Submitted:
05 February 2025
Posted:
05 February 2025
Read the latest preprint version here
Abstract
We propose a standardized naming system for vertebrate visual photoreceptors (i.e., rods and cones) that reflects our current understanding of their evolutionary history. Vertebrate photoreceptors have been studied for well over a century, but a fixed nomenclature for referring to orthologous cell types across diverse species has been lacking. Instead, photoreceptors have been variably - and often confusingly - named according to morphology, presence/absence of ‘rhodopsin,’ spectral sensitivity, chromophore usage, and/or the gene family of the opsin(s) they express. Here, we propose a unified nomenclature for vertebrate rods and cones that aligns with the naming systems of other retinal cell classes and that is based on the photoreceptor’s putative ancestral derivation. This classification is informed by the functional, anatomical, developmental and molecular identities of the neuron as a whole, including the expression of deeply conserved transcription factors required for development. The proposed names will be applicable across all vertebrates and indicative of the widest-possible range of properties, including their postsynaptic wiring, and hence will allude to their common and species-specific roles in vision. Furthermore, the naming system is open-ended to accommodate the future discovery of as-yet unknown photoreceptor types.
Keywords:
Vertebrate Retina
; Photoreceptor
; Rod
; Cone
INTRODUCTION
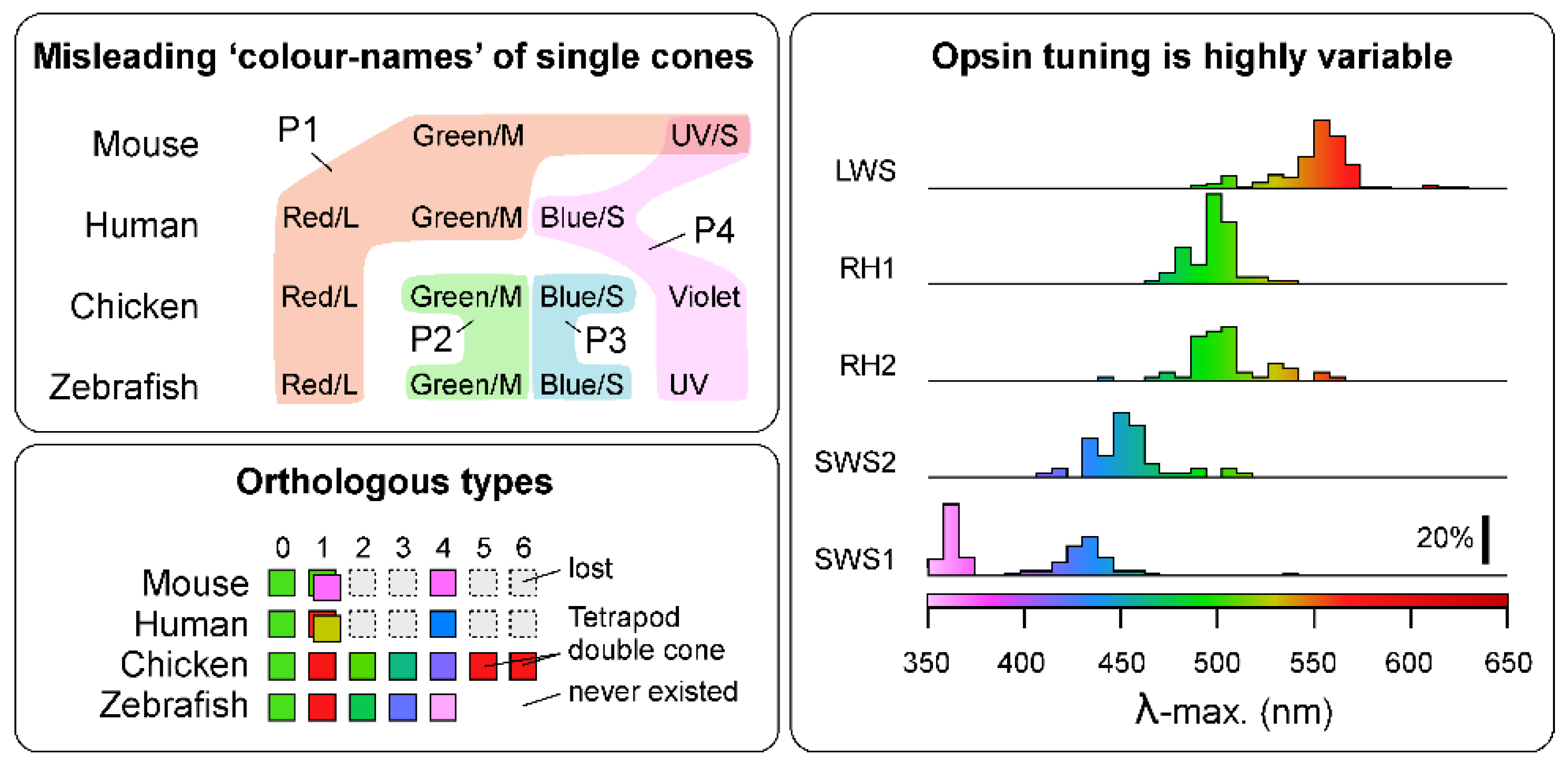
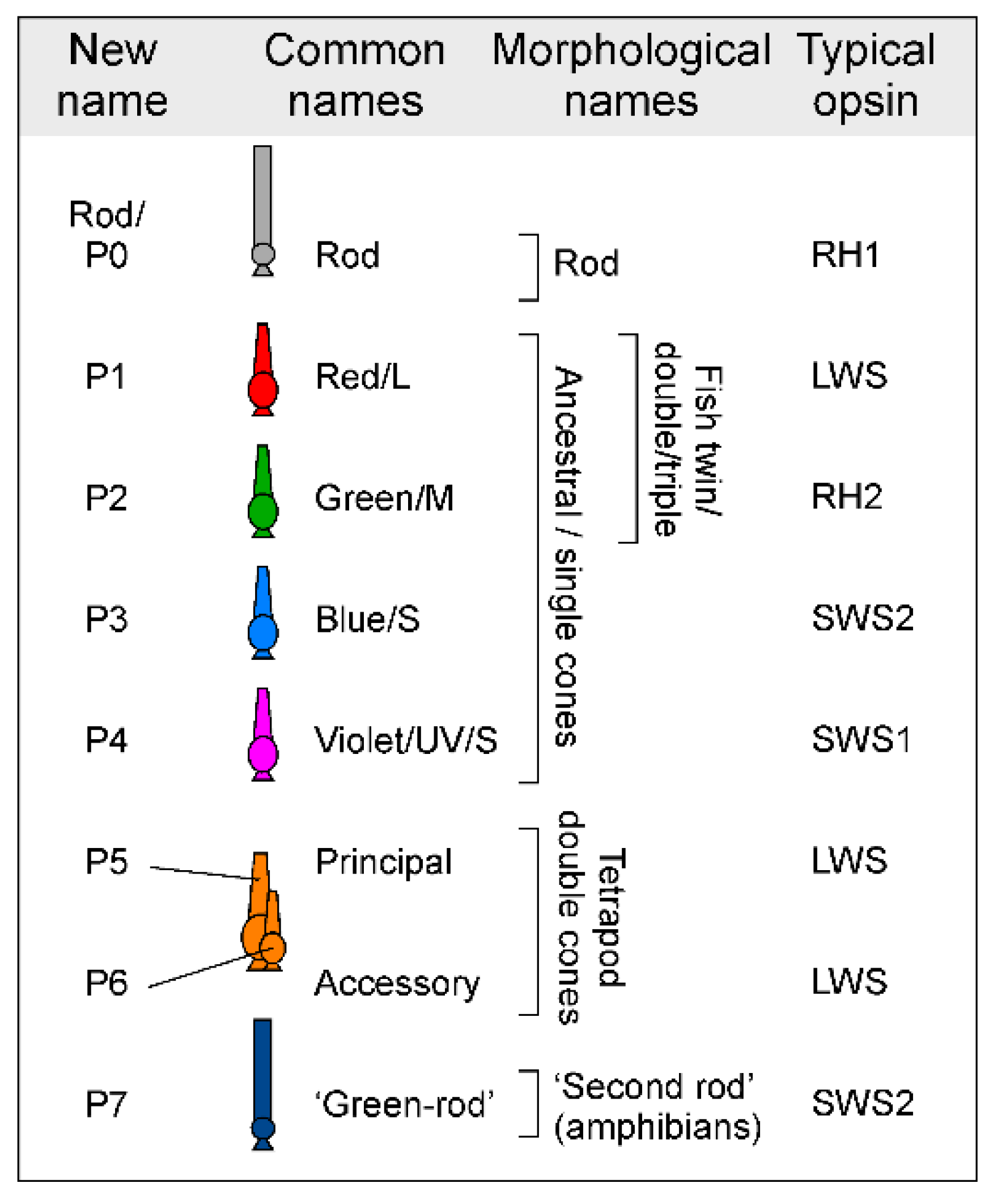
The common ancestor of extant vertebrates likely possessed a photoreceptor system consisting of one rod and four cone types, based on the shared presence of these five cell types in present-day jawed and jawless vertebrates[5,6]. Accordingly, we begin our classification with these five types (Figure 1). Following prior studies of other retinal cell classes, we propose a simple naming system in which the photoreceptor types are designated with a single letter (‘P’ for ‘photoreceptor’) followed by a number (Figure 1). Thus, cones derived from the four ancestral single cone types will be designated P1-P4 (alternative: ‘Type 1-4 photoreceptors’), and rods will be P0. Prior studies have shown that retinal cell types typically occur in regular spatial mosaics, in which cells are tiled in a non-random distribution with minimal spacing between neurons of the same type. A prior study of cone photoreceptor distributions in chicken suggested that mosaic spacing is also a feature of photoreceptor types[7]. Accordingly, we propose mosaic spacing as a working definition of photoreceptor type. Under this definition, regional variation in photoreceptor morpholology or gene expression can occur in a single photoreceptor type, without needing to designate regional variants (e.g., foveal vs. peripheral cones in human) as distinct cell types. Our classification also includes the tetrapod double cone, which consists of principal (P5) and accessory (P6) members, and the so-called ‘green rod’ of amphibians (P7). The two members of the double cone likely arose in the common ancestor of tetrapods, whereas the ‘green rod’ appears to have arisen early in the amphibian lineage[8]. According to this system, zebrafish retinas possess P0-P4[9], while chicken retinas have P0-P6[10]. Eutherian mammals, which include mice and humans, have P0, P1, and P4 types[11]. Depending on context, special notation may be required to distinguish unique subtypes of photoreceptors, such as human ‘green/M’ and ‘red/L’ cones, which differ only in the expression of paralogous LWS opsin genes. In this case, we propose the designations P1-M and P1-L, for ‘M’ and ‘L’ cones, respectively. Additional photoreceptor types can readily be added to this system when they are discovered.
THE PROBLEM
Current naming systems for vertebrate ciliary photoreceptors represent a grab bag of species-specific schemes, typically based on a single functional, morphological, or molecular feature. For example, the cone types of the human eye are often referred to as red/L, green/M, and blue/S. This naming system references the wavelength of cones’ maximal spectral sensitivity as imparted by the visual pigment [1,12] (opsin + 11-cis retinal chromophore), and oil droplet filtering [13], where present. Since the 1990’s [14,15,16] vertebrate visual photoreceptors have commonly been classified by the main opsin photopigment that they express: RH1 (rods) and LWS, RH2, SWS2, SWS1 (cones). These ancient gene sub-families were already present in the last common vertebrate ancestor, and exhibit remarkable evolutionary conservation, for example in the spectral ranges of the visual pigments, and, as we explain here, in the cell types in which they are expressed. Even so, naming cone types by their photopigment can be problematic. For example, human red and green cones differ by a single feature, namely which of two paralogues of the LWS opsin gene is expressed11. Human L and M cones derive from a single ancestral photoreceptor type, and homologs of this cell type are present in the eyes of most extant vertebrates [4]. Why, then, should this photoreceptor type not share a common designation across species?
