
Submitted:
28 February 2025
Posted:
03 March 2025
You are already at the latest version
Abstract
Many countries have banned the use of antibiotics in livestock feed owing to the develop-ment of antibiotic-resistant bacteria. Therefore, it is necessary to identify suitable alterna-tives to antibiotics to maintain intestinal health and improve piglet performance after the weaning. The objective of the present research was to evaluate the effect of isolated or combined intake of potato protein concentrate (PPC) and Saccharomyces boulardii (SB) on productive performance and some morphophysiological responses of the digestive tract in the two post-weaning weeks. We probe negative control without antibiotics, PPC, or SB; positive control with only antibiotics, and three other diets with SB, PPC, or a combination of PPC-SB. The absence of antibiotics in the diet or the inclusion of PPC and SB did not af-fect performance and digestive organ development after the weaning. The piglets fed a combination of PPC-SB had a higher concentration of protein occludins in the jejunum and intestinal villi than those fed a C- diet and had a similar effect to the C+ diet. The use of SB alone or in combination with PPC increased fecal Lactobacillus and antibiotics re-duced coliform bacteria. The PPC-SB combination prevented atrophy of the jejunal villi and promoted adhesion between enterocytes in the jejunum.
Keywords:
piglets
; occludins
; intestinal permeability
; functional foods
; probiotics
1. Introduction
After birth, piglets have a low capacity to digest and absorb nutrients of plant origin; therefore, their digestive systems must mature quickly to ensure survival [1]. The post-weaning period is critical for pig production, health, and performance. Therefore, there is substantial worldwide interest in mitigating post-weaning challenges, and identifying nutritional means, management, and disease prevention to positively influence the structure and function of the gastrointestinal tract (GIT) and improve production and health outcomes [2]. Stress at weaning, in addition to the presence of new pathogens, leads to neuroendocrine, immunological, and digestive complications [3]. Consequently, in the first 24–48 h after weaning, low or even no feed intake, weight loss, and atrophy of the intestinal mucosa provoke a decrease in digestive capacity, as well as post-weaning diarrhea [4]. Diarrhea is widely related to sudden changes in diet from milk to solid feed and gastrointestinal infections. Both factors promote rapid bacterial dysbiosis with an abrupt reduction in Lactobacillus spp., which contributes to the loss of intestinal structure [5]. Antibiotics have traditionally been used as growth promoters to combat gastrointestinal complications because their use in animal feed improves the animal growth rate and reduces the incidence of diseases and mortality [6].
To reduce antibiotic usage in animal production, a global effort has been made [7] and feed-based antibiotics have been prohibited in several parts of the world. Although Mexico has a mandatory National Action Strategy against antimicrobial resistance, published in the Official Gazette of the Federation in June 2018, there is still no regulatory legislation that effectively controls antimicrobials. International pressure has promoted research into alternatives to antibiotics in pig feed [7]. The use of potato protein concentrate (PPC) in antibiotic-free diets had a positive effect on the ileal digestibility of nutrients in piglets, as well as improving the use of dry matter and energy throughout the gastrointestinal tract [8,9]. In addition to being a protein of high nutritional quality, it offers functional benefits for human health like antiallergic, antioxidant, and anticancer activity [10-13] which can also cover animal health. We are particularly interested in actions that can promote intestinal health in piglets on antibiotic-free diets [14], due to its peptides with antifungal and antimicrobial action [15].
Yeasts have been used as probiotics in animal diets to reduce the use of growth-promoting antibiotics owing to their positive effects on productive criteria, intestinal microbiota, diarrhea incidence, intestinal anti-inflammatory response, health and integrity of the intestinal mucosa, and secretion of metabolites that improve the immune response [16]; in addition to having antiviral action [17]. Advantages of a single, early-life administration of Saccharomyces boulardii on the growth performance and faecal microbiota of piglets from birth until weaning are reported [18]. Saccharomyces boulardii is used in animals feed because of its benefices reducing diarrhea in young animals, improving body immunity, intestinal barrier function, and performance. Effectively reduce the colonization and translocation of pathogenic bacteria in the digestive tract of pigs, increase IgA antibody secretion, inhibit inflammation and decrease mortality [19]. Another study with Saccharomyces cerevisiae var. boulardii RC009 observed improvements in productive parameters in the post-weaning stage and an increase in the health status of the animals, indicating that it may be a promising alternative to prophylactic antibiotics [20].
Therefore, PPC and Saccharomyces boulardii meet the definition of functional foods [21] because they are characterized by improving intestinal health and therapeutics conditions of individuals when consumed in adequate quantities.
An important function of a healthy small intestine is to serve as a physical barrier, limiting the migration of pathogenic microorganisms from the lumen to the interior of enterocytes [22]. With the increased paracellular permeability, toxins, allergens, and microorganisms can pass through the mucosa and induce inflammatory and immunological responses [23]. Occludins are essential for the selective permeability of enterocytes as they are integral membrane proteins with four transmembrane domains located exclusively at tight junctions [24].
The objective of this study was to evaluate the effects of PPC and Saccharomyces boulardii (SB), either alone or in combination, on piglet performance, digestive organ weight, pH of digestive contents, intestinal morphology of the villi and crypts, presence of occludin proteins in jejunum and colon samples, coliform and Lactobacillus populations in feces, and the incidence and severity of diarrhea during the first two weeks post-weaning.
2. Materials and Methods
This study was approved by the Bioethical Committee of the Faculty of Natural Sciences of the Autonomous University of Querétaro (approval number: 63FCN2021) and was conducted at the experimental farm of CENID Physiology at the National Institute of Agricultural and Livestock Forestry Research, National Center of Research in Animal Physiology, Mexico. Pigs were treated according to the International Guiding Principles for Biomedical Research Involving Animals [25] and Official Mexican Standards and Regulations for the Technical Specifications of Production, Care, and Use of Laboratory Animals (NOM-062-ZOO-1999) [26].
