
Submitted:
24 February 2025
Posted:
25 February 2025
You are already at the latest version
Abstract
Anatomical abnormalities in octopuses, whose behavior is facilitated by flexible neuron-rich arms, offer insights into life histories and the neurological implications of understudied conditions such as bifurcation. Although documentation is scarce, here we present in-situ videos of 9-armed O. vulgaris with a functional bifurcated R1 arm. Analysis using RDAs and GLMs investigated the impact of the bifurcated arm on behavior and examined changes during growth. Analysis revealed a differential usage of between the bifurcated arms in addition to an initial specialization of the bifurcated arms for actions below the body, decreasing overtime as the arms grew. Further, bifurcated and regrown arms were utilized more in safe behaviors than risky ones, with more severely injured arms showing a higher frequency of use in safe behaviors. These findings contribute to the growing knowledge of arm usage in octopuses, suggesting that arm bifurcation may lead to branchial neural differentiation and potentially indicate post-traumatic associated in O. vulgaris.

Keywords:
cephalopods
; Octopus vulgaris
; 9-armed Octopus
; bifurcation
; arm use
; behavior
; adaptability
; pain-associated memory
1. Introduction
The molluscan class Cephalopoda (Cuvier, 1795), and particularly individuals within the order Octopoda (Leach, 1818), have been celebrated for their complex body patterning and unique morphology, including their regeneratable arms. Despite considerable research on the regeneration process, the specific molecular pathways and morphological modifications involved remain unclear [1,2,3]. Occasionally, during regeneration, octopuses’ arms, particularly their tips, undergo splitting and branching in their growth, causing conditions such as bifurcation, wherein the arm branches once producing two arms in place of one. Although it is still uncertain, the molecular mechanism for this abnormal growth has been hypothesized as being due to a mutation in the hox group of the homeobox genes [4,5]. Homeobox genes direct the formation of many body structures in early development, while the expression of the hox group provides the basis for the anterior versus posterior axis specification [6]. In invertebrates, the mutation of these genes can lead to the growth of an extra and typically non-functional body part [7]. An alternative molecular pathway associated with abnormal growth in Cephalopods is the Hedgehog (Hh) signaling pathway [8]. Present in both embryonic and adult tissues, it similarly mediates processes such as development, morphogenesis, and growth [9]. This abnormal growth was noted during the transplantation of tissue expressing the Hh gene family onto the posterior side of a stage 17 limb bud in Sepia officinalis which, within 7 of the 14 trials, resulted in a posteriorly duplicated limb [8]. Despite the identification of these mechanisms, as so few individuals with arm abnormalities have been recorded, with even fewer found alive, very little is still known about this process. Nevertheless, as the arms of an octopus help contribute to their incredible behavioral flexibility, the addition of an extra appendage has been speculated as altering the complex suite of behaviors [10].
Although the study of this behavioral suite is comprehensive, recent reviews have pointed to a need to study individuals in-situ or within the field, as current methods use ex-situ or laboratory experiments to examine behavior [11,12]. These approaches cause concerns in providing environmental context to observed behaviors. To help fill the gaps in environmentally relevant studies and to describe and analyze this rare abnormality, this study presents videos of a living cephalopod with a fully functional bifurcated arm in the wild. This paper aims to employ behavioral quantification methods to describe and examine the behavioral repertoire of this unique individual via videos collected by citizen science. Special attention was directed towards behaviors and actions involving arm usage, and a particular emphasis was placed on the two bifurcated arms. The research questions investigated are as follows: 1) does the presence of a fully functional bifurcated arm alter the behavior of Octopus vulgaris; 2) To what extent is the bifurcated arm used in different behaviors; 3) Does the usage of the arms change over time.
2. Materials and Methods
2.1. Study’s subject
All videos in this study are of the same male O. vulgaris. It was determined to be male based on photo analysis showing a hectocotylized R3 arm. (Fig. 1A). Although its cause was not captured on video this individual was presumed to have lost arms R1, R2, R3, and tips of R4 and L1 in a potential past predator encounter. During regrowth arm R1 bifurcated producing arms R1a and R1b, while all other arms regrew normally (Fig. 1B).
Figure 1.
(A) Photo taken on May. 02, 2022, showing the hectocotylized R3 arm (blue) and a normal arm tip (pink). (B) Photo from Jan. 28, 2022, displaying terminology used for arms, with bifurcated R1 arms in orange.
Figure 1.
(A) Photo taken on May. 02, 2022, showing the hectocotylized R3 arm (blue) and a normal arm tip (pink). (B) Photo from Jan. 28, 2022, displaying terminology used for arms, with bifurcated R1 arms in orange.

2.2. Quantification of behavioral data
Using the open-source software Behavioural Observation Interactive Research Software (BORIS)[13], the occurrence and duration of behaviors were analyzed from >10 hours of video records (23 videos). To do this, an ethogram created using behaviors previously noted in various Octopus species (Table S2) was inputted into the software and specific keys were associated with each of the actions and behaviors in the ethogram. This allowed each behavior to be recorded as a singular event (from the beginning to the finish of a concrete behavior) within the entire observation of each video. Events were classified into two types based on their association with duration: point events (without durations) and state events (with durations). In this sense, inside each of the 24 videos, the events were identified and the usage/non-usage of each of the individual’s 9 arms were registered through a binomial response variable matrix. When new or altered behaviors not previously noted in the catalog were observed, they were later added to the ethogram for further consideration. Only video segments where a minimum of 3 arms were visible for the entire behavioral sequence were analyzed (see Video S1 & S2 for examples of usable and unusable segments). Videos were analyzed at a maximum speed of 50 Frames Per Second (FPS), often frame by frame, to approximate arm movements accurately. Time budget data, exported as .csv files from BORIS, were used to create the final dataset.
2.3. Data manipulation and exploration
BORIS-generated outputs underwent data manipulation and exploration using R Studio version 2022.07.2. A binomial response variable matrix was crafted to signify arm usage (1) or non-usage (0) in identified events across the 23 videos. The video record dates were transformed into a continuous variable (days) for testing arm usage over time (with video 1: day 1 to video 24:147 days). Behaviors were categorized into specific groups (Behavior Category) to address scientific questions about arm use (Table S1). Safe behaviors involved those wherein the arms were held closer to the body [14] while Risky behaviors included actions with potential predator interactions and arm extensions away from the body.
