
Submitted:
07 February 2025
Posted:
10 February 2025
You are already at the latest version
Abstract
The main objective of this study was to investigate the changes in winter oilseed rape (Brassica napus L.) under simulated prolonged drought and to assess the effects of a microbial biostimu-lant ProbioHumus and calcium, individually and in combination, in order to improve the plant's drought resistance and to identify the biochemical processes occurring in the plant tis-sues. The oilseed rape cv. 'Visby' was grown under controlled laboratory conditions. CaCO3 (hereafter Ca) (3.71 g) was added to the soil of one pot at 70 g m-2. After 21 days of cultivation, 3-4 leaf stage seedlings were sprayed with ProbioHumus 2 ml 100 ml-1 and exposed to drought for 8 days to achieve a high water deficit. Irrigation was then resumed and recovery was assessed af-ter 4 days. The results showed that the microbial biostimulant alleviated the physiological and biochemical response of oilseed rape to drought stress. ProbioHumus + Ca reduced plant wilting by increasing leaf relative water content (RWC) by 87% and induced drought tolerance by in-creasing endogenous proline content by 4-fold, photosynthetic pigment content in leaves by 10-28%, reducing H2O2 by 53% and malondialdehyde (MDA) by 45%, and by stimulating stomata opening (by 2-fold on the upper and 1.4-fold in the lower leaf surface), vs drought control. The most effective measure to increase plant survival and/or resume growth after drought was the application of a microbial biostimulant with additional calcium to the soil.
Keywords:
Brassica napus
; calcium carbonate
; commercial plant probiotics
; drought stress responses
1. Introduction
Microorganisms play an important role in adapting and/or withstanding environmental stresses. Efficient commercial microbial inoculants (probiotics) determine the successful implementation of these beneficial microbes in agriculture especially when microbes are alive in the formulation products [1,2,3]. Oilseed rape (Brassica napus L) is an economically crucial agricultural crop widely grown in many countries. However, adverse environmental conditions such as drought can limit the growth, development, and survival of oilseed rape plants, especially in regions with low rainfall [4,5]. Drought induces morphological, physiological, and biochemical changes in crops and results in reduced yield, which is a major task that needs to be addressed by finding of ecological tools to solve this problem [6]. Currently, strategies to enhance the plant′s ability to cope with drought stress include traditional breeding, genetic engineering of drought-tolerant transgenic plants, and water-saving irrigation [7,8]. Unfortunately, these strategies are very complex, time-consuming, and labor-intensive. There is a need to focus on organic and eco-friendly methods for improving crops facing drought stress. Many reports reveal that the use of plant probiotic microorganisms, also known as bioprotectants, biofertilizers, or biostimulants, have been extensively associated with stress responses in plants and play a key role in the control of plant abiotic stress [9,10]. According to the Higa and Parr concept [11], environmentally friendly effective microorganisms (EM) benefit on agricultural fields where they are utilized as fertilizer or sprayed. EM contains selected species of microorganisms including predominant populations of lactic acid bacteria and yeasts and smaller numbers of photosynthetic bacteria, actinomycetes and other types of organisms that can improve the growth, yield, and quality of crops [11]. Integrating beneficial microorganisms as biostimulants in sustainable agriculture is therefore attracting growing interest. So, one of the questions of this study was to evaluate whether foliar application of commercial plant probiotic composition ProbioHumus produced by Latvian consumers improves the response of oilseed rape to prolonged water deficit stress. Probiotics can be applied directly to the soil, via seed treatment, or foliar spraying. In addition, calcium (Ca2+) is known as the main signal transmitter in controlling plant development and the response to drought stress, especially when Ca levels are low, which has a very negative impact on the production of many crops. Furthermore, its multifaceted roles in the response to drought stress include reducing the degree of lipid peroxidation and increasing the activity of antioxidant enzymes [12,13,14,15,16]. Several studies have demonstrated the positive effects of exogenous calcium application in improving stress tolerance, which can be attributed to water status regulation, stomatal opening and closing, activity of antioxidant systems, osmolyte accumulation, and improvement of photosynthetic pigment contents [17,18,19,20,21,22]. These findings provide new insights into the mechanisms by which exogenous Ca2+ enhances the phenotypes of crops under water-limiting conditions but it should be noted that in calcareous soils added Ca should not be used. The combined use of calcium and probiotics is likely to have a synergistic effect and could be a promising approach to improve sustainable crop production and abiotic stress tolerance. The use of such substances to regulate drought tolerance in oilseed rape (Brassica napus L.) has not been investigated. We hypothesized that oilseed rape treatment with microbial biostimulants coupled with Ca can benefit growth by strengthening (intensification) the tolerance mechanisms protecting them from detrimental effects of drought stress by up-regulating the antioxidant system, osmoregulation, and secondary metabolite accumulation, and increasing the survival.
2. Materials and Methods
2.1. Plant Growth Conditions and Treatments
Winter oilseed rape (Brassica napus L.) cv. ‘Visby’ was grown in controlled laboratory conditions in a Climacell plant growing chamber (Medcenter Einrichtungen GmbH) at 20–23 °С under the illumination of 60 µmol m-2 s-1 cool white fluorescent light photon flux at the soil level and photoperiod 16/8 h and at 65% humidity. The 60 seeds were sown in each pot (40 × 17 × 13) cm with 6 L high-quality pH adjusted moss peat substrate (pH 5.5-6.5) (SuliFlor SF2, Radviliškis, Lithuania). According to the experimental design, seeds from four pots were primed in a commercial plant probiotic ProbioHumus (purchased from Baltic Probiotics, Rucava, Latvia) at 2 µL g-1 (Figure 1). Peat moisture was assessed using a soil moisture meter (Biogrod, China) and maintained at 70% by daily irrigation. Calcium carbonate CaCO3 (MKDS, Vilnius, Lithuania) (hereafter ’Ca’) was added to the soil according to the scheme, at a rate of 3.71 g per pot, calculated at 70 g m-2. After 21 days of cultivation, seedlings were sprayed according to the experimental design at the 3-4 leaf stage [23] with ProbioHumus at 2 ml 100 ml-1, containing Bacillus subtilis (103 CFU/mL), Saccharomyces cerevisiae, Bifidobacterium animalis, B. bifidum, B. longum (104 CFU/mL), Lactobacillus diacetylactis, L. casei, L. delbrueckii (105 CFU/mL), Lactococcus lactis (102 CFU/mL), Streptococcus thermophilus, Rhodopseudomonas palustris and R. sphaeroides (104 CFU/mL). For the drought stress control studies, plants were subjected to prolonged drought stress for 8 days to reach a high-water deficit. After 8 days of prolonged drought, irrigation was resumed to achieve a soil moisture content of 70%, then plant recovery was assessed.
2.2. Experimental Design
Eight pots were used for the experiment: four for watering and four for prolonged drought simulation (in two replicates). Experiment was repeated three times. Calcium was added to the soil at a rate of 3.71 g per pot, based on 70 g m−2. ProbioHumus was used for seed priming and seedling spraying 2 mL per pot according to the following scheme: 1) watered control, 2) ProbioHumus and watering, 3) Ca and watering, 4) Ca + ProbioHumus and watering 5) drought control, 6) ProbioHumus and drought, 7) Ca and drought, 8) ProbioHumus + Ca and drought.
Figure 1.
Experimental design. Sixty plants per pot were used, each variant was replicated twice, and three biological experiments were performed.
Figure 1.
Experimental design. Sixty plants per pot were used, each variant was replicated twice, and three biological experiments were performed.