Approximately 132 pigs (Fertilis × Genetiporc) were weaned at 19.8 ± 1.6 days and weighed in at 6.2 ± 0.85 kg. The piglets were moved to a weaning room with a controlled environment (30 °C and 28 ± 2 °C during the first and second week after weaning, respectively). The pens were elevated (38 cm above the floor), 115 cm wide, and 150 cm long (actual surface area 1.7 m2), contained a nipple drinker, feeder, and plastic grid floor. The piglets were assigned to one of the five experimental diets according to a randomized complete block design, with initial weight used as a blocking factor: C- (negative control, without PPC or SB), C+ (positive control, with antibiotic), SB (without antibiotic, with SB), PPC (without antibiotic, with PPC), or PPC-SB (without antibiotic, with PPC + SB). The diets were formulated using the principle of Ideal Protein, using the Standardized Ileal Digestibility of amino acids (Table 1). All diets met the nutritional requirements of the National Research Council (NRC) [27]. Feed was offered three times a day at 8:00, 12:00, and 16:00, and feed intake was recorded daily. The piglets were weighed at weaning and at the end of weeks one and two after weaning. Simultaneously, the average daily feed intake (ADFI), average daily gain (ADG), and feed efficiency (FE) were determined. All animals had ad libitum access to water.
Euthanization and sample collection: On day 14 post-weaning, six piglets from each experimental group were randomly selected for euthanasia, and samples were collected. The animals were desensitized by CO2 inhalation for three minutes, and the jugular vein was sectioned for exsanguination. Subsequently, the abdominal cavity was opened to extract the liver, pancreas, stomach, and small and large intestines, which were then emptied of their contents, washed, and weighed. The relative weights (g/Kg live weight) were calculated. The contents of the stomach, duodenum, jejunum, ileum, cecum, and colon were measured using a potentiometer. Samples of approximately 10 cm were collected from the duodenum, jejunum, ileum, and colon and preserved in buffered formalin (10%) until processing. A 2-cm wide area of the mucosa was fixed in paraffin, and 5-µm sections were cut and stained with hematoxylin-eosin to determine the depth of the crypt (from the villi base to the bottom of the crypt) (µm) and height (from apex-base) of the villi (µm). Approximately 10 villi were examined under an optical microscope with a 10× objective using a Primo Star optical microscope (Carl Zeiss, Oberkochen, Germany) [28].
Presence of occludins: To determine the presence of occludins, 10 cm of jejunum and colon were obtained from three piglets in each experimental group and selected at random. Histological sections were prepared to produce antigen-antibody reactions and quantify occludins by adapting the immunohistochemistry technique [29], which is briefly described below. The tissues were incubated with a polyclonal anti-occludin antibody (Thermo Fisher Scientific Bioss Brand, Catalog No.: BS-1495R, diluted to 1:100), and horse serum (1:1000). Subsequently, the tissues were incubated with a Goat and Rabit IDG (H+L) secondary antibody (Brand Thermo Fisher Scientific Invitrogen, Catalog No.: 32460); Finally, it was performed using 3,3'-Diaminobenzidine tetrahydrochloride hydrate (Thermo Scientific Chemicals). This reaction produced a sepia precipitate in the immunoreactive cells. The samples were fixed with entelan. Photographic analyses of three different sections of the jejunum and colon were performed on each slide. The samples were observed under a Primo Star microscope (Carl Zeiss, Oberkochen, Germany) and photographed at 40× and 10× magnifications using the ZEN program (Carl Zeiss, Jena, Germany). The photographs were analyzed using ImageJ software developed by Wayne Rasband at the U.S. National Institutes of Health (NIH), which can be downloaded from https://imagej.nih.gov/ij.
Coliform and Lactobacillus Populations: Immediately before slaughter, approximately 5 g of feces were collected directly from the anus of each animal for microbiological analysis. The samples were serially diluted and plated onto 100 µl surface plates. To quantify he total coliforms the norm NOM-113-SSA1-1994 was used [30]. Bile and violet red agar (Bioxon™) culture medium was used for coliforms and MRS agar (Difco Laborato-ries, Detroit, MI, USA) was used for Lactobacillus [31].
Incidence and severity of diarrhea: The incidence and severity of diarrhea in each pen were evaluated daily. Diarrhea incidence (DI) was measured as the number of days diarrhea was observed within a pen. Diarrhea severity (DS) was defined as a daily visual score of 0–3 [32] based on fecal consistency: 3 = severe, highly fluid diarrhea; 2 = moderate diarrhea; 1 = light, pasty diarrhea; and 0 = no diarrhea. Daily scores were added for each week and divided into seven days to obtain the diarrhea severity index for each pen.
Statistical analysis: In the performance trial, the variables were analyzed according to a random block design, with the initial weight used as a blocking factor and the pen used as the experimental unit. Other variables were analyzed according to a completely randomized design in which piglets were used as the experimental unit. The General Linear Model (GLM) procedure of the statistical software SAS (SAS Inst. Inc., Carry, NC, USA). The significance of differences between means was determined using Tukey’s test. p < 0.05 was considered as significant.
3. Results
The average daily feed intake (ADFI), average daily gain (ADG), and feed efficiency (FE) of piglets were not affected by the different diets. In the first week after weaning, all piglets, regardless of the diet, exhibited reduced feed intake. Consequently, they lost weight and had a negative FE. In the second week, all animals recovered as their performance improved with an increase in ADFI and ADG, reaching an FE very close to the ideal value of 1 (Table 2).
The relative weights of the digestive organs and the pH of the digestive contents (Table 3) did not differ (P > 0.05) between the animals of the different experimental diets.
In the duodenum, the villi were wider (P < 0.01) in the piglets that fed the SB and PPC-SB diets, and intermediate in the C+ diets (Table 4). With regards to the jejunum, these values were significantly higher (p < 0.01) in the C+ and PPC-SB groups. No differences between piglets were observed (p > 0.05) in the other intestinal segments.
The group fed a diet with antibiotics (C+), as well as those fed a mixture of the two functional feeds, (PPC-SB) exhibited higher concentrations of occludins (P < 0.05) in the jejunum (Figure 1).
In the piglets that were given feed containing only yeast (SB) or potato protein concentrate (PPC), there was a lower abundance of occludins, like animals that fed the C-diet; but no differences were observed (p > 0.05) in the concentration of occludins in the colon (Figure 2).
