
Submitted:
24 February 2025
Posted:
25 February 2025
You are already at the latest version
Abstract
Natural populations provide valuable information and resources for addressing the ge-netic characterization of biological systems. Botrytis cinerea is a necrotrophic fungus that exhibits complex responses to light. Physiological analysis of B. cinerea populations from vineyards in Castilla y León (Spain) allowed the identification of isolate Bc116. This field isolate shows a reduced pathogenicity that is conditioned by the light regime. Light also delays germination and accentuates the negative effect it exerts on the vegetative growth of B. cinerea. Bc116 also displays a marked hyperconidiation phenotype and a characteristic sclerotia production pattern. Genetic analysis demonstrates that the alternative pheno-types regarding pathogenicity, conidiation and pattern of sclerotia production co-segregate in the progeny of crosses between isolate Bc116 and a wild type field isolate, Bc448, showing that they are under the control of a single locus. By applying a strategy based on bulked segregant analysis, the mutation in Bc116 has been mapped to a 200 kb region on Chr14 and the analysis of this region identifies a 2 kb deletion affecting the bcltf1 gene, encoding the B. cinerea Light responsive Transcription Factor 1 described in the reference isolate B05.10. Transformation of Bc116 with the B05.10 bcltf1 allele restored the wild type phenotypes, providing functional evidence that the natural mutant Bc116 is altered in gene bcltf1. This study offers additional information, derived from the analysis of the ge-netic background of a natural mutant, on the physiological processes regulated by BcLTF1 and supports the key role of this TF in the pathogenicity and photobiology of B. cinerea.
Keywords:
Botrytis cinerea
; pathogenicity
; light responses
; genetic diversity
; Bulked Segregant Analysis
1. Introduction
Botrytis cinerea, the causal agent of gray mold, is a highly relevant phytosanitary problem in many cultivated plant species. It is an ubiquitous phytopathogenic fungus with a very wide host range, including species of enormous economic importance [1]. It can infect all types of organs and tissues and causes problems both in the field and during post-harvest. Its life cycle is complex. As a saprophyte it can survive in the field on senescent or decomposing plant tissues. As a necrotrophic pathogen it infects living plant tissues, causing the death of the host cells to obtain from the decomposing tissues the nutrients it needs for its growth and multiplication [2,3]. The asexual spores produced on the conidiophores of the mycelium proliferating on the colonized tissues constitute the main structure for dispersion and infection. Mycelium itself is an important source of inoculum in the field. The fungus produces resistance structures, the sclerotia, which allow the pathogen to overcome adverse environmental conditions, such as low winter temperatures. When favorable conditions are restored, the sclerotia germinate, producing mycelium that actively sporulates. The sclerotia also play a fundamental role in the sexual cycle of the pathogen, acting as a reproductive structure provided by the female parent in crosses in which they are fertilized by microconidia produced by an isolate of compatible mating type that acts as a male parent. The ascospores produced in the derived fruiting bodies, the apothecia, also contribute to the dispersion of the pathogen and, like the spores derived from the asexual phase, are infective.
The fact that a species presents a sexual phase in its life cycle offers an extremely useful experimental tool in the context of the genetic analysis of traits: the possibility to perform crosses. Early studies on the mating type of B. cinerea indicated that it is a heterothallic species [4]. The sexual type is determined by a single locus (MAT1) with two idiomorphs, MAT1-1 and MAT1-2, initially assigned arbitrarily to two tester isolates, derived from single ascospores, SAS56 and SAS405, sexually compatible, generated in the course of the optimization of methods to perform crosses under laboratory conditions [5,6]. Both sexual types are found in similar proportions in natural populations. Although infrequently, it is also possible to find pseudo-homothallic isolates in nature that can be crossed with isolates of both sexual types [6,7,8]. Even though producing apothecia from crosses in the laboratory is a routinary practice nowadays, their presence in nature is very limited, if any [9]. This observation could be indicative that the sexual phase of the fungus is infrequent in the field. However, it is difficult to explain the high levels of genetic diversity observed in natural populations of B. cinerea if the contribution of a powerful mechanism for generating genetic diversity such as meiotic recombination, characteristic of sexual reproduction, is not considered. Different evidence, largely derived from population genetics studies [10,11,12], suggest that meiotic recombination and sexual reproduction occur very frequently in nature.
The existence of genetic variation constitutes the second fundamental pillar on which genetic analysis is based. B. cinerea is well known as a very plastic organism whose natural populations present very high levels of phenotypic diversity. This variation has been described in relation to diverse physiological aspects, such as vegetative growth, secondary metabolism, resistance to fungicides, virulence and responses to light. Numerous works have also demonstrated, by analyzing molecular variation, that B. cinerea populations present a high degree of genetic variability [13,14,15,16]. In recent years, the availability of well annotated reference genomes [17,18] and the resequencing of the genome of numerous field isolates have allowed us to confirm this high genetic variability across the genome [10,11,19]. The phenotypic diversity of the individuals that make up the populations is organized around the pool of genetic variability. Sometimes, genetic variation determines notable functional alterations, even loss of function, in genetic factors with a major effect on a certain character, or subtle alterations in the function of the different genetic factors involved in determining polygenic characters. In other cases, genetic variation is silent or affects intergenic regions and does not determine functional alterations. Whatever its nature, the existence of variation and the possibility of performing crosses allow the construction of genetic maps and the development of strategies based on association analysis in segregating offspring to identify genes with a major effect or Quantitative Trait Loci (QTLs) involved in determining traits. Bulked Segregant Analysis (BSA) is a QTL mapping method based on the identification of molecular markers associated with genes involved in the determination of traits of interest [20,21]. It is particularly appropriate in the analysis of offspring populations that show clearly contrasted alternative phenotypes characteristic of the parents involved in a certain cross. Its application has reported successful results in the genetic analysis of non-aggressive mycelial field isolates of B. cinerea [22]. On the other hand, the determination of variation in populations of individuals, and ideally at the genomic level, makes it possible to carry out Genome-Wide Association Studies (GWAS) that are not dependent on the offspring of crosses. When applied to the Botrytis-Arabidopsis pathosystem, indications of a complex architecture of virulence in the fungus have been found [23]. A similar trend is observed when investigating the interaction between B. cinerea and wild and domesticated tomato genotypes [19]. Candidate polymorphisms and genes associated with virulence have been reported in both systems.
Basic research on the biology of B. cinerea and its necrotrophic lifestyle has revealed a close relationship between virulence, development and light sensing. Early work already indicated that B. cinerea can sense light of different wavelengths and that this stimulus regulates differentiation programs. Genome analysis shows that B. cinerea possesses 11 photoreceptors whose activities cover the light spectrum (reviewed by Schumacher, 2017) [24]. In nature most strains respond to light like the B05.10 sequenced reference strain and undergo photomorphogenesis, that is, light exposure stimulates the production of macroconidia while its absence stimulates the production of sclerotia [25]. These strains are classified as light-responsive strains. But natural populations also show variation regarding this ability and “blind strains” displaying the same phenotype regardless of the light regime applied, “always conidia”, “always sclerotia” or “always mycelia”, are found. These strains are assumed to be deficient in key components of the light sensing machinery. Characterization of “always conidia” strains demonstrates that this phenotype may result from alterations in different genes. In the wild strains T4 and 1750 Single Nucleotide Polymorphisms (SNPs) in gene bcvel1 (member of the VELVET complex) determining stop codons, and therefore generating truncated versions of the encoded protein, were found to be responsible for the observed phenotype [26,27]. Deletion of any VELVET complex member determined inhibition of sclerotial development, increased conidiation, increased conidial melanogenesis and, remarkably, reduced virulence [26,28,29]. This demonstrates a role of VELVET in light sensing and development and pathogenicity in B. cinerea [28,29]. A similar phenotype has been described in mutants altered in the B. cinerea White Collar Complex (WCC), the key component involved in light perception and in the coordination of light response in B. cinerea. The functional WCC is the heterodimeric Transcription Factor (TF) integrated by the bcwcl1 and bcwcl2 gene products, two GATA-type transcription factors. The ∆bcwcl1 mutants generated in the B05.10 background show hyperconidiation and lack of sclerotial development. They also show precocious and persistent conidiation. Interestingly, they show reduced virulence, but only when incubated in photoperiod conditions, not in permanent darkness. Additionally, the mutants are hypersensitive to oxidative stress caused by H2O2 [30]. By applying a random mutagenesis approach another virulence-related factor, BcLTF1 (for B. cinerea Light Responsive Transcription Factor 1), was identified and described in the B05.10 strain [31]. The mutants, generated through A. tumefaciens-Mediated Transformation (ATMT), were first selected based on their reduced virulence. Detailed physiological and transcriptomic analysis uncovered the functions of BcLTF1 in the regulation of light-dependent differentiation, the equilibrium between production and scavenging of Reactive Oxygen Species (ROS) and secondary metabolism. Specifically, the mutants were unable to produce sclerotia, were found to hyperconidiate and be hypersensitive to light and oxidative stress. bcltf1, encodes a GATA-type TF homologous to the Neurospora crassa SUB-1 [32] and the Aspergillus nidulans NsdD [33]. In these model systems, the TF has demonstrated to participate in the modulation of light responses and differentiation.
Our group is interested in the characterization of the natural populations of B. cinerea from the vineyards of Castilla y León (Spain), in the evaluation of their genetic diversity and in the identification of natural mutant strains altered in pathogenicity [16]. The result of these evaluations was the identification of non-aggressive mycelial isolates whose characterization led to the identification of a gene with a major effect on development and pathogenicity in B. cinerea, Bcin04g03490, initially catalogued as a gene encoding a TF, since the encoded protein has a Gal4-type DNA binding domain [22]. During those evaluations, a strain with the “always conidia” phenotype was also isolated, which was deficient in its ability to infect Phaseolus vulgaris and Vitis vinifera leaves. Here we describe the physiological characterization of this strain, Bc116, and the application of a BSA strategy to map the mutation and identify the altered gene. This analysis demonstrates that strain Bc116 is a natural mutant altered in gene bcltf1.
2. Results
2.1. The pathogenicity of Bc116 is Light Dependent
Former studies indicated that, when the inoculated plant tissues are incubated under standard photoperiod conditions (16h light / 8h darkness, LD) isolate Bc116 is completely unable to infect Vitis leaves and shows reduced aggressiveness on bean leaves [22]. As light has been shown to affect pathogenicity in B. cinerea we decided to evaluate the effect of the light regime on the behaviour of isolate Bc116 in comparison with the aggressive field isolate Bc448 [22]. Pathogenicity assays were carried out on V. vinifera leaves variety Juan García by inoculating mycelium plugs from colonies actively growing on MEA plates. When the inoculated leaves entered the photoperiod at the beginning of the light phase (LD), isolate Bc116 was shown to be unable to cause infection (Figure 1A-LD), as shown previously. The same behaviour was observed when incubated under permanent light conditions (LL) (Figure 1A-LL). When the leaves were incubated under permanent darkness conditions (DD), isolate Bc116 was as infectious as the aggressive isolate Bc448 (Figure 1A-DD; Figure 1B). Interestingly, when the inoculated leaves entered the photoperiod at the beginning of the dark phase (DL), isolate Bc116 was also able to cause infection (Figure 1A-DL). These results indicate that light blocks the ability of isolate Bc116 to infect the Vitis leaves, but that it is specifically the effect of light during the early stages of the plant-pathogen interaction what is conditioning the ability of the fungus to cause infection.