In general, the names of photoreceptor types fall into two major groups: those that refer to spectral properties and/or the expressed type(s) of opsin (Box 1), and those that describe morphological properties, such as their size or linkage association with other photoreceptor types (Box 2). However, ‘spectral’ (‘red/L’) and opsin (‘LWS’) terms are problematic because the properties that they reference are subject to routine variation within and between species4. Likewise, morphological terms such as ‘single’, ‘double’, ‘twin’, and ‘paired’ mean different things in different animals [3]: In fishes, the latter three refer to coupled combinations of ancestral ‘red’ and ‘green’ cones [17], commonly found as part of mosaics of variable regularity [18,19]. In contrast, ‘tetrapod double cones’ [20] are molecularly [17,21], morphologically [7,22,23] and functionally [13] distinct from the four ancestral single cones24 and likely have a distinct evolutionary origin.
To overcome such naming confusions, current progress in life sciences requires cell-type definitions that encompass a wide spectrum of features [25,26,27], ideally combining morphological [28], functional [29], developmental [17,30] and molecular traits [31]. We also emphasize the importance of deeply conserved transcription factors as type-defining features in our classification.
The vertebrate retina, with its planar structure and regularly tiled architecture has long served as a central model system for cell type taxonomy. Retinal neurons form spatially repeating units [32] that anchor efforts to define types and their plausible subdivisions across animal brains. In some species such as the mouse, the catalogue of retinal neuron types is probably close to complete. This momentous achievement has been gradually unlocked by the use of transgenic tools [33], large-scale physiological recording techniques [29,34], EM-connectomics [28,35,36], and, perhaps most importantly single cell transcriptomics [26,27,31,37,38,39], which can assign a genome-wide molecular signature to anatomical and functional catalogues.
The bipolar cells of the mouse retina were amongst the first to be ‘solved’ in this way [36,37,40], and it has taken only a few more years for the tabulation of other retinal neuron types in a number of species to become increasingly complete. The mouse retina comprises ~130 neuron types: the rod, 2 cones, 1 horizontal cell, 15 bipolar cells [35,36,37,41], ~63 amacrine cells [38] and ~45 ganglion cells [26,27,29]. Of these, all except the rods and cones are primarily known by a numbered identity (e.g., H1 horizontal cell, Type 1 bipolar cell, etc.). While aspects of these catalogues still require consensus and alignment across species, in principle a numerical system gives each type of neuron a unique and unmistakable identity. For example, a horizontal cell that is orthologous to the H1 of mice [42] also exists in lampreys [43], zebrafish [39,44], chicken [10,21,45] and humans [39,46]. This understanding allows us to make powerful inferences across the vertebrate tree of life.
Recent transcriptomic studies (e.g., Refs [17,30,47,48]) investigated molecular relationships of vertebrate photoreceptors across a broad range of species. Among other findings, this work provided a molecular confirmation of what had long been suspected4: Many vertebrate photoreceptor types appear to be orthologous across multiple evolutionarily distant species. We believe that the time has come to inscribe this knowledge into the names of photoreceptor types.
OUR PROPOSAL
As a clear and systematic nomenclature for vertebrate photoreceptors, we suggest a numbering scheme that simultaneously mirrors the cell’s ancestry, their typical relative abundance in the eye, their development, and their systematically distinct postsynaptic wiring patterns (Box 3).
Ancestral red cones [4], renamed P1, are probably the least derived type of cone. In most non-avian vertebrate eyes, barring rods, P1 are the most abundant, and probably also most important type of photoreceptor [4,20,49]. P1 typically, but not always, expresses LWS-opsin, and the only sighted vertebrates thought to lack P1 are rod-only species that typically live in extreme low-light conditions. Human daylight vision is overwhelmingly driven by P1 cones [50], which include both their ‘L’ and ‘M’ variants [11]. In mouse, P1 includes both the ‘green’ sensitive dorsal and ‘UV-sensitive’ ventral LWS cones [51,52,53].
Beyond P1, the other three ‘original’ single cones [4,6] (ancestral green/RH2, blue/SWS2, UV/SWS1) become P2-4, respectively. This order mirrors their typical spectral order [2] (cf. Box 1), and their transcriptomic relatedness to P1 [17], their anatomical wiring order in the retina (‘spectral block wiring’ [1,9]), as well as the order of their typical numerical abundance and relative sizes (P1≥P2≥P3≥P4, e.g., chicken [7], adult zebrafish [54], see also Box 2). Moreover, this sequence mirrors the numerical order of cones’ postsynaptic targets, namely horizontal cells and bipolar cells. For example, the zebrafish H1 horizontal cell is the only one that connects with P1 cones, alongside P2,3 [44]. H2 then links with P2-4 cones, while H3 links to P3,4. Likewise, the Type 1 bipolar cell of mice is the only cone-bipolar cell that preferentially targets P1 cones [36]. The numerical system also elucidates evolutionary loss of cone types, e.g., for eutherian mammals who retain only P1 and P4 [4].
