
Submitted:
08 February 2025
Posted:
10 February 2025
You are already at the latest version
Abstract
FK506-binding proteins (FKBPs) belong to the peptidyl-prolyl cis/trans isomerase (PPIase) Super-family and are involved in a wide range of biological processes including protein folding, hormone signalling, plant growth and stress responses. However, the FKBPs and their biological functions have not been identified in tea. In this study, we searched and identified 21 FKBP genes in the tea plant genome using the consensus model of the FK506 binding domain (PF00254). Their phylogeny, classification, structure, motifs, interactors and expression patterns were analysed. Comprehensive qRT-PCR analysis revealed different expression patterns of CsFKBPs in different tissues and in response to low-temperature. We performed a genome-wide analysis of the low-temperature expression pattern of this gene family and determined that overexpression of CsFKBP53 enhances low-temperature tolerance in Arabidopsis plants. Furthermore, the expression of CsFKBP53 differed significantly in different tea varieties. Our results suggest that the FKBP gene is highly conserved in plant species and that transgenic CsFKBP53 can improve low temperature resistance in plants.
Keywords:
Camellia sinensis
; FKBP gene family
; CsFKBP53
; function analysis
; low temperature
1. Introduction
Immunosuppressants are defined as receptors for immunosuppressive drugs such as cyclosporine A (CsA), tacrolimus (FK506) and rapamycin (sirolimus). In contrast, FK506-binding proteins (FKBPs) are one of two classes of immunophilin, the receptors for an immunosuppressive drugs CsA, and are clearly characterised by their peptidyl prolyl cis-trans isomerase (PPIase) activity, which catalyzes the rotation of the peptide bond before the proline residue between the cis and trans conformations [1,2,3]. All FKBPs contain one FK506 binding domain (FKBd/FKBP-C) consisting of approximately 110 amino acids at least, providing the active site for PPlase catalysis and the receptor site for proline and proline analogues [4].
As genome assembly techniques and bioinformatics have developed, FKBP family genes have been identified and characterized in different plants. But unfortunately, there has been limited attention to FKBP genes in tea plant. Some FKBP genes have been found to play an important role in abiotic stresses in different plants. ROF1 (AtFKBP62) and ROF2 (AtFKBP65) are reciprocal proteins that respond to heat stress by regulating the expression of heat stress proteins (sHSPs) associated with heat stress reparation. ROF1 binds to HSP90 through its TPR domain, which in turn binds to the heat-stimulated transcription factor HsfA2, and the ROF1-HSP90-HsfA2 complex induces the expression of sHSPs. ROF2 negatively regulates sHSP expression by binding to FKBd of ROF1 and disrupting the ROF1-HSP90-HsfA2 complex in the nucleus [5]. After knockdown the ROF1 gene, there was almost complete loss of resistance to heat stress in plants after heat stress and 2-3 days of recovery. The HsfA2 knockdown phenotype was similar to ROF1, but the ROF2 mutant and ROF1 overexpression plants gained greater heat tolerance compared to control [6,7]. AtFKBP20-1a was highly expressed in all tissues and caused a rapid increase in expression under both heat and drought stress, but AtFKBP20-1b expression only began to increase after 24h of stress treatment. Both were localized in the nucleus, while AtFKBP20-1b was also present in the cytoplasm [8]. AtFKBP15-1 and AtFKBP15-2, which are localized to the endoplasmic reticulum in Arabidopsis, are 70% homologous and both are induced by high temperature [9]. ZmFKBPs were involved in low temperature signaling in Zea Mays, and ZmFKBP12, ZmFKBP13, ZmFKBP15-3, ZmFKBP16-1 and ZmFKBP57 were rapidly and continuously repressed at low temperatures (4℃) [10], it was verified that some FKBP family members may have a positive response to low temperature induction.
The FKBP in plants and other species contains, in addition to the basic structural domain FKBd, other structural domains: the tridecapeptide repeat TPR (Tetratricopeptide repeat), the calmodulin-binding domains CaMBds (Calmodulin-binding domains), NPL and the trigger factor TIG. TPR develops an inverse parallel helical domain that is involved in the assembly of multi-protein complexes and can bind to the heat kinin HSP90. CaMBds are generally located at the C-terminus of plant bulk FKBP and can bind to the calmodulin CaM (Calmodulin), the C-terminal CaMBd can be regulated by Ca2+ ion signalling to activate its PPIase and molecular chaperone activity, suggesting that multi-structured plant FKBP may have similar calcium regulation mechanisms [11]. Research has, however, identified an NPL structural domain within the Drosophila FKBP gene family that binds to core histones to produce function in nucleosome assembly [12]. TIG is a class of multi-domain triggers associated with FKBPs, mainly consisting of the FKBD and C-terminal chaperone domains, with the central FKBD bound on both sides to the N-terminal ribosome, which has peptidyl prolyl cis-trans isomerase activity similar to that of FKBP-C [13,14,15], who is playing a role in protein export. Although it has been demonstrated that TIG can act as a molecular chaperone in prokaryotes interacting with nascent polypeptides in an open conformation, its role in plants is not yet clearly recognized, and there is speculation that it may be directly involved in plant chloroplast translation and protein folding [16,17]. It is indisputable that the multi-structural domain FKBP will also undertake more complicated and important functions due to its complex structure.
Tea is an important beverage plant with economic, health and cultural value [18]. Tea trees are more likely to grow in mountainous in China. Many believe that higher altitudes more likely produce the better product, but this can also lead to a lack of temperature and light during the growing period, making the tea more susceptible to cold and frost [19,20]. Meanwhile, the drop of temperature in spring when the buds are sprouting can also seriously affect the yield of tea plants. In conclusion, low temperature stress not only limits the geographical distribution of tea trees, but also reduces the yield and quality of tea [21], Screening for relevant genes that respond to low temperatures and promoting the molecular breeding of tea plants with excellent cold resistance traits to improve tea production has become an important issue. With the publication of genomic data for important tea tree varieties such as “Shuchazao” (Camellia Sinensis var. Assamica cv. Shuchazao, Shuchazao)and “Yunkang10” (C. Sinensis var.Assamica cv. Yunkang 10, Yunkang10) [22,23,24], makes it possible to further analyse genes related to biotic and abiotic stresses in tea plants. In this study, a key gene for low temperature stress, CsFKBP53, was screened using Yunkang10 as material, and its expression pattern and phenotypic traits of transgenic Arabidopsis were analysed to investigate the cold resistance function.
2. Results
2.1. Identification and Physical Properties Of CsFKBPs
In order to identify encoding FK506-binding proteins in Yunkang10 genome, we used FK506-binding sequence to identified, and the results retrieved from HMMER, PFAM, SMART and CD-Search. In conclusion, we identified 21 CsFKBPs in the database. We named the CsFKBPs according to their molecular weight and homology with the Arabidopsis and rice FKBP genes, based on the nomenclature rules of previous studies [25,26,27]. The gene name, gene ID, CDS length, number of free amino acids, molecular weight, theoretical PI, Subcellular Localizations and the number of FKBP - C domains are showed in Table 1. The FKBP genes in tea consist of 100-619 amino acids, with CDS lengths between 306 bp-2429 bp, molecular weights between 10.81 kDa-72.43 kDa and theoretical PI between 4.5 - 10.44. Among them, 57% of the CsFKBPs are basic. Subcellular localization predictions showed that they were located in the nucleus, chloroplast, cytoplasm, vesicles, peroxisomes and extracellular matrix, with 38% of the genes located in the nucleus and 33% in the chloroplast, indicating that CsFKBPs may function mostly in the nucleus and chloroplast.
The 21 CsFKBPs contain 6 different structural domains, of which 13 genes contain only one FKBP-C domain, and CsFKBP62 with two and CsFKBP47/72 with three (Table 1, Figure 1). In contrast to other species, the TPR domain is only found in CsFKBP42, and the position is similar with FKBPs in other species. The Tigger_C domain is only found in CsTIG, but it does not contain the FKBP_C domain. The NPL domain is found at the 5' end of CsFKBP33 and CsFKBP53. CsFKBP27, CsFKBP62 and CsFKBP72 contain a 3a0801s09 domain at the 3' end. Unfortunately, the function of this domain is not well explored at present.
2.2. Evolutions of FKBPs in Different Species
Based on the classification results of the evolutionary tree and the role of each subfamily member in other plants, we can predict the function of some of the CsFKBPs. For example, CsFKBPs in group B may play an important role in the resistance of tea plants to low temperature stress [10]. In order to explore the evolutionary relationships among tea plants and the FKBP genes of Arabidopsis and rice, we built a phylogenetic tree using the full-length amino acid sequences of the FKBPs, of which Arabidopsis and rice had 23 FKBPs (Figure 2). The results showed that the 73 FKBP genes were classified into five groups: A, B, C, D and E, based on structural domains and evolutionary relationships. 6 (28.5%) of the FKBP genes in Yunkang10 were classified in group B, the group containing the most FKBP genes in tea. groups A, C, D and E contained 5, 3, 4 and 6 CsFKBPs respectively. group A and B were placed on the same branch, which may indicate that the genes in these two groups are more closely related genetically.
2.3. Gene Structure and Conserved Motif Analysis of CsFKBPs
The combined analysis of conserved motifs of gene structure and the results of cluster analysis in this study showed that similar family members clustered together (Figure 3a). We identified 10 Motifs in 21 CsFKBPs by analysis in the MEME online software (Figure 3b/3d). The result showed that except for CsTIG, other CsFKBPs contain multiple motifs. motif1 and motif3 are present in every CsFKBP, presumably as the most conservative and basic motifs of the FKBP family. Interestingly, evolutionarily more closely related genes exhibit motif motifs containing similar motifs. Interestingly, the CsFKBPs in groups B, C, D and E showed to contain extremely similar Motif domains, except for CsTIG in group C. Gene structure information shows that members of the CsFKBP family all contain introns and exons, including at least 2 and up to 19 introns (Figure 3c). However, eight genes do not have an untranslated region (UTR). All genes contain multiple coding sequences (CDSs) except for CsFKBP17-2 which contains two CDSs.