Animals fed the antibiotic diet had a lower (p < 0.05) number of coliforms in their feces when compared to the other animals. However, the intake of the PPC diet reduced the proportion of these bacteria, leading to effects like those observed in piglets that fed the antibiotic diet. The piglets that consumed SB and PPC-SB contained more Lactobacillus (p < 0.001) than those that consumed C-, C+, and PPC (Table 5).
With regards to post-weaning diarrhea, all piglets, regardless of the diet they consumed, presented diarrhea (p > 0.05) during the experimental period (Table 5); however, with mild to moderate severity. On average, the incidence was lower in weeks 1 and 2 (4.5 vs 6.5), whereas the severity had a slight increase in week 2 (1.7 vs 1.8).
4. Discussion
The combined effects of different stress factors induce functional and structural changes in the gastrointestinal tract during the weaning period [33], resulting in low feed intake, poor initial growth, weight loss, and diarrhea [4]. Furthermore, weaning is associated with increased expression of inflammatory cytokines in the intestine, which can contribute to anatomical and functional intestinal disorders [28].
Therefore, the weight loss observed in the present work could be considered “normal” or “expected.” This suggests that the presence or absence of antibiotics or functional feeds in the diets was insufficient to overcome the problems generated by weaning, resulting in the low performance observed in piglets during the first week post-weaning. However, piglets must adapt quickly to these changes during the post-weaning period, which is why in the following week, a significant increase in ADFI was observed, increasing from an average of 101.4 g/day in the first week to 261 g/day in the second week; that is, the piglets increased their feed intake 2.29-fold. Consequently, the ADG also increased significantly; the animals went from an average negative weight gain of -22 g/day across all treatment groups to 202 g/day.
In another study [20], animals treated with Saccharomyces boulardii RC009 in the first two weeks post-weaning showed no differences in their average daily gain (ADG) compared to the untreated group, however the probiotic influence on piglets occurred during the fourth post-weaning week. Maybe we finished our experiment before the functional foods could show its benefits on the growth performance.
At the macroscopic level, the relative weights of the digestive organs indicated that the harmful effects of weaning were not detrimental to visceral development. During lim-ited protein conditions, there is a preference to maintain the size of the diges-tive organs at the expense of other tissues because the deposition of proteins in the gastro-intestinal tract is preserved [34]. The relative weights of the digestive organs were like those observed in previous studies [35].
However, the results observed at the microscopic level, such as the greater width of the villi in the duodenum and greater size in the jejunum, revealed the positive effect of the functional foods PPC and SB, specifically when they were provided in combination and without antibiotics. This contributed to an increase in the digestion and absorption surface in these intestinal segments, counteracting the atrophy of the intestinal structure observed during the post-weaning phase [36]. The jejunum plays a significant role in digestive function because it is the most considerable portion of the small intestine [37]; in addition, most of the digestion of proteins and absorption of amino acids is performed in the proximal part of the jejunum, and a small portion reaches the ileum, where the digestion and absorption of amino acids is completed [38]. The greater abundance of occludins in the jejunum observed in animals in the PPC-SB experimental treatment confirms the positive effect of the combination of these two functional foods on intestinal structure. The effects of probiotics on the intestinal barrier are commonly studied in rodents; however, reports in pigs, especially in piglets, are scarce [39].
The use of yeast in piglet diets increased the abundance of occludins in the jejunum [40]. These proteins play central roles in sealing intercellular spaces [24]. Therefore, they serve as a barrier for the passage of undesirable substances through the paracellular spaces of enterocytes, maintaining intestinal integrity and preventing harmful agents and microorganisms from passing through them.
Some authors [40] claim that the integrity of the intestinal barrier is complex, and the collectivity of the elements that compose it (structure of tight junctions, mucus, microbiota, and immune system) maintains the health of animals at the local and systemic levels. These microscopic results are relevant since maintaining the integrity of the intestinal villi and the junctions between the cells that cover the villi should be an essential function of the feed ingested by newly weaned piglets as changes in histological parameters are one of the primary causes of decreased digestion and absorption of nutrients that contribute to post-weaning diarrhea [41].
Our results indicated that diarrhea was not associated with a specific diet, as all piglets presented with this digestive disorder during the experimental period. This is probably due to the low digestive capacity of the piglets and the lack of adaptation of the intestine to the new diets, which leaves many dietary substrates, especially of protein origin, to be metabolized by the microbiota of the large intestine. This can increase the levels of potentially toxic substances, such as amines, indoles, and phenols [42], as well as ammonia and branched-chain fatty acids [43], which have been implicated in the pathogenesis of post-weaning diarrhea. Fortunately, in the present study, diarrhea was of moderate severity and did not compromise the health of the animals, and they managed to recover their body weight in the second week post-weaning, and the development of the digestive organs was not affected.
Previous studies support the idea that functional foods can help reduce the incidence of diarrhea in post-weaning pigs [8]. However, the results vary between authors, and it is not conclusive. Some of them do not report a significant reduction in diarrhea as we reported in this work [20,35,44,45].
A sudden change in feed causes bacterial dysbiosis in the GIT with increased coli-form populations, which may explain the more significant quantities of CFUs of coliform/g feces observed in animals [5]. The abrupt reduction of Lactobacillus spp. at weaning, as mentioned by other authors [5,46], was also observed in animals fed the C and PPC diets. In the case of those fed SB alone or a combination of PPC at SB, a more significant number of CFUs of Lactobacillus/g of feces and a greater Lactobacillus:Coliform ratio (0.79:1, 0.70:1, and 0.47:1 for the animals) were observed for the SB, PPC-SB, and C- diets, respectively). The inclusion of antibiotics and, to a lesser extent, PPC controlled this increase in coliforms and had a slight effect on Lactobacillus spp (Lactobacillus:Coliform ratio of 0.72:1). The combined effect of PPC and SB is probably most related to the control of pathogenic bacteria by the antimicrobial peptides observed in PPC [9,14] and enhanced antioxidant activity, anti-inflammatory responses and improved intestinal microbial ecology by Saccharomyces boulardii [47], improving intestinal health. However, this mechanism requires further verification and the presence of these antimicrobial peptides in PPC needs to be confirmed.