The effect of light was also evaluated on bean leaves. In this case, the inoculations with isolates Bc116 and Bc448 were performed on whole plants. At 24 hours post-inoculation (hpi) (Figure 2A) isolate Bc116 did not produce any necrotic lesion on the leaves when the plants were incubated under LL regime. When incubating the inoculated plants under LD conditions, a very limited infection capacity of isolate Bc116 was observed in comparison with the aggressive Bc448 isolate. At this timepoint, the necrotic lesions produced by Bc116 were slightly smaller than those caused by Bc448 under the DL and DD conditions. Microscopic observation of stained tissues correlated with these observations (Figure 2B). Monitoring the progress of infection along time showed that Bc116 can establish an interaction with the host tissues, but that infection is severely delayed in comparison with isolate Bc448: at 72 hpi, expanding necrotic lesions were generated by Bc116 in all light regimes, but the diameter of the lesions was smaller under LL and LD conditions than under DL and DD conditions (Figure 2C,D). Bc448 generated larger necrotic lesions than Bc116 under the four light regimes (Figure 2C,D). When incubation was extended, both isolates produced full maceration of plant tissues (data not shown).
2.2. Light limits Bc116 Ability to Penetrate Onion Epidermal Cells
As Bc116 shows a delay in infection in the presence of the light stimulus we decided to evaluate possible alterations in processes involved in early stages of infection. Specifically, we decided to evaluate its ability to penetrate onion epidermis cells in inoculations performed either with a spore suspension or with mycelium plugs. At 12 hpi, spores of the aggressive Bc448 isolate were found to have germinated and penetrated the cells, and they did essentially to the same extend both under LL or DD conditions (Figure 3E,G). The Bc116 spores germinated and penetrated the cells under DD conditions (Figure 3C), but under LL conditions, although germination of the spores was observed, penetration was hardly detected. Instead, non-germinated spores and short and long germ tubes were observed on the cells’ surface (Figure 3A). When inoculations were performed with mycelium plugs, Bc448 produced branching mycelium inside the epidermal cells at 24 hpi, both under LL and DD conditions (Figure 3F,H). However, Bc116 showed penetrating or expanding mycelium inside the cells only under DD conditions (Figure 3D). In longer incubations (48 and 72 hpi) both with conidia and mycelium, Bc116 showed hyphae inside the host cells in LL (data not shown). Taken together, these observations indicate that light is conditioning the ability of isolate Bc116 to penetrate the host cells and determines a delay in this process.
2.3. Light Affects Germination, Vegetative Growth Rate and Conidiation in Bc116
The infection process of B. cinerea requires the germination of conidia and subsequent penetration in the plant tissues. If Bc116 shows a limited ability to infect when the initial stages of the infection process occur under light exposure, could the germination program of Bc116 be affected by light? To evaluate it, a germination assay in liquid synthetic medium was performed. Drops of conidial suspensions were placed in the center of empty Petri dishes and samples were incubated at 22°C under LL or DD conditions. Estimations of germination were carried out after 6 h, when most of the spores have germinated and lengths of the germ tubes can easily be scored. As shown in Figure 4A, the progress of germination of the Bc448 isolate was very similar under both light conditions, since the percentages of spores in each of the three considered stages was very similar under LL and DD. In Bc116 the light exposure determined a delay in the germination process, as the percentage of conidia in stage 2 was significantly higher when the fungus was incubated under DD than under LL. The situation was the opposite for stages 0 and 1.
Bc116 has been reported to be a hyperconidiating isolate, but its capacity to sporulate has not been quantified and it has not been determined if light influences sporulation. To investigate it, we determined the production of spores by isolate Bc116 in MEA plates in comparison with the field isolate Bc448 and the reference isolate B05.10 under the three light regimes, LL, LD and DD. In B05.10 light was shown to stimulate the production of spores, resulting in higher sporulation under continuous light. In comparison, Bc116 was found, indeed, to produce very large amounts of spores, between one and two orders of magnitude higher than those produced by isolates Bc448 and B05.10, and it hyperconidiates in the three light regimes, although more efficiently under DD conditions (Figure 4B). This implies that in Bc116 the light stimulation of sporulation is not manifested. Remarkably, the Bc448 field isolate resembles the reference strain B05.10 regarding the number of spores produced, but the stimulating effect of light is not manifested either. In this regard the two field isolates Bc116 and Bc448 display a similar response.
The effect of light in saprophytic growth rate was also investigated. To this end, fungal cultures on MEA plates were initiated with a drop of a spore suspension. After 5 days of incubation at 22°C under the three light regimes, the colony diameter was estimated. Growth rate was negatively affected by light in B05.10 and Bc448, as shown by the reduction of the colony diameter (reduction of 4.46% and 4.83% for isolate Bc448 under LL and LD, respectively; reduction of 4.41% and 5.48% for B05.10 under LL and LD, respectively) (Figure 4C). Bc116 grew more slowly than the B05.10 and Bc448 isolates in the three light regimes, but light further slowed down the growth of Bc116, indicating that it negatively affects the growth of this strain. The effect of light in Bc116 under LL or LD was quantitatively similar, as the reduction of growth was 12.15% and 13.12%, respectively. Although the saprophytic growth rate was reduced in all light regimes, Bc116 showed earlier sporulation both in cultures initiated either with spores (not shown) or mycelium plugs (see Figure 5), already evident in four days old cultures.
The production of sclerotia in most B. cinerea isolates is stimulated in the absence of light and favoured by low temperatures. In these conditions Bc448 and B05.10 isolates produced large sclerotia irregularly distributed on the plates. Bc116 also produced sclerotia, but following a different pattern characterized by the appearance of large numbers of very tiny sclerotia, most deeply embedded in medium and distributed all over the plate (Figure 4D).
2.4. Bc116 is Hypersensitive to Oxidative Stress under LL Conditions
As B. cinerea isolates altered in light responses are often reported to be also altered in their capacity to cope with oxidative stress [30,31], we evaluated the sensitivity to the oxidative stress conditions derived from the exposition to H2O2 under the three light regimes considered. To this end, cultures were initiated by transferring mycelium plugs to the center of MEA plates containing 7.5 mM H2O2 and then incubated at 22°C under LL, LD and DD conditions. Sensitivity to H2O2 was quantified by measuring the colony diameter 96 hpi. The three strains displayed reduced growth in the presence of H2O2 under the three light regimes. The behaviour of B05.10 and Bc448 was very similar and the magnitude of the reduction they manifested under the three light regimes was nearly the same (31.4%, 31.3% and 34.6% for B05.10 under LL, LD and DD, respectively; and 29.9%, 29.2% and 25.7% for Bc448 under LL, LD and DD, respectively). Bc116 was found to be more sensitive to oxidative stress and particularly under LL conditions (growth reduction of 69.7%, 63.1% and 40.5% under LL, LD and DD, respectively) (Figure 5).
2.5. Genetic Analysis of Bc116
Bc116 displays alterations in several aspects related to pathogenicity, development and differentiation and responses to the light stimulus. To characterize the genetic basis of these alterations we undertook a genetic analysis. The Bc116 isolate has been found to carry the MAT 1-2 allele and the Bc448 isolate, a highly aggressive field isolate belonging to the same natural population (vineyards of Castilla y León, Spain) [16] which is being used in our studies as a reference isolate and resembles in most aspects the reference isolate B05.10, has been shown to carry the MAT 1-1 allele [22]. Therefore, Bc116 and Bc448 show contrasting phenotypes and should be sexually compatible. Crosses between the two isolates were attempted. As Bc116 produces tiny sclerotia, crosses could only be established using Bc116 as the spermatizing strain, as it produces microconidia. An offspring was collected from the cross ♂Bc116 x ♀Bc448 consisting of 222 single ascospore isolates. First, the traits “aggressiveness” and “pattern of conidiation” were scored in the full set of descendants. 122 individuals were shown to hyperconidiate and be unable to infect Vitis leaves, while 100 displayed normal sporulation pattern and caused infection. When incubated at low temperatures in DD, the non-aggressive and hyperconidiating isolates were found to produce sclerotia like the Bc116 isolate, while the aggressive isolates which sporulate normally all produced sclerotia like the Bc448 isolates (Figure S1 shows the phenotypes of a representative selection of the progeny). These observations indicate that the three traits cosegregate, and the proportions observed informed of a 1:1 segregation. Therefore, it can be concluded that the three traits considered, “aggressiveness”, “pattern of conidiation” and “pattern of sclerotia production”, are under the control of a single genetic locus which has been altered in the natural isolate Bc116.
2.6. Mapping the Altered Gene in Bc116 by BSA
A BSA strategy was applied to the segregating population from the cross ♂Bc116 x ♀Bc448 in order to identify molecular polymorphisms cosegregating with the alternative phenotypes of the traits of interest. We have previously obtained information about the sequence polymorphisms of isolate Bc448 in the form of SNPs in comparison with the genome sequence of the reference isolate B05.10 [22]. We sequenced the Bc116 genome and high-quality short reads were aligned to the B05.10 genome reference sequence to identify the SNPs in Bc116. Analysis of the list of the polymorphisms of each isolate identified 83,118 SNPs exclusive of isolate Bc448 and 114,893 SNPs exclusive of isolate Bc116. Table 1 shows the chromosomes described in the B05.10 genome and their sizes as well as the total number of SNPs identified in each chromosome in the genome of Bc448 and Bc116 and the number of SNPs exclusive of one or the other isolate. The minichromosome Chr18 described in B05.10 [18] is not present either in the Bc448 or in the Bc116 genomes.
Two groups of descendants, A and B, from the cross ♂Bc116 x ♀Bc448, were established, each one integrated by 60 individuals: the first one included individuals resembling the aggressive isolate Bc448 and the second one individuals resembling the non-aggressive isolate Bc116. Then, the distribution of the frequencies of the SNPs specific of each parental isolate was analysed in the two groups of individuals. Except in Chr14, the plots of the SNP index display values close to “0” along the entire chromosome for most chromosomes, as shown for Chr13 (Figure 6A, plots for other chromosomes not shown). In Chr14 the two SNP plots peaked around coordinate 1,500,000, reaching maximal values (close to +1) for the one corresponding to the Bc116 isolate specific SNPs and minimal values (close to -1) for the one corresponding to the Bc448 isolate specific SNPs. Both plots delimitated a genomic region of markers associated with the segregating phenotypes of about 200 kb (between coordinates 1,400,000 and 1,600,000) (Figure 6B). This region includes 41 annotated genes in the genome of the reference strain B05.10 (Table S1).
2.7. Bc116 is Altered in the Light Responsive Transcription Factor BcLTF1
The mapped region was analysed in detail searching for polymorphisms with an expected large impact that could determine a loss of function and explain the observed phenotypes. As a second criterium, genes with a predicted regulatory role were considered as best candidates, as the mutation in Bc116 is determining alterations in diverse processes. A list of the genes annotated in the mapped region, together with the polymorphisms identified, is presented in Table S1.
