
Submitted:
03 March 2025
Posted:
04 March 2025
You are already at the latest version
Abstract
To examine the role of asymptomatic infected trees in the spread of pine wilt disease (PWD), we established two study sites in a coastal black pine forest: one in a heavily damaged site and the other in a slightly damaged site. Half of the trees in each site were treated with a nematicide injection to suppress nematode activity. Tree health, assessed by resin exudation and external symptoms, was monitored for four years. In the slightly damaged site, asymptomatic infected trees emerged within 20 m of infected trees, and even with nematicide treatment, trees within 2 m of infected trees became asymptomatic infected. However, nematicide treatment allowed temporarily asymptomatic infected trees to survive or recover. These findings suggest that combining nematicide injection with the felling of neighboring infected trees can effectively suppress PWD.
Keywords:
pine wilt disease
; asymptomatic infected tree
; preventive nematicide injection
1. Introduction
Pine wilt disease (PWD) is a severe epidemic that causes large-scale mortality in pine forests. First identified in Japan in the early 1900s, PWD has since spread to East Asian countries, Portugal, and Spain, causing severe economic and ecological damage [1,2,3,4,5,6]. The pathogen responsible for PWD is the pine wood nematode (Bursaphelenchus xylophilus, hereafter referred to as PWN), which is transmitted from wilted trees to healthy ones by pine sawyer beetles (Monochamus spp.) [7]. In East Asian countries such as Japan, South Korea, and the People's Republic of China, Monochamus alternatus is the primary vector of PWN [1,2]. The potential range of PWD is projected to expand further, raising concerns due to global warming and the increasing international movement of timber [8]. In response, the European Union and several Nordic countries have imposed strict bans on the import of pine wood, causing significant economic losses even in countries where the disease has not yet been reported [9]. In Japan, PWD was first detected in the Kyushu region in the early 1900s and has progressively expanded northward. As of 2024, PWD damage has been observed throughout Japan, except in Hokkaido [1,10].
The infection mechanism of PWD was elucidated in 1972 based on the life cycles of the pathogen, the PWN, and its vector, pine sawyer beetle (Monochamus alternatus) [11,12]. After pupation, newly emerged M. alternatus adults feed on young twigs of healthy pines, creating feeding scars that serve as entry points for PWN infection [13,14,15,16]. Once inside the tree, PWNs spread through the parenchymal tissues, disrupting physiological functions and ultimately leading to tree death [17,18,19,20,21]. During the summer, M. alternatus uses recently wilted pine trees as oviposition hosts. The larvae develop inside the wood, and when the adults emerge in early summer the following year, they carry PWNs from infected wood to new host pines, initiating the next infection cycle [1,22,23,24].
Various control strategies have been developed and implemented to manage PWD, targeting both Monochamus beetles as PWN vectors and PWN itself. Among these, the most common preventive measures are chemical spraying to prevent post-emergence feeding by M. alternatus adults [25,26,27] and preventive trunk injection of nematicides into pine trees to prevent PWN infection [28,29,30]. Eradication measures include felling and removing infected trees through mechanical shredding, fumigation with insecticides, or incineration [31]. Although the infection mechanism and control strategies have been well established, in practice, PWD remains difficult to control, resulting in the widespread distribution of infected trees [31,32,33]. In recent years, emerging technologies, including remote sensing and machine learning, have been developed to improve the accuracy of detecting infected trees and to monitor the expansion of PWD on a large scale [34,35,36].
Despite extensive efforts involving insecticide spraying and the felling of infected trees, PWD persists in pine forests, remaining a major obstacle to effective control. One of the primary factors responsible for this persistence is the presence of asymptomatic infected trees, also referred to as latent infected trees, which appear externally healthy despite harboring PWN [37,38]. In susceptible pine species, PWN inoculation experiments have demonstrated that infected trees can survive for several years while harboring PWNs [39,40]. Notably, some trees have remained asymptomatic for up to 11 years [41]. Consequently, these asymptomatic infected trees are often overlooked and remain in the forest undetected, because conventional control operations primarily target visibly wilted trees. Furthermore, as these trees develop symptoms, they release volatile terpenes and ethanol, attracting adult M. alternatus beetles and facilitating PWN transmission to adjacent healthy trees [42,43]. This delayed disease onset, along with subsequent PWN transmission, poses a major problem for control and drives the continued spread of PWD.
Recent studies have provided further evidence that asymptomatic infected trees are critical factors in the spread of PWD. A trunk injection experiment conducted in Shizuoka Prefecture demonstrated that asymptomatic infected trees can act as new infection sources in the following year, highlighting the importance of their removal [44]. In Iwate Prefecture, a study on the distribution of asymptomatic infected trees and subsequent felling experiments suggested that their removal could contribute to suppressing the spread of PWD [45]. Similarly, field control efforts against PWD in South Korea underscored the problems posed by asymptomatic infected trees, emphasizing the difficulties in achieving effective disease management [46]. Furthermore, a recent study suggests that sexually mature M. alternatus beetles may feed on healthy trees near oviposition hosts during the egg-laying period, potentially transmitting PWNs and leading to the emergence of new asymptomatic infected trees [47].
This study aims to elucidate the role of asymptomatic infected trees in the progression and spatial distribution of PWD through a four-year tree vigor monitoring study initiated prior to infection onset. Special attention is given to identifying asymptomatic infected trees that become detectable following the oviposition period of M. alternatus beetles and examining their contribution to the spread of PWD. By elucidating their role in the infection cycle, we seek to develop more effective PWD management strategies.
2. Materials and Methods
2.1. Study Site
Two study sites were established in a coastal Pinus thunbergii forest in Kakegawa City, Shizuoka Prefecture, Japan (Figure 1). A complete mapping of all healthy trees at the time of site establishment was conducted. One site, referred to as the “severely damaged site,” had already undergone extensive PWD-induced mortality, resulting in a sparse forest structure. The second site, located approximately 500 m east of the severely damaged site, exhibited minimal PWD-induced damage and was thus referred to as the “slightly damaged site.” At the time of site establishment, PWD was actively spreading from the severely damaged site toward the slightly damaged site.
During the study period (2020–2023), the mean annual temperature at the site was 17.7°C, with a recorded maximum of 34.6°C and a minimum of -3.4°C (“Omaezaki” AMeDAS weather station, Japan Meteorological Agency, Figure 1). In the severely damaged site, 102 (97 healthy and five abnormally resin exudated) trees that had survived the PWD outbreak were selected for monitoring. Among them, 51 trees in the southern section were injected with a nematicide (morantel tartrate 20.0%, “Greenguard NEO”; Nisso Green Co., Ltd.) on March 26, 2020, designated as the “nematicide treatment plot.” Trees with a DBH < 20 cm received 90 mL of nematicide per tree, while those with a DBH ≥ 20 cm received 180 mL. The remaining 51 trees in the northern section were left untreated, constituting the control plot. In the slightly damaged site, 100 trees were selected, with 50 trees in the northern section designated as the "nematicide treatment plot," where nematicide was applied under the same conditions as in the severely damaged site on the same date, while the remaining 50 trees constituted the control plot. All trees in the two study sites were individually tagged with numbered tape for identification.