Due to the functional properties of potato proteins described by different authors, better results were expected in animals that consumed PPC added individually to the diet. A probable explanation for this was that during the procedure for the extraction, separation and purification of PPC there were structural changes in these proteins and therefore they lost part of their functionality. High-performance precipitation methods are used to extract proteins from potato juice; however, these can alter their functional properties leading to partial or total loss of the protein functionality, which restricts the application of PPC in animal feed [48]. Recently, there has been increased interest in PPC, and many authors have investigated alternative treatments and modification of the technologies to eliminate this threat [11,49]. In future studies, a prior evaluation of the functionality of the PPC proteins is recommended.
Fortunately, the association PPC with SB yeast had positive results for intestinal health due to the better height of the jejunum villi, the higher concentration of occludins in this tissue and Lactobacillus in the feces of the piglets that consumed the mixture of these two functional foods. Other researchers also observed the ability of Saccharomyces to associate with other functional foods to generate benefits to the host. Piglets fed a dietary supplement with blood plasma and Saccharomyces cerevisiae over 0–21 days had a great impact on the growth performance, nutrient digestibility, fecal microbial, and gas emission at the end of the experiment [44]. On the other side, the combination of Pediococcus pentosaceus RC007 (a new probiotic) and Saccharomyces boulardii RC009 promoted healthier gut microbiota, with the reduced abundance of Proteobacteria and Cyanobacteria and could be a good substitute for antibiotics in improving pig production performance [45].
5. Conclusions
It was concluded that including a combination of PPC and SB in the diet without antibiotics did not negatively affect the performance or development of digestive organs in the post-weaning period. The PPC-SB combination prevented atrophy of the jejunal villi, increasing their length, and had a similar effect as the antibiotics. Saccharomyces boulardii, alone or in combination with PPC, stimulated an increase in Lactobacillus. Simultaneously, the addition of PPC and SB to the piglet diet promoted outstanding adhesion between enterocytes in the jejunum, similarly to antibiotics, reduced intestinal permeability, and protected the function of the intestinal barrier. Likewise, it reduced the probability that microorganisms would penetrate enterocytes and enter the blood, causing systemic infections. Therefore, including these functional feeds in piglet diets contributed to maintaining intestinal health in the critical post-weaning phase.
Author Contributions
Conceptualization, T.R.S.; methodology, T.R.S., G.M.L., and U.M.C.; formal analysis, G.M.L., and U.M.C.; investigation, T.H.V. and C.N.B.; resources, T.R.S.; data curation, G.M.L. and U.M.C.; writing—original draft preparation, T.R.S., J.G.S. and G.M.L.; writing—review and editing, T.R.S., J.G.S., and T.H.V.; visualization, T.R.S.; supervision, T.H.V.; project administration, T.R.S; funding acquisition, T.R.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by “FONDEC 2021 and FOPER 2023 projects” and “The APC was funded by University Autonomous of Querétaro”.
Institutional Review Board Statement
“The animal study protocol was approved by the Bioethical Committee of the Faculty of Natural Sciences of the Autonomous University of Querétaro (approval number: 63FCN2021)”.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting this study will be shared upon reasonable request from the corresponding authors.
Acknowledgments
We would like to thank the company Lallemand Animal Nutrition (Canada) for donating Saccharomyces boulardii. We also thank Dr. Santiaga Maricela Ahumada Solorzano and M.C. Sandra Patricia Rodríguez Jiménez for their advice during development of the immunofluorescence technique.
Conflicts of Interest
“The authors declare no conflicts of interest.”
References
- Pohl, C.S.; Medland, J.E.; Moeser, A.J. Early-life stress Origins of gastrointestinal disease: animal models, intestinal pathophysiology, and translational implications. Am J Physiol Gastrointest Liver Physiol 2015, 309, G927–G941. [Google Scholar]
- Pluske, J.R. Invited review: aspects of gastrointestinal tract growth and maturation in the pre-and postweaning period of pigs. J Anim Sci 2016, 94, 399–411. [Google Scholar]
- Siemińska, I.; Pejsak, Z. Impact of stress on the functioning of the immune system, swine health, and productivity. Med Weter 2022, 78, 541–547. [Google Scholar]
- Campbell, J.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J Anim Sci Biotechnol 2013, 4, 19–22. [Google Scholar]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet Scand 2017, 59, 1–19. [Google Scholar]
- Thacker, P.A. Alternatives to antibiotics as growth promoters for use in swine production: A review. J Anim Sci Biotechnol, 2013, 4, 1–12.
- Liu, H.Y.; Zhu, C.; Zhu, M.; Yuan, L.; Li, S.; Gu, F.; Hu, P.; Chen, S.; Cai, D. Alternatives to antibiotics in pig production: looking through the lens of immunophysiology. Stress Biol 2024, 4, 1–23. [Google Scholar] [CrossRef]
- de Souza, T.C.R. , Barreyro, A.A., Rubio, S.R., González, Y. M.; García, K. E.; Soto, J.G.G.; Mariscal-Landín, G. Growth performance, diarrhoea incidence, and nutrient digestibility in weaned piglets fed an antibiotic-free diet with dehydrated porcine plasma or potato protein concentrate. Ann Anim Sci.
- Parra-Alarcón, E.; de Jesús Hijuitl Valeriano, T.; Landín, G.M.; De Souza, T.C.R. Concentrado de proteína de papa: una posible alternativa al uso de antibióticos en las dietas para lechones destetados. revisión. Rev Mex Cienc Pecu.