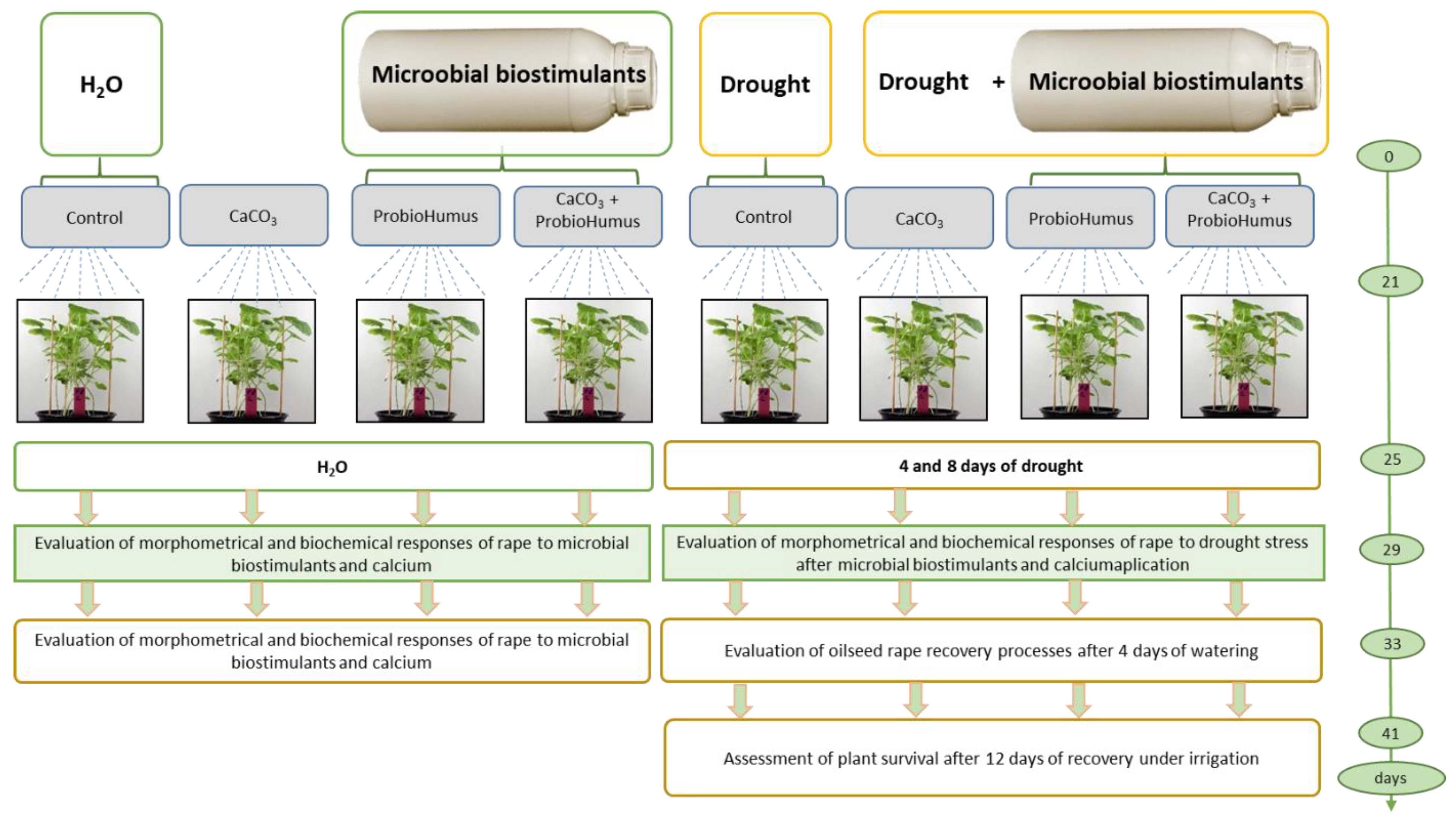
2.3. Sampling
Rapeseed seedlings were sampled three times: on the 4th day of drought (40 % peat moisture), on the 8th day of prolonged drought (20 % peat moisture) and on the 12th day of the experiment, after the resumption of the irrigation on the 8th day (70% peat moisture). The watered plants, used as controls, were sampled at the same time (pea moisture 70%). For biochemical analysis, three independent replicates were carried out using the third leaves of oilseed rape seedlings. Freshly harvested samples of oilseed rape leaves were used for ethylene emission analysis and pigment measurement. For plasma membrane ATPase (PM ATPase) activity, MDA, H2O2, and proline assays, the samples were immediately frozen in liquid nitrogen and stored in a low-temperature freezer (Skadi Green line) at -80 °C until the analysis.
2.4. Relative Water Content (RWC) Measurement
Plant water status was determined by measuring leaf RWC of control and drought-stressed plants according to [24]. The first weighing was carried out immediately after collecting the plant material – fresh weight (Wf), the second after soaking the plant material for 24 h in distilled water – saturated weight (Wt).The dry weight (Wd) was obtained by drying the leaves in a drying chamber in 70° C and then weighted. RWC have been calculated of thirty plant leaves per tratment from three biological experiments using the equation:
RWC = [(Wf − Wd)/(Wt − Wd)] × 100%
2.5. Evaluation of Stomatal Density and Opening
For stomatal measurements, histological preparations were made using the leaf impression technique on a fully expanded leaf, according to the method described by Hilu and Randall [25] A layer of acrylic (synthetic nail coating) was brushed onto the adaxial and abaxial sides of the leaf, dried for a few seconds, then carefully removed and mounted for microscopy. Stomatal density (number in mm2) and percentage of closure stomata was examined under a microscope (magnification x20 and x40) (Olympus BX51) from three microscope fields randomly selected for each leaf epidermis. Three readings were taken in random spots from each print, and the average of the three readings was used as the stomata number for each leaf.
2.6. Estimation of Photosynthetic Pigment Content
Photosynthetic pigments Chl a, Chl b, total Chl and carotenoids were extracted with N, N’-dimethyl-formamide (DMF) (Sigma-Aldrich) and estimated according the absorbance of pigments measured at 480, 664, 647 nm [26].
2.7. Measurement of MDA Content
The Hodges [27] method was used to estimate MDA. Leaf material (0.3 g) was homogenised using 5% trichloroacetic acid (TCA) (Sigma-Aldrich, St. Louis, MO, USA). The homogenates were centrifuged at 13 × g for 17 min (centrifuge MPW-351 R) and the supernatant was added to 20% TCA containing 0.50% thiobarbituric acid (TBA) (Alfa Aesar, Haverhill, MA, USA). The homogenate was placed in a water bath (Blockthermostat BT 200) at 95 °C for 30 min and then cooled on ice. The ab-sorption of the supernatant was read with a spectrophotometer at 532 nm. The optical density was measured with a spectrophotometer (Analytik Jena Specord 210 Plus, Analytik Jena, Jena, Germany) at 532 and 660 nm. The results were expressed in µmol g-1 FW. To avoid drying effects, RWC was used as a coefficient to calculate the amount of MDA and to evaluate the effective differences between treatments.
2.8. Estimation of Hydrogen Peroxide (H2O2) Content
The method of Velikova with coauthors [28] was used to determine the H2O2 content in leaves. Leaf material (0.3 g) was homogenised using 5% trichloroacetic acid (TCA) (Sigma-Aldrich, St. Louis, MO, USA) and centrifuged at 10 000 g for 5 min. The supernatant was mixed with 10 mM, pH 7.0 potassium phosphate buffer (Alfa Aesar) and 1 M potassium iodide (Alfa Aesar) in a 1:1:2 ratio. The reaction solution was incubated for 30 min at 25 °C in the dark. The absorbance of the supernatant was measured with a spectrophotometer at 390 nm. The H2O2 content was calculated from the standard curve. The results are expressed in µmol g-1 FW.
2.9. Estimation of Proline Levels
The amount of free proline in frozen rapeseed leaves was determined using ninhydrin [29]. A supernatant (0.5 g) of equal volumes of plant material, acetic acid and acidified ninhydrin were mixed and heated at 108.5 °C. The resulting chromophore was extracted with toluene. Calibration was carried out with L-proline (SigmaAldrich Chemie, GmBH, Steinheim, Germany) as standard. Absorbance was read spectrophotometrically at 520 nm using a multilayer quartz cuvette and a Rainbow microplate reader. Calculations were carried out using the SLT software (SLT Labinstruments. Proline concentrations in the samples are expressed in µmol g-1 FW.
2.10. Estimation of Ethylene Levels
Endogenous ethylene emission from fresh leaves was estimated according to Child et al. [30] with minor modifications. Leaf tissues (0.25-0.45 g fresh weight) were placed in 40-ml clear glass vials (Agilent technologies, Santa Clara, CA, USA) sealed with PTFE/Si septum caps and incubated for 24 h at 21 °C in the dark. After incubation, a 1 ml gas sample from each vial was withdrawn with a gas-tight syringe (Agilent technologies) and injected into a gas chromatograph equipped with a stainless steel column (Propac R, Sigma-Aldrich, USA) and a hydrogen flame ionisation detector. The temperatures of the injector, column and detector were 110, 90 and 150 °C respectively. Helium (AGA) was used as carrier gas. Calibration was carried out with an ethylene standard (Messer, Bad Soden, Germany). The results were expressed in nL g−1 FW h−1.
