
Submitted:
02 August 2023
Posted:
04 August 2023
You are already at the latest version
Abstract
The ability to perceive and respond to light stimuli is fundamental not only for spatial vision, but also to many other light mediated interactions with the environment. In animals, light perception is performed by specific cells known as photoreceptors and, at molecular level, by a group of GPCRs known as Opsins. Sea urchin larvae possess a group of photoreceptor cells (PRCs), deploying a Go-Opsin (Opsin3.2), which have been shown to share transcription factors and morphology with PRCs of the ciliary type, contributing to raising new questions on how this sea urchin larva PRC is specified and whether it shares a common ancestor with ciliary PRCs or it evolved independently through convergent evolution.
To answer these questions, we combined immunohistochemistry and fluorescent in situ hybridization to investigate how the Opsin3.2 PRCs develop in the sea urchin Strogylocentrotus purpuratus larva. Subsequently, we applied single cell transcriptomics to investigate the molecular signature of the Sp-Opsin3.2 cells, and show that they deploy an ancient regulatory program responsible for photoreceptors specification. Finally, we also discuss the possible functions of the Opsin3.2 cells based on their molecular fingerprint, and we suggest that they are involved in a variety of signaling pathways, including those entailing the thyrotropin-releasing hormone.
Keywords:
Opsin
; Photoreceptors
; Evolution
; Sea Urchin
1. Introduction
The ability to perceive and respond to environmental conditions, such as light and temperature variations, is fundamental to all organisms. Moreover, many of these conditions undergo cyclic variations and many organisms, including animals, developed mechanisms to anticipate and synchronize their activities to the external periodicities: e.g., circadian and circalunar rhythms [1,2]. One of the main sources of information to entrain such rhythms is light [3]. Nonetheless, light is used to drive a variety of other behaviors and physiological processes. Well-known are the roles of light in spatial vision, which requires specialized structures such as eyes or eyespots and a specific organization of the nervous system [4], and the one in non-directional UV-avoidance behaviors that have been widely described in marine larvae [5].
Light perception is executed by specialized photoreceptor cells (PRCs). In order to increase the photosensitive surface available, PRCs possess different types of membrane extensions [6,7] and can be classified accordingly. Bilaterians mainly possess two types of PRCs: ciliary photoreceptors (c-PRCs) and rhabdomeric photoreceptors (r-PRCs). These two types of photoreceptors are diversified both at morphological and molecular levels, with the latter being characterized by different transcription factor (TF) repertoires guiding their specification and distinct phototransduction cascades [7].
The molecules that allow photoreceptors to translate light stimuli into intracellular signals are proteins called opsins. Animal opsins are membrane proteins, members of the G-Protein Coupled Receptor (GPCR) family, which are able to detect light stimuli due to a Schiff base linkage to a chromophore [8,9]. The chromophore is usually 11-cis-retinal and the linkage happens at a specific lysine residue situated in the VII helix (K296 in the bovine rhodopsin, the first opsin to be fully characterized at the level of primary amino acid sequence and 3D structure and often used as reference for comparisons with other opsins [8,10]. The presence of this specific residue, therefore, can be used as a fingerprint indication to identify a GPCR as bona fide opsin.
Phylogenetic analyses have revealed a widespread distribution of opsins in metazoans and have classified them into a variety of main groups, depending on which phyla were taken into account for the analysis [9,11,12,13,14]. Recent studies have classified opsins into the following groups: ciliary opsins (or c-opsins, containing vertebrate visual pigments), rhabdomeric opsins (or r-opsins, containing for example the melanopsins), neuropsins, peropsins, RGR opsins, Go-opsins and cnidopsin [13,15,16,17,18,19].
Apart from the opsin classification, the molecular events following the opsin activation have been studied in great detail. When the chromophore absorbs a photon, it undergoes a conformational change becoming all-trans-retinal and activating the protein. The activated opsin subsequently binds a G-protein (guanine nucleotide-binding protein) activating a specific phototransduction cascade, depending on which G-protein type is involved. For instance, vertebrate visual opsins recruit Gt proteins with subsequent decrease in the intracellular cGMP levels, closure of ionic channels and final cell membrane hyperpolarization, which eventually inhibits the glutamate neurotransmitter release. In contrast, melanopsins and many invertebrate opsins, have been shown to activate a Gq-mediated transduction cascade. This involves a signal cascade initiated by the enzyme Phospholipase C (PLC) and eventually induce membrane depolarization (reviewed in [14,20]). Moreover, a phototransduction cascade mediated by Go-opsins was discovered in scallop hyperpolarizing photoreceptors [21]. In contrast, Go-Opsin1 has been reported to mediate depolarization in Platynereis [22].
Ciliary and rhabdomeric photoreceptors were taught to be typically used for vision in deuterostome and protostome eyes, respectively. However, the finding of ciliary PRCs in protostomes [23,24,25], rhabdomeric photoreceptors in deuterostomes [26,27,28], and the molecular similarities shared by PRC types among different clades suggest that Urbilateria possessed both photoreceptor types [6,29,30]. Nonetheless, how these two different types of photoreceptors evolved still remains an open question. The sea urchin Strongylocentrotus purpuratus represents an interesting case to address this questions since it belongs to deuterostomes but utilizes both rhabdomeric and ciliary opsins [31,32] and its genome encodes for nine opsins: Opsin1, Opsin2, Opsin3.1, Opsin3.2, Opsin4, Opsin5, Opsin6, Opsin7 and Opsin8 [11]. Despite the description of the wide repertoire of opsins encoded in the sea urchin genome, very little is known on their expression patterns and functions. Recently, the sea urchin extraocular photoreceptor cells in Paracentrotus lividus at mature rudiment (when the adult rudiment formed in the left side of an 8-armed larva is fully mature and ready to go through metamorphosis) and juvenile stages was investigated. In brief, while the mature rudiment expresses only Opsin2 positive and Opsin4 positive r-PRCs, juveniles express all the aforementioned Opsins (Opsin1, Opsin2, Opsin3.1, Opsin3.2, Opsin4, and Opsin5) except Opsin6 and 7 [33]. Moreover, the study of Opsin1 and Opsin4 PRCs in the juveniles of P. lividus revealed an expression pattern similar to their orthologous in S. purpuratus juveniles [27,34], suggesting conservation of their functions in the two species. Less is known about Opsin expression in sea urchins at larval stages. The Go-Opsin, known as Opsin3.2 in S. purpuratus and P. lividus, was found expressed in two cells located at the sides of the apical organ in S. purpuratus [35] and Hemicentrotus pulcherrimus [36]. The Sp-Opsin3.2 expressing PRCs, which have been previously described as non-directional [35], also express evolutionary conserved developmental transcription factors similar to what is found in c-PRCs of other animals, typically deploying a c-opsin. However, it remains to be clarified if the regulatory programs of ciliary and Go-Opsin photoreceptor cells derive from the same ancestor photoreceptor cell program and, in this case, how they diversified during evolution.
Sea urchin larvae also express the Opsin2, in a few cells localized in the oral and post oral arms, likely belonging to a mesenchymal cell population [37]. Sea urchin Opsin2 was originally identified as belonging to the echinoderm specific Echinopsin A type [11], but most recently it has been re-clustered in a new group called Bathyopsins [17]. Nonetheless, nothing else is known about the sea urchin larva Opsin2 photoreceptors in terms of regulatory state and cell identity.
The single cell omics approaches have enabled the identification of cell types and helped in reconstructing their evolution at an unprecedented level. Single cell RNA sequencing (scRNA-seq) has been successfully applied to echinoderm embryonic and larval stages to investigate animal evolution from a cell type perspective [38,39,40]. In addition, scRNA-seq of P. lividus mature rudiments has revealed the presence of distinct PRC populations utilising an evolutionary conserved photoreceptor regulatory program [33].
Here we used fluorescent in situ hybridization (FISH) and immunohistochemistry (IHC) to identify and trace, during S. purpuratus development, a Go-Opsin (Sp-Opsin3.2) positive neuronal cell type. Using single cell transcriptomics, we were able to reconstruct the molecular identity of these photoreceptor cells, contributing to disentangling their evolution and predicting their function. Moreover, we compared the profiles of the Opsin3.2 positive cells with the single cell transcriptomes of the other PRCs present in the S. purpuratus larva, the Opsin2 positive cells [37]. Through this analysis, we showed that the Opsin3.2 PRCs employ an evolutionary conserved photoreceptor genetic program and are able to produce multiple neurotransmitters, suggestive of multifunctionality; we also partially reconstructed the putative phototransduction cascade activated within these cells, as predicted by single cell transcriptomics. Furthermore, comparison between the two photoreceptors showed many similarities but also significant differences at the level of TFs expressed and activated transduction cascade. For example, while the Opsin3.2 PRCs are neurons associated to the apical plate of the larva, thus possibly from ectodermal origin, the Opsin2 PRCs have a mesodermally derived muscle and immune cell identity. Last but not least, we investigated the presence of the circadian rhythm machinery in the Opsin3.2 and Opsin2 PRCs finding no significant expression of genes that were suggested to be part of the sea urchin master clock [41].
