
Submitted:
12 February 2025
Posted:
13 February 2025
You are already at the latest version
Abstract
The dendroflora of an urban arboretum, consisting of 193 species, was ecologically characterized as bioforms, phytogeographical elements, and preferences for moisture, temperature, and soil reaction. The inventoried species are grouped in 111 genera and 45 families. The native and non-native dendroflora share 16 common families. The most representative family both in the native and non-native dendroflora is Rosaceae. The monotypic families are largely present (22.22% in the native dendroflora, and respectively 42.22% in the non-native dendroflora). The plant life-form spectrum is dominated by megaphanerophytes (49%), followed by mesophanerophytes (41%). The chorological spectrum of the native species comprises 16 chorological types and is dominated by Eurasians (32%) and Europeans (30%). The species characteristics of the Pontic-Carpathian space, to which Romania belongs, are rare in the analyzed urban park (4%). The mesophyte, mesothermal and slightly acido-neutrophilous species dominate both the native and non-native dendroflora. In the adaption process of the non-native dendroflora, 37% of species exceeded their native requirements for moisture, 41% for temperature, and 50% for soil reaction. The species requirements for temperature are associated to those for moisture and soil reaction. The results show the potential of the analysed woody species to exceed their native requirements within the acclimation and adaption process, and in this process, for the studied temperate site, the plant life-form is important, and also the plant family. The species of the analyzed urban arboretum, both native and non-native, are taxonomically and biogeographically diverse, with specific habitat requirements, suggesting their great ability in acclimating, adapting and resisting.
Keywords:
bioform
; chorology
; phytogeographic element
; geoelement
; moisture
; temperature
; soil pH
Introduction
The preoccupation of studying the urban green spaces has increased in the last decades since the scientist forecasts announce the expansion of urban settlements as the main living areas of the human population. Among the diversity of the approached subjects, the ecosystem services provided by the urban green spaces are by large interest (improvement of the quality of the environmental factors, sanogenesis, carbon sequestration, climatic buffer, aesthetic benefits). The biodiversity services brought by the urban green spaces are of interest for humanity because its survival is linked to the urban sustainability as safe and reliable living environment. The new approaches regarding the design and the management of the urban green spaces are in present established, or these should be, considering the biodiversity strategy, enhancing the role of the urban green spaces as biodiversity hotspots in the urban environment. In the urban planning of the green spaces, the species composition is a factor in rising up the potential of these areas to fulfill their roles in the sustainability of the urban environment. But, most of the time, when urban parks are established, other more pragmatic factors take precedence, such as the human investment for their long-term maintenance or the receptiveness and satisfaction of citizens with the design offered by the selected species. The settlement of the urban tree parks is an option for keeping the sustainability of cities for several well known important reasons such as higher resistance to pollution stress, efficient rates of carbon sequestration, but also for other less mediated roles like seasonal indicators due to their successive life cycles, urban identity providers (Dümpelmann 2024), or drivers of the biogeochemical cycles. The fulfillment of the ecosystem services by the urban green spaces represents the motivation for establishing these parks, and the efficiency of this objective is reflected in the selection of plant categories used for their settlement, so that the ratio between woody plants and herbaceous plants becomes important, as woody plants offer several advantages over herbaceous ones when the efficiency of ecosystem services provided by the urban green spaces are analyzed. The tree size and implicitly the biomass amount, both the underground and the above ground one, the foliar surface of the canopy represent advantages regarding the quantity of the sequestereted carbon or drought resistance (Simovic et al. 2024). In the temperate areas, the establishment of the urban green spaces with tree-dominated species composition, exploits a proven tree strategy (Niu et al. 2022) to develop height-mediated hydraulic mechanisms to fight against the freeze-thaw stress and sustain their long-term survival despite unfavorable periods. The long life span of the trees as compared to the herbaceous plants is an opportunity for a long time carbon sequestration through the above ground biomass (Lahoti et al. 2020), regulating thus the local carbon cycle and acting like long lasting carbon sinks. The herbaceous plants are more vulnerable to the urban stress factors which, associated to their short life cycles make them less desirable for reaching environmental gains in the urban areas. Previous studies showed that, the short life cycle of the annual weeds in several urban green spaces in addition to moisture deficit and drought climate favored their presence only in the favorable seasons (Heneidy et al. 2021), which make them unreliable on long time for the urban environmental sustainability. The resilience of the urban ecosystems to the various environmental threats (droughts, air pollution, freeze, soil leaching, floods) is enhanced and supported by the urban woody vegetation and is increasing with the species diversity (Hirons et al. 2021). The ecosystem services provided by the urban green spaces are the main goal of the urban sustainability and for its achievement the ratio between the woody and herbaceous plants in the urban parks is considered. The urban green spaces remain the main connection between the continuously growing urbanization areas and the natural environment, and their establishment represents a mean of mitigating specific effects of urbanization like natural habitat disruption, soil sealing and defective infiltration precipitations, species erosion.
The urbanization phenomenon is responsible for certain ecological disturbances in the urban biocoenoses, such as the biological invasion (Horvat et al. 2024) or decrease of species richness in cities. A study on urban remnant patches of forest after urban surrounding showed that the intensity of urban expansion is a factor which decreases the woody species richness in this type of urban green space (Yang et al. 2024). Therefore, the species selection in establishing of the urban green spaces remains the suitable option to increase the urban diversity and to meet its goals in providing urban resilience and sustainability.
The climate changes faced by human-inhabited urban environments need, in order to be countered, urban management measures that include green infrastructure strategies. The warnings of scientists regarding the phenomenon of global warming, fully felt in cities, should determine a proactive approach towards the measures that could be taken to reduce this phenomenon, and in this sense, the presence of trees in cities, in the form of arboretums or as street alignments, could be an exploitable solution.
Materials and Methods
Research Site
The research was conducted in the Botanic Park of Timișoara, Timiș County, Romania (45°45'18''N, 21°13'28''E) (Figure 1), located in the north of the Bega River, which flows through the city. The Botanic Park of Timișoara has been established between years 1986-1990, by the Romanian architect Silvia Grumeza, under the name Botanical Garden of Timișoara, and it was opened to visitors on June 29, 1986 (Ciupa 2018). From the beginning, it was designed as an arboretum (dendrological park). Initially, over 1,650 plant species with diverse origins were planted here (Ciupa 2018). The park covers an area of 8.41 hectares (Ciupa 2010). Since 1986, when the Botanic Park of Timișoara has been established, the human intervention at soil level is mainly for the care of the few herbaceous decorative plants, therefore nowadays the soil park is considered a semi-natural soil. Since 1995, by County Council Decision no. 19/23.02.1995, the Botanic Park was declared a protected natural area - for the conservation of biodiversity, the gene pool, the ecological reserve, and to maintain the ecological balance in Timiș County, with Timișoara City Hall designated as its manager. The Botanic Park of Timișoara is a protected area with two main objectives: (1) the conservation and development of the dendrological collection and (2) the conservation and enhancement of landscapes, with the possibility of being visited for scientific, touristic, educational, and recreational-social purposes (Ciupa 2018).
Research Methodology
The inventory of the woody plant species present in the Botanic Park of Timișoara have been provided upon request by the Timișoara City Hall – Office of Recreational Green Spaces, the authority who sourced the work ”Local Register of Green Spaces – Timișoara Municipality” (76). Totally, the dendrofloristic list comprised 193 species (73 species are native to Romania and 120 species are alien cultural species). All species have been described using the working methodology focused on three objectives:
1. Establishing the categories of plant life-forms (bioforms, biological forms) specific to the studied dendroflora according to C. Raunkiaer's classification (Raunkiaer 1934). The plant life-forms are the expression of the convergent evolution of different species, which gives them similar morphological, structural, and physiological characteristics (Sârbu et al. 2003). The method has been chosen for the purpose of the present research because the delineation of bioforms in plant ecology is based on grouping species by their survival strategies during periods with critical ecological factors, regardless of their taxonomic affiliation. The most widely accepted classification is that of C. Raunkiaer (1934), which is primarily based on how plant’s regenerative structures are protected during the unfavorable season, specifically the position of the renewal organs (buds). Thus, in the delineation of plant life forms (bioforms), the key factor is the level (relative to the soil surface) at which the tissues that ensure the plant’s perennity are found.
2. Classification of plant species into categories of phytogeographical elements (geoelements, chorology) according to a methodology provided by several Romanian authorities in the field (Cristea et al. 2004; Sanda et al. 1983; Sanda et al. 2003; Ciocârlan 2000). The geoelements serve to designate categories of plant species that are more or less distantly related phylogenetically, which, during the process of speciation, have occupied the same geographical region, and then followed specific migration paths and coenotic integration towards the formation of their current distribution ranges (Cristea et al. 2004; Sanda et al. 1983; Sanda et al. 2003; Ciocârlan 2000).
3. Identifying the ecological requirements of the studied plant species for factors such as moisture, temperature, and soil reaction (pH) according to the methodology proposed by Sanda et al. (1983). This methodology is in the sense of Ellenberg scale of plant preferences for the same ecological factors (Ellenberg 1952) and consists of attributing an ecological preference index and respectively an ecological significance description for each species depending on its preferences for moisture, temperature, and soil reaction (Table 1).
For the spontaneous dendroflora there was analyzed the chorological spectrum which shows the geographic distribution of the plant species, but this characteristic has been dropped for the non-native dendroflora because of the great heterogeneity of this group and the difficulty (limited accessible resources) in establishing the phytogeographic element for each non-native species with accuracy, which could affect the reliability of the study. Instead this, there was chosen to be analyzed the acclimation of the non-native species to the local environmental conditions only by studying other characteristics of them such as the habitat requirements for moisture, temperature and soil reaction, to conclude about the adabtability of the non-native species to the host urban ecosystem.
Results and Discussion
The dendroflora of the Botanic Park of Timișoara consists of 193 species: 73 species are native to Romania (Table 2) and 120 species are non-native species (Table 3), grouped in 111 genera and 45 families. This species richness is high and comparable to other urban woody parks in the world (Muhlisin et al. 2021; Bartoli et al. 2022), but the species richness in the urban parks should be assessed also depending on the site size, its geographical characteristics or management preferences (Nero et al. 2024). Predominant are the non-native species which represent 62,17% from the total dendroflora and the angiosperms which represent 84.46% as compared with the gymnosperms (15.54%). Seven native species belong to two gymnosperm families (Pinaceae and Taxaceae), and 23 non-native species belong to five gymnosperm families (Cupressaceae, Ginkgoaceae, Pinaceae, Taxodiaceae, and Taxaceae) (Table 2 and Table 3).
The dominance of the non-native dendroflora in the studied park is consistent with other findings regarding the urban dendroflora from abroad, showing the general tendency in the urban green areas to be dominated by the presence of the alien or exotic species (Fonseca et al. 2024; Muvengwi et al. 2024), as a consequence of the aesthetic and ornamental motivations or due to the lack of scientifically based planning among authority decisional factors since the recommendations indicate the settlement of the urban arboreta based on the native species which are more susceptible to comply with the local ecological criteria (Jang and Woo 2022) to avoid the ecological unbalances (such as alien species invasion with all its associated consequences) and other problems related to population health (like allergenic potential of the pollen). However, the non-native species contribute more to the phylogenetic diversity in the urban green spaces (Muvengwi et al. 2024). But the native tree species are nowadays more encouraged to be part of the urban dendroflora because the native species have a better plasticity in facing the local environmental stressors, a better long-time resilience, and a better ability to support the rest of the local native biodiversity (Galfrascoli et al. 2023). The native and alien dendroflora of the studied arboretum share 16 common families. The most representative family both in the native and alien dendroflora is Rosaceae, which is represented by 14 species (19%) in the native dendroflora and respectively by 20 species (10%) in the non-native dendroflora. The family Rosaceae has been also found to be the most representative in other studies regarding the biodiversity and ecological succession of the urban green spaces (Rogovskyi et al. 2023; Postarnak and Zhavoronkov 2023). The predominance of the family Rosaceae was found by Lakicevic et al. (2022) to be a characteristic of the dendroflora from the temperate climatic regions, where Romania belongs. This aspect has been previously observed by Postarnak and Zhavoronkov (2023) as a characteristic of the urban flora of other cities. The monotypic families are largely present in the dendroflora of the analyzed urban park, namely 10 families in the native dendroflora (22.22% of total) and 19 families (42.22% of total) in the non-native dendroflora. The dominance of the -native species (62.17%) in the analyzed urban park, combined with the dominance of monotypic families, are characteristics of the urban green spaces and a strategy of the urbanistic managements to increase the species richness in the urban parks, because the species richness is an indicator of their sustainability. However, despite this desirable goal, the voluntarily introduced non-native species could gradually transform into invasive species, threatening the local diversity and sometimes compromising the adjacent ecosystems (Seboko et al. 2024). A limiting factor for this inconvenient is the urban environment itself, because it offers limited space for spontaneously emerging since the great part of the urban soil is sealed and controlled. But this aspect increases the urban plant competition for the safe sites (Horvat et al. 2024) defined as appropriate sites for germination and emergence, and within this competition the herbaceous plants are more likely to succeed (Horvat et al. 2024). Thus, the urban parks, voluntarily settled by man, remain the major way to have woody plants in cities to benefit from their environmental advantages. The dominant non-native species in the park dendroflora has been investigated in the present study only for the plant life-forms and ecological requirements for moisture, temperature and soil reaction, but the implications of the dominance of the alien woody species in urban space could have also other implications. For example, although the presence of the native woody species in the urban green spaces has been found to favour the relationships between flora and fauna, supporting or enhancing the local biodiversity, there are also studies regarding diverse non-native woody taxa in urban parks which showed that the woody non-native species impoverish these relationships and lower the diversity of the native communities (Nielsen 2014). Some studies highlighted the role of the trees as ecological indicators in the urban green planning, recommending the prioritization of the native species in urban planting (Nobre Lisboa et al. 2024) to keep on the urban identity. But others suggested that the urban allochthonous woody species have generative potential to be used as seed banks for the urban greenery (Dimitrova et al. 2023).
The analysis of the plant life-form spectrum of the park dendroflora showed that the main plant life-forms are megaphanerophytes (49%), followed by mesophanerophytes (41%) (Figure 2).