Among the candidates it was detected the bcltf1 gene (Bcin14g03940), described previously as a gene encoding a TF regulating virulence and light responses in B. cinerea [31]. In the Bc116 genome a large deletion of about 2 kb in size was identified (Figure 6C) between positions 1,530,248 and 1,532,273. This deletion involves the 5’-UTR of gene bcltf1 and the first 211 nucleotides of its structural region, including the first exon, consisting of 32 nucleotides, the first intron, consisting of 91 nucleotides, and the first 88 nucleotides of the second exon. Because of this mutation, the encoded protein is expected to lack the first 54 aminoacids. The presence of the deletion and its extension was confirmed by amplifying the deleted region with primers flanking it in the reference genome (Figure 7A) and sequencing it (data not shown).
Given the functions reported for bcltf1 and the nature of the mutation identified, it was selected as the first candidate to be considered as the gene that has been altered in Bc116. To determine if the mutation identified in bcltf1 is responsible for the phenotype observed in isolate Bc116, a strategy based on functional complementation was undertaken. To this end, a 4.7 kb genomic DNA fragment containing the entire wild type allele of bcltf1, including the sequence deleted in the genome of Bc116, was amplified by PCR with primers bcltf1-c2F and bcltf1-c2R (annealing positions are indicated in Figure 6C) using as the template genomic DNA from the reference isolate B05.10. The fragment was cloned into plasmid pWAM6, giving rise to plasmid pVPM1, and from it a 7.7 kb linear fragment containing the bcltf1 wild type allele and the hygromycin resistance cassette was amplified with primers bltf1-c2R and PoliC- F5’. The fragment was transformed into Bc116 protoplasts. Two independent transformants, T2 and T7, were selected in which the bcltf1 wild type allele, together with the mutant allele, could be detected (Figure 7A). These two transformants were found to be able to infect Vitis leaves under LD conditions, behaving like the Bc448 and B05.10 isolates (Figure 7B), displayed growth rates similar to those of the wild type strains, did not show premature sporulation (Figure 7C) and did not hyperconidiate. Both transformants produced sclerotia following a pattern resembling more that of the B05.10 and Bc448 isolates than the one characteristic of Bc116 (Figure 7D). All these observations show the restoration of the wild type phenotype in T2 and T7 transformants, thus demonstrating the functional complementation.
3. Discussion
Natural variation on populations of fungal pathogens is of key importance to explain their biology. Whether induced or natural, genetic analysis of variation makes it possible to address the genetic dissection of traits. B. cinerea is considered one the most important fungal pathogens [34] and has attracted deep interest in the scientific community. In recent decades it has become the model organism to study the necrotrophic lifestyle. Experimental evidence is accumulating outlining the mode of action of a pathogen that establishes interaction with the host by manipulating and exploiting essential biological processes of the plant for its own benefit [35]. Recent research shows that B. cinerea has a remarkable capacity to sense light stimuli that the fungus integrates to take developmental decisions, and several authors consider B. cinerea as a valuable model system to expand our knowledge of fungal photobiology [24,30]. Our work, focused on the characterization of natural variation in B. cinerea populations, supports the existence of close relationships at the level of their genetic determination and regulation between the processes involved in pathogenicity and responses to light, since the identified natural mutants altered in their ability to infect the host plant, Bc116 among them, but not only [22], also show alterations in their responses to light.
Genetic analysis demonstrates that, although Bc116 shows alterations in different aspects related to pathogenicity, development and responses to light, they all depend on a single gene. This situation facilitates the consideration of procedures such as BSA to map the mutation and identify the altered gene. Although originally developed to map and identify QTLs [21], BSA is particularly well suited for the identification of genes with a major effect on the phenotype(s) of interest. In B. cinerea, our group has previously successfully applied this methodology [22]. In the work here presented, Bc116 has been shown to harbour levels of polymorphism like those reported for the Bc448 isolate [22] and for other field isolates [10,19]. By adapting the experimental framework previously considered, it has been possible to perform an association mapping covering the entire genome of our B. cinerea isolates offering a resolution allowing to map the mutation in isolate Bc116 to a 200 kb genomic region in Chr14. The availability of a fully sequenced and accurately annotated reference genome has become an unvaluable tool. The detailed analysis of the mapped region under the consideration of the two criteria highlighted, a mutation of expected high impact and affecting a genetic factor with possible regulatory function, led to the selection of bcltf1 as the most likely candidate. The restoration of the wild type phenotype by transformation with the allele derived from strain B05.10 demonstrates that bcltf1 is the altered gene in Bc116.
Former studies performed by Schumacher et al. [31] identified and described BcLTF1 as a virulence factor in B. cinerea. By means of ATMT, they generated three different mutants in B05.10 that displayed reduced aggressiveness on bean plants. The three mutants were found to harbour a T-DNA insertion in the upstream region of bcltf1. B05.10-Δbcltf1 mutants showed the same phenotype as those T-DNA mutants. These mutants were also altered in several differentiation and development processes regulated by the light stimulus. The results on the characterization of the natural mutant isolate Bc116 supports the regulatory role for BcLTF1 in virulence and light responses. In addition, it broadens the knowledge about the role of this factor in the biology of B. cinerea.
Our analysis indicates that the isolate Bc116, like the B05.10-Δbcltf1 mutants, shows a reduction in growth in all light regimes, and that this reduction is greater under permanent light conditions. It also shares with them the early sporulation and hyperconidiation phenotypes, but its capacity to produce spores is even greater. It is interesting to note that Bc116 does not show the stimulation of sporulation by light described as characteristic of B. cinerea isolates [24]. Remarkably, a similar behaviour is observed in the field isolate Bc448, supporting that, in relation to these responses, variation in natural populations is certain.
In this comparison it is more striking the ability of Bc116 to produce sclerotia. It is true that the pattern of sclerotia production is different from that shown by strain Bc448, which is very similar to that of B05.10, but under the experimental conditions used in this work to stimulate sclerotia production, DD and low temperatures for about one month, Bc116 indeed produced sclerotia. These conditions are similar to those reported for the characterization of the B05.10-∆bcltf1 mutants [31]. Therefore, it is possible to assume that the differences observed between the B05.10-∆bcltf1 mutants and Bc116 are due to their different genetic backgrounds. The fact that the pattern of sclerotia production shows segregation in the offspring of the Bc116 x Bc448 cross and that the complemented transformants show a pattern of sclerotia production resembling that of the Bc448 isolate indicates that the pattern of sclerotia formation is under the control of BcLTF1 and that segregation of other genetic factors in both parental genetic backgrounds likely determine differences in individuals. On the other hand, although the size of the sclerotia of Bc116 limits the possibility of their utilization in crosses, this isolate was successfully used as the male parental strain in our crosses, demonstrating that it is sexually competent.
The behaviour of Bc116 in inoculations on bean leaves in LD is similar to that described for the B05.10-∆bcltf1 mutants under the same conditions. A delay is observed in the progress of infection in comparison with the aggressive isolate Bc448, which in our analysis is verified macroscopically and microscopically at 24 hpi. It is a delay, because the formation of dispersive lesions is evident at 72 hpi and the plant tissues are fully colonized at later stages. These observations corroborate the descriptions made in the B05.10-∆bcltf1 mutants. However, it is interesting to note that, when extending the analysis to other light regimes, it is found that the factor limiting the mutant’s capacity to cause infection is the light exposure during the early phases of the plant-fungus interaction. In the B05.10-∆bcltf1 mutants, the ability to penetrate the onion epidermis was analysed in inoculations carried out both, with spores and with non-sporulating mycelium. No differences were observed in comparison with the wild type isolate and it was concluded that the delay in infection was due to limitations in colonization, not in penetration. However, these evaluations were carried out only in DD. Our analysis detects the same situation in DD, but it is verified that in LL, light reduces the efficiency of penetration into the onion epidermis of both, spores and mycelium. This indicates that the Bc116 isolate has limitations to cope with the effect of light during the early stages of development, which in planta involves penetration. Therefore, it may be concluded that the delay observed in the infection process of the bcltf1 mutants in LL is due, at least in part, to defects in penetration. The work by Schumacher et al. [31], found that their B05.10-∆bcltf1 mutants are hypersensitive to oxidative stress. The authors cleverly demonstrated that the delay in the infection process and the reduced growth observed in planta is due to the limited capacity of the bcltf1 mutants to cope with the oxidative stress that arises during light exposure. Bc116 is also hypersensitive to the oxidative stress produced by exposure to H2O2 during saprophytic growth, a sensitivity accentuated in LL and LD, results supporting their observations.
The impairment of the infecting capacity of Bc116 is most noticeable on Vitis leaves in LL, where a complete incapacity to establish the interaction is observed. As it is the same isolate on two different hosts, the more extreme limitation observed on Vitis should be attributed to differential properties of the host, either intrinsic or related to the defence mechanisms activated in response to the presence of the pathogen, which differentially affect the B. cinerea wild type isolate and the bcltf1 mutant. Since on bean leaves the sensitivity to oxidative stress accounts for the reduction in aggressiveness, it is possible to assume also on Vitis leaves an important role for the ROS produced by the plant tissues in response to the presence of the pathogen, which constitute one of the earliest cellular responses upon pathogen recognition [36]. The timing or the intensity of this production could be different on Vitis leaves. It will be of interest determining if other mechanisms or metabolites, specifically activated or produced on Vitis leaves, sum their effect to that of the oxidative stress conditions created during the early stages of the interaction to block the ability of the natural bclft1 mutant to cause infection.
It is striking that on Vitis leaves the effects of light exposure during the early stages of the interaction determine alterations in the fungus that are maintained over time once the inoculated tissues are transferred to dark conditions (the dark phase in LD). This implies that during the first moments of the attempted infection, the conditions created by the presence of light in the environment in which the plant tissues interact with the infective structures of the fungus determine alterations in the development program of the pathogen that are irreversible. It is interesting to note that the ROS play crucial roles in development and differentiation processes in fungi [37]. The interaction described in this work may offer an interesting experimental system to identify specific targets of the ROS that are key in the definition of these differentiation programs in B. cinerea.
In N. crassa, the most deeply investigated model system in fungal photobiology, SUB-1 functions as an early light-responsive TF which is involved in regulating some early and most late light-responses [32]. In the plant pathogen B. cinerea the homologous BcLTF1 responds to light and regulates light dependent processes [31] and, in addition, it regulates pathogenicity related functions. Our work has characterized a natural mutant, Bc116, altered in bcltf1. Its physiological and genetic analysis broadens our knowledge of the functions regulated by this TF and supports its fundamental role in the regulation of light responses, differentiation and pathogenicity in B. cinerea.
4. Materials and Methods
4.1. Organisms and Growth Conditions
The B. cinerea isolates B05.10 [38], Bc116 and Bc448 [16] were used in this study. Fungal cultures were established from frozen conidia stored on 15% glycerol (v/v) at -80°C. Fungal isolates were grown at 22°C under the light/darkness conditions required for each experiment. Light was generated by Cool White Osram L 36W/840 fluorescent bulbs.
Common bean plants (Phaseolus vulgaris L.) cv Blanca Riñón were kindly provided by Centro de la Legumbre (Pajares de la Laguna, Salamanca, Spain). Plants were grown in natural substrate for 2 weeks in the greenhouse under a 16/8 h light/darkness photoperiod. Vitis vinifera plants variety Juan García were maintained in the greenhouse in the same conditions.