2. Materials and Methods
2.1. Animal Husbandry
Adult S. purpuratus were obtained from Patrick Leahy (Kerckhoff Marine Laboratory, California Institute of Technology, Pasadena, CA, 648 USA) and maintained in circulating seawater at Stazione Zoologica Anton Dohrn in Naples at 15 °C. Gametes were obtained by vigorous shaking the adult sea urchins. The sperm was collected dry using a Pasteur pipette and stored at 4 °C until usage. To collect eggs, females were inverted over a beaker filled with diluted 9:1 (9 parts Mediterranean Sea seawater and 1 part distilled water) FMSW. About 20 ml of eggs were fertilized, adding a few drops of sperm diluted 1:10000. Embryos were transferred in FMSW and reared at 15 °C under a 12 h light/12 h dark cycle. Larval cultures were maintained by exchanging half of the FMSW with fresh FMSW 2 times per week. After 3 days post fertilization (dpf) pluteus stage, the larvae were fed 3 times per week with the unicellular micro-algae Dunaliella sp. at an approximate concentration of 1000 cells/mL.
2.2. Fluorescent In Situ Hybridization (FISH) and Immunohistochemistry (IHC)
Whole mount RNA Fluorescent in situ hybridization and combined FISH-IHC were performed as described in [42,43]. Summarizing, specimens at different developmental stages were collected and fixed in Fixative I (4% PFA in 0.1 M MOPS and 0.5 M NaCl) for at least one night at 4 °C. subsequently, samples were washed 3 times with MOPS buffer (0.1 M MOPS, 0.5 M NaCl, 0.01% Tween 20) for 15 minutes at RT, dehydrated in 70% ethanol and finally stored at −20 °C until usage. Antisense probes were transcribed from linearized DNA and labelled during transcription using Digoxigenin (Roche) or Fluorescein (Roche) labelled ribonucleotides following the manufacturer’s instructions. Trh probe, instead, was DNP labelled as described in details in [42,43]. Primer sequences used for cDNA isolation and probes synthesis are included in Supplementary Table S1. Fluorescent signal was developed via using fluorophore conjugated tyramide technology (Perkin Elmer, Cat. #NEL752001KT). For combined FISH-HIC, after tyramide amplification step, samples were incubated in blocking (containing 1 mg/ml Bovine Serum Albumin and 4% Sheep Serum in PBS) for 1 h at RT, then transferred in primary antibody diluted 1:400 in blocking O/N at 4 °C. Samples were washed 4–6 times with PBS 1×, then stained with appropriate Alexa Fluor secondary antibodies (488 rabbit, 555 rat) diluted 1:1000 in blocking, and finally washed 4–6 times with PBS 1×. DAPI (10 mg/ml stock) was added to the samples at a final dilution of 1:10000 to stain nuclei. Specimens were imaged using a Zeiss LSM 700 confocal microscope and pictures analyzed using ImageJ. To stain the Opsin3.2 positive cells we used an anti Sp-Opsin3.2 gifted by Dr. Robert D Burke. To stain the TRHergic cells we used a custom antibody produced for us by GenScript adapting the method used by [44]. Briefly, the Sp-TRH mature amidated peptide (QYPGa) was coupled to an immunogen (Keyhole limpet haemocyanin, KLH) via an N-terminal cysteine. Subsequently the KLH-CQYPGa was used to immunize rabbits. Sera were affinity purified against the antigen by the company which obtained nine antibody fractions. All the fractions were tested by Elisa with different sea urchin neuropeptides to find the most specific ones which were used to perform immunohistochemistry [45].
2.3. Larvae Dissociation
Dissociation of the five dpf Strongylocentrotus purpuratus plutei into single cells was performed as described in [40]. In brief, Larvae were collected, seawater was removed, and larvae were resuspended in Ca2+ Mg2+-free artificial sea water and then passed to dissociation buffer containing 1 M glycine and 0.02 M EDTA in Ca2+ Mg2+-free artificial sea water. Larvae were incubated for 10 min on ice and mixed gently via pipette aspiration every 2 min. From that point and onwards the progress of dissociation was monitored. Cell viability was assessed via using Propidium Iodide and Fluorescein diacetate and only specimens with cell viability ≥ 90% were further processed. Single cells were counted using a haemocytometer and diluted according to the manufacturer’s protocol (10× Genomics). Throughout this procedure samples were kept at 4 °C.
2.4. Single-Cell RNA Sequencing and Data Analysis
Single cell RNA sequencing was performed using the 10× Genomics single-cell capturing system. Specimens from two independent biological replicates were loaded on the 10× Genomics Chromium Controller. Single cell cDNA libraries were prepared using the Chromium Single Cell 3′ Reagent Kit (Chemistry v3). Libraries were sequenced by GeneCore (EMBL, Heidelberg, Germany) for 75 bp paired-end reads (Illumina NextSeq 500). ScRNA-seq output reads were aligned and analyzed using Cell Ranger Software Suite 3.0.2 (10× Genomics) The genomic index was made in Cell Ranger using the S. purpuratus genome version 3.1 [46,47]. Cell Ranger output matrices for two biological replicates were used for further analysis in the Seurat v4 R package [48]. The analysis was performed according to the Seurat scRNA-seq R package documentation [48,49]. Genes that are transcribed in less than three cells and cells that have less than a minimum of 250 transcribed genes were excluded from the analysis. Datasets were normalized and variable genes were found using the vst method with a maximum of 2000 variable features. Data integration was performed via identification of anchors between the two different objects. Nearest Neighbor (SNN) graph was computed with 36 dimensions (resolution 1.0) to identify the clusters. Uniform Manifold Approximate and Projection (UMAP) was used to perform clustering dimensionality reduction. Cluster markers were found using the genes that are detected in at least 0.01 fraction of min.pct cells in the two clusters. The 3 dpf scRNA-seq [40,50] and 5 dpf datasets were merged using the harmony package (v0.1.0._and 30 dimensions) [51] Transcripts of all genes per cell type were identified by converting a Seurat DotPlot with all these transcripts as features into a table (ggplot2 3.2.0 R package). Subsetting analysis was performed by selecting all the cells present in neurons 4 and immune cells that express Opsin3.2 and Opsin2 respectively, through the Subset function incorporated into the Seurat R package.
3. Results
3.1. Expression Pattern of Opsin3.2 (Go-Opsin) during Larval Development
To investigate the expression pattern of the Sp-Opsin3.2 gene during S. purpuratus larva development, we combined IHC and FISH performed on larvae collected at different developmental stages, from the early 2-armed pluteus (3 dpf) to the 8-armed pluteus (5 wpf). At the 3 dpf pluteus stage, Sp-Opsin3.2 FISH revealed the presence of one or two positive cells located in the area above the mouth, distally to the larval apical organ (Figure 1A–B′). The apical organ is a sensory organ present in many marine larvae involved in settlement and metamorphosis [52,53,54]. In sea urchin larvae, it is located between the oral arms, just above the mouth, and mainly consists of a group of serotonergic neurons [31,52,55]. In the 4 dpf larvae, the number of cells positive for Sp-Opsin3.2 transcripts increases to two or three (Figure 1C–D′), while at 5 dpf they reach a total number of 3 to 4 cells (Figure 1E,F). At 6-arm pluteus stage (4 wpf), we could detect the presence of Opsin3.2 ganglia (Figure 1G,G′), situated in the same region in respect to the apical organ right above the mouth and below the joint with the oral arms. These ganglia remain up to the 8-arm competent to metamorphosis pluteus larva (5 weeks post fertilization) (Figure 1G–H″).
Previous studies have demonstrated the presence of neurons that produce the Thyrotropin releasing hormone neuropeptide (TRH) in cells located in similar regions as the Opsin3.2 positive neurons [40,56]. In order to understand whether the TRH positive cells (TRHergic neurons) are in close proximity to the Opsin3.2 PRCs and if TRH could be produced by the Opsin3.2 PRCs, we performed double FISH and double IHC for Sp-Opsin3.2 and Sp-Trh. Interestingly, the co-localization of both proteins and Sp-Trh transcripts in the same neurons of the 3 dpf pluteus larva (Figure 1B,B′) indicate that Opsin3.2 PRCs are able to produce the neuropeptide TRH. Similarly, at 4 and 5 dpf pluteus stages, all the Sp-Opsin3.2 positive cells are also TRHergic, as shown by Sp-Opsin3.2/Sp-Trh double FISH combined with Sp-TRH immunostaining (Figure 1F) and TRH/Opsin3.2 double immunostaining combined with Sp-Trh FISH (Figure 1B–D′)
3.2. Characterization of the Go-Opsin3.2 and the Opsin2 Positive Cell Types at a Single Cell Resolution
ScRNA-seq is a powerful technique allowing to identify cell clusters or populations and finely disentangle their regulatory state (i.e., TFs expressed) and predict possible functions and interactions among cell types (based on which genes, signaling molecules or receptors are expressed). Such an approach was successfully applied to identify and characterize sea urchin larval cell populations [40,50,57] at early stages. Compared to the 3 dpf pluteus, more Opsin3.2 positive cells are present in the 5 dpf pluteus (See Figure 1). This observation, prompted us to perform scRNA-seq on S. purpuratus 5 dpf pluteus larvae and to combine it with the already available 3 dpf pluteus dataset to investigate PRC diversity and the Go-Opsin3.2 molecular fingerprint. Our analysis included single cell transcriptomes from 32,116 cells and resulted in the identification of 21 clusters corresponding to distinct cell types or closely related cell type families (Figure 2B,C). The quality of the clustering analysis was assessed using gene markers previously shown to label distinct cell type families at the 3 dpf S. purpuratus pluteus stage: Sp-Fbsl_2 and Sp-Btub2/3 (ciliary band); Sp-Hbn and Sp-Frizz5/8 (apical plate); Sp-Spec2a and SPU_006199 (aboral ectoderm); Sp-SynB and Sp-Chrna9_4 (neurons); Sp-Mhc and Sp-Mlckb (esophageal muscles), Sp-Nan2 and Sp-Vasa (coelomic pouches); Sp-MacpfA2 and Sp-Pks1 (immune cells); Sp-185/333B3d and Sp-Fcoll/II/IIIf (blastocoelar cells); Sp-Msp130 and Sp-Sm37 (skeletogenic cells); Sp-Hox11/13b, Sp-Cdx and Sp-Rfxc1l (posterior gut), Sp-Nkx6.1 and Sp-Pdx (pyloric sphincter); Sp-Hnf4, Sp-ManrC1a (stomach); Sp-Ptf1a and Sp-Cpa2L (exocrine pancreas-like); Sp-Ahrl and Sp-Trop1 (cardiac sphincter); Sp-Alpi and Sp-Brn1/2/4 (esophagus/oral ectoderm). Plotting of the average expression of the aforementioned gene markers resulted in a meaningful clustering reconstructing most of the well known larval cell types (Figure 2B,C).