2.11. Estimation of PM ATPase Activity
The membrane-enriched fraction (microsomes) was isolated from oilseed rape leave samples by differential centrifugation : 1) at 4.500×g for 5 min, 2) at 18.000×g for 20 min (centrifuge MPW-351 R), 3) supernatant at 92.200×g (centrifuge Thermo Scientific Sorvall WX 100 Ultra) for 1 h. Protein content was measured using the Bradford dye binding procedure [31] at 595 nm. The H+-ATPase activity of the microsomal fraction was assessed by the release of inorganic phosphate (Pi), which accumulates as a result of the hydrolysis of ATP [32]. The colour reaction for Pi measurement was carried out with ammonium molybdate and stannous chloride at 750 nm. PM ATPase activity was expressed as μmol Pi mg-1 protein h-1.
2.12. Determination of Plant Survival
After a recovery period of 12 days, rehydrated plants in each pot were estimated as dead from the drought or living. To confirm whether plants were recovered, generating green shoots at the growing point of apical meristems in the centre of leaf rosette was counted. Plant survival is presented as the percentage of plants that recovered out of the total number of plants.
2.13. One-Way Statistical Analysis by ANOVA
The results are presented as mean of three independent experiments with at least two replicates. The data were analysed using one-way analysis of variance (one-way ANOVA). The data meet the requirements of normal distribution and homogeneity of variance. Tukey’s test was performed to test the statistical significance of differences (p <0.05) between means.
3. Results
3.1. Effect of Microbial Biostimulant and Ca on Physiological and Biochemical Indicators of Control and Drought Stressed Plants
3.1.1. Leaf RWC
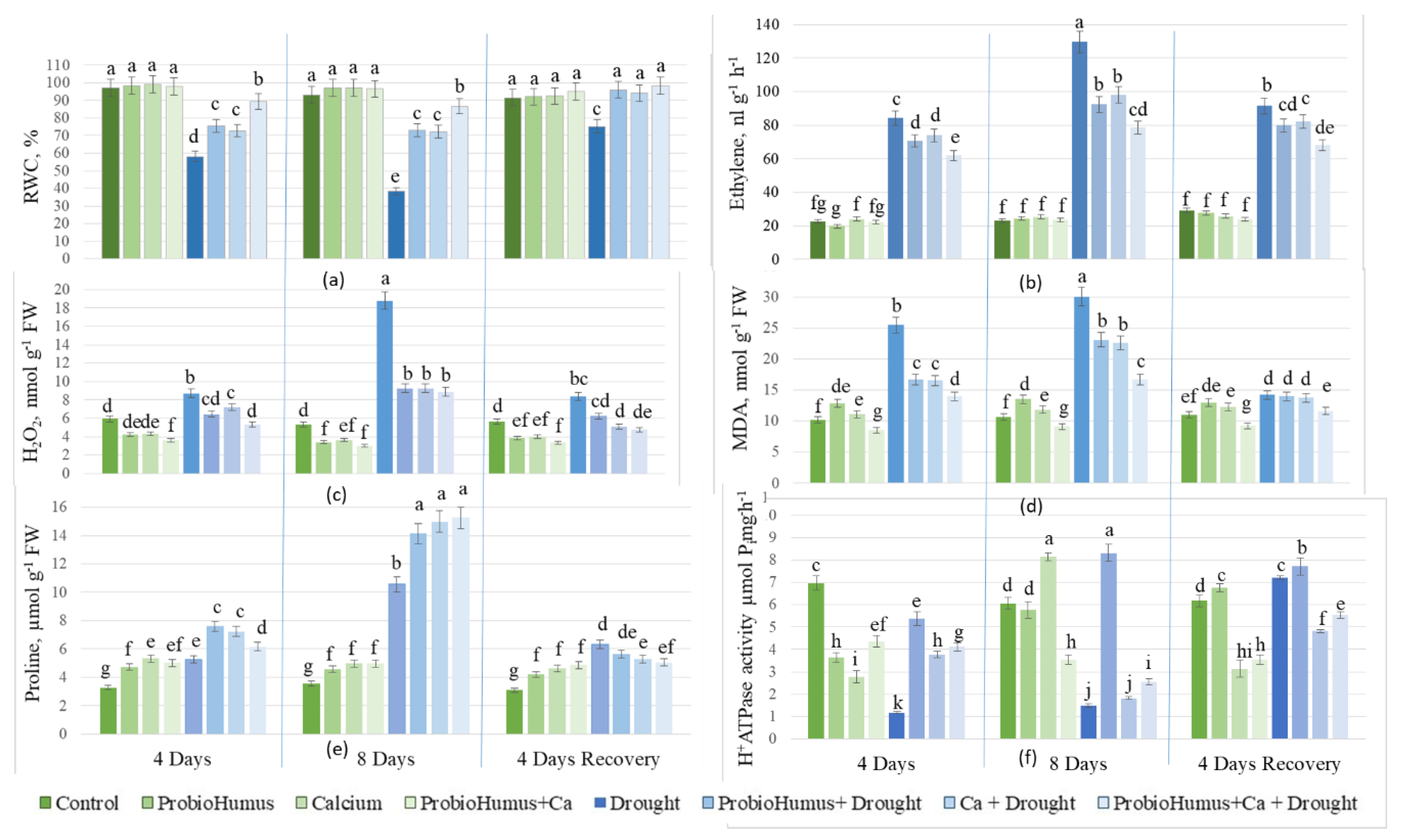
Prolonged drought significantly (p < 0.05) reduced leaf RWC to 38%. Exogenous application of ProbioHumus, Ca, and ProbioHumus + Ca reduced the effects of 8 days of drought from 38% to 73%, 72% and 87%, respectively. After 4 days of renewed irrigation, the RWC of drought-exposed ProbioHumus and Ca treated plants increased to the RWC of the regularly irrigated plants, except for the drought control, whose RWC did not recover fully (Figure 2a).
3.1.2. Ethylene Level
Prolonged drought promoted ethylene release from oilseed rape leaves compared to the irrigated control plants. After 4 and 8 days of drought, the ethylene content increased by 3.7 and 5.6 fold respectively. ProbioHumus, Ca and ProbioHumus + Ca treatments significantly reduced the ethylene emission in leaves compared to plants exposed to 8 days of drought. Drought exposure was best reduced in ProbioHumus + Ca treated plants, where the ethylene content was 41% lower than in drought-only treated plants. After 4 days of recovery by irrigation, the ethylene content of plants treated with the drought-protective compounds decreased more than that of untreated and drought stressed plants (Figure 2b). The ethylene emission of continuously irrigated plants was independent of their treatment with drought protectants.
Figure 2.
Effect of microbial biostimulant and soil calcium on RWC (a), ethylene emission (b), H2O2 (c), MDA (d), proline (e) accumulation and PM ATPase activity (f) in leaves of Brassica napus plants after 8 days of prolonged drought and after recovery from 4 days of watering. The values presented are the mean and standard deviation of thirty plant leaves (n = 30) from three biological experiments. Means marked with different letters on the same day of drought are significantly different from watered untreated control (p < 0.05).
Figure 2.
Effect of microbial biostimulant and soil calcium on RWC (a), ethylene emission (b), H2O2 (c), MDA (d), proline (e) accumulation and PM ATPase activity (f) in leaves of Brassica napus plants after 8 days of prolonged drought and after recovery from 4 days of watering. The values presented are the mean and standard deviation of thirty plant leaves (n = 30) from three biological experiments. Means marked with different letters on the same day of drought are significantly different from watered untreated control (p < 0.05).