After preliminary data inspection and visualization via triplots, noise or irrelevant behaviors were excluded based on three criteria: (1) relevance to arm use and scientific questions, (2) having more than 9 total events in all videos, and (3) completion in all videos. A new row for arm R1 was created by combining and correcting arms R1a and R1b for cohesive results, facilitating comparisons with past and future studies of arm usage in O. vulgaris, with or without bifurcated arms. In cases where behavioral categories showed no distinct trends for R1a versus R1b, R1 was used for analysis.
2.4. Preliminary analysis and arm groupings
Arm usage and association within specific behaviors involving the scientific questions were analyzed and compared for each individual arm through the metrics of the total number of occurrences and the total duration of each arm within behavioral categories previously determined to address scientific questions. The metrics were later confirmed using a Redundancy Analysis (RDA) for all behaviors and the specific behavioral categories. From the general RDA on all behaviors, the even distribution of events associated with Foraging, Exploration, and Locomotion facilitated the combination of bifurcated arms into R1 during the specific RDA’s for these categories. For Under Web Actions and Safe vs Risky behaviors, R1a and R1b were analyzed separately as these seemed to show differentiation in usage when an RDA was run with only bifurcated arms. When fitting Generalized Linear Models (GLMs) R1a and R1b were run separately for each behavioral category, besides Locomotion as there were no expected variations in usage between the two arms.
2.5. Statistical analysis
Various Redundancy Analysis (RDA; [15]) were used to visualize and test potential associations among the arms usage with behavior and behavioral categories. A preliminary RDA was run and visualized using a correlation triplot which included all behaviors, except those which did not meet the criteria previously explained. Following this initial step, additional behaviors were filtered out due to their high autocorrelation with behaviors more central to the research questions.
Before the RDA was run all events were aggregated by behavior and time, which produced a table wherein total counts for behavior within each video were made. The RDA was completed using vegan’s rda function [16], with behaviors and behavior categories as explanatory variables, and the matrix of arm usage by event in each video as a response variable (transformed using the Hellinger-transform method; [15,17]. The potential variance conferred by time, as days after the first videos was controlled by including it in the model as a condition factor.
The model was first assessed by the examination of the bimultivariate redundancy statistic - the amount of variance of the response matrix explained by the explanatory variables - equivalent to the R2 value in multiple regression [15]. However, the bimultivariate redundancy statistic of an RDA is biased like the ordinary R2 of multiple regression, and for the same reason [18]. This bias was fixed by adjusting the bimultivariate redundancy statistic using Ezekiel’s formula achieved through vegan’s function RsquareAdj [16,19]. The significant association of behaviors with arms usage was assessed using permutation tests implemented in the anova function. Lastly, the adequacy of the fitted RDA model was confirmed by assessing the linear relationships among explanatory variables using variance inflation factors (VIF; [15]).
In addition, binomial regression via Generalized Linear Models (GLMs, fitted using lme4 library in R software; [20], were used to test for potential changes in arm use over time and differences between probability of arm usage between the bifurcated arms. To conduct this, the binomial response of each arm (used/unused) was individually tested for specific behaviors or behavior categories in the function of time (from day 1 to day 147) that related to the scientific questions. Shapiro-Wilk tests were additionally used to assess the normality of the residuals. To visualize the data and test for differences between bifurcated arms, Logistic Regression Plots were used which were then compared using a two-sample z-test for proportions.
3. Results
Videos were recorded from December 13th, 2021, to May 8th, 2022, with distribution across the months as follows: December (4.17%), January (16.67%), February (37.5%), March (25%), April (12.5%), and May (4.17%) (Table 1). All the data used in the current study were collected within Ibiza, Balearic Islands, Spain.
Of these videos, the total video duration ranged from 177 to 2686.10 seconds. As not all the content in each video was usable due to the arms not being visible, the percentage analyzed differed with a minimum of 6.4% analyzed in video 13 (ultimately removed due to its low usability) and a maximum of 98.8% analyzed in video 21 (mean; 84.9%). This led to 6,642 events for analysis (Table 1).
3.1. Arm usage and their behavioral associations
When examined within all recorded events, although increased usage of anterior arms over posterior arms was detected, R3 was favored in its occurrence of use over L2 (2,472 vs 2,425 times used) albeit with a shorter duration (1,423.7 vs 1,521.3 s) (Fig. S1). On a sagittal plane, right-side arms were favored with both higher occurrences and durations of use (12,061 vs 8,051 times used, respectively). Finally, the arms with the highest usage were L1 and R1a (Fig. S1).