To identify which cell clusters express our Go-Opsin of interest, the average expression of Sp-Opsin3.2 was plotted in each cluster (Figure 3A,B). These data show that only the Neurons (4) cluster contains cells that highly express the Opsin3.2 (Figure 3B). Notably, this is also the cluster having the most significant Sp-Trh expression levels (Figure 3A). We next plotted the Sp-Opsin2 average expression and found it enriched in esophageal muscles and immune cells suggesting a mesodermal origin for these cells, as already shown in another sea urchin species [37]. Moreover, our single cell data indicate that the Opsin2 positive cells belonging to the immune cells cluster could possibly be pigment cells, as judged by the co-expression of the pigment cell marker Sp-Pks1 in (Figure 3A). For our analysis, we choose to focus on the Opsin2 PRCs belonging to the Immune cell population (Figure 3B), therefore all of the following plots including Opsin2 positive cells have been produced selecting only the Pks1/Opsin2 positive cells.
Finally, to confirm the quality of our data, we plotted the average expression of transcription factors that were found through FISH to be either co-expressed with Opsin3.2 cells or in the Opsin3.2 region [18]. The list includes, but it’s not limited to Sp-Otx, SpTbx2/3 and Sp-Six3 which have been suggested to be part of an ancestral photoreceptor specification code (Figure 3C) [6]. The average expression of the selected genes was plotted in all the cell clusters and the main accumulation was found in the Apical Plate and Neuron (4) cells, while the Immune Cells had high expression only of the transcription factor Sp-Id (IDs are a family of helix-loop-helix proteins involved in regulating cell proliferation and differentiation) and low expression levels of Sp-Tbx2/3.
3.3. Molecular Signature of Photoreceptor Cells in S. purpuratus Larvae
To gain additional information on how the Opsin3.2 cells work, we exploited the scRNA-seq data to explore their molecular signature and compared it to the molecular signature of the Opsin2 PRCs. In particular, the analysis was specifically targeted to the Opsin3.2 expressing cells and the Opsin2 positive cells belonging to the Immune cell cluster.
Go-Opsin photoreceptors in invertebrates have been found to be either depolarizing, P. dumerilii [22] or hyperpolarizing, scallop [21]. To determine which type of signaling is activated by the stimulation of the Opsin3.2 PRCs in the sea urchin larva, the expression levels of all the S. purpuratus genes that are known to be part of phototransduction cascades in other organisms were plotted in the Opsin3.2 cells (Figure 4A). Such gene expression profiles were plotted also in the Opsin2 cells belonging to the Immune cells to investigate the differences of the two cascades (Figure 4A). Unexpectedly, the G-protein type most highly expressed in the Go-Opsin PRCs are the Sp-Gs, Sp-Gi, and Sp-GbA, while the G proteins annotated as Sp-Go and Sp-GoI (https://www.echinobase.org/entry/) are lowly expressed, suggesting that the first types of G proteins activated by the Go-Opsin3.2 are Gs, Gi and GbA. Nonetheless, other putative components of the phototransduction cascade are only lowly expressed and it is therefore difficult to make any further prediction on the activated cascade. In any case, intriguing is the expression, although in a small percentage of cells and at low levels, of many genes annotated as phosphodiesterases (PDEs), which might be indicative of a hyperpolarizing cascade, similarly to what happens in mammalian eye photoreceptor cells (see the example in [14]. Looking at the genes from the phototransduction cascade expressed in the Opsin2 cells, the overall picture is relatively similar to the Opsin3.2 PRCs in terms of G-protein expressed and their expression levels, however the Opsin2 cells do not show any significant expression of the other putative genes involved in the phototransduction cascade.
Furthermore, we investigated the possibility that the Opsin3.2 protein might be involved in the entrainment of circadian clock (Figure 4B). The S. purpuratus genome encodes for several putative orthologues of the components of the canonical animal circadian clocks: e.g., Clock (Sp-Clock), Timeless (Sp-Tim), Brain and muscle Arnt-like protein (Sp-Bmal), Cryptochromes (Sp-Dcry and Sp-Vcry), Hepatic leukemia factor (Sp-Hlf) [31]. Interestingly, of the canonical core genes, the Opsin3.2 positive PRCs express only Sp-Clock and at low levels (Figure 4B). Nonetheless, they also express Sp-Shaggy which, in other species, is responsible for the phosphorylation of Tim, and therefore regulates its function. The Opsin3.2 PRCs also highly express Sp-Hlf, which was found to oscillate during the diel cycle (but not in free running condition) [41]. Finally, Sp-Sim, Sp-Slimb, Sp-Hey, and Sp-Nfil3 are expressed at low levels in the Opsin3.2 PRCs. On the contrary, the Opsin2 PRCs (Figure 4B) express only Sp-Hlf and Sp-Shaggy and at levels comparable to those found in the Opsin3.2 positive cells. In addition, they have low levels of Sp-6_4photolyase (photolyases are enzymes that repairs DNA damages caused by UV-light, [58]) and Sp-Ncoa.
Based on the evidence that the Opsin3.2 positive cells are neuron belonging to the Neuron (4) cluster, we investigated which type of neuronal genes are found in these PRCs taking advantage of the available comprehensive characterization of the larval nervous system [40] (Figure 4C). Notably, the Opsin3.2 cells express not only genes involved in photoreceptor specification (Sp-Otx, Sp-Six3, and Sp-Rx) but also genes involved in neuron/anterior neuroectoderm patterning (Sp-Nkx2.1, Sp-Hbn, Sp-Soxb1, Sp-SoxC and Sp-Brn1/2/4). Looking at the signaling molecules, the gene having the highest expression level is the Sp-Trh, neuropeptide, which is evidence for the double function of the Go-Opsin PRCs as both photoreceptor and neurosecretory cells. Moreover, it has been shown, both combining IHC and FISH and by scRNA-seq, that the TRHergic cells also produce another neuropeptide, Sp-Salmfap [56,59].
To conclude, we found also that both Opsin3.2 and Opsin3 cell populations express the TFs Sp-Otx and Sp-Id which are fundamental for general photoreceptor specification; while Sp-Rx, expressed in ciliary photoreceptors in many animals [18], is expressed only by the Opsin3.2 positive cells (Figure 4E).
4. Discussion
Main aim of this work was to characterize the Opsin3.2 photoreceptors found in S. purpuratus larvae. Nonetheless, during our analysis we encountered a second photoreceptor cell type known at this stage, the one expressing the Sp-Opsin2. Opsin2 PRCs have been already described not only in S. purpuratus [18], but also in other sea urchin species (Hemicentrotus pulcherrimus, S. Yaguchi et al., 2022) and showed to be a peculiar photoreceptor group, probably rising from a mesenchymal cell population. Indeed, our data also found a group of Opsin2 positive cells belonging to the Immune cell cluster. Comparison of the molecular signature of Opsin3.2 and Opsin2 PRCs showed many similarities in the TFs involved in their specification (both express Sp-Soxb1, Sp-Soxb2, Sp-Id, Sp-Otx, and Sp-Hbn), in the phototransduction cascade activated and in genes deployed for circadian rhythm establishment, with minor differences. Subsequently, despite the evidence that Opsin2 PRCs do not belong to a neuronal population, they still express typical neuronal markers. Major differences arose when we looked deeply into the molecular identity of the two PRCs in terms of neuropeptides, receptors and other TFs involved in neuronal specification. For example, Opsin2 do not express the Sp-Trh neuropeptide, and the Sp-Drd1 dopamine receptor. Nonetheless, further comparison of the differences between the two photoreceptor cell types was outside the scope of this work and we will not further discuss the data collected about Opsin2.
4.1. Opsin3.2 Cells Characterization during S. purpuratus Larval Development
The presence of two Opsin3.2 positive cells at the sides of the apical organ in the S. purpuratus sea urchin larva was already reported [18,35] but no information was available about their developmental origin. Therefore, first we combined immunohistochemistry and fluorescent in situ hybridization to identify the Opsin3.2 producing cells in larvae ranging from early 4-armed to late 8-armed pluteus stages (Figure 1). Interestingly, the number of Opsin3.2 cells appeared to increase during larval development in a non-stereotypical way, going from 1–2 cells at early pluteus to two clusters of up to 6 cells bilaterally distributed at either side of the apical organ, at the late pluteus stage (4–5 wpf) (Figure 1). Moreover, combined staining of Sp-Opsin3.2 and Sp-Trh provided the first evidence for a dual sensory/neurosecretory role of those cells, also displaying a bipolar morphology, with a cilium-like structure hosting the Go-Opsin molecules on one side and the axon transporting the TRH neuropeptide on the other side of the cell. Nonetheless, to prove that these cells are ciliary PRCs, the morphology of the ciliated structure requires further investigation using Electron Microscopy.