3.1.3. H2O2 and MDA Content
A prolonged drought of 8 days caused a pronounced increase in the ROS (H2O2) levels in oilseed rape leaves from 5 to 19 nmol g-1 FW (Figure 2 c) which led to lipid peroxidation as assessed by MDA, which increased from 11 to 30 nmol g-1 FW (Figure 2 d). Drought-protective compounds reduced H2O2 release almost 2-fold: ProbioHumus + Ca had the greatest effect, reducing H2O2 by 53%. MDA content also was reduced by ProbioHumus by 25%, Ca by 24%, and ProbioHumus + Ca by 45% compared to the drought control. In addition, H2O2 levels were reduced 2-fold after 4 days of recovery from irrigation compared to the 8-day drought control. The applied microbial biostimulant ProbioHumus, as well as Ca, and ProbioHumus + Ca further reduced the H2O2 levels in oilseed rape leaves after 4 days of renewed irrigation, and the MDA level was also reduced to the level of continuously irrigated plants (Figure 2c,d).
3.1.4. Endogenous Proline Content
The endogenous proline content of drought-stressed oilseed rape leaves increased with the duration of drought, reaching 5.26 µmol g-1 FW during the first 4 days of drought, and then increasing almost 3-fold during the next 4 days compared to the irrigated control plants (Figure 2e). Both ProbioHumus and Ca, individually and in combination, increased the proline content in oilseed rape leaves in both stages of simulated drought vs drought control. The highest proline content of 15.26 µmol g-1 FW was found in the leaves of plants treated with ProbioHumus + Ca + drought. When irrigation was resumed, the free proline content decreased rapidly and after four days was close to that found in unstressed plants.
3.1.5. PM ATPase Activity
Under drought stress, PM ATPase activity in oilseed rape cells decreased by up to 6-fold compared to control plants. Drought-protective compounds at the beginning of the simulated drought (after 4 days) maintained PM ATPase activity similar to that of continuously irrigated plants. In oilseed rape grown with added Ca, both with and without ProbioHumus application, PM ATPase activity decreased almost to that of the drought control after 8 days of simulated drought (Figure 2f). In contrast, plants treated with ProbioHumus recorded higher H+-ATPase activity (47.5% and 44.4% on average) during the entire drought period vs all other test variants. After 4 days of recovery by irrigation, the PM-ATPase activity of drought-treated plants increased 4.5-fold and approached that of continuously irrigated plants and also that of plants treated with drought-protective compounds (Figure 2f).
3.1.6. Photosynthetic Pigment Content
Quantitative analysis of leaf pigments in oilseed rape showed a significant decrease in chlorophylls a and b after 4 and 8 days of simulated drought compared to continuously irrigated plants. Foliar application with ProbioHumus, Ca additive in soil and ProbioHumus with soil Ca mitigated the drought-induced decrease in chlorophyll a and b concentrations, i.e. their concentrations became 10–28% higher vs the drought control. All protectants increased carotenoid content after 8 days of drought: carotenoid concentrations increased by 0.09 mg g-1 FW, 0.10 mg g-1 FW and 0.11 mg g-1 FW in plants treated with ProbioHumus, Ca and ProbioHumus + Ca respectively. Pigment content increased significantly with renewed irrigation, especially in plants treated with anti-drought treatments (Table 1). The pigment content of continuously irrigated plants was independent of the treatment with drought-protecting compounds.
3.2. Effect of Microbial Biostimulant and Ca on Stomata Density and Aperture in Leaves of Control and Drought Stressed Plants
Measurements of the change in stomatal density and aperture allow us to characterise the response of the plant to various environmental stimuli. In irrigated plants treated with ProbioHumus in combination with Ca, the density of stomata on the upper and lower leaf surface was higher than in the control samples. After 4 days of drought due to wilting, the number of stomata on the upper and lower leaf surface was reliably higher than in the irrigated control plants. In the upper side of oilseed rape leaves treated with ProbioHumus and Ca + drought, the density of stomata varied during the prolonged drought: after the first 4 days, it remained very similar to the irrigated control, and after 8 days of drought, it increased almost to that one of drought control on day 4 of drought (Figure 3a,b). Microbial biostimulants stimulated opening of stomata of the irrigated control plants: percent of open stomata both in upper and lower surface of control leaves was less vs plants affected by biostimulants (Figure 3c,d). After 4 days of drought, in the upper and lower surface of plants treated with ProbioHumus in complex with Ca, the number of open stomata was 30% and 50% higher, respectively, vs drought control. After 8 days of drought stress, percent of open stomata was the lowest in the lower surface of leaf, still microbial biostimulants increased stomata opening to that of irrigated control without treatment. In recovered plants, percent of stomatal opening also recovered to that of irrigated control at the beginning of the experiment, except of ProbioHumus + Ca test variant where stomata opening increased about 1.5–2 fold vs control plants (Figure 3c,d).
3.3. Effect of Microbial Biostimulant and Ca on Survival of Control and Drought Stressed Plants
After 8 days of simulated drought stress and 12 days of irrigation recovery, the number of survived oilseed rape plants treated with ProbioHumus and Ca was significantly higher (on average 42%) compared to plants treated with drought stress alone. The number of plants treated with ProbioHumus + drought was lower than those treated with Ca + drought and these differences were statistically significant (Table 2).
4. Discussion
In regions with low rainfall, droughts limit the growth, development and survival of oilseed rape [33,34]. Prolonged drought stress adversely affects plant morphological, physiological, biochemical and molecular processes leading to growth inhibition. Biologically active compounds can activate metabolic pathways, enhance plant adaptation to drought and overcome or delay the most important negative effects [3,35,36]. Plant-derived microbial stimulants have been shown to have a high potential to induce plant tolerance to various abiotic stresses and thus improve plant growth and productivity related parameters. [3,37]. Furthermore, literature data suggest that exogenous application of calcium has a positive effect on plants growing under drought stress conditions, promoting growth and antioxidant activity [38,39]. The idea of this study was to investigate the effect of the plant biostimulant ProbioHumus, Ca, and their combination on growth, biochemical responses, and recovery of oilseed rape after simulated prolonged drought under laboratory conditions.
Various cytological, physiological and biochemical indicators, such as stomatal density and openness, photosynthetic pigment levels, enzyme activity and concentrations of primary and secondary metabolites, and overall plant growth– can be used to assess plant responses to environmental factors, even when these responses are not externally visible. According to the literature, the RWC of leaves is considered as an indicator of the plant’s water status and dehydration [40,41]. RWC has been reported to decrease by more than 40% in various plants under stress conditions [42.43]. In our study, 8 days of prolonged drought reduced the RWC of oilseed rape leaves by up to 38%. Several studies suggest that plant microbial biostimulants and exogenous Ca2+ treatments can help increase the RWC of crops and to some extent compensate for drought-induced water deficit [44,45]. Our findings support this, as treatments withat ProbioHumus and ProbioHumus + Ca mitigate drought-induced wilting with RWC doubling in these plants. A reduction in the RWC cause stomatal closure, which in turn limits photosynthesis. Drought stress alters stomata density across different plant species, although the data are sometimes contradictory [46]. Some literature suggests that drought stress may reduce stomata density as seen in Hordeum vulgare, where reduced stomatal density may enhance water stress tolerance [47]. This is not always the case, and other research indicates that a higher density of smaller stomata can reduce water loss and improve drought tolerance in crops [48,49]. Therefore, drought can both increase stomatal density and reduce stomatal opening [50]. We observed a significant increase in the number of stomata in oilseed rape leaves during the drought. Stomatal density was highest on the lower leaf surface of plants treated with ProbioHumus + Ca + drought. These results align with findings from studies on olive leaves, where microbial inoculation led to increased stomatal density [51]. In recovered plants treated with ProbioHumus + Ca, stomatal opening was about 2-fold higher than in continuously watered control plants.
Drought has been shown to cause a reduction in chlorophyll content in the leaves of various crops such as rice and oats [52,53]. Reduced chlorophyll content due to drought stress can lead to inactivation of photosynthesis. Microbial plant biostimulants and exogenous calcium have been shown to modify pigment accumulation in plant leaves and play a key role in tolerating/resisting drought stress [54]. Our data show that the pigment content in oilseed rape leaves was significantly reduced after 4 and 8 days of prolonged drought compared to irrigated control. The foliar application of ProbioHumus, Ca in the soil and ProbioHumus+Ca mitigated the drought-induced decrease in chlorophyll a and chlorophyll b concentrations. Pigment content increased significantly with renewed irrigation, especially in plants treated with drought-protective compounds. The decrease in chlorophyll content due to drought stress is due to photo-oxidation of the pigment and chlorophyll degradation, which is considered to be a sign of oxidative stress [5].
