Submitted:
31 January 2023
Posted:
31 January 2023
You are already at the latest version
Abstract
Melatonin (MT) controls various physiological functions and enhances plant drought tolerance in response to environmental stressors, including water deficit. This study aimed to evaluate the effect of exogenous MT on the morphophysiological attributes of Ranunculus asiaticus under normal and drought conditions. R. asiaticus seedlings were divided into drought-stress and control groups and subjected to foliar application of MT at various concentrations (0, 50, 100, and 200 μM) four times during the study. The drought-stress group exhibited considerably decreased shoot length, leaf number, leaf area, fresh and dry vegetative weights, total chlorophyll and carotenoid contents, and relative water content; delayed emergence of flower stalks; and increased relative electrolyte leakage compared with well-watered plants. Conversely, foliar application of MT notably increased growth parameters compared with their no-treatment counterparts. Foliar treatment with 200 µM MT resulted in the most significant growth response in R. asiaticus under normal or drought-stress conditions. Moreover, compared with no treatment, exogenously applied MT induced the appearance of flower buds and increased relative water and proline contents as well as peroxidase activity while reducing electrolyte leakage. Regarding tolerance index percentages, higher peroxidase and proline contents indicated their suitability for use as markers for drought tolerance, supporting the effective role of exogenous MT in enhancing the adaptability of Ranunculus to drought stress.
Keywords:
Drought
; Melatonin
; Peroxidase
; Photosynthesis
; Stress
1. Introduction
Ranunculus asiaticus L., commonly known as “butter cup,” is a perennial geophyte with tuberous roots. This species belongs to the Ranunculaceae family that is endemic to the Mediterranean basin and Asia Minor. Its flowers are dark red in color and terminal, and this annual crop is cultivated for use in the floral industry [1]. The seeds and tuberous roots of the R. asiaticus plant can be used to propagate new plants [2]. In its native Mediterranean habitat, the tuberous rhizomes of R. asiaticus grow as a rosette of leaves with long petioles, after the first autumn rain rehydrates its dried tissue [3]. After six to eight leaves are formed, floral induction occurs, with flowering occurring from February to May. During summer, these plants undergo dormancy, i.e., their roots dry up and their leaves wilt and die [4]. R. asiaticus requires moderately low temperatures to grow (night/day regime, 5°C–10°C/12°C–25°C; optimum day temperature, 16°C) and moderately high light intensity. R. asiaticus is regarded as a long-day quantitative plant [3]. This plant has recently become more widely cultivated because of the emergence of numerous hybrids [5]. However, owing to the scarcity of high-quality water, particularly in arid and semiarid areas, the floral industry faces stiff competition from other water-intensive industries, such as agriculture, urban management, and human consumption. Accordingly, water allotted to the floral industry is highly valuable and should be utilized as efficiently as possible [6].
As one of the worst environmental pressures, drought is often believed to be the main cause of declining flower production in water-stressed regions [7,8,9]. The most effective water-management strategies include the study of drought tolerance in plants to select more-suitable plants and determine their water requirements. Water stress arises when physical and environmental factors lead to water scarcity for plant growth, which subsequently inhibits plant development and flower output by affecting the chlorophyll and water content [10]. Research on plant responses to drought-induced stress reveals that they are dependent on morphophysiological parameters, such as photosynthesis, water status, and biochemical changes [11,12]. The mechanisms underlying the drought-stress responses of plants are extraordinarily complex and vary considerably with the plant species, development stage, and duration of water shortage [13]. Severe drought-induced stress significantly impacts the physiological and biochemical functions of plants. The most deleterious effects of drought stress include reduction in relative water content and water potential of leaves, loss of turgor, cell size reduction, decrease in photosynthetic pigments, and disturbance of different metabolic processes [14]. Moreover, responses can include stunted growth and partial or total plant death [15]. A severe lack of water can lead to an imbalance in the redox constituents of cells, i.e., the accumulation of reactive oxygen species (ROS) overwhelms the antioxidant defenses. This in turn leads to a cascade of oxidative damage that eventually impairs plant growth and development [16] and reduces plants’ fresh and dry weight [17]. ROS production is considered harmful to cells as it can result in electron leakage, lipid peroxidation, membrane deterioration, and nucleic acid and protein damage [18]. Furthermore, it is suggested that active oxygen species, antioxidative enzymatic reactions, and accumulation of osmolytes such as proline are appropriate markers of plants to drought stress [12,19,20]. Commonly, plant hormones regulate plant growth and enhance resistance [7]; hence, identifying the possible growth promoters and their processes is critical for strengthening agricultural drought tolerance.
Melatonin (MT; N-acetyl-5-methoxytryptamine), a low molecular–weight indole-based molecule, is present in all organisms and is thought to serve as a hormone in certain plant species. MT plays a variety of physiological roles in plants, including in growth, photosynthesis, biological cycles, germination of seeds and roots, and osmoregulation [21]. MT also contributes to the maintenance of ion homeostasis [22]. Exogenous MT has been demonstrated to be absorbed by plants and to exert positive effects under several biotic and abiotic stress [23,24]. MT has been extensively studied for its possible role as a plant growth promoter; its mode of action involves cooperation with other chorismate-derived phytohormones, such as indole-3-acetic acid (IAA) [25] and salicylic acid [26]. Because of its role in scavenging ROS and as a modulator of the production and activity of antioxidants (both enzymatically and nonenzymatically or by altering the expression of genes involved in many physiological mechanisms), MT may mitigate the detrimental consequences of oxidative damage to proteins, lipids, and nucleic acids [21,27]. Under drought conditions, MT improved the germination percentage of rapeseed seedlings and increased the leaf area as well as fresh and dry weights of roots and shoots [24]. In Moringa oleifera L. MT improved photosynthetic pigments (IAA), phenolic and element contents, and antioxidant enzyme systems and decreased malondialdehyde (MDA) [14]. In maize plants, MT increased the relative water content (RWC), leaf water content, and relative saturation deficit and enhanced antioxidant activities, such as peroxidase (POD) and proline accumulation [28]. In tomato plants, a stronger root system with decrease the vulnerability to water deficit stress was observed after MT application, which resulted in reduced membrane injury, presumably due to the activation of antioxidant enzymes and their related genes which initiated ROS scavenging [29]. The drought resistance of tea seedlings significantly improved with MT application, as evidenced by reduced membrane damage; increased proline, total protein, and sugar content; and increased CAT and POD activities [30].
Nonetheless, to the best of our knowledge, no study has evaluated the relative impact of MT in improving R. asiaticus tolerance against drought stress. The present study examined the possible implications of foliar MT treatment at various concentrations in promoting drought tolerance in R. asiaticus under drought conditions based on alterations in morphological, physiological, and biochemical attributes. The main objectives under the two levels of irrigation were to (1) investigate the morphological response of R. asiaticus (plant height, leaf number, and fresh and dry weights) and emergence of flower buds after MT foliar application; (2) evaluate the influence of separate spraying applications on pigments contents (chlorophyll and carotenoids), RWC, proline accumulation, electrolyte leakage (EL), and POD activity; and (3) examine the drought tolerance index (DTI%) for all the studied parameters to determine which parameter(s) could be utilized as an indicator of drought tolerance.
2. Materials and Methods
2.1. Material and Conditions for Planting
Pot experiments were conducted under uniform environmental conditions at a research greenhouse at the Floriculture and Dendrology Department of The Hungarian University of Agriculture and Life Science (Budapest, Hungary). The average day and night temperature of the greenhouse was maintained at 20°C–15°C with 60% relative humidity. Healthy seedlings of R. asiaticus with four to five true leaves (5–6 cm in size) were transplanted 30 days after sowing into plastic pots measuring 9 × 9 × 10 cm. One plant was placed in each pot, and 160 plants per hypothesis were divided into two groups: well-watered group and drought group, with each group further subdivided into four subgroups that were treated with different concentrations of MT (20 plants/subgroup). The growing medium in each pot comprised a uniform mixture of Klassmann TS3 Baltic peat (chemical components presented in Table 1), supplemented with 3 kg/m3 Osmocote Exact Potassium Dominant (Scotts, NSW, Australia) and 1 kg/m3 (soluble carbonate). The planting process was initiated on October 1, 2021, and harvesting was performed 150 days later.
2.2. Experimental Design and Irrigation Treatments
After transplantation, plant irrigation was conducted according to the standard irrigation techniques until the plants reached the eighth true-leaf stage using 100% of field potential. Thereafter, 30 days after the transplants were established, the irrigation treatments were initiated. Using a completely randomized design (RCD), irrigation and MT foliar application were considered as two independent variables. The first factor, i.e., irrigation treatment, included two irrigation levels—well-irrigated, wherein the moisture content of the wet soil was maintained between 50 and 120 millibars, and drought-stressed, wherein the moisture content was maintained between 180 and 200 millibars. This was combined with foliar MT application at four different concentrations, 0, 50, 100, and 200 μM, at 45, 60, 75, and 90 days after R. asiaticus planting. Tensiometer readings were obtained daily using a Blumat Digital PRO Plus instrument (Blumat GmbH & Co. KG, Telfs, Austria) to assess the changes in soil moisture and soil water capacity (bar) for each plant; the tensiometers were placed at a depth of 5–8 cm. The MT doses were selected as per previously published studies by Zhang et al. and Bidabadi et al. [10,31]. MT was purchased from Thermo Fisher Scientific (Geel, Belgium) and prepared according to the protocol published by Li et al. [32] by dissolving the solute in ethanol and then diluting it with Milli-Q water (ethanol/water (v/v) = 1/10000), followed by an application using a manual pump.
2.3. Morphological Characteristics
After the fourth foliar treatment within 2 weeks, 10 plant samples per treatment group were randomly selected and gathered to estimate the following vegetative growth parameters: plant height (cm) measured from the medium surface to the shoot apex using a meter rod, the number of leaves per plant counted manually, fresh and dry vegetative weights (g) (shoots and leaves), leaf area (cm2) measured using a leaf area meter (Area Meter 350, ADC Bioscientific Ltd., UK), and flower bud emergence (days from planting).
2.4. Physiological and Biochemical Characteristics
To evaluate the chlorophyll, carotenoids, and RWC, tissues from the five uppermost fully expanded leaves were collected. To determine the proline and EL content and antioxidant activity (POD), fresh leaves were collected from at least five individual plants at the end of the experiment, instantly cooled in liquid nitrogen, and kept frozen at −80°C pending inspection..
2.5. Photosynthetic Pigments Analysis
The photosynthetic pigments in the fresh leaves (0.1 g) from each treatment group were measured after crushing and extraction in 80% acetone. The homogenates were centrifuged at 14000 ×g for 5 min, and 1.5 mL of the supernatant was used to analyze the leaf pigments. A spectrophotometer (Genesys 10S UV-VIS Spectrophotometer, USA) was used to measure the amount of light absorbed at 644 nm, 663 nm for total chlorophyll, and 480 nm for carotenoids [33].
The total chlorophyll and carotenoid contents were determined using the following equations:
Total chlorophyll (mg g−o) F.W = [20.2 (A644) + 8.02 (A663)] × V/W
Carotenoid (mg g−1) F.W = 5.01 × A480 × V/W
2.6. RWC in Leaves
RWC content was assessed using the method of Turk and Erdal [34]. The following formula was used to calculate the RWC with five replicates:
RWC = (FW − DW) / (TW − DW) × 100.
A portion of the leaves was placed in distilled water overnight and their turgid weight (TW) was determined. The leaf fragment was dried in an oven at 75°C to estimate its dry weight. The fresh weight (FW) of the fifth leaf was determined upon sampling.