During the study period, felling operations to remove PWD-infected trees were conducted in May 2021 and May 2022. Additionally, to prevent feeding by M. alternatus adults, healthy pine stands were treated annually with aerial application of fenitrothion (MEP) microcapsule formulation (MEP 23.5%, “Sumipine MC”; Sumitomo Chemical Co., Ltd.). The insecticide was applied once per year between mid- and late May at a rate of 60 L/ha using a five-fold dilution of the original MEP (23.5%) formulation. The application was adjusted based on PWD spread, with treatments continuing until 2021 in the severely damaged site and until 2023 in the slightly damaged site. Nematicide trunk injection was not conducted outside the designated experimental sites.
2.2. Tree Health Monitoring
The health status of all trees within the study sites was monitored monthly from March 2020 to November 2021 and subsequently three times per year from April 2022 to August 2023. Tree health monitoring was performed at least three times per year: once during the oviposition period of M. alternatus (July to August) [1,15,16], as well as once before and once after this period.
Tree health was evaluated using two criteria: external appearance and resin exudation. The external appearance was categorized into three classes through visual observation: healthy (green needles), discolored or partially wilted needles, and dead. Resin exudation was evaluated by drilling a hole into the cambium using either a 6 mm punch tool or a 3 mm electric drill. The amount of exuded resin was then assessed approximately 30 minutes after drilling. Resin exudation levels were categorized into five grades (+++, ++, +, −, and 0) following Oda’s method [48].
Some trees were accidentally felled during the study period. In such cases, the identification number of each felled tree was recorded, and resin exudation from the stump was assessed.
2.3. Statistical Analysis
In the slightly damaged site, the spatial relationship between oviposition target trees (i.e., trees targeted for oviposition by M. alternatus adults during the given year) and asymptomatic infected trees (i.e., trees that developed abnormal resin exudation after the oviposition period but remained asymptomatic before the following year’s oviposition period) was analyzed using the O-ring statistic [49]. The pair correlation function was calculated for both tree categories, and the degree of spatial clustering was evaluated by comparing the observed values to those of a null model. The null model was generated from 99 simulations of an inhomogeneous Poisson process with marks to produce the expected pair correlation function .
The null model encompassed all monitored trees. The area analyzed was 341 m² for the untreated (control) plot and 272 m² for the nematicide-treated plot. Deviations from random distribution were considered significant when the observed exceeded the 95% confidence envelope of the null model. All spatial analyses were performed using the spatstat package in R (version 4.3.1) [50]. There is a possibility that asymptomatic infected trees emerging around oviposition target trees located outside the plot were inadvertently included in this analysis. Although this potential limitation was recognized, it is considered unlikely to have significantly affected the conclusions of this study.
Observation data were obtained from the slightly damaged site, where the initial occurrence of PWD-infected trees was monitored. Based on the oviposition period of M. alternatus (July to August) [1,15,16], trees were classified into four categories: healthy trees, oviposition target trees, current-year asymptomatic infected trees, and previous-year asymptomatic infected trees. Oviposition target trees were defined as those that died or weakened between July and August. Asymptomatic infected trees newly emerging in a given year were defined as trees that appeared healthy with normal resin exudation levels (+ or higher) until August but subsequently exhibited reduced resin exudation (minus or lower) while maintaining a healthy external appearance from September to the following June.
The bandwidth parameter for the pair correlation function was optimized separately for each year and treatment plot based on the Berman-Diggle Cross-Validation Criterion [51]. The upper limit for the search range of was set at 5 m, equivalent to half the length of the shorter side of the plot. The bandwidth that minimized the mean squared error was selected for the analysis.
3. Results
3.1. Relationship Between Initial PWD Damage, Nematicide Treatment, and Damage Progression
The annual progression of PWD damage in each study site differed depending on whether nematicide treatment was applied (Figure 2). In the severely damaged site, 60–80% of the trees died following infection in 2020, and an additional 15–20% of the trees died in 2021. After the 2021 infection, almost all untreated (control) trees eventually died. In contrast, approximately 24% (12 trees) of the nematicide-treated trees survived, although they showed abnormal resin exudation from September to June the following year. i.e., became asymptomatic infected trees, without progressing to mortality. Among these trees, approximately 16% (eight trees) remained alive until the end of the study period.
In the slightly damaged site, where PWD damage started to occur later, nematicide treatment not only prolonged the survival of asymptomatic infected trees but also restored their resin exudation to normal levels. PWD damage increased primarily in this site in 2022, with 24% of the trees in the untreated (control) plot and 16% in the nematicide treatment plot dying that year (Figure 2). The proportion of asymptomatic infected trees peaked at 64% (32 trees in each plot) in October 2022.
By August 2023, all the trees in the untreated (control) plot had either died or developed resin exudation abnormalities. In contrast, 72% of the trees in the nematicide treatment plot remained healthy (Figure 3). Notably, of the 32 trees that had been classified as asymptomatic infected in October 2022, 31 (approximately 62% of all monitored trees) exhibited normal resin exudation by May 2023.