The megaphanerophytes dominate both the native (49%) and the non-native dendroflora (52%), but the native mesophanerophytes are almost double as species number than the non-native mesophanerophytes. The nanophanerophytes are low represented in the native dendroflora (10%) but are more representative in the non-native one (26%). In all types of ecosystems, in the competition for resources, the ratio between phanerophytes and other plant life-forms is important, including the ratio between megaphanerophytes and mesophanerophytes. In the natural ecosystems, the ratio between these two categories of plants provides information about vegetation dynamics and about the availability of certain resources (light, water, temperature), which in turn determines specific types of ecological interactions between different species that influence the structure and dynamics of biological communities (competition, parasitism, predation, extinction, ecological succession etc.). For example, a high ratio of megaphanerophytes may indicate an ecosystem dominated by mature forests, an ecosystem that tends toward or has already reached a climax stage, with a developed canopy level, a condition for high biodiversity at this level (habitat for other species, such as birds and insects, support for epiphytic species). A higher ratio of mesophanerophytes may suggest a younger successional stage or an ecosystem where disturbance conditions are present. A study regarding mountain grasslands facing with the expansion of the phanerophytes showed that these grasslands are more demanding for light and temperature, but the effect is similar with that manifested by the chamaephytes (Palaj and Kollár 2021). Other studies showed that the phanerophytes were least affected by extinction when different habitats have been analyzed (Stehlik et al. 2007). It seems that the height of the woody plants in the urban park in one of the most important factors (alongside canopy factors, and tree trunk diameter) in shaping microhabitats and competition strategies in the urban parks. In our study, the megaphanerophytes own 49% in the spontaneous dendroflora and respectively 52% in the alien dendroflora. These percentages of tall trees with long lifespan and slow growth in the park show their urban adaption on long term with all the advantages emerging from here. The tall tree species have been found to be more abundant in the urban green settlements (Yang et al. 2023a; Nobre Lisboa et al. 2024), these are preferred and recommended because of their better adaption to the urbanization: better regeneration rate, better competitiveness in resources exploitation, better resistance to urban stressors. Several studies indicated strong correlation between the tree size and urban biodiversity (Stagoll et al. 2012). The studied urban park is an anthropic ecosystem, designed as a dendrological park, and the ratio between mega- and mesophanerophytes was predetermined. The fact that mega- and mesophanerophytes dominate the park (90%) as compared to nanophanerophytes determines a certain vertical and horizontal spatial distribution important in describing habitats and microhabitats and thus the potential of this type of vegetation to support various life forms, which was also the argument for establishing the park in this form, namely to increase the urban biodiversity alongside the urban wellness. However, the selection of tree species composition greatly considers aesthetics when urban arboretums are established (Campbell-Arvai et al. 2024), therefore the ratio between woody plants with various heights expressed as mega-, meso-, and nanophanerophytes in the urban green spaces are an assumed design option, which should be a median between citizen preferences and experts recommendations. The ratio herbaceous/shrubs/woody plants is not only an issue of urban architecture or aesthetics, but is also a basis in establishing the expectations regarding the ecosystem services provided by the urban green spaces. For example, the tree size is correlated to the CO2 stoking capacity in the urban parks (Nero et al. 2024) and with the capacity to resist to freeze-thaw cycles, because the high trees have bio-physiological (conductive vessels) and physical (hydraulic features) characteristics (Niu et al. 2022) adapted to this purpose, so that the ecosystem services will not be stopped due to tree dieback. The settlement of compounding species in the urban green spaces should take into consideration the findings of several studies which showed a direct correlation between the anthropogenic built structures around the green space and the biodiversity loss (Yang et al. 2024), or factors like the species lifespan (Dümpelmann 2024), its efficiency or sensitivity in urban de-pollution (Fini et al. 2024), or the resilience in front of urbanization expansion, which confer several advantages to the woody vegetation to be chosen. The dominance of a certain plant life-form within a site has been found to be an important biomarker for a certain resource, like hemicryptophytes have been reported for the water availability in several urban habitats (Salinitro et al. 2018). The settlement of the analyzed urban park as a dendrological/arbuscular park aims to harness all the advantages provided by the very presence of the phanerophytes that constitute it: to mitigate certain undesirable environmental phenomena like air pollution, noise pollution, overheating (Lin et al. 2021), and to bring aesthetic benefits to the urban landscape.
From chorological point of view, the most numerous native woody species are Eurasian (32%) and European (30%), which shows the dominating European autochthony of the native dendroflora in the studied urban park. The species characteristic of the Pontic-Carpathian space, to which Romania belongs, are rare in the analyzed urban park (4%). There was noticed the high heterogeneity of the phytogeographic elements in the group of the native dendroflora represented by 16 chorological types (Figure 3).
The analysis of the species requirements for the factors moisture, temperature and soil reaction showed the dominance of mesophyte, mesothermal and slightly acido-neutrophilous species both in the native and non-native dendroflora (Figure 4, Figure 5, Figure 6, Figure 7, Figure 8 and Figure 9 ).
The dominance of the mesophytes in the analyzed urban park is compliant with the local temperate climate. The percentages of the mesohygrophytes differ majorly in the native dendroflora (9%) compared with the non-native one (23%) (Figure 4 and Figure 5).
The acclimation capacity of the mesohygrophytes in the studied arboretum is possible because it is sustained by the moisture regime offered by the park management. There was found in the natural forest ecosystems that changes of the moistening degree could induce the transformation of the plant cover structure, such as appearance of the hygrophytes or the increase of the mesohygrophytes role in the phytocoenosis (Morozkin et al. 2001). Other studies showed the contribution of the environmental factors upon the vegetation shifting from one type of moisture requirement to another. A study regarding the riparian vegetation in China, indicated the vegetation shifting from xerophytes to mesophytes or even to hygrophytes in areas where the flooding fluctuations in a vegetal biocoenosis where the annual herbs were the dominant life form (Liu et al. 2024).
Within the native dendroflora, six plant species from the total 73 species, meaning 8%, are microthermal (Figure 6): 2 species are mesophanerophytes (Salix viminalis and Spiraea salicifolia), and 4 species are megaphanerophytes (Alnus incana, Betula pendula, Pinus mugo, and Sorbus aucuparia) (Table 2).
The acclimation capacity of the microthermal species in cities, which are known as heat islands, in contrast to their surrounding environments, has been previously reported and is possible in the cooler microhabitats provided by the urban matrix (Géron et al. 2022; Yang et al. 2023b): ventilation corridors (You and Liang 2024), reflective building materials, shading-oriented design of the buildings or other elements of urban morphology, like building geometry or building height, suggesting that complex built context is more efficient in urban cooling due to its increased shading potential (Li et al. 2024). Salix viminalis is a microthermal species successfully adapted to the temperate climates and urban environment (Teodorescu et al. 2011), which demonstrated real contributions for the urban environment, due to its potential to grow on metal-contaminated urban soil and for its potential of metal phytoextraction (Zn and Cd) from the contaminated urban soil and thus of phytoremediation of the urban soil (Jensen et al. 2009; Grignet et al. 2020). Spiraea salicifolia is a microthermal species with high efficiency in carbon sequestration in the urban green spaces (Fan et al. 2023). In the arctic urban climates, Spiraea salicifolia has shown low resistance to winter freezing, although it develops physiological adaptations to mitigate the winter hardiness by stocking sucrose in their tissues in autumn, to be later released during the winter period (Andronova and Platonov 2022). Alnus incana and Sorbus aucuparia are important species for the urban environment, these species have bioaccumulation potential of the cesium, transferring it from the urban topsoil in their biomass (Lipatov et al. 2023), so that the adaption of these microthermal tree species to the temperate climate of the analyzed site ends in an important ecosystem services. Alnus incana is also an efficient species in particulate matter removal (Muhammad et al. 2022) and phytoremediation of the organic hydrocarbons (Hostyn et al. 2022) in the urban areas. Other studies showed competition in the urban environments between Alnus incana, Sorbus aucuparia and Betula pendula: the decrease in abundance of the Sorbus aucuparia determined the abundance increase of Alnus incana and the decrease of Betula pendula in an urban forest from Finland (Hamberg et al. 2015). Alnus incana has also other benefits for the urban soils, contributing to the nutrient cycles, through nitrogen fixation in the roots, via symbiotic Frankia, even in polluted soils (Ridgway et al. 2004). Betula pendula is a species largely encountered in the natural areas of Northeastern and Central Europe and is a pioneer species in the natural ecological successions (Krisans et al. 2022), with characteristics emerging from this quality: fast growing and thus lower mechanical properties, which make this species more vulnerable to the stressing micro-environmental, local conditions of the site, like winds (Krisans et al. 2021). From this point of view, it might be possible that Betula pendula be a more successful species in cities, in organized arboretums, where the winds are attenuated by the buildings, because some studies indicate that the local climatic conditions have more significant contribution to the mechanical stability of this species in cities than its mechanical properties, since no significant differences between the mechanical properties of this species in the urban areas versus in the forests has been found (Krisans et al. 2022). However, this mechanical sensitivity of the Betula pendula species is outweighed by its aesthetic and ecological benefits for the urban environment (Petrushkevych and Korshykov 2020). Pinus mugo, although a microthermal species, is well acclimated in cities due to its resistance to drought, and it is resistant in the arid and semi-arid regions. The mechanisms which mediate these adaptions, making the Pinus mugo appropriate to be planted in dry and drought environments are physiological and biochemical and refer to: shoot biomass, chlorophyll and carotenoid contents, water content, electrolyte leakage, sugar content, antioxidant enzymes, nutrient content, fatty acid content, protein content, osmosis (Nouri et al. 2023). The acclimative capacity of the above mentioned microthermal tree species to the conditions of the temperate climate is proven by their presence and continuity in the studied urban site as native species of the Romanian flora, and may represent a potential solution to certain urban environmental issues, and thus a source of ecosystem services for the studied urban environment.
The alien dendroflora of the studied urban park is characterized majorly through mesothermal and moderate termophilic species, as expected for a temperate climate site, but also through few microthermal - 7 (6%) species and respectively through 3 (2%) cryophyle species (Figure 7). The microthermal species are: Berberis julianae, Berberis stenophylla, Berberis thunbergii, Berberis haoi, Abies concolor, Pinus excela, Pinus wallichiana, and the cryophyle tree species are: Abies pinsapo, Picea pungens, and Tsuga canadensis. These species has been previously reported in the temperate-climate urban parks. The acclimation of the plant species with low-temperature requirements in temperate zones can be possible because the tolerance to cold stress can be lost after exposure to warmer climate, through faster processes than the acquiring one (Kalberer et al. 2006). This mechanism enables the plants species that possess it to have great responsiveness and adaptability to different types of climates and has been observed also in plants with Arctic origin (Chew et al. 2012).
Some studies have shown a decline of the native tree species across Europe (Klisz et al. 2021), and the acclimation of the non-native tree species seems to be one of the reasons of their invasive potential threatening the native species. However, an analysis about the distribution of the woody invaders from contrasting climatic origins across the urban-rural gradient in oceanic Europe, showed that the woody alien plants with warmer native requirements are more present in the urban local climates (Géron et al. 2022).
A statistically significant association (Chi-square test) was found between the moisture and temperature requirements of species in both native (χ2 (15, N=73) = 49.11, p < 0.001) and non-native dendroflora (χ2 (25, N=120) = 65.91, p < 0.001) of the studied urban park. The response of the urban vegetation to the urban temperature oscillations (frequent and prolonged heat-waves) in the actual context of climate changes is a complex phenomenon involving structural and physiological mechanisms, not always in accordance to the native characteristics of the species. The woody vegetation reacts dynamically to keep its resilience to temperature variations, by changes in the stomatal conductance, leaf water potential, photosynthesis efficiency, respiration and evapotranspiration (Esperon-Rodriguez et al. 2021) or through physical and mechanical strategies able to modify the surrounding microclimate, like reciprocal leaf shading (Wright and Francia 2024). Other mechanisms are the geographic distributional shifting, but in the case of the exotic species acclimated in the urban parks, the niche breadth (referring to the range of ecological factors tolerated by the species) of their natural distribution is not respected, so that the physiological mechanisms remain the primary strategy adopted for coping with and adapting to the local environmental conditions.
Another statistically significant associations (Chi-square test) was found between the temperature preferences and soil reaction preferences of the studied dendroflora, both native (χ2 (12, N=73) = 40.55, p < 0.001) and non-native (χ2 (25, N=120) = 59.25, p < 0.001). This correlation is important in assessing the potential of the identified species to develop adapting relationships to the urban environmental factors, because the temperature (Maes et al. 2020) and the soil pH are closely influencing the plant functional traits (Song et al. 2019) in their acclimation and adaption strategies. However, the pH variability of the urban soils is high and therefore correlations between it and other plant features are difficult to be obtained. A study concerning several Mediterranean evergreen woods showed not statistically significant correlations of the species preferences for soil pH with other plant traits like temperature and precipitations (Marcenò and Guarino 2015). Other studies carried in human-controlled ecosystems, such as arable lands, showed correlations between the plant preferences for soil pH and site elevation, and respectively between plant preferences for temperature and elevation or season (Lososova et al. 2004; Di Biase et al. 2023).
In the studied dendrological park, the main part of the woody vegetation (37% of native dendroflora and 50% of non-native dendroflora) natively prefers the slightly acido-neutrophilous soils (Figure 8 and Figure 9), but this fact does not transform the respective flora into an indicative one of soil pH, because there are several studies which indicate that the native preference of the woody species for soil reaction does not accurately describe the in situ value of soil pH (Lawesson 2003; Carpenter and Goodenough 2014) due to the extensive mechanisms of adaption to the environmental local factors exhibited by the species. For example, due to the irrigation practices imposed by the management of the urban parks (Zalacáin et al. 2019) or because of other factors like the deicing salts which are used in winter in the temperate zones to defrost the urban streets and which are indicated as the main cause of salt stress in the urban environments (Dmuchowski et al. 2021), the urban soil faces with salt accumulation, an environmental soil factor to which the urban plants must be adapted during their acclimation or to exceed the limits imposed by their native requirements.
The pH variability of the urban soils is a phenomenon widely appearing in the urban areas and this has been described in many urban sites (Rahmonov et al. 2024). Because of this limitation, correlations between the native preferences of the plants regarding the environmental factors become viable options to be considered, especially in the ecological studies regarding the acclimation and adaption of the plant species in the urban area, where the soil pH reaches high variability. For example, the plant preferences for pH have been proven to be a good predictor of species richness (Chytrý et al. 2002).
In the studied urban arboretum, within the acclimation process of the non-native dendroflora, 37% of species exceeded their native requirements for moisture, 41% for temperature, and 50% for soil reaction.
A Chi-square test has been conducted to determine whether there is an association between the studied factors in the native dendroflora of the studied urban green space. There was found significant associations between the plant life-form (megaphanerophyte, mesophanerophyte, nanophanerophyte) and the ecological plant requirements for soil reaction (χ2 (8, N=73) = 16.27, p = 0.039) (Figure 10 and Figure 11), and respectively between the plant family and the plant requirement for moisture (χ2 (110, N=73) = 139.72, p = 0.029) (Figure 12 and Figure 13).
For the non-native woody species, the Chi-square test has shown a statistically significant association between the plant life-form and the temperature preferences for all three types of plant life-forms (megaphanerophytes, mesophanerophytes and nanophanerophytes) present in the studied arboretum (Chi-square test, χ2 (10, N=120) = 19.36, p = 0.036) (Figure 14 and Figure 15).
The fact that plant life-form is associated with the plant pH requirement in the native dendroflora, while in the alien dendroflora is associated with the temperature requirement results from the evolutionary adaption strategies of the species during their historical survival in various environments. The introduced species face often shifting ecological requirements necessitating fast acclimation adaption, and the temperature is a first filter in species selection and spread, unlike native species which evolves in situ. Because of their long life span, the woody species tend to specialize more in their native soil pH because this is a factor involved in multiple ecological interactions and a driver of the ecological niches. Thus, the evolutionary pressure shapes the woody species to match the soil conditions of their habitats, ensuring the species long time resilience.
Conclusions
The species composition of the studied urban arboretum indicates high species richness. The species are taxonomically and biogeographically diverse, both native and non-native, suggesting the great ability of the identified woody species in acclimating to the temperate environment characteristic to the studied site.
The dominant dendroflora is mesophyte, mesothermal and slightly acido-neutrophilous. The evolutionary and acclimating pressure shaped the woody species of the urban park to match the habitat conditions according to their native requirements for moisture, temperature and soil reaction, or shifting them. Therefore, in the acclimation process of the non-native dendroflora, 37% of species exceeded their native requirements for moisture, 41% for temperature, and 50% for soil pH. With the same purpose, there was found that in the acclimation process, the plant life-form is relevant both in the native and alien dendroflora, and the plant family is relevant only in the native dendroflora. These findings sustain the sustainability of the studied urban park and its long time resilience, ensuring its potential in providing local ecosystem services.
Acknowledgments
The article publishing fee is supported by the University of Life Sciences “King Mihai I” from Timisoara, Romania.