Thus, when plants are exposed to stressful environmental conditions, ROS production increases and can cause severe cell damage. During drought stress, H2O2 is overproduced among all ROS [55]. In this study, a simulated prolonged drought caused a significant increase in H2O2 content in oilseed rape leaves from 5 to 19 nmol g-1 FW, resulting in an increase in lipid peroxidation, as assessed by MDA, from 11 to 30 nmol g-1 FW. On the other hand, it has been observed that low concentrations of H2O2 can be rapidly formed in plant cells in the presence of exogenous Ca2+ [16], then H2O2 molecules generate signals that trigger a series of protective physiological responses to reduce drought stress. In this study, all the drought-protecting compounds reduced H2O2 release by almost 2-fold: ProbioHumus + Ca had the highest effect, reducing H2O2 by 53% and MDA by 45% compared to the drought control. The microbial biostimulants as well as Ca alone and in complex reduced H2O2 and content in oilseed rape leaves after 4 days of renewed irrigation to the level of continuously irrigated plants. Similar data on probiotic effects on plants have been obtained in studies with other plants [56,57].
Several publications indicate that ethylene production after drought stress can reduce photosynthesis, alter growth, inhibit shoot and leaf development, and regulate ROS metabolism by modulating enzymatic antioxidants [58,59]. Interestingly, it has also been reported that low concentrations of ethylene facilitate the activation of defence signals in plants, while high concentrations inhibit defence signals in wheat [60]. We found that under drought stress, ethylene content in oilseed rape leaves increased intensively to 84.7 nl g-1 h-1, while in irrigated control plants it was only 22.7 nl g-1 h-1. Drought-protective compounds significantly reduced ethylene emission under prolonged drought. Drought exposure was best reduced in ProbioHumus + Ca treated plants, where the ethylene content was 41% lower than in drought-only treated plants, and at these concentrations ethylene can act by activating defence signals. When irrigation was resumed, the ethylene emissions of drought-stressed plants were very similar to those of continuously irrigated plants. Plants treated with drought-protective compounds showed a greater decrease in ethylene emissions than untreated and drought-stressed plants. Our data contribute to the findings of the researchers, who suggest that inoculation of probiotic microorganisms affecting ethylene content may help to remove the inhibitory effect of drought stress on plant growth. [61]. These findings were also consistent with results from [58,62] that changes in ethylene content during drought can activate the plant’s antioxidant defence system, leading to a reduction in oxidative stress and a concomitant recovery of the plants photosynthetic efficiency.
Drought stress usually correlates with oxidative and osmotic stresses leading to ion imbalance that further leads to drastic changes in cell membrane structure and various other cellular functions [63].We find data that PM ATPase is an important component of membrane integrity in response to abiotic stress [64,65] and that H+-ATPase activity can increase or decrease when plants are exposed to drought stress [66]. Also, several reports described that treatment of leaves with stress-protective compounds such as polyamines significantly increased PM ATPase activity compared to untreated plants under drought stress [67]. The results of our investigations showed that PM H+-ATPase activity in oilseed rape plants under drought stress significantly decreased, as much as 304.66%, compared to irrigated plants. In plants grown in soil with incorporated Ca, both with and without ProbioHumus, PM ATPase activity decreased in the later stage of drought. Meanwhile, higher H+-ATPase activity was recorded during the entire drought period (47.5% and 44.4% on average) in the cells of the plants that were exposed to ProbioHumus than the corresponding variant plants that were watered.
Moreover, the production and accumulation of proline is an important component of the drought resistance mechanism [68,69], which allows reducing the cell water potential and promoting water retention in the plant. The results of our study showed that proline levels were significantly higher in plants subjected to drought stress than in control plants. In the leaves of oilseed rape plants under drought stress, the proline content increased as the drought prolonged: it reached 5.26 µmol g-1 FW during the first 4 days of drought and increased to 10.56 µmol g-1. Application of probiotic ProbioHumus and Calcium increased proline content in drought stressed plants. In addition, during a period of longer drought, the synergistic effect of ProbioHumus + Ca begins to emerge, when a slightly higher amount of proline is detected, compared to the application of Probiohumus or Ca alone. This suggests that increased proline accumulation in rapeseed exposed to ProbioHumus+Ca under drought may improve the efficiency of plant osmotic regulation, which could explain the higher relative water content of plants. Several mechanisms are known by which plant probiotics induce plant tolerance to drought stress through osmoprotectants: some of them produce osmolytes acting synergistically with plant, while others induce synthesis of osmoprotectants in host plants [70]. Our results are also consistent with the authors’ data showing that Ca addition to drought-stressed plants increased proline accumulation [38,71,72] and reinforces the conclusion of [38] that calcium may be involved in osmotic adjustment by increasing proline content in plants under drought stress. While accumulating evidence suggest that both plant probiotics and calcium can increase plant tolerance to stress [73], the precise mechanism behind their combined effect on abiotic stress tolerance remain unclear. A comprehensive understanding of the molecular basis for this synergistic relationship is still lacking. The results of this study showed that both probiotics and Ca increased the survival of oilseed rape after prolonged drought exposure followed by re-irrigation, with the highest survival rates observed when probiotics when probiotics were combined with Ca. These findings underscore that the application of microbial biostimulants and calcium induces biochemical, physiological, and morphological changes in plant tissues, contributing to the enhanced survival of rape. The practical implications of this research point to the potential benefits of applying these ecological measures under field conditions.
5. Conclusions
Microbial biostimulant ProbioHumus individual and in combination with Ca, enhanced the tolerance mechanisms of oilseed rape (Brassica napus L.) against the damaging effects of drought stress. ProbioHumus and ProbioHumus + Ca controlled the drought tolerance of oilseed rape by suppressing drought-induced wilting, regulating stomatal opening, the antioxidant system, osmoregulation, and the accumulation of secondary metabolites, which ensure the survival of plants. Drought stress was best reduced in ProbioHumus + Ca treated plants, leading to a significant reduction in oxidative stress and a concomitant increase in survival.
Author Contributions
Conceptualization, V.G. and S.J.; methodology, V.Š., J.J. and R.M.; software, E.J.-B. and T.Z; formal analysis, E.J.-B. and TZ.; investigation, V.Š., R.M., and M.Z.; data curation, V.G. and J.J., writing—original draft preparation, R.M. and V.G.; writing—review and editing, V.G. and S.J., visualization, R.M. and E.J.-B.; supervision, V.G.; project administration, S.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the project of the European Commission EIP-AGRI Rural development for Operational Groups (in the sense of Art 56 of Reg.1305/2013).
Data Availability Statement
The data supporting reported results can be found in the archive of scientific reports of Nature Research Centre.
Acknowledgments
We thank Nijolė Bareikienė engineer of the Laboratory of Plant Physiology of the Nature Research Center for the support and help provided.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- de Souza Vandenberghe L.P.; Garcia, L.M.B.; Rodrigues, C.; Camara, M.C.; de Melo Pereira, G.V.; de Oliveira, J.; Soccol, C.R. Potential applications of plant probiotic microorganisms in agriculture and forestry. AIMS Microbiol. 2017, 3, 629–648. [CrossRef]
- Jiménez-Gómez, A.; Celador-Lera, L.; Fradejas-Bayón, M.; Rivas, R. Plant probiotic bacteria enhance the quality of fruit and horticultural crops. AIMS Microbiol. 2017, 3, 483–501. [CrossRef]
- Ma, Y.; Freitas, H.; Dias, M.C. Strategies and prospects for biostimulants to alleviate abiotic stress in plants. Front. Plant Sci. 2022, 13, 10242. [CrossRef]
- Sharif, P.; Seyedsalehi, M.; Paladino, O.; Van Damme, P.; Sillanpää, M.; Sharifi, A. Effect of drought and salinity stresses on morphological and physiological characteristics of canola. Int. J. Environ. Sci. Technol. 2018, 15, 1859–1866. [CrossRef]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response mechanism of plants to drought stress. Horticulturae 2021, 7, 50. [CrossRef]
- Batool, M.; El-Badri, A.M.; Hassan, M.U.; Haiyun, Y.; Chunyun, W.; Zhenkun, Y.; Jie, K.; Wang, B; Zhou, G. Drought stress in Brassica napus: effects, tolerance mechanisms, and management strategies. J. Plant Growth Regul. 2023, 42, 21-45. [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Samuel, C.; Fatai, A.; Magaji, U.; Kareem, I.; Kamarudin, Z.S.; Muhammad, I.; Kolapo, K. Drought resistance in rice from conventional to molecular breeding: A review. Int. J. Mol. Sci. 2019, 20, 3519. [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science. 2020, 368, 266-269. [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: a global perspective. Front. Plant Sci. 2017, 7, 2049. [CrossRef]
- Del Buono, D. Can biostimulants be used to mitigate the effect of anthropogenic climate change on agriculture? It is time to respond. Sci. Total Environ. 2021, 751, 141763. [CrossRef]
- Higa, T.; Parr, J.F. Beneficial and Effective Microorganisms for a Sustainable Agriculture and Environment, 1st ed.; International Nature Farming Research Center: Atami, Japan, 1994; pp. 1–16.