According to the method used by Turk and Erdal [34], EL was evaluated by placing 0.5-cm diameter disks taken from of the leaves in tubes containing 40 mL of distilled deionized water at 10°C for 24 h. On the following day, the initial electrical conductivity (EC1) of the solution was determined using a solution analyzer (Cole-Parmer Instrument Co., Chicago). To extract the total electrolytes, the samples were autoclaved for 20 min at 121°C. After incubating the samples at 21°C overnight, the final conductivity of the dead tissues (EC2) was determined. The proportion of EL was calculated of the solution of the dead tissue for each treatment using a previously published formula [35]:
EL (%) = EC1 / EC2 × 100.
2.7. POD Activity Determination
The enzyme extract was obtained by homogenizing 1 gram of leaf sample with 2 mL of phosphate buffer (pH 7.0) using a pre-cooled mortar. The mixture was then centrifuged at 12,000× g for 20 minutes at 4°C. The supernatant was used to measure the activity of POD. The activity was determined by following the procedure described by He et al. [36]. The reaction mixture was made up of 2.9 mL of 50 mM phosphate buffer (pH 5.5), 1 mL of 0.6 M hydrogen peroxide, 1 mL of 50 mM guaiacol, and 0.1 mL of the enzyme extract. The reaction was incubated at 37°C for 15 minutes and then stopped by adding 2 mL of 20% (v/v) trichloroacetic acid. The change in absorbance caused by the oxidation of guaiacol was measured at 470 nm.
2.8. Proline Content
According to a previously described method [37], proline was extracted from a 100-mg leaf sample by homogenizing it with 3% sulfosalicylic acid (5 µL/mg FW). The previous mixture was centrifuged at room temperature (25OC) for 5 min at 14000 rpm. Subsequently, 100 µl of the supernatant was placed in test tubes containing 200 µl of acidic ninhydrin solution and 200 µl of glacial acetic acid. The tubes were immersed for 1 h in a 90°C water bath, and the process was stopped by placing the tubes in an ice bath. Thereafter, 5 mL of toluene was added into the reaction mixture. Then the mixture was vortexed for 15 s. After separating the toluene and aqueous phases for at least 20 min in the dark at room temperature, the toluene phase was carefully collected into test tubes, and the absorbance of the fraction was determined at 520 nm using a spectrophotometer (Genesys 10S UV-VIS Spectrophotometer, USA).
2.9. DTI
The DTI was determined as a percentage (%) for each of the analyzed traits, as indicated in [38], with a minor change in the symbols used, as follows: DTI = (T* drought/T* cont) × 100, where T drought represents the average traits value under the stress of a water deficit, and T cont represents the average traits value under well-watered conditions [11].
T*(Drought and Control): all studied parameters.
2.10. Statistical Analyses
The current study adopted a completely randomized design. Two-way MANOVA followed by UNIANOVA was used to analyze the variables with Bonferroni correction for all dependent variables, with two between-factor levels: 1) treatment (irrigation and dry), and 2) MT concentrations (0, 50, 100, and 200 µM). In turn, one-way MANOVA was used to analyze method the DTI characteristics.
Assumption treatments: The normality of the residuals for most of the dependent variables were accepted using the Kolmogorov–Smirnov test (P > 0.05), with the exception of the cases of Num. leaf, Height DTI, Num. leaf DTI, and El. DTI was accepted by testing skewness and kurtosis, wherein the absolute value of both measures was <1 [39]. Finally, the normality was violated for the Carotenoid variable, and Log (value) data transformation was used.
The homogeneity of variances was assessed using Leven’s F test and was satisfied (P > 0.05) for most of the dependent variables, whereas those of plant height, area/leaf, DW, RWC DTI, and Carotenoids DTI were violated (P < 0.005). However, the violated values of homogeneity were not serious and were accepted by the variance ratio test (F = Max variance/Min variance) whenever the max sample size/min sample size ratio was <1.5 and the max var/min var ratio was <6 [40]. Consequently, Tukey’s post hoc test was used for factor-level comparisons [41,42]. The pairwise within-subject effect was compared using the Bonferroni method. All statistical procedures were performed using the IBM SPSS27 software [43].
3. Results
One- and two-way MANOVA overall tests revealed a highly significant multivariate main effect of factors (Wilk’s lambda < 0.001). Furthermore, in the case of two-way MANOVA, the interaction effect of factor levels was significant (Wilk’s lambda < 0.001) [44]; therefore, we compared the irrigation levels separately in terms of the hormone concentration levels, then compared the hormone concentration levels separately regarding the irrigation levels. A subsequent univariate ANOVA for different variables (Bonferroni correction) indicated significant differences for all individual variables >0.05 [45]. A post-hoc test was performed for all significant variables under the MT concentration factor and irrigation level effect. Different letters indicate different groups (Tukey/Games–Howell test, P < 0.05). Lower-case letters represent comparison of MT concentrations under fixed treatments, whereas upper-case letters represent comparison of irrigation levels under fixed MT concentration levels.
3.1. Impact of MT on Morphology under Drought Conditions
The findings of this study demonstrated that the morphological parameter values decreased significantly under drought stress. However, MT treatment considerably affected plant development and growth. Under well-irrigated conditions, foliar spraying treatment with MT (0, 50, 100, and 200 µM) substantially improved the fresh and dry vegetative weights, shoot length, leaf number, and leaf area compared with no treatment (W0MT). In untreated/stressed plants (D0MT), the shoot length decreased by 26.0%, the leaf number decreased by 31.03%, leaf area decreased by 62.2%, and shoot FW and DW decreased by 34.5% and 52.9%, respectively, compared with well-watered plants (W0MT). Compared with untreated/drought-stressed plants (D0MT), plants subjected to foliar spraying of MT (50 MT treatment) showed minor improvement in all vegetative traits, whereas those subjected to 100 MT and 200 MT treatments showed favorable effects on plant development in the form of dramatically increased shoot length (20.7% and 22.3%, respectively), leaf number (24.6% and 28.9%, respectively), leaf area (58.6% and 62.9%, respectively), shoot FW (33.5% and 42.7%, respectively), and shoot DW (25.9% and 36.7%, respectively) (Table 2; Figure 1). These effects were observed in a concentration-dependent manner.
Moreover, considering our findings, the drought-stress induced flower bud emergence early. Thus, the flower bud emergence time of control plants (D0MT) occurred 29 days early, followed by 23, 19, and 21 days early for the D50 MT, D100 MT, and D200 MT treatments, respectively, compared with properly watered plants with or without MT application (Figure 2). These outcomes imply that the foliar MT treatment had more favorable effects on plant biomass and development in drought-stressed plants and that 100 and 200 µM MT afforded more appropriate drought protection than did 50 µM MT.
3.3. Changes in Photosynthetic Pigments
The results of MT foliar applications at different doses (0, 50, 100, and 200 μM) on photosynthetic pigments in R. asiaticus plants subjected to drought stress (soil moisture, 180–200 mbar) are presented in Figure 3A,B, which revealed a dramatic decrease in the photosynthetic pigment components of leaves (total chlorophyll (Chl) and carotenoids (Car)) compared with well-irrigated plants (no stress and no MT treatment). The rate of the decrease of the chlorophyll and carotenoid content of leaves in non-MT-treated plants under drought stress was estimated at 68.29% and 51.72%, respectively, compared with nonstressed plants without MT treatment. Regarding the foliar application of MT, the data showed that the spraying of MT on leaves (particularly 200 μM MT) had a substantial impact on the chlorophyll and carotenoid content of the leaves in both conditions. Under drought circumstances, the chlorophyll and carotenoid content increased by 75 and 50%, respectively, in the 200 μM MT plants, compared with the non-MT-stressed plants (D0MT).
3.3. Exogenous MT Alters the RWC and Proline Content under Drought Stress
Plants’ accumulation of osmolytes is an efficient defense mechanism against drought-stress induced osmotic stress [46]. Therefore, the plant’s water stress levels can be evaluated by measuring the specific accumulated solutes. The relative water content (RWC) of the tissues of a plant is the most reliable indicator of its water status and capacity for survival under stressful conditions. The significant reduction of the RWC observed under drought stress was mitigated by MT treatment. Untreated/stressed plants (D0MT) showed a considerable decrease (7.1%) in RWC compared with untreated plants that had received adequate watering (W0MT). In contrast, MT-treated plants exhibited a significantly improved RWC under stressful conditions. The optimum improvement in RWC was noted at 200 µM MT, with a 6.36% increase in RWC values compared with nontreated/drought-stressed plants (D0MT) (Figure 4A).
Proline is a compatible solute that acts as a protective membrane solute and can maintain water balance, enhance cytoplasmic osmotic pressure, and protect cells during dehydration. The current investigation demonstrated that the proline content in R. asiaticus plants under drought stress was higher than those of well-irrigated plants (Figure 4B). In general, proline was impacted by drought stress and MT. Under stress conditions, the proline content in MT (50 µM)-treated plants was slightly increased by 17.8% compared with stressed plants without MT treatment (D0MT). In addition, plants treated with MT (100 and 200 µM) under stressful conditions showed significant increases in proline content (by 28.6% and 32.1%, respectively) compared with untreated/stressed plants (D0MT). Furthermore, the application of MT (0, 50, 100, and 200 µM, respectively) under nonstressed conditions led to a gradual slight increase in the proline content of well-irrigated plants (W0MT), which was less compared with plants subjected to drought conditions.
These results suggest that proline in the leaves of R. asiaticus plants adopts various pathways to counteract the negative effects of drought. Hence, applying exogenous MT may rectify these alterations and provide enhanced protection.
3.4. MT Modulates the Activity of Peroxidase Enzymes and Checks EL
Drought stress increases the generation of ROS, which in excess can damage the permeability of cell membranes, and EL has been used as an indicator of this oxidative damage [47]. The relationship between EL and POD activity was observed in our experiments (Figure 5A,B) to evaluate the function of POD in protecting membrane structure. As an activator of antioxidant enzymes, MT defends plants from oxidative stress [48]. This investigation tested the effect of drought stress on the POD activity of antioxidant enzymes in R. asiaticus plants with or without MT treatment (Figure 5A). Compared with MT-treated plants, the absence of MT significantly decreased POD activity in plants under both well-irrigated and drought conditions. Moreover, the POD activity of plants under well-watered conditions was lower than that detected in drought-stressed plants in a concentration-dependent manner. The POD activity of untreated/stressed plants (D0MT) increased by 64.6% compared with untreated/well-irrigated plants (W0MT). Under nonstressed conditions, MT treatment (MT50 and MT100) led to a gradual increase in antioxidant enzyme activity (41.2% and 94.1%, respectively), followed by a slight decrease in the presence of MT200 (70.6%), which was still higher than that observed in nontreated plants (W0MT). Conversely, after exposure to drought stress, MT-treated plants (MT50, MT100, and MT200) exhibited a progressive increase in POD activity (16.6%, 45.8%, and 58.3%, respectively) compared with untreated plants (D0MT). The effects of water deficit and exogenous MT on membrane integrity and EL are illustrated in Figure 5B. The stressful conditions significantly enhanced EL. Compared with non-MT stressed plants, plants exposed to MT at concentrations of 50, 100, and 200 µM showed a substantial reduction in EL by 5.7%, 7.7%, and 14.34%, respectively. These findings indicate that foliar MT application at 200 µM was more successful at scavenging ROS buildup.
3.5. DTI
Table 3 depicts the DTI as a percentage of all examined variables between the non-MT-treated plants grown under well-irrigated and drought-stress conditions during this study. In addition, the DTI% demonstrated that POD activity was the trait most sensitive to drought conditions, with a value of 282.35% compared with the DTI% of other traits. Proline content was the second most sensitive trait to drought, with a value of 207.40%, followed by EL (120.49%). In addition, the remaining traits exhibited DTI values <100% and >50%, with the exception of carotenoids, shoot dry weight, leaf area, and total Chl, which demonstrated the lowest responsiveness to drought conditions, with values of 49.12%, 47.20%, 37.84%, and 32%, respectively.