3.2. Spatial Overlap Between Oviposition Target Trees and Asymptomatic Infected Trees in the Slightly Damaged Site
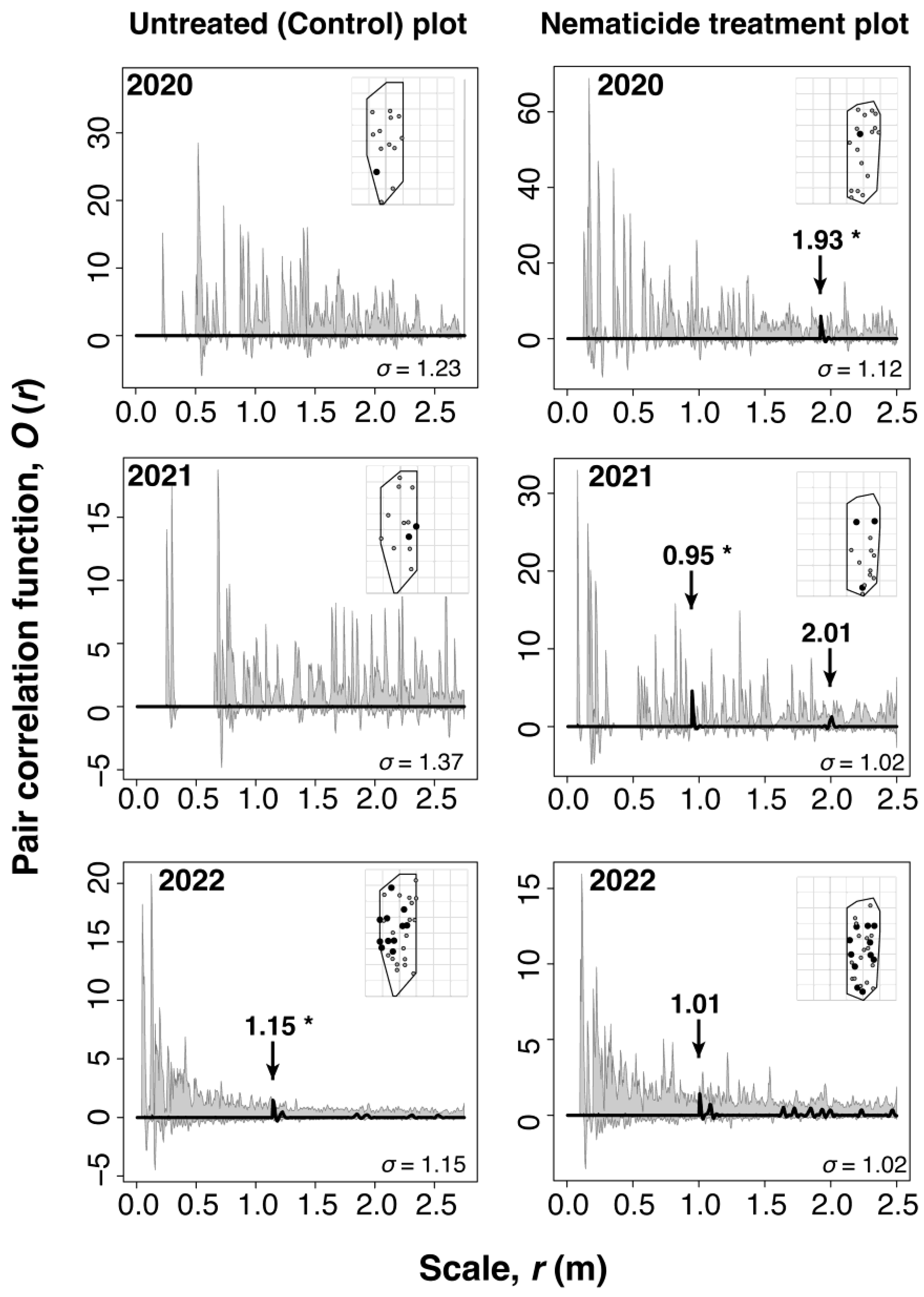
New asymptomatic infected trees emerged within a range of up to approximately 20 m from oviposition target trees, regardless of nematicide treatment (Figure 3). The spatial overlap between the two distributions was analyzed using the pair correlation function (Figure 4). Significant deviations from the null model were detected in the nematicide treatment plot before 2021, preceding the severe outbreak of PWD. This result suggests a tendency for asymptomatic infected trees to cluster around oviposition target trees in the nematicide treatment plot. In contrast, the observed in the untreated (control) plot remained at 0 for all analyzable distances until 2021, indicating no clear spatial association between the two tree categories.
When the PWD outbreak intensified in 2022, the spatial distribution pattern of asymptomatic infected trees relative to oviposition target trees reversed compared to the 2021 results. In the untreated plot, asymptomatic infected trees exhibited significant clustering around oviposition target trees. In contrast, no significant clustering was detected in the nematicide treatment plot. However, a peak in the distribution was detected at approximately 1.01 m, though it remained within the 95% confidence envelope of the null model.
4. Discussion
4.1. Suggestions for PWD Control
This study confirmed the presence of PWD-infected trees that develop symptoms after the oviposition period of M. alternatus adults, as suggested in previous research [37]. These trees play a critical role in the spread of PWD at the stand level. According to conventional theory [1,22], PWNs infect host pines via feeding scars created by immature M. alternatus adults shortly after emergence. The infected trees typically die in the summer of the same year and subsequently serve as oviposition hosts. However, trees exhibiting abnormal resin exudation (asymptomatic infected trees) and/or wilting symptoms from autumn to the following spring (trees that developed symptoms after an asymptomatic stage) (Figure 3) became new oviposition target trees in the following season, leading to the further progression of PWD. Nevertheless, they were omitted from control measures based on the conventional understanding of the infection cycle.
Felling-based control strategies, which involve removing dead trees by early summer of the following year, may overlook asymptomatic infected trees that develop symptoms after autumn. Failure to remove these trees can lead to the release of volatile terpenes and ethanol from weakened trees, attracting sexually mature M. alternatus adults and perpetuating PWD transmission [42,43]. This suggests that undetected asymptomatic infected trees are a key factor in the persistence of PWD outbreaks [38]. The observed expansion of PWD-induced mortality in 2023 in the untreated (control) plot of the slightly damaged site (Figure 3) further underscores the critical role of asymptomatic infected trees, as the number of PWD-affected trees increased rapidly within a short period.
Trunk injection of nematicides, such as morantel tartrate, which has been shown to suppress nematode multiplication for up to seven years, offers an additional benefit beyond its established preventive effect [28]. Even if PWN infection cannot be entirely prevented, nematicide treatment can extend the period from the onset of abnormal resin exudation to eventual tree mortality (Figure 2). When nematicide was applied across the stand before PWD invasion, some trees temporarily exhibited abnormal resin production but later recovered, with resin exudation returning to normal, and survived for up to three years (Figure 3).
for the spatial pattern of oviposition target trees and asymptomatic infected trees. The dashed line indicates the expected pair correlation function obtained from 99 simulations of a null model based on an inhomogeneous Poisson process with marks. The gray-shaded band corresponds to the 95% confidence envelope of the null model. Arrows highlight locations where the observed pair correlation function shows a positive deviation. Among these, values exceeding the 95% confidence envelope are considered significant deviations and are marked with an asterisk (*). The inset maps show the spatial distribution of all monitored trees, with the area used for the null model analysis outlined by solid lines. Oviposition target trees are shown as black circles, while asymptomatic infected trees are shown as black outlines with gray-filled circles. The ring width was estimated based on the Berman-Diggle Cross-Validation Criterion, and the analysis range of was set to .