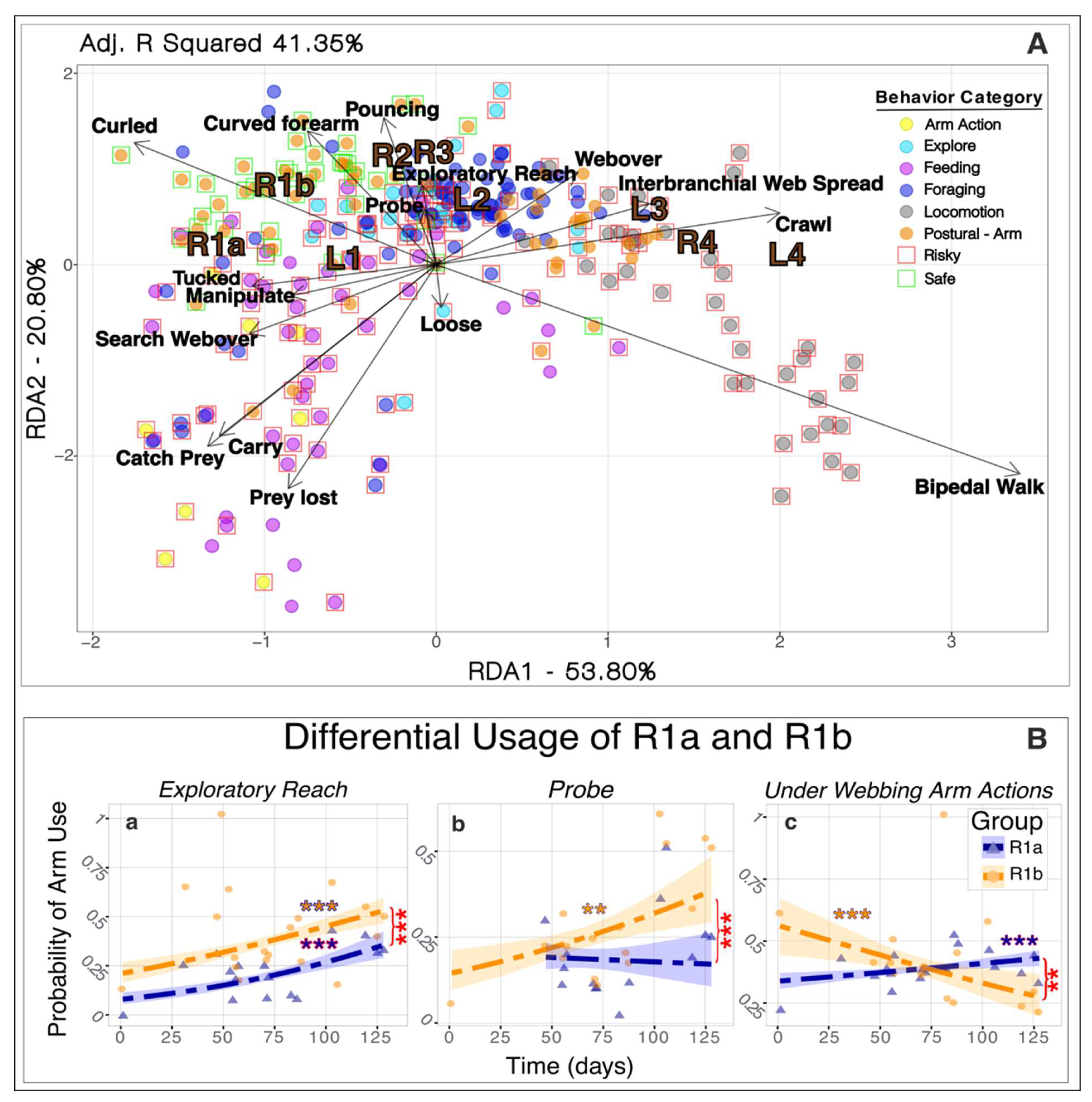
Redundancy Analysis for behaviors related to the scientific questions significantly demonstrated associations of behaviors and their categories with arms (RDA, P<0.001; Fig 2A). The RDA explained an adjusted 41.35% of the total variance of the arm usage response matrix. Of this percentage, 53.80% was explained by the first axis (RDA1), while 20.80% was explained by the second axis (RDA2), resulting in a total of 76% explained by the first two axes. Locomotion behaviors, particularly Bipedal Walk and Crawl, were more closely associated with the posterior arms, particularly L4 and R4. Bipedal Walk explained most of the variation along the first axis, suggesting its nearly exclusive use by the back two arms, while crawling was linked to other predominantly posterior arms. Foraging and Exploratory behaviors, namely, Exploratory Reach, Probe, and Pouncing were all associated with R2, L2, R3, L2, and to a lesser extent R1a and R1b; with R1b showing a significantly higher probability of use over time for Exploratory Reach and Probe than R1a (GLM, P<0.001; Fig. 2B, a-b). Feeding behaviors and behaviors that occurred underneath the webbing, especially Manipulate, were associated with the bifurcated arms and L1, though favoring R1a and L1 to R1b with a significant difference between the probability of use overtime between R1a and R1b (GLM, P<0.001; Fig 2B, c). In the postural categories, Arm(s) Curled, and Arm(s) Curved showed a heightened preference for using the bifurcated arms, however, its distribution throughout the graph is wide (Fig. 2A).
3.2. Risky and safe usage
Analysis of usage revealed that right-sided arms displayed a cautious tendency, strongly associated with Safe Behaviors, while left arms were linked to Risky Behaviors. RDA’s for the Safe Behaviors showed a significant association for both right and left-side arms (RDA, P<0.001; Fig. 2A). However, the adjusted bimultivariate redundancy statistic values (representing the amount of variance of the response matrix explained by the explanatory variables), differed exhibiting a greater explanation of the model for the right-side arms (40%) than the left-side arms (18%). This association was further exemplified via the occurrence and duration of arms within Safe Behaviors with the right side showing greater values than the left for every arm except R4 (Fig. S2A).
Figure 2.
(A) Association of filtered behaviors with arms (R1a, R1b, R2, R3, L1, L2, L3, L4). Arrows indicate the direction and magnitude of behaviors associated with arms, while each point represents events aggregated by behavioral categories. Behavioral categories are denoted by colored circles, while risky and safe events are denoted by red and green boxes. (B) Differential usage of bifurcated arms over time (in days) with logistic regression model fit for selected behaviors (a-c). Orange and blue asterisks represent significant regression curves between respective arms and use over time and the shaded areas indicate the confidence intervals of the model. Red asterisks represent significant differences between each arm’s probability of use within the respective behaviors. The significance level is represented by an asterisk as follows: ** P<0.001, ***P< 0.0001.
Figure 2.
(A) Association of filtered behaviors with arms (R1a, R1b, R2, R3, L1, L2, L3, L4). Arrows indicate the direction and magnitude of behaviors associated with arms, while each point represents events aggregated by behavioral categories. Behavioral categories are denoted by colored circles, while risky and safe events are denoted by red and green boxes. (B) Differential usage of bifurcated arms over time (in days) with logistic regression model fit for selected behaviors (a-c). Orange and blue asterisks represent significant regression curves between respective arms and use over time and the shaded areas indicate the confidence intervals of the model. Red asterisks represent significant differences between each arm’s probability of use within the respective behaviors. The significance level is represented by an asterisk as follows: ** P<0.001, ***P< 0.0001.