4. Discussion
Water deficiency is a serious environmental stressor that restricts agricultural growth and productivity, especially in arid and semiarid regions. It can have various physiological, anatomical, and morphological impacts on plants [49]. Drought stress had substantial effects on the attributes of R. asiaticus in our study such that as the drought stress increased, all growth requirements decreased, and plants flowered early (Table 2, Figure 1 and Figure 2). Restricted growth under drought stress is a morphological response of plants to prevent water loss by reducing the transpiration area [50]. Previous related studies [14,51,52,53] demonstrated that drought stress reduced the desirable characteristics of mung bean, flax, sunflower, and M. oleifera plants and decreased their water-retaining ability; the authors attributed these alterations to drought-induced diseases and ROS formation [14,54]. Plant height reductions that were observed after exposure to drought conditions (Table 2) may be explained by the decrease in cell elongation, turgor, volume, and cell growth [55]. Water deficiency reduces the amount of water in the shoots, thus triggering osmotic stress and inhibiting cell development and division, resulting in stunted plant growth as a whole [56,57]. In addition, a decrease in leaf area under stress reduces water loss and carbon assimilation, degrades the pigments used in photosynthesis, and affects and reduces photosynthesis thereby negatively impacting plant growth [58]. MT is a plant hormone that has been shown to have dual functions in plants, including promoting growth [60,61,62] and protecting against abiotic stress [14,59,60,61,62]. MT regulates the biochemical and physiological functions in plants [14,24,63,64,65]. MT is an endogenous plant regulator that acts in trace amounts [17], which explains why it potentially promoted the growth of R. asiaticus after its foliar application under normal and stress conditions (Table 2). Nevertheless, there is currently insufficient information regarding the processes underlying MT-induced tolerance to drought stress. In the current study, MT treatment gradually enhanced the vegetative growth traits under nonstress conditions. Additionally, decreased the negative effects were observed under drought-stress conditions on the characteristics of R. asiaticus, such as improved growth standards (Table 1; Figure 1). Similar impacts were noted in 100 and 200 µM MT-treated plants on enhanced shoot length and number of leaves compared to untreated/drought-stressed plants. At the same time, higher fresh and dry weights were recorded in 200 µM MT than in 100 and 50 µM MT-treated/drought-stressed plants, indicating that exogenous MT application might mitigate the detrimental effects of drought stress on R. asiaticus by enhancing their morphophysiological characteristics. This is in agreement with Imran et al. [62], Sadak et al. [66], and Altaf et al. [12], who stated that exogenous MT accelerated the growth of soybean, lupin, and tomato plants, respectively, under drought conditions. This implies that high MT levels may allow plants to endure longer stress conditions, thus significantly increasing their yield [63]. MT, an indoleamine, shares IAA’s metabolic precursor, which may explain its influence on plant growth and development [67]. Moreover, Han et al. [60] demonstrated that the exogenous administration of MT to soybean plants induced a greater level of abscisic acid accumulation and increased their resistance to drought conditions, indicating that MT and other hormones may interact in this manner. Exogenous MT upregulates the expression levels of components related to defense; for instance, an Arabidopsis transcriptome analysis in response to MT demonstrated that auxin-responsive genes were up and downregulated [24,68].
Nevertheless, in our study, stressed plants exhibited early flower bud emergence compared with plants that had received adequate irrigation (Figure 3). Furthermore, flower buds started to develop early to alter the growth of the plants in response to stressful circumstances [69]. Even in harsh environments, plants may be able to perpetuate their species in this manner [70]. Previous research also showed that shoots under drought stress developed flower buds earlier and on shorter stems than plants subjected to less stress conditions [71]. In Arabidopsis, exogenous MT application results in delayed flowering [72]. The exogenous supplementation of tissues with MT led to a delay in the flower bud emergence time because of the upregulation of the flowering locus C (FLC), which in turn suppressed the transcription of the flowering locus T (FT) [73,74].
In addition to being critical for transmitting and absorbing solar energy, photosynthetic pigments and chlorophyll are involved in vital physiological processes for maintaining plant activity [75]; therefore, their decrease may result in a reduction in photosynthesis, which could be linked to pigment oxidation, thus damaging photosynthetic pigments, impairing their biosynthesis, and limiting photosynthetic carbon assimilation [76,77]. Moreover, it reduces the amount of carbohydrates that accumulate in mature leaves, which may result in a slower rate of transfer of carbohydrates to growing organs [78]. The present results revealed that drought stress significantly (p < 0.05) decreased plant pigment contents in R. asiaticus (Figure 3A,B). Meanwhile, foliar MT at different levels increased these pigment contents under normal and stress conditions. A decrease in chlorophyll and carotenoid contents in water-deprived plants was also previously observed [79] and [80]. Total chlorophyll content reduction in water-deprived plants is connected with drought stress, which dehydrates mesophyll cells and inhibits the enzymes involved in glucose metabolism and some phases of chlorophyll synthesis, disrupting the chloroplasts to a certain extent [76,77].
Furthermore, carotenoids are essential pigments that function as photoprotectants and safety valves, thus dissipating excess energy before damaging plant cells [59]. The antioxidant properties of carotenoids in plants keep chlorophyll levels stable even when plants are subjected to oxidative stress [81]. In our investigation, MT-treated plants exhibited an increase in photosynthetic efficiency during stress in R. asiaticus. Further, 200 μM MT was found to be the best concentration for inhibiting pigment breakdown (Figure 3A,B) and increasing carotenoid content, which could protect against the degradation of chlorophyll, resulting in the maintenance of higher chlorophyll content in MT-treated plants compared with untreated plants. Similar outcomes regarding the improvement of chlorophyll levels have been documented in rice (Oryza sativa), M. oleifera, and melon (Cucumis melo L.) under abiotic stress after exogenous MT treatment [14,60,82]. Exogenous MT treatment at the appropriate concentration can improve chlorophyll biosynthesis and enhance plant biomass by reducing chlorophyll breakdown and increasing chlorophyll content, thereby enhancing photosynthesis in drought-stressed plants and consequently led to increased translocation of the pigment from the source to different tissues [74]. This effect has been attributed to the antioxidant properties of MT and its impact on the genes that encode the chlorophyll-degrading enzymes chlorophyllase, pheophorbide (an oxygenase), and red chlorophyll catabolite reductase [74].
In many plant species, relative water stress is regarded as a quick and reliable signal of stress level that is closely linked to physiological changes at the leaf and entire-plant levels [20,83]. For decorative pot plants, the RWC is an appropriate index for irrigation control [84]. Furthermore, maintaining a high RWC is a well-known strategy in breeding programs for drought resistance and is essential to maintain plant metabolism [84]. Therefore, a decreased RWC indicates turgor loss and lower water availability for cell growth. The variance in RWC observed in drought-stressed plants is attributable to the varying capability of plants to absorb water or the capacity of stomata to reduce water loss [85]. The significant difference in the RWC detected between well-irrigated and drought-stressed plants in our study demonstrated that plants were subjected to adequate water stress (Figure 4A). In addition, the increased RWC after MT application is likely attributable to the enhanced water absorption due to the safeguarding of the membrane [82,86]. It has been demonstrated that MT can improve the thickness of a plant’s cuticle, which in turn limits water loss. MT treatment has been shown to improve drought tolerance in plants by keeping their turgor and water ratio stable [87].
One of the main objectives of this study was to investigate the effect of drought stress on the accumulation of certain chemicals such as proline in leaves and their impact on drought-tolerance enhancement. As an amino acid, proline is osmotically active and regulates the storage of usable nitrogen, thus substantially contributing to the stability of membranes by enhancing cell turgor, preserving the cell osmotic balance, and guarding against cell dehydration and protein impairment [88]. These findings are consistent with previous investigations and suggest the importance of proline as a stress-protective component [12,20,82]. Proline can detoxify plants, act as an antioxidant against abiotic stresses, or stabilize macromolecules under dehydration stress by scavenging free radicals and buffering the cellular redox potential [89]. As a signaling control molecule, it initiates several pathways that aid in drought adaptation [90]. Under stress conditions, the plant water status was inversely correlated with proline accumulation in non-MT-treated stressed plants , while with MT application both of proline and RWC showed a substantial increase (Figure 4A,B), this result is in agreement with [91]. A negative relationship between RWC and proline accumulation appears to be associated with a diminished effect of proline on osmotic adjustment [81]. Moreover, MT acts as an antioxidant, thus preventing proline breakdown [22]. This study the highest proline level was detected in drought-stressed plants compared with well-watered plants. Meanwhile, exogenous MT treatments yielded a considerable but gradual increase in proline levels under drought conditions with increasing concentrations of MT, in particular treatment with 200 µM MT was the most effective (Figure 4B). Similar results were reported by Li et al. [24], Kamiab [65], and Jiang et al. [94].
In adverse environmental stressors, plants produce more ROS, which causes membrane lipid oxidation, oxidative damage, cell damage, and increased EL [93]. In this context, water deficiency as one of the aforementioned environmental stresses increases the overproduction of free radical molecules as ROS [94]. To avoid irreparable damage and preserve their integrity, cells attempt to maintain ROS homeostasis through the antioxidant machinery of the cells such as antioxidant compounds and enzymes [95,96]. The equilibrium of these antioxidant enzymes is essential to suppress ROS generation [95]. The main role of POD as a ROS scavenger is the degradation of H2O2 to water molecules after the conversion of O2− to O2 and H2O2 by superoxide dismutase (SOD) [97]. The primary function of MT in plant stress tolerance is the enhancement of antioxidant defense mechanisms, which are well-known endogenous scavengers of ROS and antioxidants [48]. In the present study, we measured the activity of POD as an antioxidant enzyme and demonstrated that different POD levels were observed in plants under normal or drought conditions treated with or without MT application. MT-treated stressed plants showed a notably higher POD activity that gradually increased with MT concentration, and 200 µM MT had the most effective POD activity (Figure 5A). This outcome is in agreement with previous studies obtained for different plant species, such as Brassica napus [24], pansy [20], and tomato [12], in which MT treatment during drought stress reduced oxidative damage and restored the damaged cellular membranes. Moreover, MT, a strong long-distance signal, can be transported from treated plant tissues to distant untreated tissues via vascular bundles, resulting in abiotic stress tolerance [48]. Additionally, the administration of MT increased transcription, reduced ROS buildup by scavenging ROS, and promotes activity levels of antioxidant enzymes [98,99]
Conversely, the stability of the cell membrane or EL has long been recognized as a marker of abiotic cell damage [100]. The loss of K+ due to drought stress is irreversible. This leakage can be attributed to a loss of membrane integrity, which reduces plants’ ability to retain K+ [101]. In this research, minimizing EL in plants treated with MT afforded a protective effect toward membrane damage under drought conditions (Figure 5B). MT has both hydrophilic and lipophilic attributes, can circulate between the lipid membrane and cytoplasm, and is found on the hydrophilic side of lipid bilayers [100,102,103]. At minimal concentrations, MT molecules align themselves parallel to the lipid tail. In contrast, at high concentrations, they align themselves parallel to the bilayer, indicating that the ability of MT to organize itself in lipid membranes is affected by its concentration [103].
Studies reported by Rodriguez et al. [104] demonstrated that MT could control the antioxidant enzyme system and improve enzyme activity in plants directly or indirectly. Under conditions of water deficit, MT can preserve the cell wall of plants by scavenging ROS; the current study supported this observation, as it found an elevation in proline content and POD level and a reduction in electrolyte leakage in MT-treated plants (Figure 4B and Figure 5A,B). Similarly, soybean plants stressed by a water deficit and treated with exogenous MT exhibited alleviation of oxidative damage in leaves via an increase in POD activity and a reduction of EL levels [62], in contrast to the levels recorded under untreated drought conditions. In the present work, 200 µM exogenous MT was the best concentration to ameliorate oxidative damage in leaves, suggesting that exogenous MT significantly preserved the cell membrane against oxidative stress during drought conditions (Figure 5B). In addition, the various doses led to distinct patterns of enhanced antioxidant enzyme activity.