In contrast, untreated asymptomatic infected trees did not recover normal resin exudation and died by the following summer (Figure 3). Despite two rounds of felling operations, PWD damage continued to spread. By 2023, all healthy trees in the untreated plot had disappeared, causing the complete collapse of the pine stand. These findings suggest that applying trunk injection prior to PWD onset in a stand can effectively reduce the occurrence of asymptomatic infected trees and mitigate the risk of severe outbreaks.
4.2. Suggestions for PWD Control Considering Asymptomatic Infected Trees
This study demonstrated that asymptomatic infected trees occur within a broader range—up to 20 m from oviposition target trees—than previously recognized [37,52]. The actual range, however, may extend further due to the influence of oviposition target trees in adjacent pine stands. Although few in number, several studies in which felling operations targeted asymptomatic infected trees have demonstrated their effectiveness in suppressing the spread of PWD. For example, at the front line of PWD spread in South Korea, all trees within a 20 m radius of an infected tree are felled to prevent the emergence of latent infections. This approach significantly mitigated PWD damage in several regions of South Korea [46]. In Japan, a similar strategy was applied in Iwate Prefecture, where all trees were subjected to resin exudation tests to identify asymptomatic infected trees. Felling both dead and asymptomatic infected trees led to a substantial reduction in PWD damage [45]. While the spatial distribution analysis in this study was constrained by the size of the experimental plots, our present results support the effectiveness of the felling radius adopted in these control strategies. Additionally, because our study could not distinguish infections originating from oviposition target trees outside the experimental plots, the actual range of newly emerging asymptomatic infected trees may have been even broader. Future advancements in early infection detection technologies, such as UAV-based remote sensing [34,35,36], could enhance the accuracy of asymptomatic infected tree identification over larger areas.
Even in plots where nematicide trunk injection was performed to maintain low levels of damage, asymptomatic infected trees tended to cluster at around 1.9 m from oviposition target trees. This proximity likely results in frequent feeding by M. alternatus adults [14,53], increasing the risk of multiple feeding scars and a higher frequency of PWN infections. However, none of the asymptomatic infected trees that emerged near oviposition target trees in nematicide-treated plots died the following year (Figure 3). This result suggests that nematicide treatment suppressed nematode multiplication, preventing asymptomatic infected trees from progressing to mortality.
Our findings suggest that preventive trunk injection of nematicide for all trees in areas not yet affected by PWD is a highly effective strategy. Even if trees become temporarily asymptomatic infected, the nematicide can promote recovery or prolong survival for several years. However, trees within a 2 m radius of initial infection sites—corresponding to neighboring trees in mature pine stands—are likely to remain at a high risk of PWN infection due to concentrated feeding by Monochamus beetles. Although trunk injection may help restore vigor and prevent mortality in asymptomatic infected trees, temporary declines in tree health (i.e., reduced resin exudation) may still attract Monochamus adults, increasing the risk of further spread of PWD.
For effective area-wide prevention of PWD, combining nematicide treatment with targeted felling of infected trees and their neighboring pines within a 2 m radius is likely to be a more effective strategy. This integrated approach could enhance PWD management and help maintain long-term forest health.
Author Contributions
Conceptualization, K.F.; methodology, K.F.; formal analysis, Y.U.; investigation, Y.U.; data curation, Y.U.; writing—original draft preparation, Y.U.; writing—review and editing, Y.U. and K.F.; visualization, Y.U. and K.F.; supervision, K.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data published in this study will be shared by the co-authors and is available upon request from the corresponding author.
Acknowledgments
We thank Shujiro Yamada, Tatsuaki Ito, Takumi Fukuda, Naoki Nozue, and Tatsumune Washiyama (Shizuoka Prefecture) for their assistance with tree health monitoring. We also thank Michimasa Yamasaki (Kyoto University) for his valuable suggestions on spatial statistical analysis. We sincerely thank Toru Kato for his extensive contributions to fieldwork over an extended period, as well as for his constructive comments and valuable participation in discussions during the preparation of this paper.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mamiya, Y. History of Pine Wilt Disease in Japan. J Nematol 1988, 20, 219–226. [Google Scholar] [PubMed]
- Zhao, B.G. Pine Wilt Disease in China. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer Japan: Tokyo, Japan, 2008; ISBN 978-4-431-75655-2. [Google Scholar]
- Hao, Z.; Huang, J.; Li, X.; Sun, H.; Fang, G. A Multi-Point Aggregation Trend of the Outbreak of Pine Wilt Disease in China over the Past 20 Years. Forest Ecology and Management 2022, 505, 119890. [Google Scholar] [CrossRef]
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First Report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Futai, K. Pine Wood Nematode, Bursaphelenchus xylophilus. Annual Review of Phytopathology 2013, 51, 61–83. [Google Scholar] [CrossRef]
- Back, M.A.; Bonifácio, L.; Inácio, M.L.; Mota, M.; Boa, E. Pine Wilt Disease: A Global Threat to Forestry. Plant Pathology 2024, 73, 1026–1041. [Google Scholar] [CrossRef]
- Akbulut, S.; Stamps, W.T. Insect Vectors of the Pinewood Nematode: A Review of the Biology and Ecology of Monochamus Species. Forest Pathology 2012, 42, 89–99. [Google Scholar] [CrossRef]
- Hirata, A.; Nakamura, K.; Nakao, K.; Kominami, Y.; Tanaka, N.; Ohashi, H.; Takano, K.T.; Takeuchi, W.; Matsui, T. Potential Distribution of Pine Wilt Disease Under Future Climate Change Scenarios. PLOS ONE 2017, 12, e0182837. [Google Scholar] [CrossRef]
- Vicente, C.; Espada, M.; Vieira, P.; Mota, M. Pine Wilt Disease: A Threat to European Forestry. Eur J Plant Pathol 2012, 133, 89–99. [Google Scholar] [CrossRef]
- Annual Report on Forest and Forestry in Japan: Fiscal Year 2023. Available online: https://www.rinya.maff.go.jp/j/kikaku/hakusyo/r5hakusyo/index.html (accessed on 14 February 2025).