4.2. Opsin3.2 cells utilize an ancestral gene regulatory toolkit
Furthermore, we investigated the molecular fingerprint of the Sp-Opsin3.2 photoreceptors and showed that they utilize an ancestral regulatory toolkit. Namely, they express TFs required for neuron (Sp-SoxC and Sp-Brn1/2/4), and anterior neuroectoderm (Sp-Hbn, Sp-Six3) specification in sea urchins [60]. Moreover, the expression of Sp-Otx and Sp-Six3 supports the ancestral module suggested by [6] to be present in the precursor of all animal photoreceptors. According to this hypothesis, the ancestral photoreceptor cells utilized a variety of Opsin types (at least 9 in the Bilaterian ancestror [17]) and gave rise to the two sister cell types known as ciliary and rhabdomeric photoreceptors. Each of them also co-opted specific TFs such as Rx in ciliary photoreceptors. Intriguingly, Sp-Go-Opsin3.2 PRCs also express the TF Sp-Rx and have been described as belonging to the ciliary-type [18]. The hypothesis for the common evolutionary origin of ciliary and rhabdomeric photoreceptors is supported by the evidence that P. dumerilii possesses rhabdomeric photoreceptors expressing not only r-opsins but also Go-Opsin1 both in the adult eye and in the larval eyespot [22]. This latter opsin type, is most commonly associated with ciliary type PRCs (e.g., in scallop and sea urchins [18,21]. This hypothesis is also supported by the existence of a r-PRCs utilizing both xenopsin and r-opsin in the larval eye of the mollusk Leptochiton asellus [19]. However, whether Urbilateria, the common ancestor of Bilateria, already possessed both ciliary and rhabdomeric photoreceptors, or still had a single bimodal ciliary/rhabdomeric precursor cell is not yet clear and only analyzing the regulatory toolkit, the morphology and the opsin repertoire of photoreceptor cells in diverse early branching bilaterians animals will help disentangling this mystery.
4.3. Opsin3.2 Cells as Sensory and Neurosecretory Unit
In addition to their photoreceptor signature, Opsin3.2 PRCs are also able to produce a variety of signaling molecules, mostly neuropeptides (TRH, An, FSALFa/Salmfap, and Nesfatin), and acetylcholine [40,56]. Therefore, they look to be both sensory and neurosecretory cells.
It has been proposed that the Urbilateria already possessed a simple nervous system containing cells which were both sensory and neurosecretory. In animals having a more complex nervous system, the sensory signal collected by a specified sensory cell is transferred to the central nervous system through a series of interneurons. Here, the information is integrated with the signals received by other sensory cells and the final output is then transferred to an effector neuron which might be a motor neuron or endocrine cells for example. In less complex nervous systems, such as the case of Urbilateria, on the contrary, one cell can both collect the sensory stimulus and elicit the response, thus producing a very simple circuit or minimal module capable of controlling behavioral or physiological process [61,62]. Among these minimal modules, photosensitive-neuroendocrine cells could control neurohormones secretion under particular light conditions (Tessmar-Raible et al. 2007). The Opsin3.2 cells could be the descendants of such cells, retained in the sea urchin larva. Intriguingly, a cluster of non-visual photoreceptors expressing the vertebrate ancient long opsin b (valopb) was found in the brain of adult Danio rerio [63], suggesting an interesting evolutionary scenario for an ancestral TRHergic/photoreceptor cell cluster, which could be tested looking for the existence of TRHergic/Opsin positive neurons in other phyla.
4.4. Opsin3.2 Cells Possible Function(s)
Considering the sensory-neurosecretory nature of the Opsin3.2 cells, it is even more interesting to understand their function. Since these are photoreceptors, the first hypothesis is that they might be involved in controlling swimming behavior in response to light. In previous works, it has been suggested that light response activated by the Go-Opsin3.2 PRCs might be directional, namely that the larva is able to distinguish from which direction light hits the photoreceptor [18]. However, in all known cases of 3D phototaxis in the water column, the cells containing shading pigments are in close juxtaposition to the photosensitive cells (as reviewed by [64]. Sea urchin pigment cells are not fixed in their position and migrate in response to infections [65,66], therefore representing an unreliable source of shading.
Nonetheless, non-directional perception of light intensity can be used by aquatic animals to adjust their vertical position in the water column [22] and to avoid high UV levels [5,29,67]. Interestingly, Yaguchi and colleagues [37] showed that Hemicentrotus pulcherrimus and Temnopleurus reevesii larvae change their swimming behavior in response to strong photoirradiation. Although the authors seem to exclude an involvement for the Go-Opsin in this response, thorough experiments including knock-down or knock-out of this opsin exposed to different wavelengths are necessary to assess the role of these PRCs in light mediated swimming behaviors. Moreover, in a previous paper, Yaguchi and collaborators [36] provide evidence for Go-Opsin PRCs controlling sphincter opening in response to light through a serotonergic signaling in H. pulcherrimus larva and it would be interesting to test if this is a conserved function also in S. purpuratus larvae.
Intriguingly, the Opsin3.2 cells in our data are also predicted to express the dopamine receptor Sp-Drd1. Since a dopamine signaling is involved in mediating phenotypic (developmental) plasticity in response to food availability [68], we can hypothesize that the Sp-Opsin3.2 PRCs could control this process. Another possibility is that the light stimulus perceived by the Go-Opsin3.2 is used to entrain a circadian clock. To investigate this possibility, we looked at the genes putatively involved in this process (Figure 3D). Our scRNA-seq data do not show a prominent expression of putative core genes in the Sp-Opsin3.2 PRCs in the 3 and 5 dpf S. purpuratus larva, thus suggesting that the Opsin3.2 cells are involved in light perception more than circadian rhythms regulation.
While all the evidence collected so far contributes to build hypotheses on the function of the Opsin3.2/TRHergic cells, functional experiments to knock-down or knock-out Opsin3.2, coupled with phenotypic and behavioral observations, are necessary to comprehend the function of the Opsin3.2/TRHergic cells in sea urchin larvae.
5. Conclusions
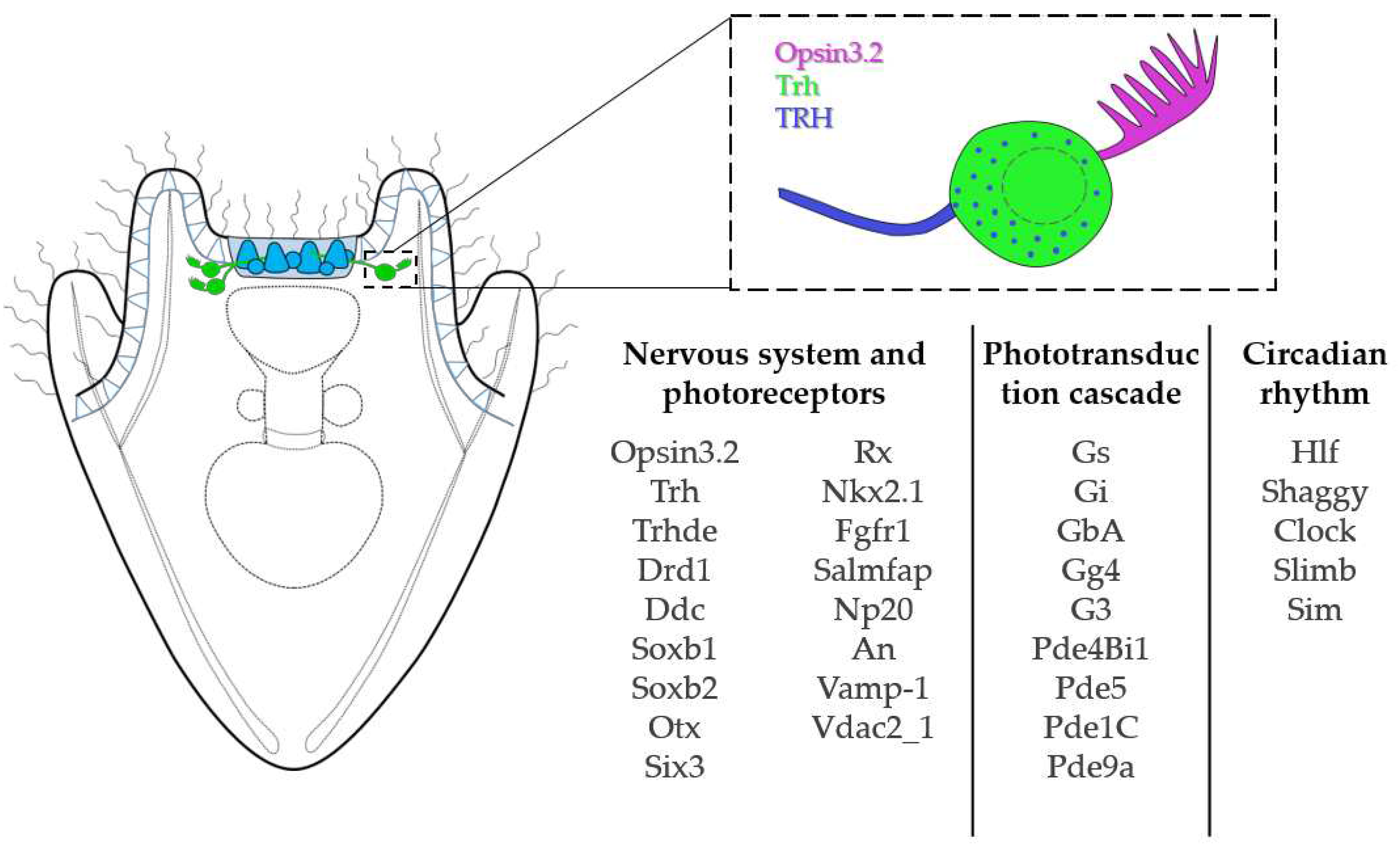
The sea urchin S. purpuratus larva feature a set of bilaterally symmetrical photoreceptor cells expressing the Go-Opsin3.2. This cell type deploys an ancient conserved regulatory module for photoreceptors specification. This makes them an important component to reconstruct photoreceptor/opsin system evolution, especially if combined with the analysis of photoreceptors in early branching animals. Additionally, the wide set of neuropeptides, signaling molecules and receptors expressed by these cells (including for example TRH, RX, Salmfap, An, Drd1) (Figure 5) strongly suggests their involvement in multiple processes. Nonetheless, knock-down and/or knock-out experiments are required to really understand in which process(es) the Opsin3.2 are involved.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. The raw reads for the sequencing data obtained for this study are available at NCBI Gene Expression Omnibus (accession numbers TBA). Supplementary Table S1: Primers used to clone the gene of interests.