Depending on the DTI% values, the characters investigated in this study could be classified into three categories. The first category consisted of the variables having a DTI% > 100%, as observed for POD, proline, and El; the second category included variables that showed a DTI% >50% and <100%, such as the time of flower bud emergence, shoot FW, number of leaves and shoot length, in decreasing order. The final category included the variables with DTI% values <50%, as observed for carotenoid content, shoot dry weight, leaf area, and chlorophyll. Based on the high DTI% values observed for POD enzyme activity, leaf proline content, and El, it is conceivable to use these characteristics as obvious markers of response to drought stress in R. asiaticus plants. Several previous works have debated the use of DTI as a drought-response indicator in Malva sylvestris, Althea rosea, Callistephus chinensis, Rudbeckia hirta [11], and Helianthus annuus [105], in which the stress could be assessed based on the stress index of various traits, such as plant height, dry matter, root length, and relative root volume and density. To the best of our knowledge, this is the first study that computed DTI% for a wide variety of R. asiaticus plant features. The results of this study may prove helpful for future studies of plant physiology and breeding.
5. Conclusions
This study revealed that the MT-induced improvement in drought stress tolerance in R. asiaticus plants was associated with biochemical changes and might be an effective practice under stress conditions. Our data showed identified protective evidence supporting that exogenous MT application in Ranunculus seedlings enhances the performance of the antioxidant defense system by diminishing the generation of ROS as demonstrated by POD activation and decreased EL. Furthermore, it can improve osmotic regulation capability by increasing osmolyte accumulation and maintaining water status to reduce drought-related damage. MT treatment also enhanced the amount of carotenoid content, which acts as a nonenzymatic antioxidant, resulting in a higher level of total chlorophyll content being maintained, thereby strengthening the vegetative growth parameters under drought conditions. Considering the DTI% values, POD enzyme activity and leaf proline content could indicate the drought stress response of Ranunculus plants. Overall, when applied at the optimal dose of 200 µM in this study, MT alleviates drought stress on the morphophysiological characteristics, improving R. asiaticus’ tolerance. This can also be applied on a broad scale as a safe and economical technique to increase plant tolerance to drought stress and other stressors in future studies.
Author Contributions
Conceptualization, E.A.E.; Formal Analysis, E.A.E., P.H., A.T-M., M.A.G.; Funding acquisition, N/A; Investigation, E.A.E., P.H., A.T-M.; Project administration, E.A.E., P.H., A.T-M.; Validation, E.A.E., P.H., A.T-M., M.A.G.; Visualization, E.A.E., P.H., A.T-M.; Writing – original draft, E.A.E.; Writing – review & editing, M.A.G.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Karlsson, M. Producing Ravishing Ranunculus. Greenh. Prod. News January 2003, 44–48. [Google Scholar]
- Margherita, B.; Giampiero, C.; Pierre, D. Field Performance of Tissue-Cultured Plants of Ranunculus Asiaticus L. Sci. Hortic. (Amsterdam). 1996, 66, 229–239. [Google Scholar] [CrossRef]
- Rauter, S.; Sun, Y.; Stock, M. Visual Quality, Gas Exchange, and Yield of Anemone and Ranunculus Irrigated with Saline Water. Horttechnology 2021, 31, 763–770. [Google Scholar] [CrossRef]
- Meynet, J.A.; Le Nard, M. Ranunculus. In The physiology of flower bulbs; De Hertogh, A.A., Nard, M.L., Eds.; Elsevier, 1993; p. 795. [Google Scholar]
- Beruto, M.; Rabaglio, M.; Viglione, S.; Labeke, M.-C. Van; Dhooghe, E. Ranunculus. In Ornamental Crops; Springer, 2018; pp. 649–671. [Google Scholar]
- Hashemi, G.S.E.; Mostafa, Z.B.; Heidarzadeh, M. Estimation of Water Requirement about Some of the Most Dominant Green Areas in Isfahan, Using Lysimeter. In Proceedings of the Third National Conference on Green Spaces and Urban Landscape, Kish Island, Municipality and Degradation Organization of the Country (In Persian); Elsevier, 2007. [Google Scholar]
- Khan, F.; Upreti, P.; Singh, R.; Shukla, P.K.; Shirke, P.A. Physiological Performance of Two Contrasting Rice Varieties under Water Stress. Physiol. Mol. Biol. Plants 2017, 23, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Tombesi, S.; Frioni, T.; Poni, S.; Palliotti, A. Effect of Water Stress “Memory” on Plant Behavior during Subsequent Drought Stress. Environ. Exp. Bot. 2018, 150, 106–114. [Google Scholar] [CrossRef]
- Caser, M.; Chitarra, W.; D’Angiolillo, F.; Perrone, I.; Demasi, S.; Lovisolo, C.; Pistelli, L.; Pistelli, L.; Scariot, V. Drought Stress Adaptation Modulates Plant Secondary Metabolite Production in Salvia Dolomitica Codd. Ind. Crops Prod. 2019, 129, 85–96. [Google Scholar] [CrossRef]
- Bidabadi, S.S.; Vander, W.J.; Sabbatini, P. Exogenous Melatonin Improves Glutathione Content, Redox State and Increases Essential Oil Production in Two Salvia Species under Drought Stress. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Rafi, Z.N.; Kazemi, F.; Tehranifar, A. Morpho-Physiological and Biochemical Responses of Four Ornamental Herbaceous Species to Water Stress. Acta Physiol. Plant. 2019, 41, 1–13. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.-X.; Naz, S.; Altaf, M.M.; Khan, L.U.; Tiwari, R.K.; Lal, M.K.; Shahid, M.A.; Kumar, R.; et al. Melatonin Improves Drought Stress Tolerance of Tomato by Modulating Plant Growth, Root Architecture, Photosynthesis, and Antioxidant Defense System. Antioxidants 2022, 11, 309. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Ashraf, U.; Hussain, S.; Shahzad, B.; Khan, I.; Wang, L. Effect of Progressive Drought Stress on Growth, Leaf Gas Exchange, and Antioxidant Production in Two Maize Cultivars. Environ. Sci. Pollut. Res. 2016, 23, 17132–17141. [Google Scholar] [CrossRef]
- Sadak, M.S.; Abdalla, A.M.; Abd Elhamid, E.M.; Ezzo, M.I. Role of Melatonin in Improving Growth, Yield Quantity and Quality of Moringa Oleifera L. Plant under Drought Stress. Bull. Natl. Res. Cent. 2020, 44, 18. [Google Scholar] [CrossRef]
- Abdul Jaleel, C.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J. Somasundaram, R. Panneerselvam, R. Drought Stress in Plants: A Review on Morphological Characteristics and Pigments Composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Munné-Bosch, S.; Mueller, M.; Schwarz, K.; Alegre, L. Diterpenes and Antioxidative Protection in Drought-Stressed Salvia Officinalis Plants. J. Plant Physiol. 2001, 158, 1431–1437. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.D. Roles of Melatonin in Abiotic Stress Resistance in Plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Maksup, S.; Roytrakul, S.; Supaibulwatana, K. Physiological and Comparative Proteomic Analyses of Thai Jasmine Rice and Two Check Cultivars in Response to Drought Stress. J. plant Interact. 2014, 9, 43–55. [Google Scholar] [CrossRef]
- Jafari, S.; Garmdareh, S.E.H.; Behzad, A. Effects of Drought Stress on Morphological, Physiological, and Biochemical Characteristics of Stock Plant (Matthiola Incana L.). Sci. Hortic. (Amsterdam). 2019, 253, 128–133. [Google Scholar] [CrossRef]
- Oraee, A.; Tehranifar, A. Evaluating the Potential Drought Tolerance of Pansy through Its Physiological and Biochemical Responses to Drought and Recovery Periods. Sci. Hortic. (Amsterdam). 2020, 265, 109225. [Google Scholar] [CrossRef]
- Tan, D.-X.; Manchester, L.C.; Liu, X.; Rosales-Corral, S.A.; Acuna-Castroviejo, D.; Reiter, R.J. Mitochondria and Chloroplasts as the Original Sites of Melatonin Synthesis: A Hypothesis Related to Melatonin’s Primary Function and Evolution in Eukaryotes. J. Pineal Res. 2013, 54, 127–138. [Google Scholar] [CrossRef]
- Sarropoulou, V.N.; Therios, I.N.; Dimassi-Theriou, K.N. Melatonin Promotes Adventitious Root Regeneration in in Vitro Shoot Tip Explants of the Commercial Sweet Cherry Rootstocks CAB-6P (Prunus Cerasus L.), Gisela 6 (P. Cerasus × P. Canescens), and MxM 60 (P. Avium × P. Mahaleb). J. Pineal Res. 2012, 52, 38–46. [Google Scholar] [CrossRef]
- Shi, H.; Jiang, C.; Ye, T.; Tan, D.X.; Reiter, R.J.; Zhang, H.; Liu, R.; Chan, Z. Comparative Physiological, Metabolomic, and Transcriptomic Analyses Reveal Mechanisms of Improved Abiotic Stress Resistance in Bermudagrass [Cynodon Dactylon (L). Pers.] by Exogenous Melatonin. J. Exp. Bot. 2015, 66, 681–694. [Google Scholar] [CrossRef]
- Li, J.; Zeng, L.; Cheng, Y.; Lu, G.; Fu, G.; Ma, H.; Liu, Q.; Zhang, X.; Zou, X.; Li, C. Exogenous Melatonin Alleviates Damage from Drought Stress in Brassica Napus L.(Rapeseed) Seedlings. Acta Physiol. Plant. 2018, 40, 1–11. [Google Scholar] [CrossRef]
- Wang, F.; Zeng, B.; Sun, Z.; Zhu, C. Relationship Between Proline and Hg2+-Induced Oxidative Stress in a Tolerant Rice Mutant. Arch. Environ. Contam. Toxicol. 2009, 56, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Llorca, M.; Muñoz, P.; Müller, M.; Munné-Bosch, S. Biosynthesis, Metabolism and Function of Auxin, Salicylic Acid and Melatonin in Climacteric and Non-Climacteric Fruits. Front. Plant Sci. 2019, 10, 136. [Google Scholar] [CrossRef]
- Liang, C.; Zheng, G.; Li, W.; Wang, Y.; Hu, B.; Wang, H.; Wu, H.; Qian, Y.; Zhu, X.-G.; Tan, D.-X.; et al. Melatonin Delays Leaf Senescence and Enhances Salt Stress Tolerance in Rice. J. Pineal Res. 2015, 59, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Gul, N.; Haq, Z.U.; Ali, H.; Munsif, F.; Hassan, S.S. ul; Bungau, S. Melatonin Pretreatment Alleviated Inhibitory Effects of Drought Stress by Enhancing Anti-Oxidant Activities and Accumulation of Higher Proline and Plant Pigments and Improving Maize Productivity. Agronomy 2022, 12, 2398. [Google Scholar] [CrossRef]
- Mushtaq, N.; Iqbal, S.; Hayat, F.; Raziq, A.; Ayaz, A.; Zaman, W. Melatonin in Micro-Tom Tomato: Improved Drought Tolerance via the Regulation of the Photosynthetic Apparatus, Membrane Stability, Osmoprotectants, and Root System. Life 2022, 12, 1922. [Google Scholar] [CrossRef] [PubMed]
- Langaroudi, I.K.; Piri, S.; Chaeikar, S.S.; Salehi, B. Evaluating Drought Stress Tolerance in Different Camellia Sinensis L. Cultivars and Effect of Melatonin on Strengthening Antioxidant System. Sci. Hortic. (Amsterdam). 2023, 307, 111517. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Yang, S.J.; Chen, Y.Y. Effects of Melatonin on Photosynthetic Performance and Antioxidants in Melon during Cold and Recovery. Biol. Plant. 2017, 61, 571–578. [Google Scholar] [CrossRef]
- Li, H.; Chang, J.; Chen, H.; Wang, Z.; Gu, X.; Wei, C.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X.; et al. Exogenous Melatonin Confers Salt Stress Tolerance to Watermelon by Improving Photosynthesis and Redox Homeostasis. Front. Plant Sci. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of Total Carotenoids and Chlorophylls a and b of Leaf Extracts in Different Solvents 1983.