After these original five photoreceptor types, the next most widespread photoreceptor type in vertebrates is the tetrapod double cone [20,23,24], and we suggest that this becomes P5,6 for the principal and accessory members, respectively. This system also mirrors their likely evolutionary order of appearance [20]. We posit that the double cone should receive two numbers (5 and 6) rather than one because (i) it consists of two neurons rather than one [17,21,23], (ii) each member makes independent connections to postsynaptic targets [22], and (iii), the two members almost certainly differ in their physiological properties and roles in vision [13,24,55]; and (iv) the two cell types are morphologically [22] and molecularly [17,30] distinct.
Beyond these established photoreceptor types (see below), many amphibians have a ‘second-rod’ type known traditionally as the ‘second’ or ‘green rod’ based on its microscopic appearance (and in contrast with the canonical or ‘red rod’) [56,57,58]. Tentatively designated P7, the ancestry of this eighth type of photoreceptor remains unresolved. However, its exclusive presence in amphibians strongly suggests that unlike P5/6, P7 may have emerged after present-day amphibians diverged from all other tetrapods. Beyond P7, further hints of yet to be defined photoreceptor types include a possible ‘extra’ type in some marsupials such as dunnarts [59] and subsets of photoreceptors found in some snakes [60] and geckos [61,62,63]. We posit that these, or others, should be added onto the end of the proposed scheme if and when appropriate.
Finally, we propose that rods be designated P0. Despite rods’ substantial molecular differentiation from cones [17], both rods and cones share a common ancestry, and the presence of rods (P0) and ancestral single cones (P1-4) in cyclostomes [6,43,64] strongly suggests that both were present in stem vertebrates. While some species feature additional rod-like photoreceptors, it remains unclear which, if any, represent genuine new types (see Box 3, Definitions).
A FINAL WORD
We acknowledge that some common names for cones are deeply engrained, both in the scientific literature but also in popular culture. For example, the terms of ‘L’, ‘M’ and ‘S’ cones of the human eye, although potentially misleading (discussed in Ref [4]), are unlikely to go away anytime soon, or perhaps ever. However, as a community we can endeavour to define the terms that we use in our communications by the simple and hopefully uncontroversial numbering scheme suggested here. Perhaps, in this way, it will eventually become more widely adopted.
- Box 1. Opsins and their spectral properties are poor indicators of cone identity.
- Comparison of humans, zebrafish, and mice illustrates the central problem. Humans and zebrafish have ‘red/L’, ‘green/M’ and ‘blue/S’ cones, while zebrafish additionally have ultraviolet (UV) cones [4], but human ‘green/M’ and ‘blue/S’ cones are evolutionarily unrelated to zebrafish ‘green/M’ and ‘blue/S’ cones [17]. The opsins of human ‘green/M’ and ‘red/L’ cones are orthologous to zebrafish ‘red/L’ (all express LWS), and human ‘blue/S’ to zebrafish UV (both express SWS1). However, this match by opsins is fortuitous in the sense that both human and zebrafish cones, where present, consistently express variants of their ancestrally linked opsins: LWS, RH2, SWS2 and SWS1 for P1-4, respectively [2]. By contrast, the opsin scheme falls apart in mice because mouse P1 cones (which are often referred to as ‘green/M’ in reference to their ‘green-shifted’ LWS opsin) co-express the ancestral UV-opsin SWS1 in the ventral retina [51,52,53]. The same ancestral neuron type P1 therefore transitions from ‘functionally green’ to ‘functionally UV’ along the dorsal-ventral axis of the retina. Moreover mice retain the ancestral UV-cone P4, which like the SWS1-coexpressing P1 cones are more concentrated in the ventral retina [65]. Mice therefore have two types of UV-sensitive cones in direct proximity. Similarly, some fish species including cichlids and salmonids are known to switch opsin expression in individual cones, such that P1 cones may express LWS or RH2 opsins, and P4 cones may express SWS1 or SWS2 opsins depending on developmental stage or environmental cues [66,67,68]. While these examples illustrate the problem, they are not outliers in the vertebrate tree of life. The identity and wavelength specificity of expressed cone opsins is subject to routine variation [1,2], both across species (e.g., Ref [69]) as well as within species (including by retinal region [51,52], life stage [70], season [71,72,73], and environment [74]). It further depends on an opsin’s associated chromophore (A1 or A2) [75,76], which also varies seasonably and according to life stage. In fact, opsins and their properties are an evolutionary hotspot, varying as species enter new visual niches [2,77]. The identity or functional properties of opsins therefore do not reliably specify the identity of the neuron that expresses them.
- A second issue is that a definition by ‘colour’ implies that wavelength selectivity is the only important characteristic of a photoreceptor. This is misleading4, because beyond wavelength selectivity, cone types systematically differ in their basic cellular physiology including their spatio-temporal properties [78,79,80], as well as in their developmental postsynaptic wiring [22,36,81] – all of which directly feed into their distinct roles in vision [49].
Figure 2.
Right, Sequence changes within vertebrate opsins can give rise to diverse spectral properties (based on Ref2). Note e.g., that a λ-max of ~500 nm is readily achieved by four out of five opsin families. Colour shadings as defined on the bottom right, by most common expressed opsin variant, independent of oil droplets, where present.
Figure 2.
Right, Sequence changes within vertebrate opsins can give rise to diverse spectral properties (based on Ref2). Note e.g., that a λ-max of ~500 nm is readily achieved by four out of five opsin families. Colour shadings as defined on the bottom right, by most common expressed opsin variant, independent of oil droplets, where present.
- Box 2. Cellular morphology is an imperfect indicator of photoreceptor identity.
- Morphological definitions of photoreceptor types based on the shape of the outer segment (i.e., ‘rod’ vs. ‘cone’), association with other photoreceptors (e.g., ‘single’ vs. ‘double’ cones) or other cellular features are as problematic as opsin-based definitions. Photoreceptors can be grouped into ‘morphological types’, namely ‘single cones’ which tend to occur in isolation, ‘twin’ cones [3] which comprise pairs made up of morphologically identical partners, and ‘double/triple cones’ consisting of asymmetric groups, often with ‘principal’ and ‘accessory’ members [3,7,24,82]. Single cones are occasionally further identified by other descriptors such as ‘long’, ‘short’ [83], and miniature [84] in reference to their size and/or vertical location in the outer retina. However, there are many factors that influence the anatomical arrangement of photoreceptors, and like opsin or spectral identity, none are reliably stable across species, or within.