References
- Andronova, M., and A. Platonov. 2022. Sucrose in the tissues of annual shoots of introduced woody plants. Lesnoy Zhurnal-Forestry Journal 1: 62–76. [Google Scholar] [CrossRef]
- Bartoli, F., V. Savo, and G. Caneva. 2022. Biodiversity of urban street trees in Italian cities: a comparative analysis. Plant Biosystems 156, 3: 649–662. [Google Scholar] [CrossRef]
- Campbell-Arvai, V., R. Vergel, M. Lindquist, N. Fox, and D. Van Berkel. 2024. Tree selection for a virtual urban park: Comparing aided and unaided decision-making to support public engagement in greenspace design. Urban Forestry & Urban Greening 99: 128447. [Google Scholar] [CrossRef]
- Carpenter, W., and A. Goodenough. 2014. How robust are community-based plant bioindicators? Empirical testing of the relationshipbetween Ellenberg values and direct environmental measures in woodland communities. Community Ecology 15: 1–11. [Google Scholar] [CrossRef]
- Chew, Y. H., A. M. Wilczek, M. Williams, S. M. Welch, J. Schmitt, and K. J. Halliday. 2012. An augmented Arabidopsis phenology model reveals seasonal temperature control of flowering time. New Phytologist 194, 3: 654–665. [Google Scholar] [CrossRef]
- Chytrý, M., L. Tichý, and J. Roleček. 2002. Local and regional patterns of species richness in Central European vegetation types along the pH/calcium gradient. Folia Geobotanica 38: 429–442. [Google Scholar] [CrossRef]
- Ciocârlan, V. 2000. Flora ilustrată a României, Editura Ceres, București. (in Romanian) [Google Scholar]
- Ciupa, V. 2010. Cadrul natural şi peisagistic al Municipiului Timişoara. Volumul I. (In Romanian). Available online: https://www.primariatm.ro/wpcontent/uploads/2020/11/Cadrul_Natural_Timisoara_vol.1.pdf (accessed on May 2024). (In Romanian).
- Ciupa, V. 2018. Timișoara-Oraș grădină, oraș al parcurilor, oraș al florilor-Monografie. In Editura ArtPress. Timișoara. (In Romanian) [Google Scholar]
- Cristea, V., D. Gafta, and F. Pedrotti. 2004. Fitosociologie. Editura Presa Universitară Clujană, Cluj-Napoca. (in Romanian) [Google Scholar]
- Di Biase, L., N. Tsafack, L. Pace, and S. Fattorini. 2023. Ellenberg indicator values disclose complex environmental filtering processes in plant communities along an elevational gradient. Biology-Basel 12, 2: 161. [Google Scholar] [CrossRef]
- Dimitrova, A., V. Stipanovic, and D. Kolevska. 2023. Collection of Experiences: 25 Years' Work on Seed Propagation of Allochthonous Woody Plants in Skopje and Their Possible Role in the Urban Landscape. Seefor-South-East European Forestry 14, 1: 53–67. [Google Scholar] [CrossRef]
- Dmuchowski, W., A. Baczewska-Dabrowska, D. Gozdowski, P. Bragoszewska, B. Gworek, I. Suwara, T. Chojnacki, A. Jóźwiak, and E. Swiezewska. 2021. Effect of salt stress in urban conditions on two Acer species with different sensitivity. PEERJ 9: e10577. [Google Scholar] [CrossRef]
- Dümpelmann, S. 2024. Tree Times: Urban Plants as Timekeepers and Seasonal Indicators. Journal of Urban History. [Google Scholar] [CrossRef]
- Ellenberg, H. 1952. Landwirtschaftliche Pflanzensoziologie II. Wiesen und Weiden und ihre standörtliche Bewertung; Ulmer: Stuttgart,Germany; pp. 1–143.17. [in German]. Available online: https://www.researchgate.net/publication/367323581_Ellenberg_Indicator_Values_Disclose_Complex_Environmental_Filtering_Processes_in_Plant_Communities_along_an_Elevational_Gradient (accessed on October 2024). (in English).
- Esperon-Rodriguez, M., S. Power, M. Tjoelker, R. Marchin, and P. Rymer. 2021. Contrasting heat tolerance of urban trees to extreme temperatures during heatwaves. Urban Forestry & Urban Greening 66: 127387. [Google Scholar] [CrossRef]
- Fan, L., J. Wang, D. Han, J. Gao, and Y. Yao. 2023. Research on Promoting Carbon Sequestration of Urban Green Space Distribution Characteristics and Planting Design Models in Xi'an. Sustainability 15, 1: 572. [Google Scholar] [CrossRef]
- Fini, A., I. Vigevani, D. Corsini, P. Wężyk, K. Bajorek-Zydron, O. Failla, E. Cagnolati, L. Mielczarek, S. Comin, M. Gibin, A. Pasquinelli, F. Ferrini, and P. Viskanic. 2024. CO2 assimilation, sequestration, and storage by urban woody species growing in parks and along streets in two climatic zones. SCIENCE OF THE TOTAL ENVIRONMENT 927: 172355. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, W., A. Martini, S. Martins, M. Oliveira, L. Duenez, and W. Alves. 2024. Exploring urban forests in Minas Gerais, Brazil: floristic diversity and biome-driven insights to green infrastructure planning. Urban Ecosystems 27, 6: 2331–2347. [Google Scholar] [CrossRef]
- Galfrascoli, G., G. Bernardello, and A. Calviño. 2023. How well do trees fit the city? Lessons from an urban tree survey in Córdoba, Argentina. Boletin de la Sociedad Argentina de Botanica 58, 4: 561–572. [Google Scholar] [CrossRef]
- Géron, C., J. Lembrechts, I. Nijs, and A. Monty. 2022. Woody invaders from contrasted climatic origins distribute differently across the urban-to-rural gradient in oceanic Europe-Is it trait-related? Urban Forestry & Urban Greening 75: 127694. [Google Scholar] [CrossRef]
- Grignet, A., A. de Vaufleury, A. Papin, and V. Bert. 2020. Urban soil phytomanagement for Zn and Cd in situ removal, greening, and Zn-rich biomass production taking care of snail exposure. Environmental Science and Pollution Research 27, 3: 3187–3201. [Google Scholar] [CrossRef]
- Hamberg, L., S. Lehvävirta, D. Kotze, and J. Heikkinen. 2015. Tree species composition affects the abundance of rowan Sorbus aucuparia L.) in urban forests in Finland. Journal of Environmental Management 151: 369–377. [Google Scholar] [CrossRef]
- Heneidy, S. Z., M. W.A. Halmy, S. M. Toto, S. K. Hamouda, A. M. Fakhry, L. M. Bidak, E. M. Eid, and Y. M. Al-Sodany. 2021. Pattern of Urban Flora in Intra-City Railway Habitats (Alexandria, Egypt): A Conservation Perspective. Biology-Basel 10, 8: 698. [Google Scholar] [CrossRef]
- Hirons, A. D., J. H.R. Watkins, T. J. Baxter, J. W. Miesbauer, A. Male-Muñoz, K. W.E. Martin, N. L. Bassuk, and H. Sjöman. 2021. Using botanic gardens and arboreta to help identify urban trees for the future. Plants People Planet 3, 2: 182–193. [Google Scholar] [CrossRef]
- Horvat, E., M. Sipek, and N. Sajna. 2024. Urban hedges facilitate spontaneous woody plants. Urban Forestry & Urban Greening 96: 128336. [Google Scholar] [CrossRef]
- Hostyn, G., C. Schwartz, J. Côme, and S. Ouvrard. 2022. Assessment for combined phytoremediation and biomass production on a moderately contaminated soil. Environmental Science and Pollution Research 29, 39: 59736–59750. [Google Scholar] [CrossRef] [PubMed]
- Jang, J., and S. Woo. 2022. Native Trees as a Provider of Vital Urban Ecosystem Services in Urbanizing New Zealand: Status Quo, Challenges and Prospects. Land 11, 1: 92. [Google Scholar] [CrossRef]
- Jensen, J., P. Holm, J. Nejrup, M. Larsen, and O. Borggaard. 2009. The potential of willow for remediation of heavy metal polluted calcareous urban soils. Environmental Pollution 157, 3: 931–937. [Google Scholar] [CrossRef]
- Kalberer, S. R., M. Wisniewski, and R. Arora. 2006. Deacclimation and reacclimation of cold-hardy plants: current understanding and emerging concepts. Plant Science 171: 3–16. [Google Scholar] [CrossRef]
- Klisz, M., R. Puchalka, M. Netsvetov, Y. Prokopuk, M. Vítková, J. Sádlo, R. Matisons, M. Mionskowski, D. Chakraborty, P. Olszewski, T. Wojda, and M. Koprowski. 2021. Variability in climate-growth reaction of Robinia pseudoacacia in Eastern Europe indicates potential for acclimatisation to future climate. Forest Ecology and Management 492: 119194. [Google Scholar] [CrossRef]
- Krisans, O., L. Caksa, R. Matisons, S. Rust, D. Elferts, A. Seipulis, and A. Jansons. 2022. A static pulling test is a suitable method for comparison of the loading resistance of silver birch Betula pendula Roth.) between urban and peri-urban forests. Forests 13, 1: 127. [Google Scholar] [CrossRef]
- Krisans, O., R. Matisons, M. Kitenberga, J. Donis, S. Rust, D. Elferts, and A. Jansons. 2021. Wind resistance of Eastern Baltic silver birch (Betula pendula Roth.) suggests its suitability for periodically waterlogged sites. Forests 12, 1: 21. [Google Scholar] [CrossRef]
- Lahoti, S., A. Lahoti, R. K. Joshi, and O. Saito. 2020. Vegetation structure, species composition, and carbon sink potential of urban green spaces in Nagpur City, India. Land 9, 4: 107. [Google Scholar] [CrossRef]
- Lakicevic, M., K. M. Reynolds, S. Orlovic, and R. Kolarov. 2022. Measuring dendrofloristic diversity in urban parks in Novi Sad (Serbia). Trees, Forests and People 8: 100239. [Google Scholar] [CrossRef]
- Lawesson, J. E. 2003. pH optima for Danish forest species compared with Ellenberg reaction values. Folia Geobotanica 38: 403–418. [Google Scholar] [CrossRef]
- Li, S., Y. Zhu, H. Wan, Q. Xiao, M. Teng, W. Xu, X. Qiu, X. Wu, and C. Wu. 2024. Effectiveness of potential strategies to mitigate surface urban heat island: A comprehensive investigation using high-resolution thermal observations from an unmanned aerial vehicle. Sustainable Cities and Society 113: 105716. [Google Scholar] [CrossRef]
- Lin, B. B., A. Ossola, W. Ripple, M. Alberti, E. Andersson, X. Bai, C. Dobbs, T. Elmqvist, K. Evans, N. Frantzeskaki, R. Fuller, K. Gaston, D. Haase, C. Jim, C. Konijnendijk, H. Nagendra, J. Niemela, T. McPhearson, W. Moomaw, S. Parnell, D. Pataki, and P. Tan. 2021. Integrating solutions to adapt cities for climate change. The Lancet Planetary Health 5: 479–486. [Google Scholar] [CrossRef] [PubMed]
- Lipatov, D., V. Varachenkov, D. Manakhov, S. Mamikhin, and A. Shcheglov. 2023. 137Cs pollution in soils and plants of urban ecosystems near the Elektrostal Heavy Machinery Plant. Biology Bulletin 50, 12: 3383–3393. [Google Scholar] [CrossRef]
- Liu, Y., X. Duan, X. Li, W. Yi, G. Chen, J. Yang, D. Deng, X. Guo, Z. Yang, G. Huang, M. Hu, and C. Ye. 2024. Anti-seasonal flooding drives substantial alterations in riparian plant diversity and niche characteristics in a unique hydro-fluctuation zone. Ecology and Evolution 14, 8: e70036. [Google Scholar] [CrossRef]
- Lososová, Z., M. Chytrý, S. Cimalová, Z. Kropáč, Z. Otýpková, P. Pyšek, and L. Tichý. 2004. Weed vegetation of arable land in Central Europe: Gradients of diversity and species composition. Journal of Vegetation Science 15: 415–422. [Google Scholar] [CrossRef]
- Maes, S. L., M. P. Perring, L. Depauw, M. Bernhardt-Römermann, H. Blondeel, G. Brūmelis, J. Brunet, G. Decocq, J. den Ouden, S. Govaert, W. Härdtle, R. Hédl, T. Heinken, S. Heinrichs, L. Hertzog, B. Jaroszewicz, K. Kirby, M. Kopecký, D. Landuyt, F. Máliš, T. Vanneste, M. Wulf, and K. Verheyen. 2020. Plant functional trait response to environmental drivers across European temperate forest understorey communities. Plant Biology 22, 3: 410–424. [Google Scholar] [CrossRef]
- Marcenò, C., and R. Guarino. 2015. A test on Ellenberg indicator values in the Mediterranean evergreen woods Quercetea ilicis. Rendiconti Lincei-Scienze Fisiche e Naturali 26, 3: 345–356. [Google Scholar] [CrossRef]
- Morozkin, A., S. Kalimullina, L. Salova, and T. Shpak. 2001. Status of forest ecosystems in the impact zone of the Nizhnekamsk industrial complex. Eurasian Soil Science 34, 12: 1323–1330. [Google Scholar]
- Muhammad, S., K. Wuyts, and R. Samson. 2022. Selection of plant species for particulate matter removal in urban environments by considering multiple ecosystem (dis)services and environmental suitability. Atmosphere 13, 12: 1960. [Google Scholar] [CrossRef]
- Muhlisin, J. I., B. Gunawan, and M. F. Cahyandito. 2021. Vegetation diversity and structure of urban parks in Cilegon City, Indonesia, and local residents’ perception of its function. Biodiversitas 22, 7: 2589–603. [Google Scholar]
- Muvengwi, J., H. Ndagurwa, E. Witkowski, and M. Mbiba. 2024. Woody species composition, diversity, and ecosystem services of yards along an urban socioeconomic gradient. Science of The Total Environment 912: Article 168976. [Google Scholar] [CrossRef]
- Nero, B., E. Kuusaana, A. Ahmed, and B. Campion. 2024. Carbon storage and tree species diversity of urban parks in Kumasi, Ghana. City and Environment Interactions 24: Article 100156. [Google Scholar] [CrossRef]
- Nielsen, A. B., M. van den Bosch, S. Maruthaveeran, and C. Konijnendijk van den Bosch. 2014. Species richness in urban parks and its drivers: A review of empirical evidence. Urban Ecosystems 17: 305–327. [Google Scholar] [CrossRef]
- Niu, C., W. Shou, L. Ma, and J. Qian. 2022. Tree height-related hydraulic strategy to cope with freeze-thaw stress in six common urban tree species in North China. Phyton-International Journal of Experimental Botany 91, 4: 811–825. [Google Scholar] [CrossRef]
- Nobre Lisboa, M. A., L. V. Alves da Silva, A. da Silva Nascimento, A. de Oliveira Silva, M. R. Alves Teixeira, M. F.R. Ferreira, S. Cardoso Fereira, A. C. Vieira da Silva, A. Viana Corales, and J. Tovares Calixto. 2024. Diversity, structure, and carbon sequestration potential of the woody flora of urban squares in the Brazilian semiarid region. Trees Forests and People 16: 100561. [Google Scholar] [CrossRef]
- Nouri, K., A. Nikbakht, M. Haghighi, N. Etemadi, M. Rahimmalek, and A. Szumny. 2023. Screening some pine species from North America and dried zones of western Asia for drought stress tolerance in terms of nutrients status, biochemical and physiological characteristics. Frontiers in Plant Science 14: 1281688. [Google Scholar] [CrossRef]
- Palaj, A., and J. Kollár. 2021. Expansion of phanerophytes above the timberline in the Western Carpathians. Biologia 76, 7: 1991–2003. [Google Scholar] [CrossRef]
- Petrushkevych, Y., and I. Korshykov. 2020. Ecological and biological characteristics of Betula pendula in the conditions of urban environment. Regulatory Mechanisms in Biosystems 11, 1: 29–36. [Google Scholar]
- Postarnak, Y., and V. Zhavoronkov. 2023. Urban dendroflora of dry subtropics of the northwestern part of the greater Caucasus on the example of the city of Gelendzhik. Russian Journal of Earth Sciences 23, 5: ES0203. [Google Scholar] [CrossRef]
- Primăria Timișoara–Registrul local al spațiilor verzi-Municipiul Timișoara-Parcul Botanic, Executant S.C. Geotop S.R.L., Beneficiar Primăria Timișoara, Faza ”Actualizarea lucrării «Cadastru verde existent (parcuri) pe baza datelor existente la beneficiar» ”(in Romanian), no document datation. (accessed on 8 May 2024).