- Turk, H.; Erdal, S. Melatonin Alleviates Cold-Induced Oxidative Damage in Maize Seedlings by up-Regulating Mineral Elements and Enhancing Antioxidant Activity. J. Plant Nutr. Soil Sci. 2015, 178, 433–439. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-Induced Responses of Photosynthesis and Antioxidant Metabolism in Higher Plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Ren, Y.; Chen, X.; Chen, H. Protective Roles of Nitric Oxide on Seed Germination and Seedling Growth of Rice (Oryza Sativa L.) under Cadmium Stress. Ecotoxicol. Environ. Saf. 2014, 108, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Ábrahám, E.; Hourton-Cabassa, C.; Erdei, L.; Szabados, L. Methods for Determination of Proline in Plants. In; 2010; Vol. 639, pp. 317–331.
- Sbei, H.; Shehzad, T.; Harrabi, M.; Okuno, K. Salinity Tolerance Evaluation of Asian Barley Accessions (Hordeum Vulgare L.) at the Early Vegetative Stage. J. Arid L. Stud. 2014, 24, 183–186. [Google Scholar]
- West, S.G.; Finch, J.F.; Curran, P.J. Structural Equation Models with Nonnormal Variables: Problems and Remedies. In Structural equation modeling: Concepts, issues, and applications; Hoyle, R.H., Ed.; Sage Publications, Inc, 1995; pp. 56–75.
- Brown, M.B.; Forsythe, A.B. Robust Tests for the Equality of Variances. J. Am. Stat. Assoc. 1974, 69, 364–367. [Google Scholar] [CrossRef]
- Garson, G.D. Testing Statistical Assumptions. In Asheboro, NC: Statistical Associates Publishing; 2012.
- Tabachnick, B.; Fidell, L. IBM SPSS Statistics for Windows. Using Multivar. Stat. Pearson,Boston. IBM Corp. Released 2020. IBM SPSS Stat. Wind. 27.0. Armonk, NY IBM Corp. 2013.
- Armonk, N.Y. IBM Corp. Released 2020. IBM SPSS Statistics for Windows, Version 27.0. Armonk, NY: IBM Corp. Google Search 2020.
- Olson, C.L. On Choosing a Test Statistic in Multivariate Analysis of Variance. Psychol. Bull. 1976, 83, 579. [Google Scholar] [CrossRef]
- Barbara, G.T.; Linda, S.F. Using Multivariate Statistics; , 7th Edition. NY, NY: Pearson.ISBN-13: 9780135350904 ISBN-10: 0134790545., 2013.
- Sahin, U.; Ekinci, M.; Ors, S.; Turan, M.; Yildiz, S.; Yildirim, E. Effects of Individual and Combined Effects of Salinity and Drought on Physiological, Nutritional and Biochemical Properties of Cabbage (Brassica Oleracea Var. Capitata). Sci. Hortic. (Amsterdam). 2018, 240, 196–204. [Google Scholar] [CrossRef]
- Zhang, H.-J.; Zhang, N.; Yang, R.-C.; Wang, L.; Sun, Q.-Q.; Li, D.-B.; Cao, Y.-Y.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin Promotes Seed Germination under High Salinity by Regulating Antioxidant Systems, ABA and GA 4 Interaction in Cucumber ( Cucumis Sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.-D. Roles of Melatonin in Abiotic Stress Resistance in Plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, M.; Liu, K.; Sui, N. Effects of Drought Stress on Seed Germination and Seedling Growth of Different Maize Varieties. J. Agric. Sci. 2015, 7, 231. [Google Scholar] [CrossRef]
- Shi, L.; Wang, Z.; Kim, W.S. Effect of Drought Stress on Shoot Growth and Physiological Response in the Cut Rose ‘Charming Black’ at Different Developmental Stages. Hortic. Environ. Biotechnol. 2019, 60, 1–8. [Google Scholar] [CrossRef]
- Elewa, T.A.; Sadak, M.S.; Saad, A.M. Proline Treatment Improves Physiological Responses in Quinoa Plants under Drought Stress. Biosci. Res. 2017, 14, 21–33. [Google Scholar]
- Sadiq, M.; Akram, N.A.; Ashraf, M. Impact of Exogenously Applied Tocopherol on Some Key Physio-Biochemical and Yield Attributes in Mungbean [Vigna Radiata (L.) Wilczek] under Limited Irrigation Regimes. Acta Physiol. Plant. 2018, 40, 1–14. [Google Scholar] [CrossRef]
- Dawood, M.G.; El-Awadi, M.E.; Sadak, M.S.; El-Lethy, S.R. Comparison between the Physiological Role of Carrot Root Extract and Β$-Carotene in Inducing Helianthus Annuus L. Drought Tolerance. Asian J Biol Sci 2019, 12, 231–241. [Google Scholar]
- Dawood, M.G.; Sadak, M.S. Physiological Role of Glycinebetaine in Alleviating the Deleterious Effects of Drought Stress on Canola Plants (Brassica Napus L.). Middle East J. Agric. Res 2014, 3, 943–954. [Google Scholar]
- Banon, S.; Ochoa, J.; Franco, J.A.; Alarcon, J.; Sanchez-Blanco, M.; Bañon, S.; Ochoa, J.; Franco, J.A.; Alarcón, J.J.; Sánchez-Blanco, M.J. Hardening of Oleander Seedlings by Deficit Irrigation and Low Air Humidity. Env. Exp Bot. 2006, 56, 36–43. [Google Scholar] [CrossRef]
- Bakry, B.A.; El-Hariri, D.M.; Sadak, M.S.; El-Bassiouny, H.M.S. others Drought Stress Mitigation by Foliar Application of Salicylic Acid in Two Linseed Varieties Grown under Newly Reclaimed Sandy Soil. J Appl Sci Res 2012, 8, 3503–3514. [Google Scholar]
- Alam, M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Trehalose-Induced Drought Stress Tolerance: A Comparative Study among Different Brassica Species. POJ 2014, 7, 271–283. [Google Scholar]
- Naeem, M.; Naeem, M.; Ahmad, R.; Ahmad, R.; Ashraf, M.; Ihsan, M.; Nawaz, F.; Athar, H.; Ashraf, M.; Abbas, H.; et al. Improving Drought Tolerance in Maize by Foliar Application of Boron: Water Status, Antioxidative Defense and Photosynthetic Capacity. Arch Agron Soil Sci 2018, 64, 626–639. [Google Scholar] [CrossRef]
- Szafrańska, K.; Reiter, R.J.; Posmyk, M.M. Melatonin Application to Pisum Sativum L. Seeds Positively Influences the Function of the Photosynthetic Apparatus in Growing Seedlings during Paraquat-Induced Oxidative Stress. Front. Plant Sci. 2016, 7, 1663. [Google Scholar] [CrossRef]
- Han, Q.; Huang, B.; Ding, C.; Zhang, Z.; Chen, Y.; Hu, C.; Zhou, L.; Huang, Y.; Liao, J.; Yuan, S. Osystem II in Cold-Stressed Rice Seedlings. Front. Plant Sci. 2017, 8, 785. [Google Scholar] [CrossRef]
- Kamiab, F. Exogenous Melatonin Mitigates the Salinity Damages and Improves the Growth of Pistachio under Salinity Stress. J. Plant Nutr. 2020, 43, 1468–1484. [Google Scholar] [CrossRef]
- Imran, M.; Latif Khan, A.; Shahzad, R.; Aaqil Khan, M.; Bilal, S.; Khan, A.; Kang, S.; Lee, I. Exogenous Melatonin Induces Drought Stress Tolerance by Promoting Plant Growth and Antioxidant Defence System of Soybean Plants. AoB Plants 2021, 13, 1–12. [Google Scholar] [CrossRef]
- Wei, W.; Li, Q.T.; Chu, Y.N.; Reiter, R.J.; Yu, X.M.; Zhu, D.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Zhang, J.S.; et al. Melatonin Enhances Plant Growth and Abiotic Stress Tolerance in Soybean Plants. J. Exp. Bot. 2015, 66, 695–707. [Google Scholar] [CrossRef]
- El-Awadi, M.; Sadak, M.; Dawood, M.; Khater, M.; Elashtokhy, M. Amelioration the Adverse Effects of Salinity Stress by Using γ- Radiation in Faba Bean Plants. Bull NRC 2017, 41, 293–310. [Google Scholar]
- Ahmad, S.; Cui, W.; Kamran, M.; Ahmad, I.; Meng, X.; Wu, X.; Su, W.; Javed, T.; El Hamed, A.; Zhikuan, S.; et al. Exogenous Application of Melatonin Induces Tolerance to Salt Stress by Improving the Photosynthetic Efficiency and Antioxidant Defense System of Maize Seedling. J. Plant Growth Regul. 2021, 40, 1270–1283. [Google Scholar] [CrossRef]
- Sadak, M.S.; Ramadan, A.A.E.-M. Impact of Melatonin and Tryptophan on Water Stress Tolerance in White Lupine (Lupinus Termis L.). Physiol. Mol. Biol. Plants 2021, 27, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ruiz, J.; Cano, A.; Arnao, M.B. Melatonin: A Growth-Stimulating Compound Present in Lupin Tissues. Planta 2004, 220, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Weeda, S.; Zhang, N.; Zhao, X.; Ndip, G.; Guo, Y.; Buck, G.; Fu, C.; Ren, S. Arabidopsis Transcriptome Analysis Reveals Key Roles of Melatonin in Plant Defense Systems. PLoS One 2014, 9, e93462. [Google Scholar] [CrossRef]
- Sharp, R.G.; Else, M.A.; Cameron, R.W.; Davies, W.J. Water Deficits Promote Flowering in Rhododendron via Regulation of Pre and Post Initiation Development. Sci. Hortic. (Amsterdam). 2009, 120, 511–517. [Google Scholar] [CrossRef]
- Wada, K.C.; Takeno, K. Stress-Induced Flowering. Plant Signal. Behav. 2010, 5, 944–947. [Google Scholar] [CrossRef]
- Chimonidou-Pavlidou, D. Effect of Irrigation and Shading at the Stage of Flower Bud Appearance. Acta Hortic. 2001, 245–251. [Google Scholar] [CrossRef]
- Shi, H.; Wei, Y.; Wang, Q.; Reiter, R.; He, C. Melatonin Mediates the Stabilization of DELLA Proteins to Repress the Floral Transition in Arabidopsis. J Pineal Res 2016, 60, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Hu, Q.; Liu, Y.; Cheng, P.; Cheng, H.; Liu, W.; Xing, X.; Guan, Z.; Fang, W.; Chen, S.; et al. Strigolactone Represses the Synthesis of Melatonin, Thereby Inducing Floral Transition in Arabidopsis Thaliana in an FLC-Dependent Manner. J Pineal Res 2019, 67, e12582. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.B.A.J.H.; Arnao, M.B.; Hernández-Ruiz, J. Melatonin in Flowering, Fruit Set and Fruit Ripening. Plant Reprod. 2020, 33, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Protective Effect of Melatonin against Chlorophyll Degradation during the Senescence of Barley Leaves. J. Pineal Res. 2009, 46, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Din, J.; SU, K.; J, A.; AR, G. Physiological and Agronomic Response of Canola Varieties to Drought Stress. J An Plant Sci. 2011, 21, 78–82. [Google Scholar]
- Pandey, H.; Baig, M.; Bhatt, R. Effect of Moisture Stress on Chlorophyll Accumulation and Nitrate Reductase Activity at Vegetative and Flowering Stage in Avena Species. Agric Sci Res J 2012, 2, 111–118. [Google Scholar]
- Anjum, F.; Yaseen, M.; Rasul, E.; Wahid, A.; S, A. Water Stress in Barley. I. Effect on Chemical Composition and Chlorophyll Content. Pak J Agric Sci 2003, 40, 45–49. [Google Scholar]
- Liang, B.; Ma, C.; Zhang, Z.; Wei, Z.; Gao, T.; Zhao, Q.; Ma, F.; Li, C. Long-Term Exogenous Application of Melatonin Improves Nutrient Uptake Fluxes in Apple Plants under Moderate Drought Stress; Elsevier B.V., 2018; Vol. 155; ISBN 8629870826.
