economic impact of COVID-19. In COVID-19 in the Environment; Rawtani, D., Hussain, C.M., Khatri, N., Eds.; Elsevier, 2022; pp. 153–190. [Google Scholar] [CrossRef]
- Watson, O.J.; Barnsley, G.; Toor, J.; Hogan, A.B.; Winskill, P.; Ghani, A.C. Global impact of the first year of COVID-19 vaccination: a mathematical modelling study. The Lancet Infectious Diseases 2022, 22, 1293–1302. [Google Scholar] [CrossRef]
- Ferdinands, J.M.; Rao, S.; Dixon, B.E.; Mitchell, P.K.; DeSilva, M.B.; Irving, S.A.; Lewis, N.; Natarajan, K.; Stenehjem, E.; Grannis, S.J.; et al. Waning of vaccine effectiveness against moderate and severe covid-19 among adults in the US from the VISION network: test negative, case-control study. Bmj 2022, 379, e072141. [Google Scholar] [CrossRef] [PubMed]
- Sobczak, M.; Pawliczak, R. COVID-19 vaccination efficacy in numbers including SARS-CoV-2 variants and age comparison: a meta-analysis of randomized clinical trials. Annals of clinical microbiology and antimicrobials 2022, 21, 32. [Google Scholar] [CrossRef]
- Bai, Z.; Cao, Y.; Liu, W.; Li, J. The SARS-CoV-2 Nucleocapsid Protein and Its Role in Viral Structure, Biological Functions, and a Potential Target for Drug or Vaccine Mitigation. Viruses 2021, 13, 1115. [Google Scholar] [CrossRef]
- Cubuk, J.; Alston, J.J.; Incicco, J.J.; Singh, S.; Stuchell-Brereton, M.D.; Ward, M.D.; Zimmerman, M.I.; Vithani, N.; Griffith, D.; Wagoner, J.A.; et al. The SARS-CoV-2 nucleocapsid protein is dynamic, disordered, and phase separates with RNA. Nature communications 2021, 12, 1936. [Google Scholar] [CrossRef]
- Nakanaga, K.; Yamanouchi, K.; Fujiwara, K. Protective effect of monoclonal antibodies on lethal mouse hepatitis virus infection in mice. Journal of virology 1986, 59, 168–171. [Google Scholar] [CrossRef]
- Dangi, T.; Class, J.; Palacio, N.; Richner, J.M.; Penaloza MacMaster, P. Combining spike- and nucleocapsid-based vaccines improves distal control of SARS-CoV-2. Cell reports 2021, 36, 109664. [Google Scholar] [CrossRef]
- Tso, F.Y.; Lidenge, S.J.; Poppe, L.K.; Pena, P.B.; Privatt, S.R.; Bennett, S.J.; Ngowi, J.R.; Mwaiselage, J.; Belshan, M.; Siedlik, J.A.; et al. Presence of antibody-dependent cellular cytotoxicity (ADCC) against SARS-CoV-2 in COVID-19 plasma. PloS one 2021, 16, e0247640. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wang, M.; Zhang, X.; Li, S.; Lu, Q.; Zeng, H.; Hou, H.; Li, H.; Zhang, M.; Jiang, F.; et al. Antibody-dependent cellular cytotoxicity response to SARS-CoV-2 in COVID-19 patients. Signal transduction and targeted therapy 2021, 6, 346. [Google Scholar] [CrossRef]
- Bahnan, W.; Wrighton, S.; Sundwall, M.; Blackberg, A.; Larsson, O.; Hoglund, U.; Khakzad, H.; Godzwon, M.; Walle, M.; Elder, E.; et al. Spike-Dependent Opsonization Indicates Both Dose-Dependent Inhibition of Phagocytosis and That Non-Neutralizing Antibodies Can Confer Protection to SARS-CoV-2. Frontiers in immunology 2021, 12, 808932. [Google Scholar] [CrossRef]