- Rahmonov, O., A. Kowal, M. Rahmonov, and S. Pytel. 2024. Variability of concentrations of potentially toxic metals in the topsoil of urban forest parks (Southern Poland). Forests 15, 6: 1020. [Google Scholar] [CrossRef]
- Raunkiaer, C. 1934. The life forms of plants and statistical plant geography. Oxford University Press: London. [Google Scholar]
- Ridgway, K. P., L. A. Marland, A. F. Harrison, J. Wright, J. P.W. Young, and A. H. Fitter. 2004. Molecular diversity of Frankia in root nodules of Alnus incana grown with inoculums from polluted urban soils. FEMS Microbiology Ecology 50, 3: 255–263. [Google Scholar] [CrossRef] [PubMed]
- Rogovskyi, S., L. Ishchuk, and H. Ishchuk. 2023. Chornobyl's current dendroflora: analysis of natural successions in the abandoned urban phytocoenoses. Trakya University Journal of Natural Sciences 24, 2: 5–21. [Google Scholar] [CrossRef]
- Salinitro, M., A. Alessandrini, A. Zappi, D. Melucci, and A. Tassoni. 2018. Floristic diversity in different urban ecological niches of a southern European city. Scientific Reports 8: 15110. [Google Scholar] [CrossRef]
- Sanda, V., A. Popescu, M. I. Doltu, and N. Doniţă. 1983. Caracterizarea ecologică şi fitocenologică a speciilor spontane din flora României. Studii şi comunicări, 25, Muzeul Brukenthal, Sibiu. (in Romanian) [Google Scholar]
- Sanda, V., and S. Stefanut. 2003. Atlas Florae Romaniae. I. Pinophytina. Editura Vergiliu, București (In Romanian).
- Sârbu, A., D. Smarandache, and G. Pascale. 2003. Îndrumător de practică botanică: Munţii Bucegi-Baiului. Editura Universităţii din Bucureşti. (In Romanian)
- Seboko, T., C. Shackleton, and S. Ruwanza. 2024. Urban residents' knowledge of and attitudes and willingness to control woody invasive alien plants in their domestic gardens in South Africa. People and Nature 6, 5: 2077–2090. [Google Scholar] [CrossRef]
- Simovic, M., K. E. Mueller, S. M. McMahon, and J. S. Medeiros. 2024. Functional traits and size interact to influence growth and carbon sequestration among trees in urban greenspaces. Functional Ecology 38: 967–983. [Google Scholar] [CrossRef]
- Song, G., J. Wang, T. Han, Q. Wang, H. Ren, H. Zhu, X. Wen, and D. Hui. 2019. Changes in plant functional traits and their relationships with environmental factors along an urban-rural gradient in Guangzhou, China. Ecological Indicators 106: 105558. [Google Scholar] [CrossRef]
- Stagoll, K., D. B. Lindenmayer, E. Knight, J. Fischer, and A. D. Manning. 2012. Large trees are keystone structures in urban parks. Conservation Letters 5, 2: 115–22. [Google Scholar] [CrossRef]
- Stehlik, I., J. Caspersen, L. Wirth, and R. Holderegger. 2007. Floral free fall in the Swiss lowlands: environmental determinants of local plant extinction in a peri-urban landscape. JOURNAL OF ECOLOGY 95, 4: 734–744. [Google Scholar] [CrossRef]
- Teodorescu, T., W. Guidi, and M. Labrecque. 2011. The use of non-dormant rods as planting material: A new approach to establishing willow for environmental applications. Ecological Engineering 37, 9: 1430–1433. [Google Scholar] [CrossRef]
- Wright, A., and R. Francia. 2024. Plant traits, microclimate temperature and humidity: A research agenda for advancing nature-based solutions to a warming and drying climate. Journal of Ecology 112, 11: 2462–2470. [Google Scholar] [CrossRef]
- Yang, J., C. Cen, Z. Wang, and M. Jian. 2024. Impacts of spatiotemporal urban expansion on the species richness and functional traits of adults and sapling woody trees and shrubs of urban remnant forest patches. Ecological Indicators 166: 112498. [Google Scholar] [CrossRef]
- Yang, J., Z. Wang, Y. Pan, and Y. Zheng. 2023a. Woody plant functional traits and phylogenetic signals correlate with urbanization in remnant forest patches. Ecology and Evolution 13, 8: e10366. [Google Scholar] [CrossRef] [PubMed]
- Yang, S., L. Wang, T. Stathopoulos, and A. M. Marey. 2023b. Urban microclimate and its impact on built environment-A review. Building and Environment 283: 110334. [Google Scholar] [CrossRef]
- You, W., and Y. Liang. 2024. Numerical investigation of different building configurations for improving outdoor spatial ventilation conditions in strip-type residential neighbourhoods. Urban Climate 56: 102012. [Google Scholar] [CrossRef]
- Zalacáin, D., S. Martínez-Pérez, R. Bienes, A. García-Díaz, and A. Sastre-Merlín. 2019. Salt accumulation in soils and plants under reclaimed water irrigation in urban parks of Madrid (Spain). Agricultural Water Management 213: 468–476. [Google Scholar] [CrossRef]
Figure 1.
The Botanic Park of Timişoara (45°45'18''N, 21°13'28''E) (Google Maps capture).
Figure 2.
Spectrum of plant life-forms in the native and non-native dendroflora of the Botanic Park from Timișoara City.
Figure 2.
Spectrum of plant life-forms in the native and non-native dendroflora of the Botanic Park from Timișoara City.
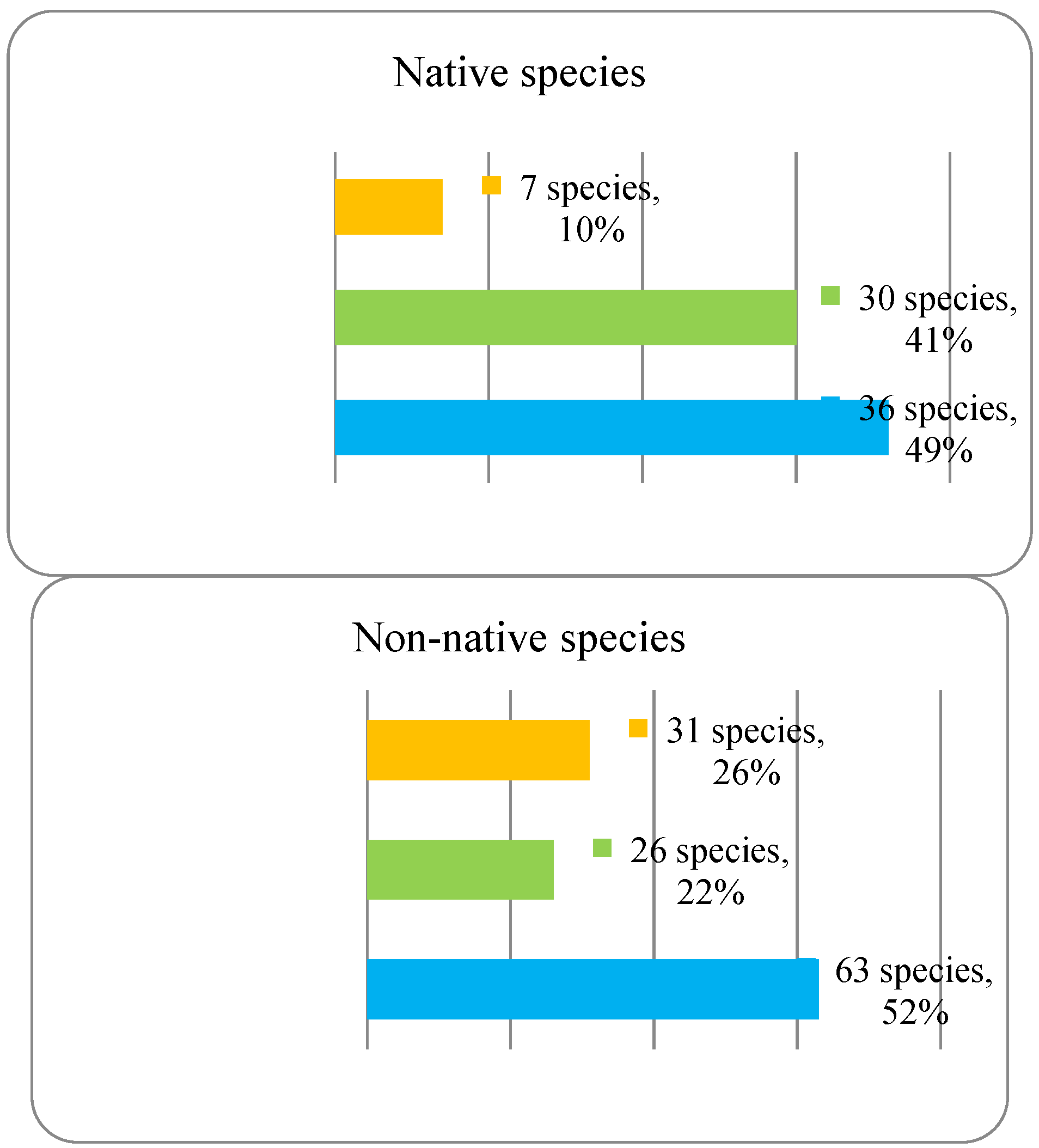
Figure 3.
Chorological spectrum regarding the geographic origin of the native woody species in the Botanic Park from Timișoara City.
Figure 3.
Chorological spectrum regarding the geographic origin of the native woody species in the Botanic Park from Timișoara City.
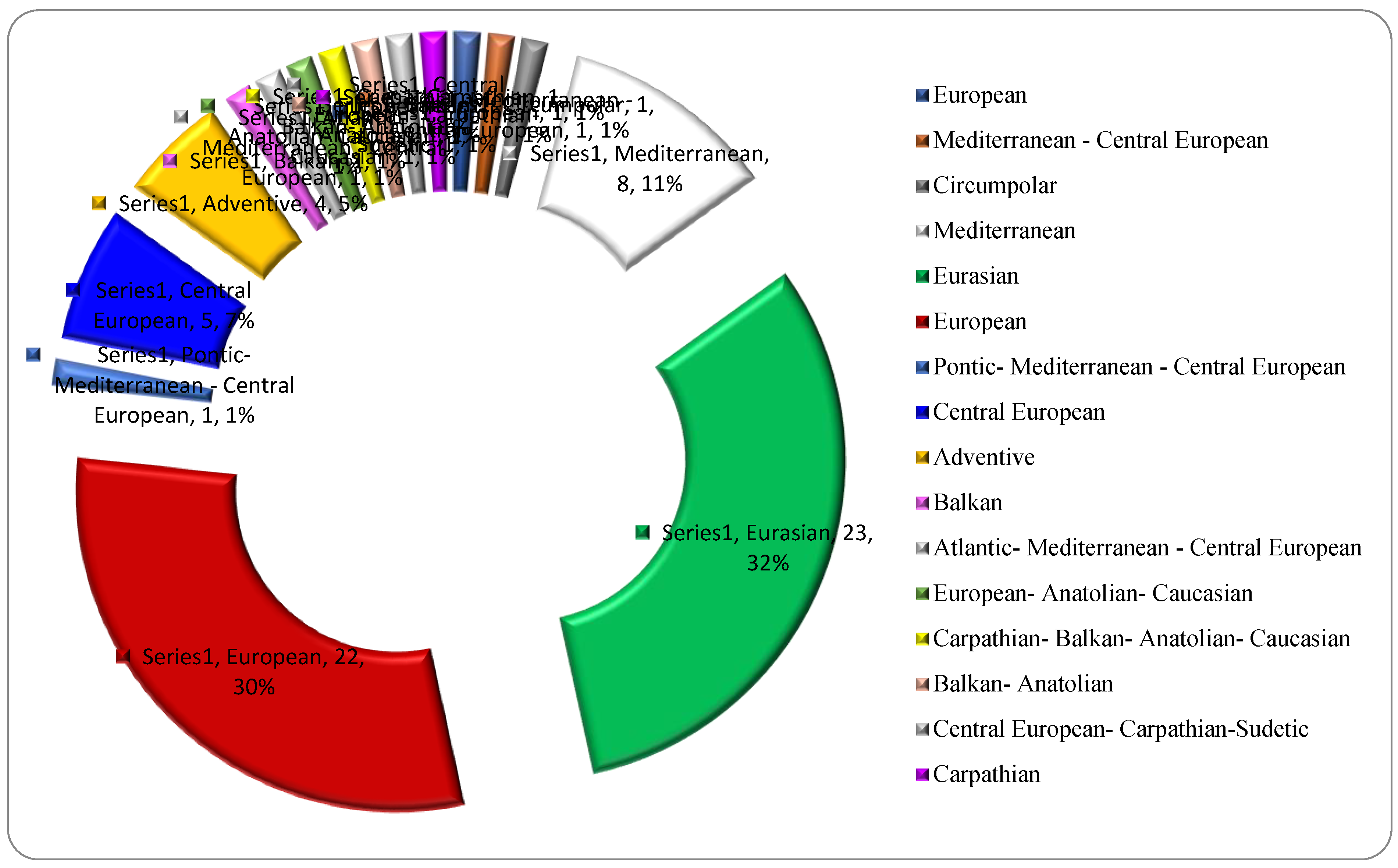
Figure 4.
The spectrum of species requirements for moisture in the native dendroflora of the Botanic Park from Timișoara City.
Figure 4.
The spectrum of species requirements for moisture in the native dendroflora of the Botanic Park from Timișoara City.
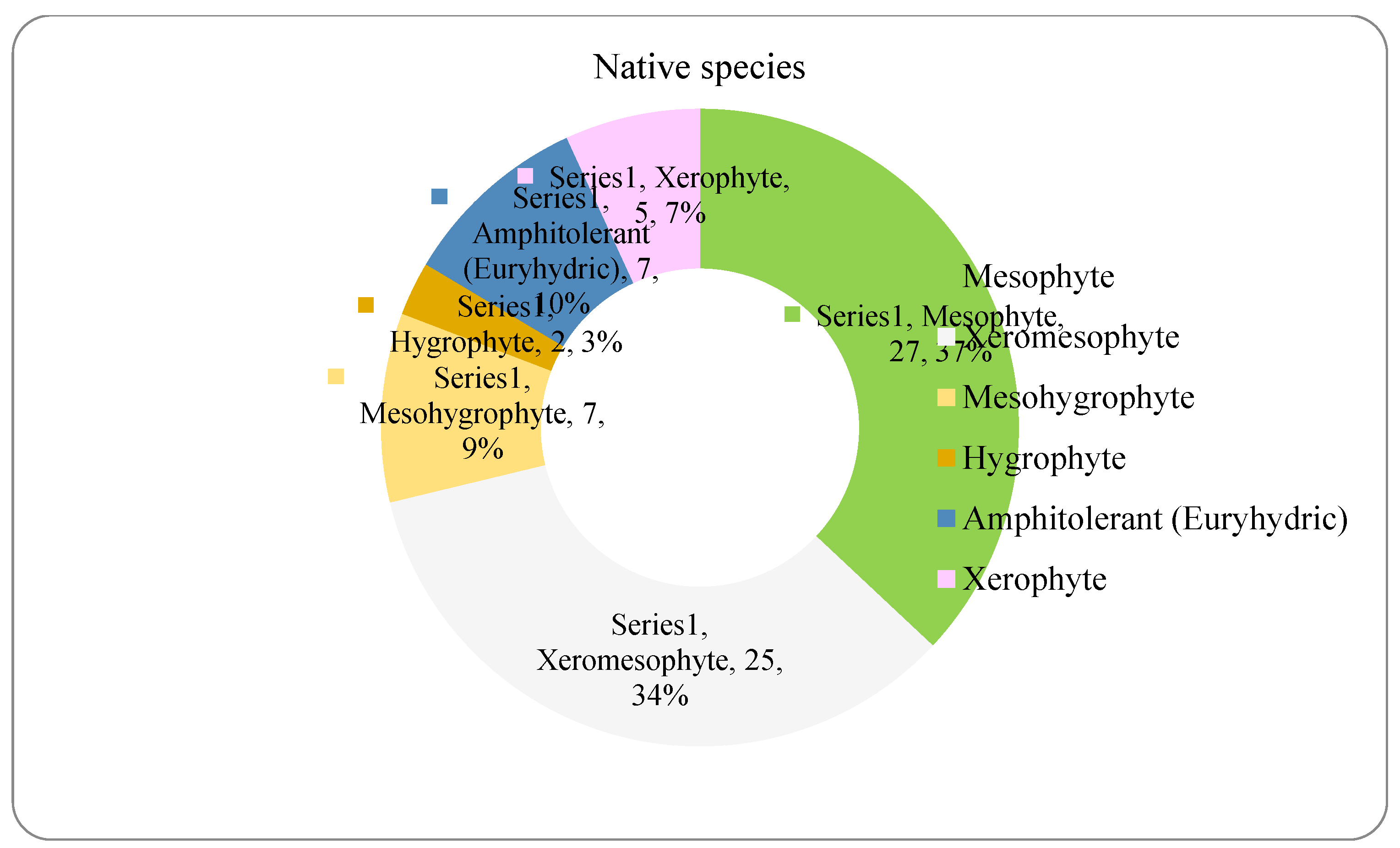
Figure 5.