2.4. Expression Patterns of CsFKBPs Under Different Cold Stresses
We analyzed the expression of 21 CsFKBPs in yunkang10 old leaf tissues by qRT-PCR, among which four members had no expression detected, CsFKBP11, CsFKBP12-1, CsFKBP33, and CsFKBP53a respectively (Figure 4a). The results showed that the expression trends of different members were variable. Genes that responded significantly to low temperature stress included CsFKBP16-3, CsFKBP42, and CsFKBP53, which showed an overall increasing trend in expression under more severe low temperature conditions, indicating that they may exercise important functions when subjected to low temperature stress. Among them, CsFKBP53 showed an increase in expression after both low temperature stresses and reached 2-fold of CK after 6 h and 3 days at 4℃ and even more than 4-fold after 3 days of treatment at 10℃. In addition, we compared the expression patterns of homologous genes in shuchazao and formed a heat map and found that their expression profiles were similar (Figure 4b).
2.5. Sequence and Phylogenetic Analyses and Sub-Cellular Localization of CsFKBP53
In order to investigate the evolutionary relationship between FKBP53 homologs in different species, we compared the reported FKBP53 sequences in Arabidopsis thaliana, rice, strawberry, peach, tomato, maize and tea(Figure 5a). The results show that the FKBP53s all contain an FKBP-C structural domain and multiple amino acid sites are conserved. In addition, we also used to analysed the affinities among CsFKBP53 and FKBP53 of other species, and found that CsFKBP53 was the closest to SlFKBP53, with a homology of 43.21 % (Figure 5b). The results of three-dimensional structure analysis showed that the three-dimensional structures of the seven proteins, CsFKBP53, AtFKBP53, FaFKBP53-1, FaFKBP53-2, SlFKBP53, OsFKBP53b, and PpFKBP53b, were all mainly composed of β-folding, and all of them showed a highly similar state (Figure 5c). These results may indicate that FKBP53 proteins in different species may have similar functions.
2.6. The Expression Pattern Analysis and Subcellular Localization of CsFKBP53
We analyzed the expression of CsFKBP53 in different tissues of Yunkang 10(YK10) and in old leaf tissues of different tea trees, including Yunkang 8(YK8), Yunkang 9(YK9), Zijuan(ZJ), Tieguanyin(TGY), Shilixiang(SLX), Nannuodayecha(NN), and Longjing 43(LJ43). The results are shown in Figure 6a/6b, which indicate that CsFKBP53 is the higher express in the tissues of roots and old leaves, and it has a high degree of tissue expression specificity. The expression was about 3 times higher than that in young stem and young leaf tissues, and about 5 times higher than that in flowers and buds; the highest expression was found in the highly cold-resistant varieties Shilixiang, Yunkang 8, and Yunkang 9, whereas no expression signals were detected in Tieguanyin. We also found that CsFKBP53 expression was up-regulated after 10 °C and 4°C treatments(Figure 6c/6d). During the 10℃ treatment, the difference between the 3H treatment and the 0H control was not obvious. However, the expression increased significantly after 24H. The expression pattern was also up-regulated in the 4℃ treatment relative to the control. During the 4℃ treatment, the expression peaked at 24 H, 48H and 72H, which proved the low-temperature correlation of CsFKBP53. CsFKBP53 was transformed into the onion lower epidermal tissues by constructing a CsFKBP53-GFP fusion protein transient expression vector, and the fusion protein was observed to be localized in the nucleus of the cells using laser confocal microscopy (Figure 6e). While the control showed green fluorescent signal in cell tissues such as nucleus and cells.
2.7. Transgenic CsFKBP53 Improves Low-Temperature Resistance of Arabidopsis
To further validate the cold-resistant function of CsFKBP53, it was transfected into wild-type Arabidopsis thaliana by heterologous expression, and a total of 9 overexpression lines were obtained and their expression was examined (Figure 7b). The results showed that no fluorescent signals were detected in the wild-type control (WT), whereas the expression of CsFKBP53 was highly expressed in the overexpression lines OE-1, OE-6 and OE-9. In addition, we cultured T2 generation seedlings and WT plants and placed them in a 4℃ incubator after one week of germination for 48 H of low-temperature stress treatment, while 10 WT plants cultured at 25℃ under ambient conditions were taken as the control, and the phenotypic changes in the growth process were observed and recorded (Figure 7a). The results showed that the phenotypes of the plants changed significantly after low-temperature treatment. Compared with 25 ℃ WT, the leaf margins of WT at 4 ℃ were curled, the colour of some leaves was purple, and the root development of plants was obviously hindered, whereas overexpression of Arabidopsis thaliana could still grow normally after low-temperature treatment, and the growth of root length was 25 ℃ WT>4 ℃ OE>4 ℃ WT, which seriously affected the root growth of wild-type Arabidopsis thaliana, and the root growth of transgenic plants was vigorous after low-temperature treatment (Figure 7c). The root growth of transgenic plants was vigorous after low temperature treatment.
The T3 generation transgenic plants were subjected to 4 ℃ low temperature treatment for 24 H to detect changes in the activities of POD, SOD and CAT indexes. The results showed that relative to 25 ℃ WT, POD activity (173.09-318.55), SOD activity (76.38-140.52) and CAT activity (132.44-296.04) were significantly increased in the three transgenic lines after 4 ℃ low-temperature treatment, and showed a more obvious increasing trend than WT (Figure 7d).
3. Discussion
In recent years, some researchers have identified the FKBP (FK506 binding protein) genes in Arabidopsis [28,29,30], rice [31,32,33], apple [25], strawberry [34], tomato [13], grape [35], peach [36] and maize [10] and found that they play important functions in plant growth and development as well as in abiotic stress response. They were found to play important functions in plant growth and development as well as in abiotic stress response. However, as an economically important plant, the CsFKBP family of genes in tea tree has not yet been reported. In the present study, bioinformatic analysis revealed that tea and peach had the same number of FKBP family members, who with 21 family members (Table 1). Whereas Arabidopsis, rice, strawberry and grape had 23, tomato and maize had 24, and apple had 28. The difference in the number of FKBP family members may indicate that the FKBP family genes have undergone genetic divergence during the evolution of different species. The differences in the number of FKBP family members may indicate that gene duplication or loss events have occurred during the evolution of different species. There is a wide range of variation in the size of plant FKBP family members, for example, Arabidopsis AtFKBP12 is a single domain (SD) FKBP consisting of only one FKBd, whereas wheat TaFKBP77 is an FKBP with multiple structural domains (Multidomain, MD) [37]. In this study, the length of the 21 CsFKBPs CDS sequences ranged from 306 bp (CsFKBP11) to 2429 bp (CsFKBP53a), encoding amino acid sequences ranging from 100 aa to 619 aa, with a large variability among the different CsFKBP proteins.
Moreover, in addition to the FKPC-C domain, other structural domains have been identified in the FKBP family: the Tetratricopeptide repeat (TPR), the calmodulin-binding domains (CaMds), Tigger-C and NPL. These structural domains have complex functions and may be involved in protein complex assembly, stimulating other structural domains, promoting chloroplast translation and participating in protein folding. The results of this study show that tea tree CsFKBP protein is similar to other species in that they contains FKBP-C, TPR and TIG structural domains (Figure 1). Seventeen of these proteins contained one FKBP-C domain and three contained multiple FKBP-C domains. However, the SlpA and 3a0801s09 domains in CsFKBP proteins have not been reported in other species, and their specific functions need to be further investigated. Interestingly, Motif 3 is present in all FKBP members except CsTIG (Figure 3b). This indicates that Motif 3 corresponds to the FKBP-C structural domain, which is the most conserved segment of the FKBP-C structural domain of CsFKBP proteins. To investigate the clustering of FKBP proteins in tea, Arabidopsis and rice, a phylogenetic tree was constructed based on the amino acid sequences of FKBP proteins from each species. 73 proteins were divided into five groups, with CsFKBP proteins having the most distributed members in GroupB (Figure 2/3a). The 21 CsFKBP proteins were also clustered and found to be classified into seven groups. CsFKBP with similar clusters had similar gene structures, conserved structural domains, and this result was similar to the clustering results of Arabidopsis FKBP proteins [30].
10℃ is the limiting temperature at which tea plants are subjected to low temperature stress, while tea plants are already under low temperature stress when the temperature is below 4℃ [38,39]. Transcriptome analysis showed that the CsFKBP family of genes in tea tree showed a more pronounced expression trend under low temperature stress conditions. We also examined the expression of CsFKBPs at different periods of 4℃ treatment and 10℃ treatment by qRT-PCR (Figure 4a). The results showed that these genes showed different trends with increasing time of low temperature treatment, with the most obvious trend of increasing CsFKBP53 expression. Further analysis of the spatial and temporal expression patterns of CsFKBP53 under low temperature stress showed that CsFKBP53 expression began to show an increasing trend after 3 H treatment at 10℃, and reached the highest expression at 24 H and 72 H. The response of CsFKBP53 to low temperature was more rapid at 4℃ treatment, and the expression reached a peak at 6 H treatment. The expression of CsFKBP53 reached the peak at 6 H and then began to decrease, but the expression increased to about 2.5 times that of the control group at 48 H (Figure 4b). In conclusion, CsFKBP53 showed a positive trend in response to low temperature stress, and its expression increased more positively at lower growth temperatures.
Zhao [40] found that the low temperature stress response gene CsWRKY37 was most highly expressed in the roots of tea plants. In contrast, CsFKBP53 also showed higher expression in more lignified roots in the tissue expression pattern analysis of Yunkang10. The results of the tissue expression pattern analysis may prove that the low temperature stress-related genes are more biased to exercise their functions in tea tree roots that are less damaged under low temperature conditions (Figure 6a). The results of Wang [41] et al. found that tea tree varieties with common leaf colour were cold resistant than purple leaf varieties, with Longjing 43 showing stronger cold resistance while Zijuan tea was less cold resistant. Yunkang species varieties is a national breed selected by the Yunnan Agricultural Institute through asexual lines, and is generally highly resistant to cold and can be grown at an absolute temperature of -5 °C. The low temperature correlation was verified by the higher expression of CsFKBP53 in the old leaf tissue of the highly cold resistant tea tree varieties (Figure 6b). expression showed a positive correlation with the cold resistance of tea tree varieties.
