
Submitted:
09 January 2025
Posted:
13 January 2025
You are already at the latest version
Abstract
This study aims to investigate whether γ -aminobutyric acid (GABA) and proline, alone and in combination, can influence the growth of oilseed rape under drought stress and after the resumption of irrigation, and to determine whether the exogenous application of these acids improves the response of oilseed rape to prolonged water deficit, and, if so, which biochemical processes are involved in the plant tissue. The research was carried out under controlled laboratory conditions. After 21 days of plant cultivation, at the 3–4 leaf stage aqueous solutions of GABA (0.1 mM) and proline (0.1 mM) were used to spray seedlings and then subjected to prolonged drought stress for 8 days to achieve a severe water deficit, next, irrigation was resumed, and recovery was assessed after 4 days. According to the results, both amino acids effectively alleviated the drought-induced stress as reflected by higher relative water content (RWC) levels, endogenous proline and photo-synthetic pigment content in leaves, and survival capacity to resume growth after drought. Plants exposed to the GABA improved membrane integrity and maintained PM ATPase activity during long-term drought stress while decreasing ethylene, H2O2, and MDA levels. Proline affected these biochemical reactions during simulated drought but to a lesser degree. The combined treatment of GABA and proline stimulated oilseed rape recovery compared with drought control after resumed watering. A significant increase in gene expression was observed upon GABA treatment compared to untreated control. Molecular studies of drought-responsive genes revealed that the gene expression level of plants treated with proline and GABA simultaneously was almost always intermediate between gene expression in proline-treated and GABA-treated plants. Based on these findings, we proposed that application of GABA is an alternative to proline technique for improving oilseed rape tolerance to drought and could be used to increase the amount and quality of crops.
Keywords:
Brassica napus
; γ-aminobutyric acid
; recovered growth
; water deficit
; drought sensitive gene expression
1. Introduction
Drought stress adversely affects plant growth and development by disturbing many physiological and biochemical processes. Oilseed rape is highly sensitive to drought and global climate change, which is causing severe and prolonged droughts in some parts of the world, is expected to reduce the survival and productivity of oilseed rape [1]. In central and eastern Europe, decreasing summer rainfall and increasing warm temperature extremes are hampering plant growth and development [2]. Numerous studies have indicated that exogenous application of hormones, polyamines, nutrients, antioxidants, osmoprotectants, etc., improves drought tolerance in plants [3,4,5,6]. According to these studies, such compounds can enhance parameters such as morphology, photosynthetic capacity, RWC, and promote osmolyte accumulation. In recent years, more studies have shown that the exogenous application of the proteogenic amino acid proline has been successful in increasing plant resistance to various abiotic stresses [7,8,9]. Furthermore, we find evidence that γ-aminobutyric acid (GABA) is associated with plant stress response and control of plant resistance to abiotic stresses [10,11,12]. In addition, GABA is a non-protein amino acid involved in a variety of physiological processes; including protective effects on plants against drought stress, increasing osmolyte content and leaf turgor, and reducing oxidative damage [12,13]. Thus, proline, a proteogenic amino acid that acts as an osmoprotector, subcellular stabiliser and antioxidant, and GABA, a non-protein amino acid that acts as a signalling molecule, both play an important role in the protection of the cells in the event of water shortage. Another common feature of these amino acids is that their main biosynthetic pathway is from glutamate. In addition, the literature shows that treatment with exogenous GABA can improve the drought tolerance of cultivated plants (white clover, sunflower, cucumber) by increasing the free proline content, the activity of the enzymes peroxidase and catalase, the rate of photosynthesis and the efficiency of water use [14,15]. Thus, GABA plays a positive role in physiological regulation when plants are exposed to unfavourable environmental stressors; hence, regulating GABA levels could be a promising approach to improve plant stress tolerance. At present, there is evidence that endogenous GABA concentrations in plants are relatively low but can increase in response to a range of stress conditions [13]. However, the effect of GABA application on drought tolerance in commercially important oilseed rape has not been investigated. Therefore, one of the questions of this study was: does exogenous application of GABA improve the response of oilseed rape to prolonged water deficit and, if so, by which biochemical processes is this reflected in the plant tissues?
Investigations of crop plants show various adaptive and acclimatization strategies to drought stress, which range from seemingly simple morphological or physiological traits that serve as important stress tolerance markers to major upheavals in gene expression in which many transcription factors are induced [16,17]. Moreover, crop plant adaptation to drought is a very complex process, altering the expression level of numerous genes. The factors mitigating drought stress (GABA, proline) should change the expression of drought-induced genes under water deprivation conditions. So, such genes could be a valuable marker for the evaluation of drought stress diminishing. Recently a set of genes, which alter their expression level in response to simulated drought, was identified and confirmed by qRT-PCR in oilseed rape, therefore bringing them as putative candidates for water stress tolerance [18]. Among them, strong increase in expression showed KIN2-like gene which is similar to LEA (late embryogenesis abundant) and KIN1/2 genes. It is known that genes encoding LEA proteins are expressed in seeds and vegetative organs during abiotic stress [19]. Also, the expression of KIN2 gene in Arabidopsis strongly increases in response to drought [20]. Two other genes, which expression increased in response to water deficit in oilseed rape were genes encoding peroxygenase 3 (caleosin 3) and EDL3 (EID1-like F-box protein 3) [18,21] acting via the proteasome degradation mechanism [22]. Also, the increase in expression in response to simulated drought in oilseed rape showed the gene of bZIP transcription factor ABI5 [18]. The transcription factor is mostly active in the earliest stages of plant development and is important under abiotic stress conditions, mainly drought and salt [23]. ABI5 could be used as a marker gene in drought tolerance studies if significant changes in expression could be detected under drought conditions compared to the untreated amino acid control. Taken together, the existing knowledge led us to the decision to evaluate KIN2-like, peroxygenase 3 (caleosin 3), EDL3 and ABI5 gene expression after proline and GABA and their combined treatments in the adaptation of oilseed rape to drought, which have never been studied. Thus, in order to increase drought tolerance in oilseed rape, the effect of GABA and proline and their co-treatment on the induction of resistance was investigated.
2. Results
2.1. Selection of Active GABA Concentrations
Plants treated with 0.1 mM GABA at 3 and 12.5 ml per pot had RWC of 58 % and 60 %, respectively, whereas drought-only treated plants had RWC of 42 % and 38 % (high stress) on the 8th day of drought. It should be noted that all concentrations of GABA had not remarkable influence the RWC of leaves of continuously watered plants. Thus, with a treatment of 0.1 mM GABA at 12.5 mL per pot, plants were less wilted compared to the drought control (Table 1). The choice of GABA concentration and dose to improve the growth of oilseed rape under simulated prolonged drought conditions showed that the most suitable concentration was 0.1 mM at 12.5 ml per pot (Table 2). It was observed that this dose of GABA enhanced the final average weight and survival of recovered seedlings after 4 days of resumed irrigation (Table 1 and Table 2). Thus, treatment with 0.1 mM GABA 12.5 ml per pot, was selected for further experiments because it promotes plant RWC and drought-stressed plants growth recovery.
2.2. Selection of Active Mix GABA + Proline Concentrations
Two variants of mix were selected for complex treatment of proline 1 mM + GABA 0.1 mM and proline 0.1 mM and GABA 0.1 mM on fresh weight of oilseed rape seedlings (per plant) (Table 3) and on plant survival after 8 days of prolonged drought recovered after 12 days of irrigation (Table 3). Treatment with 0.1 mM GABA and 0.1 mM proline 12.5 ml per pot, was selected for further experiments because it promoted drought-stressed plants growth recovery.
2.3. RWC
The RWC of leaves of plants exposed to drought for 4 to 8 days gradually decreased from 60% to 39%, indicating that the plants were highly stressed. Exogenous application of proline, GABA, and a mixture of proline + GABA effectively reduced drought-induced wilting of oilseed rape leaves (Figure 1). A positive effect of all applied compounds on RWC of plants was also observed after a 4-day recovery period. No significant differences in RWC were observed between all applied treatments when growing oilseed rape under favorable conditions.
2.4. Photosyntetic Pigments
The chlorophyll a and b content in rapeseed leaves significantly decreased under drought stress, 26 % and 47 % lower after 4 days of drought and 57 % and 67 % lower after 8 days of drought than the corresponding control plants. The carotenoid amount also decreased significantly when the plants were exposed to drought stress. The concentration of carotenoids decreased to 0.29 mg g-1 FW and 0.23 mg g-1 FW on drought days 4 and 8, respectively, when 0.34 mg g-1 FW was recorded in leaves of irrigated plants at that time. In plants exposed to exogenous GABA, proline and a mixture of these amino acids before drought stress, a higher content of chlorophyll a, chlorophyll b and carotenoids were detected at both drought and during recovery after drought (Table 4).
2.5. Ethylene
The ethylene emission study showed that during drought stress, the ethylene production of oilseed rape plants increased intensively, reaching 114.79 nl g-1 h-1 on the 8th day of drought, compared to 59.83 nl g-1 h-1 in the irrigated control. Proline and GABA compounds applied alone significantly reduced ethylene formation during drought stress: 82.70 and 60.53 nl g-1 h-1 on the 8th day of drought respectively. After resumption of irrigation, ethylene emission of Proline-treated plants, and especially GABA, decreased much faster and reached the level of irrigated plants within 4 days (Figure 1)
2.6. Impact of Two Amino Acids on Biochemical Responses of Oilseed Rape Exposed to Prolonged Drought
2.6.1. H2O2
H2O2 concentrations increased significantly on days 4 and 8 of the drought, with 8.66 and 16.03 nmol g-1 FW, respectively, compared with 4.07-4.79 nmol g-1 FW in the well-watered control plants (Figure 1). Pre-drought treatment of oilseed rape plants with GABA, proline and a mixture of proline+GABA resulted in a decrease in H2O2 content. On the eighth day of the drought, the lowest H2O2 content (10.48 nmol g-1 FW) was recorded in oilseed rape plants treated with GABA before the drought. All the treatments had a positive effect and the oilseed rape plants recovered from the drought stress.