The spectrum of species requirements for moisture in the non-native dendroflora of the Botanic Park from Timișoara City.
Figure 5.
The spectrum of species requirements for moisture in the non-native dendroflora of the Botanic Park from Timișoara City.
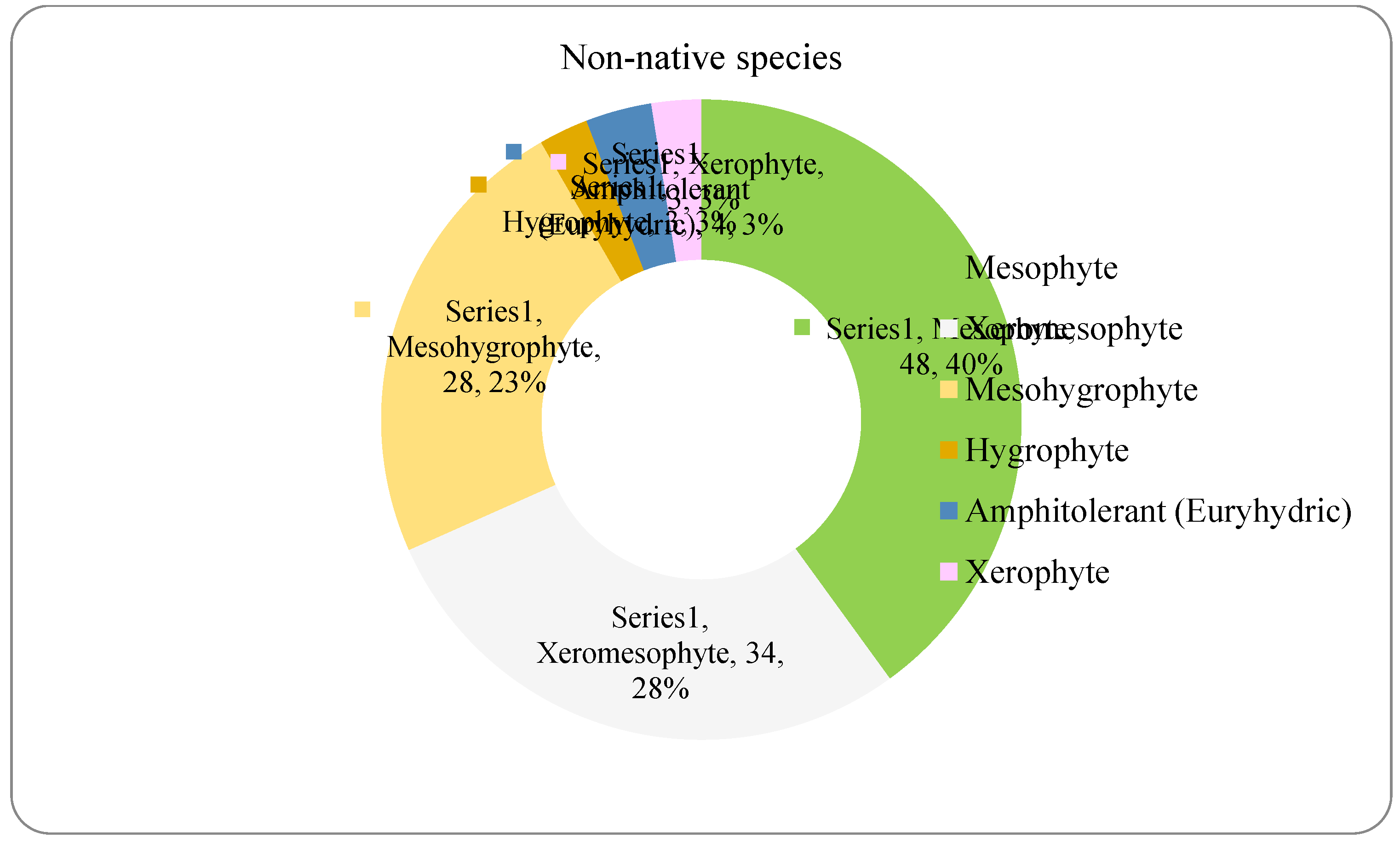
Figure 6.
The spectrum of species requirements for temperature in the native dendroflora of the Botanic Park from Timișoara City.
Figure 6.
The spectrum of species requirements for temperature in the native dendroflora of the Botanic Park from Timișoara City.
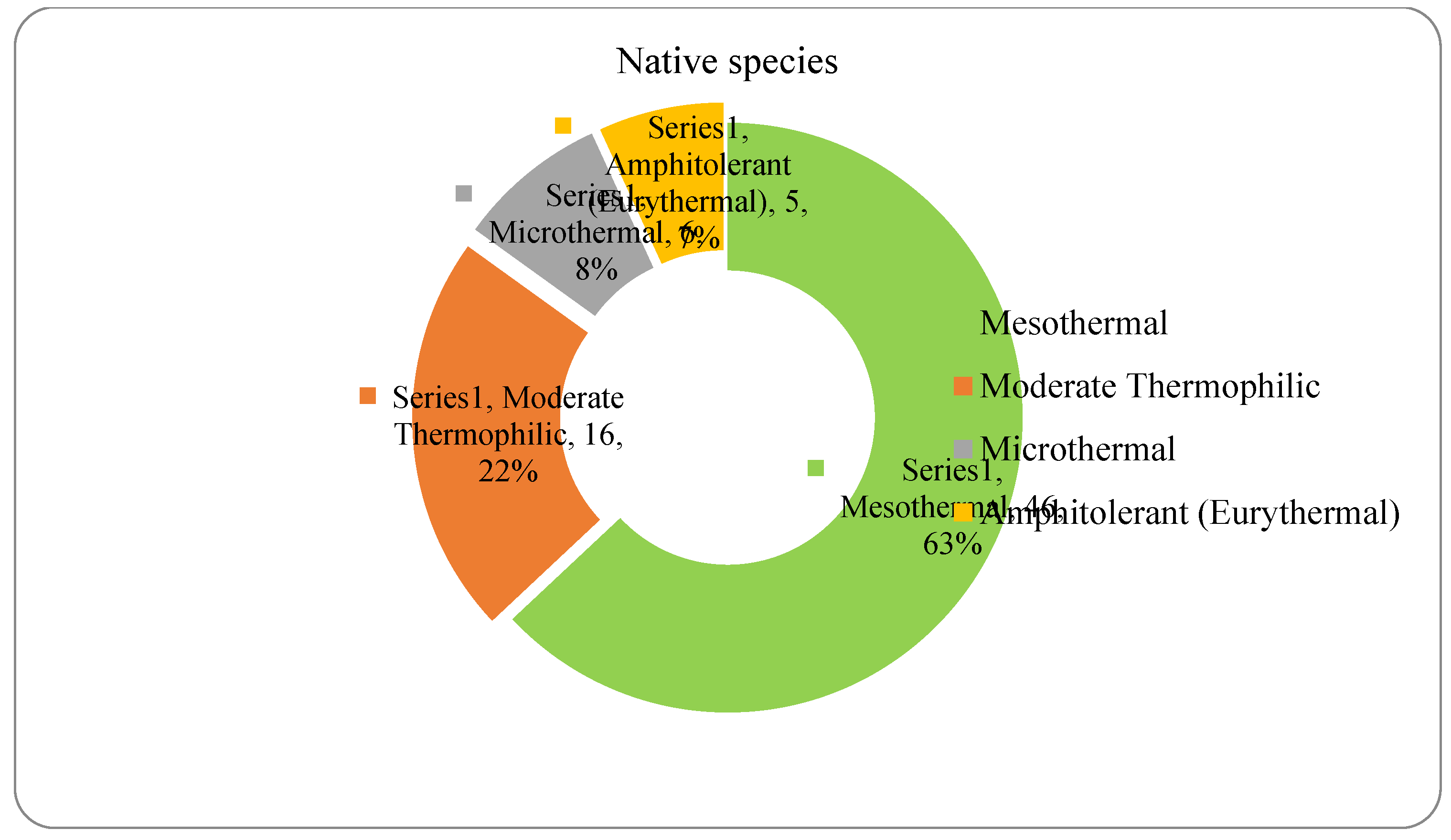
Figure 7.
The spectrum of species requirements for temperature in the non-native dendroflora of the Botanic Park from Timișoara City.
Figure 7.
The spectrum of species requirements for temperature in the non-native dendroflora of the Botanic Park from Timișoara City.
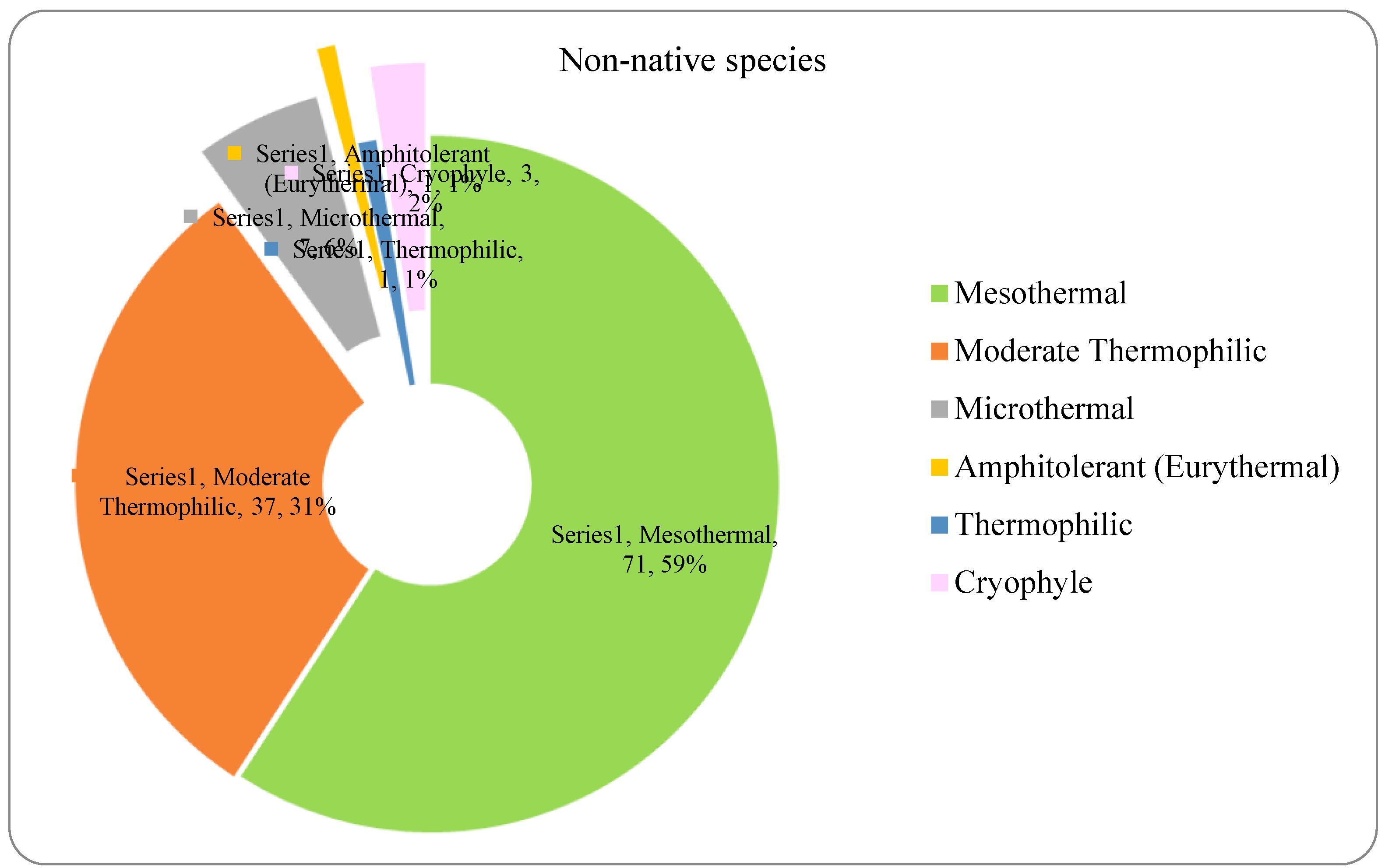
Figure 8.
The spectrum of species requirements for soil reaction (pH) in the native dendroflora of the Botanic Park from Timișoara City.
Figure 8.
The spectrum of species requirements for soil reaction (pH) in the native dendroflora of the Botanic Park from Timișoara City.