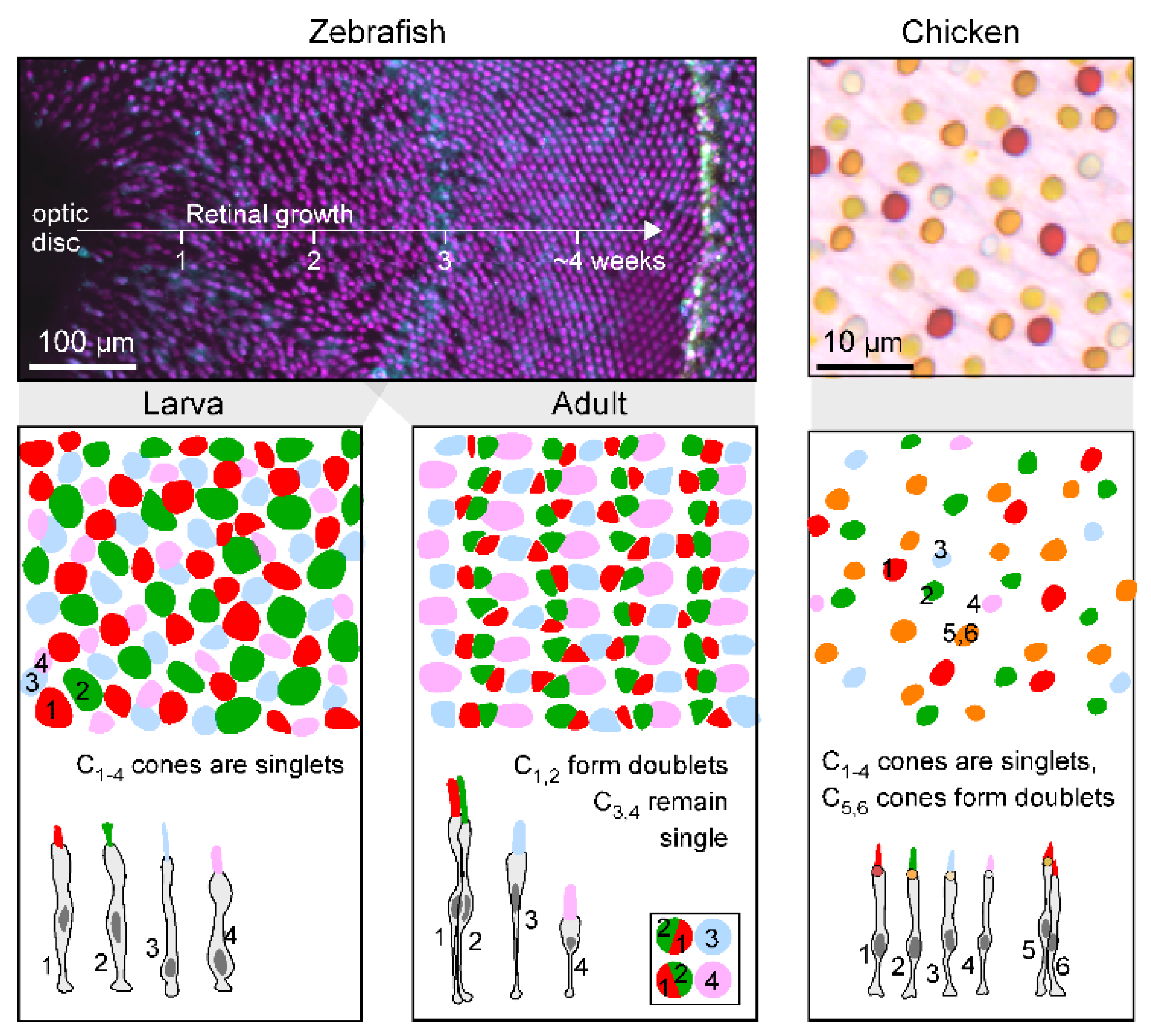
Zebrafish and chicken serve as two well studied examples. Adult zebrafish have a crystalline photoreceptor mosaic with P1-4 cones arranged at a fixed 2:2:1:1 stoichiometry in the adult [9,85]. These six neurons are arranged in a tight lattice whereby P1,2 pairs alternate rows with C3 and C4. Fishes have different patterns of C1-4 cones that vary in spectral content and regularity depending on developmental stage and retinal location. C1,2 pairs have been generally referred to as ‘paired cones’. If the two members have different morphology (usually C1>C2) or visual pigment content, they have been called ‘double cones’. If they appear morphologically equal and contain the same visual pigment, they have been termed ‘twin cones’ [86].
In non-tetrapod vertebrates, all the above terms describe subsets of the same four ancestral cone types [17], P1-4. One way to ascertain this is during development: In zebrafish, the retina does not start out in a crystalline arrangement that forces some cone types into pairs. Instead, larval cones are arranged independently [44,85,87]. This developmental history is inscribed into the adult retina as the ‘larval patch’ near the optic disc, which never rearranges [85,88]. Beyond this developmental evidence, the conclusion that all (zebra)fish cones are subsets of the same four ancestral ‘single’ cones is overwhelmingly supported by single cell transcriptomics [17,89]. By contrast, many tetrapods, with the notable exception of eutherian mammals, do have an ‘extra’ pair of cone-photoreceptors, called ‘the tetrapod double cone’20 (P5,6). Unlike fish ‘double/twin/paired’ cones, this cone pair exists in parallel to P1,2. Birds and most reptiles therefore have six ancestrally distinct types of cones. The four ‘original’ cones P1-4 plus the two members of the tetrapod double cone P5 and P6. This view is supported by extensive morphological [7,13,22,23], behavioural [24,90] and molecular evidence [17,21,91].
Figure 3.
Left, Developmental transition of zebrafish cone patterning from a mosaic with low regularity to a highly regular row lattice (top, mod from Ref [85]) and schematic summary of cone type arrangements at each stage (bottom, mod. from refs [9,44,92]). Right, oil droplets in chicken retina show five independent cone-type mosaics (top) and schematic summary of cone types (bottom, both mod. from Ref [7]).
Figure 3.
Left, Developmental transition of zebrafish cone patterning from a mosaic with low regularity to a highly regular row lattice (top, mod from Ref [85]) and schematic summary of cone type arrangements at each stage (bottom, mod. from refs [9,44,92]). Right, oil droplets in chicken retina show five independent cone-type mosaics (top) and schematic summary of cone types (bottom, both mod. from Ref [7]).
- Box 3. Naming yet-to-be-identified photoreceptor types.
We recognize that the list of photoreceptor types presented in Figure 1 is not exhaustive. For example, evidence suggests that the little skate, deep-sea fishes, marsupials, geckos, and snakes may possess unusual photoreceptor types that do not fall within any of the categories we propose. Furthermore, the photoreceptor types of only a minute fraction of the nearly 67,000 extant vertebrate species have been studied in any detail. Thus, the present classification system is intended to be open-ended, permitting the addition of more photoreceptor types as they are characterized.
The goal of our definition is to include ‘extra’ types that exist in parallel to the ancestral ones (such as tetrapod double cones P5,6 or the amphibian ‘second rod’ P7), but to exclude those that represent within-retina variation (e.g., human P1-L versus P1-M).
More generally, the proposed definition of photoreceptors into ancestral types is centrally anchored in their distinct transcriptomic signatures [17] across vertebrate ‘model’ species that are thought to be broadly representative for their clade: Humans (primates), squirrel (rodents), chicken (birds), anole lizard (non-avian reptiles), and zebrafish (teleosts). In these species, single cell transcriptomic data conforms with long standing insights into a photoreceptor’s morphological and functional properties (reviewed in Refs [4,9,10,12,20]). Together, this wealth of data renders it unlikely that their cell-type definitions will need to be revised in the light of possible future evidence. We therefore posit that these species, and others where corresponding insights exist (e.g., Ref [43]), can serve as a reference when defining cone types in other species.
Figure 4.
Non-exhaustive list of possible marker genes for rods and cones (based on Ref [17]). Note that the second rod of amphibians P7 is omitted because single-cell transcriptomic data remains outstanding.
Figure 4.
Non-exhaustive list of possible marker genes for rods and cones (based on Ref [17]). Note that the second rod of amphibians P7 is omitted because single-cell transcriptomic data remains outstanding.