3.3. Bifurcated arms’ specialized use
The association of Under Web Actions with bifurcated arms was observed in the general RDA (Fig. 2A). Analysis of actions involving arm movements under the webbing—Search Webover, Manipulate, and Arm(s)-Tucked Under—revealed a preference for anterior arms, except for arm R2. Arm R1a was the most frequently used (414 observations), followed by R1b (302 observations) and anterior left arms (Fig. S2B). Arm L4 was the least utilized posterior arm (37 observations). The use of arm R1a in these behaviors significantly increased (GLM, P<0.001; Fig. 2B, c) as the arm grew over time, while significantly decreasing over time for R1b (GLM, P<0.001; Fig. 2B, c).
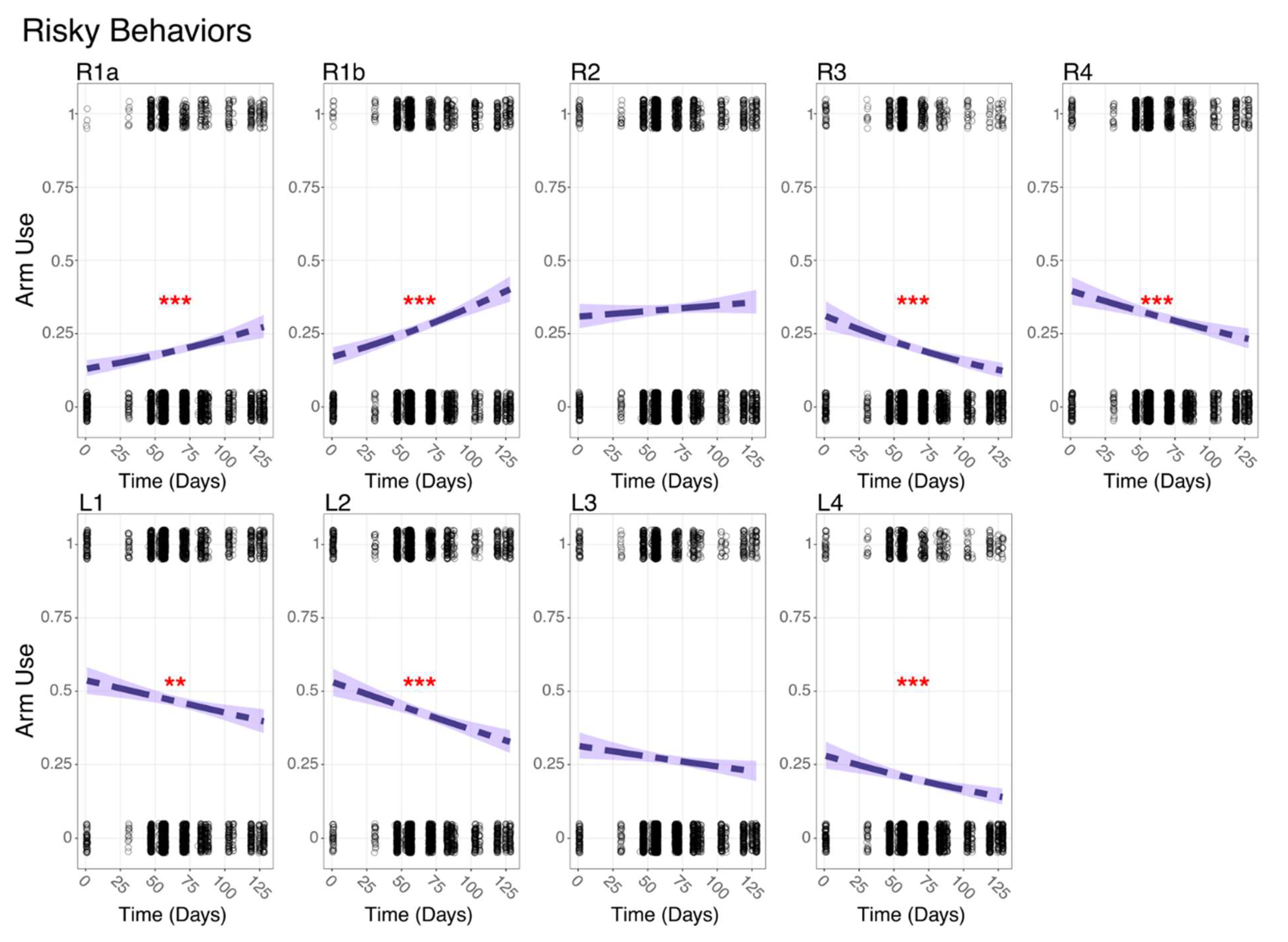
Figure 3.
Summary plots of Risky Behavior usage over time for each arm. The logistic regression models were fitted to assess the change in behavior usage over time. Points represent observed data, and shaded areas represent the confidence intervals of the fitted models. The significance level is represented by an asterisk as follows: ** P≤0.001, ***P≤ 0.0001.
Figure 3.
Summary plots of Risky Behavior usage over time for each arm. The logistic regression models were fitted to assess the change in behavior usage over time. Points represent observed data, and shaded areas represent the confidence intervals of the fitted models. The significance level is represented by an asterisk as follows: ** P≤0.001, ***P≤ 0.0001.
3.4. Behavioral change over time
In Risky Behaviors, characterized by extended arm use or interactions with other species, R1a and R1b showed a significant increase in usage over time (GLM, P<0.001; Fig. 3), while R2 decreased without statistical significance. All other arms, except L3 whose decrease was unsignificant, exhibited a significant decrease in usage in Risky Behaviors over time (GLM, P<0.001; Fig. 3). In Safe Behaviors, the anterior arms on octopus's right side (R1a, R1b and R2) exhibited significant decreases in usage over time (GLM, P<0.0001; Fig S3), while the posterior arms showed no significant change. On the left side, only L4 demonstrated a significant increase over time (GLM, P<0.001; Fig. S3).