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud. J.P.; Thuleau, P.; Mazars, C. Calcium sensors as key hubs in plant responses to biotic and abiotic stresses. Front. Plant Sci. 2016, 7, 327. [CrossRef]
- Weigand, C.; Kim, S.H.; Brown, E.; Medina, E.; Mares, M.; Miller, G.; Harper, J.F.; Choi, W.G. A ratiometric calcium reporter CGf reveals calcium dynamics both in the single cell and whole plant levels under heat stress. Front. Plant Sci. 2021, 12, 777975. [CrossRef]
- Iqbal, Z.; Memon, A.G.; Ahmad, A.; Iqbal, M.S. Calcium mediated cold acclimation in plants: underlying signaling and molecular mechanisms. Front. Plant Sci. 2022, 13, 855559. [CrossRef]
- Kang, X.; Zhao, L.; Liu, X. Calcium signaling and the response to heat shock in crop plants. Int. J. Mol. Sci. 2024, 25, 324. [CrossRef]
- Demidchik, V.; Shabala, S.; Isayenkov, S., Cuin, T.A.; Pottosin, I. Calcium transport across plant membranes: mechanisms and functions. New Phytol. 2018, 220, 49-69. [CrossRef]
- Xu, T.; Niu, J.; Jiang, Z. Sensing mechanisms: calcium signaling mediated abiotic stress in plants. Front. Plant Sci. 2022, 13, 925863. [CrossRef]
- Ayyaz, A.; Zhou, Y.; Batool, I.; Hannan, F.; Huang, Q.; Zhang, K.; Shahzad, K.; Sun, Y.; Farooq, M.A.; Zhou, W. Calcium nanoparticles and abscisic acid improve drought tolerance, mineral nutrients uptake and inhibitor-mediated photosystem II performance in Brassica napus. J. Plant Growth Regul. 2024, 43, 516–537. [CrossRef]
- Wang, J.; Geng, Y.; Zhang, J.; Li, L.; Guo, F.; Yang, S.; Zou, J.; Wan, S. Increasing calcium and decreasing nitrogen fertilizers improves peanut growth and productivity by enhancing photosynthetic efficiency and nutrient accumulation in acidic red soil. Agronomy 2023, 13, 1924. [CrossRef]
- Liang, C. Exogenous Ca2+ alleviates waterlogging-caused damages to pepper. Photosynthetica 2016, 54, 620–629. [CrossRef]
- Dong, Q.; Wallrad, L.; Almutairi, B.O.; Kudla, J. Ca2+ signaling in plant responses to abiotic stresses. J. Integr. Plant Biol. 2022, 64, 287–300. [CrossRef]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Differential responses of phenolic compounds of Brassica napus under drought stress. Iran. J. Plant Physiol. 2018, 8, 2417–2425.
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants. BBCH Monograph, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Bonn, Germany, 2001; pp. 115–117.
- Weng, M.; Cui, L.; Liu, F.; Zhang, M.; Shan, L.; Yang, S.; Deng, X.-P. Effects of drought stress on antioxidant enzymes in seedlings of different wheat genotypes. Pak. J. Bot. 2015, 47, 49–56.
- Hilu, K.W.; Randall, J.L. A convenient method for examining the epidermis of grass leaves. Taxon 1984, 33, 413–415.
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [CrossRef]
- Hodges, D.; DeLong, J.; Forney, C.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66.
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [CrossRef]
- Child, R.D.; Chauvaux, N.; John, K.; Van Onckelen, H.A.; Ulvskov, P. Ethylene biosynthesis in oilseed rape pods in relation to pod shatter. J. Exp. Bot. 1998, 49, 829–838. [CrossRef]
- Bradford, M.M. A rapid method for the quantification of microgram quantities of proteins utilising the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254.
- Darginavičienė, J.; Pašakinskienė, I.; Maksimov, G.; Rognli, O.A.; Jurkonienė, S.; Šveikauskas, V.; Bareikienė, N. Changes in plasmalemma K+ Mg2+-ATPase dephosphorylating activity and H+ transport in relation to freezing tolerance and seasonal growth of Festuca pratensis Huds. J. Plant Physiol. 2008, 165, 825–832.
- Raza, A. Eco-physiological and biochemical responses of rapeseed (Brassica napus L.) to abiotic stresses: consequences and mitigation strategies. J Plant Growth Regul. 2021, 40, 1368–1388. [CrossRef]
- Kopecká, R.; Kameniarová, M.; Černý, M.; Brzobohatý, B.; Novák, J. Abiotic stress in crop production. Int J Mol Sci. 2023, 24, 6603. [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [CrossRef]
- Kaur, G.; Asthir, B. Molecular responses to drought stress in plants. Biol. Plant. 2017, 61, 201–209. [CrossRef]
- Ansari, M.; Devi, B.M.; Sarkar, A.; Chattopadhyay, A.; Satnami, L.; Balu, P.; Choudhary, M.; Shahid, M.A.; Jailani, A.A.K. Microbial exudates as biostimulants: role in plant growth promotion and stress mitigation. J. Xenobiot. 2023, 13, 572–603. [CrossRef]
- Khushboo, B.K.; Singh, P.; Raina, M.; Sharma, V.; Kumar, D. Exogenous application of calcium chloride in wheat genotypes alleviates negative effect of drought stress by modulating antioxidant machinery and enhanced osmolyte accumulation. In Vitro Cell. Dev. Biol. Plant. 2018, 54, 495–507. [CrossRef]
- Pathak, J.; Ahmed, H.; Kumari, N.; Pandey, A.; Rajneesh, Sinha, R.P. Role of calcium and potassium in amelioration of environmental stress in plants. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress: Biochemical and Molecular Perspectives, 1st ed.; Roychoudhury, A., Tripathi, D.K., Eds.; Wiley-Blackwell: New Jersey, USA, 2020, pp. 535–562.
- Vanani, F. R.; Shabani, L.; Sabzalian, M. R.; Dehghanian, F.; Winner, L. Comparative physiological and proteomic analysis indicates lower shock response to drought stress conditions in a self-pollinating perennial ryegrass. Plos One, 2020, 15, e0234317. [CrossRef]
- Qayyum, A.; Al Ayoubi, S.; Sher, A.; Bibi, Y.; Ahmad, S.; Shen, Z.; Jenks, M.A. Improvement in drought tolerance in bread wheat is related to an improvement in osmolyte production, antioxidant enzyme activities, and gaseous exchange. Saudi J. Biol. Sci. 2021, 28, 5238–5249. [CrossRef]
- Patanè, C.; Cosentino, S.L.; Romano, D.; Toscano, S. Relative water content, proline, and antioxidant enzymes in leaves of long shelf-life tomatoes under drought stress and rewatering. Plants 2022, 11, 3045. [CrossRef]
- Masheva, V.; Spasova-Apostolva, V.; Aziz, S.; Tomlekova, N. Variations in proline accumulation and relative water content under water stress characterize bean mutant lines (P. vulgaris L.). Bulg. J. Agric. Sci. 2022, 28, 430–436.