- Beals, K.A. Potatoes, Nutrition and Health. Am. J. Potato Res 2019, 96, 102–110. [Google Scholar]
- Fu, Y.; Liu, W.N.; Soladoye, O.P. Towards potato protein utilisation: Insights into separation, functionality and bioactivity of patatin. Int J Food Sci Technol 2020, 55, 2314–2322. [Google Scholar]
- Galves, C.; Galli, G.; Kurozawa, L. Potato protein: Current review of structure, technological properties, and potential application on spray drying microencapsulation. Crit Rev Food Sci Nutr 2023, 6323, 6564–6579. [Google Scholar]
- Bhutto, R.A.; Khanal, S.; Wang, M.; Iqbal, S.; Fan, Y.; Yi, J. Potato protein as an emerging high-quality: Source, extraction, purification, properties functional, nutritional, physicochemical, and processing, applications, and challenges using potato protein. Food Hydrocoll 2024, 110415, 1–6. [Google Scholar]
- Escobar-García, K.; de Souza, T. C. R.; Díaz-Muñoz, M.; Bautista-Marín, S.E. Nivel dietético de concentrado de proteína de papa y su efecto sobre la concentración intestinal de citocinas y ácidos grasos volátiles en lechones destetados. Rev Mex Cienc Pecu 2025, 16, 1–235. [Google Scholar] [CrossRef]
- Bártová, V.; Bárta, J.; Jarošová, M. Antifungal and antimicrobial proteins and peptides of potato Solanum tuberosum L. tubers and their applications. Appl Microbiol Biotechnol 2019, 103, 5533–5547. [Google Scholar] [CrossRef]
- Sanders, M.E.; Benson, A.K.; Lebeer, S.; Merenstein, D.; Klaenhammer, T.R. Shared mechanisms among probiotic taxa: Implications for general probiotic claims. Curr Opin Biotechnol 2018, 49, 207–216. [Google Scholar] [CrossRef]
- Fijan, S. Probiotics and their antimicrobial effect. Microorganisms 2023, 112, 528–531. [Google Scholar] [CrossRef]
- Luise, D.; Spinelli, E.; Correa, F.; Nicodemo, A.; Bosi, P.; Trevisi, P. The effect of a single, early-life administration of a probiotic on piglet growth performance and faecal microbiota until weaning. Ital J Anim Sci 2021, 20, 1373–1385. [Google Scholar] [CrossRef]
- Pang, Y.; Zhang, H.; Wen, H.; Wan, H.; Wu, H.; Chen, Y.; Liu, X. Yeast probiotic and yeast products in enhancing livestock feeds utilization and performance: An overview. J Fungi 811, 1191–1202. [CrossRef]
- Parada, J.; Magnoli, A.; Isgro, M.C. , Poloni, V., Fochesato, A., Martínez, M.P., Carranza A., Cavaglieri, L. In-feed nutritional additive probiotic Saccharomyces boulardii RC009 can substitute for prophylactic antibiotics and improve the production and health of weaning pigs. Vet World, 1035. [Google Scholar]
- Alkalbani, N.S.; Osaili, T.M.; Al-Nabulsi, A.A.; Olaimat, A.N.; Liu, S.Q.; Shah, N.P.; Apostolopoulos, V.; Ayyash, M.M. Assessment of yeasts as potential probiotics: A review of gastrointestinal tract conditions and investigation methods. J Fungi 2022, 84, 365–387. [Google Scholar] [CrossRef]
- Touchette, K.J.; Carroll, J.A.; Allee, G.L.; Matteri, R.L.; Dyer, C.J.; Beausang, L.A.; Zannelli, M.E. Effect of spray-dried plasma and lipopolysaccharide exposure on weaned pigs: I. Effects on the immune axis of weaned pigs. J Anim Sci 2002, 802, 494–501. [Google Scholar] [CrossRef]
- Spreeuwenberg, M.A.M.; Verdonk, J.M.A.J.; Gaskins, H.R.; Verstegen, M.W.A. Small intestine epithelial barrier function is compromised in pigs with low feed intake at weaning. J Nutr 2001, 1315, 1520–1527. [Google Scholar] [CrossRef]
- Saitou, M.; Furuse, M.; Sasaki, H.; Schulzke, J.; Fromm, M.; Takano, H.; Noda, T.; Tsukita, S. Complex phenotype of mice lacking occludin, a component of tight junction strands. Mol Biol Cell 2000, 1112, 4131–4142. [Google Scholar] [CrossRef]
- CIOMS. International Guiding Principles for Biomedical Research Involving Animals. In: Organization WH editor. International Guiding Principles for Biomedical Research Involving Animals. Council for International Organization of Medical Sciences. Geneva; 1985.
- Diario Oficial de la Federación. Publication date: , 2001. Available online:http://legismex.mty.itesm.mx/normas/zoo/zoo062.pdf accessed on 09 august 2024. 22 August.
- NRC. Nutrient Requirements of Swine, 11th ed.; National Academic Press: Washington, DC, USA, 2012. [Google Scholar]
- Bautista-Marín, S.; Escobar-García, K.; Molina-Aguilar, C.; Mariscal-Landín, G.; Aguilera-Barreyro, A.; Díaz-Muñoz, M.; de Souza, T.R. Antibiotic-free diet supplemented with live yeasts decreases inflammatory markers in the ileum of weaned piglets. S Afr J Anim Sci 2020, 503, 353–365. [Google Scholar]
- Pita-López, W.; Gomez-Garay, M.; Blanco-Labra, A.; Aguilera-Barreyro, A.; Souza, T.C.R.; Olvera-Ramírez, A.M.; Ferriz-Martínez, R.A.; García-Gasca, T. Tepary bean Phaseolus acutifolius lectin fraction provokes reversible adverse effects on rats’ digestive tract. Toxicol Res 2020, 95, 714–725. [Google Scholar]
- Diario Oficial de la Federación. Publication date: , 1995. NOM-113-SSA1-1994. Bienes y servicios. C. Available online on http://www.ordenjuridico.gob.mx/Documentos/Federal/wo69536.pdf accessed on 09 august 2024. 25 August.
- De Man, J.C.; Rogosa, M.; Sharpe, M.E.A. Medium for the Cultivation of Lactobacilli. J Appl Bacteriol 1960, 23, 130–135. [Google Scholar]
- Ball, R.O.; Aherne, F.X. Influence of dietary nutrient density, level of feed intake and weaning age on young pigs. II. Apparent nutrient digestibility and incidence and severity of diarrhea. Can J Anim Sci 1987, 674, 1105–1115. [Google Scholar] [CrossRef]
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors influencing the structure and function of the small intestine in the weaned pig: a review. Livest Prod Sci 1997, 511, 215–236. [Google Scholar]
- Ebner, S.; Schoknecht, P.; Reeds, P.; Burrin, D. Growth and metabolism of gastrointestinal and skeletal muscle tissues in protein-malnourished neonatal pigs. Am J Physiol Regul Integr Comp Physiol 1994, 2666, R1736–R1743. [Google Scholar] [CrossRef]
- Hijuitl-Valeriano, T.J. Efecto del nivel de inclusión de concentrado de proteína de papa en la dieta de lechones sobre algunos indicadores de la salud intestinal. Master ina Helth and Produ, Universidad Autónoma de Querétaro. Querétaro, Qro: Universidad Autónoma de Querétaro; 2021. Available online: https://ri-ng.uaq.mx/browse?type=author&value=Teresita+De+Jesus+Hijuitl+Valeriano&value_lang=es_ES.