3.5. Behavioral variations and undescribed behaviors
During video analysis, modifications to previously described behaviors were observed. The protective posture Retroflex [21,22], characterized by upturned arms, showed an alteration in this individual where bifurcated arms were crossed to form an X, termed Retroflex X (Fig. S4). Additionally, adjustments were noted in the feeding behavior of Web-Overs [23] also referred to as Envelope [24]. Due to the lack of intrabronchial webbing development between bifurcated arms, creating a gap in Web-Overs, the studied individual adapted Web-Overs tactics. This involved tucking bifurcated arms underneath the body and the crossing of arms L1 and R2 over the bifurcated region or the repositioning of bifurcated arms for optimal coverage (Fig. S5; Video S3). Notably, these adjustments were largely employed after a large fish escaped through the gap created by the shorter bifurcated arms during a non-adjusted Web-Over (Video S4).
4. Discussion
Although arm bifurcation has been morphologically described, to the author’s knowledge this study presents the best-recorded case of any living cephalopod species with bifurcated arms and the only case wherein the functionality of the bifurcated arms is described. Using observations from an octopus in the wild, we have shown that the usage of bifurcated cephalopod arms follows patterns observed in normal arms, while also displaying slightly specialized usage which has been hypothesized to be related to their smaller initial size. Further, there existed differentiated usage of the two bifurcated arms, with R1a being used more in Feeding Behaviors and R1b being used more in Exploratory Behaviors. These results might allude to a degree of pain-associated memory and that bifurcation may potentially lead to branchial neural differentiation in the affected arms.
4.1. Patterns of arm use
Despite being morphologically and structurally similar, patterns of association between octopus’ behaviors and certain arms have been previously described [22,25,26]. Authors have noted specialized functionality within stereotypic motor-programs [27,28,29], which constitute the majority of the actions made by the seemingly equipotential 8 arms. The primary research question aimed to assess the impact of a bifurcated arm on behavioral associations amongst arms. All patterns of usage, besides Risky and Safe usage, showed predominant differences when examined on a posterior-anterior axis. In the context of locomotion, the individual exhibited a consistent preference for posterior arms, aligning with prior studies [21,22,31-33]. However, in Foraging, Feeding, and Exploration, there was no clear distinction between anterior and posterior arms, despite increased anterior arm usage. Further analyses yielded contrasting results, necessitating additional tests for confirmation. This variability in anterior and posterior arm usage may be attributed to the heightened use of R3 in exploratory reach and a right-side distribution of usage. Contrary to previous findings [14,30,34], anterior arms like R1a and R1b were initially not associated with reaches but were utilized for actions closer to or underneath the body (Video S5). Instead, R3 compensated for the smaller bifurcated arms, particularly in Foraging and Exploratory behaviors, displayed by its increased initial usage. Over time, as R1a and R1b grew, their usage increased in Exploration and Foraging Behaviors, while R3’s usage within Exploration decreased (Video S6). The specialized nature of the hectocotylized R3 arm, in conjunction with these findings, may suggest that food accumulation outweighed the potential reproductive hindrance associated with the loss of R3. These findings differ from the results observed by earlier studies on arm loss in octopuses, where male octopuses often hold their R3 arms closer to their body and use them less in Risky actions [35,36]. Thus, there might exist anatomical and behavioral distinctions in younger octopuses like the one studied, prioritizing growth over mating.
From the analysis of behavior and arm association conducted in this study, we can corroborate previous findings that octopods, even those with abnormal limb assemblages, simplify the usage of arms by associating them with behavioral categories [22,25,30]. Specifically, this individual demonstrated to slightly favor anterior arms when involved in tasks related to foraging or reaching, and posterior arms for actions involving benthic locomotion. A behavioral confirmation noted as being important in laboratory studies due to a lack of environmental relevance [30]. These patterns of association provide additional evidence of the slight specialization that exists within seemingly equipotential arms. Furthermore, the choice of specialization indeed seems to be able to change to best fit the needs of the individual, indicating a heightened degree of individualistic behavioral flexibility. Here the subjects' decreased usage of bifurcated arms in under-webbing actions, decreased use of R3 and left-sided arms in Risky actions, and increased usage of bifurcated arms in Risky actions over-time exhibit this personalization.
In 8 armed octopuses, the mirroring of limb specialization on a bilaterally symmetrical axis has been shown; with the R2 and L2 arms being dominant for visually evoked prey capture [25] and R4 and L4 being preferentially used during Locomotion behaviors such as Bipedal Walking [32,37,38]. As two R1 arms are present in this individual, normal assumptions of arm relatedness were examined. It was hypothesized that R1a and L1 would be the most functionally similar due to their positioning on the body. However, R1b was shown to be more closely related in its usage to L1 rather than R1a. R1a exhibited lower usage in Exploratory Reach compared to other anterior arms, exhibited well in its nonuse during an interaction with a hermit crab with 3 anemones on its shell - note the curling of R1a as all adjacent arms reach out (Video S7). Instead, R1a demonstrated specialization in Feeding Behaviors, with increased utilization beneath the body and webbing. This suggests that while R1b may function as a typical arm in an 8-armed octopus, the unique anatomy of this individual led to R1a being utilized as a specialized tool.
The continual investigation of specific axial nerve cord transmission pathways has highlighted the bidirectionality of the Octopus’s nervous system, with the bulk of motor control responsibility embedded within the neural circuitry of the arms [28,29,39,40]. Given this, the presence of an extra appendage could surely alter the signals being communicated between the central and peripheral sensory systems. Further, as the brain does not support somatotopic representation, to achieve arm-specific actions local proprioceptive and tactile input from the periphery is integrated with specific central commands to activate embedded motor programs within the arm site labeled by the initial peripheral input [41,42]. Thus, the distinctive usage of R1a and R1b could stem from the differentiation of the branchial nerve or elsewhere along the axial nerve cord as specific central commands would have been integrated within two unique peripheral sites, generating individual motor programs and thus different behaviors.
4.2. Persistence of memory
The significant association of arms R1a, R1b, R2, and R3 in Safe Behaviors, best exemplified through the main posture taken by the R1 arms (Fig. 4A and B), and their decreased usage in Risky Behaviors compared to undamaged arms, may suggest the persistence of a potential trauma or pain-related memory. Post-traumatic protection and avoidance learning have already been observed in cephalopods and octopuses [43,44,45].