Plants must regulate various physiological and biochemical processes in order to alleviate low temperature stress [42]. Low temperature stress reduces the growth rate of plants, which is reflected in the phenotypic effects of low temperature stress on leaf growth and root growth vigour [43]. In terms of the enzymatic activity of antioxidant enzymes, low temperature stress resulted in excessive accumulation of ROS, which in turn promoted the expression of a series of antioxidant enzyme genes and induced an increase in the content of catalase (CAT), peroxidase (POD) and superoxide dismutase (SOD) in the plant to eliminate ROS accumulation to avoid cell damage [44]. In this study, we successfully transferred CsFKBP53 into wild-type Arabidopsis and obtained three highly expressed transgenic lines (Figure 7d). The growth of the three transgenic lines under low temperature stress was significantly better than that of the wild-type control, as evidenced by longer root length and reduced leaf purplishness (Figure 7a). In addition, the results of POD, SOD and CAT assays showed that the activities of all three physiological indicators were significantly increased after the transgenic CsFKBP53 Arabidopsis plants were treated with low temperature at 4 ℃ for 24H. This indicated that the transgenic CsFKBP53 enhanced the activity of POD, SOD and CAT in Arabidopsis and improved the permeability of cell membranes, which ultimately led to the enhanced cold resistance of the transgenic plants.
4. Materials and Methods
4.1. Plant Materials and Stress Treatments
To evaluate the functions of FKBPs, Two-years-old “Yunkang10” were used to plant material, who is from horticulture greenhouse in Yunnan University (Yunnan, China). Different tea plant organs, including shoots, roots, stems, young leaves, old leaves, flowers, and buds were collected, immediately frozen in liquid nitrogen immediately and stored at −80°C. Collected “Yunkang8” (C. Sinensis var .Assamica cv. Yunkang8, Yunkang8), "Yunkang9"(C. Sinensis var. Assamica cv. Yunkang9, Yunkang9), "Longjin43" (C. Sinensis var. Sinensis cv. Longjin43, Longjin43), "Tieguanyin", "Zijuan" (C. Sinensis var .Assamica cv. Zijuan, Zijuan), "Nannuo", and "Shilixiang" old leaf tissues were stored at -80°C. Arabidopsis seeds were grown in a greenhouse in a controlled environment for 16H per day (25 ± 3°C) and 8H at night (20 ± 3°C) at 70% relative humidity. Take a batch of YK10 tea plant, part of which were treated at low temperature (4°C/10°C) in a low-temperature incubator, and the rest were used as controls. Collect at different times (0, 3, 6, 9, 12, 24, 48, and 72H), immediately freeze in liquid nitrogen, and then store it at −80℃ for RNA extraction. N. benthamiana used in this study was grown in growth chamber pots at 20°C at night and 24°C during the day, with a photoperiod set to 16 hours of light and 8 hours of dark.
4.2. Identification and Physical Properties of FKBP Genes in Tea
Genome sequencing and assembly of tea plant Yunkang10 were completed, the protein and CDS sequences were published in TPIA(https://tpdb.shengxin.ren/index.html) [45,46]. Hidden Markov Model profles in FK506-binding domain was downloaded from the Pfam database. Sequence information for the AtFKBP and OsFKBP genes was obtained from the TAIR(https://www.arabidopsis.org/) and RGAP websites, respectively. The hidden Markov model profile in the FK506 binding domain (PF00254) was downloaded from the Pfam (https://pfam.xfam.org/) , The HMMER V3.3.2(http://hmmer.org/)was used to search through the amino acid sequences of the Yunkang10 genome (e value <1e-5) and the sequences were used as candidate sequences for the FKBP gene of tea [47]. The above sequences were used as candidates for the FKBP gene of tea tree, and the structural domains were verified using the SMART [48](http://smart.embl.de/), and the CD-Search database in NCBI(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi), respectively. Physicochemical properties such as isoelectric point, molecular weight and theoretical PI values of all members of the CsFKBP gene family were predicted using ExPASY(https://web.expasy.org/protparam/).
4.3. Evolution of CsFKBP Protein in Tea
We constructed an evolutionary tree for the Arabidopsis, Rice, Longjing43, Shuchazao, and Yunkang10 FKBP genes by the neighbor-joining (NJ) method (execution parameters: Poisson correction, pairwise deletion and bootstrap (1000 repetitions)) using MEGA 7.0 software [49].
4.4. Gene Structure and Motif Analysis
The exon/intron structure of each CsFKBP gene was analysed using GSDS2.0 (http://gsds.gao-lab.org/) to align the coding gene sequence with the corresponding CsFKBP gene sequences retrieved from the Yunkang10 genome [50]. Protein conserved motifs of CsFKBPs were visualized by setting 10 Motifs in the MEME online tool(https://meme-suite.org/meme/index.html). Gene structures and Chromosomal location were analysed using TBtools [51].
4.5. Total RNA Extraction and Expression Analysis
We extracted total RNA from frozen tea tree organs, including shoots, roots, young stems, new leaves, old leaves, flowers and buds, using a silica matrix column method reagent plant total RNA extraction kit (TIANGEN, Beijing, China) according to the instructions for use. Qrt-PCR reactions were performed using SYBR® Premix Ex Taq TM II (TakaRa, Shanghai, China) in a 20µL reaction system, using 1µL cDNA and 1µL primer, with CsACTIN as an internal reference control. qRT PCR reactions were programmed under the following conditions: 95°C pre denaturation for 30s, followed by denaturation at 95°C for 5s, annealing at 60°C for 34s and extension at 72°C for 15s, maintained for 40 cycles. Primers specific for the CsFKBPs and CsActin used for qRT PCR analysis are shown in Supplementary Table S1. three replicates of each PCR run were performed. Relative changes in gene expression were analysed according to the threshold cycle (Ct) using the 2-ΔΔCᴛ method previously reported [52]. The same method was used for the analysis of FKBP gene expression at different time periods under low temperature treatment conditions.
4.6. Gene Cloning and Vector Construction
The open reading frame (ORF) of CsFKBP53 was amplified using PrimeSTAR® Max DNA Polymerase (Takara, Shanghai, China) with an amplification program of denaturation at 95°C for 4 min, followed by 33 cycles of 98°C for 10s, 58°C for 15s and 72°C for 10s. After recovered it using Takara Mini BEST Agarose Gel DNA Extraction Kit Ver.4.0, and ligated it into the pEASY®-Blunt Cloning Vector (TransGen, Beijing, China). The sequencing was done at Sangon Biotechnology Co., Ltd(Kunming, China).
4.7. Protein Subcellular Localization Studies
We predicted the subcellular location of CsFKBP53 subcellular localization using WOLF PSORT(https://wolfpsort.hgc.jp/) [4]. The full-length sequence of CsFKBP53 with the terminator deleted was ligated into a transient expression vector containing green fluorescent protein (GFP) reporter gene to generate a 35S::CsFKBP53::GFP fusion vector. The recombinant plasmid and the control 35S::GFP were transformed into Agrobacterium tumefaciens GV3101. After adding kana (50mg/ml) and rifampicin (100mg/ml), incubate at 28°C for 24H and 200rpm. Centrifuge at 4000rpm for 10 minutes when the concentration to 1.0. The precipitate was resuspended with a suspension buffer containing 10 mM MgCl2, 10 mM MES and 100 µM acetosyringone (AS) and OD600 was then adjusted to 0.6-0.8. The resuspension was injected into the lower epidermis of well grown leaves of N. benthamiana. The GFP signal was observed using a laser confocal microscope (OLYMPUS, JAPAN) after 72 H of dark light incubation. The GFP channel signal was observed at 500-540nm while the DAPI and TD signals were at 650-750nm.
4.8. Overexpress and Functional Analysis of CsFKBP53 Under Low Temperature Treatment
The ORF of CsFKBP53 was ligated onto the overexpression vector P30-3MYC using double digestion, and the plasmid was extracted and transferred into Agrobacterium tumefaciens GV3101 using the freeze-thaw method. The transfer into Arabidopsis thaliana was done by flush infestation. In this study, four transgenic positive plants were screened for resistance to Kana on the P30-3MYC vector using Clumbia-0 (Col-0) as the wild type, followed by extraction of genomic DNA by PCR amplification using 35S and gene-specific primers and validation (Supplementary Table S2). We then analysed the expression levels of four transgenic strains overexpressing CsFKBP53 by qRT-PCR. The results showed that CsFKBP53 was more significantly expressed in the overexpression lines OE-1/6/9. We selected overexpression plants for further analysis and consulted a previous method where transgenic plants were exposed to -6°C for 2 H and then incubated at 25°C for 4d to record survival rates [53].
5. Conclusions
The FKBP family has been identified and functionally analysed in a variety of plants, but little research has been done on the CsFKBPs in tea. In this paper, 21 CsFKBPs were identified and their evolutionary relationships, gene structures, conserved structural domains and expression patterns under low-temperature treatment were analysed, and CsFKBP53, was screened for low-temperature stress. The CsFKBP53 gene enhanced the cold resistance of Arabidopsis plants. But the research on the genetic transformation system of tea tree is incomplete, the gene in tea plant can only be verified by transforming other plants. As the research on the transformation system of tea plant is gradually improved, the target gene can be transferred into tea tree tissues through plant tissue culture technology, which can further understand the function of CsFKBP53 gene, and then breed high quality tea plant varieties with remarkable cold resistance and excellent tea quality.
6. Patents
This section is not mandatory but may be added if there are patents resulting from the work reported in this manuscript.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Methodology, M.-H.X., J.T., C.-N.L., W.-Q. Z and Q.L.; Software, J.T; Data curation, M.-H.X. and Q.L.; Writing-original draft, M.-H.X., and J.T; Writing-review & editing, D.-D.L.; Project administration, D.-D.L. and F.Y.; Funding acquisition, D.-D.L and F.Y. All authors have read and agreed to the published version of the manuscript.All authors have read and agreed to the published version of the manuscript.