- Kiyohara, T.; Tokushige, Y. Inoculation Experiments of a Nematode, Bursaphelenchus sp., onto Pine Trees. Journal of the Japanese Forestry Society 1971, 53, 210–218, (in Japanese with English summary). [Google Scholar] [CrossRef]
- Mamiya, Y.; Enda, N. Transmission of Bursaphelenchus lignicolus (Nematoda: Aphelenchoididae) By Monochamus alternatus (Coleoptera: Cerambycidae). Nematologica 1972, 18, 159–162. [Google Scholar] [CrossRef]
- Kishi, Y. Invasion of Pine Trees by Bursaphelenchus lignicolus M. & K. (Nematoda: Aphelenchoidae) from Monochamus alternatus HOPE (Coleoptera: Cerambycidae). Journal of the Japanese Forestry Society 1978, 60, 179–182. (in Japanese). [Google Scholar] [CrossRef]
- Togashi, K. Transmission Curves of Bursaphelenchus xylophilus (Nematoda : Aphelenchoididae) from Its Vector, Monochamus alternatus (Coleoptera : Cerambycidae), to Pine Trees with Reference to Population Performance. Applied Entomology and Zoology 1985, 20, 246–251. [Google Scholar] [CrossRef]
- Linit, M.J. Transmission of Pinewood Nematode Through Feeding Wounds of Monochamus carolinensis (Coleoptera: Cerambycidae). J Nematol 1990, 22, 231–236. [Google Scholar]
- Togashi, K. Vector-Nematode Relationships and Epidemiology in Pine Wilt Disease. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer Japan: Tokyo, Japan, 2008; ISBN 978-4-431-75654-5. [Google Scholar]
- Togashi, K. Variation in External Symptom Development of Pine Wilt Disease in Field Grown Pinus thunbergii. Journal of the Japanese Forestry Society 1989, 71, 442–448. [Google Scholar] [CrossRef]
- Kuroda, K. Mechanism of Cavitation Development in the Pine Wilt Disease. European Journal of Forest Pathology 1991, 21, 82–89. [Google Scholar] [CrossRef]
- Fukuda, K. Physiological Process of the Symptom Development and Resistance Mechanism in Pine Wilt Disease. Journal of Forest Research 1997, 2, 171–181. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Q.; Zhou, Z.; Yin, H.; Xie, Y.; Wei, Y. Two Terpene Synthases in Resistant Contribute to Defence Against Bursaphelenchus xylophilus. Plant, Cell & Environment 2021, 44, 257–274. [Google Scholar] [CrossRef]
- Feng, Y.; Li, Y.; Li, D.; Liu, Z.; Wang, X.; Zhang, W.; Wen, X.; Zhang, X. Bursaphelenchus xylophilus Venom Allergen Protein BxVAP2 Responds to Terpene Stress, Triggers Plant Defense in Nicotiana benthamiana. Forests 2024, 15, 1929. [Google Scholar] [CrossRef]
- Linit, M.J. Nemtaode-Vector Relationships in the Pine Wilt Disease System. J Nematol 1988, 20, 227–235. [Google Scholar]
- Zhao, L.; Zhang, S.; Wei, W.; Hao, H.; Zhang, B.; Butcher, R.A.; Sun, J. Chemical Signals Synchronize the Life Cycles of a Plant-Parasitic Nematode and Its Vector Beetle. Current Biology 2013, 23, 2038–2043. [Google Scholar] [CrossRef]
- Kirino, H.; Maehara, N.; Shinya, R. How Did Bursaphelenchus Nematodes Acquire a Specific Relationship with Their Beetle Vectors, Monochamus? Front. Physiol. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K. Inhibitory Activity of Fenitrothion Against Maturation Feeding of Japanese Pinesawyer, Monochamus alternatus. Japanese Journal of Applied Entomology and Zoology 1988, 32, 245–251. [Google Scholar] [CrossRef]
- Kwon, T.-S.; Song, M.-Y.; Shin, S.-C.; Park, Y.-S. Effects of Aerial Insecticide Sprays on Ant Communities to Control Pine Wilt Disease in Korean Pine Forests. Applied Entomology and Zoology 2005, 40, 563–574. [Google Scholar] [CrossRef]
- Suh, D.Y.; Jung, J.-K.; Lee, S.K.; Seo, S.-T. Effect of Aerial Spraying of Thiacloprid on Pine Sawyer Beetles (Monochamus alternatus) and Honey Bees (Apis mellifera) in Pine Forests. Entomological Research 2021, 51, 83–89. [Google Scholar] [CrossRef]
- Fujishita, A. Trunk Injection of Chemicals for the Control of the Pine Wilt Disease Caused by Bursaphelenchus xylophilus. Bulletin of the Shizuoka Prefecture Forestry Experiment Station 1985, 13, 23–34, (in Japanese with English summary). [Google Scholar]
- Sousa, E.; Naves, P.; Vieira, M. Prevention of Pine Wilt Disease Induced by Bursaphelenchus xylophilus and Monochamus galloprovincialis by Trunk Injection of Emamectin Benzoate. Phytoparasitica 2013, 41, 143–148. [Google Scholar] [CrossRef]
- Li, M.; Wang, M.; Yang, T.; Xu, M.; Li, Y.; Pei, Y.; Tang, J.; Zheng, Z.; Sun, Z.; Cheng, G.; Li, X.; Li, H.; Wang, L; Chen, F. Optimized Emamectin Benzoate Trunk Injection: Addressing Temperature Limitations for Pine Wilt Disease Control. Pest Management Science 2025, 81, 892–902. [Google Scholar] [CrossRef]
- Yoshimura, A.; Kawasaki, K.; Takasu, F.; Togashi, K.; Futai, K.; Shigesada, N. Modeling the Spread of Pine Wilt Disease Caused by Nematodes with Pine Sawyers as Vector. Ecology 1999, 80, 1691–1702. [Google Scholar] [CrossRef]
- Togashi, K.; Shigesada, N. Spread of the Pinewood Nematode Vectored by the Japanese Pine Sawyer: Modeling and Analytical Approaches. Popul Ecol 2006, 48, 271–283. [Google Scholar] [CrossRef]