- Fielding, C.A.; Sabberwal, P.; Williamson, J.C.; Greenwood, E.J.D.; Crozier, T.W.M.; Zelek, W.; Seow, J.; Graham, C.; Huettner, I.; Edgeworth, J.D.; et al. SARS-CoV-2 host-shutoff impacts innate NK cell functions, but antibody-dependent NK activity is strongly activated through non-spike antibodies. eLife 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Rieke, G.J.; van Bremen, K.; Bischoff, J.; ToVinh, M.; Monin, M.B.; Schlabe, S.; Raabe, J.; Kaiser, K.M.; Finnemann, C.; Odainic, A.; et al. Natural Killer Cell-Mediated Antibody-Dependent Cellular Cytotoxicity Against SARS-CoV-2 After Natural Infection Is More Potent Than After Vaccination. The Journal of infectious diseases 2022, 225, 1688–1693. [Google Scholar] [CrossRef] [PubMed]
- Diez, J.M.; Romero, C.; Cruz, M.; Vandeberg, P.; Merritt, W.K.; Pradenas, E.; Trinite, B.; Blanco, J.; Clotet, B.; Willis, T.; et al. Anti-Severe Acute Respiratory Syndrome Coronavirus 2 Hyperimmune Immunoglobulin Demonstrates Potent Neutralization and Antibody-Dependent Cellular Cytotoxicity and Phagocytosis Through N and S Proteins. The Journal of infectious diseases 2022, 225, 938–946. [Google Scholar] [CrossRef]
- Hagemann, K.; Riecken, K.; Jung, J.M.; Hildebrandt, H.; Menzel, S.; Bunders, M.J.; Fehse, B.; Koch-Nolte, F.; Heinrich, F.; Peine, S.; et al. Natural killer cell-mediated ADCC in SARS-CoV-2-infected individuals and vaccine recipients. European journal of immunology 2022, 52, 1297–1307. [Google Scholar] [CrossRef]
- Lamerton, R.E.; Marcial-Juarez, E.; Faustini, S.E.; Perez-Toledo, M.; Goodall, M.; Jossi, S.E.; Newby, M.L.; Chapple, I.; Dietrich, T.; Veenith, T.; et al. SARS-CoV-2 Spike- and Nucleoprotein-Specific Antibodies Induced After Vaccination or Infection Promote Classical Complement Activation. Frontiers in immunology 2022, 13, 838780. [Google Scholar] [CrossRef]
- Vojdani, A.; Vojdani, E.; Kharrazian, D. Reaction of Human Monoclonal Antibodies to SARS-CoV-2 Proteins With Tissue Antigens: Implications for Autoimmune Diseases. Frontiers in immunology 2020, 11, 617089. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
The monitoring of performance of hamsters immunized with anti-N (B.1) or control sera after B.1 (Wuhan) challenge: (a) individual body weight dynamics; (b) AUCs of weight loss; (c) cumulative mean wellness score at day 6 after the challenge.
Figure 2.
The monitoring of performance of hamsters immunized with anti-N (B.1) or control sera after B.1 (Wuhan) challenge: (a) individual body weight dynamics; (b) AUCs of weight loss; (c) cumulative mean wellness score at day 6 after the challenge.