2.6.2. MDA
The accumulation of MDA in rapeseed leaves increased continuously during drought: from 20.96 nmol g-1 FW detectable on the 4th day of drought to 38.28 nmol g-1 FW on the 8th day of drought. Meanwhile, lipid peroxidation in drought-stressed oilseed rape plants that were sprayed with GABA before stress was the lowest: 16.41 and 24.39 nmol g-1 FW, on days 4 and 8, respectively (Figure 1). In the test variant proline+GABA, lower MDA content was determined only on the 8th day of drought compared to plants affected only by drought. As plants recovered from drought, a similar decrease in MDA levels was recorded in plants treated with amino acids and their mixture.
2.6.3. Free Proline
The accumulation of free endogenous proline in plants exposed to 4 days of drought stress was 8.32 µmol g-1 FW and after 8 days of drought increased to 12.38 µmol g-1 FW, wheal in control plants it was 0.61 0.68 µmol g-1 FW (Figure 1). After 8 days of drought, the highest amount of free proline was detected in the variant sprayed with GABA and proline (sprayed separately) 16.6 and16.3 µmol g-1 FW, respectively. The results showed that the free proline content significantly increases as the drought period continues and decreases with the start of irrigation resumption.
2.6.4. PM ATPase Activity
Drought slightly increased PM ATPase activity on the fourth day after the start of drought treatment. It should be noted here that plants sprayed with GABA retained similar enzyme activity to those not exposed to drought. The drought significantly reduced the PM ATPase activity of oilseed rape leaves on the 8th day of the drought, except for GABA, which increased the activity by an average of 53 % compared to drought control and reached 20.01 μmol Pi mg-1 protein h-1. On the 4th day of irrigation resumption, the PM ATPase activity of GABA- and proline-treated plants increased by 44 % and 56 %, respectively, compared to the drought control, and was significantly higher than that of continuously irrigated plants (Figure 1).
2.6.5. Effect of Amino Acids on Four Genes Expression Levels of Oilseed Rape Exposed to Prolonged Drought
After 4 and 8 days of drought, the expression of all four genes were increased in the drought affected oilseed rape plants with no additional treatments compared to the watered control. The weakest increase in expression showed ABI5 gene after eight days of drought treatment, but remarkable strong upregulation showed KIN2-like gene after 4 days of drought stress (Figure 2). Amino acids proline, GABA and their mix reduced the effect of drought. Moreover, there were also differences in gene expression levels at different days after treatment. The proline treatment almost did not increase gene expression of all four genes tested, compared to corresponding watered control (except EDL3, which expression increased by 3,7 times) after four days of drought. Meanwhile GABA treated plants at the same time showed increased gene expression compared to the corresponding watered control and the expression was always significantly higher compared to the drought affected proline treated plants. After eight days of drought treatment, we observed the strong increase of gene expression in proline treated plants compared to corresponding watered and untreated drought controls and it always exceeded gene expression levels in GABA treated plants. This occurred even though the increase in gene expression in GABA treated plants was almost always higher after eight days, compared to the four days of drought stress (except of caleosin/peroxygenase 3 gene expression).
2.6.6. Impact of Exogenous GABA and Proline on Survival of Plants
After 8 days of simulated drought stress and 4 days of irrigation recovery, the number of oilseed rape plants treated with GABA and proline was significantly higher (on average 42%) compared to plants treated with drought stress alone. The number of plants treated with GABA + proline was lower than those treated with GABA and proline alone and these differences were statistically significant (Figure 3).
3. Discussion
Prolonged drought reduces the relative water content of plants and leads to various metabolic and photosynthetic disturbances [13,24]. In response to drought stress, the accumulation of some amino acids, such as GABA and proline, can provide plant protection against drought damage by maintaining cellular water balance and cell membrane stability through osmotic regulation. In addition, as drought limits cell growth due to a reduction in turgor pressure, this abiotic stress can affect the formation of above-ground plant mass [15]. In current study, we showed that water deficit negatively affected fresh weight (FW) parameters of oilseed rape shoots and was dependent on prolonged drought. Shoot FW was significantly higher when drought-stressed plants were sprayed with 0.1 mM GABA or 1 mM proline. According to the literature, the RWC is considered as an indicator of plant water status and dehydration [25,26]. It predicts the physiological state of the plant and indicates the plant’s ability to retain water under adverse conditions [27,28], so, can be used as a criterion for evaluating the effects of various compounds used to enhance plant drought tolerance [29]. In our study, RWC was a primally reliable indicator for evaluating the effect of GABA and proline in response to prolonged drought of oilseed rape. The results showed that the RWC of drought stressed oilseed rape leaves was significantly reduced in agreement with other studies [30,31,32,33]. Under well-watered conditions, foliar treatment with amino acids did not affected leaf RWC, but it significantly increased by 30% and 58% in GABA and proline + GABA treated plants after 4 and 8 days of prolonged drought, respectively. At 4 days of rewatering, the RWC of previously drought-stressed plants increased to the nonstress level, which was not significantly different among GABA, proline, GABA + proline, treatments, indicating that all plants were fully rehydrated on rewatering.
Numerous studies have shown that oxidative stress caused by water deficit significantly reduces photosynthesis and pigments in plants [29,34,35,36]. This was confirmed by our results, where chlorophyll a and b content in oilseed rape leaves decreased to 0.54 mg g-1 FW and 0.11 mg g-1 FW after 8 days of drought, while in the irrigated control it was 1.27 mg g-1 and 0.33 mg g-1 FW, respectively. Moreover, exogenous application of GABA and proline prior to the drought resulted in a significant improvement of photosynthetic pigments, not only during prolonged drought, but also during the recovery of the plants from the resumption of watering. This confirms the results of other researchers, where exogenous GABA treatment significantly improved chlorophyll and carotenoid content in pepper [29], while proline treatment improved chlorophyll content in the leaves of wheat [9], barley [37], and rice [38] under drought conditions.
Furthermore, several researchers found that decrease in chlorophyll content was due to over-production of reactive oxygen species (ROS), degradation in nutritional balance and inactivation of enzyme activities, as well as a decline in cellular water contents [30,39]. H2O2 is ROS produced by cellular metabolism and is an indicator of the ROS scavenging capacity of plants under stress. It is often found in the literature that plants under drought stress produce ROS such as H2O2 [40,41]. Our study showed that H2O2 concentration increased 3.3-fold during 8 days of drought compared to watered control. Similar drought-induced oxidative stress has been reported in other authors’ studies on oilseed rape plants [31,33]. In the present current study, pre-drought spraying of oilseed rape with GABA, proline and proline+GABA resulted in a decrease in H2O2 content under prolonged drought simulation and had a positive effect on plant recovery resuming irrigation. Meanwhile, we find evidence that exogenous stress protecting compounds significantly reduced H2O2 accumulation under drought stress conditions [33,42]. Excessive concentrations of free radicals, including H2O2, can cause damage to cell membranes, ion leakage and osmotic imbalances, so it is essential to maintain their levels [30,31]. The results indicate that GABA might have a role in preventing oxidative damage caused by increased accumulation of H2O2 during prolonged drought stress.
In addition, we recorded high levels of H2O2 in drought-stressed oilseed rape plants, leading to an increase in the production of MDA, an indicator of oxidative stress. The accumulation of MDA in oilseed rape leaves increased steadily during the drought, from 20.96 nmol g-1 FW detected on the 4th day of the drought to 38.28 nmol g-1 FW on the 8th day of the drought. These findings were consistent with the results obtained in the literature, which indicate that drought induces overproduction of ROS and oxidative stress in a wide range of plant species. According to Saha et al. [43], a significant increase in MDA content was observed in rice plants after the 8th day of drought. Hasanuzzaman et al. [31] monitored the extent of lipid peroxidation (MDA content) in drought-stressed oilseed rape plants. There are several studies that exogenous amino acids (GABA, proline) can effectively reduce MDA levels during drought stress in cultivated plants [10,44]. Our studies showed that exogenous treatment of oilseed rape with GABA or proline resulted in lower oxidative damage: on the 4th day of drought, MDA levels were 22 % and 16 % lower than in control plants, respectively. As the drought persisted longer, lipid peroxidation was significantly lower in the leaves of both proline (31 %) and GABA (36 %) treated plants, alone and in their combination (21 %).
Several reports suggest that PM ATPase plays a key role in plant growth and development and is an important component of membrane integrity in response to abiotic stress [45,46,47]. There is also contrary evidence in the literature indicating that H+-ATPase activity can increase [48] or decrease [49] when plants are exposed to drought stress. Furthermore, we find relevant data that treatment of leaves with stress-protective compounds such as polyamines significantly increased PM ATPase activity by 43.79 % compared to untreated plants under drought stress. The results of our study showed that H+-ATPase activity was significantly reduced in the drought control samples. Also, drought on the 8th day of drought resulted in a significant decrease in the PM ATPase activity of rapeseed leaves, with the exception of GABA, which increased its activity by an average of 53% compared to the drought control, reaching 20.01 μmol Pi mg-1 protein h-1. On the fourth day of irrigation renewal, the PM ATPase activity of GABA- and proline-treated plants increased by 44% and 56%, respectively, compared to the drought control and was significantly higher than that of continuously irrigated plants. Furthermore, it was found that complex application of canola with GABA and proline had a positive effect on the H+-ATPase activity, but it was still lower.
A range of osmotically active molecules such as proline accumulated to balance the water relations under drought stress and in this case is most extensively studied osmolyte [50,51,52]. Numerous studies have reported that exogenous application of proline has shown positive effects on plants growing under drought stress, promoting growth and antioxidant activity [53,54]. Moreover, we find data that exogenous application of amino acids GABA can increase free endogenous proline levels in plant tissues [56], but effects of this amino acid on oilseed rape in improving resistance to drought has not been investigated. In our research we found that exogenous GABA treatment reduced the concentration of proline in rapeseed leaves under drought stress, while after a prolonged drought, plants sprayed with exogenous GABA had a significantly higher endogenous proline content than untreated plants. It was also found that the plants exposed to exogenous proline, according to the amount of free proline in the leaf tissues, were slightly lower than those exposed to GABA. The results on the free proline content showed that the proline content significantly increases as the drought period continues and decreases with the start of watering. Therefore, we proposed the positive effects of exogenous GABA and proline on alleviating the oxidative stress associate with proline accumulation and homeostasis in oilseed rape plants under drought conditions.