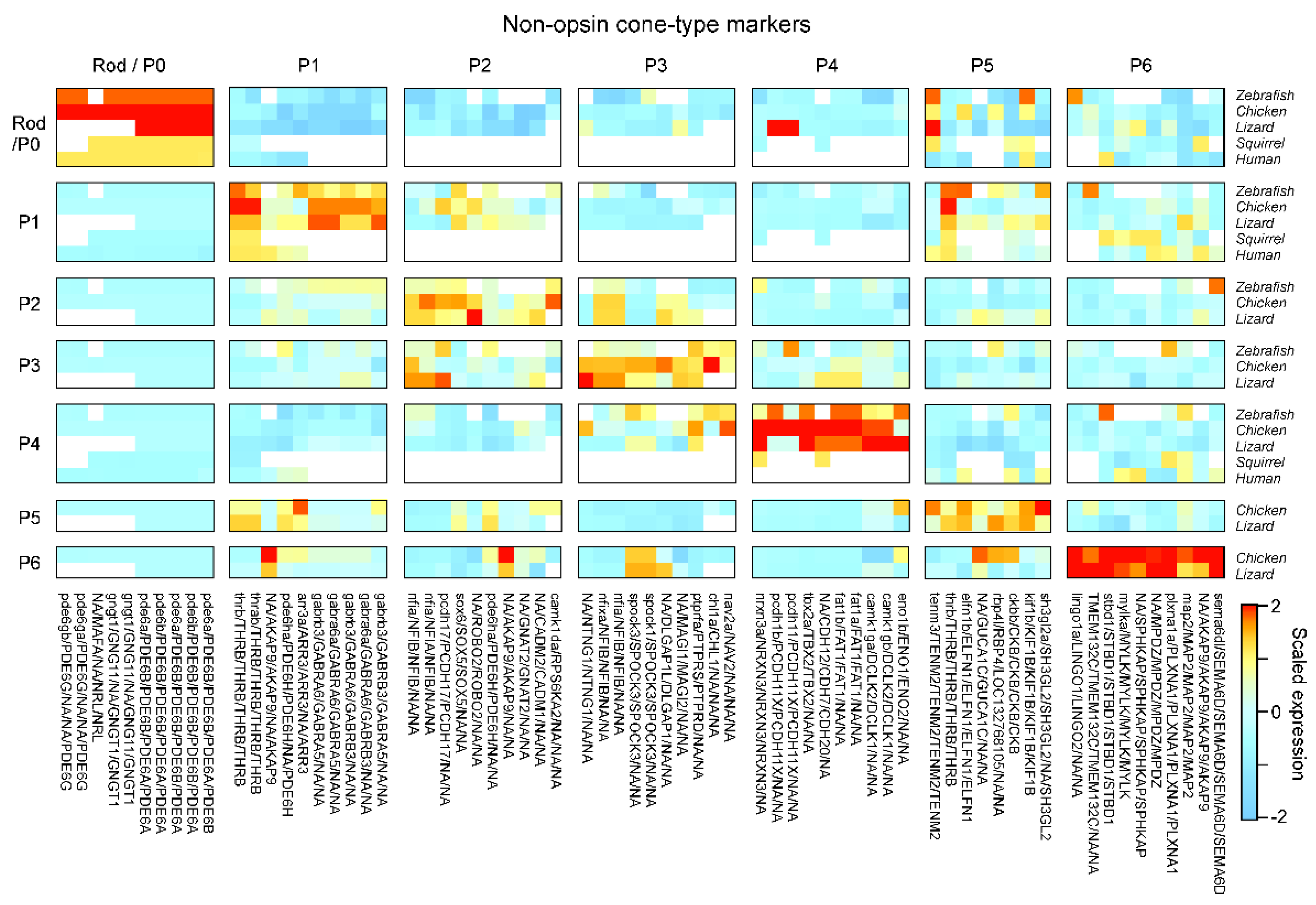
In addition, transcriptomic datasets are in good agreement with functional genetic experiments on the development of cone identity. These experiments have shown that specific transcription factors control fate decisions in photoreceptor progenitors and are thus required for the generation of each cone type or for the expression of their unique set of genes; transcriptomic datasets confirm that each cone type retains specific expression of these transcription factors throughout their lifespan, likely actively controlling cone identity. Furthermore, evidence suggests that transcription factors and their cognate binding sites throughout the genome are interlocked and therefore highly resistant to evolutionary change. Thus, transcription factors and cis-regulatory ‘grammars’ are often relatively fixed, ancestral features of a cell type which are more persistent in evolution than the modules of ‘effector genes’ which they regulate, as the latter are more directly subject to present-day adaptive pressures [93,94]. Some examples of deeply conserved photoreceptor transcription factors follow:
- Generation of P5 depends on THRB; and similar to P1, P5 also expresses SAMD7.
- P6 expresses FOXQ2 and SKOR1, similar to P3 and P3,4, respectively.
Aside from the above, inferences about photoreceptor identity can nevertheless emerge from limited data such as those used in classical definitions (Figure 1) and reference to typical patterns of photoreceptor properties across the vertebrate tree of life (reviewed e.g., in Refs [4,13,20,24]). For example:
- Numerical abundance is usually P1≥P2>P3≥P4. If present, P5,6 is usually P1>P5/6>P4, except in birds, where the more typical pattern is P5/6>P1.
- If cone types are missing, the likely order of loss is P2=P3>P4>P1. In non-eutherian tetrapods, P5,6 is usually present. P5,6 are not known to occur individually.
- Postsynaptic wiring appears to conform to ‘spectral blocks’ in the sense that ‘intermediate’ cones, if present, do not tend to be skipped. For example, a bipolar cell is unlikely to connect with P1 and P3 without also contacting P2. In this order, rods and P5,6 appear to group with P1 (i.e., P0/P5,6-P1-P2-P3-P4). In birds, P6 appears to additionally group with P3 [45].
- The spectral appearance of pigmented oil droplets, if present, generally correlates with cone-type identity, with P1-P4 exhibiting long to short-wavelength filtering properties, respectively, matching the spectral sensitivity of the corresponding opsins. P4 usually has a clear oil droplet, devoid of light-absorbing carotenoid pigments. P5 tend to have spectrally intermediate oil droplets and P6 tend to have either absent or minute droplets, while frequently retaining carotenoid pigmentation in the mitochondrial aggregates of the ellipsoid13.
Author Contributions
Original draft: TB and JCC, editing: all authors.
Conflicts of Interest
The Authors declare no conflict of interest.
References
- Baden, T.; Osorio, D. The Retinal Basis of Vertebrate Color Vision. Annu. Rev. Vis. Sci. 2019, 5, 177–200. [Google Scholar] [CrossRef]
- Hagen, J.F.; Roberts, N.S.; Johnston, R.J. The evolutionary history and spectral tuning of vertebrate visual opsins. Dev. Biol. 2022, 493, 40–66. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; et al. Retinal twin cones or retinal double cones in fish: misnomer or different morphological forms? Int. J. Neurosci. 2005, 115, 981–987. [Google Scholar] [CrossRef]
- Baden, T. Ancestral photoreceptor diversity as the basis of visual behaviour. Nat. Ecol. Evol. 2024, 8, 374–386. [Google Scholar] [CrossRef]
- Collin, S.P.; Knight, M.A.; Davies, W.L.; Potter, I.C.; Hunt, D.M.; Trezise, A.E. Ancient colour vision: multiple opsin genes in the ancestral vertebrates. Curr. Biol. 2003, 13, R864–R865. [Google Scholar] [CrossRef] [PubMed]
- Warrington, R.E.; Davies, W.I.L.; Hemmi, J.M.; Hart, N.S.; Potter, I.C.; Collin, S.P.; Hunt, D.M. Visual opsin expression and morphological characterization of retinal photoreceptors in the pouched lamprey (Geotria australis, Gray). J. Comp. Neurol. 2020, 529, 2265–2282. [Google Scholar] [CrossRef] [PubMed]
- Kram, Y.A.; Mantey, S.; Corbo, J.C. Avian Cone Photoreceptors Tile the Retina as Five Independent, Self-Organizing Mosaics. PLOS ONE 2010, 5, e8992. [Google Scholar] [CrossRef]
- Takahashi, Y.; Hisatomi, O.; Sakakibara, S.; Tokunaga, F.; Tsukahara, Y. Distribution of blue-sensitive photoreceptors in amphibian retinas. FEBS Lett. 2001, 501, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Baden, T. Circuit mechanisms for colour vision in zebrafish. 2021, 31, R807–R820. [Google Scholar] [CrossRef]
- Seifert, M.; Baden, T.; Osorio, D. The retinal basis of vision in chicken. Semin. Cell Dev. Biol. 2020, 106, 106–115. [Google Scholar] [CrossRef]
- Peng, Y.-R.; Shekhar, K.; Yan, W.; Herrmann, D.; Sappington, A.; Bryman, G.S.; van Zyl, T.; Do, M.T.H.; Regev, A.; Sanes, J.R. Molecular Classification and Comparative Taxonomics of Foveal and Peripheral Cells in Primate Retina. Cell 2019, 176, 1222–1237.e22. [Google Scholar] [CrossRef] [PubMed]
- Brainard, D.H. Color and the Cone Mosaic. Annu. Rev. Vis. Sci. 2015, 1, 519–546. [Google Scholar] [CrossRef]
- Toomey, M.B.; Corbo, J.C. Evolution, Development and Function of Vertebrate Cone Oil Droplets. Front. Neural Circuits 2017, 11, 97. [Google Scholar] [CrossRef]
- Okano, T.; Kojima, D.; Fukada, Y.; Shichida, Y.; Yoshizawa, T. Primary structures of chicken cone visual pigments: vertebrate rhodopsins have evolved out of cone visual pigments. Proc. Natl. Acad. Sci. 1992, 89, 5932–5936. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S.; Yokoyama, R. ADAPTIVE EVOLUTION OF PHOTORECEPTORS AND VISUAL PIGMENTS IN VERTEBRATES. Annu. Rev. Ecol. Syst. 1996, 27, 543–567. [Google Scholar] [CrossRef]
- Bowmaker, J.K. Evolution of vertebrate visual pigments. Vis. Res. 2008, 48, 2022–2041. [Google Scholar] [CrossRef]
- Tommasini, D.; Yoshimatsu, T.; Baden, T.; Shekhar, K. Comparative transcriptomic insights into the evolutionary origin of the tetrapod double cone. 2024, arXiv:2024.11.04.621990. [Google Scholar] [CrossRef]
- Engström, K. Cone Types and Cone Arrangements in Teleost Retinae1. Acta Zoöl. 1963, 44, 179–243. [Google Scholar] [CrossRef]
- Lyall, A.H. Cone Arrangements in Teleost Retinae. J. Cell Sci. 1957, S3-98, 189–201. [Google Scholar] [CrossRef]
- Baden, T. From water to land: Evolution of photoreceptor circuits for vision in air. PLOS Biol. 2024, 22, e3002422. [Google Scholar] [CrossRef]
- Yamagata, M.; Yan, W.; Sanes, J.R.; Biology, C.; States, U. A cell atlas of the chick retina based on single-cell transcriptomics. eLife 2021, 10. [Google Scholar] [CrossRef]