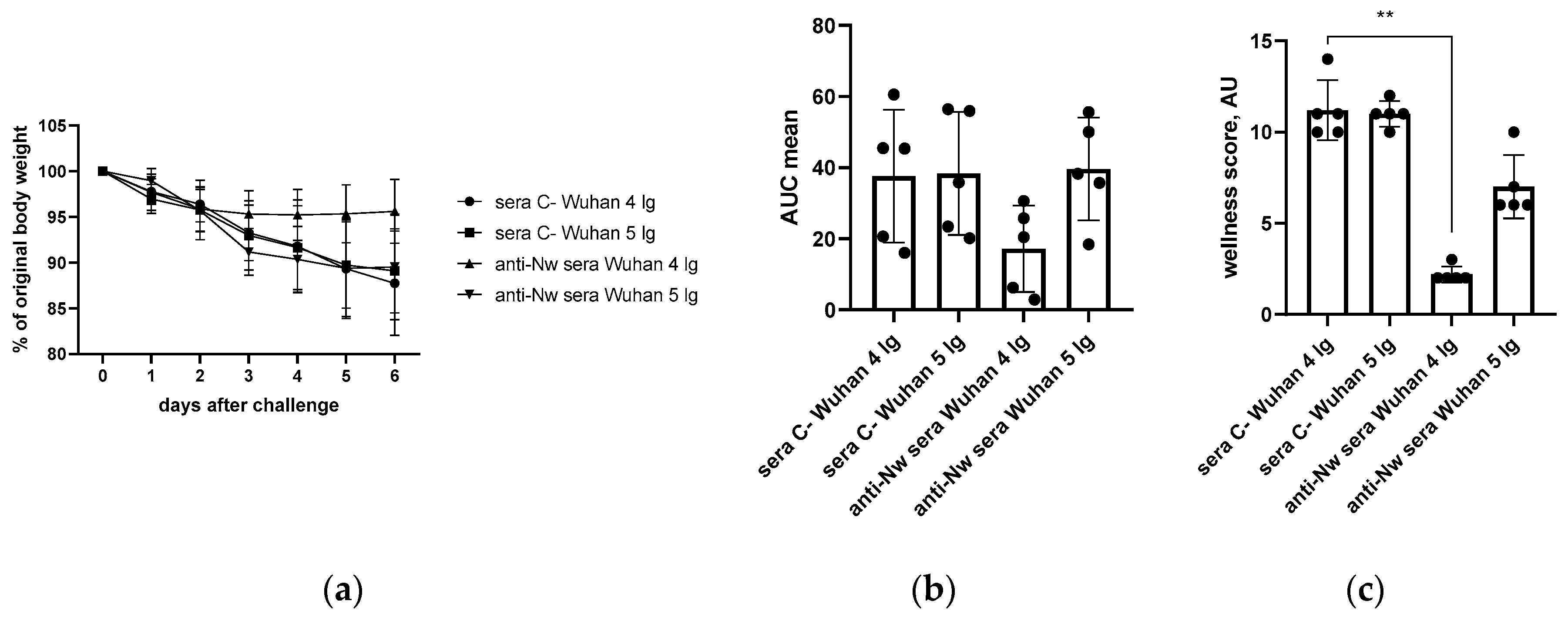
Figure 6.
Analysis of changes in alveolar septum thickness in naïve or immunized hamsters as a result of challenge infection with Wuhan (B.1) (a) or Omicron (BA.2.86.1.1.18) (b) viruses. The calculations are performed at 200x magnification.
Figure 6.
Analysis of changes in alveolar septum thickness in naïve or immunized hamsters as a result of challenge infection with Wuhan (B.1) (a) or Omicron (BA.2.86.1.1.18) (b) viruses. The calculations are performed at 200x magnification.