- Safari, M.; Mousavi-Fard, S.; Rezaei Nejad, A.; Sorkheh, K.; Sofo, A. Exogenous Salicylic Acid Positively Affects Morpho-Physiological and Molecular Responses of Impatiens Walleriana Plants Grown under Drought Stress. Int. J. Environ. Sci. Technol. 2022, 19, 969–984. [Google Scholar] [CrossRef]
- Emiliani, J.; D’Andrea, L.; Lorena Falcone Ferreyra, M.; Maulión, E.; Rodriguez, E.; Rodriguez-Concepción, M.; Casati, P. A Role for β,β-Xanthophylls in Arabidopsis UV-B Photoprotection. J. Exp. Bot. 2018, 69, 4921–4933. [Google Scholar] [CrossRef]
- Luis Castañares, J.; Alberto Bouzo, C. Effect of Exogenous Melatonin on Seed Germination and Seedling Growth in Melon (Cucumis Melo L.) Under Salt Stress. Hortic. Plant J. 2019, 5, 79–87. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Chen, A.; Wang, L.; Guo, X.; Niu, Y.; Liu, S.; Mi, G.; Gao, Q. Reducing Basal Nitrogen Rate to Improve Maize Seedling Growth, Water and Nitrogen Use Efficiencies under Drought Stress by Optimizing Root Morphology and Distribution. Agric. Water Manag. 2019, 212, 328–337. [Google Scholar] [CrossRef]
- SoltysKalina, D.; Plich, J.; Strzelczyk-Zyta, D.; Sliwka, J.; Marczewski, W. Effect of Drought Stress on the Leaf Relative Water Content and Tuber Yield of a Half-Sib Family of ‘Katahdin’-Derived Potato Cultivars. Breed. Sci. 2016, 66, 328–331. [Google Scholar] [CrossRef]
- Keyvan, S. The Effects of Drought Stress on Yield, Relative Water Content, Proline, Soluble Carbohydrates and Chlorophyll of Bread Wheat Cultivars. J Anim Plant Sci. 2010, 8, 1051–1060. [Google Scholar]
- Hu, W.; Zhang, J.; Wu, Z.; Loka, D.A.; Zhao, W.; Chen, B.; Wang, Y.; Meng, Y.; Zhou, Z.; Gao, L. Effects of Single and Combined Exogenous Application of Abscisic Acid and Melatonin on Cotton Carbohydrate Metabolism and Yield under Drought Stress. Ind. Crops Prod. 2022, 176, 114302. [Google Scholar] [CrossRef]
- Liu, J.; Wang, W.; Wang, L.; Sun, Y. Exogenous Melatonin Improves Seedling Health Index and Drought Tolerance in Tomato. Plant Growth Regul. 2015, 77, 317–326. [Google Scholar] [CrossRef]
- Chegah, S.; Chehrazi, M.; Albaji, M. Effects of Drought Stress on Growth and De- Velopment Frankinia Plant (Frankinia Leavis). Bulg. J. Agric. Sci. 2013, 19, 659–665. [Google Scholar]
- Seki, M.; Umezawa, T.; Urano, K.; Shinozaki, K. Regulatory Metabolic Networks in Drought Stress Responses. Curr. Opin. Plant Biolol. 2007, 10, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Marcińska, I.; Czyczyło-Mysza, I.; Skrzypek, E.; Filek, M.; Grzesiak, S.; Grzesiak, M.T.; Janowiak, F.; Hura, T.; Dziurka, M.; Dziurka, K.; et al. Impact of Osmotic Stress on Physiological and Biochemical Characteristics in Drought-Susceptible and Drought-Resistant Wheat Genotypes. Acta Physiol. Plant. 2013, 35, 451–461. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, L.; Wang, X.; Wang, Z.; Zhang, H.; Chen, J.; Liu, X.; Wang, Y.; Li, C. Beneficial Effects of Exogenous Melatonin on Overcoming Salt Stress in Sugar Beets (Beta Vulgaris L.). Plants 2021, 10, 886. [Google Scholar] [CrossRef]
- Jiang, D.; Lu, B.; Liu, L.; Duan, W.; Meng, Y.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Dong, H.; et al. Exogenous Melatonin Improves the Salt Tolerance of Cotton by Removing Active Oxygen and Protecting Photosynthetic Organs. BMC Plant Biol. 2021, 21, 331. [Google Scholar] [CrossRef] [PubMed]
- Kar, R. Plant Responses to Water Stress: Role of Reactive Oxygen Species. Plant Signal. Behav. 2011, 6, 1741–1745. [Google Scholar] [CrossRef] [PubMed]
- Ozgur, R.; Uzilday, B.; Sekmen, A.H.; Turkan, I. Reactive Oxygen Species Regulation and Antioxidant Defence in Halophytes. Funct. Plant Biol. 2013, 40, 832. [Google Scholar] [CrossRef] [PubMed]
- Yildiztugay, E.; Ozfidan-Konakci, C.; Kucukoduk, M.; Tekis, S.A. The Impact of Selenium Application on Enzymatic and Non-Enzymatic Antioxidant Systems in Zea Mays Roots Treated with Combined Osmotic and Heat Stress. Arch. Agron. Soil Sci. 2017, 63, 261–275. [Google Scholar] [CrossRef]
- Khan, A.; Numan, M.; Khan, A.L.; Lee, I.-J.; Imran, M.; Asaf, S.; Al-Harrasi, A. Melatonin: Awakening the Defense Mechanisms during Plant Oxidative Stress. Plants 2020, 9, 407. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Fan, J.; Xie, Y.; Amombo, E.; Liu, A.; Gitau, M.M.; Khaldun, A.B.M.; Chen, L.; Fu, J. Comparative Photosynthetic and Metabolic Analyses Reveal Mechanism of Improved Cold Stress Tolerance in Bermudagrass by Exogenous Melatonin. Plant Physiol. Biochem. 2016, 100, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Varghese, N.; Alyammahi, O.; Nasreddine, S.; Alhassani, A.; Gururani, M.A. Melatonin Positively Influences the Photosynthetic Machinery and Antioxidant System of Avena Sativa during Salinity Stress. Plants 2019, 8, 610. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chen, Y.; Tan, D.-X.; Reiter, R.J.; Chan, Z.; He, C. Melatonin Induces Nitric Oxide and the Potential Mechanisms Relate to Innate Immunity against Bacterial Pathogen Infection in Arabidopsis. J. Pineal Res. 2015, 59, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Saneoka, H.; Moghaieb, R.E.; Premachandra, G.S.; Fujita, K. Nitrogen Nutrition and Water Stress Effects on Cell Membrane Stability and Leaf Water Relations in Agrostis Palustris Huds. Environ. Exp. Bot. 2004, 52, 131–138. [Google Scholar] [CrossRef]
- Parkash, V.; Singh, S. A Review on Potential Plant-Basedwater Stress Indicators for Vegetable Crops. Sustain. 2020, 12. [Google Scholar] [CrossRef]
- Catala, A. The Ability of Melatonin to Counteract Lipid Peroxidation in Biological Membranes. Curr. Mol. Med. 2007, 7, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Chen, Y.-E.; Zhao, Y.-Q.; Ding, C.-B.; Liao, J.-Q.; Hu, C.; Zhou, L.-J.; Zhang, Z.-W.; Yuan, S.; Yuan, M. Exogenous Melatonin Alleviates Oxidative Damages and Protects Photosystem II in Maize Seedlings under Drought Stress. Front. Plant Sci. 2019, 10, 677. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolin, I.; Herrera, F.; Martin, V.; Reiter, R.J. Regulation of Antioxidant Enzymes: A Significant Role for Melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Ahmad, R.; Ashraf, M.Y.; Ashraf, M.; Waraich, E.A. Sunflower (Helianthus Annuus L.) Response to Drought Stress at Germination and Seedling Growth Stages. Pak. J. Bot 2009, 41, 647–654. [Google Scholar]
Figure 1.
Morphological changes in R. asiaticus plants under well-irrigated and drought-stress conditions after exogenous MT treatment. M (exogenous melatonin) at different concentrations (M0, M50, M100, and M200); 0, 50, 100, and 200 µM, respectively. Well-irrigated, plants under well irrigation conditions; Drought, plants under drought conditions.
Figure 1.
Morphological changes in R. asiaticus plants under well-irrigated and drought-stress conditions after exogenous MT treatment. M (exogenous melatonin) at different concentrations (M0, M50, M100, and M200); 0, 50, 100, and 200 µM, respectively. Well-irrigated, plants under well irrigation conditions; Drought, plants under drought conditions.

Figure 2.
Effect of exogenous melatonin on the emergence of flower buds in R. asiaticus plants cultivated under well-irrigated and drought-stress conditions. MT0, well-watered and drought-stressed plants without MT treatment; MT50, MT100, and MT200, treatment with MT at different concentrations (50, 100, and 200 μM, respectively) under well-irrigated and drought circumstances. Lower-case letters: comparison of MT concentrations under fixed irrigation treatments; upper-case letters: comparison of the irrigation treatment groups under fixed MT concentration levels (Tukey/Games–Howell test, P < 0.05). The values represent the mean ± standard deviation (SD) of at least 10 replicates.
Figure 2.
Effect of exogenous melatonin on the emergence of flower buds in R. asiaticus plants cultivated under well-irrigated and drought-stress conditions. MT0, well-watered and drought-stressed plants without MT treatment; MT50, MT100, and MT200, treatment with MT at different concentrations (50, 100, and 200 μM, respectively) under well-irrigated and drought circumstances. Lower-case letters: comparison of MT concentrations under fixed irrigation treatments; upper-case letters: comparison of the irrigation treatment groups under fixed MT concentration levels (Tukey/Games–Howell test, P < 0.05). The values represent the mean ± standard deviation (SD) of at least 10 replicates.