- Günther, A.; Haverkamp, S.; Irsen, S.; Watkins, P.V.; Dedek, K.; Mouritsen, H.; Briggman, K.L. Species–specific circuitry of double cone photoreceptors in two avian retinas. Commun. Biol. 2024, 7, 1–11. [Google Scholar] [CrossRef]
- Günther, A.; Dedek, K.; Haverkamp, S.; Irsen, S.; Briggman, K.L.; Mouritsen, H. Double Cones and the Diverse Connectivity of Photoreceptors and Bipolar Cells in an Avian Retina. J. Neurosci. 2021, 41, 5015–5028. [Google Scholar] [CrossRef] [PubMed]
- Kelber, A. Bird colour vision – from cones to perception. Curr. Opin. Behav. Sci. 2019, 30, 34–40. [Google Scholar] [CrossRef]
- Zeng, H. What is a cell type and how to define it? Cell 2022, 185, 2739–2755. [Google Scholar] [CrossRef] [PubMed]
- Goetz, J.; Jessen, Z.F.; Jacobi, A.; Mani, A.; Cooler, S.; Greer, D.; Kadri, S.; Segal, J.; Shekhar, K.; Sanes, J.R.; et al. Unified classification of mouse retinal ganglion cells using function, morphology, and gene expression. Cell Rep. 2022, 40, 111040–111040. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Xu, Q.; Su, J.; Tang, L.; Hao, Z.-Z.; Xu, C.; Liu, R.; Shen, Y.; Sang, X.; Xu, N.; et al. Linking transcriptomes with morphological and functional phenotypes in mammalian retinal ganglion cells. Cell Rep. 2022, 40, 111322. [Google Scholar] [CrossRef]
- Bae, J.A.; Mu, S.; Kim, J.S.; Turner, N.L.; Tartavull, I.; Kemnitz, N.; Jordan, C.S.; Norton, A.D.; Silversmith, W.M.; Prentki, R.; et al. Digital Museum of Retinal Ganglion Cells with Dense Anatomy and Physiology. Cell 2018, 173, 1293–1306.e19. [Google Scholar] [CrossRef] [PubMed]
- Baden, T.; Berens, P.; Franke, K.; Rosón, M.R.; Bethge, M.; Euler, T. The functional diversity of retinal ganglion cells in the mouse. Nature 2016, 529, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; et al. Avian photoreceptor homologies and the origin of double cones. Curr. Biol. 2025, in press.
- Rheaume, B.A.; Jereen, A.; Bolisetty, M.; Sajid, M.S.; Yang, Y.; Renna, K.; Sun, L.; Robson, P.; Trakhtenberg, E.F. Single cell transcriptome profiling of retinal ganglion cells identifies cellular subtypes. Nat. Commun. 2018, 9, 2759. [Google Scholar] [CrossRef] [PubMed]
- Keeley, P.W.; Eglen, S.J.; Reese, B.E. Author response for "From Random to Regular: Variation in the Patterning of Retinal Mosaics". 2020. [Google Scholar] [CrossRef]
- Wässle, H.; Puller, C.; Müller, F.; Haverkamp, S. Cone Contacts, Mosaics, and Territories of Bipolar Cells in the Mouse Retina. J. Neurosci. 2009, 29, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Field, G.D.; Gauthier, J.L.; Sher, A.; Greschner, M.; Machado, T.A.; Jepson, L.H.; Shlens, J.; Gunning, D.E.; Mathieson, K.; Dabrowski, W.; et al. Functional connectivity in the retina at the resolution of photoreceptors. Nature 2010, 467, 673–677. [Google Scholar] [CrossRef]
- Helmstaedter, M.; Briggman, K.L.; Turaga, S.C.; Jain, V.; Seung, H.S.; Denk, W. Connectomic reconstruction of the inner plexiform layer in the mouse retina. Nature 2013, 500, 168–174. [Google Scholar] [CrossRef]
- Behrens, C.; Schubert, T.; Haverkamp, S.; Euler, T.; Berens, P. ; Germany Connectivity map of bipolar cells and photoreceptors in the mouse retina. eLife 2016, 5. [Google Scholar] [CrossRef]
- Shekhar, K.; Lapan, S.W.; Whitney, I.E.; Tran, N.M.; Macosko, E.Z.; Kowalczyk, M.; Adiconis, X.; Levin, J.Z.; Nemesh, J.; Goldman, M.; et al. Comprehensive Classification of Retinal Bipolar Neurons by Single-Cell Transcriptomics. Cell 2016, 166, 1308–1323.e30. [Google Scholar] [CrossRef]
- Yan, W.; Laboulaye, M.A.; Tran, N.M.; Whitney, I.E.; Benhar, I.; Sanes, J.R. Mouse Retinal Cell Atlas: Molecular Identification of over Sixty Amacrine Cell Types. J. Neurosci. 2020, 40, 5177–5195. [Google Scholar] [CrossRef]
- Hahn, J.; Monavarfeshani, A.; Qiao, M.; Kao, A.H.; Kölsch, Y.; Kumar, A.; Kunze, V.P.; Rasys, A.M.; Richardson, R.; Wekselblatt, J.B.; et al. Evolution of neuronal cell classes and types in the vertebrate retina. Nature 2023, 624, 415–424. [Google Scholar] [CrossRef]
- Franke, K.; Berens, P.; Schubert, T.; Bethge, M.; Euler, T.; Baden, T. Inhibition decorrelates visual feature representations in the inner retina. Nature 2017, 542, 439–444. [Google Scholar] [CrossRef]
- Della Santina, L.; Kuo, S.P.; Yoshimatsu, T.; Okawa, H.; Suzuki, S.C.; Hoon, M.; Tsuboyama, K.; Rieke, F.; Wong, R.O. Glutamatergic Monopolar Interneurons Provide a Novel Pathway of Excitation in the Mouse Retina. Curr. Biol. 2016, 26, 2070–2077. [Google Scholar] [CrossRef] [PubMed]
- Chapot, C. A.; Euler, T. & Schubert, T. How do horizontal cells ‘talk’ to cone photoreceptors? Different levels of complexity at the cone-horizontal cell synapse. J. Physiol. 2017, 595, 5495–5506. [Google Scholar]
- Wang, J.; Zhang, L.; Cavallini, M.; Pahlevan, A.; Sun, J.; Morshedian, A.; Fain, G.L.; Sampath, A.P.; Peng, Y.-R. Molecular characterization of the sea lamprey retina illuminates the evolutionary origin of retinal cell types. Nat. Commun. 2024, 15, 1–21. [Google Scholar] [CrossRef]
- Yoshimatsu, T.; Bartel, P.; Schröder, C.; Janiak, F.K.; St-Pierre, F.; Berens, P.; Baden, T. Ancestral circuits for vertebrate color vision emerge at the first retinal synapse. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef]
- Guenther, A.; et al. Morphology and connectivity of retinal horizontal cells in two avian species. 2025.01.27. 2025, arXiv:2025.01.27.634460. [Google Scholar] [CrossRef]
- Cameron Cowan, A.S.; Renner, M.; De Gennaro, M.; Roma, G.; Nigsch, F.; Roska Correspondence, B.; Cowan, C.S.; Gross-Scherf, B.; Goldblum, D.; Hou, Y.; et al. Cell Types of the Human Retina and Its Organoids at Single-Cell Resolution. Cell 2020, 182. [Google Scholar] [CrossRef]
- Volkov, L.I.; Ogawa, Y.; Somjee, R.; Vedder, H.E.; Powell, H.E.; Poria, D.; Meiselman, S.; Kefalov, V.J.; Corbo, J.C. Samd7 represses short-wavelength cone genes to preserve long-wavelength cone and rod photoreceptor identity. Proc. Natl. Acad. Sci. 2024, 121. [Google Scholar] [CrossRef] [PubMed]
- Angueyra, J.M.; Kunze, V.P.; Patak, L.K.; Kim, H.; Kindt, K.; Li, W.; Neurophysiology, U.O.R.; Function; Disorders, O. C.; States, U. Transcription factors underlying photoreceptor diversity. eLife 2023, 12. [Google Scholar] [CrossRef]
- Fornetto, C.; Euler, T.; Baden, T. Vertebrate vision is ancestrally based on competing cone circuits. 2024.11.19. 2024, arXiv:2024.11.19.624320. [Google Scholar] [CrossRef]
- Hofer, H.; Carroll, J.; Neitz, J.; Neitz, M.; Williams, D.R. Organization of the Human Trichromatic Cone Mosaic. J. Neurosci. 2005, 25, 9669–9679. [Google Scholar] [CrossRef]