- Wang, M.; Wang, L.; Tan, X.; Wang, L. ; Xiong, X; Wang, Q.; Yang, H.; Yin, Y. The developmental changes in intestinal epithelial cell proliferation, differentiation, and shedding in weaning piglets. Anim Nutr.
- Schubert, D.C.; Mößeler, A.; Ahlfänger, B.; Langeheine, M.; Brehm, R.; Visscher, C.; El-Wahab, A.A.; Kamphues, J. Influences of exocrine pancreatic insufficiency on nutrient digestibility, growth parameters as well as anatomical and histological morphology of the intestine in a juvenile pig model. Front Med 9, 1–12.
- Silk, D. Digestion and Absorption of Dietary Protein in Man. Proc. Nutr. Soc 1980, 391, 61–70. [Google Scholar]
- Yang, F.; Wang, A.; Zeng, X.; Hou, C.; Liu, H.; Qiao, S. Lactobacillus reuteri I5007 modulates tight junction protein expression in IPEC-J2 cells with LPS stimulation and in newborn piglets under normal conditions. BMC Microbiol 2015, 151, 1–11. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, S.; Zhang, J.; Li, H.; Yang, D.; Huang, S.; Wei, Z.; Liang, X.; Wang, Z. Supplementation with yeast culture improves the integrity of intestinal tight junction proteins via NOD1/NF-ΚB P65 pathway in weaned piglets and H2O2-challenged IPEC-J2 cells. J Funct Foods 2020, 72, 104058. [Google Scholar]
- Barszca, M.; Skomiał, J. The development of the small intestine of piglets – chosen aspects. J Anim Feed Sci. 2011, 20, 3–15. [Google Scholar]
- De Lange, C.F.M.; Pluske, J.; Gong, J.; Nyachoti, C.M. Strategic use of feed ingredients and feed additives to stimulate gut health and development in young pigs. Livest Sci 2010, 134, 124–134. [Google Scholar]
- García, K.E.; de Souza, T.C.R.; Landín, G.M.; Barreyro, A.A.; Santos, M.G.B.; Soto, J.G.G. Microbial fermentation patterns, diarrhea incidence, and performance in weaned piglets fed a low protein diet supplemented with probiotics. FNS 2014, 518, 1776. [Google Scholar]
- Sampath, V.; Heon Baek, D.; Shanmugam, S.; Kim, I.H. H. Dietary inclusion of blood plasma with yeast Saccharomyces cerevisiae supplementation enhanced the growth performance, nutrient digestibility, Lactobacillus count, and reduced gas emissions in weaning pigs. Animals 2021, 113, 759–771. [Google Scholar]
- Parada, J.; Magnoli, A.; Poloni, V.; Corti Isgro, M.; Rosales Cavaglieri, L.; Luna, M. J.; Carranza, A.; Cavaglieri, L. Pediococcus pentosaceus RC007 and Saccharomyces boulardii RC009 as antibiotic alternatives for gut health in post-weaning pigs. J Appl Microbiol 2024, 135, lxae282. [Google Scholar]
- Gresse, R.; Chaucheyras Durand, F.; Dunière, L.; Blanquet-Diot, S.; Forano, E. Microbiota composition and functional profiling throughout the gastrointestinal tract of commercial weaning piglets. Microorganisms 2019, 79, 343–366. [Google Scholar]
- Zhang, W.; Bao, C.; Wang, J.; Zang, J.; Cao, Y. Administration of Saccharomyces boulardii mafic-1701 improves feed conversion ratio, promotes antioxidant capacity, alleviates intestinal inflammation and modulates gut microbiota in weaned piglets. J Anim Sci Biotechnol 2020, 11, 1–11. [Google Scholar]
- Hussain, M.; Qayum, A.; Xiuxiu, Z.; Liu, L.; Hussain, K.; Yue, P.; Yue, S. Potato protein: An emerging source of high quality and allergy free protein, and its possible future based products. Food Res. Int. 2021, 148, 110583. [Google Scholar]
- Hu, C.; He, Y.; Zhang, W.; He, J. Potato proteins for technical applications: Nutrition, isolation, modification and functional properties-A review. IFSET 2024, 91, 103533. [Google Scholar]
Figure 1.
Concentration of occludins in the piglet’s jejunum. a,bValues of bars with different letters differ significantly (P <0.05). C: Diet without antibiotics. C+ Diet supplemented with antibiotics. PPC: diet with potato protein concentrate. SB: diet with Saccharomyces boulardii. PPC-SB: diet with potato protein concentrate and Saccharomyces boulardii.
Figure 1.
Concentration of occludins in the piglet’s jejunum. a,bValues of bars with different letters differ significantly (P <0.05). C: Diet without antibiotics. C+ Diet supplemented with antibiotics. PPC: diet with potato protein concentrate. SB: diet with Saccharomyces boulardii. PPC-SB: diet with potato protein concentrate and Saccharomyces boulardii.

Figure 2.
Concentration of occludins in the piglet’s colon. C: Diet without antibiotics. C+ Diet supplemented with antibiotics. PPC: diet with potato protein concentrate. SB: diet with Saccharomyces boulardii. PPC-SB: diet with potato protein concentrate and Saccharomyces boulardii.
Figure 2.
Concentration of occludins in the piglet’s colon. C: Diet without antibiotics. C+ Diet supplemented with antibiotics. PPC: diet with potato protein concentrate. SB: diet with Saccharomyces boulardii. PPC-SB: diet with potato protein concentrate and Saccharomyces boulardii.