Several publications indicate that under drought stress conditions, increased level of ethylene in many plant species [56,57]. Ethylene production after drought stress can reduce photosynthesis, inhibit root growth, and reduce shoot/leaf expansion [58]. Our data contribute to the suggestion that exogenous amino acids affecting ethylene content may help to remove the inhibitory effect of drought stress on plant growth. Ethylene emission analysis showed that during drought stress, the amount of ethylene produced by rapeseed plants increased intensively and reached 114.79 nl g-1 h-1 on the 8th day of drought, while in the irrigated control plants it was 59.83 nl g-1 h-1. Meanwhile, GABA or proline significantly reduced ethylene formation during drought stress. After resuming watering, the ethylene emission of GABA-treated plants decreased much faster and reached the level of irrigated plants within 4 days. A mixture of proline+GABA resulted in an increase in ethylene accumulation, but still was lower than in drought control. These data agreed with the results obtained by Sharma et al. [59] that changes in ethylene content during drought can activate the plant antioxidant defense system, resulting in a reduction of oxidative stress, with accompanied recovery of plant growth and photosynthetic efficiency.
Drought stress affects and alters the expression of many genes. The four putative drought responsive genes, KIN2-like, caleosin/peroxygenase 3, EDL 3 (EID 1 like F-box protein 3), and ABI 5 were chosen as a markers to understand the patterns of action of proline and GABA on the genetic level in the response of rapeseed to drought stress. It is known that these genes increase their expression in response to drought stress conditions [18,20,21,22,60]. As expected, the expression of all four genes were increased in the simulated drought affected oilseed rape plants with no additional treatments compared to the watered control. The weakest increase in expression showed ABI5 gene after eight days of drought treatment, but remarkable strong upregulation showed KIN2-like gene after 4 days of drought stress, what is consistent with the data obtained by Zhang et al. [19]. Also, this is consistent with our obtained data showing that plants without additional treatment suffered from drought after 4 and 8 days of drought application. Proline, GABA and their mix reduced the effect of drought. Interestingly, although both amino acids diminished the effect of drought to similar levels, the response at the gene expression level differed between proline and GABA treated plants. It seems that proline and GABA act differently under drought conditions at the genetic level. It is known that proline can protect plants from environmental stresses in multiple ways: acting as osmolyte and molecular chaperone, through ROS scavenging activity, or as a donor of electrons in mitochondria [61]. So, it can be speculated that application of exogenous proline to rapeseed initially enhances the proline metabolic pool inside, which in turn increases plant drought tolerance to such level, that plants do not need to increase the expression of the genes necessarily for resistance to drought. But later exogenous proline’s drought protective properties weaken, and the drought-dependent genes are upregulated. The induction of drought responsive genes could be promoted also by proline catabolism, which could be enhanced in the case of exogenous proline application. Indeed, it was shown, that some products of proline catabolism could induce osmotically regulated genes in rice [62]. The different situation is with GABA. Although the GABA like proline can protect plants from drought acting through various physiological and biochemical mechanisms [12], it seems that such GABA action is not enough for plant to protect against water deficit, so the genes responsible for drought resistance were upregulated already in the early stages of drought. From the other hand the lower expression level of drought-induced genes under the GABA treatment compared to proline treated plants after eight days of drought could indicate, that GABA function as a drought protective molecule at physiological and biochemical level may continue longer comparing to proline. So, in this case the plants did not need to maintain very strong drought-inducible gene expression in order to achieve the same protection against drought. Finally in the plants treated with proline and GABA simultaneously, gene expression level was almost always intermediate between gene expression in proline treated and GABA treated plants separately. It seems that their action is not additive. In the recovery phase the expression of tested genes was often at low levels, not even reaching the level of corresponding watered control, indicating that tested genes are not important for plant recovery. The exception made KIN 2-like gene, which in additionally untreated recovering plants showed surprisingly high expression. Interestingly, GABA treated plant’s gene expression in the recovery phase almost always was slightly higher compared to the other variants, though this may not be related to the recovery from drought conditions.
Thus, in oilseed rape, various molecular, biochemical, physiological and morphological processes are damaged under drought stress, but the plant response to amino acid application can induce biochemical and cellular changes in the tissues that ensure survival. We found that exogenous GABA and proline improved the survival of oilseed rape seedlings after simulated drought stress, and the number of plants surviving after resumed irrigation was significantly higher compared with plants exposed to drought alone.
4. Materials and Methods
4.1. Research Object and Growth Conditions
Winter oilseed rape (Brassica napus L.) cv. ‘Visby’ seeds were sown in the pots with a peat moss (pH 5.5–6.5). Plants were grown under controlled conditions in a Climacell plant growing chamber (Medcenter Einrichtungen GmbH) at 23 °С and 16/8 hours (light/darkness) photoperiod at a light intensity of 75 μmol m−2 s−1. Soil moisture was maintained at ~70%. Moisture was measured with a soil moisture meter (Biogrod, China).
4.2. Treatments
4.2.1. Spraying with Amino Acids
Aqueous solutions of amino acids L-proline (Roth), GABA (Sigma-Aldrich) and a mixture of L-proline and GABA (Sigma-Aldrich) were used to spray the seedlings at the 3-4 leaf stage, BBCH scale 13-19 [63]. The concentration range was chosen based on literature data and our previous experiments [64,65,66]. Spraying of the plants was carried out using a hand-held sprayer, at a dose of 3 ml and 12.5 ml in each pot. Control plants were sprayed with the same volume of distilled water.
4.2.2. Drought Simulation
For the drought stress control studies, plants were subjected to prolonged drought stress for 8 days to reach a high water deficit. During the simulated drought, irrigation was interrupted to allow the soil to dry out gradually.
4.2.3. Irrigation Renewal
After 8 days of prolonged drought, irrigation was resumed to achieve a soil moisture content of 70%. Plant recovery was assessed after 4 days.
4.3. Determination of the Active GABA Concentration
To determine the active GABA concentration, seeds of oilseed rape were sown in 16x3 pots with peat substrate. Each experimental unit consisted of 14 seeds. Pots without GABA served as control. Three treatments of the study of GABA impact were used: 1) control, H2O 3 ml per pot; 2) GABA 0.1 mmol 3 ml per pot; 3) GABA 1 mmol 3 ml per pot; 4) GABA 10 mmol·3 ml per pot; 5) control, H2O 12.5 ml per pot; 6) GABA 0.1 mmol 12.5 ml per pot; 7) GABA 1 mmol 12.5 ml per pot; 8) GABA 10 mmol·12.5 ml per pot, other eight treatments were the same plus drought treatment. Seedlings at the BBCH 13–14 third-fourth leaves stage were foliar sprayed with water solutions of GABA, control was sprayed with water.
4.4. Experimental Design
Eight pots were used for the experiment, four for rational watering and four for drought simulation. The experiment was repeated three times with eight pots each. Aqueous solutions of L-proline 0.1 mmol, GABA 0.1 mM 12.5 mL and mix of both amino acids with the same concentration 12.5 mL per pot were sprayed according to the following scheme: (1) control watering, (2) proline and watering, (3) GABA and watering, (4) proline + GABA and watering, (5) drought, (6) proline and drought. (7) GABA and drought, (8) proline + GABA and drought (Figure 4).
4.5. Sampling
Plant leaves were sampled three times for testing: on day 4 of drought (at 40% soil moisture), on day 8 of drought (at 20% soil moisture), and on day 4 of plant recovery after watering (at 70% soil moisture). Watered plants used as controls were sampled at the same time. Thirty seedlings were sampled for morphometric measurements. For biochemical analysis, three independent replicates were performed using fully expanded leaves of oilseed rape plants. Freshly harvested leave samples were used for ethylene emission analysis and pigment measurement. For H+-ATPase activity, MDA, H2O2 and proline assays, samples were immediately frozen in liquid nitrogen and stored in a low-temperature freezer (Skadi Green line, ES) at -80 °C until analysis. Surviving seedlings were counted immediately after 41 days of cultivation.
4.6. Determination of RWC
The RWC was determined according to Hodges et al. [67] as amended [28]. Leaves from the same part of each plant were picked and weighed (FW), then placed in distilled water for 24 hours to allow the leaves to become saturated with water and weighed again (SW), then wrapped in craft paper, dried at 80 ºC to a constant weight, and the dry weight was determined (DW). The RWC of the leaves was determined according to the formula:
RWC (%) = (FW − DW)/(SW − DW) × 100%,
4.7. Determination of Plant Survival
After resumed watering, plant survival, i.e., the proportion of plants that retained the ability to grow at the stem apex, was determined.
4.8. Assessment of Biochemical Parameters
4.8.1. Photosynthetic Pigments
The photosynthetic pigments were extracted from fresh leaves with N, N’-dimethyl-formamide (DMF) (Sigma-Aldrich). Light absorption was measured at 480, 664, 647 nm. Chlorophyll a and b contents were calculated according to Wellburn [68].
4.8.2. MDA Content
According to the method of Li [69], 0.3 g of leaves was crushed in a mortar containing 5 mL of 1% trichloroacetic acid (TCA). The extract was centrifuged at 10,000 g for 5 min at 4 °C. Then, 0.3 mL of the supernatant was mixed with 1.2 mL of MDA solution which contained 20% TCA and 0.5% thiobarbituric acid (TBA). The resulting mixture was placed in a warm water bath at 95 °C for 30 min. It was then cooled immediately and centrifuged again at 10,000 g for 10 min. The absorbance of the supernatant was read using a spectrophotometer at 532 nm.
4.8.3. H2O2 Content
To measure the leaf H2O2 content based on the Alexieva et al. [70] method, 0.15 g of leaves was ground in a mortar containing 5 mL of 0.1% (v/v) TCA. The obtained extract was centrifuged at 10,000 g for 5 min. Then, 25 μL of the supernatant was mixed with 250 μL of 100 mM phosphate buffer (pH 7) and 500 μL of 1 M potassium iodide (KI). Finally, the absorbance was read at 390 nm with a spectrophotometer.