4.2. Germination, Conidiation and Saprophytic Growth Experiments
To study the fungal germination patterns the strains were grown on MEA (Malt Extract Agar, Difco) plates for 3 weeks at 22°C and permanent darkness. Conidia were harvested and suspensions of 5x105 conidia/mL were prepared in PDB (Potato Dextrose Broth, Difco) at half concentration. 60 µL drops of the conidial suspensions were placed in the center of empty Petri dishes that were incubated without agitation inside a wet chamber at 22°C under continuous light or continuous darkness. Samples were imaged at 6 hpi using a MD-E3-6.3 camera (MicrosCopiaDigital, Industrial Digital Camera) adapted to a microscope Leica DLMB (Leica Microsystems, Bensheim). The percentage of germinated conidia was quantified according to the previously described classification of the conidia developmental stages [39]. Five plates per condition and strain were analyzed in each experiment and experiments were repeated three times.
To analyse conidiation rates, the fungal isolates were grown on MEA for 4 days at 22°C and permanent darkness. Agar plugs of 5 mm in diameter from the edge of the colony were taken and placed in the center of Petri dishes containing MEA. The plates were maintained at 22°C and different light conditions (LL, 16/8 h LD, DD) for 3 weeks and then the conidia were harvested from each plate. The number of conidia produced was estimated using a Thoma cell counting chamber. Three plates per strain and experiment were analysed and three independent biological experiments were carried out.
The fungal saprophytic growth was determined on MEA. Fungal isolates were grown, and conidia suspension were prepared as described above for the germination assays; 10 µL drops of the conidial suspension were placed in the center of Petri dishes containing MEA. The plates were incubated at 22°C for 5 days and then the diameter of the colony was measured. Different light conditions were assayed, LL, LD and DD. Three plates per strain and light condition were inoculated in each biological experiment and three independent experiments were performed. For the evaluation of the effect of the oxidative stress the isolates were grown on MEA supplemented with 7.5 mM of H2O2 using agar mycelium plugs of 5 mm in diameter as initial inoculum and measuring the colony diameter 96 hpi.
The capacity of fungal isolates to produce sclerotia was evaluated on MEA plates. Mycelium plugs taken from the edge of actively growing colonies were placed in the center of Petri dishes containing the media and incubated at 22°C for 4-5 days under permanent darkness. Afterwards, plates were incubated at 2-4°C for 4 weeks and then the number, size and distribution of sclerotia were analysed.
4.3. Penetration Analysis
The ability of fungal isolates to penetrate host tissues was analyzed on onion epidermal cells. Strips of onion epidermis were cut and placed on a slide with the hydrophobic layer side-up. 10 µL drops of a conidial suspension at 5x104 conidia/mL prepared in water, or non-sporulating mycelium plugs of 3 mm in diameter taken from the edge of actively growing colonies, were placed on the strips. The samples were maintained at 22°C and the light conditions required inside closed plastic boxes to ensure a high humidity environment. At the time of analysis, the mycelium plugs were removed. In both inoculations, 10 µL of lactophenol blue solution (Fluka, SIGMA) were placed on each inoculation spot and incubated for 15 min at room temperature. Afterwards, the staining solution was removed and the samples were washed with distilled water. Samples were imaged using a MD-E3-6.3 camera (MicrosCopiaDigital, Industrial Digital Camera) adapted to a microscope Leica DLMB (Leica Microsystems, Bensheim).
4.4. Inoculation Assays
The pathogenicity of fungal isolates was studied according to previously described methods [16,22]. The inoculation on common beans was performed with whole plants that were placed inside transparent plastic boxes with water on the bottom; the inoculation on Vitis was carried out on detached leaves whose petioles were inserted in wet floral foam and then placed inside plastic trays with wet paper on the bottom. In both systems 5 mm in diameter plugs of fresh mycelium taken from the edge of fungal colonies actively growing on MEA plates were placed on non-wounded leaves. Four plugs and one isolate were used per leaf in the case of common bean inoculations, and four plugs and two isolates were used per leaf in the case of Vitis leaves. At least 5 leaves were inoculated per fungal isolate and condition in each experiment. The experiments were repeated in a randomized design at least three times. The inoculated materials were maintained in closed boxes or trays at 22°C and the light conditions required in each case. Aggressiveness was evaluated by measuring the diameter of the lesions 3 and 4 dpi for common bean plants and Vitis leaves, respectively.
For the staining of inoculated common bean leaves, the mycelium plugs were removed 24 hpi and the area of the inoculation cut. Samples were immersed into a solution of lactophenol blue:ethanol (1:2) and incubated for 1 min at 100°C, cooled at room temperature and incubated again for 30 sec at the same temperature. After cooling at room temperature, the staining solution was removed and the samples were washed with absolute ethanol. Images were acquired using a Leica DFC495 camera adapted to a Leica 205FA stereomicroscope (Leica Microsystems, Bensheim, Germany) and analyzed using the LAS software v3.6.0 (Leica Microsystems, Bensheim, Germany).
4.5. Standard Molecular Techniques
Fungal genomic DNA was isolated from mycelium cultured on cellophane sheets placed onto MEA plates. All the DNA purifications followed previously described procedures [40].
The PCR reactions were carried out using the DNA Polymerase from Biotools with the exception of the amplification of the bcltf1 allele which was performed using the Phusion High Fidelity DNA polymerase from ThermoFisher Scientific. In both cases the reactions were performed according to manufacturer’s recommendations. All the primers used in this work are listed in Table S2.
The Gateway BP Clonase II Enzyme Mix (ThermoFisher Scientific) was used in the cloning reactions of the bcltf1 allele in plasmid pWAM6 [22].
4.6. B. cinerea Transformation
B. cinerea protoplasts were transformed using the method described by ten Have et al. [41], with modifications as specified by Reis et al. [42] and Leisen et al. [43]. Protoplasts were generated using a 1% concentration of Vinotaste Pro (Lamothe Abiet, Canejan, France), an enzymatic blend of chitinases and glucanases, along with 0.1% Yatalase (Takara, Saint-Germain-en-Laye, France).
4.7. Construction of Bc116-bcltf1 Complemented Transformants
The wild type allele of bcltf1 from the B05.10 strain was cloned into plasmid pWAM6 [22], which contains a hygromycin (Hph) resistance cassette from pOHT [44], using the Gateway cloning technology. To this end, the B05.10-bcltf1 allele was amplified as a 4.7 kb fragment with specific oligonucleotides harboring extensions with attB sequences (Table S2). Upon recombination facilitated by the BP clonase, plasmid pVPM1 was generated. From it, a 7.7 kb linear fragment containing the B05.10-bcltf1 allele and the Hph cassette was amplified using specific oligonucleotides (Table S2). The PCR product was used to transform Bc116, and transformants able to grow on selective media were successively transferred to fresh selective plates. Monosporic cultures were obtained from the selected transformants.
4.8. Crosses
Crosses between Bc448 and Bc116 were performed following the procedures established by Faretra et al. [5,6]. Mature apothecia were collected and crushed in water to release the ascospores. The suspension was filtered through glasswool and plated on MEA plates. Individual ascospore germlings were transferred 24 h later to fresh MEA plates for propagation.
4.9. Sequencing and Determination of Polymorphisms
For this study, four data sets were used. The genome of Bc448 was previously sequenced [22] (Accession number SRR13700579). Genomic DNA of the parental strain Bc116 and of both descendant groups—those resembling Bc448 and those resembling Bc116—in the cross Bc116 x Bc448 were sequenced using Illumina technology by Novogene (Cambridge, UK) (Accession numbers: Bc116 Genome – SAMN46863733; Pool aggressive progeny (A) – SAMN46863735; and Pool non-aggressive progeny (B) – SAMN46863734). The sequences were mapped to the genome of the reference isolate, B05.10 (ASM14353v4), and polymorphisms were subsequently extracted using the tools available in Geneious Prime® 2023.1.1 (Biomatters, Auckland, New Zealand).
4.10. BSA
For the BSA analysis a list of SNPs of each parental isolate in comparison with the B05.10 reference genome was generated with Geneious Prime® 2023.1.1 (Biomatters, Auckland, New Zealand). From these, two lists of SNPs exclusive of either Bc448 or Bc116, were derived. From the progeny, two groups of individuals were selected: Group A, consisting of 60 individuals resembling the parental isolate Bc448, and Group B, consisting of 60 individuals resembling the parental isolate Bc116. Genomic DNA was extracted from each individual isolate, and equal amounts of DNA from the 60 individuals in each group were pooled together to form two bulk DNA samples. These genomic DNA pools were sequenced by Illumina, and the frequencies of the polymorphisms specific of either Bc448 or Bc116 were determined in each pool. For the association mapping analysis, only high-quality SNPs (quality score >33) were considered. The distribution of polymorphisms specific to each parental isolate in the two progeny groups was analyzed by calculating the difference in the frequency of each polymorphism between DNA pool B and DNA pool A (f “polymorphism x” in B - f “polymorphism x” in A). This difference generates a SNP index (Y-axis) that was plotted against the chromosomal coordinates (X-axis) of the reference genome B05.10. For markers unlinked to the locus responsible for the phenotypic difference, allelic frequencies were expected to be similar in both pools, causing the SNP index plot for these chromosomal regions to fluctuate around the “0” value. Conversely, for markers linked to the locus of interest, allelic frequencies differed significantly between the DNA pools, with larger differences observed for markers more closely linked to the locus. The SNP index reached maximum values (close to +1) for Bc116-specific alleles predominantly found in the non-aggressive, hyperconidiating progeny DNA pool and minimum values (close to -1) for Bc448-specific alleles predominantly found in the aggressive progeny DNA pool.
4.11. Statistical Analysis
All the statistical analysis were performed with the help of the software Statistix 10 (Analytical Software, Tallahassee, Florida, USA).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Analysis of the phenotype of representative individuals of the progeny derived from the cross Bc116 x Bc448; Table S1: Analysis of the region in Chr14 mapped by BSA; Table S2: Oligonucleotides used in this work.
Author Contributions
Conceptualization, E.P.B. and J.M.D.-M.; experiments: V.C.-d.C., V.P.M.N, A.G.G., P.I.O. and E.P.B.; technical assistance, J.M.G.F.; writing—original draft preparation, V.C.-d.C.; V.P.M.N, J.M.D.-M and E.P.B.; writing of the article: V.C.-d.C., V.P.M.N., J.M.D.-M. and E.P.B.; funding acquisition, J.M.D.-M. and E.P.B. All authors have read and agreed to the published version of the manuscript.
Funding
Grant PID2022-139876NB-I00 funded by MCIN/AEI/10.13039/501100011033, FEDER EU. V.P.M.N. was beneficiary of an FPI predoctoral research contract linked to this project. A.G.G. and P.I.O were beneficiaries of predoctoral research contracts from Junta de Castilla y León (Spain) (2022 and 2023 call, respectively). .
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets presented in this study are publicly available in online repositories (SRA of NCBI) under the BioProject PRJNA1224713. (https://www.ncbi.nlm.nih.gov/sra/PRJNA1224713). Accession numbers are: Bc116 Genome – SAMN46863733; Pool aggressive progeny (A) – SAMN46863735; and Pool non-aggressive progeny (B) – SAMN46863734.