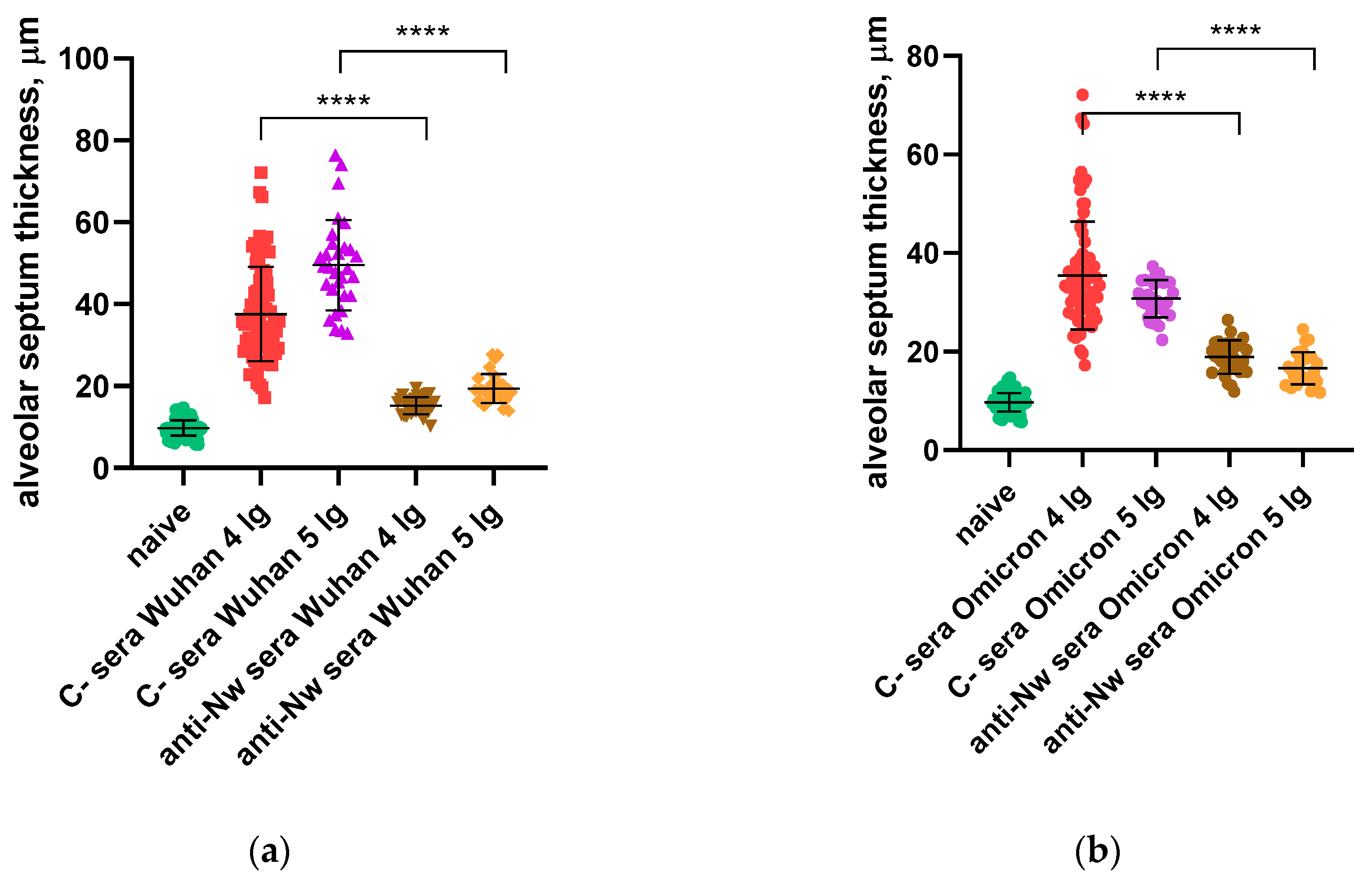
Figure 7.
SARS-CoV-2 (B.1) neutralization assay by mouse monoclonal anti-N antibodies (NCLs) and anti-RBD antibody at MOI 0.01.
Figure 7.
SARS-CoV-2 (B.1) neutralization assay by mouse monoclonal anti-N antibodies (NCLs) and anti-RBD antibody at MOI 0.01.
