Submitted:
14 February 2025
Posted:
14 February 2025
You are already at the latest version
Abstract
Background/Objectives: Bisphenol A (BPA) is a pervasive environmental endocrine disruptor, and the lifestyle associated with a high-fat diet is becoming increasingly common. The precise relationship between these factors and the high incidence of insulin resistance and type 2 diabetes worldwide in recent years remains unclear. Given the critical role of skeletal muscle in maintaining systemic glucose metabolism, this study aimed to explore the potential effects of co-exposure to BPA and a high-fat diet on insulin signaling molecules and GLUT4 translocation in mouse gastrocnemius muscle and C2C12 myotubes through both in vivo and in vitro experiments. Methods: We observed the effects of 90 days of BPA and high-fat diet treatment on animal insulin resistance, as well as the expression levels of potential signaling molecules and proteins related to GLUT4 translocation. In vitro experiments involved co-treatment of C2C12 myotubes with BPA and palmitic acid to observe their effects on insulin signaling molecules, GLUT4 translocation, and insulin resistance. Results: Animals co-exposed to BPA and a high-fat diet exhibited a significant increase in insulin resistance indices and impaired oral glucose tolerance. During this process, the expression levels of insulin signaling molecules were markedly reduced, and GLUT4 translocation was significantly blunted. In vitro cellular experiments further demonstrated that BPA and palmitic acid could inhibit the expression levels of GLUT4 on the cell membrane, i.e., the translocation of GLUT4 from the nucleus to the cell membrane was significantly suppressed. Conclusions: Co-exposure to BPA and a high-fat diet significantly affected the expression levels of insulin signaling molecules in gastrocnemius tissue and C2C12 cells, which may be closely related to their effects on inducing insulin resistance and affecting GLUT4 translocation.
Keywords:
Bisphenol A
; High-Fat Diet
; Palmitic Acid
; Glucose Transporter Type 4
; Insulin Resistance
1. Introduction
Diabetes Mellitus is a common chronic metabolic disease. Model predictions suggest that by 2045, the number of diabetic patients worldwide could reach 629 million [1]. Among these, Type 2 Diabetes Mellitus (T2D) is the most prevalent form.
The etiology of T2D is highly complex, potentially involving genetics, behavior, environment, nutritional factors, and their intricate interplay [2]. With the improvement of living conditions and the proliferation of emerging dietary methods such as takeout food, a high-fat diet lifestyle has become a significant health threat, and the incidence of related diseases has been increasing annually. Research has found that a High Fat Diet (HFD) may induce T2DM by disrupting insulin-mediated metabolic functions [3,4,5]. Additionally, studies indicate that Endocrine Disrupting Chemicals (EDCs) may cause insulin resistance and T2DM by altering glucose homeostasis and insulin secretion [6,7]. Bisphenol A (BPA), a typical EDC, is widely used in electronic devices, plastic bottles, linings of metal food cans, thermal paper, and for synthesizing polycarbonate, epoxy resins, and other polymeric materials [8]. Due to its widespread application and resistance to degradation [9], humans are universally and continuously exposed to BPA [10,11]. Epidemiological studies have shown a correlation between BPA exposure and an increased risk of insulin resistance [12].
Insulin resistance (IR), a key process and primary pathological mechanism in the development of T2DM, is characterized by elevated fasting blood glucose, reduced insulin sensitivity, and impaired glucose tolerance [13,14]. Since skeletal muscle accounts for approximately 80% of postprandial glucose uptake [15,16], it is considered a critical organ for regulating blood glucose levels, and skeletal muscle insulin resistance is often regarded as a key link and a major factor affecting the onset of T2DM. Studies have shown that the insulin signaling pathway and the translocation of Glucose Transporter-4 (GLUT4) play an essential role in glucose uptake and metabolism [17,18,19]. Rab proteins have been identified as participants in the mobilization of insulin-stimulated GLUT4 storage vesicles (GSVs) in adipocytes and skeletal muscle cells, with Rab8A and Rab13 being predominant in skeletal muscle [20]. Research has found that high concentrations of BPA can affect the function of Rab proteins [21], but the impact of environmentally relevant doses of BPA on Rab proteins remains unclear. Furthermore, SNARE complex proteins play a crucial role in GLUT4 transport and plasma membrane fusion [22,23], with SNAP-23, syntaxin-4, and VAMP-2 being highly expressed in skeletal muscle tissue and controlling GLUT4 transport in muscle cells [24,25]. Their dysfunction and changes in SNARE protein levels can cause T2DM [26].
In recent years, a high-fat diet has become a major lifestyle associated with an increase in the incidence of diabetes [27]. Foods rich in fat may be related to simultaneous exposure to BPA and HFD. Therefore, it is necessary to investigate whether BPA and a high-fat diet could exacerbate insulin resistance and explore the toxic effects of the combined exposure to BPA and HFD on insulin signaling pathway-related molecules. Utilizing C2C12 cells to further study the impact of co-exposure to BPA and palmitic acid on GLUT4 translocation will provide data support for a better understanding of the association and mechanisms between the combined exposure to BPA and a high-fat diet and impaired muscle glucose metabolism.
2. Materials and Methods
2.1. Animal Experiment Design
Sixty 6-8 week-old male SPF-grade C57BL/6J mice were purchased from the Guangdong Provincial Medical Laboratory Animal Center. After one week of adaptive feeding, the animals were randomly divided into six groups of ten based on body weight. The factorial design based on two factors (i.e., exposure to BPA or not and exposure to a high-fat diet or not) resulted in four groups: the control group (non-high-fat, no BPA, C0), the high-fat group (high-fat, no BPA, C1), the 50 mg/kg/day BPA group (non-high-fat, with BPA, C2), and the 50 mg/kg/day BPA + high-fat group (with high-fat and BPA, TM). Additionally, to investigate the dose-response relationship of BPA under high-fat conditions, two more groups were added: the 5 mg/kg/day BPA + high-fat group (TL) and the 500 mg/kg/day BPA + high-fat group (TH). Regular chow was obtained from the Guangdong Provincial Medical Laboratory Animal Center, and the high-fat diet (45% energy from fat) was purchased from Nantong Troph Animal Feed Co., Ltd. BPA was dissolved in corn oil and administered by gavage once daily at 9 AM for 90 days. Mouse body weight was measured at fixed times each week, and the trends were recorded and analyzed. During the experiment, animals had free access to water, and the light/dark cycle was 12 hours/12 hours (changing at 7:00 AM and 7:00 PM), with a temperature maintained at (22 ± 2) °C and a relative humidity of 40-70%. At the end of 90 days, animals were anesthetized with pentobarbital sodium, blood was collected via the inferior vena cava, serum was separated and stored at -80°C, and skeletal muscle (gastrocnemius) was isolated for various parameter measurements.
2.2. Fasting Blood Glucose Measurement
At day 90 of the experiment, after a 12-hour fast, mouse tail vein serum was collected and tested using a Roche glucometer.
2.3. Fasting Serum Insulin Measurement
After 90 days of feeding, mice were euthanized, blood was drawn from the inferior vena cava, and centrifuged at 4°C at 12,000 rpm for 10 minutes. The supernatant was used to measure serum insulin with an insulin ELISA kit.
2.4. Mouse HOMA-IR Measurement
Insulin resistance (IR) was assessed by combining fasting insulin (FI) and fasting glucose (FG) to reflect the relationship between blood glucose and insulin levels, with the HOMA-IR index as the standard [28]. The formula is: HOMA-IR = FI (mIU/L) × FG (mmol/L) / 22.5.
2.5. Glucose Tolerance Test
The glucose tolerance test (GTT) was performed at day 90 (animals were fasted for 12 hours before the experiment). The tail tip of the mouse was cut with scissors, and blood was squeezed onto the Roche glucometer to measure the fasting blood glucose, i.e., the 0-minute glucose value. Then, glucose was injected intraperitoneally (1g/kg body weight), and timing started after the injection. Blood glucose values were measured and recorded at 15, 30, 60, 90, and 120 minutes post-injection.
2.6. Immunohistochemistry for Protein Expression
Immunohistochemistry (IHC) was performed on tissues to observe the localization and expression level of GLUT4. Paraffin sections were deparaffinized and hydrated, then incubated with GLUT4 antibody at 4°C overnight, followed by incubation with goat anti-rabbit secondary antibody at room temperature for 1 hour. Staining was performed according to the DAB staining kit, and sections were observed and analyzed after being mounted with neutral gum.