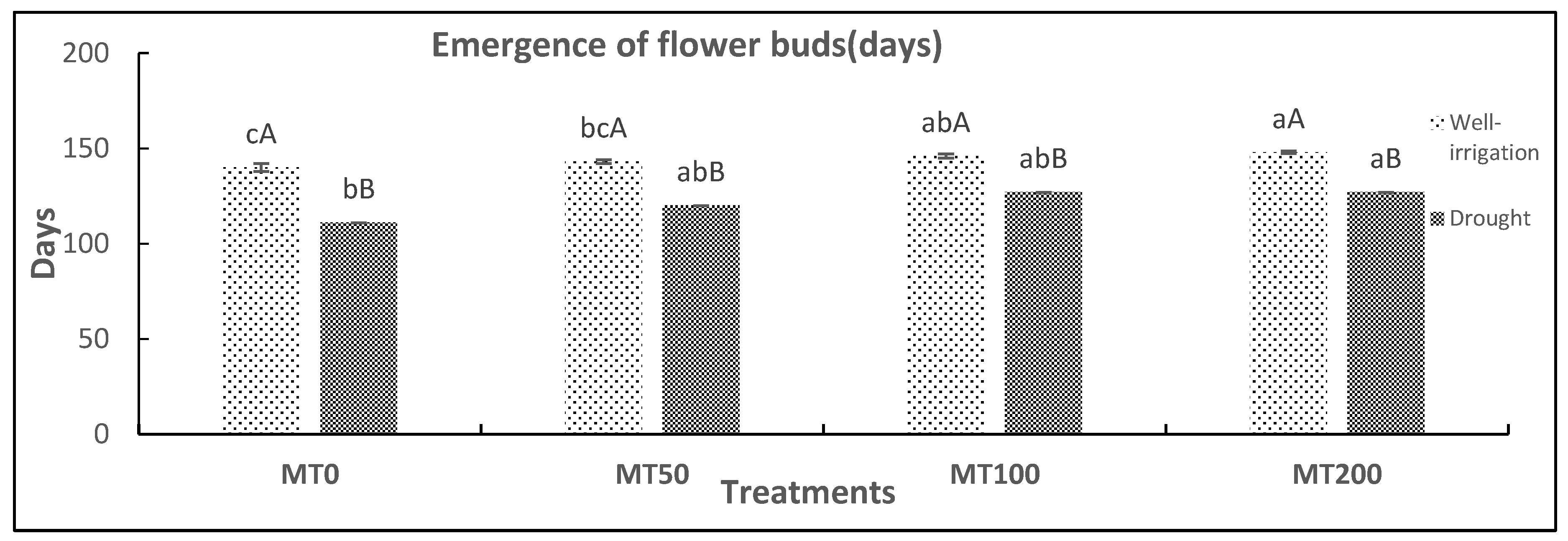
Figure 3.
Effect of exogenous melatonin on the total chlorophyll (mg g−1, FW) (A) and carotenoid (mg g−1, FW) (B) content of R. asiaticus under drought-stress conditions. A) Chlorophyll content in leaves (mg/g fresh weight). B) Carotenoid content in leaves (mg/g fresh weight). MT0, well-watered and drought-stressed plants without MT treatment; MT50, MT100, and MT200, treatment with MT at different concentrations (50, 100, and 200 μM, respectively) under well-irrigated and drought conditions. Lower-case letters: comparison of MT concentrations under fixed irrigation treatments; upper-case letters: comparison of the irrigation treatment groups under fixed MT concentration levels (Tukey/Games–Howell test, P < 0.05). The values represent the mean ± SD of at least five replicates.
Figure 3.
Effect of exogenous melatonin on the total chlorophyll (mg g−1, FW) (A) and carotenoid (mg g−1, FW) (B) content of R. asiaticus under drought-stress conditions. A) Chlorophyll content in leaves (mg/g fresh weight). B) Carotenoid content in leaves (mg/g fresh weight). MT0, well-watered and drought-stressed plants without MT treatment; MT50, MT100, and MT200, treatment with MT at different concentrations (50, 100, and 200 μM, respectively) under well-irrigated and drought conditions. Lower-case letters: comparison of MT concentrations under fixed irrigation treatments; upper-case letters: comparison of the irrigation treatment groups under fixed MT concentration levels (Tukey/Games–Howell test, P < 0.05). The values represent the mean ± SD of at least five replicates.
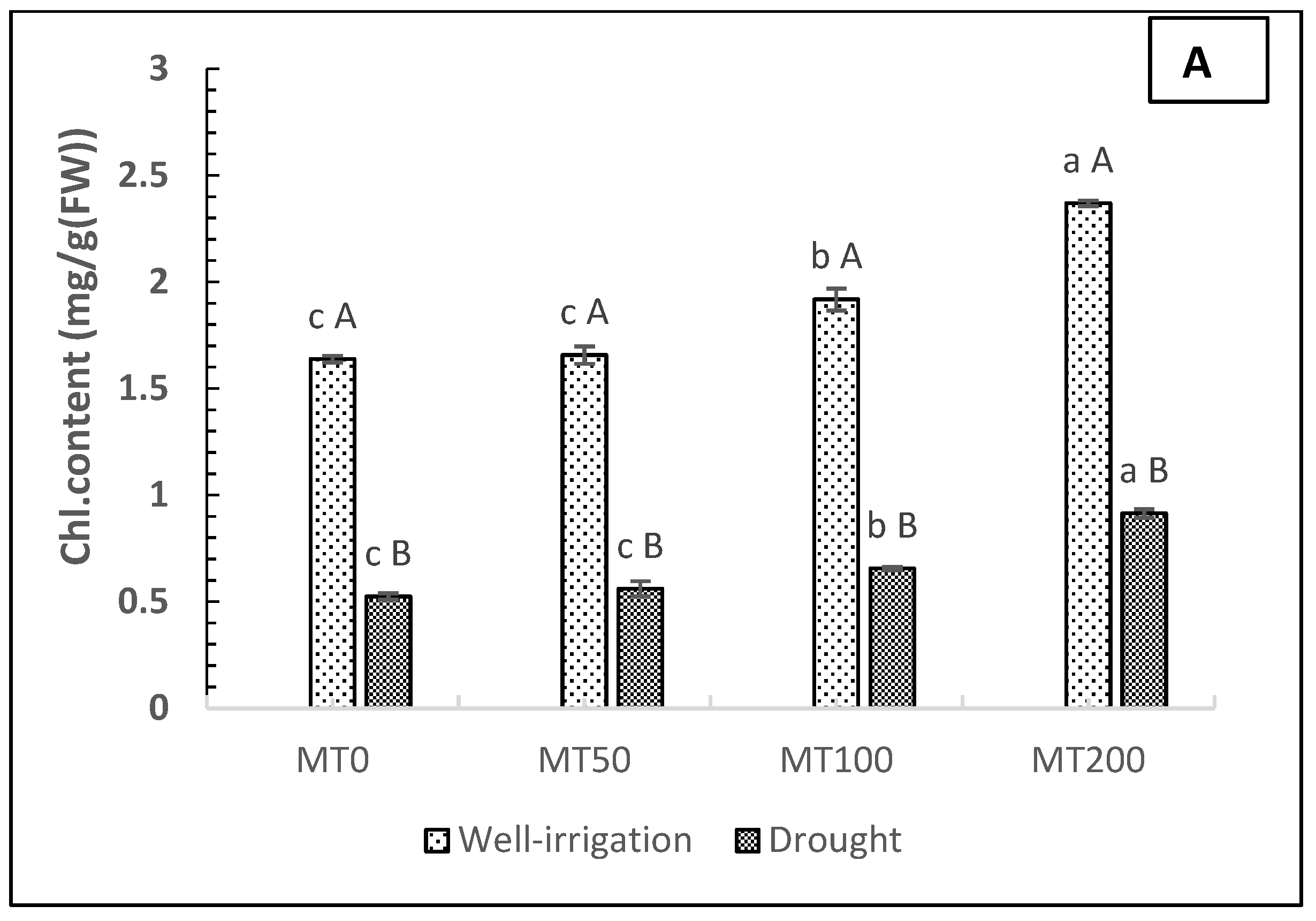
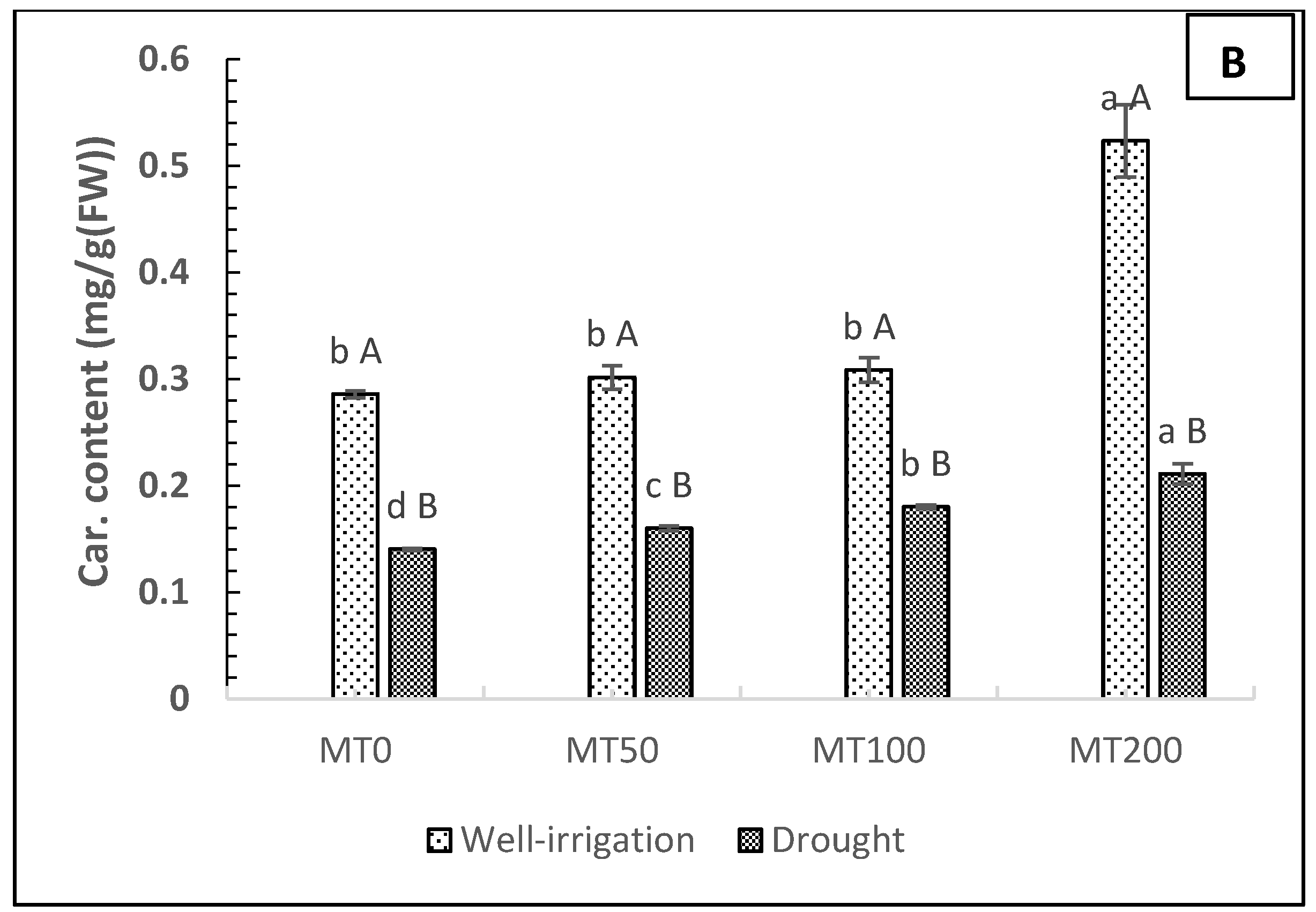
Figure 4.
Effect of exogenous melatonin on the relative water content (%) (A) and proline content (µmol g−E/FW) (B) of R. asiaticus under drought-stress conditions. MT0, well-watered and drought-stressed plants without MT treatment; MT50, MT100, and MT200, treatment with the different MT concentrations (50, 100, and 200 μM, respectively) under nonstress and stress conditions. The values represent the mean ± SD of at least five replicates. Lower-case letters: comparison of MT concentrations under fixed irrigation treatments; upper-case letters: comparison of the irrigation treatment groups under fixed MT concentration levels (Tukey/Games–Howell test, P < 0.05).
Figure 4.