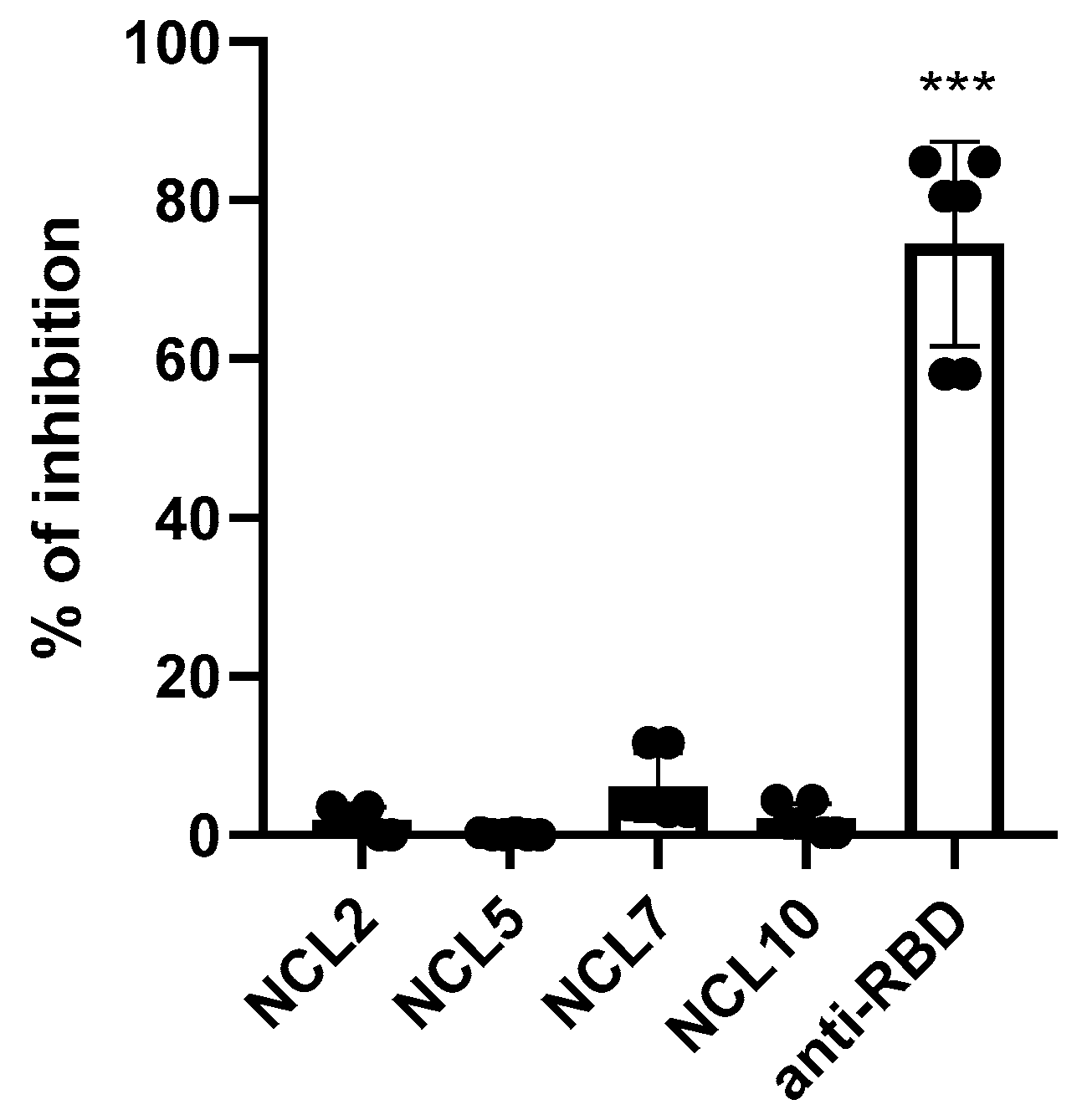
Figure 8.
Analysis of the functional activity of anti-N antibodies: (a) CDC induction; (b) representative CDC histograms.
Figure 8.
Analysis of the functional activity of anti-N antibodies: (a) CDC induction; (b) representative CDC histograms.