4.8.4. Ethylene Emission
Ethylene emission from freshly harvested leaves was estimated using the method of Child et al. [71]. Weighed leaf samples were placed in 40 ml clear glass vials (Agilent technology) sealed with PTFE/Si septa caps and incubated for 24 hours at 21 °C in the dark. After incubation, 1 ml of the gas sample from each vial was withdrawn with a gas-tight syringe (Agilent technology) and injected into a gas chromatograph equipped with a stainless steel column (Propac R) and a hydrogen flame ionisation detector. The injector, column and detector temperatures were 110, 90 and 150 °C respectively. Helium (AGA) was used as carrier gas. Calibration was carried out with ethylene standard (Messer). The results are expressed in nl g-1 FW h-1.
4.8.5. H+-ATPase Activity Assay
A membrane-enriched fraction of microsomes was isolated from plant samples. Protein content was measured using the Bradford dye binding procedure at 595 nm. The H+-ATPase activity of the microsomal fraction was assessed by the re-isolation of inorganic phosphate (Pi), which accumulates as a result of ATP hydrolysis [72]. The colour reaction to measure Pi was performed with ammonium molybdate and stannous chloride at 750 nm. Enzyme activity was expressed as μmol Pi mg-1 protein h-1.
4.8.6. Proline
Free proline content was assessed with a ninhydrin-based method using a cuvette spectrophotometer (Analytik Jena Specord 210 Plus). Proline was extracted by heating plant aliquots of ground plant material (0.5 g) for 20 min in pure ethanol. The resulting mixture was left overnight at 4°C, and then centrifuged at 14000 g (5 min). Color reaction mix was ninhydrin 1% (w/v) in acetic acid 60% (v/v), ethanol 20% (v/v) [73]. The absorbance was read spectrophotometrically at 520 nm. The corresponding content of proline was determined using the standard curve. Results were expressed as µmol of proline µmol g-1 FW.
4.9. Molecular Techniques
4.9.1. RNA Extraction and Reverse Transcription
Free proline content was assessed with a ninhydrin-based method using a cuvette spectrophotometer (Analytik Jena Specord 210 Plus). Proline was extracted by heating plant aliquots of ground plant material (0.5 g) for 20 min in pure ethanol. The resulting mixture was left overnight at 4°C, and then centrifuged at 14000 g (5 min). Color reaction mix was ninhydrin 1% (w/v) in acetic acid 60% (v/v), ethanol 20% (v/v) [73]. The absorbance was read spectrophotometrically at 520 nm. The corresponding content of proline was determined using the standard curve. Results were expressed as µmol of proline µmol g-1 FW.
4.9.2. Real-Time Quantitative PCR
Real-time quantitative PCR was carried out using Maxima SYBR Green qPCR Master Mix (2X) kit (Thermo Scientific) following the manufacturer’s recommendations. Cycling conditions comprised one cycle at 95 °C for 2 min and 40 cycles at 95 °C for 15 s followed by 60 °C for 1 min. PCR amplification was performed in an Azure Cielo (USA) real-time PCR machine. Gene expression was calculated using 2-ΔΔCt method [74].
4.9.3. Primers
The sequences of primers used in the work were taken from [18]. Two, under different experimental conditions, commonly used reference genes of actin 7 (ACT7) and ubiquitin-conjugating enzyme 21 (UBC21) [75], were used for gene expression analysis. The sequences of all primers used in the work are listed in the Table 5.
4.10. Statistical Analysis
The results are presented as mean ± standard deviation (SD) of three independent experiments with at least three replicates. The data were analysed using analysis of variance (ANOVA). Tukey’s test was performed to test the statistical significance of differences (p <0.05) between means. Post hoc test Duncan’s Multiple Range Test (DMRT) was used for comparing the means of different groups after performing an analysis of variance.
5. Conclusions
Exogenously added amino acids (GABA and proline) reduced drought-induced damage to oilseed rape. GABA and proline applied alone or in combination, alleviated drought stress in oilseed rape by improving water status, photosynthetic pigments and osmoprotective substances accumulation, and membrane integrity in leaf tissue. Plants exposed to amino acids produced large amounts of endogenous proline when exposed alone. In this study, different effects of GABA, proline and GABA+proline on plant drought tolerance and recovery from drought were found. GABA contributed mainly to the enhancement of plant tolerance to drought stress, whereas proline was more effective in promoting drought recovery compared to GABA. The tested amino acids reduced the simulation of ethylene emission in drought stressed seedlings leaves. A higher amount of ethylene emission was observed under GABA + proline exposure. Molecular studies of drought responsive genes revealed that GABA and proline can protect plants from drought acting through KIN2-like, caleosin/peroxygenase 3, EDL 3 and ABI 5 gene expression. Therefore, the application of GABA is an alternative to proline technique for improving oilseed rape tolerance to drought and could be used to increase the amount and quality of crops.
Author Contributions
Conceptualization, S.J. and V.G.; methodology, R.M., J.J. and V.Š.; formal analysis, R.M., E.J.-B. and T.Ž.; investigation, R.M., J.J. and E.J.-B.; data curation, E.J.-B., L.K. and V.Š.; writing—original draft preparation, R.M., V.Š. and V.G.; writing—review and editing, S.J., V.G. and L.K.; visualization, R.M., T.Ž. and S.J.; supervision, L.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Research Council of Lithuania (LMTLT), agreement No S-LU-22-4 and by the Ministry of Education and Science of Ukraine (Joint Ukrainian-Lithuanian R&D project, MESU M/49-2023).
Data Availability Statement
The data supporting the reported results can be found in the archive of scientific reports of the Nature Research Centre.
Acknowledgments
We thank engineer Nijolė Bareikienė and all the staff of the Laboratory of Plant Physiology of the Nature Research Centre for the support and help provided.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| GABA | γ -aminobutyric acid |
| RWC | Relative water content |
| MDA | malondialdehide |
| FW | fresh weight |
| DW | dried weight |
| SW | saturated weight |
| TCA | Trichloroacetic acid |
References
- Tesfamariam, E.H.; Annandale, J.G.; Steyn, J.M. Water stress effects on winter canola growth and yield. Agron. J. 2010, 102, 658–666. [Google Scholar] [CrossRef]
- Ionita, M.; Nagavciuc, V.; Kumar, R.; Rakovec, O. On the curious case of the recent decade, mid-spring precipitation deficit in Central Europe. NPJ Clim. Atmos. Sci. 2020, 3, 49. [Google Scholar] [CrossRef]
- Hassan, U.M.; Aamer, M.; Umer Chattha, M.; Haiying, T.; Shahzad, B.; Barbanti, L. ; Critical role of zinc in plants facing the drought stress. Agriculture 2020, 10, 396. [Google Scholar] [CrossRef]
- Huan, L.; Jin-Qiang, W.; Qing, L. Photosynthesis product allocation and yield in sweet potato with spraying exogenous hormones under drought stress. J. Plant Physiol. 2020, 253, 153265. [Google Scholar] [CrossRef] [PubMed]
- Doneva, D.; Pál, M.; Brankova, L.; Szalai, G.; Tajti, J.; Khalil, R.; Peeva, V. The effects of putrescine pre-treatment on osmotic stress responses in drought-tolerant and drought-sensitive wheat seedlings. Physiol. Plant. 2021, 171, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Rady, M.O.A.; Marey, R.A.; Abd El-Mageed, T.A. Exogenously applied proline enhances growth and productivity of drought stressed onion by improving photosynthetic efficiency, water use efficiency and up-regulating osmoprotectants. Sci. Hortic. 2020, 272, 109580. [Google Scholar] [CrossRef]
- Trovato, M.; Funck, D.; Forlani, G.; Okumoto, S.; Amir, R. Amino acids in plants: Regulation and functions in development and stress defense. Front. Plant Sci. 2021, 12, 772810. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of exogenous proline to abiotic stress tolerance in plants: A review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Farooq, M.; Nawaz, A.; Chaudhry, M.; Indrasti, R.; Rehman, A. Improving resistance against terminal drought in bread wheat by exogenous application of proline and gamma-aminobutyric acid. J. Agron. Crop Sci. 2017, 203, 464–472. [Google Scholar] [CrossRef]
- Shelp, B.J.; Aghdam, M.S.; Flaherty, E.J. γ-Aminobutyrate (GABA) Regulated Plant Defense: Mechanisms and Opportunities. Plants 2021, 10, 1939. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.M.; Alabdallah, N.M.; Alharbi, B.M.; Waseem, M.; Yao, G.; Liu, X.D. GABA: a key player in drought stress resistance in plants. Int. J. Mol. Sci. 2021, 22, 10136. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, J.; Peng, Y.; Huang, B. Metabolic pathways regulated by abscisic acid, salicylic acid and γ-aminobutyric acid in association with improved drought tolerance in creeping bentgrass (Agrostis stolonifera). Physiol. Plant. 2017, 159, 42–58. [Google Scholar] [CrossRef] [PubMed]
- Yong, B.; Xie, H.; Li, Z.; Li, Y.P.; Zhang, Y.; Nie, G.; Peng, Y. Exogenous application of GABA improves PEG-induced drought tolerance positively associated with GABA-shunt, polyamines, and proline metabolism in white clover. Front. Physiol. 2017, 8, 1107. [Google Scholar] [CrossRef] [PubMed]