Acknowledgments
The authors thank Lucía Albornos Llorente for her valuable advice in microscopy analysis.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Elad, Y.; Pertot, I.; Cotes Prado, A.M.; Stewart, A. Plant hosts of Botrytis spp. In Botrytis – the fungus, the pathogen and its management in agricultural systems. Fillinger, S.; Elad, Y.; Eds. Springer International Publishing, 2016; pp. 413–486. [CrossRef]
- van Kan, J.A.L. Licensed to kill: The lifestyle of a necrotrophic plant pathogen. Trends in Plant Sci. 2006, 11, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A.L. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Groves, J.W.; Loveland, C.A. The connection between Botryotinia fuckeliana and Botrytis cinerea. Mycologia. 1953, 45, 415–425. [Google Scholar] [CrossRef]
- Faretra, F.; Antonacci, E.; Pollastro, S. Improvement of the technique used for obtaining apothecia of Botryotinia fuckeliana (Botrytis cinerea) under controlled conditions. Ann Microbiol. 1988, 38, 29–40. [Google Scholar]
- Faretra, F.; Antonacci, E.; Pollastro, S. Sexual Behaviour and mating system of Botryotinia fuckeliana, teleomorph of Botrytis cinerea. Microbiology. 1988, 134, 2543–2550. [Google Scholar] [CrossRef]
- Lorenz, D.H.; Eichhorn, K.W. Untersuchungen an Botryotinia fuckeliana Whetz.; dem Perfektstadium von Botrytis cinerea Pers. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz. 1983, 90, 1–11. [Google Scholar]
- Beever, R.E.; Parkes, S.L. Mating behaviour and genetics of fungicide resistance of Botrytis cinerea in New Zealand. New Zeal. J. Crop Hort. 1993, 21, 303–310. [Google Scholar] [CrossRef]
- Beever, R.E.; Weeds, P.L. Taxonomy and genetic variation of Botrytis and Botryotinia. In Botrytis: Biology, pathology and control. Elad, Y.; Williamson, B.; Tudzynski, P.; Delen, N.; Eds. Springer Netherlands, 2007; pp. 29–52. [CrossRef]
- Atwell, S.; Corwin, J.; Soltis, N.; Subedy, A.; Denby, K.; Kliebenstein, D.J. Whole genome resequencing of Botrytis cinerea isolates identifies high levels of standing diversity. Front Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Atwell, S.; Corwin, J.A.; Soltis, N.; Zhang, W.; Copeland, D.; Feusier, J.; et al. Resequencing and association mapping of the generalist pathogen Botrytis cinerea. BioRxiv. 2018, p. 489799. [CrossRef]
- Walker, A.S.; Gladieux, P.; Decognet, V.; Fermaud, M.; Confais, J.; Roudet, J.; et al. Population structure and temporal maintenance of the multihost fungal pathogen Botrytis cinerea: Causes and implications for disease management. Environ. Microbiol. 2015, 17, 1261–1274. [Google Scholar] [CrossRef]
- Kerssies, A.; Bosker-van Zessen, A.I.; Wagemakers, C.A.M.; van Kan, J.A.L. Variation in pathogenicity and DNA polymorphism among Botrytis cinerea isolates sampled inside and outside a glasshouse. Plant Dis. 1997, 81, 781–786. [Google Scholar] [CrossRef]
- Giraud, T.; Fortini, D.; Levis, C.; Leroux, P.; Brygoo, Y. RFLP markers show genetic recombination in Botryotinia fuckeliana (Botrytis cinerea) and transposable elements reveal two sympatric species. Mol Biol Evol. 1997, 14, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Fournier, E.; Giraud, T.; Albertini, C.; Brygoo, Y. Partition of the Botrytis cinerea complex in France using multiple gene genealogies. Mycologia. 2005, 97, 1251–1267. [Google Scholar] [CrossRef] [PubMed]
- Acosta Morel, W.; Marques-Costa, T.M.; Santander-Gordón, D.; Fernández, F.A.; Zabalgogeazcoa, I.; de Aldana, B.R.V.; et al. Physiological and population genetic analysis of Botrytis field isolates from vineyards in Castilla y León, Spain. Plant Pathol. 2019, 68, 523–536. [Google Scholar] [CrossRef]
- Amselem, J.; Cuomo, C.A.; van Kan, J.A.L.; Viaud, M.; Benito, E.P.; Couloux, A.; et al. Genomic analysis of the necrotrophic fungal pathogens Sclerotinia sclerotiorum and Botrytis cinerea. PLOS Genetics. 2011, 7, e1002230. [Google Scholar] [CrossRef]
- van Kan, J.A.L.; Stassen, J.H.M.; Mosbach, A.; Van Der Lee, T.A.J.; Faino, L.; Farmer, A.D.; et al. A gapless genome sequence of the fungus Botrytis cinerea. Mol Plant Pathol. 2017, 18, 75–89. [Google Scholar] [CrossRef]
- Soltis, N.E.; Atwell, S.; Shi, G.; Fordyce, R.; Gwinner, R.; Gao, D.; et al. Interactions of tomato and Botrytis cinerea genetic diversity: Parsing the contributions of host differentiation, domestication, and pathogen variation. The Plant Cell. 2019, 31, 502–519. [Google Scholar] [CrossRef]
- Michelmore, R.W.; Paran, I.; Kesseli, R.V. Identification of markers linked to disease-resistance genes by bulked segregant analysis: A rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci U S A. 1991, 88, 9828–9832. [Google Scholar] [CrossRef]
- Magwene, P.M.; Willis, J.H.; Kelly, J.K. The statistics of bulk segregant analysis using next generation sequencing. PLOS Computational Biology. 2011, 7, e1002255. [Google Scholar] [CrossRef]
- Acosta Morel, W.; Anta Fernández, F.; Baroncelli, R.; Becerra, S.; Thon, M.R.; van Kan, J.A.L.; et al. A major effect gene controlling development and pathogenicity in Botrytis cinerea identified through genetic analysis of natural mycelial non-pathogenic isolates. Front Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Soltis, N.E.; Caseys, C.; Zhang, W.; Corwin, J.A.; Atwell, S.; Kliebenstein, D.J. Pathogen genetic control of transcriptome variation in the Arabidopsis thaliana - Botrytis cinerea pathosystem. Genetics. 2020, 215, 253–266. [Google Scholar] [CrossRef]
- Schumacher, J. How light affects the life of Botrytis. Fungal Genetics and Biology. 2017, 106, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, J. Tools for Botrytis cinerea: New expression vectors make the gray mold fungus more accessible to cell biology approaches. Fungal Genetics and Biology. 2012, 49, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, J.; Gautier, A.; Morgant, G.; Studt, L.; Ducrot, P.H.; Le Pêcheur, P.; et al. A functional bikaverin biosynthesis gene cluster in rare strains of Botrytis cinerea is positively controlled by VELVET. PLoS ONE. 2013, 8, e53729. [Google Scholar] [CrossRef]
- Schumacher, J.; Simon, A.; Cohrs, K.C.; Traeger, S.; Porquier, A.; Dalmais, B.; et al. The VELVET complex in the gray mold fungus Botrytis cinerea: Impact of BcLAE1 on differentiation, secondary metabolism, and virulence. Mol Plant Microbe Interact. 2015, 28, 659–674. [Google Scholar] [CrossRef]
- Yang, Q.; Chen, Y.; Ma, Z. Involvement of BcVeA and BcVelB in regulating conidiation, pigmentation and virulence in Botrytis cinerea. Fungal Genetics and Biology. 2013, 50, 63–71. [Google Scholar] [CrossRef]
- Canessa, P.; Schumacher, J.; Hevia, M.A.; Tudzynski, P.; Larrondo, L.F. Assessing the effects of light on differentiation and virulence of the plant pathogen Botrytis cinerea: Characterization of the White Collar Complex. PLoS ONE. 2013, 8, e84223. [Google Scholar] [CrossRef]
- Schumacher, J.; Simon, A.; Cohrs, K.C.; Viaud, M.; Tudzynski, P. The transcription factor BcLTF1 regulates virulence and light responses in the necrotrophic plant pathogen Botrytis cinerea. PLOS Genetics. 2014, 10, e1004040. [Google Scholar] [CrossRef]
- Chen, C.H.; Ringelberg, C.S.; Gross, R.H.; Dunlap, J.C.; Loros, J.J. Genome-wide analysis of light-inducible responses reveals hierarchical light signalling in Neurospora. EMBO J. 2009, 28, 1029–1042. [Google Scholar] [CrossRef]
- Han, K.H.; Han, K.Y.; Yu, J.H.; Chae, K.S.; Jahng, K.Y.; Han, D.M. The nsdD gene encodes a putative GATA-type transcription factor necessary for sexual development of Aspergillus nidulans. Mol. Microbiol. 2001, 41, 299–309. [Google Scholar] [CrossRef]
- Dean, R.; van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Veloso, J. and van Kan, J.A.L. Many shades of grey in Botrytis-host plant interactions. Trends Plant Sci. 2018, 23, 613–622. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef]
- Heller, J.; Tudzynski, P. Reactive oxygen species in phytopathogenic fungi: Signaling, development, and disease. Annu Rev Phytopathol. 2011, 49, 369–390. [Google Scholar] [CrossRef]
- Büttner, P.; Koch, F.; Voigt, K.; Quidde, T.; Risch, S.; Blaich, R.; et al. Variations in ploidy among isolates of Botrytis cinerea: Implications for genetic and molecular analyses. Curr Genet. 1994, 25, 445–450. [Google Scholar] [CrossRef]
- Anta-Fernández, F.; Santander-Gordón, D.; Becerra, S.; Santamaría, R.; Díaz-Mínguez, J.M.; Benito, E.P. Nitric oxide metabolism affects germination in Botrytis cinerea and is connected to nitrate assimilation. J Fungi. 2022, 8, 699. [Google Scholar] [CrossRef]
- Raeder, U.; Broda, P. Rapid preparation of DNA from filamentous fungi. Lett. Appl. Microbiol. 1985, 1, 17–20. [Google Scholar] [CrossRef]
- ten Have, A.; Mulder, W.; Visser, J.; van Kan, J.A. The endopolygalacturonase gene Bcpg1 is required for full virulence of Botrytis cinerea. Mol Plant Microbe Interact. 1998, 11, 1009–1016. [Google Scholar] [CrossRef]
- Reis, H.; Pfiffi, S.; Hahn, M. Molecular and functional characterization of a secreted lipase from Botrytis cinerea. Mol. Plant Pathol. 2005, 6, 257–267. [Google Scholar] [CrossRef]
- Leisen, T.; Bietz, F.; Werner, J.; Wegner, A.; Schaffrath, U.; Scheuring, D.; et al. CRISPR/Cas with ribonucleoprotein complexes and transiently selected telomere vectors allows highly efficient marker-free and multiple genome editing in Botrytis cinerea. PLOS Pathogens. 2020, 16, e1008326. [Google Scholar] [CrossRef]
- Hilber, U.W.; Bodmer, M.; Smith, F.D.; Köller, W. Biolistic transformation of conidia of Botryotinia fuckeliana. Curr Genet. 1994, 25, 124–127. [Google Scholar] [CrossRef]
Figure 1.