2.7. Cell Experiment Design
C2C12 myoblasts were cultured in high-glucose DMEM medium containing 10% fetal bovine serum, 100U/ml penicillin, and 100μg/mL streptomycin at 37°C in an incubator with 5% CO2. When the cells reached 80% confluence, the medium was changed to DMEM medium containing 2% horse serum, 100U/ml penicillin, and 100μg/mL streptomycin to induce differentiation. After 5 days of induction, multinucleated myotubes were formed. After the C2C12 myoblasts differentiated into mature myotubes, they were treated with PA and BPA in DMEM medium containing 3% FBS for 48 hours. Dose groups were determined by preliminary experiments such as the CCK8 cell viability assay and the 2-NBDG glucose uptake assay as follows: control group (containing 2% BSA and 1% DMSO), PA group (200μM PA + 1% DMSO), BPA group (102nM BPA + 2% BSA), PA + BPA group (200μM PA + 102nM BPA). This study used C2C12 myoblasts with fewer than 8 passages.
2.8. 2-NBDG Glucose Uptake Assay
After cell treatment, 2-NBDG was added to Krebs buffer containing 2% FA-free bovine serum albumin, and the cells were incubated with 100nM insulin for 2 hours. Cells were then lysed with cell lysis buffer for 10 minutes, and the supernatant was centrifuged at 4°C at 16,000×g for 15 minutes. Fluorescence intensity was measured using a fluorescence plate reader at 475nm/550nm (excitation wavelength/emission wavelength). BCA quantification and protein correction were performed for each well to determine the glucose uptake of control and experimental group myotubes.
2.9. Cellular Immunofluorescence Staining for GLUT4
After cell treatment, cells were fixed with 4% paraformaldehyde at room temperature for 1 hour, permeabilized with 0.2% Triton X-100, and then blocked with 30% FBS at 37°C for 30 minutes. Anti-GLUT4 antibody was added and incubated overnight, followed by incubation with fluorescent goat anti-rabbit IgG at 37°C for 1 hour. Nuclei were stained with DAPI for 10 minutes, and cells were observed and photographed under a fluorescence microscope.
2.10. Western Blotting for AKT-GLUT4 Signaling Molecules
Proteins from gastrocnemius tissue or C2C12 cells were extracted using tissue lysis buffer or cell lysis buffer. Proteins were incubated with monoclonal antibodies for AKT (CST, 4685), p-AKTSer473 (CST, 4060), p-AKTThr308 (CST, 13038), Rab8A (ab188674), Rab13 (SAB4200057), Syntaxin4 (ab184545), VAMP2 (ab181869), GLUT4 (ab33780), GSK3β (CST, 12456), and pGSK3βSer9 (CST, 9322), followed by incubation with horseradish peroxidase-labeled goat anti-rabbit secondary antibodies. Bands were developed, and band density was analyzed using Image J. The relative expression of proteins was calculated as the ratio of the band density of the protein to that of GAPDH, with the control group normalized to 1 for analysis.
2.11. Statistical Analysis
All quantitative data are expressed as mean ± standard error of the mean (SEM). Repeated measures ANOVA was used for mouse body weight, and 2×2 factorial ANOVA was used to assess the interaction effects of low-dose BPA and high-fat diet in the in vivo animal experiments for groups C0, C1, C2, and TM, as well as the in vitro cell experiments for the four groups. One-way ANOVA was used for multiple group comparisons, and LSD-t tests were used for pairwise comparisons of multiple sample means. A P-value of less than 0.05 was considered statistically significant. Image analysis was performed using Image J (National Institutes of Health, Germany), and data analysis was conducted using GraphPad Prism 7 (GraphPad Software, USA) software for graphing.
3. Results
3.1. BPA in Combination with a High-Fat Diet Exacerbates Insulin Resistance
Repeated measures ANOVA results showed a significant interaction effect between different treatment methods and changes over time on the body weight of male and female animals (F(male) = 4.794, F(female) = 5.167, P < 0.01); the effect of different treatments on mouse body weight was statistically significant (F(male) = 5.873, F(female) = 16.000, P < 0.001). Animals in the high-fat diet treatment groups (C1, TL, TM, TH) had significantly higher body weights compared to those in the normal diet groups (C0, C2), as shown in Figure 1A-1B.
Glucose tolerance test results indicated that the blood glucose levels of animals in the normal diet group gradually returned to the initial value, while the blood glucose levels of animals in the other groups remained higher than the initial value. Analysis of the area under the curve (AUC) showed that the AUC for the glucose tolerance test of animals in groups C1, TL, TM, and TH were all higher than that of the control group, suggesting that co-exposure to BPA and a high-fat diet can lead to abnormal glucose tolerance in animals. Factorial analysis results showed an interaction between BPA and high-fat diet treatment on blood glucose levels in male mice (F = 7.770, P < 0.05).
Calculation of insulin resistance and insulin sensitivity indices based on fasting blood glucose and fasting serum insulin values showed that compared to the control group, all groups exhibited increased insulin resistance and decreased insulin sensitivity. These results indicate that co-exposure to BPA and a high-fat diet exacerbated insulin resistance in mice.
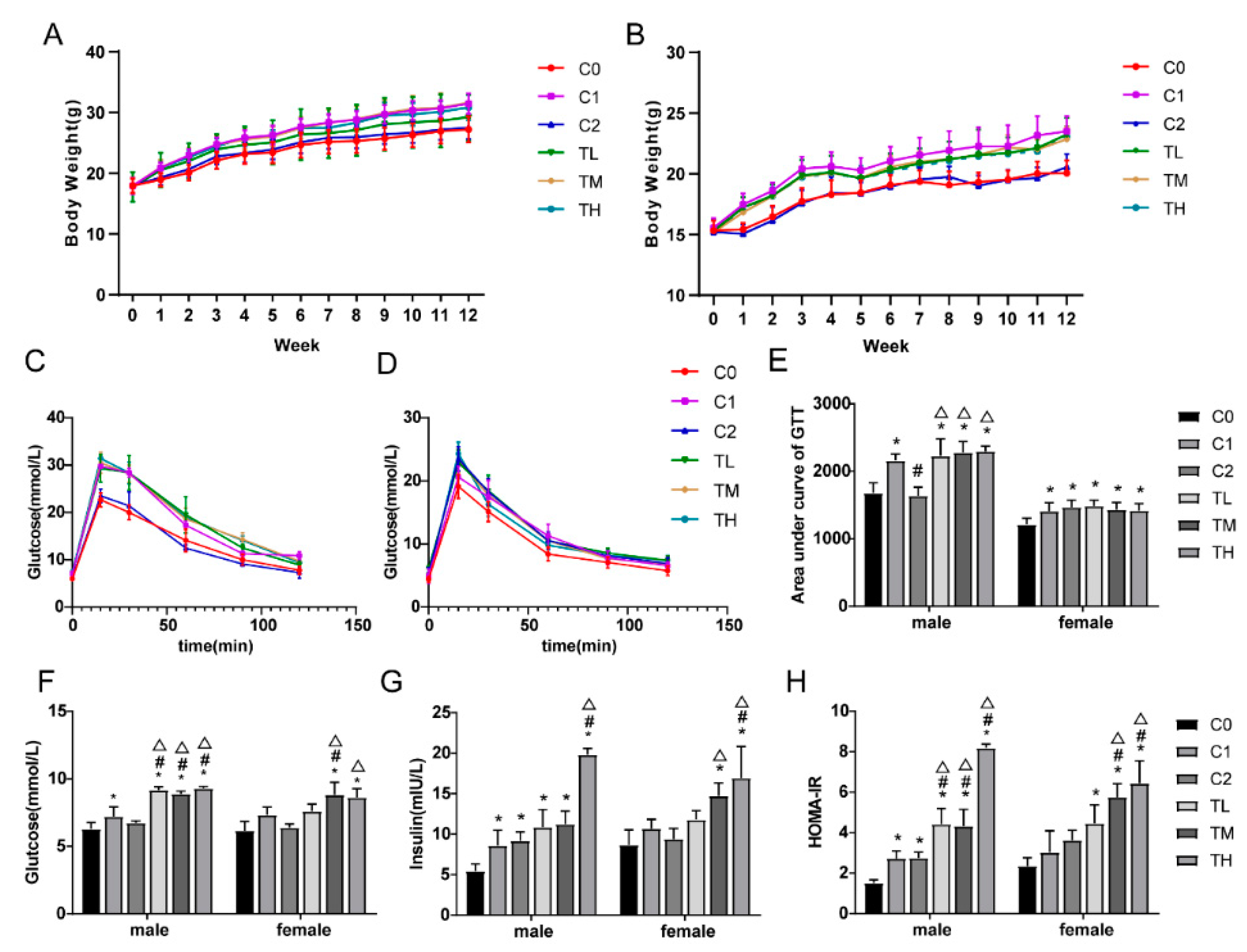
Figure 1.