- Razik, A.E.S.; Alharbi, B.M.; Pirzadah, T.B.; Alnusairi, G.S.H.; Soliman, M.H.; Hakeem, K.R. γ-Aminobutyric acid (GABA) mitigates drought and heat stress in sunflower (Helianthus annuus L.) by regulating its physiological, biochemical, and molecular pathways. Physiol. Plant. 2020, 172, 505–527. [Google Scholar] [CrossRef]
- Buffagni, V.; Vurro, F.; Janni, M.; Gullì, M.; Keller, A.A.; Marmiroli, N. Shaping durum wheat for the future: Gene expression analyses and metabolites profiling support the contribution of BCAT genes to drought stress response. Front. Plant Sci. 2020, 11, 891. [Google Scholar] [CrossRef]
- Al-Quraan, N.A.; Samarah, N.H.; Tanash, A.A. Effect of drought stress on wheat (Triticum durum) growth and metabolism: Insight from GABA shunt, reactive oxygen species, and dehydrin genes expression. Funct. Plant Biol. 2024, 51, 22177. [Google Scholar] [CrossRef]
- Zhang, J.; Mason, A.S.; Wu, J.; Liu, S.; Zhang, X.; Luo, T.; Redden, R.; Batley, J.; Hu, L.; Yan, G. Identification of putative candidate genes for water stress tolerance in canola (Brassica napus). Front. Plant Sci. 2015, 6, 1058. [Google Scholar] [CrossRef]
- Hundertmark, M.; Hincha, D.K. LEA (late embryogenesis abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics 2008, 9, 118. [Google Scholar] [CrossRef]
- Kurkela, S.; Borg-Franck, M. Structure and expression of kin2, one of two cold- and ABA-induced genes of Arabidopsis thaliana. Plant Mol. Biol. 1992, 19, 689–692. [Google Scholar] [CrossRef]
- Partridge, M.; Murphy, D.J. Roles of a membrane-bound caleosin and putative peroxygenase in biotic and abiotic stress responses in Arabidopsis. Plant Physiol. Biochem. 2009, 47, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Koops, P.; Pelser, S.; Ignatz, M.; Klose, C.; Marrocco-Selden, K.; Kretsch, T. EDL3 is an F-box protein involved in the regulation of abscisic acid signalling in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 5547–5560. [Google Scholar] [CrossRef] [PubMed]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The role and regulation of ABI5 (ABA-Insensitive 5) in plant development, abiotic stress responses and phytohormone. Front. Plant Sci. 2016, 7, 1884. [Google Scholar] [CrossRef]
- Vanani, F.R. Comparative physiological and proteomic analysis indicates lower shock response to drought stress conditions in a self-pollinating perennial ryegrass. PLoS ONE 2020, 15, e0234317. [Google Scholar] [CrossRef]
- Kamanga, R.M.; Mbega, E.; Ndakidemi, P. Drought tolerance mechanisms in plants: physiological responses associated with water deficit stress in Solanum lycopersicum. ACST 2018, 6, 362. [Google Scholar] [CrossRef]
- Qayyum, A.; Al Ayoubi, S.; Sher, A.; Bibi, Y.; Ahmad, S.; Shen, Z.; Jenk,s M. A. Improvement in drought tolerance in bread wheat is related to an improvement in osmolyte production, antioxidant enzyme activities, and gaseous exchange. Saudi. J. Biol. Sci. 2021, 28, 5238–5249. [Google Scholar] [CrossRef]
- Sultan, M.A.R.F.; Hui, L.; Yang, L.J.; Xian, Z.H. Assessment of drought tolerance of some Triticum L. species through physiological indices. Czech J. Genet. Plant Breed. 2012, 48, 178–184. [Google Scholar] [CrossRef]
- Weng, M.; Cui, L.; Liu, F.; Zhang, M.; Shan, L.; Yang, S.; Deng, X.P. Effects of drought stress on antioxidant enzymes in seedlings of different wheat genotypes. Pak. J. Bot. 2015, 47, 49–56. [Google Scholar]
- Iqbal, B.; Hussain, F.; Khan, M.S.; Iqbal, T.; Shah, W.; Ali, B.; Al Syaad, K.M.; Ercisli, S. Physiology of gamma-aminobutyric acid treated Capsicum annuum L. (Sweet pepper) under induced drought stress. PLoS ONE 2023, 18, e0289900. [Google Scholar] [CrossRef]
- Akram, N.A.; Iqbal, M.; Muhammad, A.; Ashraf, M.; Al-Qurainy, F.; Shafiq, S. Aminolevulinic acid and nitric oxide regulate oxidative defense and secondary metabolisms in canola (Brassica napus L.) under drought stress. Protoplasma 2018, 255, 163–174. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Khan, M.I.R.; Fujita, M. Silicon-mediated regulation of antioxidant defense and glyoxalase systems confers drought stress tolerance in Brassica napus L. S. Afr. J. Bot. 2018, 115, 50–57. [Google Scholar] [CrossRef]
- Sharif, P.; Seyedsalehi, M.; Paladino, O.; Van Damme, P.; Sillanpää, M.; Sharifi, A.A. Effect of drought and salinity stresses on morphological and physiological characteristics of canola. Int. J. Environ. Sci. Technol. 2018, 15, 1859–1866. [Google Scholar] [CrossRef]
- Ayyaz, A.; Farooq, M.A.; Dawood, M.; Majid, A.; Javed, M.; Athar, H.U.R.; Zafar, Z.U. Exogenous melatonin regulates chromium stress-induced feedback inhibition of photosynthesis and antioxidative protection in Brassica napus cultivars. Plant Cell Rep. 2021, 40, 2063–2080. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; He, C.G.; Wang, Y.J.; Bi, Y.F.; Jiang, H. Effect of drought and heat stresses on photosynthesis, pigments, and xanthophyll cycle in alfalfa (Medicago sativa L.). Photosynthetica 2020, 58, 5. [Google Scholar] [CrossRef]
- Mihaljević, I.; Vuletić, V.M.; Šimić, D.; Tomaš, V.; Horvat, D.; Josipović, M.; Zdunić, Z.; Dugalić, K.; Vuković, D. Comparative study of drought stress effects on traditional and modern apple cultivars. Plants 2021, 10, 561. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Selwey, W.A.; Ibrahim, A.A.; Shady, M.; Alsadon, A.A. Foliar applications of ZnO and SiO2 nanoparticles mitigate water deficit and enhance potato yield and quality traits. Agronomy 2023, 13, 466. [Google Scholar] [CrossRef]
- Abdelaal, K.A.; Attia, K.A.; Alamery, S.F.; El-Afry, M.M.; Ghazy, A.I.; Tantawy, D.S.; Al-Doss, A.A.; El-Shawy, E.E.; Abu-Elsaoud, A.M.; Hafez, Y.M. Exogenous application of proline and salicylic acid can mitigate the injurious impacts of drought stress on barley plants associated with physiological and histological characters. Sustainability 2020, 12, 1736. [Google Scholar] [CrossRef]
- Hanif, S.; Saleem, M.F.; Sarwar, M.; Irshad, M.; Shakoor, A. ; Biochemically triggered heat and drought stress tolerance in rice by proline application. J. Plant Growth Regul. 2021, 40, 305–312. [Google Scholar] [CrossRef]
- Razzaq, H.; Tahir, M.H.N.; Sadaqat, H.A.; Sadia, B. Screening of sunflower (Helianthus annus L.) accessions under drought stress conditions, an experimental assay. J. Soil Sci. Plant Nutr. 2017, 17, 662–671. [Google Scholar] [CrossRef]
- Qureshi, M.K.; Munir, S.; Shahzad, A.N.; Rasul, S.; Nouman, W.; Aslam, K. Role of reactive oxygen species and contribution of new players in defense mechanism under drought stress in rice. Int. J. Agric. Biol. 2018, 20, 1339–1352. [Google Scholar]
- Barzotto, G.R.; Cardoso, C.P.; Jorge, L.G.; Campos, F.G.; Boaro, C.S.F. Hydrogen peroxide signal photosynthetic acclimation of Solanum lycopersicum L. cv Micro-Tom under water deficit. Sci. Rep. 2023, 13, 13059. [Google Scholar] [CrossRef]
- Todorova, D.; Sergiev, I.; Katerova, Z.; Shopova, E.; Dimitrova, L.; Brankova, L. Assessment of the biochemical responses of wheat seedlings to soil drought after application of selective herbicide. Plants 2021, 10, 733. [Google Scholar] [CrossRef] [PubMed]
- Saha, I.; De, A.K.; Sarkar, B.; Ghosh, A.; Dey, N.; Adak, M.K. Cellular response of oxidative stress when sub1A QTL of rice receives water deficit stress. Plant Sci. Today 2018, 5, 84–94. [Google Scholar] [CrossRef]
- Vijayakumari, K.; Puthur, J.T. γ-Aminobutyric acid (GABA) priming enhances the osmotic stress tolerance in Piper nigrum Linn. plants subjected to PEG-induced stress. Plant Growth Regul. 2015, 78, 57–67.
- Pottosin, I.; Olivas-Aguirre, M.; Dobrovinskaya, O.; Zepeda-Jazo, I.; Shabala, S. Modulation of ion transport across plant membranes by polyamines: understanding specific modes of action under stress. Front. Plant Sci. 2021, 11, 616077. [Google Scholar] [CrossRef]
- Li, J.; Guo, Y.; Yang, Y. The molecular mechanism of plasma membrane H+-ATPases in plant responses to abiotic stress. J. Genet. Genomics 2022, 49, 715–725. [Google Scholar] [CrossRef]
- Na, C.; Ziwen, Z.; Yeyun, L.; Xianchen, Z. Exogenously applied Spd and Spm enhance drought tolerance in tea plants by increasing fatty acid desaturation and plasma membrane H+-ATPase activity. Plant Physiol. Biochem. 2022, 170, 225–233. [Google Scholar] [CrossRef]
- Feng, X.; Liu, W.; Zeng, F.; Chen, Z.; Zhang, G.; Wu, F. K+ uptake, H+-ATPase pumping activity and Ca2+ efflux mechanism are involved in drought tolerance of barley. Environ. Exp. Bot. 2016, 129, 57–66. [Google Scholar] [CrossRef]
- Chang, N.; Ziwen, Z.; Yeyun, L.; Xianchen, Z. Exogenously applied Spd and Spm enhance drought tolerance in tea plants by increasing fatty acid desaturation and plasma membrane H+-ATPase activity. Plant Physiol. Biochem. 2022, 170, 225–233. [Google Scholar]
- Zivcak, M.; Brestic, M.; Sytar, O. Osmotic adjustment and plant adaptation to drought stress. In Drought Stress Tolerance in Plants. Physiol. Biochem.; 2016; 1, 105–143. 1.
- Khanna-Chopra, R.; Semwal, V.K.; Lakra, N.; Pareek, A. Proline–A key regulator conferring plant tolerance to salinity and drought. In Plant Tolerance to Environmental Stress; CRC Press: 2019; 59–80.