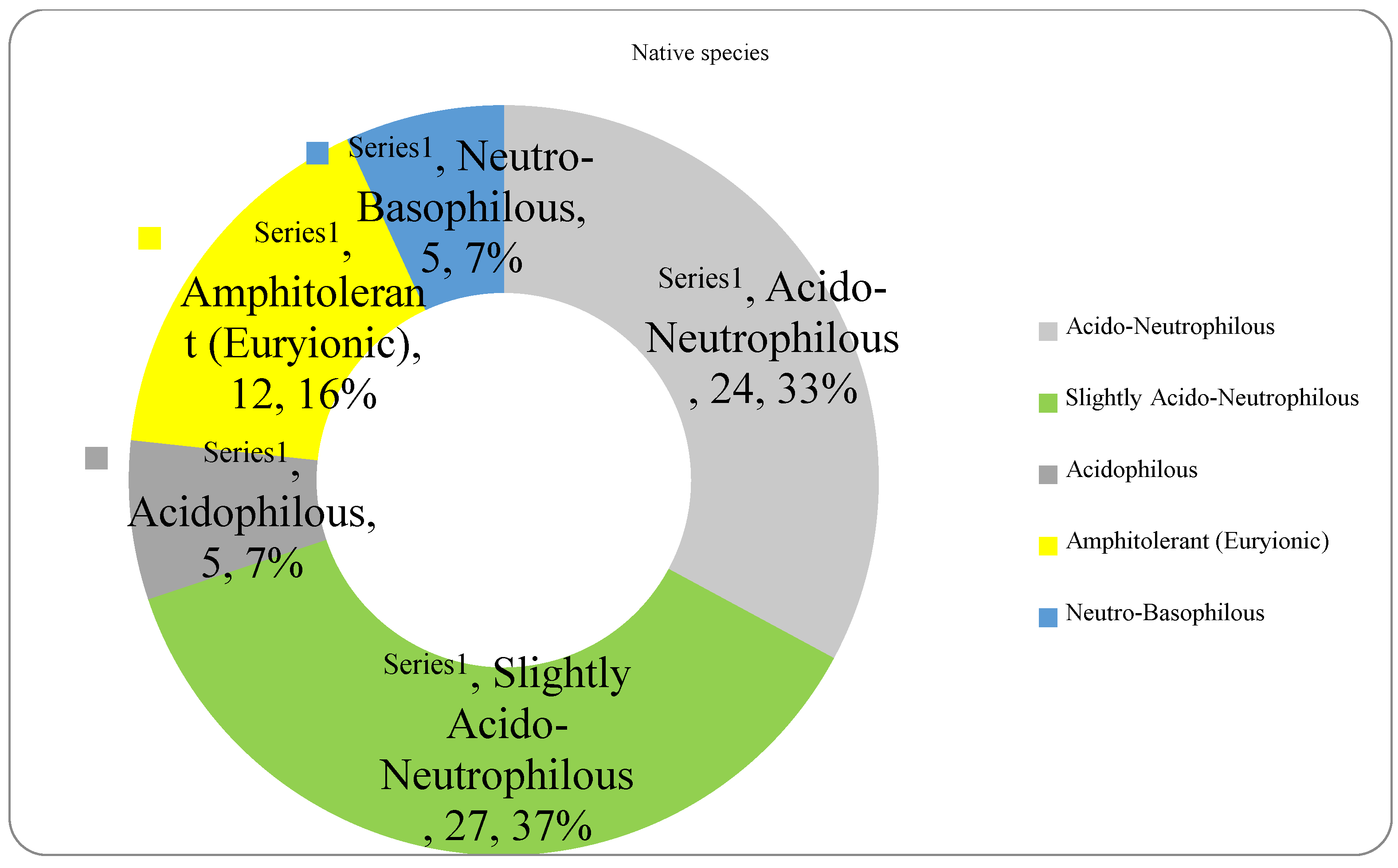
Figure 9.
The spectrum of species requirements for soil reaction (pH) in the non-native dendroflora of the Botanic Park from Timișoara City.
Figure 9.
The spectrum of species requirements for soil reaction (pH) in the non-native dendroflora of the Botanic Park from Timișoara City.
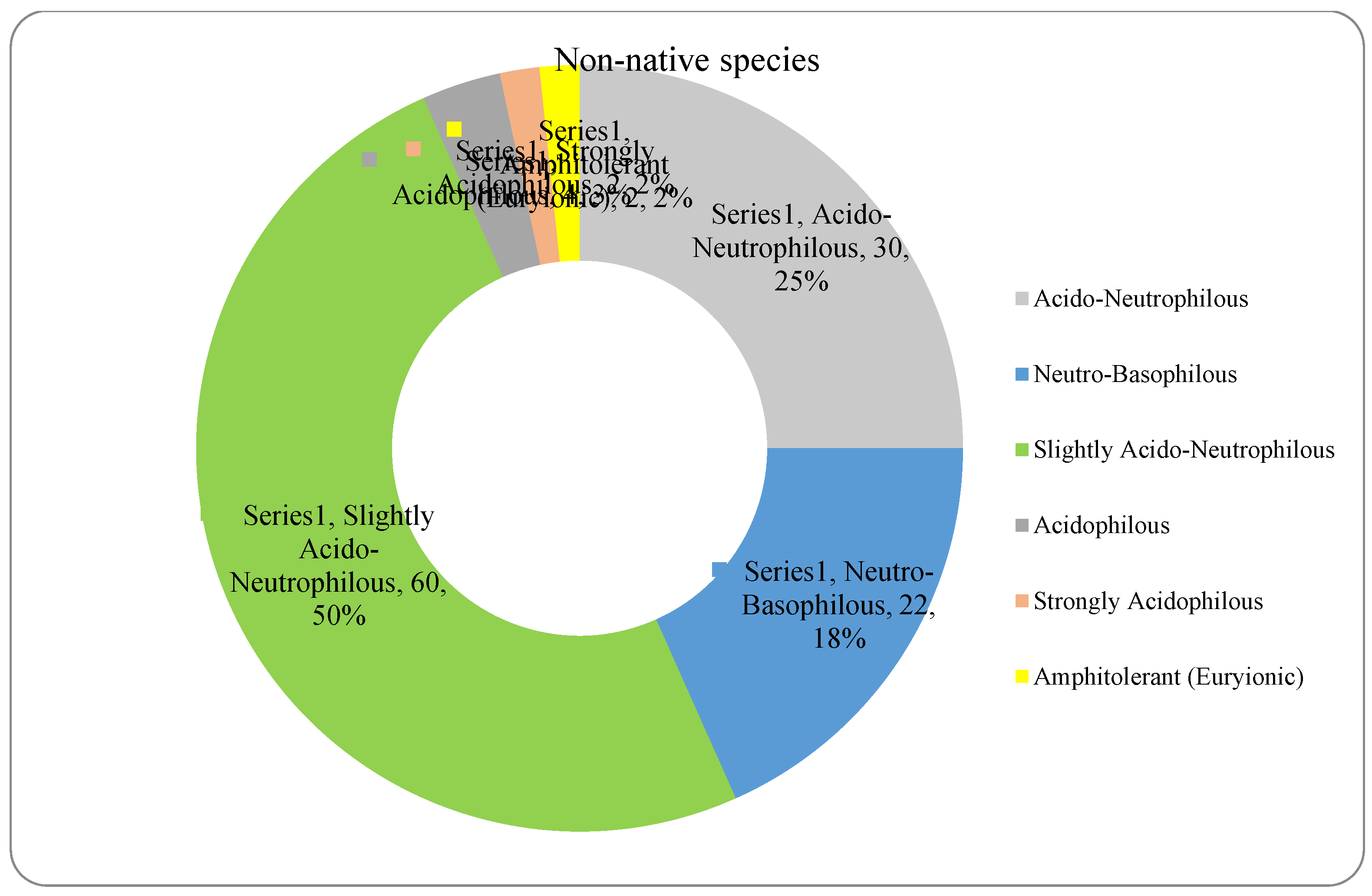
Figure 10.
Contingence diagram regarding the significant association (Chi-square test, χ2 (8, N=73) = 16.27, p = 0.039) and distribution of the soil pH-requirement among the plant life-form spectrum in the native woody species.
Figure 10.
Contingence diagram regarding the significant association (Chi-square test, χ2 (8, N=73) = 16.27, p = 0.039) and distribution of the soil pH-requirement among the plant life-form spectrum in the native woody species.
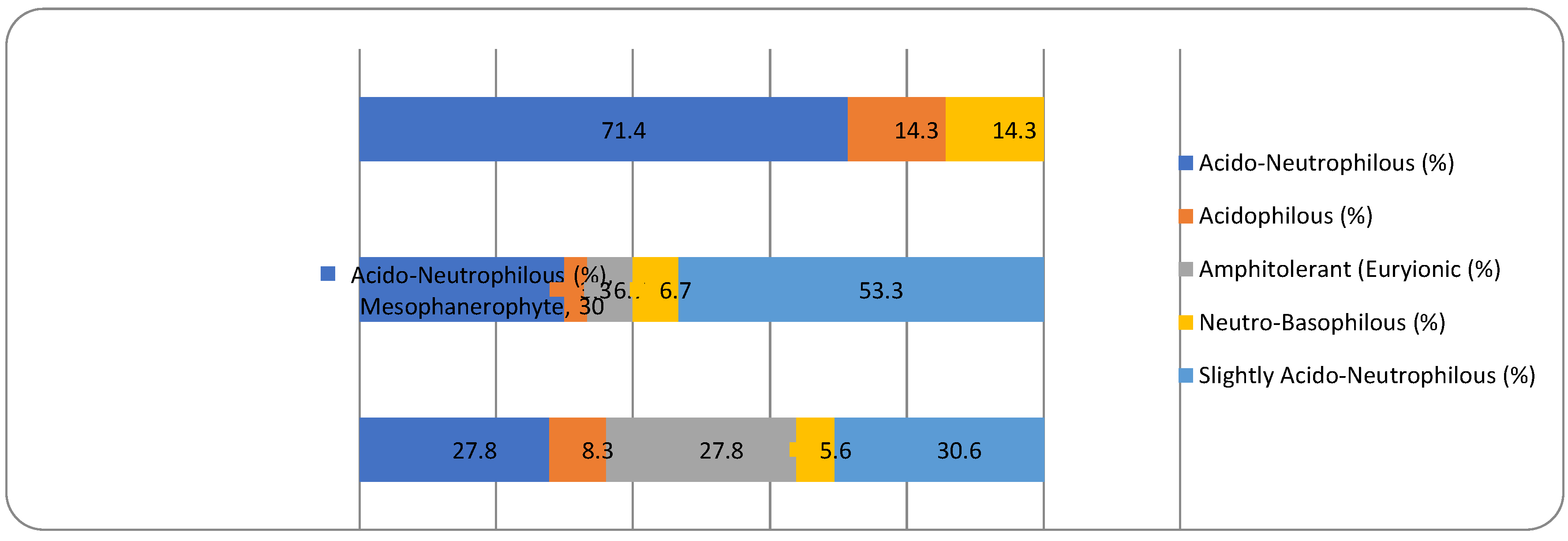
Figure 11.
Contingence diagram regarding the significant association (Chi-square test, χ2 (8, N=73) = 16.27, p = 0.039) and distribution of the plant life-forms among the soil pH-requirement spectrum in the native woody species.
Figure 11.
Contingence diagram regarding the significant association (Chi-square test, χ2 (8, N=73) = 16.27, p = 0.039) and distribution of the plant life-forms among the soil pH-requirement spectrum in the native woody species.
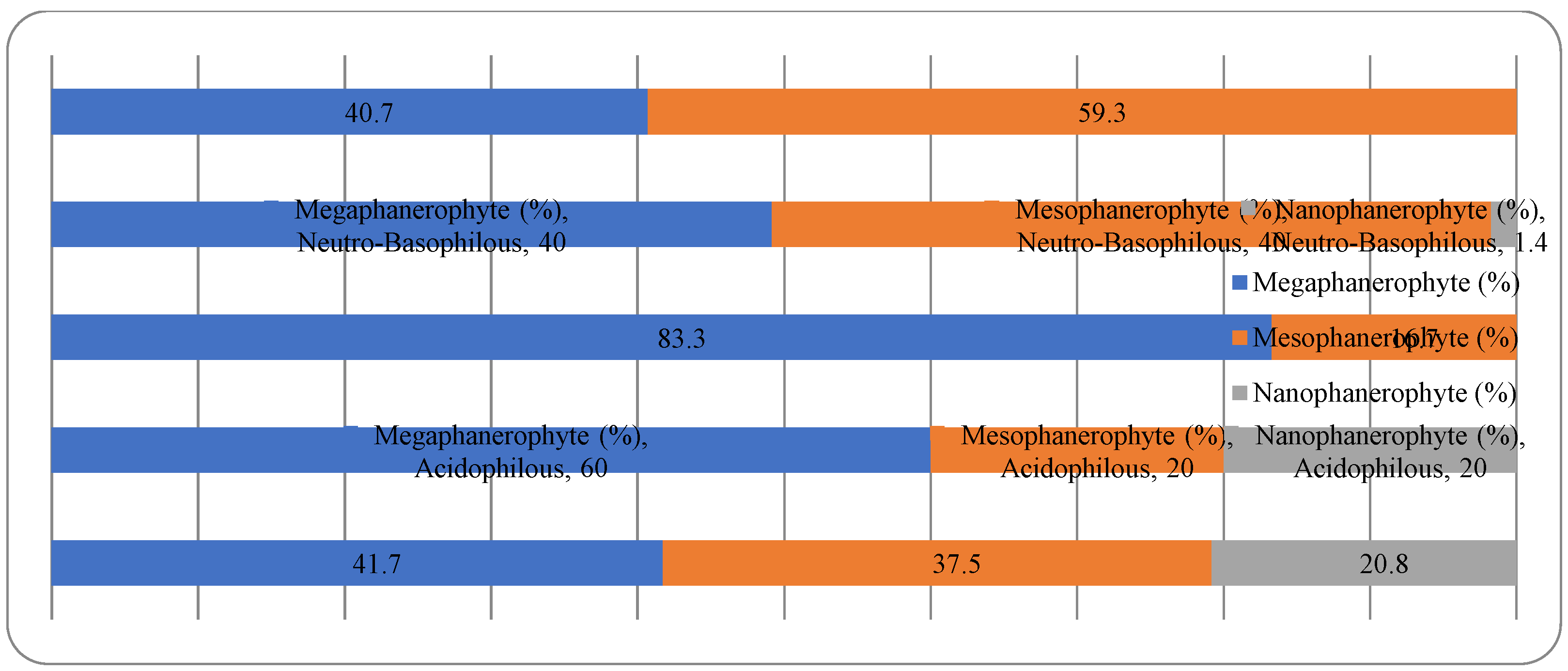
Figure 12.
Contingence diagram regarding the significant association (Chi-square test, χ2 (110, N=73) = 139.72, p = 0.029) and distribution of the plant moisture-requirements among the plant-families spectrum in the native woody species.
Figure 12.
Contingence diagram regarding the significant association (Chi-square test, χ2 (110, N=73) = 139.72, p = 0.029) and distribution of the plant moisture-requirements among the plant-families spectrum in the native woody species.

Figure 13.
Contingence diagram regarding the significant association (Chi-square test, χ2 (110, N=73) = 139.72, p = 0.029) and distribution of the plant-families among the plant moisture-requirement spectrum in the native woody species.
Figure 13.
Contingence diagram regarding the significant association (Chi-square test, χ2 (110, N=73) = 139.72, p = 0.029) and distribution of the plant-families among the plant moisture-requirement spectrum in the native woody species.

Figure 14.
Contingence diagram regarding the significant association (Chi-square test, χ2 (10, N=120) = 19.36, p = 0.036) and distribution of the plant temperature-requirement among the plant life-form spectrum in the non-native woody species.
Figure 14.
Contingence diagram regarding the significant association (Chi-square test, χ2 (10, N=120) = 19.36, p = 0.036) and distribution of the plant temperature-requirement among the plant life-form spectrum in the non-native woody species.