BPA in combination with a high-fat diet exacerbates insulin resistance. After 12 weeks of feeding with BPA and a high-fat diet, gastrocnemius tissue was collected for experimentation. Results are presented as mean ± SD, n = 10. (A) Male mouse body weight; (B) Female mouse body weight; (C) Oral glucose tolerance test (OGTT) results in male mice; (D) Oral glucose tolerance test (OGTT) results in female mice; (E) Area under the glucose tolerance curve for both male and female mice; (F) Fasting blood glucose levels in both male and female mice; (G) Fasting insulin levels in both male and female mice; (H) Insulin resistance index levels in both male and female mice. Compared to the C0 group, * P<0.05; compared to the C1 group, # P<0.05; compared to the C2 group, △ P<0.05.
Figure 1.
BPA in combination with a high-fat diet exacerbates insulin resistance. After 12 weeks of feeding with BPA and a high-fat diet, gastrocnemius tissue was collected for experimentation. Results are presented as mean ± SD, n = 10. (A) Male mouse body weight; (B) Female mouse body weight; (C) Oral glucose tolerance test (OGTT) results in male mice; (D) Oral glucose tolerance test (OGTT) results in female mice; (E) Area under the glucose tolerance curve for both male and female mice; (F) Fasting blood glucose levels in both male and female mice; (G) Fasting insulin levels in both male and female mice; (H) Insulin resistance index levels in both male and female mice. Compared to the C0 group, * P<0.05; compared to the C1 group, # P<0.05; compared to the C2 group, △ P<0.05.
3.2. Co-Exposure to BPA and a High-Fat Diet Affects the Expression Levels of Insulin Signaling Molecules in Gastrocnemius Tissue
The expression levels of key molecules in insulin signaling transduction in animal gastrocnemius tissue showed a trend of gradual decrease in the expression levels of pAKTSer473 in the TL, TM, and TH groups compared to the control group. Image analysis and statistical analysis revealed a significant reduction in the expression level of pAKTSer473 in the TH group (P<0.05), with a statistically significant difference (Figure 2A-2C). The expression levels of key molecules regulating the translocation and fusion of GLUT4 with the plasma membrane, including Rab8A, VAMP2, and Syntaxin4, also showed the same trend, with statistically significant differences compared to the control group, as shown in Figure 2D-2G.
Furthermore, as shown in Figure 2D, 2H, the expression levels of GLUT4 in male animals of the TM and TH groups were reduced compared to the control group (P<0.05). Immunohistochemical results also showed weakened expression of GLUT4 in the gastrocnemius tissue of the TM and TH groups, significantly lower than the corresponding control groups (Figure 2I-2J).
Activation of the AKT signaling pathway can promote the phosphorylation of GSK3β, thereby inhibiting the activity of glycogen synthase, leading to reduced glycogen synthesis, which can cause an increase in blood glucose levels and is one of the key factors in the development of insulin resistance. In this study, the expression level of GSK3β in male mice of the TM group was higher than that of the control group, with a statistically significant difference (P<0.05).
Compared with the control group, the protein expression levels of pGSK3βSer9 in the gastrocnemius tissue of animals in the TL, TM, and TH groups were significantly reduced in both male and female mice (P<0.05). Especially in the TM and TH groups of female mice, the expression levels of pGSK3βSer9 protein were not only lower than the C0 group (blank control group) but also significantly lower than the C1 group (high-fat diet treatment group), with statistically significant differences (P<0.05). These findings are consistent with the trend of increased fasting blood glucose levels, further supporting the hypothesis that BPA and a high-fat diet may exacerbate insulin resistance, with specific data shown in Figure 2K-2M.
3.3. BPA in Combination with Palmitic Acid Significantly Reduces Glucose Uptake Levels in C2C12 Cells
To assess the impact of BPA and palmitic acid (PA) exposure on cell viability, differentiated C2C12 cells were treated with various concentrations of BPA (1-105 nM) and PA (100-700 μM) for 48 hours, and cell activity was evaluated using the CCK8 assay. The results are shown in Figure 3A, where compared to the control group, cell activity significantly decreased with BPA at concentrations of 103, 104, and 105 nM (P<0.05). At 400 μM, PA reduced cell activity to 61% (Figure 3B). Therefore, BPA concentrations of 102 nM, 103 nM, 104 nM, and PA concentrations of 200 μM, 300 μM, and 450 μM were selected for glucose uptake experiments to determine the optimal combined dosage. Enzyme-linked immunosorbent assay (ELISA) analysis results showed (Figure 3C, 3D) that compared to the insulin-stimulated control group, glucose uptake significantly decreased after treatment with 104 nM BPA and 300 μM, 450 μM PA for 48 hours (P<0.05). Considering multiple factors such as cell activity and glucose uptake, a combination of 300 μM PA and 102 nM, 103 nM BPA for 48 hours was chosen, at which point, glucose uptake in all groups was significantly lower than in the control group (P<0.05), indicating that 300 μM PA and 102 nM, 103 nM BPA can induce insulin resistance (IR) in C2C12 cells (Figure 3E). Further research results indicated that after 48 hours of exposure to 200 μM PA and BPA, glucose uptake was also significantly lower than in the control group (P<0.05), suggesting that BPA can reduce the concentration of PA required to induce insulin resistance (Figure 3F). When the exposure time was reduced to 24 hours, there was no significant change in cell glucose uptake (P>0.05), as shown in Figure 3G. These findings suggest that BPA can reduce the concentration of PA required for exposure, and their combined action induces IR in C2C12 myocytes. Based on the above results, the dosage and exposure time of PA and BPA were determined, using 200 μM PA and 102 nM BPA for 48 hours of exposure to C2C12 myotubes. The results showed that compared to the insulin-stimulated control group, the glucose uptake of the co-exposed group was significantly reduced (P<0.05), as seen in Figure 3-18H. According to the CCK8 cell viability assay results, the cell activity of the groups exposed to 200 μM PA alone and 102 nM BPA alone was above 80%, and after 48 hours of co-exposure to 200 μM PA and 102 nM BPA, cell activity decreased to 73%, with statistically significant differences in cell activity among the groups (P<0.05), as shown in Figure 3-18I.