Specifically, it has been demonstrated that this association can result from visual cues, where octopuses avoided locations linked to noxious stimuli. Further, octopuses exhibited wound-directed grooming behaviors at injury sites, indicating the central nervous systems’ processing of both injury location and pain quality [44]. In the study subjects' arm injuries (Fig. 4C), with severity of injury following the pattern of R1>R2>R3>L1, avoidance of usage within risky behaviors and usage within safe behaviors aligned with this injury severity pattern, possibly suggesting a memory-driven usage response. This is further implied by the darkened coloration that arms R1-R3 assumed during various foraging outings; a display associated with precursing attacks on conspecifics, predators, and prey [46]. As the display is thought to draw attention to the darkened skin, within this context, it could be interpreted as either a warning or show of aggression to predators or alternatively as a lure to attract prey. Despite its interpretation, as chromatophores, which are responsible for this red coloration, are controlled by a set of lobes in the brain [47], the active darkening of the arms could imply the individual’s understanding, either visually or cognitively, of the previous site of injury. While the injury's exact cause is unknown, interactions with predators during foraging, particularly affecting those used commonly in foraging and feeding behaviors, often result in the loss of these looser arms [14], and thus may explain the decreased usage of the R1 arms in Foraging.
Figure 4.
Most common posture displayed by the octopus throughout all the videos; note the curling of the R1a and R1b arms, resembling a curled mustache. (A) was taken on Dec. 07, 2021, while (B) was taken on April 19, 2022. (C) was taken on December 07, 2021, wherein the regrowth of the tip of arm L1 and the regrowth of the smaller bifurcated arms and arms R2 and R3, respectively shown in purple and green boxes.
Figure 4.
Most common posture displayed by the octopus throughout all the videos; note the curling of the R1a and R1b arms, resembling a curled mustache. (A) was taken on Dec. 07, 2021, while (B) was taken on April 19, 2022. (C) was taken on December 07, 2021, wherein the regrowth of the tip of arm L1 and the regrowth of the smaller bifurcated arms and arms R2 and R3, respectively shown in purple and green boxes.

5. Conclusions
Efficient and sustainable management of a species can only occur if first there is a thorough understanding of the species itself. This is true for all species, with members of the class Cephalopoda (Cuvier, 1795) being no exception. Insights into ecology, life history, and behavior can provide data necessary to help inform mitigation and conservation tactics. As cephalopods are short-lived, exhibit rapid growth, and strong life-history plasticity (including the ability to alter their RNA to accommodate for temperature changes), they have a unique ability to quickly adapt to changes in the environment. An adaptability that has helped cause the proliferation of cephalopod populations despite many other marine species being in decline. Therefore, the study of this proliferation through ecology, life history, and behavioral flexibility, as we have done, might provide a unique opportunity to gain insights into conservation not only for cephalopods themselves but for all marine species. Further, as Octopuses provide a valuable system for studying regeneration and adaptive control of each of its arms, a deeper understanding of their underlying neurophysiological mechanisms can facilitate significant discoveries and bio-engendering strategies that can be applied to a wide range of taxa (including higher vertebrates) as well as other scientific disciplines, such as regenerative medicine and robotics.
While we have begun to explore bifurcated arm use in O. vulgaris, many questions remain. One avenue of research could investigate what happens to neurons in a lost octopus arm—whether they are redistributed throughout the body or simply lost. If neurons are lost, do they slowly regrow, and does the presence of an additional arm result in double the usual number of regrown neurons? Alternatively, if neurons are redistributed throughout the body, then distributed back into the arms as they grow, when an arm is bifurcated is it possible that the neurons are only distributed into one arm and not the other? Is so, this redistribution could potentially explain why bifurcation normally results in a non-functional limb.
Although further research is required to answer these questions, they suggest intriguing possibilities. Could an octopus with newly bifurcated arms be managing with less innervation and effectively learning to use these limbs anew? Or are these limbs fully innervated but simply inhibited by their length? Although these curiosities could prove to be very interesting as future lines of research, at present they will remain just that, curiosities.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: duration and occurrence of arm use for all events; Figure S2: arm occurrences for all events within behavioral events noted as Safe and Under Webbing actions; Figure S3: raw data plots of Safe Behavior usage over time for each arm; Figure S4: normal Retroflex behavior displayed by an Octopus vulgaris; Figure S5: alternative Webover; Table S1: behaviors and their different groupings; Table S2: ethogram of all behaviors, including ones not used in the final analysis; Video S1: example of a non-usable video; Video S2: example of a usable video; Video S3: alteration to Webover behavior; Video S4: large fish escapes through the gap created by the smaller bifurcated arms; Video S5: individuals first foraging outing captured on film; Video S6: later foraging outing wherein the bifurcated arms are used more in exploration and other Risky behaviors; Video S7: interaction with a hermit crab. .
Author Contributions
Conceptualization, Sam Soule, Miguel Cabanellas-Reboredo, Ángel González, Hidde Juijn and Jorge Hernández-Urcera; Data curation, Sam Soule; Formal analysis, Sam Soule; Funding acquisition, Ángel González and Jorge Hernández-Urcera; Investigation, Sam Soule, Miguel Cabanellas-Reboredo, Ángel González, Hidde Juijn and Jorge Hernández-Urcera; Methodology, Hidde Juijn; Project administration, Ángel González and Jorge Hernández-Urcera; Resources, Miguel Cabanellas-Reboredo and Jorge Hernández-Urcera; Software, Sam Soule; Supervision, Miguel Cabanellas-Reboredo and Jorge Hernández-Urcera; Validation, Miguel Cabanellas-Reboredo, Hidde Juijn and Jorge Hernández-Urcera; Visualization, Hidde Juijn; Writing – original draft, Sam Soule; Writing – review & editing, Sam Soule, Miguel Cabanellas-Reboredo, Ángel González, Hidde Juijn and Jorge Hernández-Urcera.
Funding
Ministry of Science, Innovation and Universities, Spain/ ECOSUMA Project (PID2019-110088RB-I00) and Erasmus+ Funding Programme.