- Naeem, M.; Naeem, M.S.; Ahmad, R.; Ahmad, R. Foliar-applied calcium induces drought stress tolerance in maize by manipulating osmolyte accumulation and antioxidative responses. Pak. J. Bot. 2017, 49, 427–434.
- Yadav, V.K.; Yadav, R.C.; Choudhary, P.; Sharma, S.K.; Bhagat, N. Mitigation of drought stress in wheat (Triticum aestivum L.) by inoculation of drought tolerant Bacillus paramycoides DT-85 and Bacillus paranthracis DT-97. J. Appl. Biol. Biotechnol. 2022, 10, 59–69. [CrossRef]
- Zhang, J.; Zhang, S.; Cheng, M.; Jiang, H.; Zhang, X.; Peng, C.; Lu, X.; Zhang, M.; Jin, J. Effect of drought on agronomic traits of rice and wheat: a meta-analysis. Int. J. Environ. Res. Public Health 2018, 15, 839. [CrossRef]
- Hughes, J.; Hepworth, C.; Dutton, C.; Dunn, J.A.; Hunt, L.; Stephens, J.; Waugh, R.; Cameron, D.D.; Gray, J.E. Reducing stomatal density in Barley improves drought tolerance without impacting on yield. Plant Physiol. 2017, 174, 776–787. [CrossRef]
- Zhao, W-L.; Chen, Y-J.; Brodribb T.J.; Cao, K-F. Weak coordination between vein and stomatal densities in 105 angiosperm tree species along altitudinal gradients in Southwest China. Funct. Plant Biol. 2016, 43, 1126–1133. [CrossRef]
- Bi, Y.; Pei, W. Exploring drought-responsive crucial genes in Sorghum. iScience 2022, 105347.
- Li, Y.; Li, H.; Li, Y.; Zhang, S. Improving water-use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought-resistant wheat. Crop J. 2017, 5, 231–239.
- Khalil, H.A.; El-Ansary, D.O. Morphological, physiological and anatomical responses of two olive cultivars to deficit irrigation and mycorrhizal inoculation. Eur. J. Hort. Sci. 2020, 85, 5–62. [CrossRef]
- Ali, H.E.; Saad, Z.H.; Elsayed, H. The effect of drought on chlorophyll, proline and chemical composition of three varieties of egyptian rice. J. Biol. Chem. 2020, 15, 21–30.
- Xie, H.; Li, M.; Chen, Y.; Zhou, Q.; Liu, W.; Liang, G.; Jia, Z. Important physiological changes due to drought stress on oat. Front. Ecol. Evol. 2021, 9, 644726. [CrossRef]
- Fadiji, A.E.; Babalola, O.O.; Santoyo, G.; Perazzolli, M. The potential role of microbial biostimulants in the amelioration of climate change-associated abiotic stresses on crops. Front. Microbiol. 2022, 12, 829099. [CrossRef]
- Hasanuzzaman, M; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [CrossRef]
- Lalarukh, I.; Al-Dhumri, S.A.; Al-Ani, L.K.T.; Hussain, R.; Al Mutairi, K.A.; Mansoora, N.; Amjad, S.F.; Abbas, M.H.H.; Abdelhafez, A.A.; Poczai, P.; et al. A Combined use of rhizobacteria and moringa leaf extract mitigates the adverse effects of drought stress in wheat (Triticum aestivum L.). Front. Microbiol. 2022, 13, 813415. [CrossRef]
- Azeem, M.; Haider, M.Z.; Javed, S.; Saleem, M.H.; Alatawi, A. Drought stress amelioration in maize (Zea mays L.) by inoculation of Bacillus spp. strains under sterile soil conditions. Agriculture 2022, 12, 50.
- Chen, H.; Bullock Jr, D.A.; Alonso, J.M.; Stepanova, A.N. To fight or to grow: The balancing role of ethylene in plant abiotic stress responses. Plants 2021, 11, 33.
- Jasrotia, S.; Jastoria, R. Role of ethylene in combating biotic stress. In Ethylene in Plant Biology, 1st ed.; Singh, S., Husain, T., Singh, V.P., Tripathi, D.K., Prasad, S.M., Dubey, N.K., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2022; Volume 1, pp. 388–397.
- Poór, P.; Nawaz, K.; Gupta, R.; Ashfaque, F.; Khan, M.I.R. Ethylene involvement in the regulation of heat stress tolerance in plants. Plant Cell Rep. 2022, 41, 675–698. [CrossRef]
- Camaille, M.; Fabre, N.; Clément, C.; Ait Barka, E. Advances in wheat physiology in response to drought and the role of plant growth promoting Rhizobacteria to trigger drought tolerance. Microorganisms 2021, 9, 687. [CrossRef]
- Fatma, M.; Asgher, M.; Iqbal, N.; Rasheed, F.; Sehar, Z.; Sofo, A.; Khan, N.A. Ethylene signaling under stressful environments: analyzing collaborative knowledge. Plants 2022, 11, 2211. [CrossRef]
- Bernardo, L.; Carletti, P.; Badeck, F.W.; Rizza, F.; Morcia, C.; Ghizzoni, R.; Rouphael, Y.; Colla, G.; Terzi, V.; Lucini, L. Metabolomic responses triggered by arbuscular mycorrhiza enhance tolerance to water stress in wheat cultivars. Plant Physiol. Biochem. 2019, 137, 203–212. [CrossRef]
- Pottosin, I.; Olivas-Aguirre, M.; Dobrovinskaya, O.; Zepeda-Jazo, I.; Shabala, S. Modulation of ion transport across plant membranes by polyamines: understanding specific modes of action under stress. Front. Plant Sci. 2021, 11, 616077. [CrossRef]
- Li, Y.; Zeng, H.; Xu, F.; Yan, F.; Xu, W. H+-ATPases in plant growth and stress responses. Annu. Rev. Plant Biol. 2022. 73, 495–521.
- Feng, X.; Liu, W.; Zeng, F.; Chen, Z., Zhang, G.; Wu, F. K+ uptake, H+-ATPase pumping activity and Ca2+ efflux mechanism are involved in drought tolerance of barley. Environ. Exp. Bot. 2016, 129, 57-66. [CrossRef]
- Havshøi, N.W.; Fuglsang, A.T. A critical review on natural compounds interacting with the plant plasma membrane H+-ATPase and their potential as biologicals in agriculture. J. Integr. Plant Biol. 2022, 64, 268–286. [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: effects, mechanisms and management. Sustain. Agric. 2009, 53–188.
- Abdelaal, K.A.A.; Attia, K.A.; Alamery, S.F.; El-Afry, M.M.; Ghazy, A.I.; Tantawy, D.S.; Al-Doss, A.A.; El-Shawy, E.-S.E.; M. Abu-Elsaoud, A.; Hafez, Y.M. Exogenous application of proline and salicylic acid can mitigate the injurious impacts of drought stress on barley plants associated with physiological and histological characters. Sustainability 2020, 12, 1736. [CrossRef]
- Vurukonda, S.S.; Vardharajula S.; Shrivastava, M.; SkZ A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [CrossRef]
- Nayyar, H.; Walia, D.P. Water stress induced proline accumulation in contrasting wheat genotypes as affected by calcium and abscisic acid. Plant Biol. 2003, 46, 275–279. [CrossRef]
- Jaleel, C.A.; Manivannan, B.; Sankar, A.; Kishorekumar M.; Panneerselvam, R. Calcium chloride effects on salinity-induced oxidative stress, proline metabolism and indole alkaloid accumulation in Catharanthus roseus. C. R. Biol. 2007, 330, 674–683.
- Iqbal, M.; Naveed, M.; Sanaullah, M.; Brtnicky, M.; Hussain, M. I.; Kucerik, J.; Mustafa, A. Plant microbe mediated enhancement in growth and yield of canola (Brassica napus L. plant through auxin production and increased nutrient acquisition. Journal of Soils and Sediments, 2023, 23, 1233–1249. [CrossRef]
Figure 3.
Effect of microbial biostimulant and soil calcium on stomata density and apperture in leaves of Brassica napus plants after 8 days of prolonged drought and after recovery from 4 days of watering. The values presented are the mean and standard deviation of thirty plant leaves (n = 30) from three biological experiments. Means marked with different letters on the same day of drought are significantly different (p < 0.05).