- Xia, C.; Chon, T.-S.; Takasu, F.; Choi, W.I.; Park, Y.-S. Simulating Pine Wilt Disease Dispersal With an Individual-Based Model Incorporating Individual Movement Patterns of Vector Beetles. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Yu, R.; Luo, Y.; Zhou, Q.; Zhang, X.; Wu, D.; Ren, L. Early Detection of Pine Wilt Disease Using Deep Learning Algorithms and UAV-based Multispectral Imagery. Forest Ecology and Management 2021, 497, 119493. [Google Scholar] [CrossRef]
- Tahir, S.; Hassan, S.S.; Yang, L.; Ma, M.; Li, C. Detection Methods for Pine Wilt Disease: A Comprehensive Review. Plants 2024, 13, 2876. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cai, J.; Wang, T.; Zhao, J.; Gadekallu, T.R.; Fang, K. Detection of Pine Wilt Disease Using AAV Remote Sensing With an Improved YOLO Model. IEEE Journal of Selected Topics in Applied Earth Observations and Remote Sensing 2024, 17, 19230–19242. [Google Scholar] [CrossRef]
- Futai, K. Role of Asymptomatic Carrier Trees in Epidemic Spread of Pine Wilt Disease. J For Res 2003, 8, 253–260. [Google Scholar] [CrossRef]
- Futai, K. Two Infectious Diseases: “COVID-19” and “Pine Wilt Disease. ” Forests 2024, 15, 1724. [Google Scholar] [CrossRef]
- Halik, S.; Bergdahl, D.R. Long-Term Survival of Bursaphelenchus xylophilus in Living Pinus sylvestris in an Established Plantation. European Journal of Forest Pathology 1994, 24, 357–363. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Futai, K. Asymptomatic Carrier Trees in Pine Stands Naturally Infected with Bursaphelenchus xylophilus. Nematology 2007, 9, 243–250. [Google Scholar] [CrossRef]
- Bergdahl, D.R.; Halik, S. Persistence of the Pine Wood Nematode in Asymptomatic Scots Pines. In The Pinewood Nematode, Bursaphelenchus xylophilus; Mota, M, Vieira, P., Eds.; Brill: Berlin, Germany, 2004; ISBN 978-90-474-1309-7. [Google Scholar]
- Takeuchi, Y.; Kanzaki, N.; Futai, K. Volatile Compounds in Pine Stands Suffering from Pine Wilt Disease: Qualitative and Quantitative Evaluation. Nematology 2006, 8, 869–879. [Google Scholar] [CrossRef]
- Yang, R.; Li, D.; Yi, S.; Wei, Y.; Wang, M. Odorant-Binding Protein 19 in Monochamus alternatus Involved in the Recognition of a Volatile Strongly Emitted from Ovipositing Host Pines. Insect Science 2024, 31, 134–146. [Google Scholar] [CrossRef]
- Kato, T.; Kenmochi, A.; Yamada, Y.; Futai, K. Confirmation of the Presence of Asymptomatic Carrier Trees of Pine Wilt Disease After Thorough Control Procedure, and the Effectiveness of Trunk Injection to Suppress Disease Development. Journal of the Japanese Forest Society 2019, 101, 46–51. [Google Scholar] [CrossRef]
- Koiwa, T. Survey of Asymptomatic Carrier Trees of Pine Wilt Disease in Iwate Prefecture, Northeastern Japan. Journal of the Japanese Forest Society 2019, 101, 35–45. [Google Scholar] [CrossRef]
- Cho, Y.; Jung, K. Countermeasures Against Asymptomatic Carriers in Pine Wilt Disease in Korea. Journal of the Japanese Forest Society 2019, 101, 26–29. [Google Scholar] [CrossRef]
- Ishiguro, H.; Futai, K. When Coming to a Wilting Pine Tree, Monochamus alternatus Causes Pinewood Nematode Infection on the Surrounding Healthy Trees. Tree and Forest Health 2022, 26, 59–64, (in Japanese with English summary). [Google Scholar] [CrossRef]
- Oda, K. Target trees of pine wilt disease and its diagnostic method. Forest Pests 1967, 16, 263–266. (in Japanese). [Google Scholar]
- Wiegand, T.; A. Moloney, K. Rings, Circles, and Null-Models for Point Pattern Analysis in Ecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- Baddeley, A.; Turner, R. Spatstat: An R Package for Analyzing Spatial Point Patterns. Journal of Statistical Software 2005, 12, 1–42. [Google Scholar] [CrossRef]
- Berman, M.; Diggle, P. Estimating Weighted Integrals of the Second-Order Intensity of a Spatial Point Process. Journal of the Royal Statistical Society: Series B (Methodological) 1989, 51, 81–92. [Google Scholar] [CrossRef]
- Togashi, K. Spatial Pattern of Pine Wilt Disease Caused by Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) Within a Pinus thunbergii Stand. Res Popul Ecol 1991, 33, 245–256. [Google Scholar] [CrossRef]
- Naves, P.M.; Camacho, S.; De Sousa, E.M.; Quartau, J.A. Transmission of the Pine Wood Nematode Bursaphelenchus xylophilus Through Feeding Activity of Monochamus galloprovincialis (Col., Cerambycidae). Journal of Applied Entomology 2007, 131, 21–25. [Google Scholar] [CrossRef]
Figure 1.
Location of the study site and climatic conditions (temperature and precipitation) during the study period. Temperature and precipitation data were obtained from the AMeDAS observation station in Omaezaki, operated by the Japan Meteorological Agency. The study site is represented by a black circle, while the observation station is indicated by a white circle.
Figure 1.
Location of the study site and climatic conditions (temperature and precipitation) during the study period. Temperature and precipitation data were obtained from the AMeDAS observation station in Omaezaki, operated by the Japan Meteorological Agency. The study site is represented by a black circle, while the observation station is indicated by a white circle.
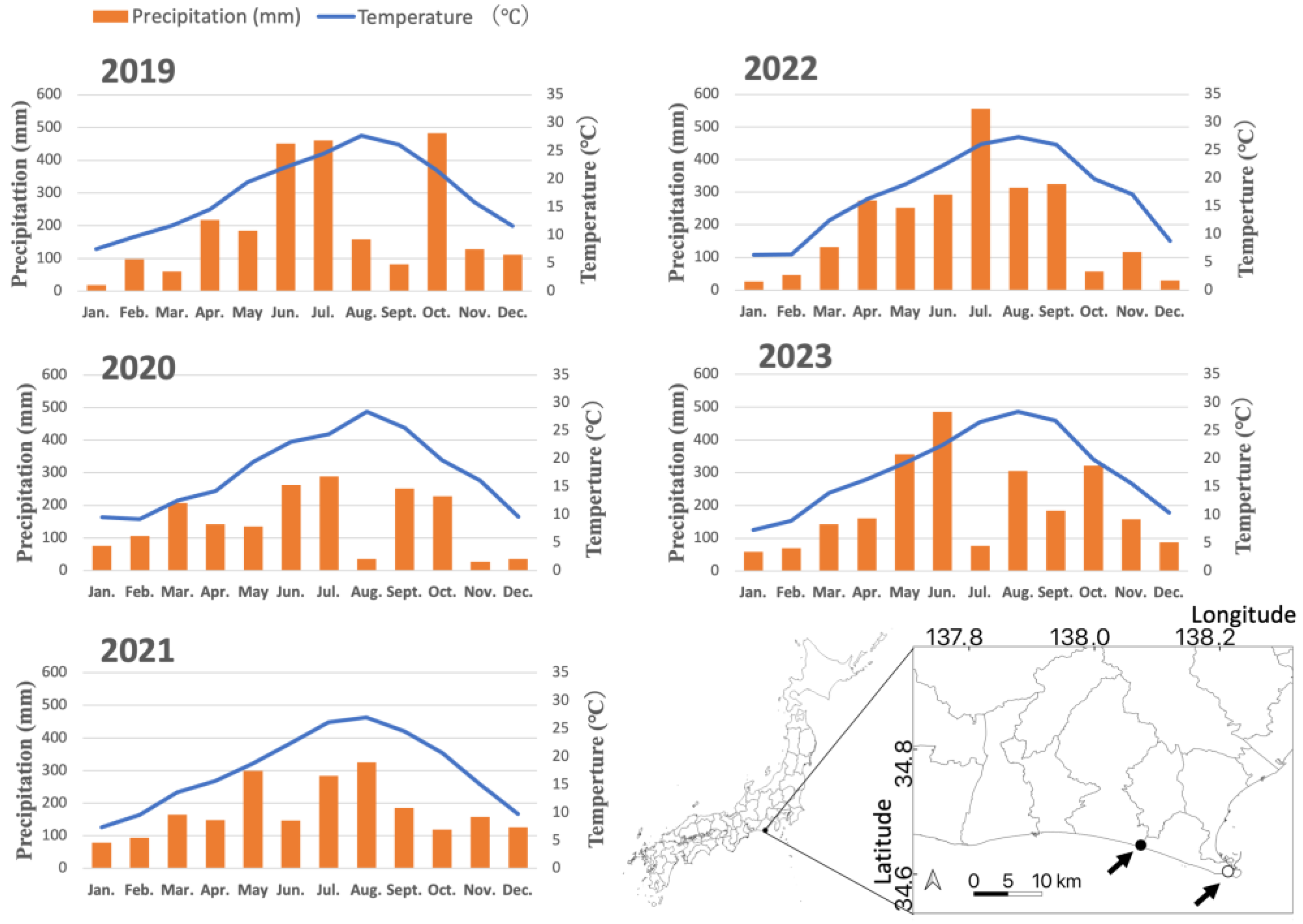
Figure 2.