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef]
- Bekka, S.; Abrous-Belbachir, O.; Djebbabar, R. Effects of exogenous proline on the physiological characteristics of Triticum aestivum L. and Lens culinaris Medik. under drought stress. Acta Agric. Slovenica 2018, 111, 477–491. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Rady, M.O.A.; Marey, R.A.; Abd El-Mageed, T.A. Exogenously applied proline enhances growth and productivity of drought stressed onion by improving photosynthetic efficiency, water use efficiency and up-regulating osmoprotectants. Sci. Hortic. 2020, 272, 109580. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Y.; Huang, B. Physiological effects of γ-aminobutyric acid application on improving heat and drought tolerance in creeping bentgrass. J. Am. Soc. Hortic. Sci. 2016, 141, 76–84. [Google Scholar] [CrossRef]
- Khan, M.I.; Iqbal, N.; Masood, A.; Per, T.S.; Khan, N.A. Salicyclic acid alleviates adverse effects of heat stress on photosynthesis through changes in proline production and ethylene formation. Plant Signal Behav. 2013, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Fahad, S. Phytohormones enhanced drought tolerance in plants: a coping strategy. Environ. Sci. Pollut. Res. 2018, 25, 33103–33118. [Google Scholar] [CrossRef]
- Dubois, M.; Van den Broeck, L.; Inzé, D. The pivotal role of ethylene in plant growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Sidhu, G.P.S.; Kumar, R.; Kohli, S.K.; Yadav, P.; Kapoor, D.; Bali, A.S. ; Shahzad, B; Khanna, K, Kumar, S, Thukral, A.K., Eds.; Bhardwaj, R. Abiotic stress management in plants: role of ethylene. Molecular Plant Abiotic Stress: Biology and Biotechnology, 2019, 185–208. [Google Scholar]
- Kobayashi, F.; Maeta, E.; Terashima, A.; Shigeo, T. Positive role of a wheat HvABI5 ortholog in abiotic stress response of seedlings. Physiol. Plant. 2008, 134, 74–86. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: a multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Iyer, S.; Caplan, A. Products of proline catabolism can induce osmotically regulated genes in rice. Plant Physiol. 1998, 116, 203–211. [Google Scholar] [CrossRef]
- Meier, U. Growth stages of mono and dicotyledonous plants. In BBCH Monograph; Meier, U., Ed.; Julius Kühn-Institut: Quedlinburg, Germany, 2018; pp. 85–88. [Google Scholar]
- Ferreira, R.A.; Borella, J.; Hüther, C.M.; Drought-induced stress in leaves of Coix lacryma-jobi L. under exogenous application of proline and GABA amino acids. Braz. J. Bot. 2020, 43, 513–521. [Google Scholar] [CrossRef]
- Zeng, W.; Hassan, M.J.; Kang, D.; Peng, Y.; Li, Z. Photosynthetic maintenance and heat shock protein accumulation relating to γ-aminobutyric acid (GABA)-regulated heat tolerance in creeping bentgrass (Agrostis stolonifera). S. Afr. J. Bot. 2021, 141, 405–413. [Google Scholar] [CrossRef]
- Jurkonienė, S.; Mockevičiūtė, R.; Gavelienė, V.; Šveikauskas, V.; Zareyan, M.; Jankovska-Bortkevič, E.; Jankauskienė, J.; Žalnierius, T.; Kozeko, L. Proline enhances resistance and recovery of oilseed rape after a simulated prolonged drought. Plants 2023, 12, 2718. [Google Scholar] [CrossRef] [PubMed]
- Hodges, D.; DeLong, J.; Forney, C.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Li, H.S. Principles and Techniques of Plant Physiological Biochemical Experiment, 1st ed.; Higher Education Press: Beijing, China, 2000; pp. 94–196. [Google Scholar]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress marker in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Child, R.D.; Chauvaux, N.; John, K.; Van Onckelen, H.A.; Ulvskov, P. Ethylene biosynthesis in oilseed rape pods in relation to pod shatter. J. Exp. Bot. 1998, 49, 829–836. [Google Scholar] [CrossRef]
- Darginavičienė, J.; Pašakinskienė, I.; Maksimov, G.; Rognli, O.A.; Jurkonienė, S.; Šveikauskas, V.; Bareikienė, N. Changes in plasmalemma K+, Mg2+-ATPase dephosphorylating activity and H+ transport in relation to freezing tolerance and seasonal growth of Festuca pratensis Huds. J. Plant Physiol. 2008, 165, 825–832. [Google Scholar] [CrossRef]
- Carillo, P.; Gibon, Y. Extraction and determination of proline. Available online: http://prometheuswiki.publish.csiro.au/tikiindex.php?page=Extraction$+$and$+$determination$+$of$+$proline (accessed on 10 July 2016).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time Quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Han, P.P.; Qin, L.; Li, Y.S.; Liao, X.S.; Xu, Z.X.; Hu, X.J.; Xie, L.H.; Yu, C.B.; Wu, Y.F.; Liao, X. Identification of suitable reference genes in leaves and roots of rapeseed (Brassica napus L.) under different nutrient deficiencies. J. Integr. Agric. 2017, 16, 809–819. [Google Scholar] [CrossRef]
Figure 1.
Impact of GABA and proline application on RWC, ethylene emission, H2O2 level, MDA content, PM ATPase activity, and endogenous proline content of oilseed rape leaves after simulated prolonged drought and recovery by watering. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
Figure 1.
Impact of GABA and proline application on RWC, ethylene emission, H2O2 level, MDA content, PM ATPase activity, and endogenous proline content of oilseed rape leaves after simulated prolonged drought and recovery by watering. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
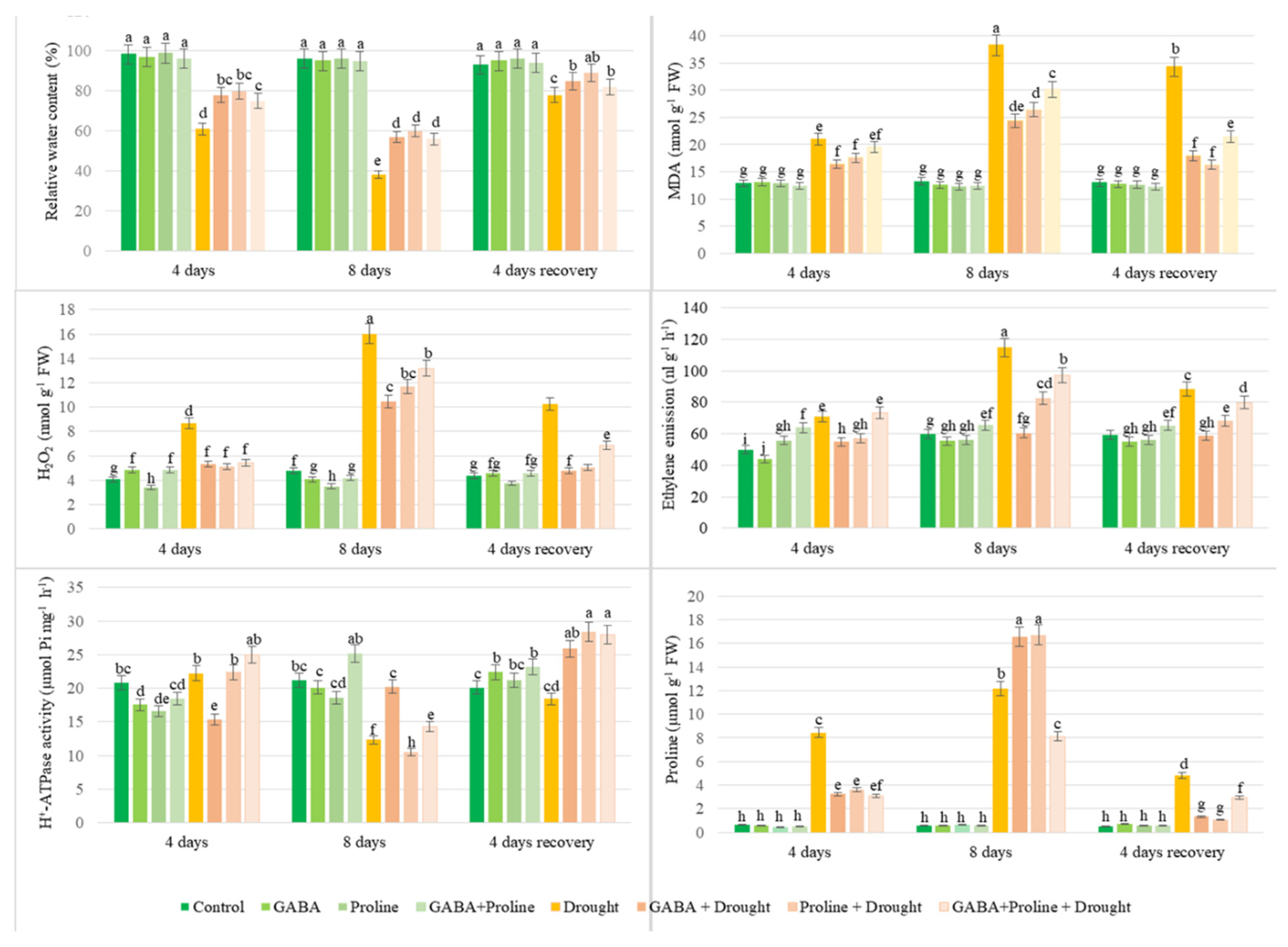
Figure 2.
The effect of exogenous GABA and proline application on oilseed rape drought sensitive gene expression under prolonged drought conditions.
Figure 2.
The effect of exogenous GABA and proline application on oilseed rape drought sensitive gene expression under prolonged drought conditions.