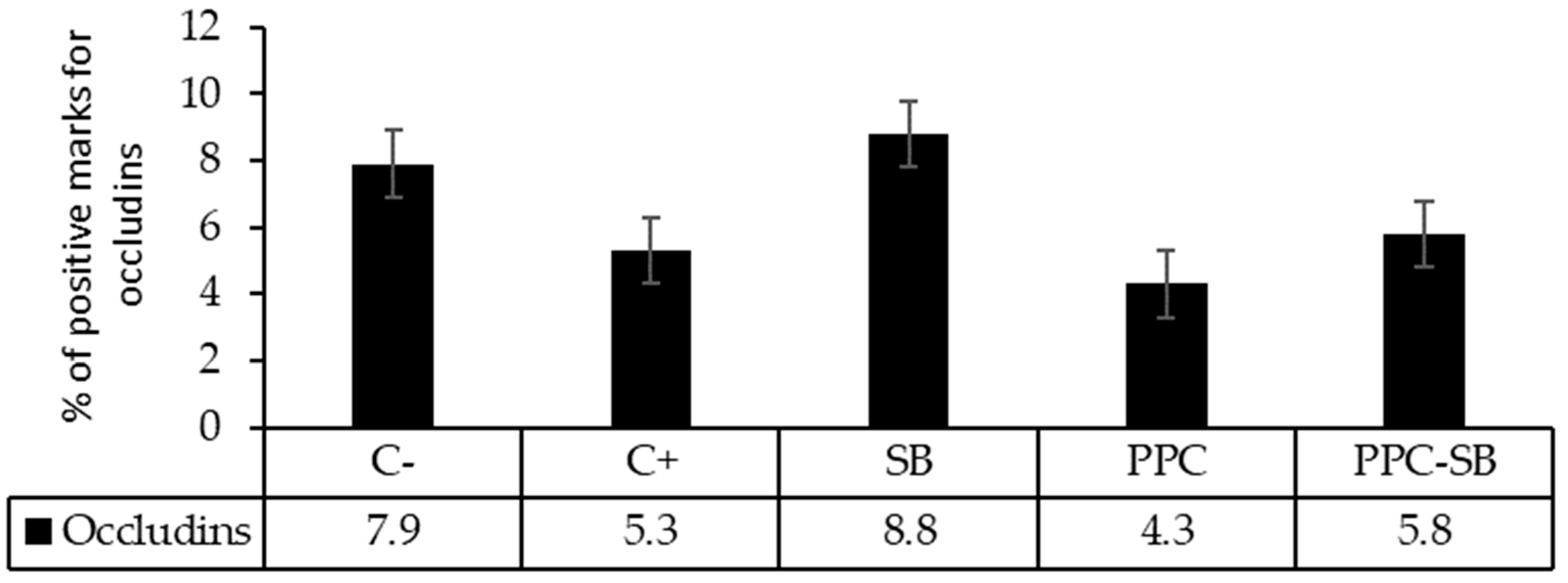

Table 1.
Centesimal composition of the experimental diets.
| Ingredients (%) | Experimental diets | ||||
|---|---|---|---|---|---|
| C- | C+ | SB | PPC | PPC-SB | |
| Maize | 44.7 | 44.56 | 44.68 | 43.78 | 43.77 |
| Soybean Meal | 12 | 12 | 12 | 12 | 12 |
| Soybean Isolate | 8.32 | 8.34 | 8.33 | 3.74 | 3.74 |
| Potato Protein Concentrate | 6 | 6 | |||
| Antibiótic1 | 0.05 | ||||
| Yeast2 | 0.01 | 0.01 | |||
| Menhaden Fish Meal | 5 | 5 | 5 | 5 | 5 |
| Sweet Whey Milk | 24.69 | 24.69 | 24.69 | 24.69 | 24.69 |
| Maize oil | 2.45 | 2.52 | 2.45 | 2.23 | 2.23 |
| L-Lysine HCl | 0.4 | 0.4 | 0.4 | 0.27 | 0.27 |
| L-Threonine | 0.12 | 0.12 | 0.12 | 0.02 | 0.02 |
| DL-Methionine | 0.19 | 0.19 | 0.19 | 0.14 | 0.14 |
| L-Tryptophan | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 |
| L-Valine | 0.02 | 0.02 | 0.02 | ||
| Salt | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 |
| Calcium Carbonate | 0.6 | 0.6 | 0.6 | 0.54 | 0.54 |
| Dicalcium Phosphate | 0.61 | 0.61 | 0.61 | 0.69 | 0.69 |
| Titanium Dioxide | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 |
| Vitamins Premix3 | 0.07 | 0.07 | 0.07 | 0.07 | 0.07 |
| Minerals Premix4 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
C: Diet without antibiotics. C+ Diet supplemented with antibiotics. PPC: diet with potato protein concentrate. SB: diet with Saccharomyces boulardii. PPC-SB: diet with potato protein concentrate and Saccharomyces boulardii. 1Linco Spectin premix: 2.2 g lincomycin, 2.2 g spectinomycin (Zoetis, USA). 2Yeast: Saccharomyces boulardii (Levucell SB, CNCM I-1079, Lallemand Animal Nutrition, Canadá). 3Per kg feed: CaCO3,1.43 mg; CuSO4.5H2O, 55.5 mg; FeSO4.H2O, 333.3 mg; C2H8N22HI, 1.01 mg; MnSO4.H2O, 135 mg; Na2SeO3, 0.5 mg; ZnSO4.H2O, 338 mg. 4Per kg feed: vitamin A, 6563 IU; vitamin D3, 893 IU; vitamin E, 33.5 IU; vitamin K, 1.2 mg; riboflavin, 3.6 mg; vitamin B12,18 μg; choline, 356 mg; niacin, 17 mg; pantothenic acid, 14 mg; thiamine, 1.4 mg; pyridoxine, 2.8 mg; biotin, 127 μg; folic acid, 0.9 mg.
Table 2.