Institutional Review Board Statement
This is an observational study. The IIM-CSIC Research Ethics Committee has confirmed that no ethical approval is required.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
Acknowledgments
We are grateful to Manuel E. Garci for his comments on earlier versions of this manuscript. We also thank Anna Thomasdotter for the revisions and artistic input.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Imperadore, P.; Fiorito, G. Cephalopod tissue regeneration: Consolidating over a century of knowledge. Front. Physiol. 2018, 9, 593. [Google Scholar] [CrossRef] [PubMed]
- Lange, M. On the regeneration and finer structure of the arms of the cephalopods. J. Exp. Zool. 2005, 31, 1–57. [Google Scholar] [CrossRef]
- Voight, J. Movement, injuries and growth of members of a natural population of the Pacific pygmy octopus, Octopus digueti. J. Zool. 2009, 228, 247–264. [Google Scholar] [CrossRef]
- Alejo-Plata, C.; Valencia-Méndez, O. Arm abnormality in Octopus hubbsorum (Mollusca: Cephalopoda: Octopodidae). Am. Malacol. Bull. 2014, 32, 1–3. [Google Scholar] [CrossRef]
- Lee, P.N.; Callaerts, P.; De Couet, H.G.; Martindale, M.Q. Cephalopod Hox genes and the origin of morphological novelties. Nature 2003, 424, 1061–1065. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.F. Developmental Biology, 10th ed.; Sinauer Associates, Inc.: Sunderland, MA, 2014. [Google Scholar]
- Lodish, H.; Berk, A.; Kaiser, C.A.; Krieger, M.; Scott, M.P.; Bretscher, A.; Ploegh, H.; Matsudaira, P. Molecular Cell Biology, 5th ed.; W.H. Freeman and Co.: New York, 2003. [Google Scholar]
- Tarazona, O.A.; Lopez, D.H.; Slota, L.A.; Cohn, M.J. Evolution of limb development in cephalopod mollusks. eLife 2019, 8, e43828. [Google Scholar] [CrossRef]
- Grimaldi, A.; Tettamanti, G.; Acquati, F.; Bossi, E.; Guidali, M.L.; Banfi, S.; Monti, L.; Valvassori, R.; de Eguileor, M. A hedgehog homolog is involved in muscle formation and organization of Sepia officinalis (Mollusca) mantle. Dev. Dyn. 2008, 237, 659–671. [Google Scholar] [CrossRef]
- González, Á.F.; Guerra, Á. First observation of a double tentacle bifurcation in cephalopods. Mar. Biodivers. Rec. 2008, 1, e44. [Google Scholar] [CrossRef]
- Nakajima, R.; Shigeno, S.; Zullo, L.; De Sio, F.; Schmidt, M.R. Cephalopods between science, art, and engineering: A contemporary synthesis. Front. Commun. /: https, 3389. [Google Scholar]
- O’Brien, C.E.; Roumbedakis, K.; Winkelmann, I.E. The current state of cephalopod science and perspectives on the most critical challenges ahead from three early-career researchers. Front. Physiol. /: https, 3389. [Google Scholar]
- Friard, O.; Gamba, M. BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Voss, K.M. Armed and dangerous: Patterns and drivers of octopus arm loss [Unpublished. Master’s Thesis, ]. University of California Santa Cruz, Santa Cruz, CA, 2022. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, 2011. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, B.; Simpson, G.; Solymos, P.; Stevens, H.; Wagner, H. Vegan: Community ecology package. R Package Version 2015, 2, 1–2. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Peres-Neto, P.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef] [PubMed]
- Ezekiel, M. Methods of Correlation Analysis; Wiley: New York, 1930. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Huffard, C.L. Ethogram of Abdopus aculeatus (d’Orbigny, 1834) (Cephalopoda: Octopodidae): Can behavioural characters inform octopodid taxonomy and systematics? J. Molluscan Stud. 2007, 73, 185–193. [Google Scholar] [CrossRef]
- Mather, J.A. How do octopuses use their arms? J. Comp. Psychol. 1998, 112, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Mather, J.A. Foraging, feeding and prey remains in middens of juvenile Octopus vulgaris (Mollusca: Cephalopoda). J. Zool. 1991, 224, 27–39. [Google Scholar] [CrossRef]
- Fiorito, G.; Gherardi, F. Prey-handling behaviour of Octopus vulgaris (Mollusca, Cephalopoda) on bivalve preys. Behav. Process. 1999, 46, 75–88. [Google Scholar] [CrossRef]
- Bidel, F.; Bennett, N.C.; Wardill, T.J. Octopus bimaculoides’ arm recruitment and use during visually evoked prey capture. Curr. Biol. 2022, 32, 4727–4733e3. [Google Scholar] [CrossRef]
- Villanueva, R.; Perricone, V.; Fiorito, G. Cephalopods as predators: A short journey among behavioral flexibilities, adaptations, and feeding habits. Front. Physiol. 2017, 8. [Google Scholar] [CrossRef]
- Flash, T.; Zullo, L. Biomechanics, motor control and dynamic models of the soft limbs of the octopus and other cephalopods. J. Exp. Biol. 2023, 226 (Suppl_1). [Google Scholar] [CrossRef]
- Sumbre, G.; Gutfreund, Y.; Fiorito, G.; Flash, T.; Hochner, B. Control of octopus arm extension by a peripheral motor program. Science 2001, 293, 1845–1848. [Google Scholar] [CrossRef] [PubMed]
- Sumbre, G.; Fiorito, G.; Flash, T.; Hochner, B. Octopuses use a human-like strategy to control precise point-to-point arm movements. Curr. Biol. 2006, 16, 767–772. [Google Scholar] [CrossRef]