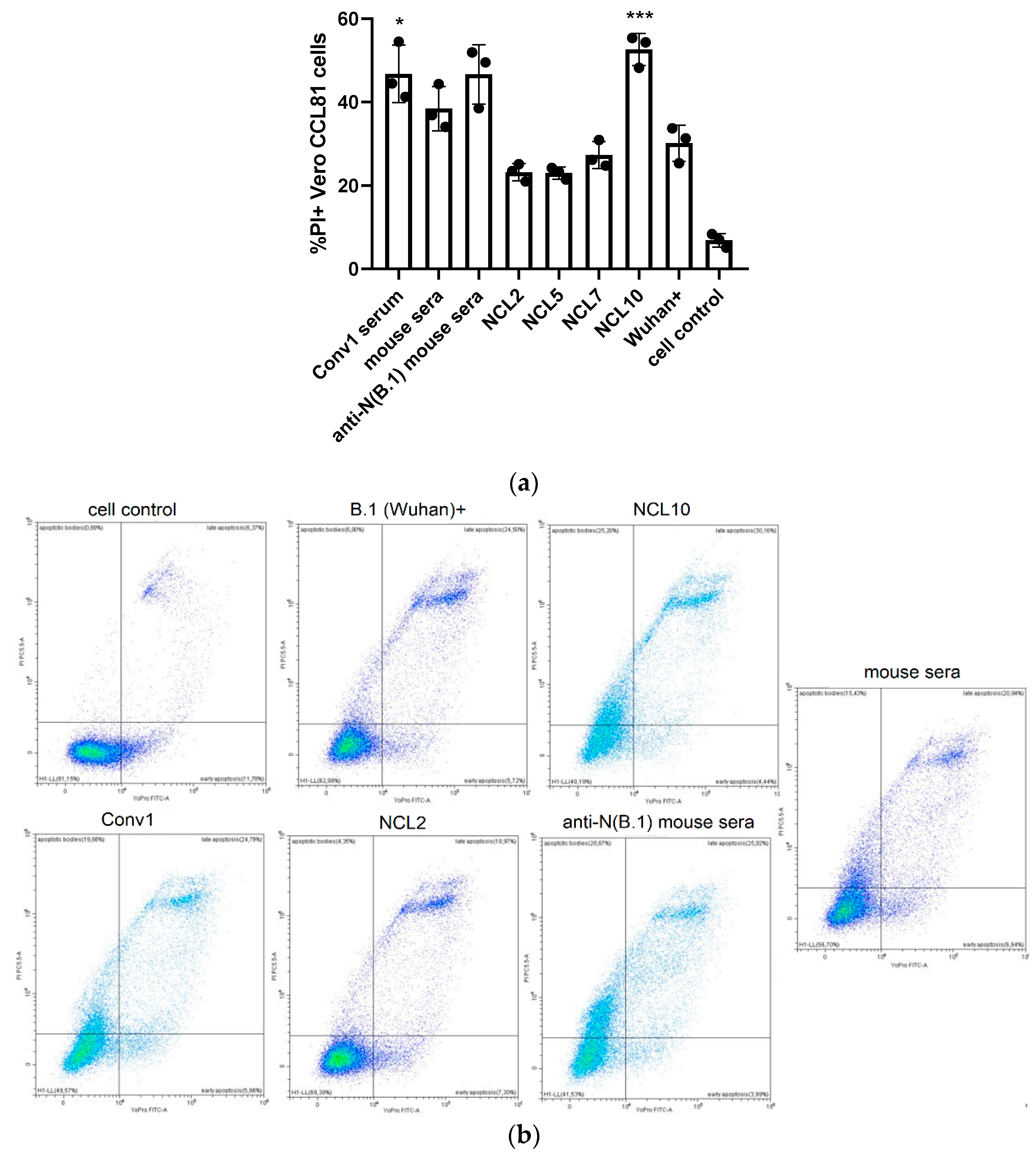
Figure 9.
Analysis of the functional activity of anti-N antibodies: NK-cells degranulation assay (ADCC evaluation).
Figure 9.
Analysis of the functional activity of anti-N antibodies: NK-cells degranulation assay (ADCC evaluation).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