Piglets’ performance parameters.
| Items | Experimental diets | p | SEM | ||||
|---|---|---|---|---|---|---|---|
| C- | C+ | SB | PPC | PPC-SB | |||
| ADFI (g/day) | |||||||
| Week 1 | 105 | 96 | 98 | 103 | 105 | NS | 3.1 |
| Week 2 | 255 | 258 | 256 | 265 | 271 | NS | 5.3 |
| ADG (g/day) | |||||||
| Week 1 | -28 | -21 | -30 | -13 | -15 | NS | 4.4 |
| Week 2 | 210 | 230 | 215 | 219 | 224 | NS | 5.4 |
| FE | |||||||
| Week 1 | -0.270 | 0.249 | -0.345 | -0.194 | -0.191 | NS | 0.052 |
| Week 2 | 0.821 | 0.914 | 0.835 | 0.830 | 0.833 | NS | 0.015 |
C: Diet without antibiotics. C+ Diet supplemented with antibiotics. PPC: diet with potato protein concentrate. SB: diet with Saccharomyces boulardii. PPC-SB: diet with potato protein concentrate and Saccharomyces boulardii. 1Linco Spectin premix: 2.2 g lincomycin, 2.2 g spectinomycin (Zoetis, USA). 2Yeast: Saccharomyces boulardii (Levucell SB, CNCM I-1079, Lallemand Animal Nutrition, Canadá). ADFI: Average daily feed intake. ADG: Average daily gain. FE: Feed efficiency. p: Probability. SEM: Standard error of the mean. NS: Non-Significant.
Table 3.
Morphophysiological characteristics of the digestive system.
| Items | Experimental diets | p | SEM | ||||
|---|---|---|---|---|---|---|---|
| C- | C+ | SB | PPC | PPC-SB | |||
| Body weight (Kg) | 8.100 | 8.527 | 7.715 | 8.238 | 7.933 | NS | 0.177 |
| Relative body weight (g*BW-1) | |||||||
| Pancreas | 2.2 | 2.1 | 1.9 | 2.3 | 2.1 | NS | 0.065 |
| Liver | 29 | 32 | 30 | 33 | 28 | NS | 0.818 |
| Estomach | 8.8 | 8.1 | 7.0 | 7.9 | 7.9 | NS | 0.312 |
| Small Intestine | 61 | 57 | 58 | 65 | 60 | NS | 1.285 |
| Large Intestine | 17.7 | 19.8 | 18.1 | 18.2 | 19.8 | NS | 0.794 |
| pH of Contents | |||||||
| Estomach | 2.8 | 3.7 | 4.1 | 3.5 | 3.7 | NS | 0.163 |
| Yeyune | 5.4 | 5.5 | 5.6 | 5.5 | 5.7 | NS | 0.064 |
| Ileum | 5.9 | 6.0 | 6.1 | 5.8 | 5.9 | NS | 0.081 |
| Ceacum | 5.4 | 5.4 | 5.4 | 5.4 | 5.5 | NS | 0.061 |
| Colon | 5.5 | 5.5 | 5.5 | 5.4 | 5.6 | NS | 0.047 |
C: Diet without antibiotics. C+ Diet supplemented with antibiotics. PPC: diet with potato protein concentrate. SB: diet with Saccharomyces boulardii. PPC-SB: diet with potato protein concentrate and Saccharomyces boulardii. ADFI: Average daily feed intake. ADG: Average daily gain. FE: Feed efficiency. p: Probability. SEM: Standard error of the mean. NS: Non-Significant.
Table 4.
Morphology of intestinal villi and crypts.
| Items | Experimental diets | p | SEM | ||||
|---|---|---|---|---|---|---|---|
| C- | C+ | SB | PPC | PPC-SB | |||
| Duodenum | |||||||
| VH (μm) | 344 | 369 | 363 | 331 | 375 | NS | 6.8 |
| VW (μm) | 115bc | 130ab | 138a | 108c | 138a | 0.009 | 2.7 |
| CD (μm) | 197 | 223 | 213 | 227 | 240 | NS | 8.3 |
| Jejunum | NS | ||||||
| VH (μm) | 378b | 439a | 337b | 343b | 435a | 0.003 | 8.0 |
| VW (μm) | 112 | 123 | 111 | 94 | 109 | NS | 4.9 |
| CD (μm) | 236 | 247 | 202 | 244 | 234 | NS | 6.7 |
| Ileum | |||||||
| VH (μm) | 298 | 331 | 332 | 366 | 361 | NS | 9.9 |
| VW (μm) | 108 | 111 | 107 | 117 | 116 | NS | 3.1 |
| CD (μm) | 208 | 187 | 194 | 203 | 214 | NS | 5.3 |
| Colon CD (μm) | 320 | 312 | 305 | 299 | 337 | NS | 6.3 |
C: Diet without antibiotics. C+ Diet supplemented with antibiotics. PPC: diet with potato protein concentrate. SB: diet with Saccharomyces boulardii. PPC-SB: diet with potato protein concentrate and Saccharomyces boulardii. VH: villus height. VW: Villus width. CD: Crypt depth. SEM: standard error of the mean. p: Probability. NS: Non-Significant.
Table 5.
Bacteria population and diarrhea incidence and severity.
| Items | Experimental diets | p | SEM | ||||
|---|---|---|---|---|---|---|---|
| C- | C+ | SB | PPC | PPC-SB | |||
| Coliforms (UFC/g) | 7.4a | 5.7b | 7.5a | 7.2ab | 8.1a | 0.04 | 0.23 |
| Lactobacillus (UFC/g) | 3.5b | 4.1b | 5.9a | 3.8b | 5.7a | 0.001 | 0.19 |
| ID | |||||||
| Week 1 (days) | 4.6 | 4.1 | 4.3 | 4.9 | 4.8 | NS | 0.14 |
| Week 2 (days) | 1.8 | 1.7 | 1.6 | 1.7 | 1.8 | NS | 0.05 |
| SD | |||||||
| Week 1 | 6.3 | 6.5 | 6.6 | 6.5 | 6.4 | NS | 0.12 |
| Week 2 | 1.7 | 1.9 | 1.9 | 1.9 | 1.7 | NS | 0.06 |
C: Diet without antibiotics. C+ Diet supplemented with antibiotics. PPC: diet with potato protein concentrate. SB: diet with Saccharomyces boulardii. PPC-SB: diet with potato protein concentrate and Saccharomyces boulardii. SEM: standard error of the mean. p: Probability. NS: Non-Significant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