3.4. Co-Exposure to BPA and Palmitic Acid Inhibits Insulin Signaling and GLUT4 Translocation in C2C12 Cells
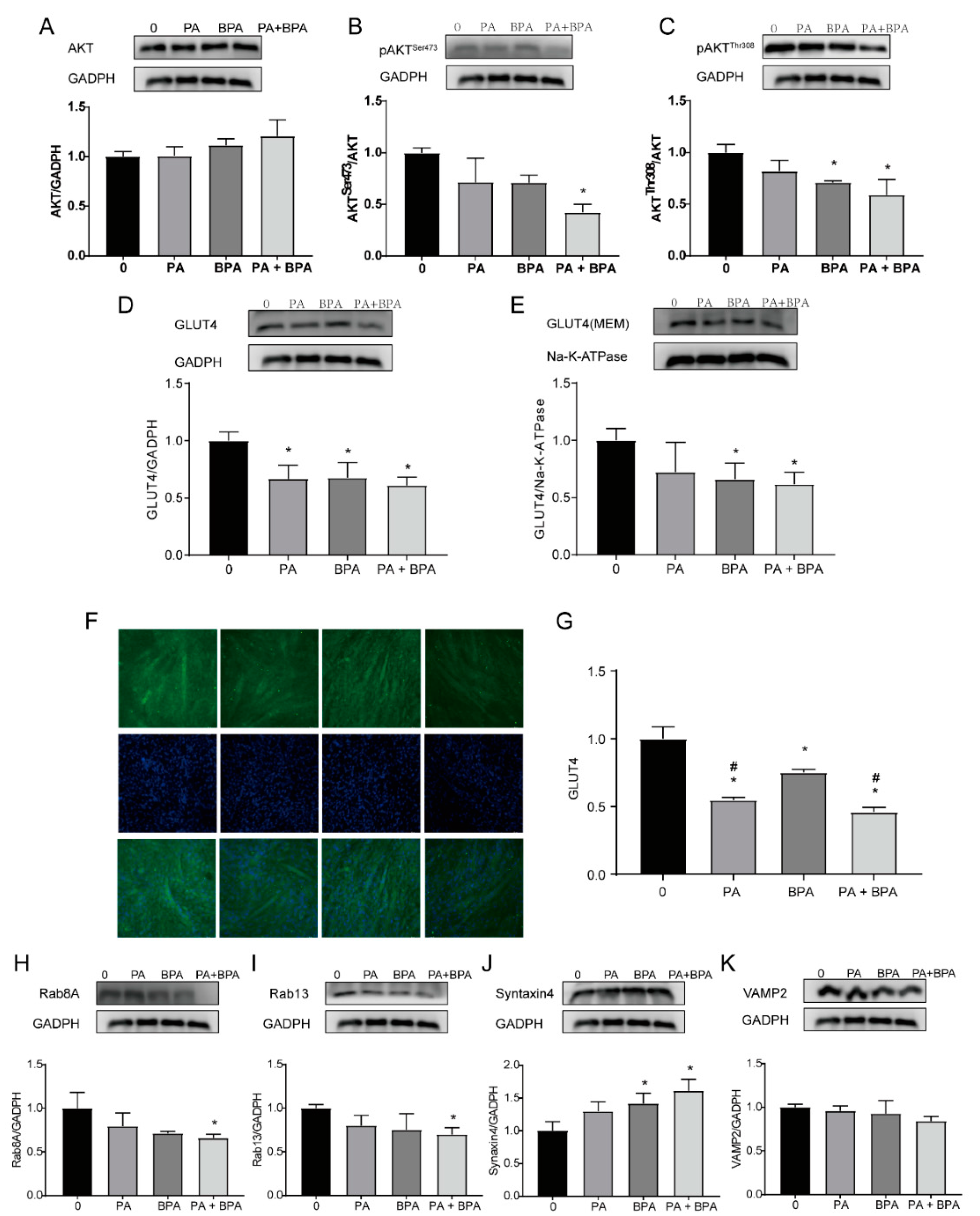
The PI3K/AKT signaling pathway is an important regulatory route for glucose uptake [17,29]. To investigate the potential molecular mechanisms by which BPA and PA affect insulin sensitivity, this study examined the expression levels of AKT and p-AKT, as well as their downstream signaling molecules and proteins related to GLUT4 translocation. The results are shown in Figure 4A-4C. Compared to the control group, the expression levels of p-AKT (Ser473 and Thr308) were significantly reduced in the group co-exposed to 200 μM PA and 102 nM BPA (P<0.01), and the expression level of AKTThr308 in the group exposed to 102 nM BPA alone also significantly decreased (P=0.018).
As shown in Figure 4D, the total GLUT4 expression levels in all experimental groups were significantly lower than those in the control group (P<0.05). After separating the cell membrane using a plasma membrane extraction kit, the results showed that the expression levels of GLUT4 on the cell membrane in the group exposed to 102 nM BPA alone (P<0.05) and in the group co-exposed to 200 μM PA and 102 nM BPA (P<0.05) were significantly lower than those in the control group (as shown in Figure 4E), suggesting that co-exposure to BPA and PA significantly inhibited the expression of GLUT4 on the cell membrane, i.e., reduced the translocation of GLUT4 from the cytoplasm to the plasma membrane. Immunofluorescence staining of C2C12 myotubes showed that the expression levels of GLUT4 in the experimental groups were significantly lower than those in the control group (P<0.05), and compared to the group exposed to 102 nM BPA alone, the expression levels of GLUT4 in the group exposed to 200 μM PA alone (P=0.0026) and in the group co-exposed to 200 μM PA and 102 nM BPA (P=0.0002) were significantly decreased, as seen in Figure 4F-4G.
Downstream molecules of AKT, such as AS160, can activate Rab proteins, and Rab8A and Rab13, along with VAMP2 and Syntaxin4 proteins, can activate GLUT4 vesicles and facilitate their fusion with the plasma membrane to release glucose molecules. In this study, it was found that compared to the control group, the protein expression levels of Rab8A and Rab13 in the group co-exposed to 200 μM PA and 102 nM BPA were significantly reduced (P<0.05). The expression levels of Syntaxin4 protein in the group exposed to 102 nM BPA alone (P=0.0281) and in the group co-exposed to 200 μM PA and 102 nM BPA (P=0.0034) were significantly higher than those in the control group, as shown in Figure 4H-4K.
4. Discussion
With changes in lifestyle and dietary habits, excessive energy intake coupled with insufficient physical activity leading to obesity and its associated insulin resistance pose a serious threat to human health. On the other hand, environmental endocrine disruptors (EDCs), represented by BPA, are produced and used in large quantities; BPA, a monomer and plasticizer, is one of the highest-volume chemicals globally, with an annual production exceeding 100 million pounds [8]. Studies have reported that BPA can be measured in human serum, urine, amniotic fluid, follicular fluid, placental tissue, and umbilical cord [30]. Literature suggests that long-term exposure to BPA and a high-fat diet are both associated with the occurrence of insulin resistance (IR) or type 2 diabetes mellitus (T2DM), yet the combined effects and mechanisms of these two factors remain unclear.
To clarify the relationship and mechanism between the combined effects of a high-fat diet and BPA on animal IR, this study observed the impact of co-exposure to a high-fat diet and low doses of BPA on tissue or cellular GLUT4 expression and insulin resistance. In the animal experiment design, BPA concentrations were selected with reference to the U.S. tolerable daily intake of 50 μg/kg·d and the European proposed tolerable daily intake of 4 μg/kg·d, and combined with literature [31], three doses of BPA were ultimately chosen: 5 μg/kg·d, 50 μg/kg·d, and 500 μg/kg·d. The results showed a significant interaction effect between BPA and high-fat diet treatment on animal body weight and blood glucose levels, and clear abnormalities were observed in the levels of blood lipids and insulin signaling pathway-related molecules.
In vitro cellular experiments involved the induction of C2C12 myoblasts to differentiate into myotubes. 2-NBDG, a fluorescent indicator for direct and rapid measurement of glucose uptake levels, is commonly used as an index of insulin resistance in cellular experiments [32,33]. HOMA-IR is a statistical parameter for assessing insulin resistance and insulin sensitivity using fasting blood glucose levels and fasting serum insulin levels. In this study, the TM and TH groups exhibited insulin resistance and impaired insulin sensitivity, consistent with other similar studies [34]. These results confirm that long-term co-exposure to BPA and a high-fat diet can induce insulin resistance, but the mechanisms are not fully understood, especially in skeletal muscle, an important peripheral tissue for maintaining blood glucose homeostasis and a primary target organ for insulin action. The expression profile of protein molecules and the activation or suppression of the insulin signaling system require further study. Among these, the activation of the glucose transport system (GLUT4) is the first step in glucose metabolism, transporting glucose into muscle cells. Under normal conditions, about 90% of GLUT4 is expressed in the cytoplasm, and only about 10% is expressed on the cell membrane. Promoting the translocation of GLUT4 from the cytoplasm to the cell membrane is an effective way to increase glucose transport, and one of the main factors stimulating GLUT4 translocation in skeletal muscle is insulin, thus regulating the insulin PI3K-AKT signaling pathway can effectively regulate GLUT4 translocation [35,36,37]. The study results showed that the expression level of pAKTSer473 in the gastrocnemius muscle tissue of the experimental animals was lower than that of the control group, and the in vitro experimental results also showed that the expression levels of pAKTSer473 and pAKTThr308 in myotube cells were significantly reduced after exposure to BPA and palmitic acid, suggesting that the combined treatment of high fat and BPA can cause disorders in the insulin signaling pathway, thereby exacerbating the development of insulin resistance [38,39]. In addition, the Rab protein family member Rab8A is a key factor regulated by insulin and involved in the translocation of GLUT4 in muscle cells. The results of this study indicate that compared with the control group, the expression levels of Rab8A in the gastrocnemius muscle cells of animals and cells cultured in vitro were significantly reduced after co-treatment with a high-fat diet and BPA, suggesting that the combined treatment of high fat factors and BPA may affect the expression of Rab8A protein and thus have a certain impact on its function in regulating GLUT4 translocation.