Progression of pine wilt disease (PWD) damage in each study plot. Graphs from top to bottom: control plot of the severely damaged site, nematicide treatment plot of the severely damaged site, control plot of the slightly damaged site, and nematicide treatment plot of the slightly damaged site. The gray-shaded bands represent the July−August period, corresponding to the oviposition period of the pine sawyer beetle (Monochamus alternatus) [1,15,16]. Trees removed due to Monochamus beetle eradication or accidental felling were also included among the dead trees. Healthy trees are shown in yellow-green, trees with abnormal resin secretion (asymptomatic infected trees) in beige and dead trees in dark brown.
Figure 2.
Progression of pine wilt disease (PWD) damage in each study plot. Graphs from top to bottom: control plot of the severely damaged site, nematicide treatment plot of the severely damaged site, control plot of the slightly damaged site, and nematicide treatment plot of the slightly damaged site. The gray-shaded bands represent the July−August period, corresponding to the oviposition period of the pine sawyer beetle (Monochamus alternatus) [1,15,16]. Trees removed due to Monochamus beetle eradication or accidental felling were also included among the dead trees. Healthy trees are shown in yellow-green, trees with abnormal resin secretion (asymptomatic infected trees) in beige and dead trees in dark brown.
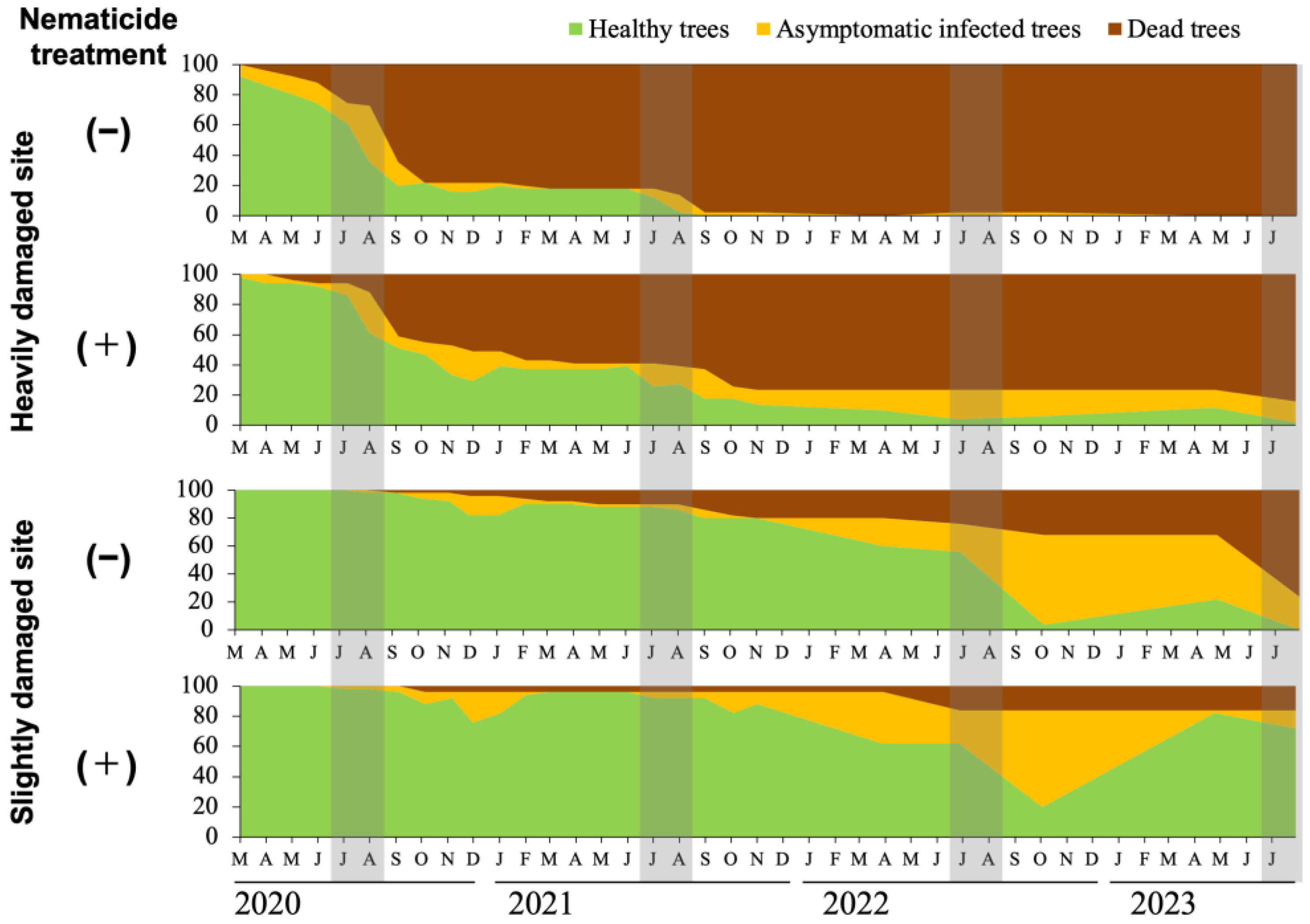
Figure 3.