- Applebury, M. L.; et al. The Murine Cone Photoreceptor: A Single Cone Type Expresses Both S and M Opsins with Retinal Spatial Patterning. Neuron 2000, 27, 513–523. [Google Scholar] [CrossRef]
- Baden, T.; Schubert, T.; Chang, L.; Wei, T.; Zaichuk, M.; Wissinger, B.; Euler, T. A Tale of Two Retinal Domains: Near-Optimal Sampling of Achromatic Contrasts in Natural Scenes through Asymmetric Photoreceptor Distribution. Neuron 2013, 80, 1206–1217. [Google Scholar] [CrossRef]
- Szél, Á.; Röhlich, P.; Gaffé, A.R.; Juliusson, B.; Aguirre, G.; Van Veen, T. Unique topographic separation of two spectral classes of cones in the mouse retina. J. Comp. Neurol. 1992, 325, 327–342. [Google Scholar] [CrossRef]
- Lagman, D.; Callado-Pérez, A.; Franzén, I.E.; Larhammar, D.; Abalo, X.M. Transducin Duplicates in the Zebrafish Retina and Pineal Complex: Differential Specialisation after the Teleost Tetraploidisation. PLOS ONE 2015, 10, e0121330–e0121330. [Google Scholar] [CrossRef]
- Seifert, M.; Roberts, P.A.; Kafetzis, G.; Osorio, D.; Baden, T. Birds multiplex spectral and temporal visual information via retinal On- and Off-channels. Nat. Commun. 2023, 14, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-Y.; Hassin, G.; Witkovsky, P. Blue-sensitive rod input to bipolar and ganglion cells of theXenopus retina. Vis. Res. 1983, 23, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Rozenblit, F.; Gollisch, T. What the salamander eye has been telling the vision scientist's brain. Semin. Cell Dev. Biol. 2020, 106, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Donner, K.; Yovanovich, C.A. A frog’s eye view: Foundational revelations and future promises. Semin. Cell Dev. Biol. 2020, 106, 72–85. [Google Scholar] [CrossRef] [PubMed]
- A Cowing, J.; A Arrese, C.; Davies, W.L.; Beazley, L.D.; Hunt, D.M. Cone visual pigments in two marsupial species: the fat-tailed dunnart (Sminthopsis crassicaudata) and the honey possum (Tarsipes rostratus). Proc. R. Soc. B: Biol. Sci. 2008, 275, 1491–1499. [Google Scholar] [CrossRef]
- Hauzman, E. Adaptations and evolutionary trajectories of the snake rod and cone photoreceptors. Semin. Cell Dev. Biol. 2020, 106, 86–93. [Google Scholar] [CrossRef]
- Dunn, R.F. Studies on the retina of the gecko Coleonyx variegatus. J. Ultrastruct. Res. 1966, 16, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Kojima, K.; Matsutani, Y.; Yanagawa, M.; Imamoto, Y.; Yamano, Y.; Wada, A.; Shichida, Y.; Yamashita, T. Evolutionary adaptation of visual pigments in geckos for their photic environment. Sci. Adv. 2021, 7, eabj1316. [Google Scholar] [CrossRef] [PubMed]
- Röll, B. Gecko vision—retinal organization, foveae and implications for binocular vision. Vis. Res. 2001, 41, 2043–2056. [Google Scholar] [CrossRef] [PubMed]
- Fain, G.L. Lamprey vision: Photoreceptors and organization of the retina. 2019, 106, 5–11. [Google Scholar] [CrossRef]
- Nadal-Nicolás, F.M.; Kunze, V.P.; Ball, J.M.; Peng, B.T.; Krishnan, A.; Zhou, G.; Dong, L.; Li, W.; Section, R.N.; States, U. True S-cones are concentrated in the ventral mouse retina and wired for color detection in the upper visual field. eLife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Carleton, K.L.; Yourick, M.R. Axes of visual adaptation in the ecologically diverse family Cichlidae. Semin. Cell Dev. Biol. 2020, 106, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.L.; Gan, K.J.; Flamarique, I.N. Thyroid Hormone Induces a Time-Dependent Opsin Switch in the Retina of Salmonid Fishes. Investig. Opthalmology Vis. Sci. 2009, 50, 3024–3032. [Google Scholar] [CrossRef]
- Nandamuri, S.P.; Yourick, M.R.; Carleton, K.L. Adult plasticity in African cichlids: Rapid changes in opsin expression in response to environmental light differences. Mol. Ecol. 2017, 26, 6036–6052. [Google Scholar] [CrossRef] [PubMed]
- Cortesi, F.; Mitchell, L.J.; Tettamanti, V.; Fogg, L.G.; de Busserolles, F.; Cheney, K.L.; Marshall, N.J. Visual system diversity in coral reef fishes. Semin. Cell Dev. Biol. 2020, 106, 31–42. [Google Scholar] [CrossRef]
- Frau, S.; Novales Flamarique, I.; Keeley, P.; Muñoz-Cueto, J. & Reese, B. Straying from the flatfish retinal plan: Cone photoreceptor patterning in the common sole (Solea solea) and the Senegalese sole (Solea senegalensis). J. Comp. Neurol. 2020, 528. [Google Scholar]
- Arbogast, P.; Flamant, F.; Godement, P.; Glösmann, M. & Peichl, L. Thyroid Hormone Signaling in the Mouse Retina. PLoS ONE 2016, 11, e0168003. [Google Scholar]
- Roberts, M.R.; Srinivas, M.; Forrest, D.; de Escobar, G.M.; Reh, T.A. Making the gradient: Thyroid hormone regulates cone opsin expression in the developing mouse retina. Proc. Natl. Acad. Sci. 2006, 103, 6218–6223. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, T. Thyroid hormone and seasonal regulation of reproduction. 2013, 34, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Dalton, B.E.; Lu, J.; Leips, J.; Cronin, T.W.; Carleton, K.L. Variable light environments induce plastic spectral tuning by regional opsin coexpression in the African cichlid fish, Metriaclima zebra. Mol. Ecol. 2015, 24, 4193–4204. [Google Scholar] [CrossRef]
- Morshedian, A.; et al. Cambrian origin of the CYP27C1-mediated vitamin A1-to-A2 switch, a key mechanism of vertebrate sensory plasticity. R. Soc. Open Sci. 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Enright, J.M.; Toomey, M.B.; Sato, S.-Y.; Temple, S.E.; Allen, J.R.; Fujiwara, R.; Kramlinger, V.M.; Nagy, L.D.; Johnson, K.M.; Xiao, Y.; et al. Cyp27c1 Red-Shifts the Spectral Sensitivity of Photoreceptors by Converting Vitamin A1 into A2. Curr. Biol. 2015, 25, 3048–3057. [Google Scholar] [CrossRef]
- Carleton, K.L.; Escobar-Camacho, D.; Stieb, S.M.; Cortesi, F.; Marshall, N.J. Seeing the rainbow: mechanisms underlying spectral sensitivity in teleost fishes. J. Exp. Biol. 2020, 223. [Google Scholar] [CrossRef]
- Herzog, T.; et al. A heterogeneous population code at the first synapse of vision. 2024, arXiv:2024.05.03.592379. [Google Scholar] [CrossRef]
- Yoshimatsu, T.; Schröder, C.; Nevala, N.E.; Berens, P.; Baden, T. Fovea-like Photoreceptor Specializations Underlie Single UV Cone Driven Prey-Capture Behavior in Zebrafish. Neuron 2020, 107, 320–337.e6. [Google Scholar] [CrossRef]
- Baudin, J.; Angueyra, J.M.; Sinha, R.; Rieke, F.; States, U. S-cone photoreceptors in the primate retina are functionally distinct from L and M cones. eLife 2019, 8. [Google Scholar] [CrossRef]
- Li, Y.N.; Tsujimura, T.; Kawamura, S.; Dowling, J.E. Bipolar cell–photoreceptor connectivity in the zebrafish (Danio rerio) retina. J. Comp. Neurol. 2012, 520, 3786–3802. [Google Scholar] [CrossRef]
- Ali, M.-Ather. & Anctil, Michel. Retinas of Fishes : An Atlas. Springer: Berlin Heidelberg, 1976.