In the GLUT4-mediated glucose transport process, in addition to the translocation of GLUT4 vesicles, the fusion of these vesicles with the plasma membrane is equally crucial for the utilization of glucose by skeletal muscle and the maintenance of blood glucose homeostasis. Specifically, the docking of GLUT4 vesicles with the plasma membrane depends on the interaction between vesicle membrane protein VAMP2 and plasma membrane protein Syntaxin4 [40,41]. Syntaxin4 anchors GLUT4 vesicles on the plasma membrane, while VAMP2 promotes the fusion of the plasma membrane with vesicles, accelerating the process of glucose exocytosis [42]. This study observed that in the gastrocnemius muscle, the expression level of VAMP2 in the group co-treated with a high-fat diet and bisphenol A (BPA) was significantly lower than that in the control group. Furthermore, in C2C12 myotubes, the group co-treated with PA (likely referring to a high-fat diet) and BPA also showed similar results. Particularly in male rats, the TH (likely referring to co-treatment with a high-fat diet and BPA) group exhibited significantly lower VAMP2 expression levels compared to the C0, C1, and C2 groups; in the TM (likely referring to BPA alone treatment) group, the expression level was also significantly lower than in the C0 and C1 groups. This suggests that the combined exposure to BPA and a high-fat diet may significantly inhibit the expression of the VAMP2 protein.
In contrast, the expression level of Syntaxin4 in the PA+BPA group was significantly higher than in the control group, with a similar trend observed in the gastrocnemius muscle, where the upregulation of Syntaxin4 protein expression was most pronounced in the TH group. This upregulation may reflect a physiological compensatory mechanism aimed at counteracting the observed reduction in glucose uptake stimulated by insulin in this study. Therefore, the overexpression of Syntaxin4 protein may be an adaptive response of cells to compensate for defects in GLUT4 translocation [43,44].
Using experimental techniques such as Western Blot, immunohistochemistry, and immunofluorescence, we found that in both gastrocnemius tissue and C2C12 myotubes, the total expression level of GLUT4 in the group co-treated with a high-fat diet and BPA was significantly lower than that in the control group. Immunohistochemical and immunofluorescence experiments further showed that compared to the groups treated with BPA or a high-fat diet alone, the expression level of GLUT4 in the co-treated group was significantly reduced. In addition, the expression level of GLUT4 on the plasma membrane, separated from C2C12 myotubes using a plasma membrane separation kit, confirmed this point, with the PA+BPA group showing significantly reduced GLUT4 expression compared to the control group. These results suggest that in vivo animal experiments, the combined exposure to BPA and a high-fat diet has a more pronounced inhibitory effect on GLUT4 expression, and in vitro cellular experiments further confirmed this finding, indicating that the combined action of BPA and a high-fat diet severely hinders GLUT4 translocation.
The storage capacity of glycogen in skeletal muscle is limited because the activity of glycogen synthase is inhibited, leading to impaired glycogen accumulation [45]. Impaired glycogen synthesis stimulated by insulin is a common pathological feature in all states of insulin resistance. Obese and diabetic patients, as representatives of metabolic syndrome, have severely impaired activity of glycogen synthase stimulated by insulin [46,47,48]. Studies have shown that glycogen synthase kinase 3β (GSK3β) is a protein that can be phosphorylated by AKT and plays a key role in the translocation of GLUT4 from intracellular vesicles to the plasma membrane. When GSK3β is phosphorylated by AKT, it inhibits the activity of glycogen synthase, reduces the synthesis of liver glycogen, thereby causing an increase in blood glucose levels, impairs insulin signaling, and ultimately leads to insulin resistance [49]. The reduction in pGSK3βSer9 levels may be associated with impaired GLUT4 translocation [[50]. This study observed that in male rats, the expression level of GSK3β protein in the BPA-only treated group (TM group) was significantly higher than that in the high-fat diet-only treated group (C2 group), and no significant difference in GSK3β protein expression levels was observed between other groups. This suggests that a high-fat diet may exacerbate the expression of GSK3β in the gastrocnemius muscle after BPA exposure. Additionally, the expression level of pGSK3βSer9 in the group co-treated with a high-fat diet and BPA was significantly lower than that in the control group, especially in the TM and TH groups of female rats, where pGSK3βSer9 levels were significantly lower than in the high-fat diet-only treated group (C1 group). These results suggest that BPA may exacerbate the reduction of pGSK3βSer9 in the gastrocnemius muscle after exposure to a high-fat diet.
Overall, the expression levels of GSK3β and pGSK3βSer9 in the gastrocnemius muscle may be influenced by different factors, with a high-fat diet having a dominant effect on the expression of GSK3β in male rats, and BPA having a dominant effect on the expression of pGSK3βSer9 in female rats. Therefore, the results of this study indicate that the combined action of BPA and a high-fat diet may more severely damage insulin signaling, reduce the expression level of pGSK3βSer9, and thus affect GLUT4 translocation.
5. Conclusions
In summary, the co-exposure to BPA and a high-fat diet may significantly affect the translocation process in the gastrocnemius muscle and C2C12 myotubes of adult mice, mainly by disrupting the insulin signaling pathway mediated by AKT. This disruption leads to a series of changes in protein expression, especially those related to the translocation of GLUT4 from the cytoplasm to the plasma membrane. These changes exacerbate the occurrence of insulin resistance and inhibit the translocation of GLUT4, which may adversely affect blood glucose regulation and metabolic health.
Author Contributions
Conceptualization, W.Z.; methodology, ZQ.L.,WX.L.JY.W.GY.Y.and B.H.;validation, WX.L.,M.C.and JT.L.; formal analysis,B.H.and Y.Z; funding acquisition, W.Z. All authors have read and agreed to the published version of the manuscript.”
Funding
This research was funded by the National Key Research and Development Program of China(2018YFC1603102), Basic Research Project of Key Laboratory of Guangzhou(2024A03J0558) and the Key Project of Medicine Discipline of Guangzhou(No.20212023012).
Institutional Review Board Statement
Informed consent was obtained from all subjects involved in the study
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Huang, Y.; Karuranga, S.; Malanda, B.; Williams, D. Call for data contribution to the IDF Diabetes Atlas 9th Edition 2019. Diabetes Res. Clin. Pr. 2018, 140, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Al Mansour, M.A. The Prevalence and Risk Factors of Type 2 Diabetes Mellitus (DMT2) in a Semi-Urban Saudi Population. Int. J. Environ. Res. Public Heal. 2019, 17, 7. [Google Scholar] [CrossRef] [PubMed]
- Woods, S.C.; Seeley, R.J.; Rushing, P.A.; D'Alessio, D.; Tso, P. A Controlled High-Fat Diet Induces an Obese Syndrome in Rats. J. Nutr. 2003, 133, 1081–1087. [Google Scholar] [CrossRef] [PubMed]
- KAHN S E, HULL R L, UTZSCHNEIDER K M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–6. [CrossRef]
- Small, L.; Brandon, A.E.; Turner, N.; Cooney, G.J. Modeling insulin resistance in rodents by alterations in diet: what have high-fat and high-calorie diets revealed? Am. J. Physiol. Metab. 2018, 314, E251–E265. [Google Scholar] [CrossRef]
- Hinault, C.; Caroli-Bosc, P.; Bost, F.; Chevalier, N. Critical Overview on Endocrine Disruptors in Diabetes Mellitus. Int. J. Mol. Sci. 2023, 24, 4537. [Google Scholar] [CrossRef]
- Weiss, M.C.; Shih, Y.-H.; Bryan, M.S.; Jackson, B.P.; Aguilar, D.; Hanis, C.L.; Argos, M.; Sargis, R.M. Relationships Between Urinary Metals and Diabetes Traits Among Mexican Americans in Starr County, Texas, USA. Biol. Trace Element Res. 2022, 201, 529–538. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Geens, T.; Goeyens, L.; Covaci, A. Are potential sources for human exposure to bisphenol-A overlooked? Int. J. Hyg. Environ. Heal. 2011, 214, 339–347. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Chahoud, I.; Heindel, J.J.; Padmanabhan, V.; Paumgartten, F.J.; Schoenfelder, G. Urinary, Circulating, and Tissue Biomonitoring Studies Indicate Widespread Exposure to Bisphenol A. Cienc. Saude Coletiva 2012, 17, 407–434. [Google Scholar] [CrossRef]
- RUSSO G, BARBATO F, MITA D G, et al. Occurrence of Bisphenol A and its analogues in some foodstuff marketed in Europe. Food and chemical toxicology : an international journal published for the British Industrial Biological Research Association 2019, 131, 110575. [CrossRef] [PubMed]
- Hafezi, S.A.; Abdel-Rahman, W.M. The Endocrine Disruptor Bisphenol A (BPA) Exerts a Wide Range of Effects in Carcinogenesis and Response to Therapy. Curr. Mol. Pharmacol. 2019, 12, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Lee, K. Usefulness of the metabolic syndrome criteria as predictors of insulin resistance among obese Korean women. Public Health Nutr 2009, 13, 181–186. [Google Scholar] [CrossRef] [PubMed]
- FREEMAN A M, ACEVEDO L A, PENNINGS N. Insulin Resistance [M]. StatPearls. Treasure Island (FL) ineligible companies. Disclosure: Luis Acevedo declares no relevant financial relationships with ineligible companies. Disclosure: Nicholas Pennings declares no relevant financial relationships with ineligible companies.; StatPearls Publishing.