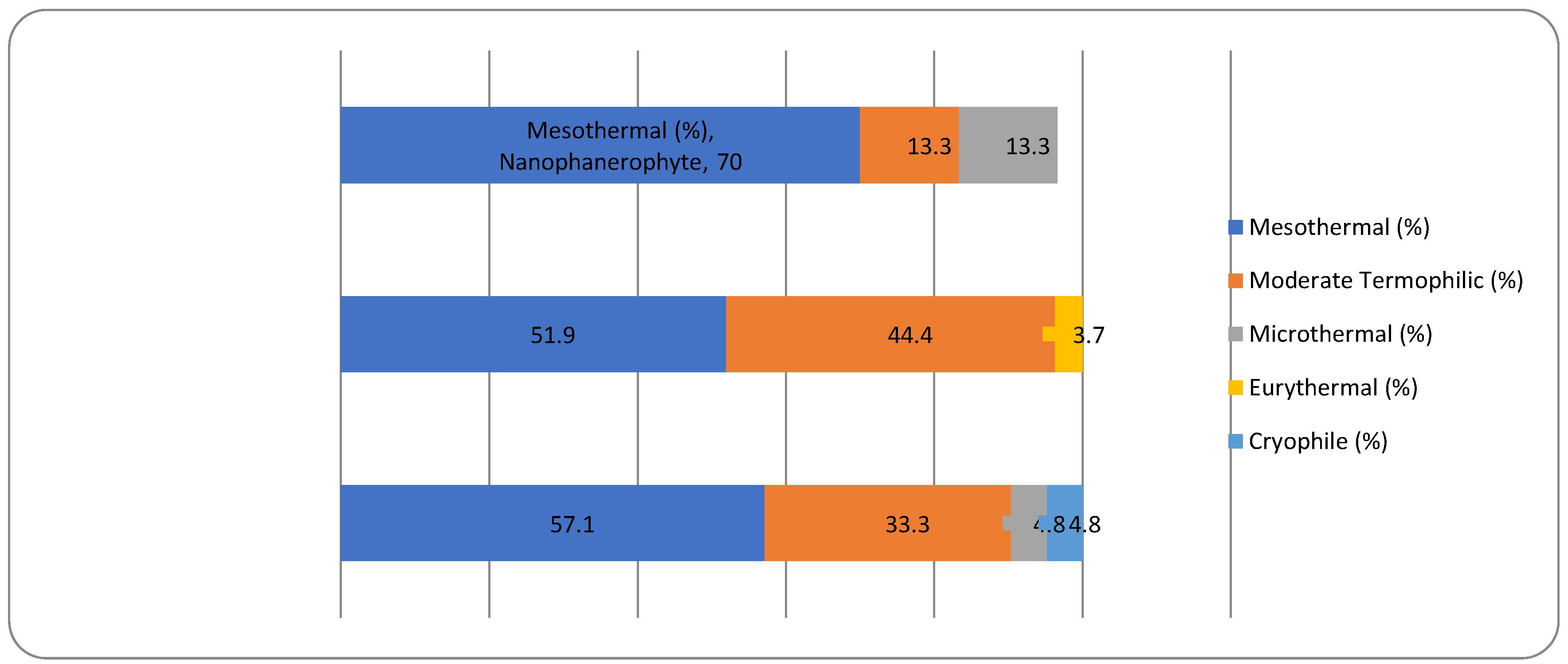
Figure 15.
Contingence diagram regarding the significant association (Chi-square test, χ2 (10, N=120) = 19.36, p = 0.036) and distribution of the life-forms among the plant temperature-requirement spectrum in the non-native woody species.
Figure 15.
Contingence diagram regarding the significant association (Chi-square test, χ2 (10, N=120) = 19.36, p = 0.036) and distribution of the life-forms among the plant temperature-requirement spectrum in the non-native woody species.
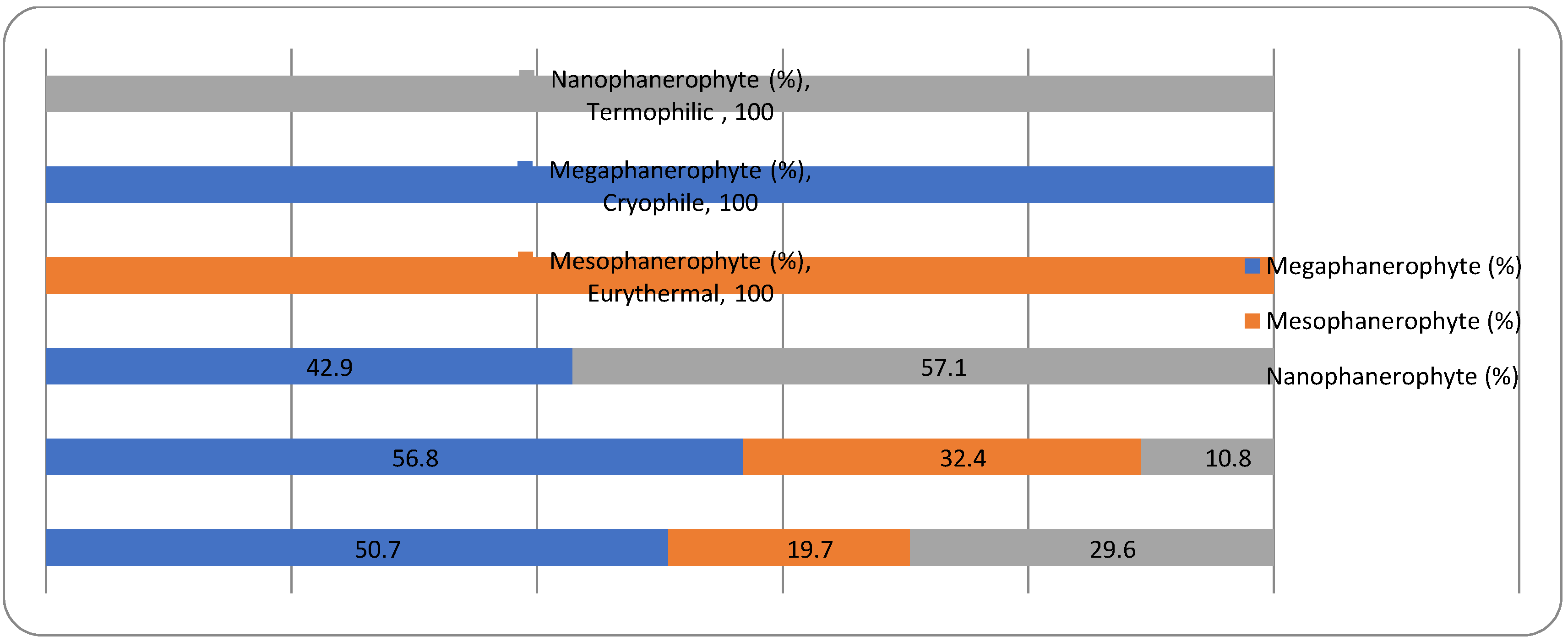

Table 1.
The significance of the ecological requirements of plant species for the factors moisture, temperature, and soil reaction (Sanda et al. 1983; Sanda et al. 2003).
Table 1.
The significance of the ecological requirements of plant species for the factors moisture, temperature, and soil reaction (Sanda et al. 1983; Sanda et al. 2003).
| Ecological preference index | Ecological significance description for moisture | Ecological significance description for temperature | Ecological significance description for soil reaction |
|---|---|---|---|
| 0 | Amphitolerant (Euryhydric) | Amphitolerant (Eurythermal) | Amphitolerant (Euryionic) |
| 1 - 1,5 | Xerophyte | Cryophile | Strongly Acidophilous |
| 2 - 2,5 | Xeromesophyte | Microthermal | Acidophilous |
| 3 - 3,5 | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 4 - 4,5 | Mesohygrophyte | Moderately Thermophilic | Slightly Acido-Neutrophilous |
| 5 - 5,5 | Hygrophyte | Thermophilic | Neutro-Basophilous |
| 6 | Hydrophyte | - | - |
Table 2.
Ecological traits of the native dendroflora of the Botanic Park of Timișoara City.
| No. | Species | Family | Monophyletic group | Plant chorology (phytogeographic elements) | Plant life-forms | Moisture requirement | Temperature requirement | Soil reaction requirement |
|---|---|---|---|---|---|---|---|---|
| 1 | Sambucus nigra | Adoxaceae | Angiosperm | European | Mesophanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 2 | Viburnum lantana | Adoxaceae | Angiosperm | Mediterranean - Central European | Mesophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 3 | Viburnum opulus | Adoxaceae | Angiosperm | Circumpolar | Mesophanerophyte | Mesohygrophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 4 | Cotinus coggygria | Anacardiaceae | Angiosperm | Mediterranean | Mesophanerophyte | Xeromesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 5 | Alnus glutinosa | Betulaceae | Angiosperm | Eurasian | Megaphanerophyte | Hygrophyte | Mesothermal | Acido-Neutrophilous |
| 6 | Alnus incana | Betulaceae | Angiosperm | Eurasian | Megaphanerophyte | Mesohygrophyte | Microthermal | Slightly Acido-Neutrophilous |
| 7 | Betula pendula | Betulaceae | Angiosperm | Eurasian | Megaphanerophyte | Mesophyte | Microthermal | Acidophilous |
| 8 | Carpinus betulus | Betulaceae | Angiosperm | European | Megaphanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 9 | Corylus avellana | Betulaceae | Angiosperm | European | Mesophanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 10 | Corylus colurna | Betulaceae | Angiosperm | Eurasian | Megaphanerophyte | Mesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 11 | Fagus sylvatica | Betulaceae | Angiosperm | European | Megaphanerophyte | Mesophyte | Mesothermal | Amphitolerant (Euryionic) |
| 12 | Buxus sempervirens | Buxaceae | Angiosperm | Eurasian | Nanophanerophyte | Xeromesophyte | Mesothermal | Acido-Neutrophilous |
| 13 | Lonicera xylosteum | Caprifoliaceae | Angiosperm | Eurasian | Mesophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 14 | Cornus alba | Cornaceae | Angiosperm | Eurasian | Nanophanerophyte | Mesohygrophyte | Moderate Thermophilic | Acido-Neutrophilous |
| 15 | Cornus mas | Cornaceae | Angiosperm | Pontic- Mediterranean - Central European | Mesophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 16 | Cornus sanguinea | Cornaceae | Angiosperm | Central European | Mesophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 17 | Hippophae rhamnoides | Elaeagnaceae | Angiosperm | Eurasian | Mesophanerophyte | Amphitolerant (Euryhydric) | Mesothermal | Slightly Acido-Neutrophilous |
| 18 | Amorpha fruticosa | Fabaceae | Angiosperm | Adventive | Mesophanerophyte | Mesophyte | Moderate Thermophilic | Amphitolerant (Euryionic) |
| 19 | Cercis siliquastrum | Fabaceae | Angiosperm | Eurasian | Mesophanerophyte | Xeromesophyte | Moderate Thermophilic | Acido-Neutrophilous |
| 20 | Laburnum anagyroides | Fabaceae | Angiosperm | Balkan | Mesophanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 21 | Robinia pseudoacacia | Fabaceae | Angiosperm | Adventive | Megaphanerophyte | Xeromesophyte | Moderate Thermophilic | Amphitolerant (Euryionic) |
| 22 | Sarothamnus scoparius | Fabaceae | Angiosperm | Atlantic- Mediterranean - Central European | Nanophanerophyte | Xeromesophyte | Mesothermal | Acidophilous |
| 23 | Castanea sativa | Fagaceae | Angiosperm | Mediterranean | Megaphanerophyte | Xeromesophyte | Moderate Thermophilic | Acidophilous |
| 24 | Quercus cerris | Fagaceae | Angiosperm | Mediterranean | Megaphanerophyte | Xeromesophyte | Mesothermal | Acido-Neutrophilous |
| 25 | Quercus macranthera | Fagaceae | Angiosperm | European- Anatolian- Caucasian | Megaphanerophyte | Mesophyte | Moderate Thermophilic | Neutro-Basophilous |
| 26 | Quercus petraea | Fagaceae | Angiosperm | European | Megaphanerophyte | Xeromesophyte | Mesothermal | Amphitolerant (Euryionic) |
| 27 | Quercus robur | Fagaceae | Angiosperm | European | Megaphanerophyte | Mesophyte | Mesothermal | Amphitolerant (Euryionic) |
| 28 | Ribes nigrum | Grossulariaceae | Angiosperm | Eurasian | Mesophanerophyte | Amphitolerant (Euryhydric) | Amphitolerant (Eurythermal) | Acido-Neutrophilous |
| 29 | Hypericum androsaemum | Hypericaceae | Angiosperm | Eurasian | Nanophanerophyte | Xeromesophyte | Moderate Thermophilic | Acido-Neutrophilous |
| 30 | Juglans regia | Juglandaceae | Angiosperm | Carpathian- Balkan- Anatolian- Caucasian | Megaphanerophyte | Mesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 31 | Tilia cordata | Malvaceae | Angiosperm | European | Megaphanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 32 | Tilia platyphyllos | Malvaceae | Angiosperm | Central European | Megaphanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 33 | Tilia tomentosa | Malvaceae | Angiosperm | Balkan | Megaphanerophyte | Xeromesophyte | Mesothermal | Acido-Neutrophilous |
| 34 | Morus alba | Moraceae | Angiosperm | Adventive | Megaphanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 35 | Fraxinus excelsior | Oleaceae | Angiosperm | European | Megaphanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 36 | Fraxinus ornus | Oleaceae | Angiosperm | Mediterranean | Mesophanerophyte | Xerophyte | Mesothermal | Neutro-Basophilous |
| 37 | Jasminum fruticans | Oleaceae | Angiosperm | Mediterranean | Mesophanerophyte | Xerophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 38 | Syringa vulgaris | Oleaceae | Angiosperm | Balkan- Anatolian | Mesophanerophyte | Xerophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 39 | Abies alba | Pinaceae | Angiosperm | Central European | Megaphanerophyte | Mesohygrophyte | Mesothermal | Amphitolerant (Euryionic) |
| 40 | Larix decidua | Pinaceae | Angiosperm | Central European- Carpathian-Sudetic | Megaphanerophyte | Xeromesophyte | Amphitolerant (Eurythermal) | Amphitolerant (Euryionic) |
| 41 | Picea abies | Pinaceae | Angiosperm | European | Megaphanerophyte | Amphitolerant (Euryhydric) | Amphitolerant (Eurythermal) | Amphitolerant (Euryionic) |
| 42 | Pinus mugo | Pinaceae | Angiosperm | European | Megaphanerophyte | Amphitolerant (Euryhydric) | Microthermal | Amphitolerant (Euryionic) |
| 43 | Pinus nigra | Pinaceae | Angiosperm | Carpathian | Megaphanerophyte | Xerophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 44 | Pinus sylvestris | Pinaceae | Gymnosperm | Eurasian | Megaphanerophyte | Amphitolerant (Euryhydric) | Amphitolerant (Eurythermal) | Amphitolerant (Euryionic) |
| 45 | Rhamnus cathartica | Rhamnaceae | Angiosperm | Eurasian | Mesophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 46 | Frangula rupestris | Rhamnaceae | Angiosperm | European | Nanophanerophyte | Mesohygrophyte | Mesothermal | Acido-Neutrophilous |
| 47 | Ziziphus jujuba | Rhamnaceae | Angiosperm | Mediterranean | Mesophanerophyte | Xerophyte | Moderate Thermophilic | Neutro-Basophilous |
| 48 | Cotoneaster integerrimus | Rosaceae | Angiosperm | Eurasian | Nanophanerophyte | Xeromesophyte | Mesothermal | Neutro-Basophilous |
| 49 | Crataegus laevigata | Rosaceae | Angiosperm | Central European | Mesophanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 50 | Crataegus monogyna | Rosaceae | Angiosperm | European | Mesophanerophyte | Xeromesophyte | Mesothermal | Acido-Neutrophilous |
| 51 | Crataegus pentagyn. | Rosaceae | Angiosperm | Mediterranean | Mesophanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 52 | Malus sylvestris | Rosaceae | Angiosperm | European | Mesophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 53 | Prunus avium | Rosaceae | Angiosperm | European | Mesophanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 54 | Prunus cerasifera | Rosaceae | Angiosperm | Eurasian | Mesophanerophyte | Xeromesophyte | Moderate Thermophilic | Amphitolerant (Euryionic) |
| 55 | Prunus padus | Rosaceae | Angiosperm | Eurasian | Megaphanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 56 | Pyrus pyraster | Rosaceae | Angiosperm | European | Mesophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 57 | Rosa canina | Rosaceae | Angiosperm | European | Nanophanerophyte | Xeromesophyte | Mesothermal | Acido-Neutrophilous |
| 58 | Sorbus aria | Rosaceae | Angiosperm | European | Megaphanerophyte | Mesophyte | Mesothermal | Neutro-Basophilous |
| 59 | Sorbus aucuparia | Rosaceae | Angiosperm | European | Megaphanerophyte | Mesophyte | Microthermal | Acidophilous |
| 60 | Sorbus torminalis | Rosaceae | Angiosperm | European | Megaphanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 61 | Spiraea salicifolia | Rosaceae | Angiosperm | Eurasian | Mesophanerophyte | Mesohygrophyte | Microthermal | Acidophilous |
| 62 | Salix viminalis | Salicaceae | Angiosperm | Eurasian | Mesophanerophyte | Hygrophyte | Microthermal | Slightly Acido-Neutrophilous |
| 63 | Populus alba | Salicaceae | Angiosperm | Eurasian | Megaphanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 64 | Acer campestre | Sapindaceae | Angiosperm | European | Megaphanerophyte | Xeromesophyte | Mesothermal | Acido-Neutrophilous |
| 65 | Acer monspessulanum | Sapindaceae | Angiosperm | Mediterranean | Megaphanerophyte | Xeromesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 66 | Acer platanoides | Sapindaceae | Angiosperm | Eurasian | Megaphanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 67 | Acer pseudoplatanus | Sapindaceae | Angiosperm | Central European | Megaphanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 68 | Acer tataricum | Sapindaceae | Angiosperm | European | Mesophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 69 | Ailanthus altissima | Sapindaceae | Angiosperm | Adventive | Megaphanerophyte | Amphitolerant (Euryhydric) | Amphitolerant (Eurythermal) | Amphitolerant (Euryionic) |
| 70 | Tamarix ramosissima | Tamaricaceae | Angiosperm | Eurasian | Mesophanerophyte | Amphitolerant (Euryhydric) | Mesothermal | Slightly Acido-Neutrophilous |
| 71 | Taxus baccata | Taxaceae | Gymnosperm | European | Mesophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 72 | Ulmus glabra | Ulmaceae | Angiosperm | Eurasian | Megaphanerophyte | Mesohygrophyte | Mesothermal | Acido-Neutrophilous |
| 73 | Ulmus minor | Ulmaceae | Angiosperm | Eurasian | Megaphanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
Table 3.