Figure 3.
Effect of microbial biostimulant and soil calcium on stomata density and apperture in leaves of Brassica napus plants after 8 days of prolonged drought and after recovery from 4 days of watering. The values presented are the mean and standard deviation of thirty plant leaves (n = 30) from three biological experiments. Means marked with different letters on the same day of drought are significantly different (p < 0.05).
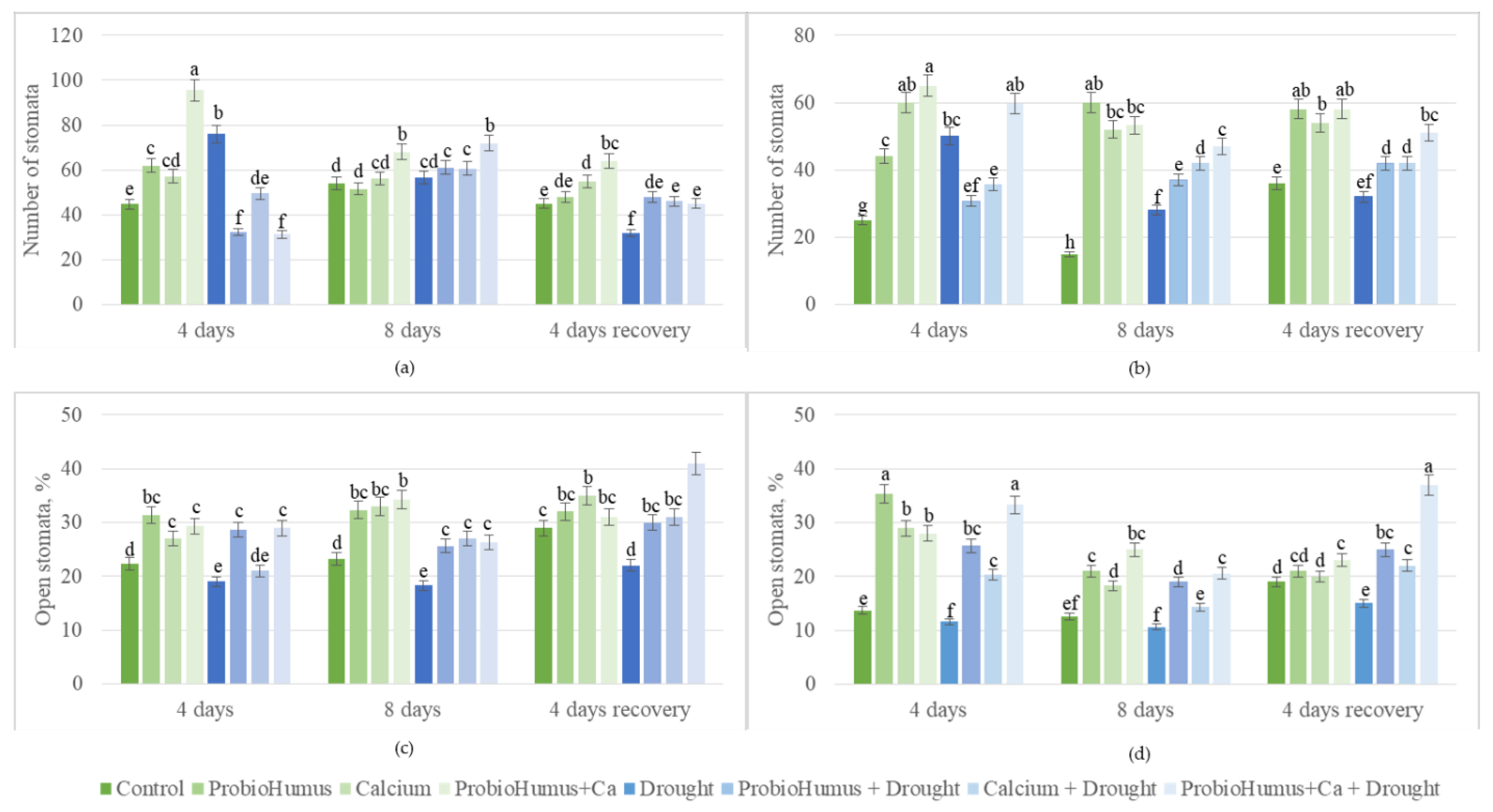

Table 1.
Effect of microbial biostimulant and soil calcium on chlorophyl and carotenoid contents in leaves of Brassica napus after 8 days of prolonged drought and after recovery from 4 days of watering. The results refer to the control group and the drought-stressed group. The values presented are the mean of thirty plants (n = 30) from three biological experiments.
Table 1.
Effect of microbial biostimulant and soil calcium on chlorophyl and carotenoid contents in leaves of Brassica napus after 8 days of prolonged drought and after recovery from 4 days of watering. The results refer to the control group and the drought-stressed group. The values presented are the mean of thirty plants (n = 30) from three biological experiments.
| Treatment | Pigment Contents (mg g−1 FW) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chlorophyll a | Chlorophyll b | Chlorophyll a/b | Carotenoid | |||||||||
| 4 Days | 8 Days | 4 Days Recovery | 4 Days | 8 Days | 4 Days Recovery | 4 Days | 8 Days | 4 Days Recovery | 4 Days | 8 Days | 4 Days Recovery | |
| Control | 1.42 a | 1.41 a | 1.43 a | 0.37 a | 0.38 a | 0.38 a | 3.84 b | 3.71 d | 3.76 c | 0.36 a | 0.36 a | 0.37a |
| ProbioHumus | 1.46 a | 1.45 a | 1.47 a | 0.39 a | 0.41 a | 0.39 a | 3.74 c | 3.54 e | 3.77 c | 0.38 a | 0.37 a | 0.37 a |
| Calcium | 1.44 a | 1.45 a | 1.46 a | 0.38 a | 0.38 a | 0.37 a | 3.79 c | 3.82 c | 3.95 b | 0.38 a | 0.37 a | 0.37 a |
| ProbioHumus+Ca | 1.43 a | 1.42 a | 1.47 a | 0.39 a | 0.40 a | 0.41 a | 3.67 c | 3.55 e | 3.59 d | 0.39 a | 0.36 a | 0.38 a |
| Drought | 0.99 d | 0.61 c | 0.75 d | 0.25 c | 0.14 c | 0.18 d | 3.96 a | 4.36 a | 4.17 a | 0.31 b | 0.22 c | 0.27 b |
| ProbioHumus+ Drought | 1.24 b | 0.75 b | 0.99 c | 0.32 b | 0.18 b | 0.28 b | 3.88 b | 4.17 b | 3.54 d | 0.36 a | 0.31 b | 0.34 a |
| Ca + Drought | 1.09 c | 0.72 b | 0.83 cd | 0.28 c | 0.17 b | 0.23 c | 3.89 b | 4.24 b | 3.61 d | 0.32 ab | 0.32 b | 0.35 a |
| ProbioHumus+Ca + Drought | 1.23 b | 0.75 b | 1.11 b | 0.32 b | 0.19 b | 0.31 b | 3.84 b | 3.95 c | 3.58 d | 0.37 a | 0.34 ab | 0.35 a |
Means with different letters in the same day of drought are significantly different from watered untreated control (p < 0.05).
Table 2.
Effect of microbial biostimulant and soil calcium. on Brassica napus plant survival after 8 days of prolonged drought, and after recovery from 12 days of watering. The results refer to the control group and the drought-stressed group. The values presented are the mean of thirty plants (n = 30) from three biological experiments.
Table 2.
Effect of microbial biostimulant and soil calcium. on Brassica napus plant survival after 8 days of prolonged drought, and after recovery from 12 days of watering. The results refer to the control group and the drought-stressed group. The values presented are the mean of thirty plants (n = 30) from three biological experiments.
| Treatment | Number of survived plants (%) | |
|---|---|---|
| Control, H2O | 100 a | |
| ProbioHumus | 100 a | |
| Ca | 100 a | |
| ProbioHumus + Ca | 100 a | |
| Drought | 34 d | |
| ProbioHumus + Drought | 60 c | |
| Ca + Drought | 69 bc | |
| ProbioHumus + Ca + Drought | 76 b |
Means with different letters in the same day of drought are significantly different (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