- Copyright © 2024, StatPearls Publishing LLC. 2024.
- Ferrannini, E.; Simonson, D.C.; Katz, L.D.; Reichard, G.; Bevilacqua, S.; Barrett, E.J.; Olsson, M.; DeFronzo, R.A. The disposal of an oral glucose load in patients with non-insulin-dependent diabetes. Metabolism 1988, 37, 79–85. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Tripathy, D. Skeletal Muscle Insulin Resistance Is the Primary Defect in Type 2 Diabetes. Diabetes Care 2009, 32 (Suppl. 2), S157–S163. [Google Scholar] [CrossRef]
- Zhu, Y.; Pereira, R.O.; O'Neill, B.T.; Riehle, C.; Ilkun, O.; Wende, A.R.; Rawlings, T.A.; Zhang, Y.C.; Zhang, Q.; Klip, A.; et al. Cardiac PI3K-Akt Impairs Insulin-Stimulated Glucose Uptake Independent of mTORC1 and GLUT4 Translocation. Mol. Endocrinol. 2013, 27, 172–184. [Google Scholar] [CrossRef]
- SHARMA B R, KIM H J, RHYU D Y. Caulerpa lentillifera extract ameliorates insulin resistance and regulates glucose metabolism in C57BL/KsJ-db/db mice via PI3K/AKT signaling pathway in myocytes. Journal of translational medicine 2015, 13, 62. [CrossRef]
- LI W, LIANG X, ZENG Z, et al. Simvastatin inhibits glucose uptake activity and GLUT4 translocation through suppression of the IR/IRS-1/Akt signaling in C2C12 myotubes. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie 2016, 83, 194–200.
- Sun, Y.; Bilan, P.J.; Liu, Z.; Klip, A. Rab8A and Rab13 are activated by insulin and regulate GLUT4 translocation in muscle cells. Proc. Natl. Acad. Sci. USA 2010, 107, 19909–19914. [Google Scholar] [CrossRef]
- Mullainadhan, V.; Viswanathan, M.P.; Karundevi, B. Effect of Bisphenol-A (BPA) on insulin signal transduction and GLUT4 translocation in gastrocnemius muscle of adult male albino rat. Int. J. Biochem. Cell Biol. 2017, 90, 38–47. [Google Scholar] [CrossRef]
- FOSTER L J, KLIP A. Mechanism and regulation of GLUT-4 vesicle fusion in muscle and fat cells. American journal of physiology Cell physiology 2000, 279, C877–90. [CrossRef] [PubMed]
- Duman, J.G.; Forte, J.G. What is the role of SNARE proteins in membrane fusion? Am. J. Physiol. Cell Physiol. 2003, 285, C237–C249. [Google Scholar] [CrossRef] [PubMed]
- Pessin, J.E.; Thurmond, D.C.; Elmendorf, J.S.; Coker, K.J.; Okada, S. Molecular Basis of Insulin-stimulated GLUT4 Vesicle Trafficking. Location! Location! Location! J. Biol. Chem. 1999, 274, 2593–2596. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, L.H.; Gould, G.W. The Vesicle- and Target-SNARE Proteins That Mediate Glut4 Vesicle Fusion Are Localized in Detergent-insoluble Lipid Rafts Present on Distinct Intracellular Membranes. J. Biol. Chem. 2002, 277, 49750–49754. [Google Scholar] [CrossRef]
- Garvey, W.T.; Maianu, L.; Zhu, J.H.; Brechtel-Hook, G.; Wallace, P.; Baron, A.D. Evidence for defects in the trafficking and translocation of GLUT4 glucose transporters in skeletal muscle as a cause of human insulin resistance. J. Clin. Investig. 1998, 101, 2377–2386. [Google Scholar] [CrossRef]
- HU F, B. Globalization of diabetes: the role of diet, lifestyle, and genes. Diabetes care, 2011, 34, 1249–57. [Google Scholar] [CrossRef]
- Tahapary, D.L.; Pratisthita, L.B.; Fitri, N.A.; Marcella, C.; Wafa, S.; Kurniawan, F.; Rizka, A.; Tarigan, T.J.E.; Harbuwono, D.S.; Purnamasari, D.; et al. Challenges in the diagnosis of insulin resistance: Focusing on the role of HOMA-IR and Tryglyceride/glucose index. Diabetes Metab. Syndr. Clin. Res. Rev. 2022, 16, 102581. [Google Scholar] [CrossRef]
- Miyata, S.; Yada, T.; Ishikawa, N.; Taheruzzaman, K.; Hara, R.; Matsuzaki, T.; Nishikawa, A. Insulin-like growth factor 1 regulation of proliferation and differentiation of Xenopus laevis myogenic cells in vitro. Vitr. Cell. Dev. Biol. - Anim. 2016, 53, 231–247. [Google Scholar] [CrossRef]
- Chen, D.; Kannan, K.; Tan, H.; Zheng, Z.; Feng, Y.-L.; Wu, Y.; Widelka, M. Bisphenol Analogues Other Than BPA: Environmental Occurrence, Human Exposure, and Toxicity—A Review. Environ. Sci. Technol. 2016, 50, 5438–5453. [Google Scholar] [CrossRef]
- Marmugi, A.; Ducheix, S.; Lasserre, F.; Polizzi, A.; Paris, A.; Priymenko, N.; Bertrand-Michel, J.; Pineau, T.; Guillou, H.; Martin, P.G.P.; et al. Low doses of bisphenol a induce gene expression related to lipid synthesis and trigger triglyceride accumulation in adult mouse liver. Hepatology 2012, 55, 395–407. [Google Scholar] [CrossRef]
- Zou, C.; Wang, Y.; Shen, Z. 2-NBDG as a fluorescent indicator for direct glucose uptake measurement. J. Biochem. Biophys. Methods 2005, 64, 207–215. [Google Scholar] [CrossRef]
- Luo, W.; Ai, L.; Wang, B.-F.; Zhou, Y. High glucose inhibits myogenesis and induces insulin resistance by down-regulating AKT signaling. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie 2019, 120, 109498. [Google Scholar] [CrossRef]
- Ding, S.; Fan, Y.; Zhao, N.; Yang, H.; Ye, X.; He, D.; Jin, X.; Liu, J.; Tian, C.; Li, H.; et al. High-fat diet aggravates glucose homeostasis disorder caused by chronic exposure to bisphenol A. J. Endocrinol. 2014, 221, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Mora, S.; Pessin, J.E. An adipocentric view of signaling and intracellular trafficking. Diabetes/Metabolism Res. Rev. 2002, 18, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Fryer, L.G.D.; Foufelle, F.; Barnes, K.; Baldwin, S.A.; Woods, A.; Carling, D. Characterization of the role of the AMP-activated protein kinase in the stimulation of glucose transport in skeletal muscle cells. Biochem. J. 2002, 363, 167–74. [Google Scholar] [CrossRef]
- HAYASHI T, HIRSHMAN M F, KURTH E J, et al. Evidence for 5' AMP-activated protein kinase mediation of the effect of muscle contraction on glucose transport. Diabetes, 1998, 47, 1369–73.