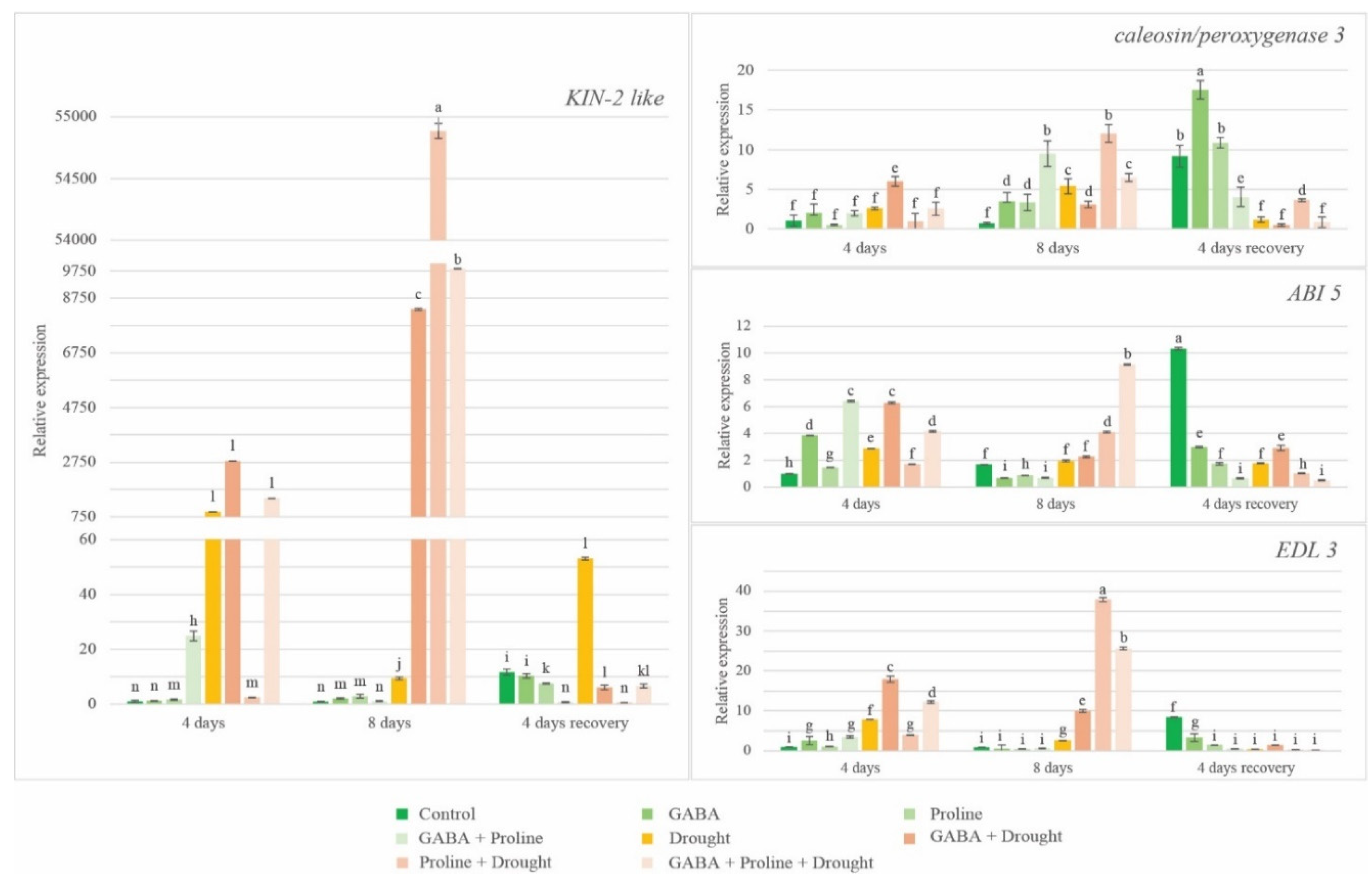
Figure 3.
Effect of exogenous GABA and proline on Brassica napus plant survival after 8 days of prolonged drought re-covered after irrigation.
Figure 3.
Effect of exogenous GABA and proline on Brassica napus plant survival after 8 days of prolonged drought re-covered after irrigation.
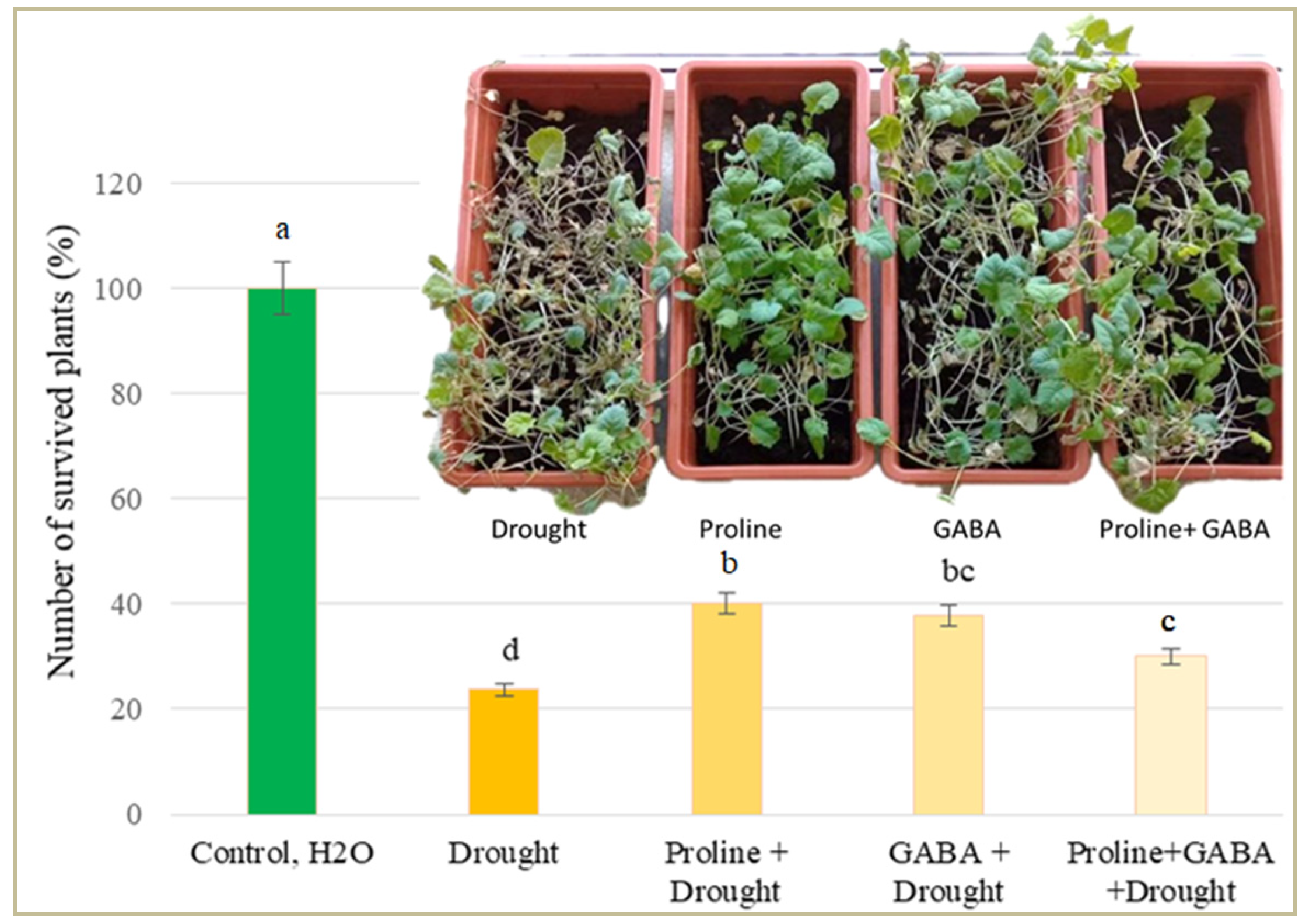
Figure 4.
Effect of exogenous GABA and proline on Brassica napus plant survival after 8 days of prolonged drought re-covered after irrigation.
Figure 4.
Effect of exogenous GABA and proline on Brassica napus plant survival after 8 days of prolonged drought re-covered after irrigation.
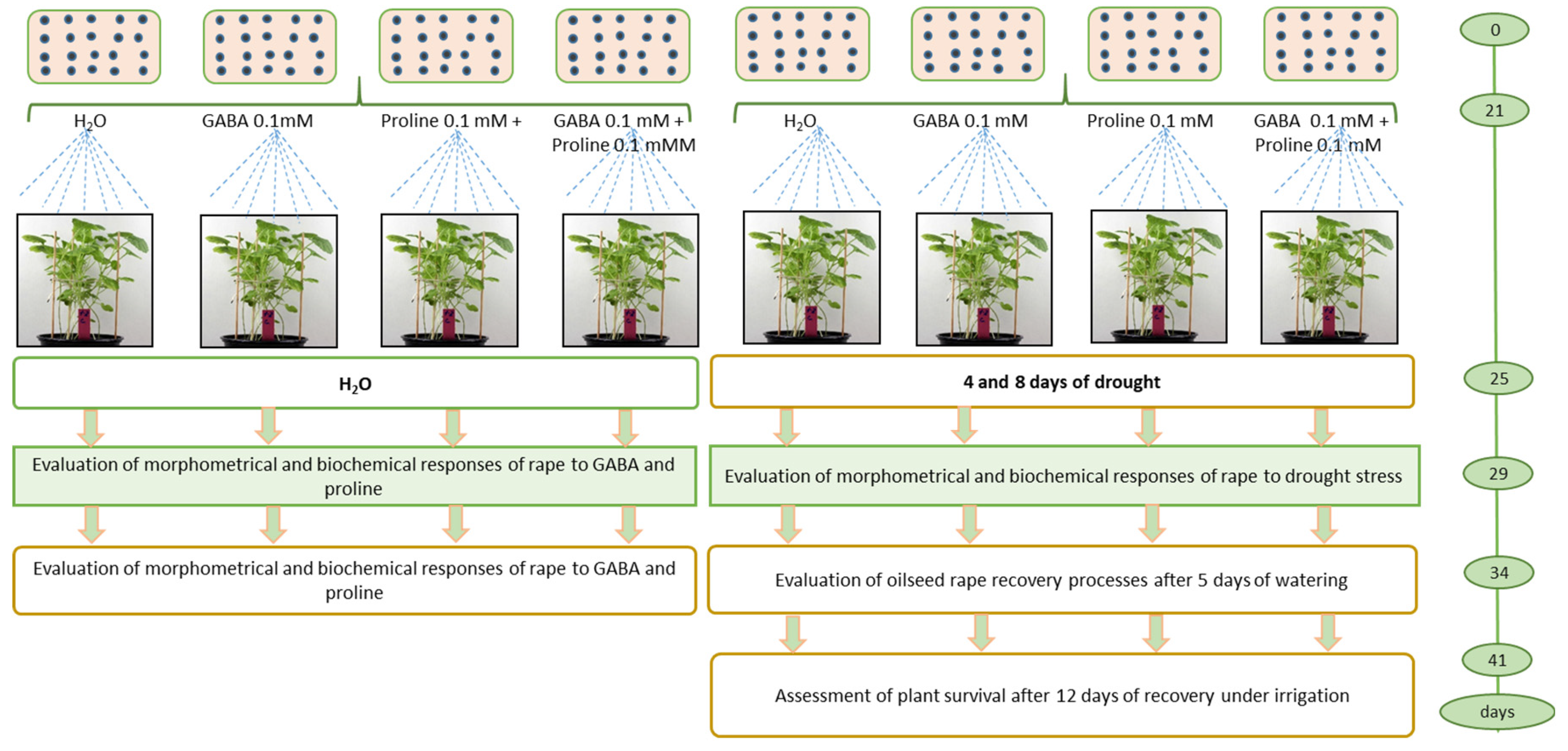

Table 1.