Effect of exogenous melatonin on the relative water content (%) (A) and proline content (µmol g−E/FW) (B) of R. asiaticus under drought-stress conditions. MT0, well-watered and drought-stressed plants without MT treatment; MT50, MT100, and MT200, treatment with the different MT concentrations (50, 100, and 200 μM, respectively) under nonstress and stress conditions. The values represent the mean ± SD of at least five replicates. Lower-case letters: comparison of MT concentrations under fixed irrigation treatments; upper-case letters: comparison of the irrigation treatment groups under fixed MT concentration levels (Tukey/Games–Howell test, P < 0.05).
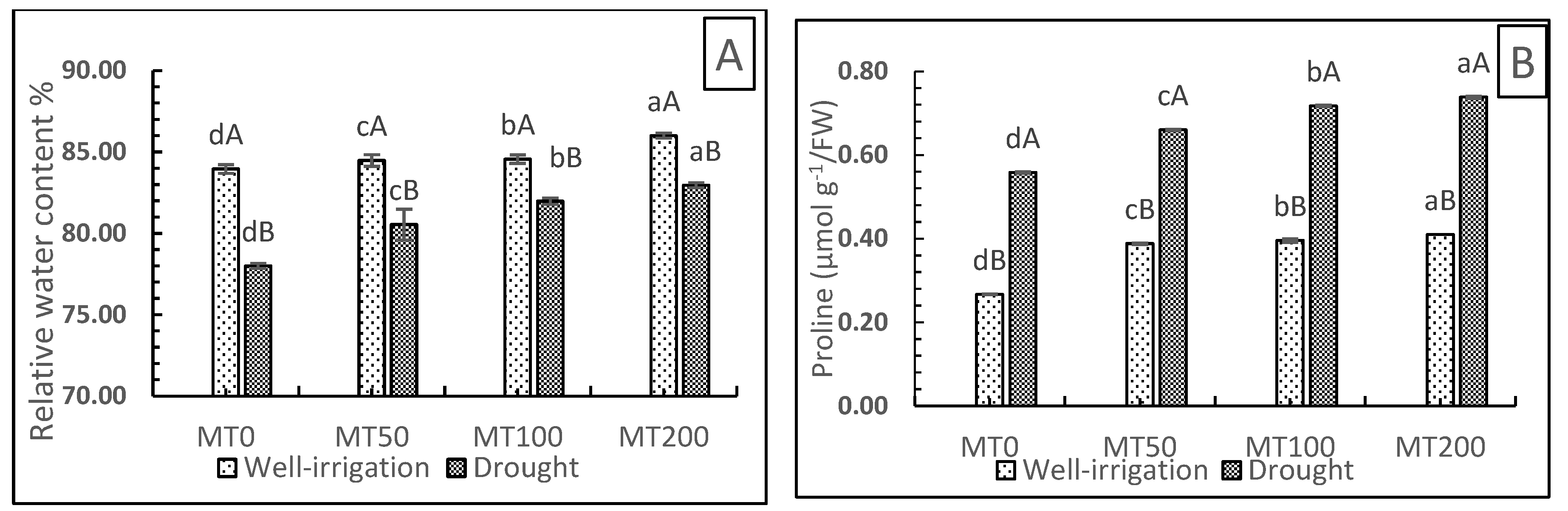
Figure 5.
Effect of exogenous melatonin on peroxidase activity [POD (U mg−O FW)] (A) and electrolyte leakage [EL (%)] (B) in R. asiaticus plants subjected to drought-stress conditions. MT0, well-watered and drought-stressed plants without MT treatment; MT50, MT100, and MT200, treatment with different MT concentrations (50, 100, and 200 μM, respectively) under nonstressed and stressful conditions. The values represent the mean± SD of at least five replicates. Lower-case letters: comparison of MT concentrations under fixed irrigation treatments; upper-case letters: comparison of the irrigation treatment groups under fixed MT concentration levels (Tukey/Games–Howell test, P < 0.05).
Figure 5.
Effect of exogenous melatonin on peroxidase activity [POD (U mg−O FW)] (A) and electrolyte leakage [EL (%)] (B) in R. asiaticus plants subjected to drought-stress conditions. MT0, well-watered and drought-stressed plants without MT treatment; MT50, MT100, and MT200, treatment with different MT concentrations (50, 100, and 200 μM, respectively) under nonstressed and stressful conditions. The values represent the mean± SD of at least five replicates. Lower-case letters: comparison of MT concentrations under fixed irrigation treatments; upper-case letters: comparison of the irrigation treatment groups under fixed MT concentration levels (Tukey/Games–Howell test, P < 0.05).
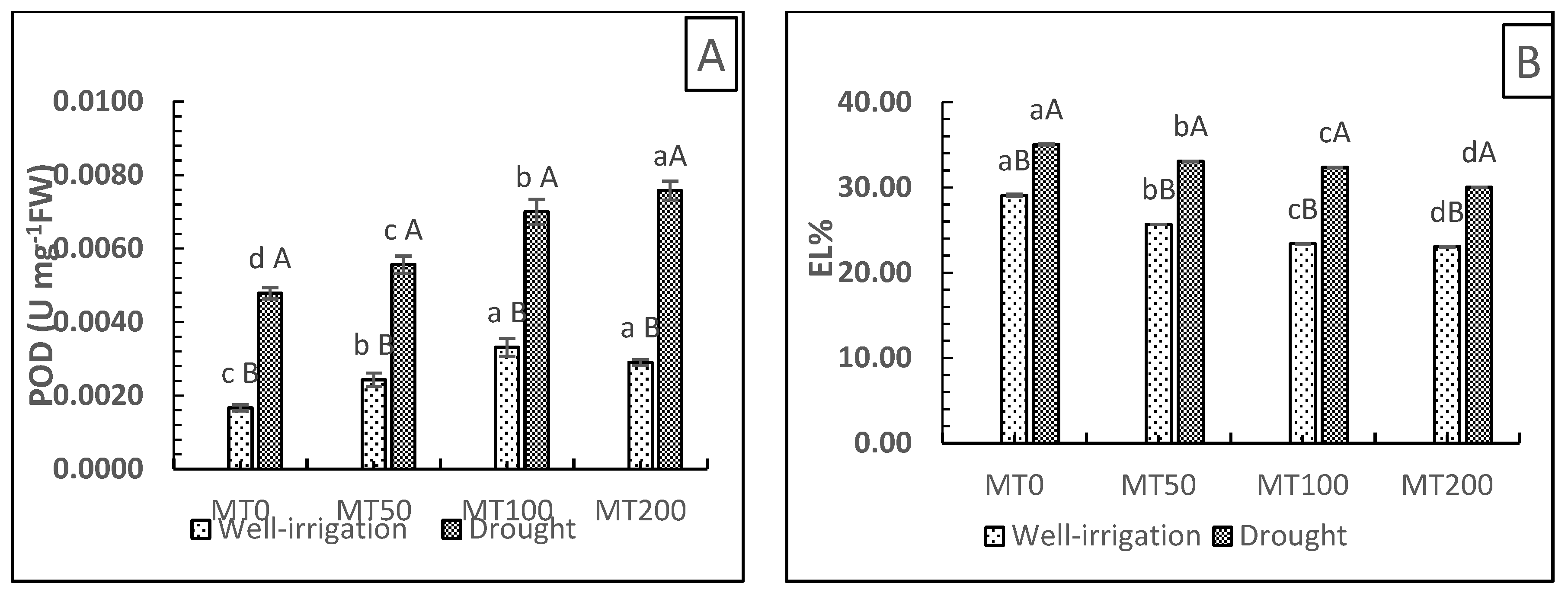

Table 1.
Klassmann TS3 Baltic peat chemical components.
| Substrate type | Peat moss |
|---|---|
| pH | 6 |
| EC (cm/mmhos) | 0.4–0.5 |
| Humidity % | 50-65 |
| Organic matter (OM)% | >90 |
| Macroelement (1 kg/m3) | |
| N:P:K | 14:10:18 |
| Microelement (mg/kg) | |
| Zn | 32.45 |
| Cu | 15.6 |
| Cd | 0.42 |
| Pb | 15.28 |
| Mo | 0.10 |
| Ni | 8.75 |
| Cr | 0.70 |
| Hg | <0.01 |
| As | 0.112 |
| Co | 1.11 |
| Cl | <0.2 |
| Na | <0.1 |
Table 2.
Effects of exogenous melatonin application on the vegetative growth traits of R. asiaticus plants with/without drought stress.
Table 2.
Effects of exogenous melatonin application on the vegetative growth traits of R. asiaticus plants with/without drought stress.
| Treatments | Shoot length (cm) | No. of leaves | Area/leaf (cm2) | Fresh weight (g) | Dry weight (g) | |
|---|---|---|---|---|---|---|
| Well-irrigated (W) | ||||||
| MT (µM) | 0 | 18.87 ± 0.18 bA | 6.67 ± 0.49 bA | 47.97 ± 0.05 dA | 18.19 ± 0.39 cA | 3.12 ± 0.19 bA |
| 50 | 19.07 ± 0.27 bA | 7.33 ± 0.49 aA | 54.49 ± 0.11 cA | 18.15 ± 0.15 cA | 3.12 ± 0.07 bA | |
| 100 | 21.49 ± 0.35 aA | 7.67 ± 0.49 aA | 55.31 ± 0.08 bA | 18.82 ± 0.26 bA | 3.17 ± 0.14 bA | |
| 200 | 21.55 ± 0.34 aA | 7.73 ± 0.46 aA | 55.60 ± 0.18 aA | 21.73 ± 0.27 aA | 3.12 ± 0.19 aA | |
| Drought (D) | ||||||
| MT(µM) | 0 | 13.97 ± 0.47 cB | 4.60 ± 0.51 cB | 18.15 ± 0.20 dB | 11.92 ± 0.17 dB | 1.47 ± 0.06 dB |
| 50 | 15.75 ± 0.37 bB | 5.20 ± 0.56 bB | 21.61 ± 0.16 cB | 13.99 ± 0.09 cB | 1.63 ± 0.09 cB | |
| 100 | 16.78 ± 0.30 aB | 5.73 ± 0.70 abB | 28.78 ± 0.22 bB | 15.92 ± 0.13 bB | 1.85 ± 0.06 bB | |
| 200 | 17.09 ± 0.39 aB | 5.93 ± 0.46 aB | 29.56 ± 0.17 aB | 17.01 ± 0.10 aB | 2.01 ± 0.12 aB | |
MT (µM) (exogenous melatonin) at different concentrations; 0, 50, 100, and 200 µM. Well-irrigated, plants under well irrigation conditions; Drought, plants under drought conditions. The values represent the mean ± standard deviation (SD) of at least 10 replicates. Lower-case letters: comparison of MT concentrations under fixed irrigation treatments; upper-case letters: comparison of the irrigation treatment groups under fixed MT concentration levels (Tukey/Games–Howell test, P < 0.05).
Table 3.
Drought tolerance index (DTI%) of all investigated R. asiaticus characteristics.
| Traits | DTI (%) |
|---|---|
| Shoot length | 74.02 ± 2.83 ef |
| Leaf number | 69.37 ± 9.35 ef |
| Leaf area | 37.84 ± 0.39 gh |
| Shoot fresh weight | 65.53 ± 1.73 f |
| Shoot dry weight | 47.20 ± 3.48 g |
| Time of flower bud emergence | 79.31 ± 2.30 ed |
| Total chlorophyll content | 32.00 ± 1.02 h |
| Carotenoid content | 49.12 ± 0.58 g |
| Relative water content (RWC) | 92.90 ± 0.45 d |
| Proline content | 207.40 ± 0.30 b |
| Peroxidase activity (POD) | 282.35 ± 10.53 a |
| Electrolyte leakage (EL) | 120.49 ± 0.72 c |
The values represent the mean ± standard deviation (SD) of at least five replicates. Different letters indicate significantly different groups (post hoc Tukey’s test, P < 0.05). Lower-case letters: comparison of the tolerance index of all estimated traits under drought stress without MT application.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