- Kidani, T.; Kamei, S.; Miyawaki, J.; Aizawa, J.; Sakayama, K.; Masuno, H. Bisphenol A Downregulates Akt Signaling and Inhibits Adiponectin Production and Secretion in 3T3-L1 Adipocytes. J. Atheroscler. Thromb. 2010, 17, 834–843. [Google Scholar] [CrossRef]
- Ropero, A.B.; Soria, B.; Nadal, A. A Nonclassical Estrogen Membrane Receptor Triggers Rapid Differential Actions in the Endocrine Pancreas. Mol. Endocrinol. 2002, 16, 497–505. [Google Scholar] [CrossRef]
- Bryant, N.J.; Gould, G.W. SNARE Proteins Underpin Insulin-Regulated GLUT4 Traffic. Traffic 2011, 12, 657–664. [Google Scholar] [CrossRef]
- Jaldin-Fincati, J.R.; Pavarotti, M.; Frendo-Cumbo, S.; Bilan, P.J.; Klip, A. Update on GLUT4 Vesicle Traffic: A Cornerstone of Insulin Action. Trends Endocrinol. Metab. 2017, 28, 597–611. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Tamori, Y.; Kanda, H.; Yoshikawa, M.; Tateya, S.; Nishino, N.; Kasuga, M. The t-SNAREs syntaxin4 and SNAP23 but not v-SNARE VAMP2 are indispensable to tether GLUT4 vesicles at the plasma membrane in adipocyte. Biochem. Biophys. Res. Commun. 2010, 391, 1336–1341. [Google Scholar] [CrossRef] [PubMed]
- KAHN B B, FLIER J S. Regulation of glucose-transporter gene expression in vitro and in vivo. Diabetes care, 1990, 13, 548–64.
- Foster, L.J.; Yaworsky, K.; Trimble, W.S.; Klip, A. SNAP23 promotes insulin-dependent glucose uptake in 3T3-L1 adipocytes: possible interaction with cytoskeleton. Am. J. Physiol. Physiol. 1999, 276, C1108–C1114. [Google Scholar] [CrossRef]
- Cohen, P.; Goedert, M. GSK3 inhibitors: development and therapeutic potential. Nat. Rev. Drug Discov. 2004, 3, 479–487. [Google Scholar] [CrossRef]
- GULLI G, FERRANNINI E, STERN M, et al. The metabolic profile of NIDDM is fully established in glucose-tolerant offspring of two Mexican-American NIDDM parents. Diabetes, 1992, 41, 1575–86.
- COHEN, P. The Croonian Lecture 1998. Identification of a protein kinase cascade of major importance in insulin signal transduction. Philosophical transactions of the Royal Society of London Series B, Biological sciences, 1999, 354, 485–95. [Google Scholar]
- WILLIAMS K V, PRICE J C, KELLEY D E. Interactions of impaired glucose transport and phosphorylation in skeletal muscle insulin resistance: a dose-response assessment using positron emission tomography. Diabetes, 2001, 50, 2069–79.
- Carnagarin, R.; Dharmarajan, A.M.; Dass, C.R. Molecular aspects of glucose homeostasis in skeletal muscle – A focus on the molecular mechanisms of insulin resistance. Mol. Cell. Endocrinol. 2015, 417, 52–62. [Google Scholar] [CrossRef]
- Sanvee, G.M.; Panajatovic, M.V.; Bouitbir, J.; Krähenbühl, S. Mechanisms of insulin resistance by simvastatin in C2C12 myotubes and in mouse skeletal muscle. Biochem. Pharmacol. 2019, 164, 23–33. [Google Scholar] [CrossRef]
Figure 2.
Effects of BPA and a high-fat diet on the expression levels of signaling molecules in gastrocnemius tissue of male and female mice. Protein expression levels of AKT (A and B), pAKTSer473 (C), Rab8A (E), VAMP2 (F), Syntaxin4 (G), GLUT4 (H), GSK3β (L), and pGSK3βSer9 (M); results of GLUT4 immunohistochemistry (I); and quantitative analysis of immunohistochemistry results (J). Compared to the C0 group, * P<0.05; compared to the C1 group, # P<0.05; compared to the C2 group, △ P<0.05.
Figure 2.
Effects of BPA and a high-fat diet on the expression levels of signaling molecules in gastrocnemius tissue of male and female mice. Protein expression levels of AKT (A and B), pAKTSer473 (C), Rab8A (E), VAMP2 (F), Syntaxin4 (G), GLUT4 (H), GSK3β (L), and pGSK3βSer9 (M); results of GLUT4 immunohistochemistry (I); and quantitative analysis of immunohistochemistry results (J). Compared to the C0 group, * P<0.05; compared to the C1 group, # P<0.05; compared to the C2 group, △ P<0.05.
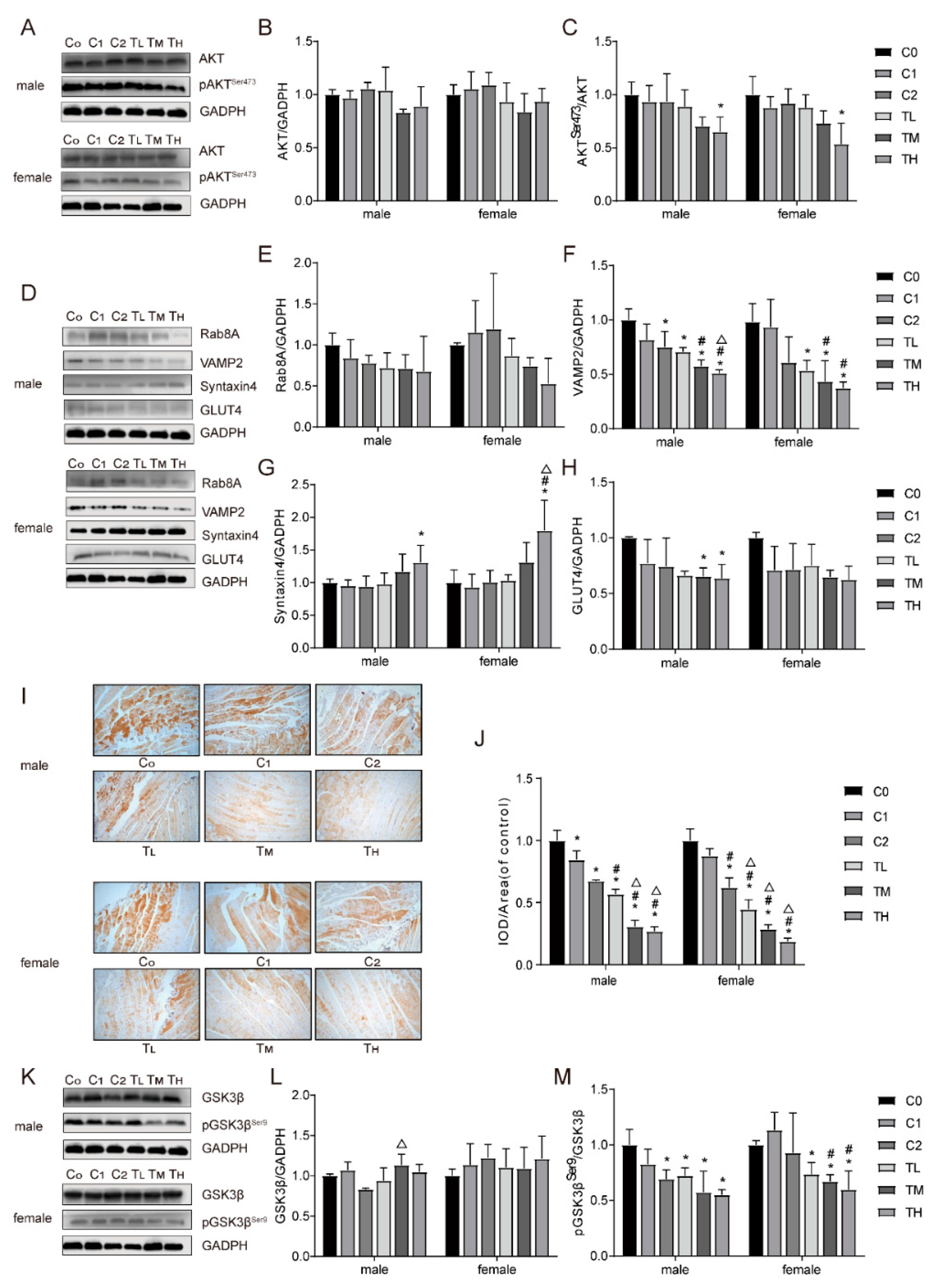
Figure 3.