Author Contributions
Conceptualization, M.I.A.; Investigation, M.C., P.P. and D.V.; Formal analysis, M.C., P.P., D.V., R.A. and M.I.A.; Software, P.P. and D.V.; Supervision, M.I.A.; Validation, P.P. and M.C.; Visualization, P.P. and M.C.; Writing original draft, M.C. and P.P.; Writing—review & editing, M.C., P.P., R.A., D.V. and M.I.A. All authors have read and agreed to the published version of the manuscript.
Funding
M.C. was supported by SZN PhD fellowships for the Open University and by Human Frontiers Science Program grant number (RGP0002/2019). P.P. was supported by Marie Curie ITN EvoCELL (H2020 Grant Number: 766053 to M.I.A) and by Human Frontiers Science Program grant number (RGP0002/2019).
Acknowledgments
The authors would like to thank Peter Halmay and Patrick Leahy (KML, Caltech, Pasadena, CA) for providing adult S. purpuratus and Davide Caramiello (SZN) for animal maintenance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Häfker, N.S.; Andreatta, G.; Manzotti, A.; Falciatore, A.; Raible, F.; Tessmar-Raible, K. Rhythms and Clocks in Marine Organisms. Annual Review of Marine Science 2023, 15, 509–538. [Google Scholar] [CrossRef] [PubMed]
- Vitaterna, M.H.; Takahashi, J.S.; Turek, F.W. Overview of Circadian Rhythms. Alcohol Res Health 2001, 25, 85–93. [Google Scholar] [PubMed]
- Dawson, A.; King, V.M.; Bentley, G.E.; Ball, G.F. Photoperiodic Control of Seasonality in Birds. Journal of Biological Rhythms 2001, 16, 365–380. [Google Scholar] [CrossRef]
- DeValois, R.L.; DeValois, K.K. Spatial Vision; Academic Press: Cambridge, MA, USA, 2008; ISBN 9780199872220. [Google Scholar]
- Verasztó, C.; Gühmann, M.; Jia, H.; Rajan, V.B.V.; Bezares-Calderón, L.A.; Piñeiro-Lopez, C.; Randel, N.; Shahidi, R.; Michiels, N.K.; Yokoyama, S.; et al. Ciliary and Rhabdomeric Photoreceptor-Cell Circuits Form a Spectral Depth Gauge in Marine Zooplankton. eLife 2018, 7, 1–19. [Google Scholar] [CrossRef]
- Arendt, D. Evolution of Eyes and Photoreceptor Cell Types. International Journal of Developmental Biology 2003, 47, 563–571. [Google Scholar]
- Arendt, D.; Wittbrodt, J. Reconstructing the Eyes of Urbilateria. Philosophical Transactions of the Royal Society B: Biological Sciences 2001, 356, 1545–1563. [Google Scholar] [CrossRef] [PubMed]
- Hargrave, P.A.; McDowell, J.H.; Curtis, D.R.; Wang, J.K.; Juszczak, E.; Fong, S.L.; Mohana Rao, J.K.; Argos, P. The Structure of Bovine Rhodopsin. Biophysics of Structure and Mechanism 1983, 9, 235–244. [Google Scholar] [CrossRef]
- Terakita, A. The Opsins. Genome Biology 2005, 6. [Google Scholar] [CrossRef]
- Schertler, G.F.X.; Villa, C.; Henderson, R. Projection Structure of Rhodopsin. Nature 1993, 362, 770–772. [Google Scholar] [CrossRef]
- D’Aniello, S.; Delroisse, J.; Valero-Gracia, A.; Lowe, E.K.; Byrne, M.; Cannon, J.T.; Halanych, K.M.; Elphick, M.R.; Mallefet, J.; Kaul-Strehlow, S.; et al. Opsin Evolution in the Ambulacraria. Marine Genomics 2015, 24, 177–183. [Google Scholar] [CrossRef]
- Peirson, S.N.; Halford, S.; Foster, R.G. The Evolution of Irradiance Detection: Melanopsin and the Non-Visual Opsins. Philosophical Transactions of the Royal Society B: Biological Sciences 2009, 364, 2849–2865. [Google Scholar] [CrossRef]
- Porter, M.L.; Blasic, J.R.; Bok, M.J.; Cameron, E.G.; Pringle, T.; Cronin, T.W.; Robinson, P.R. Shedding New Light on Opsin Evolution. Proceedings of the Royal Society B: Biological Sciences 2011, 279, 3–14. [Google Scholar] [CrossRef]
- Shichida, Y.; Matsuyama, T. Evolution of Opsins and Phototransduction. Philosophical Transactions of the Royal Society B: Biological Sciences 2009, 364, 2881–2895. [Google Scholar] [CrossRef]
- Arendt, D. The Enigmatic Xenopsins. eLife 2017, 6. [Google Scholar] [CrossRef]
- Delroisse, J.; Mallefet, J.; Flammang, P. De Novo Adult Transcriptomes of Two European Brittle Stars: Spotlight on Opsin-Based Photoreception. PLoS ONE 2016, 11, e0152988. [Google Scholar] [CrossRef]
- Ramirez, M.D.; Pairett, A.N.; Pankey, M.S.; Serb, J.M.; Speiser, D.I.; Swafford, A.J.; Oakley, T.H. The Last Common Ancestor of Most Bilaterian Animals Possessed at Least Nine Opsins. Genome Biology and Evolution 2016, 8, 3640–3652. [Google Scholar] [CrossRef] [PubMed]
- Valencia, J.E.; Feuda, R.; Mellott, D.O.; Burke, R.D.; Peter, I.S. Ciliary Photoreceptors in Sea Urchin Larvae Indicate Pan-Deuterostome Cell Type Conservation. BMC Biology 2021, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Vöcking, O.; Kourtesis, I.; Tumu, S.C.; Hausen, H. Co-Expression of Xenopsin and Rhabdomeric Opsin in Photoreceptors Bearing Microvilli and Cilia. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Shichida, Y.; Imai, H. Visual Pigment: G-Protein-Coupled Receptor for Light Signals. Cellular and Molecular Life Sciences 1998, 54, 1299–1315. [Google Scholar] [CrossRef]
- Kojima, D.; Terakita, A.; Ishikawa, T.; Tsukahara, Y.; Maeda, A.; Shichida, Y. A Novel G(o)-Mediated Phototransduction Cascade in Scallop Visual Cells. Journal of Biological Chemistry 1997, 272, 22979–22982. [Google Scholar] [CrossRef]
- Gühmann, M.; Jia, H.; Randel, N.; Verasztó, C.; Bezares-Calderón, L.A.; Michiels, N.K.; Yokoyama, S.; Jékely, G. Spectral Tuning of Phototaxis by a Go-Opsin in the Rhabdomeric Eyes of Platynereis. Current Biology 2015, 25, 2265–2271. [Google Scholar] [CrossRef] [PubMed]
- Arendt, D.; Tessmar-Raible, K.; Snyman, H.; Dorresteijn, A.W.; Wittbrodf, J. Ciliary Photoreceptors with a Vertebrate-Type Opsin in an Invertebrate Brain. Science 2004, 306, 869–871. [Google Scholar] [CrossRef]
- Kozmik, Z.; Ruzickova, J.; Jonasova, K.; Matsumoto, Y.; Vopalensky, P.; Kozmikova, I.; Strnad, H.; Kawamura, S.; Piatigorsky, J.; Paces, V.; et al. Assembly of the Cnidarian Camera-Type Eye from Vertebrate-like Components. Proceedings of the National Academy of Sciences of the United States of America 2008, 105, 8989–8993. [Google Scholar] [CrossRef]
- Pearse, J.S.; Pearse, V.B. Vision in Cubomedusan Jellyfishes. Science 1978, 199, 458. [Google Scholar] [CrossRef] [PubMed]
- Braun, K.; Kaul-Strehlow, S.; Ullrich-Lüter, E.; Stach, T. Structure and Ultrastructure of Eyes of Tornaria Larvae of Glossobalanus Marginatus. Organisms Diversity and Evolution 2015, 15, 423–428. [Google Scholar] [CrossRef]