Evaluation of the effect of light on pathogenicity of isolates Bc116 and Bc448 on Vitis leaves. (A) Vitis leaves were inoculated with mycelium plugs of both strains and incubated under the indicated light regimes during 96 h. The 16/8 h photoperiod regime was initiated either in the light phase (LD) or in the darkness phase (DL). (B) Quantification of aggressiveness of isolates Bc116 and Bc448 on Vitis leaves under different light conditions (permanent light -LL-, white bars; 16/8 h light/darkness photoperiod -LD-, grey bars; 8/16 h darkness/light photoperiod -DL-, striped bars; permanent darkness -DD-, black bars) estimated as the medium lesion diameter. Bars show the media ± standard deviation (SD) of three independent biological experiments. The letters over each bar represent significant differences between all the conditions assayed that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05).
Figure 1.
Evaluation of the effect of light on pathogenicity of isolates Bc116 and Bc448 on Vitis leaves. (A) Vitis leaves were inoculated with mycelium plugs of both strains and incubated under the indicated light regimes during 96 h. The 16/8 h photoperiod regime was initiated either in the light phase (LD) or in the darkness phase (DL). (B) Quantification of aggressiveness of isolates Bc116 and Bc448 on Vitis leaves under different light conditions (permanent light -LL-, white bars; 16/8 h light/darkness photoperiod -LD-, grey bars; 8/16 h darkness/light photoperiod -DL-, striped bars; permanent darkness -DD-, black bars) estimated as the medium lesion diameter. Bars show the media ± standard deviation (SD) of three independent biological experiments. The letters over each bar represent significant differences between all the conditions assayed that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05).
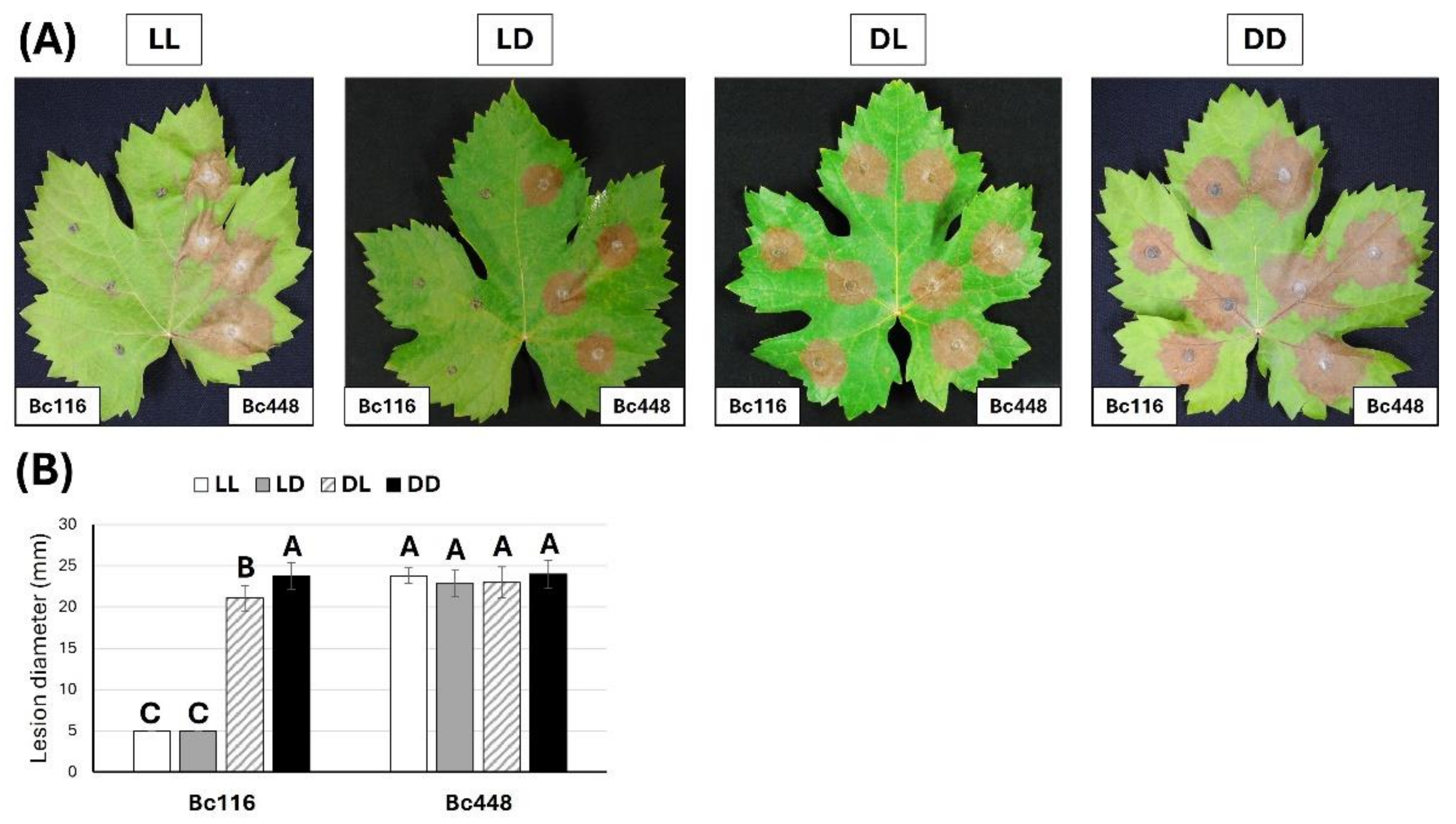
Figure 2.
Evaluation of the effect of light on pathogenicity of isolates Bc116 and Bc448 on bean leaves. Bean leaves were inoculated with mycelium plugs of both strains and incubated under the indicated light regimes. (A) Aspect of the inoculated leaves 24 hpi. (B) Lesions generated by Bc116 and Bc448 24 hpi. At the indicated time point the mycelium plugs were removed, and the plant tissues were stained with lactophenol blue. Images were taken using a Leica DFC495 camera adapted to a Leica 205FA stereomicroscope (Leica Microsystems, Bensheim, Germany). White scale bars, 2 mm; yellow scale bars, 5 mm. (C) Aspect of the inoculated leaves 72 hpi. (D) Quantification of aggressiveness of isolates Bc116 and Bc448 on bean leaves under different light conditions (LL, white bars; 16/8 h LD, grey bars; 8/16 h DL, striped bars; DD, black bars) estimated as the medium lesion diameter 72 hpi. Bars show the media ± SD of three independent biological experiments. The letters over each bar represent significant differences between all the conditions assayed that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05).
Figure 2.
Evaluation of the effect of light on pathogenicity of isolates Bc116 and Bc448 on bean leaves. Bean leaves were inoculated with mycelium plugs of both strains and incubated under the indicated light regimes. (A) Aspect of the inoculated leaves 24 hpi. (B) Lesions generated by Bc116 and Bc448 24 hpi. At the indicated time point the mycelium plugs were removed, and the plant tissues were stained with lactophenol blue. Images were taken using a Leica DFC495 camera adapted to a Leica 205FA stereomicroscope (Leica Microsystems, Bensheim, Germany). White scale bars, 2 mm; yellow scale bars, 5 mm. (C) Aspect of the inoculated leaves 72 hpi. (D) Quantification of aggressiveness of isolates Bc116 and Bc448 on bean leaves under different light conditions (LL, white bars; 16/8 h LD, grey bars; 8/16 h DL, striped bars; DD, black bars) estimated as the medium lesion diameter 72 hpi. Bars show the media ± SD of three independent biological experiments. The letters over each bar represent significant differences between all the conditions assayed that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05).
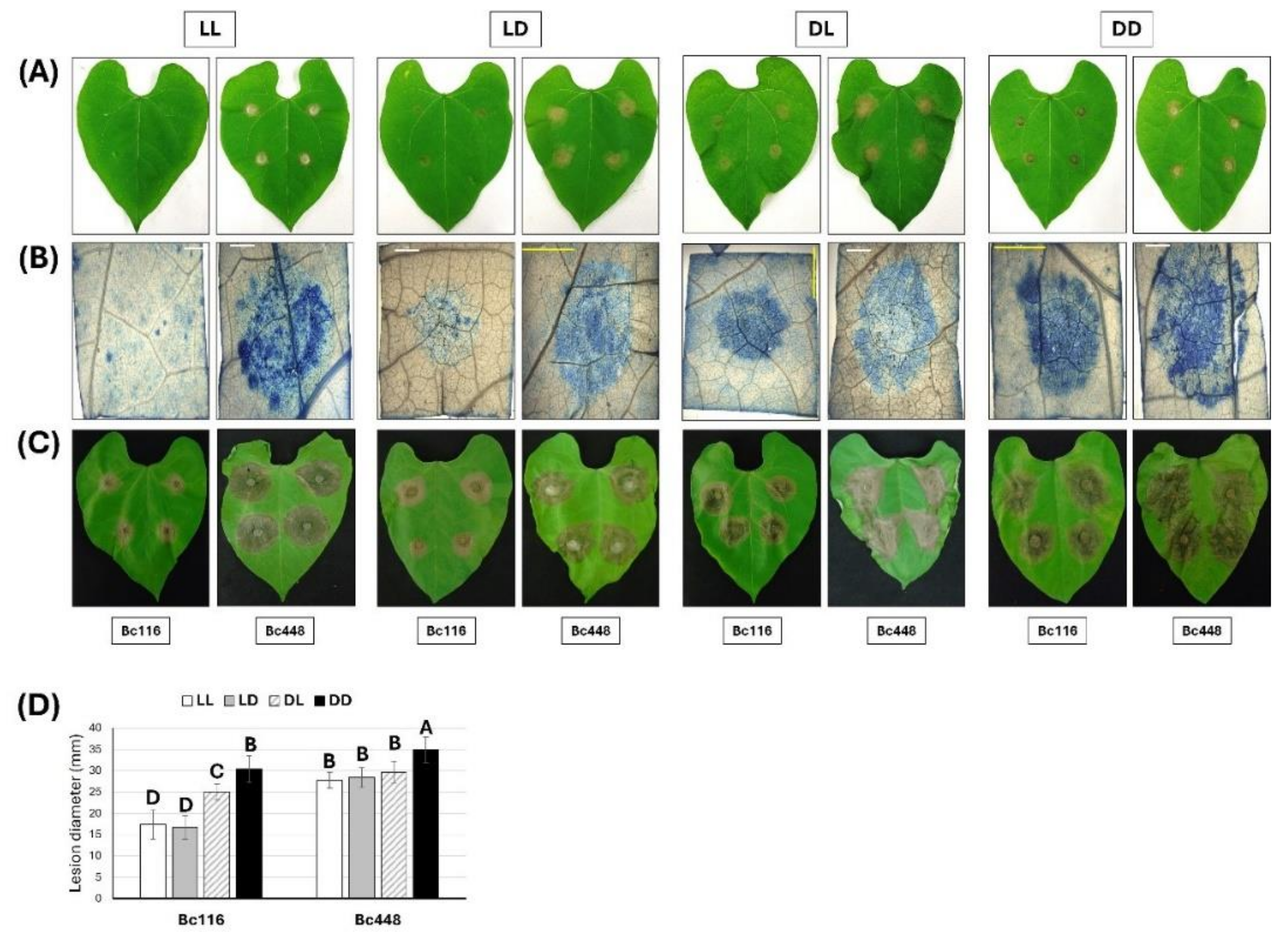
Figure 3.