- Meier, A.; Nelson, R.; Connaughton, V.P. Color Processing in Zebrafish Retina. Front. Cell. Neurosci. 2018, 12, 327. [Google Scholar] [CrossRef]
- Stell, W.K.; Hárosi, F.I. Cone structure and visual pigment content in the retina of the goldfish. Vis. Res. 1976, 16, 647–IN4. [Google Scholar] [CrossRef]
- Allison, W.T.; Barthel, L.K.; Skebo, K.M.; Takechi, M.; Kawamura, S.; Raymond, P.A. Ontogeny of cone photoreceptor mosaics in zebrafish. J. Comp. Neurol. 2010, 518, 4182–4195. [Google Scholar] [CrossRef]
- Flamarique, I.N.; Hawryshyn, C.W. No Evidence of Polarization Sensitivity in Freshwater Sunfish from Multi-unit Optic Nerve Recordings. Vis. Res. 1997, 37, 967–973. [Google Scholar] [CrossRef]
- Zimmermann, M.J.; Nevala, N.E.; Yoshimatsu, T.; Osorio, D.; Nilsson, D.-E.; Berens, P.; Baden, T. Zebrafish Differentially Process Color across Visual Space to Match Natural Scenes. Curr. Biol. 2018, 28, 2018–2032.e5. [Google Scholar] [CrossRef]
- Shand, J.; Archer, M.A.; Collin, S.P. Ontogenetic changes in the retinal photoreceptor mosaic in a fish, the black bream,Acanthopagrus butcheri. J. Comp. Neurol. 1999, 412, 203–217. [Google Scholar] [CrossRef]
- Ogawa, Y.; Corbo, J.C. Partitioning of gene expression among zebrafish photoreceptor subtypes. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Lind, O.; Chavez, J.; Kelber, A. The contribution of single and double cones to spectral sensitivity in budgerigars during changing light conditions. Journal of Comparative Physiology A 2013, 200, 197–207. [Google Scholar] [CrossRef]
- Enright, J.M.; Lawrence, K.A.; Hadzic, T.; Corbo, J.C. Transcriptome profiling of developing photoreceptor subtypes reveals candidate genes involved in avian photoreceptor diversification. J. Comp. Neurol. 2014, 523, 649–668. [Google Scholar] [CrossRef]
- Raymond, P.A.; Barthel, L.K. A moving wave patterns the cone photoreceptor mosaic array in the zebrafish retina. Int. J. Dev. Biol. 2004, 48, 935–945. [Google Scholar] [CrossRef]
- Ogawa, Y.; et al. Conservation of cis-regulatory codes over half a billion years of evolution. 2024, arXiv:2024.11.13.623372. [Google Scholar] [CrossRef]
- Roberts, R.J.V.; Pop, S.; Prieto-Godino, L.L. Evolution of central neural circuits: state of the art and perspectives. Nat. Rev. Neurosci. 2022, 23, 725–743. [Google Scholar] [CrossRef] [PubMed]
- Swaroop, A.; Kim, D.; Forrest, D. Transcriptional regulation of photoreceptor development and homeostasis in the mammalian retina. Nat. Rev. Neurosci. 2010, 11, 563–576. [Google Scholar] [CrossRef]
- Xie, S.; et al. Knockout of Nr2e3 prevents rod photoreceptor differentiation and leads to selective L-/M-cone photoreceptor degeneration in zebrafish. Biochim. Biophys. Acta BBA - Mol. Basis Dis. 2019, 1865, 1273–1283. [Google Scholar] [CrossRef] [PubMed]
- Hudson, L.G.; Santon, J.B.; Glass, C.K.; Gill, G.N. Ligand-activated thyroid hormone and retinoic acid receptors inhibit growth factor receptor promoter expression. Cell 1990, 62, 1165–1175. [Google Scholar] [CrossRef]
- Ng, L.; Hurley, J.B.; Dierks, B.; Srinivas, M.; Saltó, C.; Vennström, B.; Reh, T.A.; Forrest, D. A thyroid hormone receptor that is required for the development of green cone photoreceptors. Nat. Genet. 2001, 27, 94–98. [Google Scholar] [CrossRef]
- Suzuki, S.C.; Bleckert, A.; Williams, P.R.; Takechi, M.; Kawamura, S.; Wong, R.O.L. Cone photoreceptor types in zebrafish are generated by symmetric terminal divisions of dedicated precursors. Proc. Natl. Acad. Sci. 2013, 110, 15109–15114. [Google Scholar] [CrossRef]
- Ogawa, Y.; Shiraki, T.; Asano, Y.; Muto, A.; Kawakami, K.; Suzuki, Y.; Kojima, D.; Fukada, Y. Six6 and Six7 coordinately regulate expression of middle-wavelength opsins in zebrafish. Proc. Natl. Acad. Sci. 2019, 116, 4651–4660. [Google Scholar] [CrossRef]
- Ogawa, Y.; Shiraki, T.; Kojima, D.; Fukada, Y. Homeobox transcription factor Six7 governs expression of green opsin genes in zebrafish. Proc. R. Soc. B: Biol. Sci. 2015, 282, 20150659. [Google Scholar] [CrossRef]
- Ogawa, Y.; Shiraki, T.; Fukada, Y.; Kojima, D. Foxq2 determines blue cone identity in zebrafish. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, A.; Yamamoto, A.; Yaguchi, J. & Yaguchi, S. cis-Regulatory analysis for later phase of anterior neuroectoderm-specific foxQ2 expression in sea urchin embryos. genesis 2019, 57, e23302. [Google Scholar]
- Wakefield, M.J.; Anderson, M.; Chang, E.; Wei, K.-J.; Kaul, R.; Graves, J.A.M.; Grützner, F.; Deeb, S.S. Cone visual pigments of monotremes: Filling the phylogenetic gap. Vis. Neurosci. 2008, 25, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Davies, W.L.; Carvalho, L.S.; Cowing, J.A.; Beazley, L.D.; Hunt, D.M.; Arrese, C.A. Visual pigments of the platypus: A novel route to mammalian colour vision. Curr. Biol. 2007, 17, R161–R163. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Delfin, K.; Morris, A.C.; Snelson, C.D.; Gamse, J.T.; Gupta, T.; Marlow, F.L.; Mullins, M.C.; Burgess, H.A.; Granato, M.; Fadool, J.M. Tbx2b is required for ultraviolet photoreceptor cell specification during zebrafish retinal development. Proc. Natl. Acad. Sci. 2009, 106, 2023–2028. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