- Ullrich-Luter, E.M.; Dupont, S.; Arboleda, E.; Hausen, H.; Arnone, M.I. Unique System of Photoreceptors in Sea Urchin Tube Feet. Proceedings of the National Academy of Sciences 2011, 108, 8367–8372. [Google Scholar] [CrossRef]
- Vopalensky, P.; Pergner, J.; Liegertova, M.; Benito-Gutierrez, E.; Arendt, D.; Kozmik, Z. Molecular Analysis of the Amphioxus Frontal Eye Unravels the Evolutionary Origin of the Retina and Pigment Cells of the Vertebrate Eye. Proceedings of the National Academy of Sciences of the United States of America 2012, 109, 15383–15388. [Google Scholar] [CrossRef]
- Nilsson, D.E. Eye Evolution and Its Functional Basis. Visual Neuroscience 2013, 30, 5–20. [Google Scholar] [CrossRef]
- Fain, G.L.; Hardie, R.; Laughlin, S.B. Phototransduction and the Evolution of Photoreceptors. Current Biology 2010, 20, R114–R124. [Google Scholar] [CrossRef]
- Burke, R.; Angerer, L.; Elphick, M.; Humphrey, G.; Yaguchi, S.; Kiyama, T.; Liang, S.; Mu, X.; Agca, C.; Klein, W.; et al. A Genomic View of the Sea Urchin Nervous System. Developmental Biology 2006, 300, 434–460. [Google Scholar] [CrossRef]
- Raible, F.; Tessmar-Raible, K.; Arboleda, E.; Kaller, T.; Bork, P.; Arendt, D.; Arnone, M.I. Opsins and Clusters of Sensory G-Protein-Coupled Receptors in the Sea Urchin Genome. Developmental Biology 2006, 300, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Paganos, P.; Ullrich-Lüter, E.; Caccavale, F.; Zakrzewski, A.; Voronov, D.; Fournon-Berodia, I.; Cocurullo, M.; Lüter, C.; Arnone, M.I. A New Model Organism to Investigate Extraocular Photoreception: Opsin and Retinal Gene Expression in the Sea Urchin Paracentrotus Lividus. Cells 2022, 11, 2636. [Google Scholar] [CrossRef] [PubMed]
- Ullrich-Lüter, E.M.; D’Aniello, S.; Arnone, M.I. C-Opsin Expressing Photoreceptors in Echinoderms. Integrative and Comparative Biology 2013, 53, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Valero-Gracia, A.; Petrone, L.; Oliveri, P.; Nilsson, D.-E.; Arnone, M.I. Non-Directional Photoreceptors in the Pluteus of Strongylocentrotus Purpuratus. Frontiers in Ecology and Evolution 2016, 4, 127. [Google Scholar] [CrossRef]
- Yaguchi, J.; Yaguchi, S. Sea Urchin Larvae Utilize Light for Regulating the Pyloric Opening. BMC Biology 2021, 19, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yaguchi, S.; Taniguchi, Y.; Suzuki, H.; Kamata, M.; Yaguchi, J. Planktonic Sea Urchin Larvae Change Their Swimming Direction in Response to Strong Photoirradiation. PLOS Genetics 2022, 18, e1010033. [Google Scholar] [CrossRef]
- Foster, S.; Oulhen, N.; Wessel, G. A Single Cell RNA Sequencing Resource for Early Sea Urchin Development. Development (Cambridge) 2020, 147. [Google Scholar] [CrossRef]
- Massri, A.J.; Greenstreet, L.; Afanassiev, A.; Berrio, A.; Wray, G.A.; Schiebinger, G.; McClay, D.R. Developmental Single-Cell Transcriptomics in the Lytechinus Variegatus Sea Urchin Embryo. Development (Cambridge) 2021, 148. [Google Scholar] [CrossRef]
- Paganos, P.; Voronov, D.; Musser, J.; Arendt, D.; Arnone, M.I. Single Cell Rna Sequencing of the Strongylocentrotus Purpuratus Larva Reveals the Blueprint of Major Cell Types and Nervous System of a Nonchordate Deuterostome. eLife 2021, 10, 1–29. [Google Scholar] [CrossRef]
- Petrone, L. Circadian Clock and Light Input System in the Sea Urchin Larva; UCL (University College London), 2015. [Google Scholar]
- Paganos, P.; Caccavale, F.; La Vecchia, C.; D’Aniello, E.; D’Aniello, S.; Arnone, M.I. FISH for All: A Fast and Efficient Fluorescent In Situ Hybridization (FISH) Protocol for Marine Embryos and Larvae. Frontiers in Physiology 2022, 13. [Google Scholar] [CrossRef]
- Perillo, M.; Paganos, P.; Spurrell, M.; Arnone, M.I.; Wessel, G.M. Methodology for Whole Mount and Fluorescent RNA In Situ Hybridization in Echinoderms: Single, Double, and Beyond. In Methods in Molecular Biology; Humana Press Inc., 2021; Volume 2219, pp. 195–216. [Google Scholar]
- Conzelmann, M.; Jékely, G. Antibodies against Conserved Amidated Neuropeptide Epitopes Enrich the Comparative Neurobiology Toolbox. EvoDevo 2012, 3, 23. [Google Scholar] [CrossRef] [PubMed]
- Wood, N.J. Investigating the Roles of Neuropeptides in the Development of the Sea Urchin, Strongylocentrotus Purpuratus. Doctoral, UCL (University College London), 2020; pp. 1–349. [Google Scholar]
- Sodergren, E.; Weinstock, G.M.; Davidson, E.H.; Cameron, R.A.; Gibbs, R.A.; Angerer, R.C.; Angerer, L.M.; Arnone, M.I.; Burgess, D.R.; Burke, R.D.; et al. The Genome of the Sea Urchin Strongylocentrotus Purpuratus. Science 2006, 314, 941–952. [Google Scholar] [CrossRef] [PubMed]
- Kudtarkar, P.; Cameron, R.A. Echinobase: An Expanding Resource for Echinoderm Genomic Information. Database 2017, 2017, 74. [Google Scholar] [CrossRef] [PubMed]
- Stuart, T.; Butler, A.; Hoffman, P.; Hafemeister, C.; Papalexi, E.; Mauck, W.M.; Hao, Y.; Stoeckius, M.; Smibert, P.; Satija, R. Comprehensive Integration of Single-Cell Data. Cell 2019, 177, 1888–1902.e21. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating Single-Cell Transcriptomic Data across Different Conditions, Technologies, and Species. Nat Biotechnol 2018, 36, 411–420. [Google Scholar] [CrossRef]
- Paganos, P.; Ronchi, P.; Carl, J.; Mizzon, G.; Martinez, P.; Benvenuto, G.; Arnone, M.I. Integrating Single Cell Transcriptomics and Volume Electron Microscopy Confirms the Presence of Pancreatic Acinar-like Cells in Sea Urchins. Front. Cell Dev. Biol. 2022, 10, 991664. [Google Scholar] [CrossRef]
- Korsunsky, I.; Millard, N.; Fan, J.; Slowikowski, K.; Zhang, F.; Wei, K.; Baglaenko, Y.; Brenner, M.; Loh, P.; Raychaudhuri, S. Fast, Sensitive and Accurate Integration of Single-Cell Data with Harmony. Nat Methods 2019, 16, 1289–1296. [Google Scholar] [CrossRef]
- Byrne, M.; Nakajima, Y.; Chee, F.C.; Burke, R.D. Apical Organs in Echinoderm Larvae: Insights into Larval Evolution in the Ambulacraria. Evolution and Development 2007, 9, 432–445. [Google Scholar] [CrossRef]
- Hadfield, M.; Meleshkevitch, E.; Boudko, D. The Apical Sensory Organ of a Gastropod Veliger Is a Receptor for Settlement Cues. The Biological Bulletin 2016, 198, 67–76. [Google Scholar] [CrossRef]
- Sinigaglia, C.; Busengdal, H.; Lerner, A.; Oliveri, P.; Rentzsch, F. Molecular Characterization of the Apical Organ of the Anthozoan Nematostella Vectensis. Developmental Biology 2015, 398, 120–133. [Google Scholar] [CrossRef]