Effect of GABA Treatment on RWC of Leaves with a Continuous Increase in Water Deficit (Simulating Drought) and on the fresh weight of oilseed rape recovering from 8 days prolonged drought after 4 days of irrigation.
Table 1.
Effect of GABA Treatment on RWC of Leaves with a Continuous Increase in Water Deficit (Simulating Drought) and on the fresh weight of oilseed rape recovering from 8 days prolonged drought after 4 days of irrigation.
| Treatment (3 ml) | RWC, % | Average Fresh Weight per Plant | ||||
|---|---|---|---|---|---|---|
| 4 days | 8 days | g | % | |||
| Control, H2O | 76.8 ± 0.1 | 79.1 ± 5.3 | 0.97 ± 0.31 | 100 | ||
| GABA 0.1 mM | 78.1 ± 2.6 | 77.6 ± 2.1 | 0.97 ± 0.30 | 100 | ||
| GABA 1 mM | 76.9 ± 5.0 | 80.7 ± 2.5 | 0.83 ± 0.38 | 86 | ||
| GABA 10 mM | 74.4 ± 2.8 | 73.0 ± 0.8 | 0.95 ± 0.50 | 98 | ||
| Drought | 62.9 ± 2.4 | 42.8 ± 1.0 | 0.68 ± 0.27 | 100 | ||
| GABA 0.1 mM + Drought | 71.1 ± 0.9 | 58.4 ± 1.5 | 0.82 ± 0.18 | 121 | ||
| GABA 1 mM + Drought | 67.2 ± 3.0 | 56.1 ± 8.2 | 0.79 ± 0.17 | 116 | ||
| GABA 10 mM + Drought | 65.9 ± 1.6 | 54.3 ± 7.0 | 0.83 ± 0.17 | 122 | ||
| Treatment (12.5 ml) | ||||||
| Control, H2O | 76.2 ± 3.2 | 77.0 ± 1.3 | 1.13 ± 0.34 | 100 | ||
| GABA 0.1 mM | 76.6 ± 4.0 | 80.2 ± 2.0 | 1.18 ± 0.28 | 104 | ||
| GABA 1 mM | 75.4 ± 4.0 | 79.8 ± 2.0 | 1.18 ± 0.32 | 104 | ||
| GABA 10 mM | 75.4 ± 1.6 | 80.1 ± 3.5 | 1.16 ± 0.44 | 103 | ||
| Drought | 60.4 ± 6.9 | 38.0 ± 6.2 | 0.78 ± 0.52 | 100 | ||
| GABA 0.1 mM + Drought | 63.5 ± 5.1 | 60.0 ± 5.1 | 0.99 ± 0.41 | 127 | ||
| GABA 1 mM + Drought | 59.3 ± 2.1 | 56.5 ± 2.1 | 0.91 ± 0.41 | 117 | ||
| GABA 10 mM + Drought | 63.1 ± 2.5 | 54.0 ± 2.5 | 0.93 ± 0.42 | 119 | ||
Mean (±SE) was calculated from three replicates for each treatment. Values in a column with different letters are significantly different at p ≤ 0.05 applying Duncan’s Multiple Range Test (DMRT).
Table 2.
Effect of GABA on Brassica napus plant survival after 8 days of prolonged drought recovered after 12 days of irrigation.
Table 2.
Effect of GABA on Brassica napus plant survival after 8 days of prolonged drought recovered after 12 days of irrigation.
| Treatment (12.5 ml) | Number of survived plants (%) | |
|---|---|---|
| Control, H2O | 100 a | |
| GABA 0.1 mM | 100 a | |
| GABA 1 mM | 100 a | |
| GABA 10 mM | 100 a | |
| Drought | 25 d | |
| GABA 0.1 mM + Drought | 83 bc | |
| GABA 1 mM + Drought | 75 c | |
| GABA 10 mM + Drought | 67 c |
Mean was calculated from three replicates for each treatment. Values in a column with different letters are significantly different at p ≤ 0.05 applying DMRT.
Table 3.
Effect of complex treatment of proline 1 mM + GABA 0.1 mM and proline 0.1 mM + GABA 0.1 mM on fresh weight of oilseed rape seedlings (per plant) under 8 days simulated drought conditions and survival after 8 days of prolonged drought recovered after 12 days of irrigation.
Table 3.
Effect of complex treatment of proline 1 mM + GABA 0.1 mM and proline 0.1 mM + GABA 0.1 mM on fresh weight of oilseed rape seedlings (per plant) under 8 days simulated drought conditions and survival after 8 days of prolonged drought recovered after 12 days of irrigation.
| Treatment (12.5 ml) | Average Fresh Weight per Plant | Number of survived plants (%) | |
|---|---|---|---|
| g | % | ||
| Control, H2O | 1.64 ± 0.57 | 100 | 100 |
| Proline 1 mM + GABA 0.1 mM | 1.68 ± 0.66 | 102 | 100 |
| Proline 0.1 mM + GABA 0.1 mM | 1.90 ± 0.69 | 116 | 100 |
| Drought | 0.69 ± 0.31 | 100 | 27 |
| Proline 1 mM + GABA 0.1 mM + Drought | 0.93 ± 0.29 | 135 | 64 |
| Proline 0.1 mM + GABA 0.1 mM +Drought | 1.43 ± 0.50 | 121 | 85 |
Mean (±SE) was calculated from three replicates for each treatment. Values in a column with different letters are significantly different at p ≤ 0.05 applying DMRT.
Table 4.
Effect of GABA and Proline Treatment on Photosynthetic Pigment Content of Oilseed Rape Seedlings under Drought Stress.
Table 4.
Effect of GABA and Proline Treatment on Photosynthetic Pigment Content of Oilseed Rape Seedlings under Drought Stress.
| Treatment | Pigment Contents (mg g−1 FW) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Chlorophyll a | Chlorophyll b | Carotenoids | |||||||
| 4 Days | 8 Days | 4 Days Recovery | 4 Days | 8 Days | 4 Days Recovery | 4 Days | 8 Days | 4 Days Recovery | |
| Control, H2O | 1.24a | 1.27a | 1.28a | 0.38a | 0.33a | 0.34a | 0.34 a | 0.34 b | 0.34 a |
| GABA | 1.27a | 1.25a | 1.23a | 0.37a | 0.34a | 0.32a | 0.36 a | 0.37 a | 0.35 a |
| Proline | 1.28a | 1.26a | 1.22a | 0.37a | 0.32a | 0.35a | 0.35 a | 0.37 a | 0.34 a |
| GABA+Proline | 1.28a | 1.24a | 1.29a | 0.34a | 0.32a | 0.32a | 0.36 a | 0.36 a | 0.35 a |
| Drought | 0.92c | 0.54c | 0.75d | 0.20c | 0.11c | 0.16c | 0.29 b | 0.23 c | 0.25 b |
| GABA+Drought | 1.16b | 0.72b | 1.09b | 0.27b | 0.17b | 0.27b | 0.35 a | 0.31 b | 0.34 a |
| Proline+Drought | 1.18b | 0.70b | 1.04b | 0.29b | 0.16b | 0.26b | 0.38 a | 0.32 b | 0.35 a |
| GABA+Proline+Drought | 1.14b | 0.75b | 0.99bc | 0.27b | 0.17b | 0.24b | 0.32 a | 0.33 b | 0.34 a |
Mean was calculated from three replicates for each treatment. Values in a column with different letters are significantly different at p ≤ 0.05 applying DMRT.
Table 5.
The sequences of primers used in the work.
| Gene locus in Brassica napus | Gene name | Forward primer sequence (5′ → 3′) | Reverse primer sequence (5′ → 3′) |
|---|---|---|---|
| BnaA03g27910D | KIN 2-like | GCCAGACTAAGGAGAAGACAAGT | TGTTCTTGTTCATGCCGGTTT |
| BnaA05g10200D | caleosin/peroxygenase 3 | TGCCATCACCATTACTGCCT | CGGTTCCCCTCGGTTAAGTT |
| BnaC04g09800D | EDL 3 (EID 1 like F-box protein 3), | GTGGGTGGTGGCGGAGAA | CCGCCTGTCTCCTCACGA |
| BnaA05g08020D | ABI 5 | TAAGCAGCCGAGTCTTCCAC | ACCACCGCCGTTATTAGCAT |
| BnaA02g00190D | ACT7 | CCTCTCAACCCGAAAGCCAA | CATCACCAGAGTCGAGCACA |
| BnaA06g27860D | UBC21 | TATCCTCTGCAGCCTCCTCA | CTGTCTGCCTCAGGATGAGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