Funding
This study was financed by the Yunnan Provincial Key Research and Development Plan (202403AP140045), Open Fund of Yunnan Provincial Key Laboratory of Tea Science (2021YNCX002), the Special Project for Young Top Talents of the Ten Thousand People Programme of Yunnan Province (YNWR-QNBJ-2019-058), and the 15th Graduate Student Research and Innovation Program of Yunnan University (KC-23235573).
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Harding, M.W.; Galat, A.; Uehling, D.E.; Schreiber, S.L. , A receptor for the immuno-suppressant FK506 is a cis–trans peptidyl-prolyl isomerase. Nature 1989, 341, 758–760. [Google Scholar] [CrossRef] [PubMed]
- Siekierka, J.J.; Hung, S.H.Y.; Poe, M.; Lin, C.S.; Sigal, N.H. , A cytosolic binding protein for the immunosuppressant FK506 has peptidyl-prolyl isomerase activity but is distinct from cyclophilin. Nature 1989, 341, 755–757. [Google Scholar] [CrossRef] [PubMed]
- Fischer, G.; Bang, H.; Mech, C. , [Determination of enzymatic catalysis for the cis-trans-isomerization of peptide binding in proline-containing peptides]. Biomed Biochim Acta 1984, 43, 1101–11. [Google Scholar]
- Vasudevan, D.; Gopalan, G.; Kumar, A.; Garcia, V.J.; Luan, S.; Swaminathan, K. , Plant immunophilins: a review of their structure-function relationship. Biochimica et Biophysica Acta (BBA) - General Subjects 2015, 1850, 2145–2158. [Google Scholar] [CrossRef]
- Meiri, D.; Tazat, K.; Cohen-Peer, R.; Farchi-Pisanty, O.; Aviezer-Hagai, K.; Avni, A.; Breiman, A. , Involvement of Arabidopsis ROF2 (FKBP65) in thermotolerance. Plant Mol Biol 2010, 72, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Aviezer-Hagai, K.; Skovorodnikova, J.; Galigniana, M.; Farchi-Pisanty, O.; Maayan, E.; Bocovza, S.; Efrat, Y.; von Koskull-Döring, P.; Ohad, N.; Breiman, A. , Arabidopsis immunophilins ROF1 (AtFKBP62) and ROF2 (AtFKBP65) exhibit tissue specificity, are heat-stress induced, and bind HSP90. Plant Mol Biol 2007, 63, 237–55. [Google Scholar] [CrossRef]
- Busch, W.; Wunderlich, M.; Schoffl, F. , Identification of novel heat shock factor-dependent genes and biochemical pathways in Arabidopsis thaliana. Plant J 2005, 41, 1–14. [Google Scholar] [CrossRef]
- Ahn, J.C.; Kim, D.W.; You, Y.N.; Seok, M.S.; Park, J.M.; Hwang, H.; Kim, B.G.; Luan, S.; Park, H.S.; Cho, H.S. , Classification of rice (Oryza sativa L. Japonica nipponbare) immunophilins (FKBPs, CYPs) and expression patterns under water stress. BMC Plant Biol 2010, 10, 253. [Google Scholar] [CrossRef]
- Luan, S.; Kudla, J.; Gruissem, W.; Schreiber, S.L. , Molecular characterization of a FKBP-type immunophilin from higher plants. Proc Natl Acad Sci U S A 1996, 93, 6964–9. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, H.; Li, W.; Mu, C.; Zhang, F.; Wang, L.; Meng, Z. , Genome-wide analysis and environmental response profiling of the FK506-binding protein gene family in maize (Zea mays L.). Gene 2012, 498, 212–22. [Google Scholar] [CrossRef]
- Meiri, D.; Breiman, A. , Arabidopsis ROF1 (FKBP62) modulates thermotolerance by interacting with HSP90.1 and affecting the accumulation of HsfA2-regulated sHSPs. Plant J 2009, 59, 387–99. [Google Scholar] [CrossRef] [PubMed]
- Edlich-Muth, C.; Artero, J.B.; Callow, P.; Przewloka, M.R.; Watson, A.A.; Zhang, W.; Glover, D.M.; Debski, J.; Dadlez, M.; Round, A.R.; Forsyth, V.T.; Laue, E.D. , The pentameric nucleoplasmin fold is present in Drosophila FKBP39 and a large number of chromatin-related proteins. J Mol Biol 2015, 427, 1949–63. [Google Scholar] [CrossRef]
- Waseem, M.; Ahmad, F.; Habib, S.; Gao, Y.; Li, Z. , Genome-wide identification of FK506-binding domain protein gene family, its characterization, and expression analysis in tomato (Solanum lycopersicum L.). Gene 2018, 678, 143–154. [Google Scholar] [CrossRef]
- Romano, P.; Gray, J.; Horton, P.; Luan, S. , Plant immunophilins: functional versatility beyond protein maturation. New Phytol 2005, 166, 753–69. [Google Scholar] [CrossRef] [PubMed]
- Kramer, G.; Rutkowska, A.; Wegrzyn, R.D.; Patzelt, H.; Kurz, T.A.; Merz, F.; Rauch, T.; Vorderwülbecke, S.; Deuerling, E.; Bukau, B. , Functional dissection of Escherichia coli trigger factor: unraveling the function of individual domains. J Bacteriol 2004, 186, 3777–84. [Google Scholar] [CrossRef]
- Geisler, M.; Bailly, A. , Tête-à-tête: the function of FKBPs in plant development. Trends Plant Sci 2007, 12, 465–73. [Google Scholar] [CrossRef] [PubMed]
- Driessen, A.J.; Manting, E.H.; van der Does, C. , The structural basis of protein targeting and translocation in bacteria. Nat Struct Biol 2001, 8, 492–8. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.H.; Tong, W.; Wu, Q.; Wei, S.; Zhao, J.; Zhang, Z.Z.; Wei, C.L.; Wan, X.C. , Tea plant genomics: achievements, challenges and perspectives. Hortic Res 2020, 7, 7. [Google Scholar] [CrossRef]
- Han, W.; Li, X.; Ahammed, G.J. In Stress Physiology of Tea in the Face of Climate Change, Springer Singapore, 2018; 2018.
- Wang, M.; Yang, J.; Li, J.; Zhou, X.; Xiao, Y.; Liao, Y.; Tang, J.; Dong, F.; Zeng, L. , Effects of temperature and light on quality-related metabolites in tea [Camellia sinensis (L.) Kuntze] leaves. Food Res Int 2022, 161, 111882. [Google Scholar] [CrossRef]
- Peng, J.; Li, N.; Di, T.; Ding, C.; Li, X.; Wu, Y.; Hao, X.; Wang, Y.; Yang, Y.; Wang, X.; Wang, L. , The interaction of CsWRKY4 and CsOCP3 with CsICE1 regulates CsCBF1/3 and mediates stress response in tea plant (Camellia sinensis). Environmental and Experimental Botany 2022, 199, 104892. [Google Scholar] [CrossRef]
- Liu, M.-Y.; Burgos, A.; Ma, L.; Zhang, Q.; Tang, D.; Ruan, J. , Lipidomics analysis unravels the effect of nitrogen fertilization on lipid metabolism in tea plant (Camellia sinensis L.). BMC Plant Biology 2017, 17, 165. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.; Wei, C.; Yang, H.; Zhang, L.; Chen, Q.; Deng, W.; Wei, S.; Zhang, J.; Fang, C.; Ho, C.; Wan, X. , Transcriptomic and phytochemical analysis of the biosynthesis of characteristic constituents in tea (Camellia sinensis) compared with oil tea (Camellia oleifera). BMC Plant Biol 2015, 15, 190. [Google Scholar] [CrossRef]
- Xia, E.H.; Zhang, H.B.; Sheng, J.; Li, K.; Zhang, Q.J.; Kim, C.; Zhang, Y.; Liu, Y.; Zhu, T.; Li, W.; Huang, H.; Tong, Y.; Nan, H.; Shi, C.; Shi, C.; Jiang, J.J.; Mao, S.Y.; Jiao, J.Y.; Zhang, D.; Zhao, Y.; Zhao, Y.J.; Zhang, L.P.; Liu, Y.L.; Liu, B.Y.; Yu, Y.; Shao, S.F.; Ni, D.J.; Eichler, E.E.; Gao, L.Z. , The Tea Tree Genome Provides Insights into Tea Flavor and Independent Evolution of Caffeine Biosynthesis. Mol Plant 2017, 10, 866–877. [Google Scholar] [CrossRef]
- Dong, Q.; Mao, K.; Duan, D.; Zhao, S.; Wang, Y.; Wang, Q.; Huang, D.; Li, C.; Liu, C.; Gong, X.; Ma, F. , Genome-wide analyses of genes encoding FK506-binding proteins reveal their involvement in abiotic stress responses in apple. BMC Genomics 2018, 19, 707. [Google Scholar] [CrossRef]
- Gollan, P.J.; Bhave, M. , Genome-wide analysis of genes encoding FK506-binding proteins in rice. Plant Mol Biol 2010, 72, 1–16. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Li, L.; Luan, S. , Immunophilins and parvulins. Superfamily of peptidyl prolyl isomerases in Arabidopsis. Plant Physiol 2004, 134, 1248–67. [Google Scholar] [CrossRef] [PubMed]