Light affects the ability of isolate Bc116 to penetrate epidermal onion cells. Onion epidermal strips were inoculated with spores (panels A, C, E and G) or with mycelium plugs (panels B, D, F and H) of isolates Bc116 and Bc448 and incubated under LL or DD conditions. Penetration was evaluated visually at 12 hpi for the inoculations performed with spore suspensions and at 24 hpi for the inoculations performed with mycelium plugs. Samples were stained with lactophenol blue. Yellow arrowheads indicate hyphae penetrating host cells. Scale bars, 50 µm in panels A, C, E and G; 100 µm in panels B, D, F and H.
Figure 3.
Light affects the ability of isolate Bc116 to penetrate epidermal onion cells. Onion epidermal strips were inoculated with spores (panels A, C, E and G) or with mycelium plugs (panels B, D, F and H) of isolates Bc116 and Bc448 and incubated under LL or DD conditions. Penetration was evaluated visually at 12 hpi for the inoculations performed with spore suspensions and at 24 hpi for the inoculations performed with mycelium plugs. Samples were stained with lactophenol blue. Yellow arrowheads indicate hyphae penetrating host cells. Scale bars, 50 µm in panels A, C, E and G; 100 µm in panels B, D, F and H.
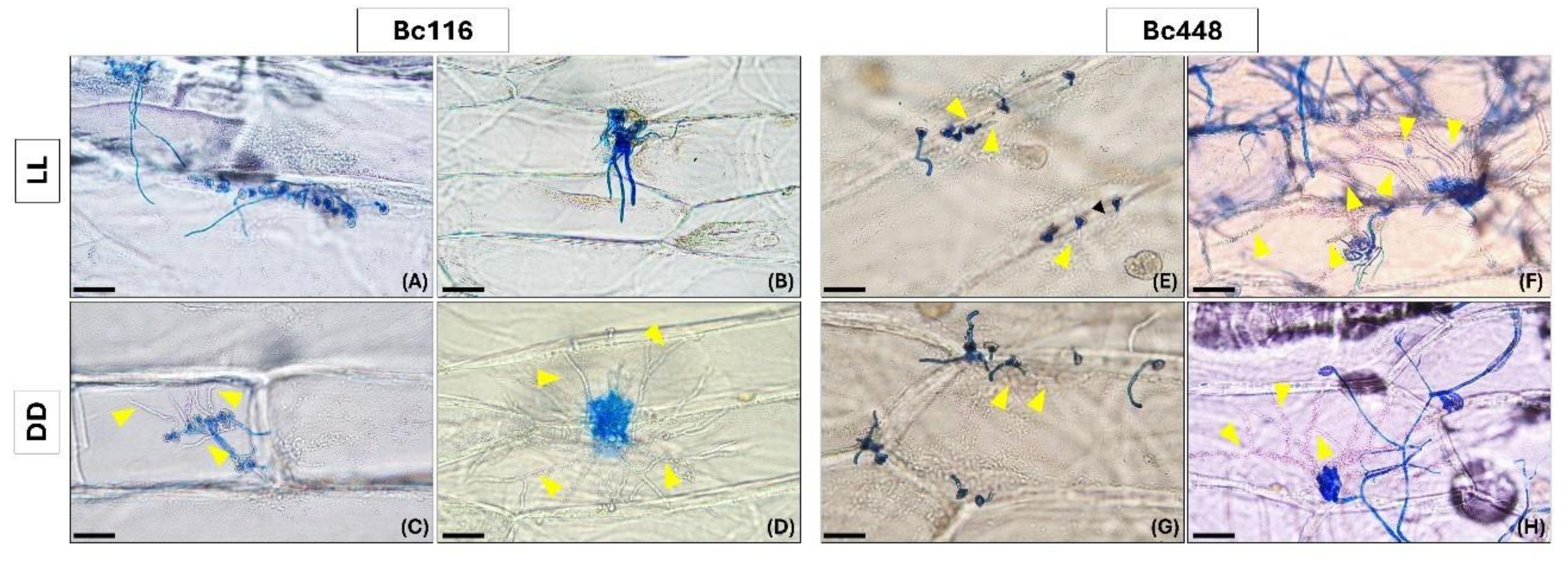
Figure 4.
Effect of light on physiology of isolates Bc116 and Bc448. (A) Germination of conidia on liquid medium in static culture. 60 µL of a 5x105 sp/mL conidial suspension were prepared and placed in the center of empty Petri dishes. The plates were incubated at 22°C under different light conditions (LL, white bars; DD, black bars). The number of conidia in each stage (stages 0, 1, 2) was determined 6 hpi. The bars show the media ± SD of three independent biological experiments. The letters over each bar represent significant differences between conditions assayed for each fungal isolate that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05). Scale bars in images of spores, 10 µm. (B) Effect of light on the conidiation rate of isolates Bc116, Bc448 and B05.10. Mycelium plugs of each fungal isolate were placed in the center of MEA plates and incubated for three weeks at 22°C under different light conditions (LL, white bars; 16/8 LD, grey bars; DD, black bars). Afterwards, conidia were harvested, and the concentration was estimated using a Thoma cell counting chamber. The bars show the media ± SD of three independent biological experiments. The letters over each bar represent significant differences between conditions assayed for each fungal isolate that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05). (C) Saprophytic growth on synthetic media of isolates Bc116, Bc448 and B05.10. 10 µL of a 5x105 sp/mL conidial suspension of each fungal isolate were placed in the center of MEA plates and the diameter of the colony was estimated 5 days post-inoculation (dpi). Plates were incubated at 22°C and different light conditions (LL, white bars; 16/8 h LD, grey bars; DD, black bars). Bars show the media ± SD of three independent biological experiments. The letters over each bar represent significant differences between all the conditions assayed that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05). (D) Production of sclerotia by isolates Bc116, Bc448 and B05.10. Mycelium plugs were placed in the center of MEA plates that were incubated at 2-4°C and DD for 4 weeks.
Figure 4.
Effect of light on physiology of isolates Bc116 and Bc448. (A) Germination of conidia on liquid medium in static culture. 60 µL of a 5x105 sp/mL conidial suspension were prepared and placed in the center of empty Petri dishes. The plates were incubated at 22°C under different light conditions (LL, white bars; DD, black bars). The number of conidia in each stage (stages 0, 1, 2) was determined 6 hpi. The bars show the media ± SD of three independent biological experiments. The letters over each bar represent significant differences between conditions assayed for each fungal isolate that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05). Scale bars in images of spores, 10 µm. (B) Effect of light on the conidiation rate of isolates Bc116, Bc448 and B05.10. Mycelium plugs of each fungal isolate were placed in the center of MEA plates and incubated for three weeks at 22°C under different light conditions (LL, white bars; 16/8 LD, grey bars; DD, black bars). Afterwards, conidia were harvested, and the concentration was estimated using a Thoma cell counting chamber. The bars show the media ± SD of three independent biological experiments. The letters over each bar represent significant differences between conditions assayed for each fungal isolate that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05). (C) Saprophytic growth on synthetic media of isolates Bc116, Bc448 and B05.10. 10 µL of a 5x105 sp/mL conidial suspension of each fungal isolate were placed in the center of MEA plates and the diameter of the colony was estimated 5 days post-inoculation (dpi). Plates were incubated at 22°C and different light conditions (LL, white bars; 16/8 h LD, grey bars; DD, black bars). Bars show the media ± SD of three independent biological experiments. The letters over each bar represent significant differences between all the conditions assayed that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05). (D) Production of sclerotia by isolates Bc116, Bc448 and B05.10. Mycelium plugs were placed in the center of MEA plates that were incubated at 2-4°C and DD for 4 weeks.

Figure 5.
Sensitivity to oxidative stress of isolates Bc116, Bc448 and B05.10. Mycelium plugs from the edge of actively growing colonies were placed in the center of MEA plates. The oxidative stress was provided by the addition of 7.5 mM H2O2 to the media. Plates were incubated at 22°C under different light conditions (LL, white bars; 16/8 h LD, grey bars; DD, black bars) and the colony diameter was estimated 96 hpi. Bars show the media ± SD of three independent biological experiments. The letters over each bar represent significant differences between the conditions assayed for each fungal isolate that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05). Representative images of colony morphology and colony diameter for each strain and condition are shown.
Figure 5.
Sensitivity to oxidative stress of isolates Bc116, Bc448 and B05.10. Mycelium plugs from the edge of actively growing colonies were placed in the center of MEA plates. The oxidative stress was provided by the addition of 7.5 mM H2O2 to the media. Plates were incubated at 22°C under different light conditions (LL, white bars; 16/8 h LD, grey bars; DD, black bars) and the colony diameter was estimated 96 hpi. Bars show the media ± SD of three independent biological experiments. The letters over each bar represent significant differences between the conditions assayed for each fungal isolate that were tested using an ANOVA analysis followed by a Tukey’s HSD test (P < 0.05). Representative images of colony morphology and colony diameter for each strain and condition are shown.

Figure 6.
Mapping the genome region linked to the Bc116 phenotype by BSA in the cross Bc116 x Bc448. (A, B) The figure plots the SNP index of Bc448 specific variants or of Bc116 specific variants (Y-axis) across Chr13 (A) or Chr14 (B) coordinates (X-axis). The box in panel (B) delimitates a region in Chr14 where the SNP index reaches maximal values for Bc116 specific variants and minimal values for Bc448 specific variants. (C) Detailed analysis of the mapped region indicating the genes within this region annotated in the B05.10 genome. The blue graph over the linear representation of the mapped area represents sequencing reads coverage when Bc116 genome reads are aligned with the B05.10 genome. A region with no coverage is observed around the 5’-upstram region of gene bcltf1. The annealing positions of the oligonucleotides P1 (bcltf1-c2F) and P2 (bcltf1-c2R), used to amplify both the B05.10 and Bc116 alleles are indicated.
Figure 6.
Mapping the genome region linked to the Bc116 phenotype by BSA in the cross Bc116 x Bc448. (A, B) The figure plots the SNP index of Bc448 specific variants or of Bc116 specific variants (Y-axis) across Chr13 (A) or Chr14 (B) coordinates (X-axis). The box in panel (B) delimitates a region in Chr14 where the SNP index reaches maximal values for Bc116 specific variants and minimal values for Bc448 specific variants. (C) Detailed analysis of the mapped region indicating the genes within this region annotated in the B05.10 genome. The blue graph over the linear representation of the mapped area represents sequencing reads coverage when Bc116 genome reads are aligned with the B05.10 genome. A region with no coverage is observed around the 5’-upstram region of gene bcltf1. The annealing positions of the oligonucleotides P1 (bcltf1-c2F) and P2 (bcltf1-c2R), used to amplify both the B05.10 and Bc116 alleles are indicated.