Progression of PWD damage and spatial distribution of affected trees in control and nematicide treatment plots of the slightly damaged site. Each grid unit represents 5 m on both the x- and y-axes. Oviposition target trees are defined as those that were either dead or asymptomatic infected trees (i.e., those with a resin value of minus or lower according to Oda’s method [48]) during the beetle’s oviposition period (July to August) of the given year. These trees are shown as black circles. New asymptomatic infected trees, defined as those that emerged from September of that year until June of the following year, are shown as grey-filled circles with a black border. Trees that became asymptomatic infected trees in a previous year or earlier and remained alive in the given year are shown as gray circles. Healthy trees, defined as those that exhibited no external symptoms and had normal resin secretion (+ or higher) [48], are shown as white circles. Additionally, trees removed during Monochamus beetle eradication efforts in May 2021 and May 2022, as well as those accidentally felled, were excluded from subsequent plots.
Figure 3.
Progression of PWD damage and spatial distribution of affected trees in control and nematicide treatment plots of the slightly damaged site. Each grid unit represents 5 m on both the x- and y-axes. Oviposition target trees are defined as those that were either dead or asymptomatic infected trees (i.e., those with a resin value of minus or lower according to Oda’s method [48]) during the beetle’s oviposition period (July to August) of the given year. These trees are shown as black circles. New asymptomatic infected trees, defined as those that emerged from September of that year until June of the following year, are shown as grey-filled circles with a black border. Trees that became asymptomatic infected trees in a previous year or earlier and remained alive in the given year are shown as gray circles. Healthy trees, defined as those that exhibited no external symptoms and had normal resin secretion (+ or higher) [48], are shown as white circles. Additionally, trees removed during Monochamus beetle eradication efforts in May 2021 and May 2022, as well as those accidentally felled, were excluded from subsequent plots.
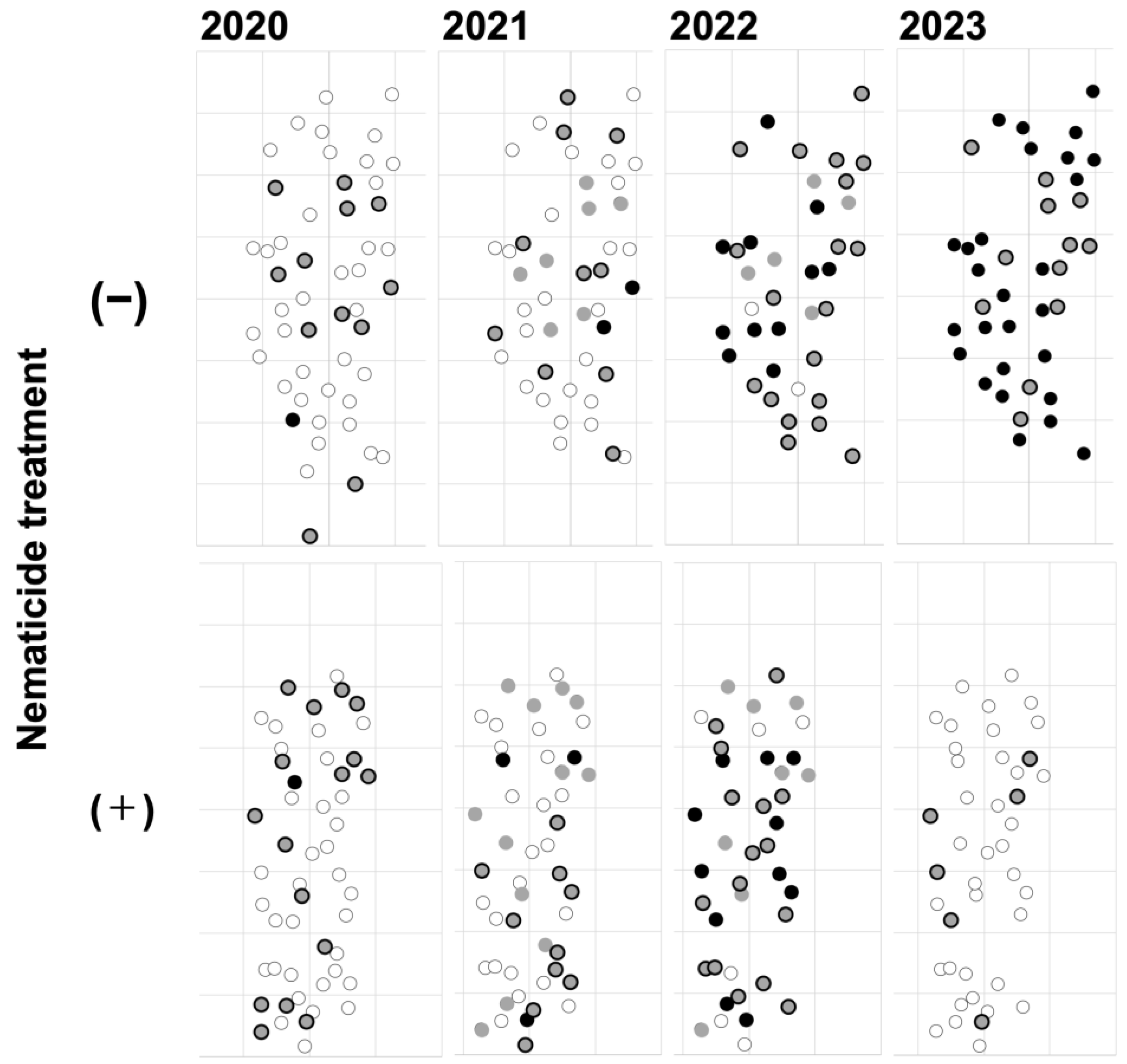
Figure 4.
Analysis of the spatial patterns of oviposition target trees and asymptomatic infected trees in the slightly damaged site using O-ring statistics. The left panels represent the control (untreated) plot, while the right panels correspond to the nematicide treatment plot. Spatial patterns were compared across different years. The thick solid line represents the observed pair correlation function
Figure 4.
Analysis of the spatial patterns of oviposition target trees and asymptomatic infected trees in the slightly damaged site using O-ring statistics. The left panels represent the control (untreated) plot, while the right panels correspond to the nematicide treatment plot. Spatial patterns were compared across different years. The thick solid line represents the observed pair correlation function
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