- Aldape, R.A.; Futer, O.; DeCenzo, M.T.; Jarrett, B.P.; Murcko, M.A.; Livingston, D.J. , Charged surface residues of FKBP12 participate in formation of the FKBP12-FK506-calcineurin complex. J Biol Chem 1992, 267, 16029–32. [Google Scholar] [CrossRef]
- Harrar, Y.; Bellini, C.; Faure, J.D. , FKBPs: at the crossroads of folding and transduction. Trends Plant Sci 2001, 6, 426–31. [Google Scholar] [CrossRef]
- Thirumalaikumar, V.P.; Gorka, M.; Schulz, K.; Masclaux-Daubresse, C.; Sampathkumar, A.; Skirycz, A.; Vierstra, R.D.; Balazadeh, S. , Selective autophagy regulates heat stress memory in Arabidopsis by NBR1-mediated targeting of HSP90.1 and ROF1. Autophagy 2021, 17, 2184–2199. [Google Scholar] [CrossRef]
- Cheung, M.Y.; Auyeung, W.K.; Li, K.P.; Lam, H.M. , A Rice Immunophilin Homolog, OsFKBP12, Is a Negative Regulator of Both Biotic and Abiotic Stress Responses. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Gollan, P.J.; Bhave, M.; Aro, E.M. , The FKBP families of higher plants: Exploring the structures and functions of protein interaction specialists. FEBS Lett 2012, 586, 3539–47. [Google Scholar] [CrossRef]
- Nigam, N.; Singh, A.; Sahi, C.; Chandramouli, A.; Grover, A. , SUMO-conjugating enzyme (Sce) and FK506-binding protein (FKBP) encoding rice (Oryza sativa L.) genes: genome-wide analysis, expression studies and evidence for their involvement in abiotic stress response. Mol Genet Genomics 2008, 279, 371–83. [Google Scholar] [CrossRef]
- Leng, X.; Liu, D.; Zhao, M.; Sun, X.; Li, Y.; Mu, Q.; Zhu, X.; Li, P.; Fang, J. , Genome-wide identification and analysis of FK506-binding protein family gene family in strawberry (Fragaria × ananassa). Gene 2014, 534, 390–9. [Google Scholar] [CrossRef] [PubMed]
- Shangguan, L.; Kayesh, E.; Leng, X.; Sun, X.; Korir, N.K.; Mu, Q.; Fang, J. , Whole genome identification and analysis of FK506-binding protein family genes in grapevine (Vitis vinifera L.). Mol Biol Rep 2013, 40, 4015–31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Han, J.; Liu, D.; Wen, X.; Li, Y.; Tao, R.; Peng, Y.; Fang, J.; Wang, C. , Genome-wide identification and analysis of FK506-binding protein gene family in peach (Prunus persica). Gene 2014, 536, 416–24. [Google Scholar] [CrossRef] [PubMed]
- Kurek, I.; Aviezer, K.; Erel, N.; Herman, E.; Breiman, A. , The wheat peptidyl prolyl cis-trans-isomerase FKBP77 is heat induced and developmentally regulated. Plant Physiol 1999, 119, 693–704. [Google Scholar] [CrossRef]
- Lou, W.; Zhao, Y.; Huang, X.; Zhu, T.; Yang, M.; Deng, S.; Zhou, Z.; Zhang, Y.; Sun, Q.; Chen, S. , Frost risk assessment based on the frost-induced injury rate of tea buds: A case study of the Yuezhou Longjing tea production area, China. European Journal of Agronomy 2023, 147, 126839. [Google Scholar] [CrossRef]
- Hao, X.; Wang, B.; Wang, L.; Zeng, J.; Yang, Y.; Wang, X. , Comprehensive transcriptome analysis reveals common and specific genes and pathways involved in cold acclimation and cold stress in tea plant leaves. Scientia Horticulturae 2018, 240, 354–368. [Google Scholar] [CrossRef]
- Zhao, H.; Mallano, A.I.; Li, F.; Li, P.; Wu, Q.; Wang, Y.; Li, Y.; Ahmad, N.; Tong, W.; Li, Y.; Zhang, Z.; Xia, E. , Characterization of CsWRKY29 and CsWRKY37 transcription factors and their functional roles in cold tolerance of tea plant. Beverage Plant Research 2022, 2, 1–13. [Google Scholar] [CrossRef]
- Wang, W.-L.; Cui, X.; Wang, Y.-X.; Liu, Z.-W.; Zhuang, J. , Members of R2R3-type MYB transcription factors from subgroups 20 and 22 are involved in abiotic stress response in tea plants. Biotechnology & Biotechnological Equipment 2018, 32, 1141–1153. [Google Scholar]
- Guo, J.; Sun, B.; He, H.; Zhang, Y.; Tian, H.; Wang, B. , Current Understanding of bHLH Transcription Factors in Plant Abiotic Stress Tolerance. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Ranawake, A.L.; Manangkil, O.E.; Yoshida, S.; Ishii, T.; Mori, N.; Nakamura, C. , Mapping QTLs for cold tolerance at germination and the early seedling stage in rice (Oryza sativa L.). Biotechnol Biotechnol Equip 2014, 28, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Ou, W.; Lu, S.; Zhong, Q. , Differential responses of antioxidative system to chilling and drought in four rice cultivars differing in sensitivity. Plant Physiol Biochem 2006, 44, 828–36. [Google Scholar] [CrossRef]
- Xia, E.H.; Li, F.D.; Tong, W.; Li, P.H.; Wu, Q.; Zhao, H.J.; Ge, R.H.; Li, R.P.; Li, Y.Y.; Zhang, Z.Z.; Wei, C.L.; Wan, X.C. , Tea Plant Information Archive: a comprehensive genomics and bioinformatics platform for tea plant. Plant Biotechnol J 2019, 17, 1938–1953. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.; Tong, W.; Hou, Y.; An, Y.; Chen, L.; Wu, Q.; Liu, Y.; Yu, J.; Li, F.; Li, R.; Li, P.; Zhao, H.; Ge, R.; Huang, J.; Mallano, A.I.; Zhang, Y.; Liu, S.; Deng, W.; Song, C.; Zhang, Z.; Zhao, J.; Wei, S.; Zhang, Z.; Xia, T.; Wei, C.; Wan, X. , The Reference Genome of Tea Plant and Resequencing of 81 Diverse Accessions Provide Insights into Its Genome Evolution and Adaptation. Mol Plant 2020, 13, 1013–1026. [Google Scholar] [CrossRef]
- Wheeler, T.J.; Eddy, S.R. , nhmmer: DNA homology search with profile HMMs. Bioinformatics 2013, 29, 2487–9. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. , 20 years of the SMART protein domain annotation resource. Nucleic Acids Res 2018, 46, D493–d496. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. , MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol Biol Evol 2016, 33, 1870–4. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. , GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–7. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. , TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. , Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Mallano, A.I.; Li, F.; Li, P.; Wu, Q.; Wang, Y.; Li, Y.; Ahmad, N.; Tong, W.; Li, Y.; Zhang, Z.; Xia, E. , Characterization of <i>CsWRKY29</i> and <i>CsWRKY37</i> transcription factors and their functional roles in cold tolerance of tea plant. Beverage Plant Research 2022, 2, 1–13. [Google Scholar]
Figure 1.
Structural domain analysis of CsFKBPs. Functional domain distribution of 21 FKBPs in tea. We used the CD-Search program to identify the six conserved domains shown in the coloured boxes in the CsFKBP protein sequence. Blue box: FKBP-C domain; orange box: TPR domain; yellow box: NPL domain; green box: Slp A domain; purple box: Tigger_C domain; and red box:3a0801s09. The functional domains are arranged in order corresponding to their position in the individual protein sequence.
Figure 1.
Structural domain analysis of CsFKBPs. Functional domain distribution of 21 FKBPs in tea. We used the CD-Search program to identify the six conserved domains shown in the coloured boxes in the CsFKBP protein sequence. Blue box: FKBP-C domain; orange box: TPR domain; yellow box: NPL domain; green box: Slp A domain; purple box: Tigger_C domain; and red box:3a0801s09. The functional domains are arranged in order corresponding to their position in the individual protein sequence.
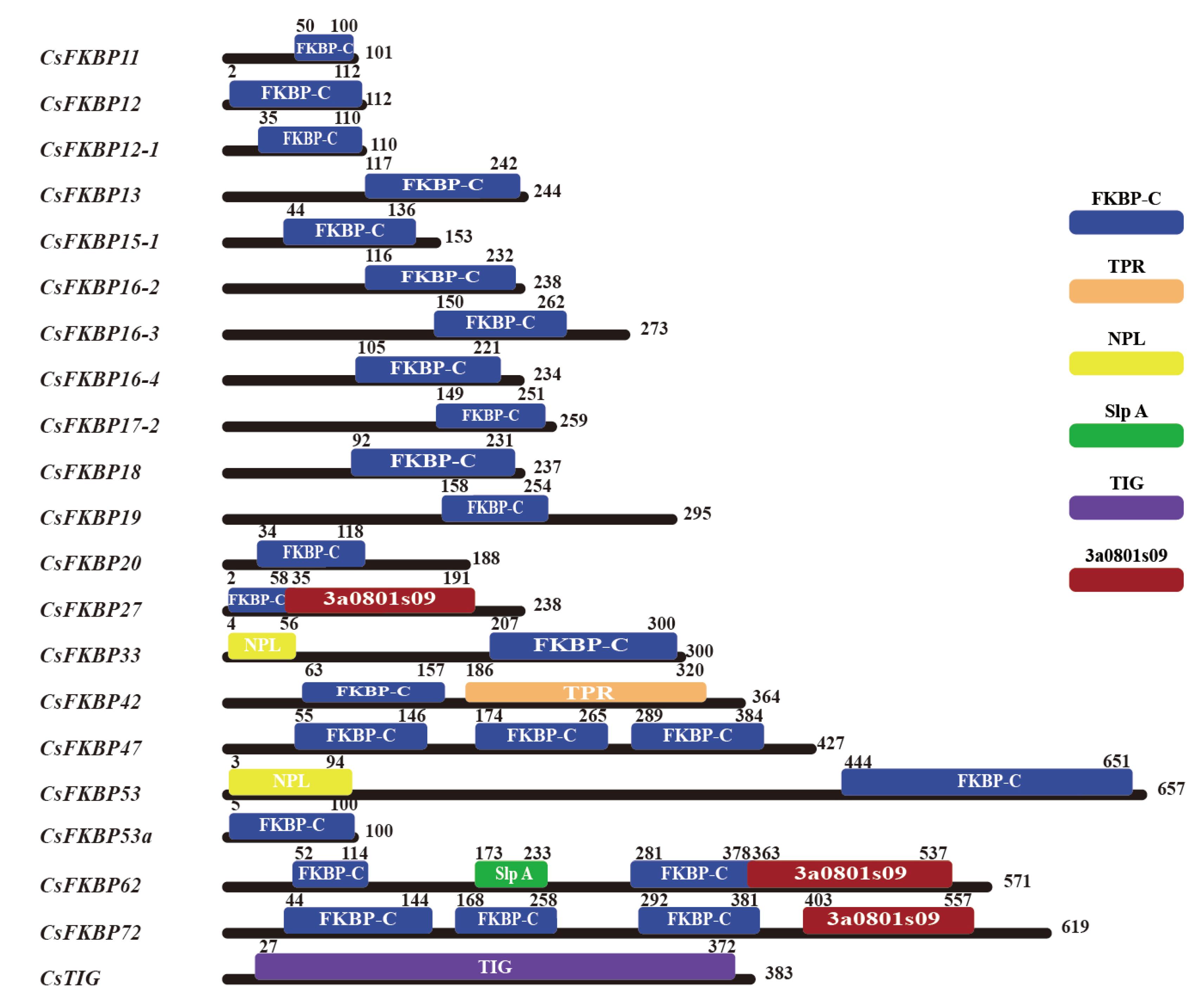
Figure 2.