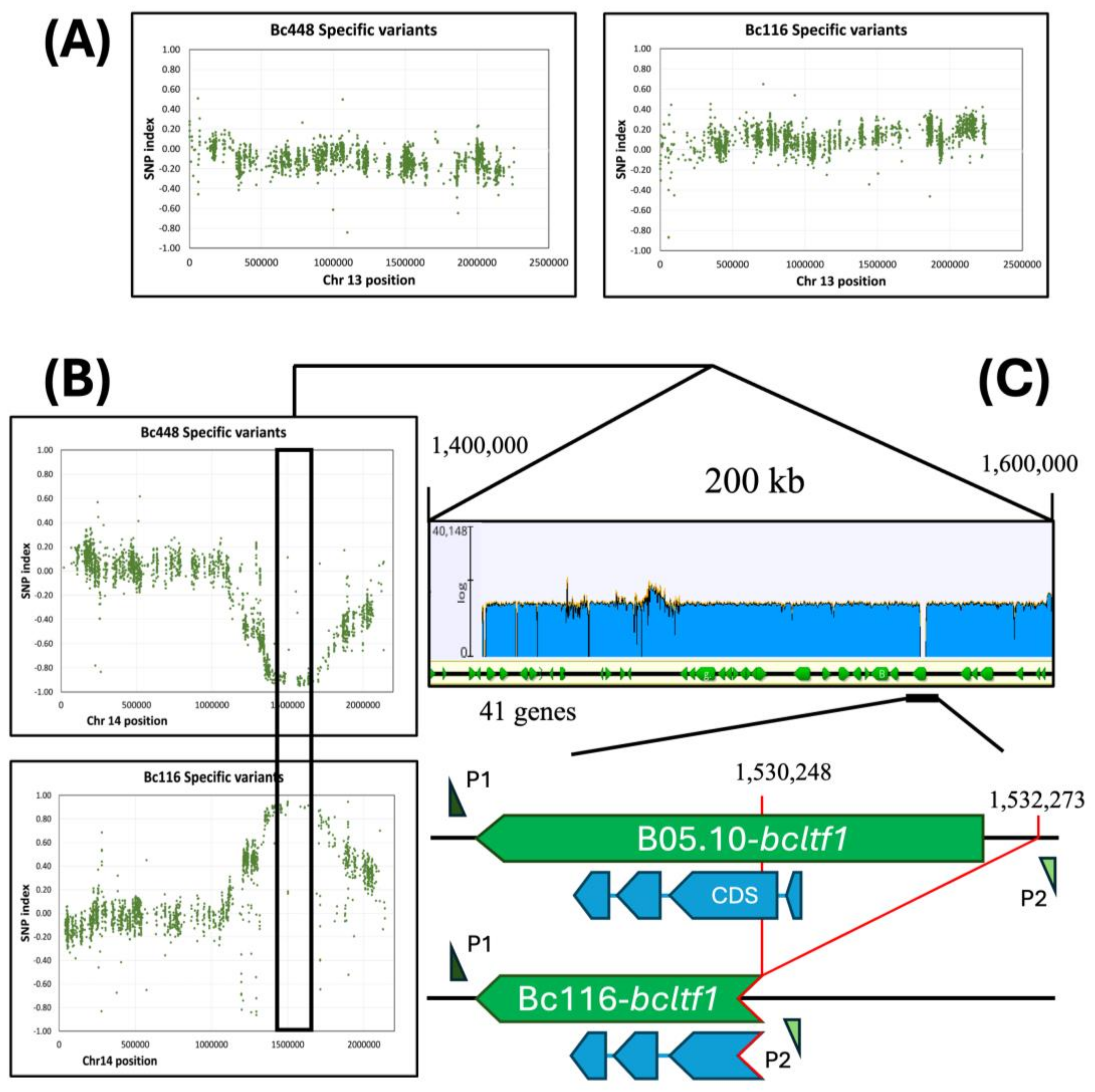
Figure 7.
Genetic and phenotypic analysis of Bc116-bcltf1 complemented transformants. (A) PCR based analysis of transformants obtained with the B05.10 bcltf1 allele. Reactions were carried out with genomic DNA of B05.10, Bc116, Bc448 or transformants T2 and T7 as template. The sizes of the diagnostic bands are indicated. MM: GeneRuler 1 kb Plus DNA Ladder (ThermoFisher Scientific). (B) Inoculations on Vitis leaves of Bc116, B05.10, Bc448 isolates and transformants T2 and T7. Leaves were inoculated with mycelium plugs of each isolate and incubated at 22°C under a 16/8 h photoperiod (LD). Images were taken 96 hpi. (C) Phenotype of the fungal isolates during growth in MEA plates (4 days at 22°C under LD conditions). (D) Pattern of production of sclerotia of the fungal isolates on MEA plates. Plates were incubated at 2-4°C under DD for 4 weeks.
Figure 7.
Genetic and phenotypic analysis of Bc116-bcltf1 complemented transformants. (A) PCR based analysis of transformants obtained with the B05.10 bcltf1 allele. Reactions were carried out with genomic DNA of B05.10, Bc116, Bc448 or transformants T2 and T7 as template. The sizes of the diagnostic bands are indicated. MM: GeneRuler 1 kb Plus DNA Ladder (ThermoFisher Scientific). (B) Inoculations on Vitis leaves of Bc116, B05.10, Bc448 isolates and transformants T2 and T7. Leaves were inoculated with mycelium plugs of each isolate and incubated at 22°C under a 16/8 h photoperiod (LD). Images were taken 96 hpi. (C) Phenotype of the fungal isolates during growth in MEA plates (4 days at 22°C under LD conditions). (D) Pattern of production of sclerotia of the fungal isolates on MEA plates. Plates were incubated at 2-4°C under DD for 4 weeks.
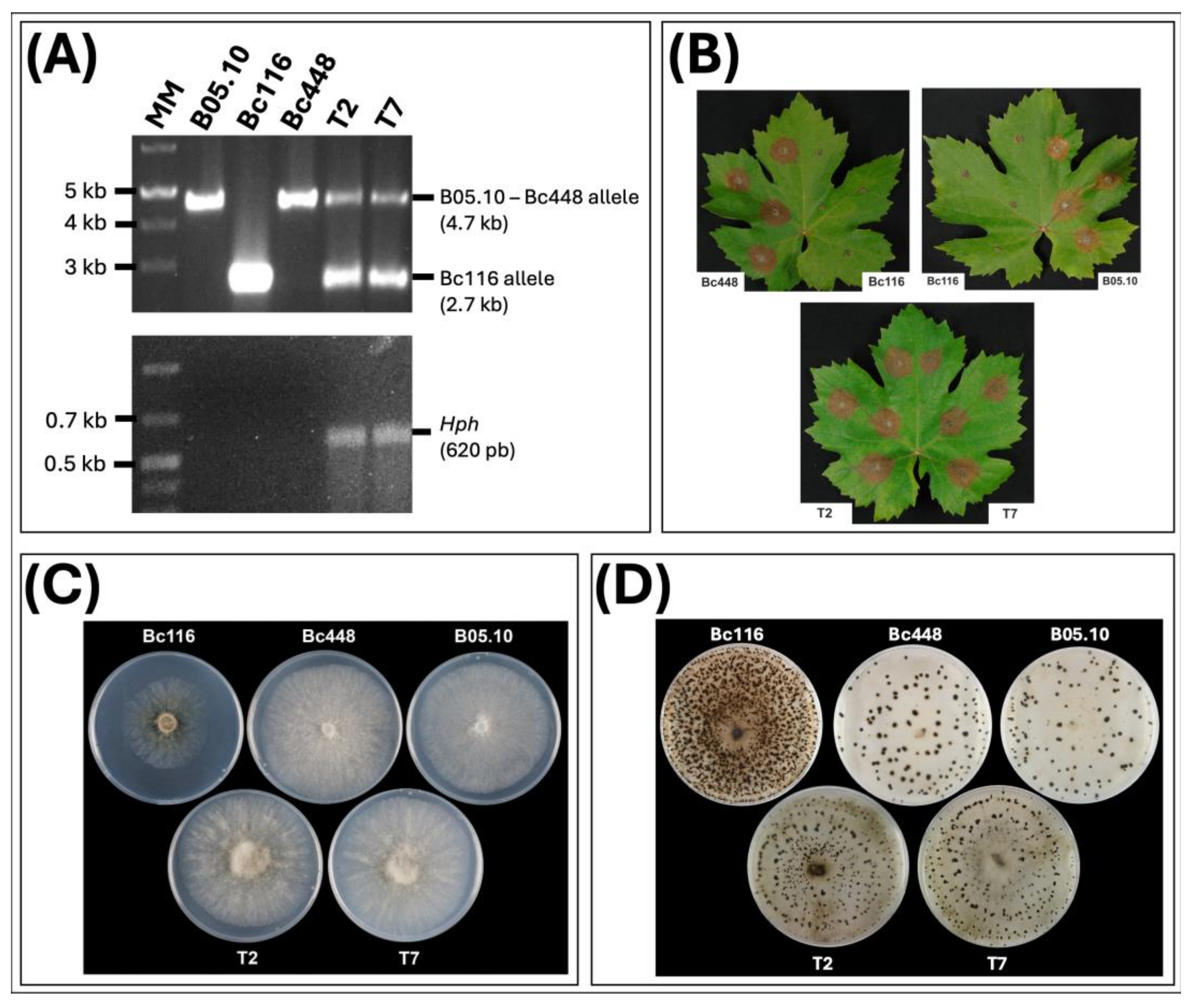

Table 1.
Total number of SNPs identified in the genomes of isolates Bc448 and Bc116 in comparison with the B05.10 genome. The number of SNPs exclusive of each isolate is presented.
Table 1.
Total number of SNPs identified in the genomes of isolates Bc448 and Bc116 in comparison with the B05.10 genome. The number of SNPs exclusive of each isolate is presented.
| B05.10 Genome | Bc448 SNPs | Bc116 SNPs | |||
|---|---|---|---|---|---|
| Chr size (kb) | Total number | Exclusive | Total number | Exclusive | |
| Chr 1 | 4109 | 23260 | 9129 | 26619 | 12128 |
| Chr 2 | 3341 | 17103 | 5335 | 19466 | 7698 |
| Chr 3 | 3227 | 13364 | 4379 | 15161 | 6176 |
| Chr 4 | 2472 | 16024 | 7069 | 18731 | 9776 |
| Chr 5 | 2959 | 11864 | 4260 | 12701 | 5097 |
| Chr 6 | 2726 | 17544 | 5529 | 19339 | 7324 |
| Chr 7 | 2652 | 17831 | 5715 | 18805 | 6689 |
| Chr 8 | 2617 | 14368 | 5583 | 15433 | 6649 |
| Chr 9 | 2548 | 16793 | 5866 | 18543 | 7616 |
| Chr 10 | 2419 | 13662 | 4484 | 15798 | 6620 |
| Chr 11 | 2360 | 12485 | 3928 | 15300 | 6743 |
| Chr 12 | 2353 | 13996 | 5298 | 14802 | 6104 |
| Chr 13 | 2258 | 10921 | 3949 | 13106 | 6134 |
| Chr 14 | 2138 | 14089 | 5718 | 15952 | 7581 |
| Chr 15 | 2028 | 11469 | 3246 | 14335 | 6112 |
| Chr 16 | 1970 | 10795 | 3556 | 13634 | 6395 |
| Chr 17 | 247 | 107 | 74 | 54 | 51 |
| SUM | 42424 | 235675 | 83118 | 267779 | 114893 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