- Bisgrove, B.W.; Burke, R.D. Development of Serotonergic Neurons in Embryos of the Sea Urchin, Strongylocentrotus Purpuratus. Development Growth and Differentiation 1986, 28, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Wood, N.J.; Mattiello, T.; Rowe, M.L.; Ward, L.; Perillo, M.; Arnone, M.I.; Elphick, M.R.; Oliveri, P. Neuropeptidergic Systems in Pluteus Larvae of the Sea Urchin Strongylocentrotus Purpuratus: Neurochemical Complexity in a “Simple” Nervous System-Supplementary. Frontiers in Endocrinology 2018, 9, 628. [Google Scholar] [CrossRef] [PubMed]
- Cocurullo, M.; Paganos, P.; Wood, N.J.; Arnone, M.I.; Oliveri, P. Molecular and Cellular Characterization of the TH Pathway in the Sea Urchin Strongylocentrotus Purpuratus. Cells 2023, 12, 272. [Google Scholar] [CrossRef]
- Li, J.; Liu, Z.; Tan, C.; Guo, X.; Wang, L.; Sancar, A.; Zhong, D. Dynamics and Mechanism of Repair of Ultraviolet-Induced (6-4) Photoproduct by Photolyase. Nature 2010, 466, 887–890. [Google Scholar] [CrossRef] [PubMed]
- Paganos, P.; Voronov, D.; Musser, J.; Arendt, D.; Arnone, M.I. Single Cell Rna Sequencing of the Strongylocentrotus Purpuratus Larva Reveals the Blueprint of Major Cell Types and Nervous System of a Nonchordate Deuterostome. eLife 2021, 10, 1–29. [Google Scholar] [CrossRef]
- Angerer, L.M.; Yaguchi, S.; Angerer, R.C.; Burke, R.D. The Evolution of Nervous System Patterning: Insights from Sea Urchin Development. Development 2011, 138, 3613–3623. [Google Scholar] [CrossRef]
- Hartenstein, V. The Neuroendocrine System of Invertebrates: A Developmental and Evolutionary Perspective. Journal of Endocrinology 2006, 190, 555–570. [Google Scholar] [CrossRef]
- Tessmar-Raible, K.; Raible, F.; Christodoulou, F.; Guy, K.; Rembold, M.; Hausen, H.; Arendt, D. Conserved Sensory-Neurosecretory Cell Types in Annelid and Fish Forebrain: Insights into Hypothalamus Evolution. Cell 2007, 129, 1389–1400. [Google Scholar] [CrossRef]
- Hang, C.Y.; Kitahashi, T.; Parhar, I.S. Localization and Characterization of Val-Opsin Isoform-Expressing Cells in the Brain of Adult Zebrafish. Journal of Comparative Neurology 2014, 522, 3847–3860. [Google Scholar] [CrossRef]
- Jékely, G.; Colombelli, J.; Hausen, H.; Guy, K.; Stelzer, E.; Nédélec, F.; Arendt, D. Mechanism of Phototaxis in Marine Zooplankton. Nature 2008, 456, 395–399. [Google Scholar] [CrossRef]
- Buckley, K.M.; Schuh, N.W.; Heyland, A.; Rast, J.P. Analysis of Immune Response in the Sea Urchin Larva. In Methods in Cell Biology; Academic Press: Cambridge, MA, USA, 2019; Volume 150, pp. 333–355. [Google Scholar]
- Buckley, K.M.; Rast, J.P. Immune Activity at the Gut Epithelium in the Larval Sea Urchin. Cell and Tissue Research 2019, 377, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Leech, D.; Johnsen, S. Behavioral Responses–UVR Avoidance and Vision. In UV Effects in Aquatic Organisms and Ecosystems; Helbling, W., Z.H., Eds.; Royal Society of Chemistry, 2003; pp. 457–481. [Google Scholar]
- Adams, D.K.; Sewell, M.A.; Angerer, R.C.; Angerer, L.M. Rapid Adaptation to Food Availability by a Dopamine-Mediated Morphogenetic Response. Nature Communications 2011, 2, 592. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Molecular characterization of the Opsin3.2 cells at different developmental stages. (A) FISH of Sp-Opsin3.2 stains 1 cell at 3 dpf. (B) FISH for Sp-Trh was combined with double TRH/Opsin3.2 immunolocalization at 3 dpf, highlighting the bipolar structure of these sensory/neurosecretory neurons with the Opsin3.2 localized on one a ciliated-like structure located on the external side of the larva. The TRH peptide, instead, is concentrated in the opposite side of the cell and it appears to be transported along the projections of the cells (directed toward the larva apical organ). (B′) Details of the cell showed in B on the left. (C) FISH of Sp-Opsin3.2 at 4 dpf. (D,D′) FISH for Sp-Trh was combined with double TRH/Opsin3.2 immunolocalization at 4 dpf. All the 3 cells have a bipolar organization. (E,F) Double FISH of Sp-Opsin3.2 and Sp-Trh at 5 dpf stains 3–4 cells. FISH of Sp-Opsin3.2 at 4 wpf (G,G′) and 5 wpf (H–H″) detect two cluster of cells located at the base of the oral arms. All images are stacks of merged confocal Z sections. Nuclei are shown in white.
Figure 1.
Molecular characterization of the Opsin3.2 cells at different developmental stages. (A) FISH of Sp-Opsin3.2 stains 1 cell at 3 dpf. (B) FISH for Sp-Trh was combined with double TRH/Opsin3.2 immunolocalization at 3 dpf, highlighting the bipolar structure of these sensory/neurosecretory neurons with the Opsin3.2 localized on one a ciliated-like structure located on the external side of the larva. The TRH peptide, instead, is concentrated in the opposite side of the cell and it appears to be transported along the projections of the cells (directed toward the larva apical organ). (B′) Details of the cell showed in B on the left. (C) FISH of Sp-Opsin3.2 at 4 dpf. (D,D′) FISH for Sp-Trh was combined with double TRH/Opsin3.2 immunolocalization at 4 dpf. All the 3 cells have a bipolar organization. (E,F) Double FISH of Sp-Opsin3.2 and Sp-Trh at 5 dpf stains 3–4 cells. FISH of Sp-Opsin3.2 at 4 wpf (G,G′) and 5 wpf (H–H″) detect two cluster of cells located at the base of the oral arms. All images are stacks of merged confocal Z sections. Nuclei are shown in white.
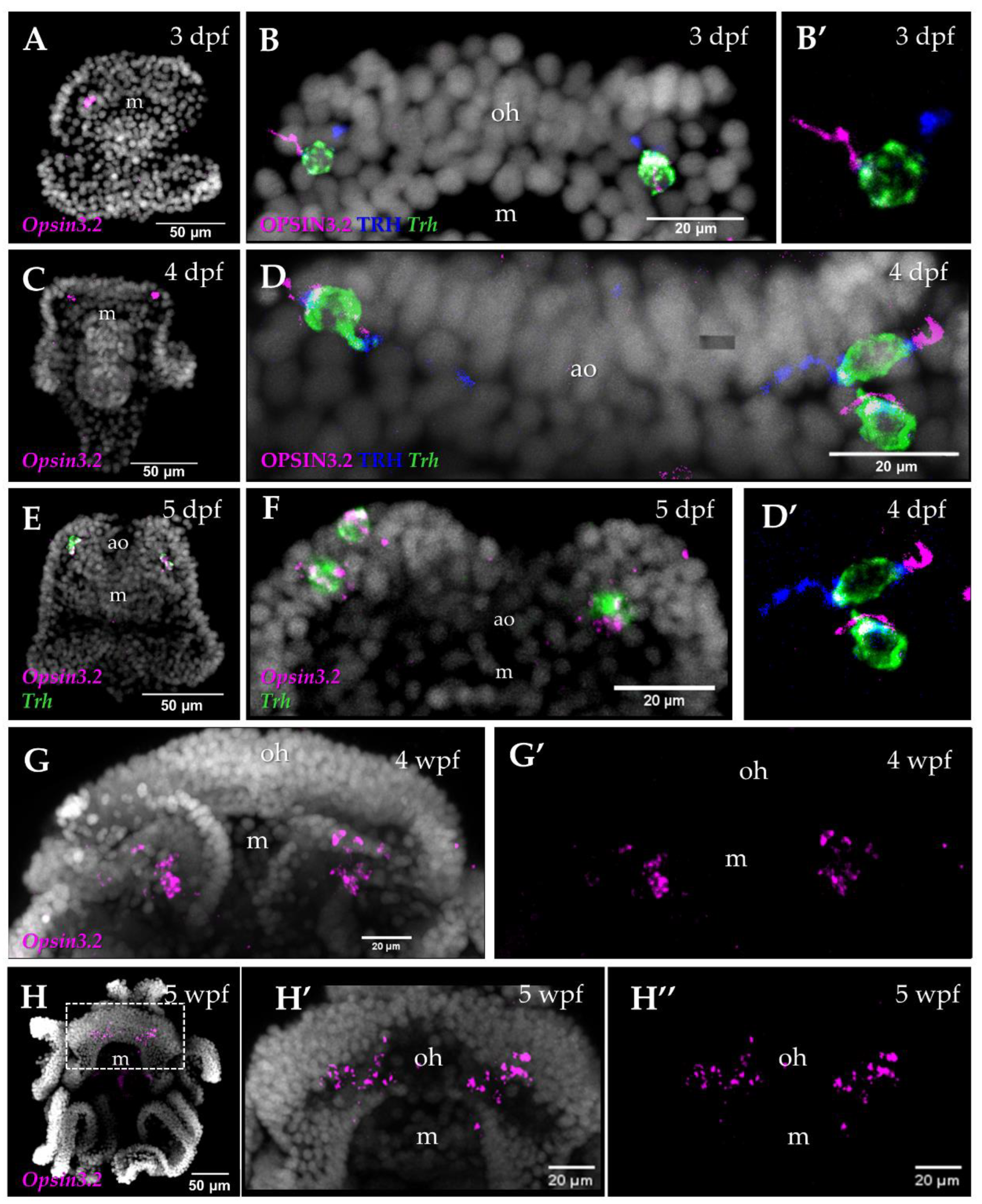
Figure 2.
Cell type atlas of the S. purpuratus larvae at 3 and 5 dpf pluteus stages. (A) Schematic representation of the scRNA-seq pipeline. (B) UMAP representing the cell clustering obtained through harmony of the 3 and 5 dpf pluteus single cell datasets and their localization in a schematic representation of a 4-armed S. purpuratus larva. (C) Dotplot showing the average expression of a subset of genes used as markers to annotate the different cell clusters.
Figure 2.
Cell type atlas of the S. purpuratus larvae at 3 and 5 dpf pluteus stages. (A) Schematic representation of the scRNA-seq pipeline. (B) UMAP representing the cell clustering obtained through harmony of the 3 and 5 dpf pluteus single cell datasets and their localization in a schematic representation of a 4-armed S. purpuratus larva. (C) Dotplot showing the average expression of a subset of genes used as markers to annotate the different cell clusters.