Phylogenetic and evolutionary analysis of FKBPs. Phylogenetic trees were generated using the neighbour-joining method based on the full-length amino acid sequences of FKBP proteins in different species, including Yunkang10, Arabidopsis and rice. The different colors and symbols represent different species. Blue box: rice; orange circle: A.thaliana; green triangle: Yunkang10.
Figure 2.
Phylogenetic and evolutionary analysis of FKBPs. Phylogenetic trees were generated using the neighbour-joining method based on the full-length amino acid sequences of FKBP proteins in different species, including Yunkang10, Arabidopsis and rice. The different colors and symbols represent different species. Blue box: rice; orange circle: A.thaliana; green triangle: Yunkang10.
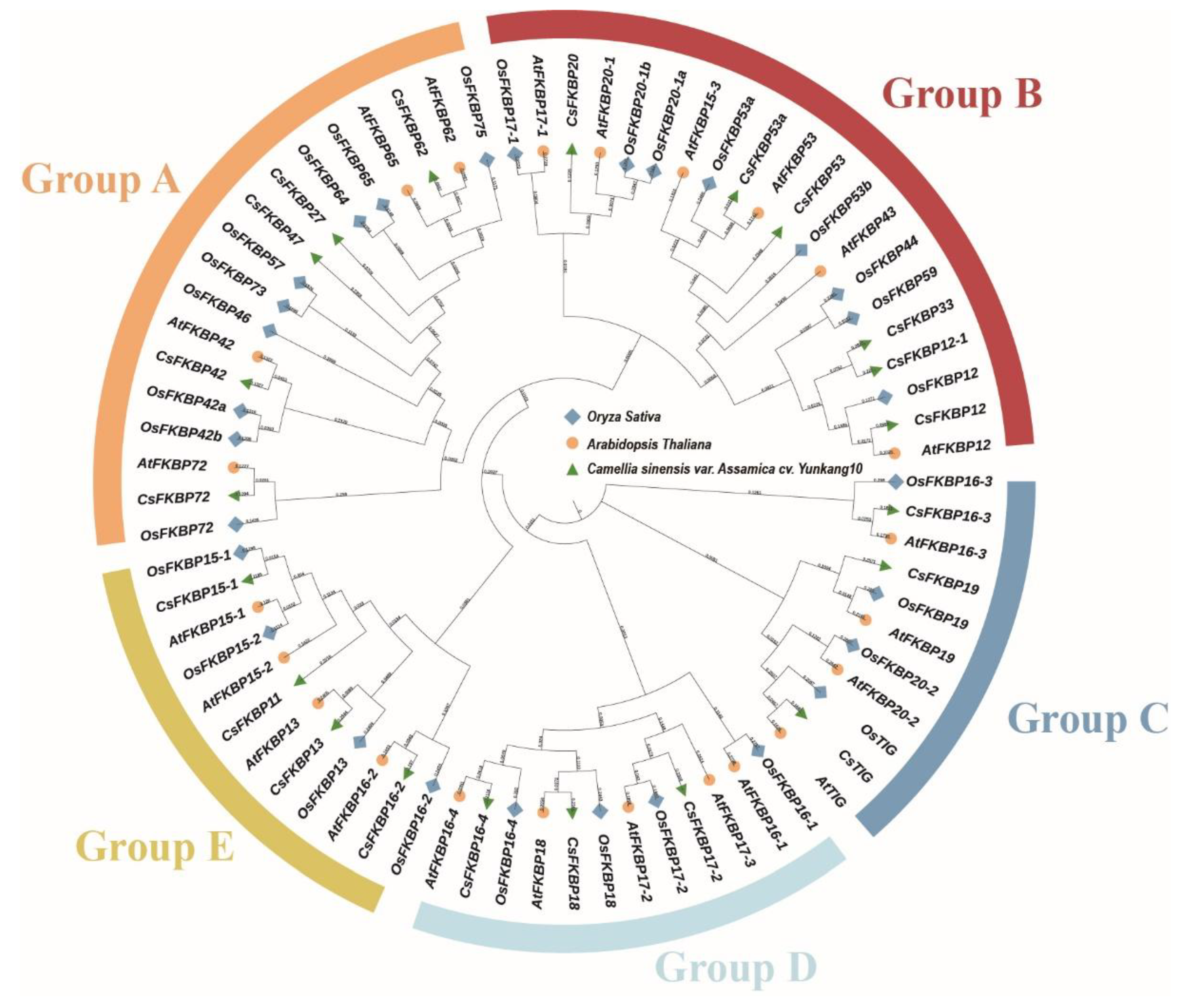
Figure 3.
Evolutionary tree, conserved motif distribution and gene structure of CsFKBP genes. (a) The evolutionary tree was built based on the 21 CsFKBPs protein sequences. (b) Conserved motifs of the members of the CsFKBP proteins. (c) The structure of CsFKBP proteins. Blue for CDS, orange for UTR, and straight lines for introns. (d) Conserved motifs of the CsFKBP protein in tea. The ordinate indicates amino acid conservation, The height of amino acid letters indicates the frequency of occurrence. The abscissa represents the position of the amino acid in the sequence.
Figure 3.
Evolutionary tree, conserved motif distribution and gene structure of CsFKBP genes. (a) The evolutionary tree was built based on the 21 CsFKBPs protein sequences. (b) Conserved motifs of the members of the CsFKBP proteins. (c) The structure of CsFKBP proteins. Blue for CDS, orange for UTR, and straight lines for introns. (d) Conserved motifs of the CsFKBP protein in tea. The ordinate indicates amino acid conservation, The height of amino acid letters indicates the frequency of occurrence. The abscissa represents the position of the amino acid in the sequence.
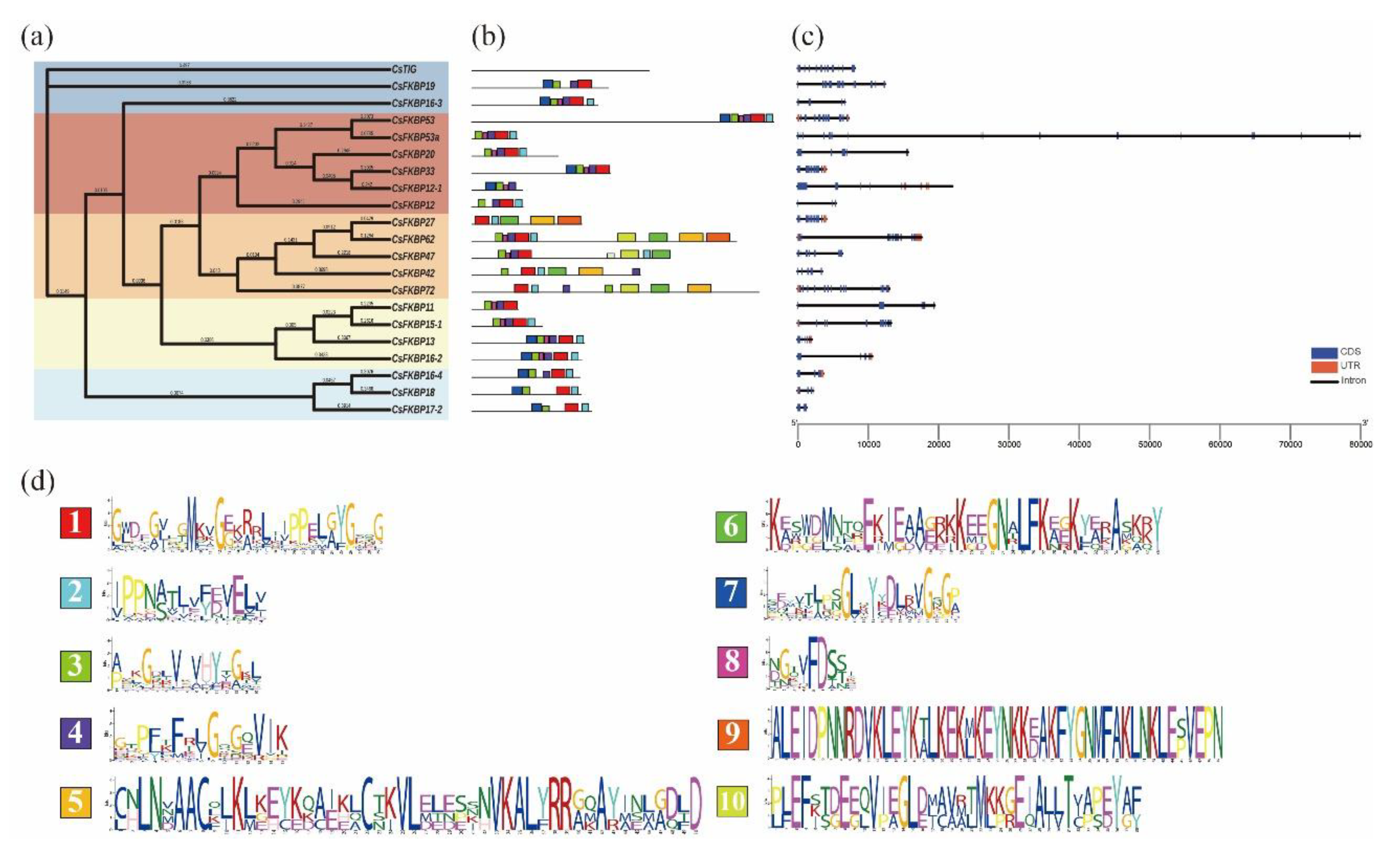
Figure 4.