Effects of 48-hour BPA and palmitic acid (PA) treatment on glucose uptake in C2C12 myotubes. C2C12 myotubes were exposed to BPA (1-105 nM, control group with 0 nM BPA; 0.1% DMSO) and PA (100-700 μM, control group with 0 μM PA; 2% BSA) for 48 hours to measure cytotoxicity with the CCK8 assay (A, B); the effect of 48-hour BPA exposure on glucose uptake (C); the effect of 48-hour PA exposure on glucose uptake (D); the effect of co-exposure to BPA (102 and 103 nM) and PA (300 μM) for 48 hours on glucose uptake (E); the effect of co-exposure to BPA (102 and 103 nM) and PA (200 μM) for 48 hours on glucose uptake (F); the effect of co-exposure to BPA (102 and 103 nM) and PA (200 μM) for 24 hours on glucose uptake (G); the effect of co-exposure to BPA (102 nM) and PA (200 μM) for 48 hours on glucose uptake (H); results are presented as mean ± SD, n = 3. Compared to the insulin-stimulated control group, * P<0.05; compared to the corresponding concentration group without insulin stimulation, # P<0.05. Cell viability after co-exposure to BPA (102 nM) and PA (200 μM) for 48 hours (I). Compared to the control group, & P<0.05; compared to the 200 μM PA group, $ P<0.05; compared to the 102 nM BPA group, P<0.05.
Figure 3.
Effects of 48-hour BPA and palmitic acid (PA) treatment on glucose uptake in C2C12 myotubes. C2C12 myotubes were exposed to BPA (1-105 nM, control group with 0 nM BPA; 0.1% DMSO) and PA (100-700 μM, control group with 0 μM PA; 2% BSA) for 48 hours to measure cytotoxicity with the CCK8 assay (A, B); the effect of 48-hour BPA exposure on glucose uptake (C); the effect of 48-hour PA exposure on glucose uptake (D); the effect of co-exposure to BPA (102 and 103 nM) and PA (300 μM) for 48 hours on glucose uptake (E); the effect of co-exposure to BPA (102 and 103 nM) and PA (200 μM) for 48 hours on glucose uptake (F); the effect of co-exposure to BPA (102 and 103 nM) and PA (200 μM) for 24 hours on glucose uptake (G); the effect of co-exposure to BPA (102 nM) and PA (200 μM) for 48 hours on glucose uptake (H); results are presented as mean ± SD, n = 3. Compared to the insulin-stimulated control group, * P<0.05; compared to the corresponding concentration group without insulin stimulation, # P<0.05. Cell viability after co-exposure to BPA (102 nM) and PA (200 μM) for 48 hours (I). Compared to the control group, & P<0.05; compared to the 200 μM PA group, $ P<0.05; compared to the 102 nM BPA group, P<0.05.
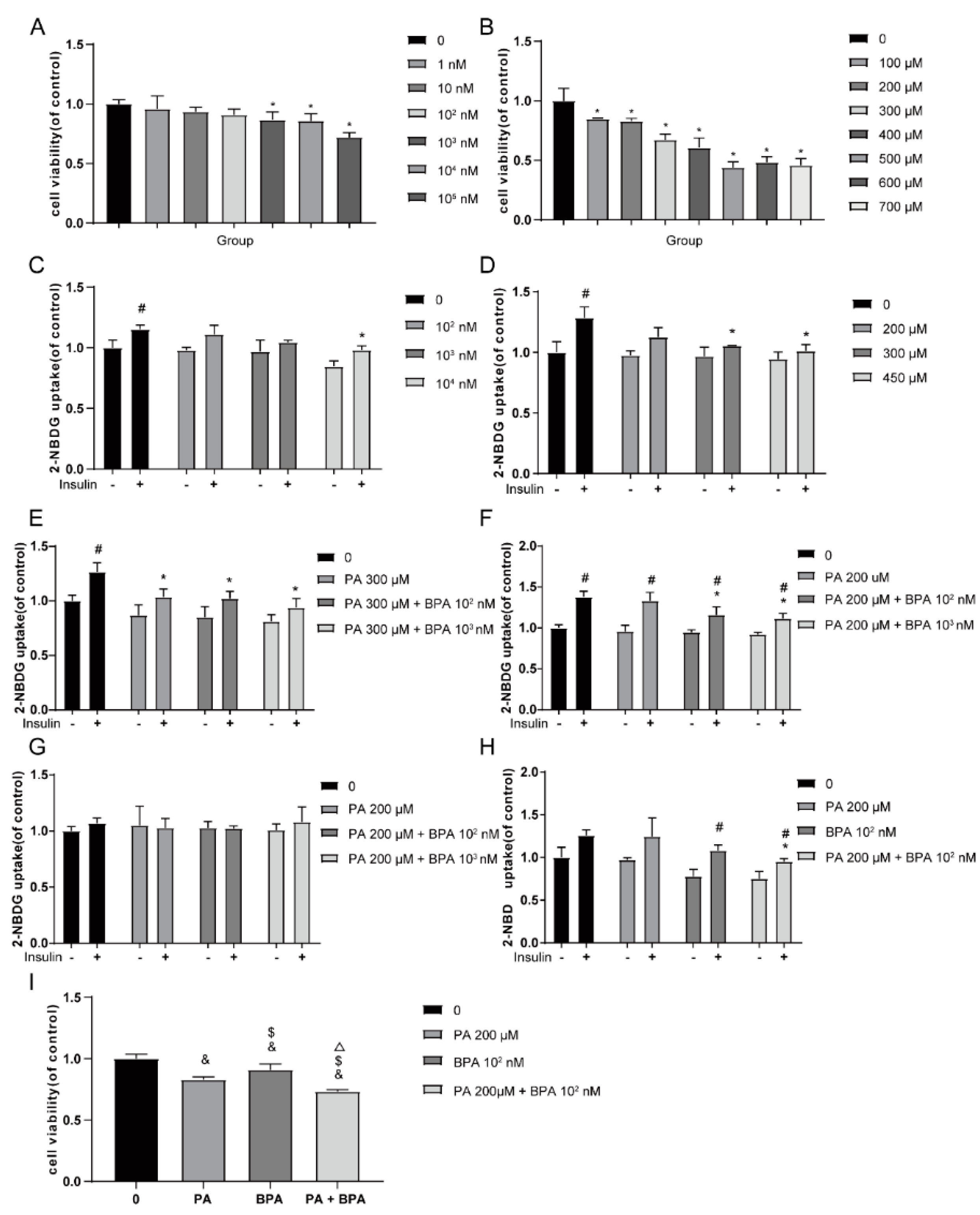
Figure 4.
Effects of BPA and palmitic acid on signaling molecules and GLUT4 translocation in C2C12 myotubes. Protein expression levels of AKT (A), pAKTSer473 (B), pAKTThr308 (C), total GLUT4 (D), GLUT4 on the membrane (E), Rab8A (H), Rab13 (I), Syntaxin4 (J), and VAMP2 (K); immunofluorescence staining with an anti-GLUT4 antibody after treating C2C12 cells with BPA and palmitic acid for 48 hours, scale bar: 100 μm (F); quantitative analysis of GLUT4 immunofluorescence (G). Compared to the control group, * P<0.05; compared to the 102 nM BPA group, # P<0.05.
Figure 4.
Effects of BPA and palmitic acid on signaling molecules and GLUT4 translocation in C2C12 myotubes. Protein expression levels of AKT (A), pAKTSer473 (B), pAKTThr308 (C), total GLUT4 (D), GLUT4 on the membrane (E), Rab8A (H), Rab13 (I), Syntaxin4 (J), and VAMP2 (K); immunofluorescence staining with an anti-GLUT4 antibody after treating C2C12 cells with BPA and palmitic acid for 48 hours, scale bar: 100 μm (F); quantitative analysis of GLUT4 immunofluorescence (G). Compared to the control group, * P<0.05; compared to the 102 nM BPA group, # P<0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