Ecological traits of the non-native dendroflora of the Botanic Park of Timișoara City.
| No. | Species | Family | Monophyletic group | Plant life-forms | Moisture requirement | Temperature requirement | Soil reaction requirement |
|---|---|---|---|---|---|---|---|
| 1 | Liquidambar styraciflua | Altingiaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 2 | Rhus semialata | Anacardiaceae | Angiosperm | Mesophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 3 | Rhus typhina | Anacardiaceae | Angiosperm | Mesophanerophyte | Xeromesophyte | Amphitolerant (Eurythermal) | Acido-Neutrophilous |
| 4 | Kalopanax septemlobus | Araliaceae | Angiosperm | Megaphanerophyte | Mesophyte | Moderate Thermophilic | Acido-Neutrophilous |
| 5 | Berberis julianae | Berberidaceae | Angiosperm | Nanophanerophyte | Xeromesophyte | Microthermal | Strongly Acidophilous |
| 6 | Berberis stenophylla | Berberidaceae | Angiosperm | Nanophanerophyte | Xeromesophyte | Microthermal | Strongly Acidophilous |
| 7 | Berberis thunbergii | Berberidaceae | Angiosperm | Nanophanerophyte | Xeromesophyte | Microthermal | Amphitolerant (Euryionic) |
| 8 | Berberishaoi | Berberidaceae | Angiosperm | Nanophanerophyte | Xeromesophyte | Microthermal | Acidophilous |
| 9 | Mahonia aquifolium | Berberidaceae | Angiosperm | Nanophanerophyte | Mesophyte | Mesothermal | Amphitolerant (Euryionic) |
| 10 | Catalpa bignonioides | Bignoniaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 11 | Catalpa ovata | Bignoniaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 12 | Kolkwitzia amabilis | Caprifoliaceae | Angiosperm | Mesophanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 13 | Lonicera fragrantissima | Caprifoliaceae | Angiosperm | Nanophanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 14 | Lonicera tatarica | Caprifoliaceae | Angiosperm | Nanophanerophyte | Mesophyte | Mesothermal | Acidophilous |
| 15 | Symphoricarpos albus | Caprifoliaceae | Angiosperm | Nanophanerophyte | Mesohygrophyte | Mesothermal | Acido-Neutrophilous |
| 16 | Weigela florida | Caprifoliaceae | Angiosperm | Nanophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 17 | Euonymus bungeanus | Celastraceae | Angiosperm | Nanophanerophyte | Mesophyte | Mesothermal | Neutro-Basophilous |
| 18 | Cercidiphyllum japonicum | Cercidiphyllaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Moderate Thermophilic | Acido-Neutrophilous |
| 19 | Chamaecyparis lawsoniana | Cupressaceae | Gymnosperm | Megaphanerophyte | Mesohygrophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 20 | Chamaecyparis pisifera | Cupressaceae | Gymnosperm | Megaphanerophyte | Mesohygrophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 21 | Cryptomeria japonica | Cupressaceae | Gymnosperm | Megaphanerophyte | Mesohygrophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 22 | Cupressus arizonica | Cupressaceae | Gymnosperm | Megaphanerophyte | Xeromesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 23 | Juniperus chinensis | Cupressaceae | Gymnosperm | Mesophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 24 | Juniperus horizontalis | Cupressaceae | Gymnosperm | Nanophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 25 | Juniperus virginiana | Cupressaceae | Gymnosperm | Mesophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 26 | Thuja occidentalis | Cupressaceae | Gymnosperm | Megaphanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 27 | Thuja occidentalis var. fastigiata | Cupressaceae | Gymnosperm | Megaphanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 28 | Thuja orientalis | Cupressaceae | Gymnosperm | Megaphanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 29 | Thuja plicata | Cupressaceae | Gymnosperm | Megaphanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 30 | Diospyros lotus | Ebenaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 31 | Albizia julibrissin | Fabaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Moderate Thermophilic | Amphitolerant (Euryionic) |
| 32 | Caragana arborescens | Fabaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 33 | Cercis chinensis | Fabaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Acidophilous |
| 34 | Gleditsia triacanthos | Fabaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Mesothermal | Neutro-Basophilous |
| 35 | Gleditsia triacanthos var. inermis | Fabaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Mesothermal | Neutro-Basophilous |
| 36 | Gymnocladus dioicus | Fabaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Neutro-Basophilous |
| 37 | Robinia hispida | Fabaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Neutro-Basophilous |
| 38 | Sophora japonica | Fabaceae | Angiosperm | Megaphanerophyte | Mesophyte | Moderate Thermophilic | Neutro-Basophilous |
| 39 | Quercus rubra | Fagaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 40 | Quercus macrocarpa | Fagaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Mesothermal | Neutro-Basophilous |
| 41 | Ginkgo biloba | Ginkgoaceae | Gymnosperm | Megaphanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 42 | Deutzia scabra | Hydrangeaceae | Angiosperm | Mesophanerophyte | Mesohygrophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 43 | Philadelphus coronarius | Hydrangeaceae | Angiosperm | Mesophanerophyte | Mesohygrophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 44 | Philadelphus wilsonii | Hydrangeaceae | Angiosperm | Mesophanerophyte | Mesohygrophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 45 | Hypericum patulum | Hypericaceae | Angiosperm | Mesophanerophyte | Mesohygrophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 46 | Carya ovata | Juglandaceae | Angiosperm | Megaphanerophyte | Mesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 47 | Juglans nigra | Juglandaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 48 | Pterocarya fraxinifolia | Juglandaceae | Angiosperm | Nanophanerophyte | Hygrophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 49 | Punica granatum | Lythraceae | Angiosperm | Nanophanerophyte | Xeromesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 50 | Liriodendron tulipifera | Magnoliaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 51 | Magnolia kobus | Magnoliaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Mesothermal | Acido-Neutrophilous |
| 52 | Hibiscus syriacus | Malvaceae | Angiosperm | Nanophanerophyte | Mesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 53 | Broussonetia papyrifera | Moraceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 54 | Ficus carica | Moraceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 55 | Maclura pomifera | Moraceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 56 | Morus nigra | Moraceae | Angiosperm | Megaphanerophyte | Mesophyte | Moderate Thermophilic | Neutro-Basophilous |
| 57 | Chionanthus retusus | Oleaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 58 | Forsythia × intermedia | Oleaceae | Angiosperm | Nanophanerophyte | Mesophyte | Mesothermal | Neutro-Basophilous |
| 59 | Fraxinus americana | Oleaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Neutro-Basophilous |
| 60 | Ligustrum ovalifolium | Oleaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 61 | Paeonia suffruticosa | Paeoniaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 62 | Paulownia tomentosa | Paulowniaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Moderate Thermophilic | Neutro-Basophilous |
| 63 | Abies concolor | Pinaceae | Gymnosperm | Megaphanerophyte | Mesohygrophyte | Microthermal | Slightly Acido-Neutrophilous |
| 64 | Abies pinsapo | Pinaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Cryophile | Acido-Neutrophilous |
| 65 | Picea pungens | Pinaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Cryophile | Acido-Neutrophilous |
| 66 | Pinus excelsa | Pinaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Microthermal | Slightly Acido-Neutrophilous |
| 67 | Pinus strobus | Pinaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 68 | Pinus wallichiana | Pinaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Microthermal | Slightly Acido-Neutrophilous |
| 69 | Pseudotsuga menziesii | Pinaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Mesothermal | Acido-Neutrophilous |
| 70 | Pseudotsuga menziesii var. glauca | Pinaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Mesothermal | Acido-Neutrophilous |
| 71 | Tsuga canadensis | Pinaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Cryophile | Acido-Neutrophilous |
| 72 | Platanus × acerifolia | Platanaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 73 | Phyllostachys aurea | Poaceae | Angiosperm | Mesophanerophyte | Hygrophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 74 | Frangula betulifolia | Rhamnaceae | Angiosperm | Mesophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 75 | Securinega suffruticosa | Rhamnaceae | Angiosperm | Nanophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 76 | Chaenomeles japonica | Rosaceae | Angiosperm | Nanophanerophyte | Mesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 77 | Cotoneaster bullatus | Rosaceae | Angiosperm | Nanophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 78 | Cotoneaster melanocarpus | Rosaceae | Angiosperm | Nanophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 79 | Crataegus multiflora | Rosaceae | Angiosperm | Mesophanerophyte | Mesohygrophyte | Moderate Thermophilic | Neutro-Basophilous |
| 80 | Crataegus phaenopyrum | Rosaceae | Angiosperm | Mesophanerophyte | Mesohygrophyte | Moderate Thermophilic | Neutro-Basophilous |
| 81 | Cydonia oblonga | Rosaceae | Angiosperm | Mesophanerophyte | Mesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 82 | Kerria japonica | Rosaceae | Angiosperm | Mesophanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 83 | Malus domestica | Rosaceae | Angiosperm | Megaphanerophyte | Amphitolerant (Euryhydric) | Mesothermal | Slightly Acido-Neutrophilous |
| 84 | Malus floribunda | Rosaceae | Angiosperm | Megaphanerophyte | Amphitolerant (Euryhydric) | Mesothermal | Slightly Acido-Neutrophilous |
| 85 | Prunus domestica | Rosaceae | Angiosperm | Megaphanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 86 | Prunus dulcis | Rosaceae | Angiosperm | Megaphanerophyte | Xerophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 87 | Prunus laurocerasus | Rosaceae | Angiosperm | Mesophanerophyte | Mesophyte | Moderate Thermophilic | Acido-Neutrophilous |
| 88 | Prunus serrulata | Rosaceae | Angiosperm | Mesophanerophyte | Mesophyte | Moderate Thermophilic | Acido-Neutrophilous |
| 89 | Prunus tomentosa | Rosaceae | Angiosperm | Mesophanerophyte | Mesophyte | Moderate Thermophilic | Acido-Neutrophilous |
| 90 | Pyracantha coccinea | Rosaceae | Angiosperm | Mesophanerophyte | Xeromesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 91 | Rhodotypos kerrioides | Rosaceae | Angiosperm | Nanophanerophyte | Mesophyte | Mesothermal | Acido-Neutrophilous |
| 92 | Rosa rugosa | Rosaceae | Angiosperm | Nanophanerophyte | Xeromesophyte | Mesothermal | Acido-Neutrophilous |
| 93 | Sorbaria sorbifolia | Rosaceae | Angiosperm | Nanophanerophyte | Mesohygrophyte | Mesothermal | Acido-Neutrophilous |
| 94 | Spiraea bumalda | Rosaceae | Angiosperm | Nanophanerophyte | Mesohygrophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 95 | Spiraea × vanhouttei | Rosaceae | Angiosperm | Nanophanerophyte | Mesohygrophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 96 | Phellodendron amurense | Rutaceae | Angiosperm | Megaphanerophyte | Amphitolerant (Euryhydric) | Mesothermal | Acido-Neutrophilous |
| 97 | Ptelea trifoliata | Rutaceae | Angiosperm | Mesophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 98 | Tetradium daniellii | Rutaceae | Angiosperm | Mesophanerophyte | Mesophyte | Moderate Thermophilic | Acido-Neutrophilous |
| 99 | Tetradium ruticarpum | Rutaceae | Angiosperm | Mesophanerophyte | Mesophyte | Moderate Thermophilic | Acido-Neutrophilous |
| 100 | Zanthoxylum piperitum | Rutaceae | Angiosperm | Nanophanerophyte | Xerophyte | Thermophilic | Slightly Acido-Neutrophilous |
| 101 | Salix babylonica | Salicaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 102 | Salix matsudana | Salicaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Mesothermal | Neutro-Basophilous |
| 103 | Acer ginnala | Sapindaceae | Angiosperm | Mesophanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 104 | Acer laetum | Sapindaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 105 | Acer negundo | Sapindaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Mesothermal | Neutro-Basophilous |
| 106 | Acer palmatum | Sapindaceae | Angiosperm | Mesophanerophyte | Xeromesophyte | Moderate Thermophilic | Slightly Acido-Neutrophilous |
| 107 | Acer saccharinum | Sapindaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Mesothermal | Neutro-Basophilous |
| 108 | Aesculus hippocastanum | Sapindaceae | Angiosperm | Megaphanerophyte | Mesohygrophyte | Moderate Thermophilic | Neutro-Basophilous |
| 109 | Koelreuteria paniculata | Sapindaceae | Angiosperm | Megaphanerophyte | Xeromesophyte | Moderate Thermophilic | Neutro-Basophilous |
| 110 | Xanthoceras sorbifolium | Sapindaceae | Angiosperm | Mesophanerophyte | Mesophyte | Moderate Thermophilic | Neutro-Basophilous |
| 111 | Buddleja davidii | Scrophulariaceae | Angiosperm | Nanophanerophyte | Mesophyte | Moderate Thermophilic | Neutro-Basophilous |
| 112 | Lycium halimifolium | Solanaceae | Angiosperm | Nanophanerophyte | Xerophyte | Mesothermal | Neutro-Basophilous |
| 113 | Taxodium distichum | Taxodiaceae | Gymnosperm | Megaphanerophyte | Hygrophyte | Mesothermal | Acido-Neutrophilous |
| 114 | Taxus baccata var. globosa | Taxaceae | Gymnosperm | Nanophanerophyte | Xeromesophyte | Mesothermal | Acido-Neutrophilous |
| 115 | Celtis occidentalis | Ulmaceae | Angiosperm | Megaphanerophyte | Amphitolerant (Euryhydric) | Moderate Thermophilic | Neutro-Basophilous |
| 116 | Viburnum burejaeticum | Viburnaceae | Angiosperm | Nanophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 117 | Viburnum carlesii | Viburnaceae | Angiosperm | Nanophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 118 | Viburnum orientale | Viburnaceae | Angiosperm | Mesophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 119 | Viburnum rhytidophyllum | Viburnaceae | Angiosperm | Mesophanerophyte | Mesophyte | Mesothermal | Slightly Acido-Neutrophilous |
| 120 | Wisteria sinensis | Wisteriaceae | Angiosperm | Megaphanerophyte | Mesophyte | Moderate Thermophilic | Acido-Neutrophilous |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