The expression of CsFKBPs under different cold stresses. (a)The expression pattern of CsFKBPs. CK was the control, CA1-6h was 6 h of 10°C stress, CA1-3d was 3 days of 10°C treatment and CA2-3d was 3 days of 4°C treatment(P<0.05). (b)Heat map of the expression of CsFKBPs in Shuchazao.
Figure 4.
The expression of CsFKBPs under different cold stresses. (a)The expression pattern of CsFKBPs. CK was the control, CA1-6h was 6 h of 10°C stress, CA1-3d was 3 days of 10°C treatment and CA2-3d was 3 days of 4°C treatment(P<0.05). (b)Heat map of the expression of CsFKBPs in Shuchazao.
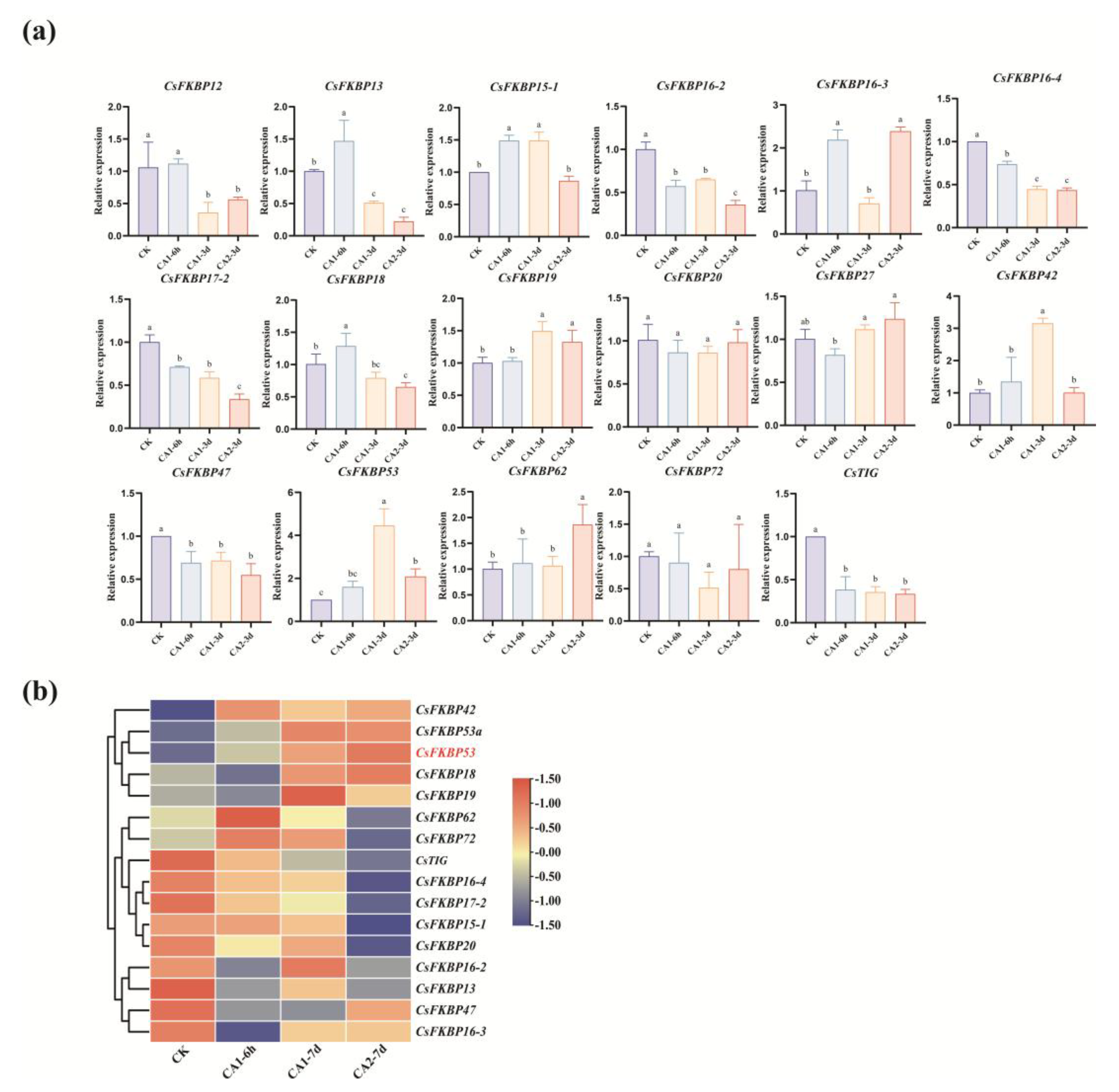
Figure 5.
Multiple sequence alignment, cluster analysis and protein structure analysis of conserved structural domains of FKBP53 proteins. (a) Multiple sequence alignment of CsFKBP53 with homologous proteins of other species. α-helices and β-folds are marked above the sequence with red and blue straight lines respectively. The FKBP-C structural domain is shown in the black box, with the same amino acids shown on a dark blue background. (Similarity: dark blue=100%; pink>75%; cyan>50%) (b) Phylogenetic analysis of CsFKBP53 and other species with homologous proteins. (c) Three-dimensional structure of CsFKBP53 with homologous proteins of other species.
Figure 5.
Multiple sequence alignment, cluster analysis and protein structure analysis of conserved structural domains of FKBP53 proteins. (a) Multiple sequence alignment of CsFKBP53 with homologous proteins of other species. α-helices and β-folds are marked above the sequence with red and blue straight lines respectively. The FKBP-C structural domain is shown in the black box, with the same amino acids shown on a dark blue background. (Similarity: dark blue=100%; pink>75%; cyan>50%) (b) Phylogenetic analysis of CsFKBP53 and other species with homologous proteins. (c) Three-dimensional structure of CsFKBP53 with homologous proteins of other species.
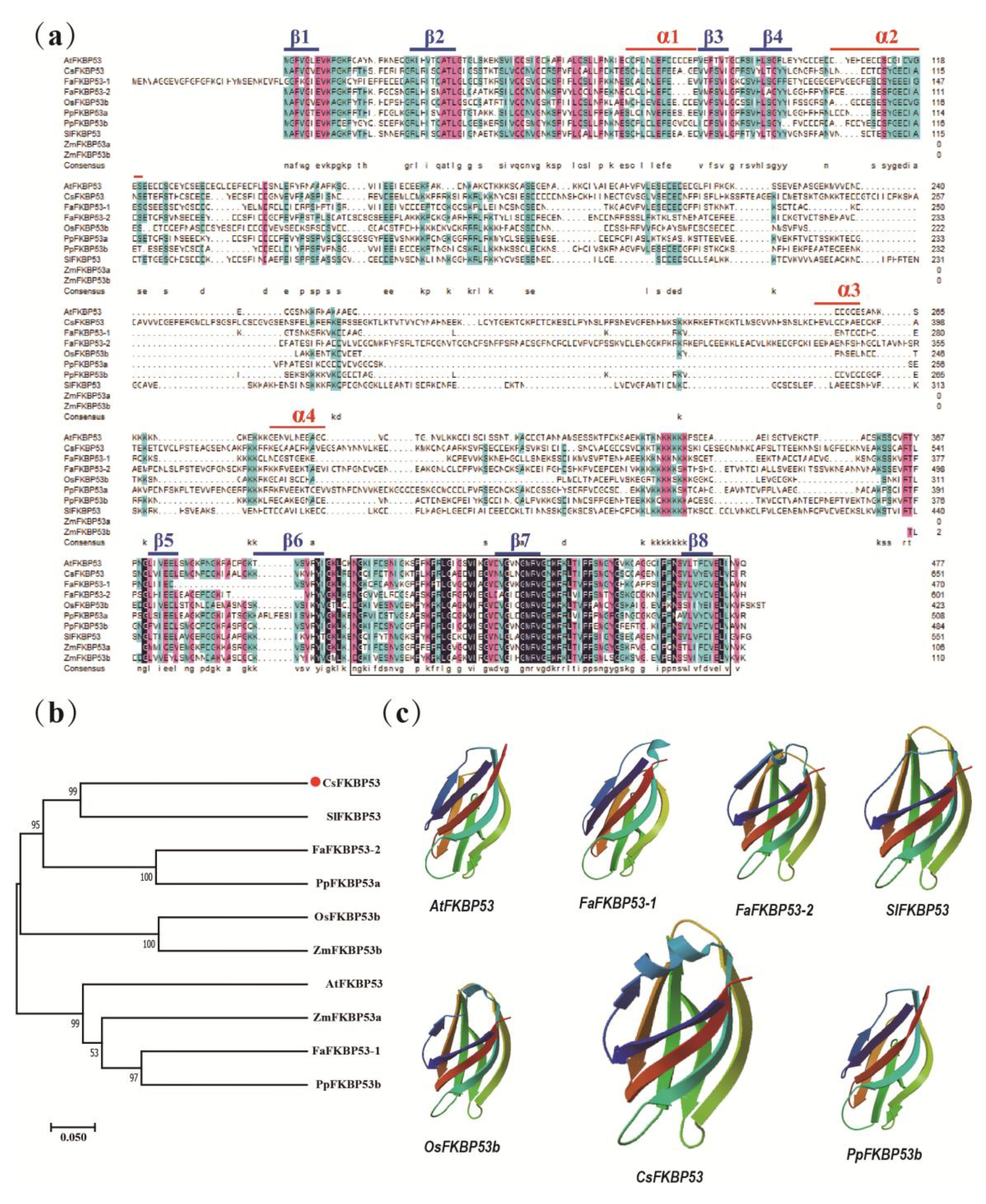
Figure 6.