- Byrne, R.; Kuba, M.; Meisel, D.; Griebel, U.; Mather, J. Does Octopus vulgaris have preferred arms? J. Comp. Psychol. 2006, 120, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, E.B.L.; Buresch, K.C.; Boinapally, P.; Hanlon, R.T. Octopus arms exhibit exceptional flexibility. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Levy, G.; Flash, T.; Hochner, B. Arm coordination in octopus crawling involves unique motor control strategies. Curr. Biol. 2015, 25, 1195–1200. [Google Scholar] [CrossRef]
- Scheel, D.; Godfrey-Smith, P.; Lawrence, M. Signal use by octopuses in agonistic interactions. Curr. Biol. 2016, 26, 377–382. [Google Scholar] [CrossRef]
- Voss, K.M.; Mehta, R.S. Asymmetry in the frequency and proportion of arm truncation in three sympatric California Octopus species. Zoology 2021, 147, 125940. [Google Scholar] [CrossRef]
- Huffard, C.L.; Caldwell, R.L.; Boneka, F. Mating behavior of Abdopus aculeatus (d’Orbigny, 1834) (Cephalopoda: Octopodidae) in the wild. Mar. Biol. 2008, 154, 353–362. [Google Scholar] [CrossRef]
- Weertman, W.L.; Scheel, D. Hold it close: Male octopus hold their hectocotylus closer to their body. Mar. Biol. 2024, 171. [Google Scholar] [CrossRef]
- Hernández-Urcera, J.; Garci, M.E.; Cabanellas-Reboredo, M. Bipedal locomotion by Octopus vulgaris. Mar. Biodivers. 2020, 50. [Google Scholar] [CrossRef]
- Huffard, C.L.; Boneka, F.; Full, R.J. Underwater bipedal locomotion by octopuses in disguise. Science 2005, 307, 1927–1927. [Google Scholar] [CrossRef] [PubMed]
- Gutnick, T.; Zullo, L.; Hochner, B.; Kuba, M.J. Use of peripheral sensory information for central nervous control of arm movement by Octopus vulgaris. Curr. Biol. 2020, 30, 4322–4327e3. [Google Scholar] [CrossRef] [PubMed]
- Hochner, B. How nervous systems evolve in relation to their embodiment: What we can learn from octopuses and other molluscs. Brain Behav. Evol. 2013, 82, 19–30. [Google Scholar] [CrossRef]
- Zullo, L.; Sumbre, G.; Agnisola, C.; Flash, T.; Hochner, B. Nonsomatotopic organization of the higher motor centers in Octopus. Curr. Biol. 2009, 19, 1632–1636. [Google Scholar] [CrossRef]
- Zullo, L.; Eichenstein, H.; Maiole, F.; Hochner, B. Motor control pathways in the nervous system of Octopus vulgaris arm. J. Comp. Physiol. A 2019, 205, 271–279. [Google Scholar] [CrossRef]
- Budelmann, B.; Young, J. Brain pathways of the brachial nerves of Sepia and Loligo. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1987, 315, 345–352. [Google Scholar] [CrossRef]
- Crook, R.J. Conditioned place preference reveals tonic pain in octopus. bioRxiv, 2634. [Google Scholar] [CrossRef]
- Crook, R.J. Behavioral and neurophysiological evidence suggests affective pain experience in octopus. iScience 2021, 24. [Google Scholar] [CrossRef]
- Côté, I.M.; Verde, A. Visual signals of the East Pacific red octopus (Octopus rubescens) during conspecific interactions. In Proc. 38th Am. Acad. Underwater Sci. Symp.; Vancouver, BC, Canada, 2019.
- Messenger, J.B. Cephalopod chromatophores: Neurobiology and natural history. Biol. Rev. Camb. Philos. Soc. 2001, 76, 473–528. [Google Scholar] [CrossRef]

Table 1.
Video analysis information. Video Number with* indicates that this video was removed from the analysis given its lack of usable video segments. Totals are indicated in bold.
Table 1.
Video analysis information. Video Number with* indicates that this video was removed from the analysis given its lack of usable video segments. Totals are indicated in bold.
| VIDEO NUMBER | RECORDING DATE (yy-mm-dd) | DAY CORRELATION (d) | VIDEO DURATION (hrs:min:sec) | PERCENT ANALYZED (%) |
|---|---|---|---|---|
| 1 | 21-12-13 | 1 | 0:16:18 | 96.9 |
| 2 | 22-01-12 | 31 | 0:19:51 | 90.9 |
| 3 | 22-01-26 | 45 | 0:5:42 | 91.1 |
| 4 | 22-01-28 | 47 | 0:32:47 | 98.0 |
| 5 | 22-01-30 | 49 | 0:3:48 | 97.4 |
| 6 | 22-02-03 | 53 | 0:23:44 | 76.0 |
| 7 | 22-02-05 | 55 | 0:28:24 | 95.7 |
| 8 | 22-02-06 | 56 | 0:44:36 | 98.8 |
| 9 | 22-02-07 | 57 | 0:32:39 | 95.1 |
| 10 | 22-02-20 | 70 | 0:28:25 | 89.1 |
| 11 | 22-02-21 | 71 | 0:33:27 | 76.6 |
| 12 | 22-02-22 | 72 | 0:41:54 | 79.9 |
| 13* | 22-02-23 | 73 | 0:9:42 | 6.4 |
| 14 | 22-02-27 | 77 | 0:4:26 | 88.1 |
| 15 | 22-03-03 | 81 | 0:4:31 | 86.4 |
| 16 | 22-03-05 | 83 | 0:40:34 | 93.8 |
| 17 | 22-03-08 | 86 | 0:42:43 | 96.5 |
| 18 | 22-03-10 | 88 | 0:42:06 | 38.1 |
| 19 | 22-03-25 | 103 | 0:25:41 | 96.2 |
| 20 | 22-03-28 | 106 | 0:30:22 | 77.5 |
| 21 | 22-04-10 | 119 | 0:44:46 | 98.8 |
| 22 | 22-04-16 | 125 | 0:41:51 | 93.4 |
| 23 | 22-04-19 | 128 | 0:39:36 | 91.4 |
| 24* | 22-05-08 | 147 | 0:2:57 | 0 |
| Total | - | 147 | 10.46:85 | - |
| Mean | - | 75.96 | 2:42:05 | 84.9 |
Of these events 38% were Postural Arm Components, 24.8% were Foraging behaviors, 20% were Exploration, 8% were Locomotion behaviors, 5.4% were Feeding, 0.8% were Arm Actions, 0.8% were Body Patterning, 0.3% were Alert behaviors, and 0.03% were Cleaning behaviors (see Table S1 for classification of Behavioral Categories). The behavioral categories mainly focused on in this study - Postural Arm Components, Foraging, Exploration, Locomotion, and Feeding - accounted for 96.1% of all the total events.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