The relative expression and subcellular localization of CsFKBP53. (a) The relative expression levels of CsFKBP53 in different tissues. (b) The relative expression levels of CsFKBP53 in different tea species. (c) and (d) are the relative expression levels of CsFKBP53 after 4 ℃ and 10℃ cold stress. (e) The localization of CsFKBP53-GFP in the lower epidermal cells of round onions was observed by laser confocal microscopy using 35s-GFP as a control.GFP: green fluorescence channel; Bright Field: bright field; Merged: GFP and Bright Field merged channel, scale bar = 100 μm.
Figure 6.
The relative expression and subcellular localization of CsFKBP53. (a) The relative expression levels of CsFKBP53 in different tissues. (b) The relative expression levels of CsFKBP53 in different tea species. (c) and (d) are the relative expression levels of CsFKBP53 after 4 ℃ and 10℃ cold stress. (e) The localization of CsFKBP53-GFP in the lower epidermal cells of round onions was observed by laser confocal microscopy using 35s-GFP as a control.GFP: green fluorescence channel; Bright Field: bright field; Merged: GFP and Bright Field merged channel, scale bar = 100 μm.
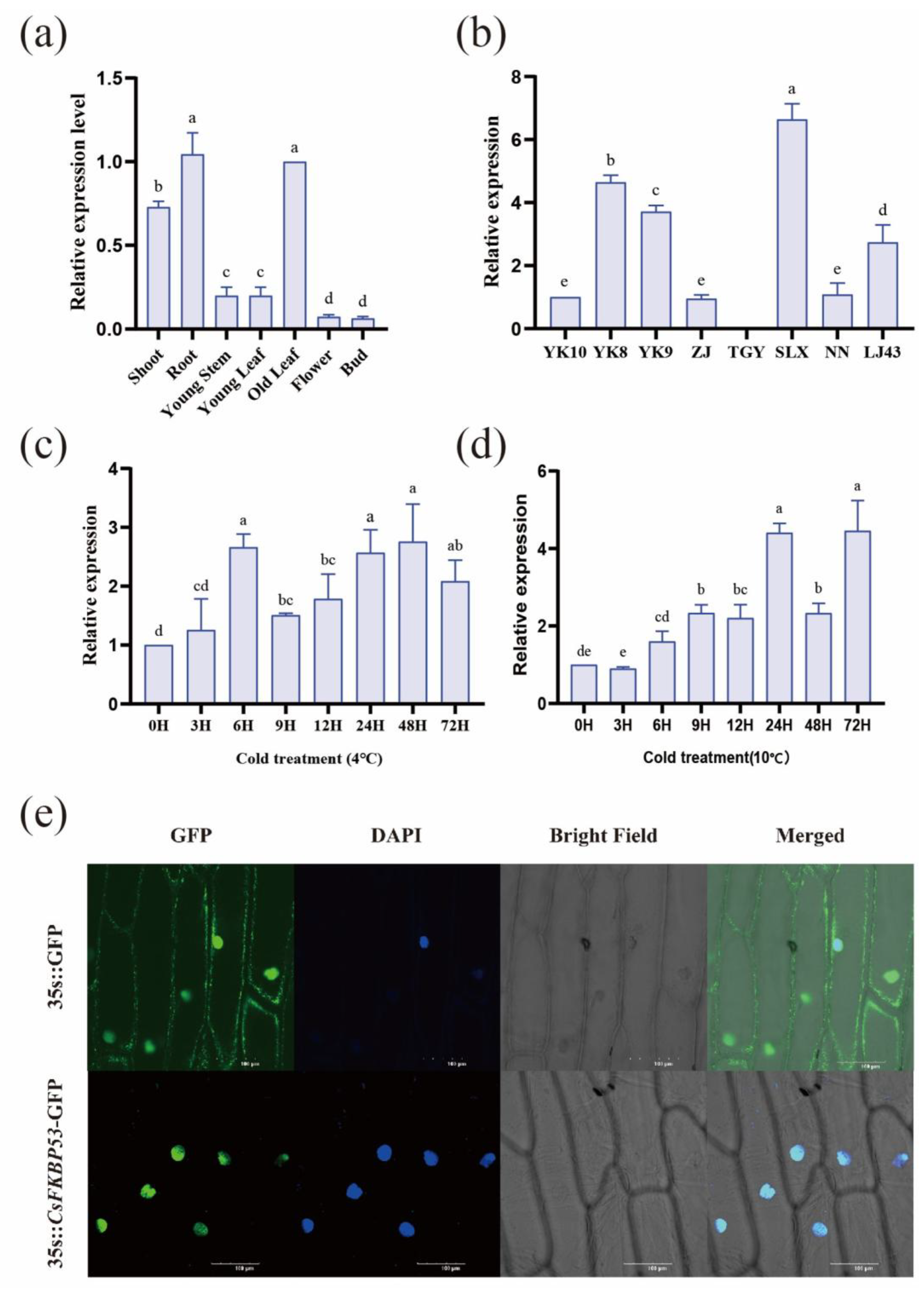
Figure 7.
Cold sensitivity of CsFKBP53-OE plants. (a) Phenotypic comparison of Arabidopsis plants. WT-4 °C is the 4 °C-treated 48 H wild-type control, OE-4 °C is the 4 °C-treated 48 H CsFKBP53 overexpression plants, and WT-25 °C is the 25 °C wild-type control. (b) Expression analysis of CsFKBP53 transgenic plants. Expression of CsFKBP53 transgenic plants was analysed by qRT-PCR. Leaf tissues from nine overexpression lines and wild-type control WT were collected for qRT-PCR analysis. (c) Arabidopsis plant root length statistics, three replicates of each plant were taken for measurement, and the vertical line in the figure indicates the standard error. Significance analysis was performed using SPSS 22 for one-way ANOVA, and different lowercase letters indicate significant differences. (p < 0.05) (d). Results of POD, SOD and CAT activity assays in Arabidopsis plants.
Figure 7.
Cold sensitivity of CsFKBP53-OE plants. (a) Phenotypic comparison of Arabidopsis plants. WT-4 °C is the 4 °C-treated 48 H wild-type control, OE-4 °C is the 4 °C-treated 48 H CsFKBP53 overexpression plants, and WT-25 °C is the 25 °C wild-type control. (b) Expression analysis of CsFKBP53 transgenic plants. Expression of CsFKBP53 transgenic plants was analysed by qRT-PCR. Leaf tissues from nine overexpression lines and wild-type control WT were collected for qRT-PCR analysis. (c) Arabidopsis plant root length statistics, three replicates of each plant were taken for measurement, and the vertical line in the figure indicates the standard error. Significance analysis was performed using SPSS 22 for one-way ANOVA, and different lowercase letters indicate significant differences. (p < 0.05) (d). Results of POD, SOD and CAT activity assays in Arabidopsis plants.

Table 1.
List of CsFKBP genes identifies in tea plant.
| Gene Name | Gene ID | FKBP-C domain number | CDS Length/bp | Amino acid/aa | Molecular Weight/kD | Theoretical pl | Subcellular Localizations | |
|---|---|---|---|---|---|---|---|---|
| CsFKBP11 | CsasTrans141447 | 1 | 306 | 101 | 11.02 | 8.87 | Nuclear | |
| CsFKBP12 | CsasTrans179814 | 1 | 692 | 112 | 11.93 | 7.75 | ChloroPlast | |
| CsFKBP12-1 | CsasTrans156117 | 1 | 332 | 110 | 12.26 | 10.13 | Nuclear | |
| CsFKBP13 | CsasTrans097150 | 1 | 1075 | 244 | 25.74 | 8.7 | Peroxisome | |
| CsFKBP15-1 | CsasTrans095919 | 1 | 959 | 153 | 16.34 | 7.68 | Vacuole | |
| CsFKBP16-2 | CsasTrans065843 | 1 | 1113 | 238 | 25.46 | 9.44 | Cytoplasm | |
| CsFKBP16-3 | CsasTrans159019 | 1 | 1132 | 273 | 29.67 | 8.47 | ChloroPlast | |
| CsFKBP16-4 | CsasTrans093053 | 1 | 1200 | 234 | 24.74 | 9.68 | ChloroPlast | |
| CsFKBP17-2 | CsasTrans181995 | 1 | 1220 | 259 | 27.69 | 6.18 | ChloroPlast | |
| CsFKBP18 | CsasTrans153703 | 1 | 1036 | 237 | 25.4 | 9.58 | ChloroPlast | |
| CsFKBP19 | CsasTrans009318 | 1 | 1424 | 295 | 32.24 | 6.79 | Extracell | |
| CsFKBP20 | CsasTrans077854 | 1 | 2327 | 188 | 20.5 | 7.76 | Nuclear | |
| CsFKBP27 | CsasTrans003218 | 1 | 1236 | 238 | 27.09 | 8.64 | ChloroPlast | |
| CsFKBP33 | CsasTrans135390 | 1 | 900 | 300 | 33.82 | 4.52 | Nuclear | |
| CsFKBP42 | CsasTrans194578 | 1 | 1493 | 364 | 41.76 | 5.66 | Nuclear | |
| CsFKBP47 | CsasTrans171260 | 3 | 1920 | 427 | 46.72 | 4.76 | Cytoplasm | |
| CsFKBP53 | CsasTrans084696 | 1 | 2329 | 651 | 72.43 | 5.61 | Nuclear | |
| CsFKBP53a | CsasTrans162628 | 1 | 2429 | 100 | 10.81 | 10.44 | ChloroPlast | |
| CsFKBP62 | CsasTrans005298 | 2 | 2600 | 571 | 63.81 | 5.17 | Peroxisome | |
| CsFKBP72 | CsasTrans105671 | 3 | 2428 | 619 | 69.99 | 5.35 | Nuclear | |
| CsTIG | CsasTrans154119 | 0 | 1151 | 383 | 43.06 | 5.09 | Nuclear |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