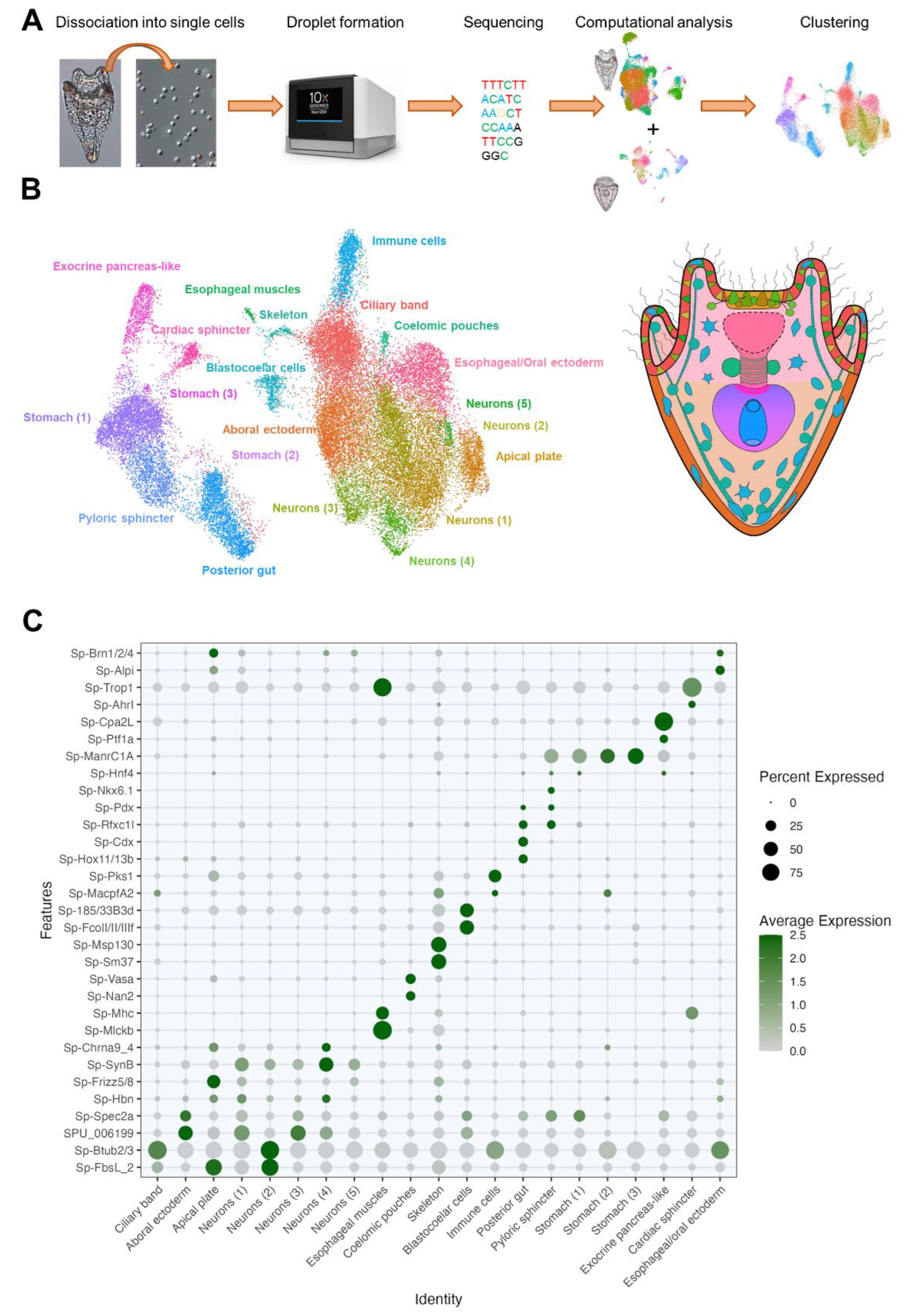
Figure 3.
Identification of the Opsin3.2 and Opsin2 PRCs. (A) Dotplot showing the expression of Sp-Osin3.2 and Sp-Osin2 and of the neuropeptide Sp-Trh and of the Immune cells marker Sp-Pks1. (B) UMAP illustrating that Opsin3.2 PRCs have been isolated from the Neuron (4) cluster while Opsin2 have been selected from the Immune cells cluster. (C) Dotplot showing the expression pattern of genes selected from Valencia et al., 2021.
Figure 3.
Identification of the Opsin3.2 and Opsin2 PRCs. (A) Dotplot showing the expression of Sp-Osin3.2 and Sp-Osin2 and of the neuropeptide Sp-Trh and of the Immune cells marker Sp-Pks1. (B) UMAP illustrating that Opsin3.2 PRCs have been isolated from the Neuron (4) cluster while Opsin2 have been selected from the Immune cells cluster. (C) Dotplot showing the expression pattern of genes selected from Valencia et al., 2021.
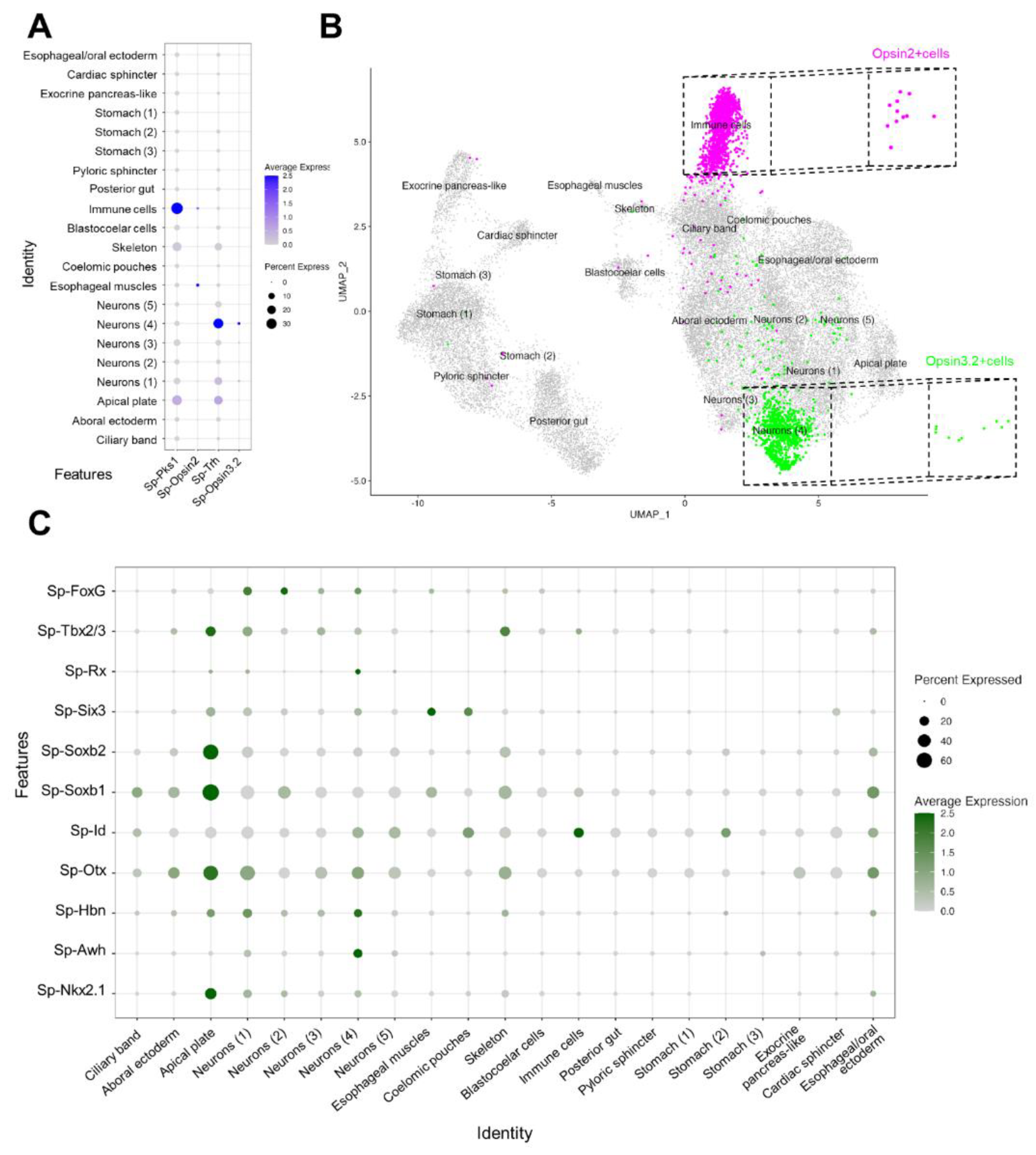
Figure 4.
Molecular signature of S. purpuratus PRCs at pluteus stages. (A) Dotplot showing the average expression of genes putatively involved in the phototransuction cascade activated by the opsins stimulation. (B) Dotplot showing the average expression of genes involved in the regulation circadian rhythms. (C) Dotplot showing the average expression of genes selected to further describe the molecular identity of the Opsin3.2 and Opsin2 PRCs. (D) Dotplot showing the percentage of TFs common to the two PRCs. (E) Dotplot showing the average expression of known TFs involved photoreceptor specification.
Figure 4.
Molecular signature of S. purpuratus PRCs at pluteus stages. (A) Dotplot showing the average expression of genes putatively involved in the phototransuction cascade activated by the opsins stimulation. (B) Dotplot showing the average expression of genes involved in the regulation circadian rhythms. (C) Dotplot showing the average expression of genes selected to further describe the molecular identity of the Opsin3.2 and Opsin2 PRCs. (D) Dotplot showing the percentage of TFs common to the two PRCs. (E) Dotplot showing the average expression of known TFs involved photoreceptor specification.
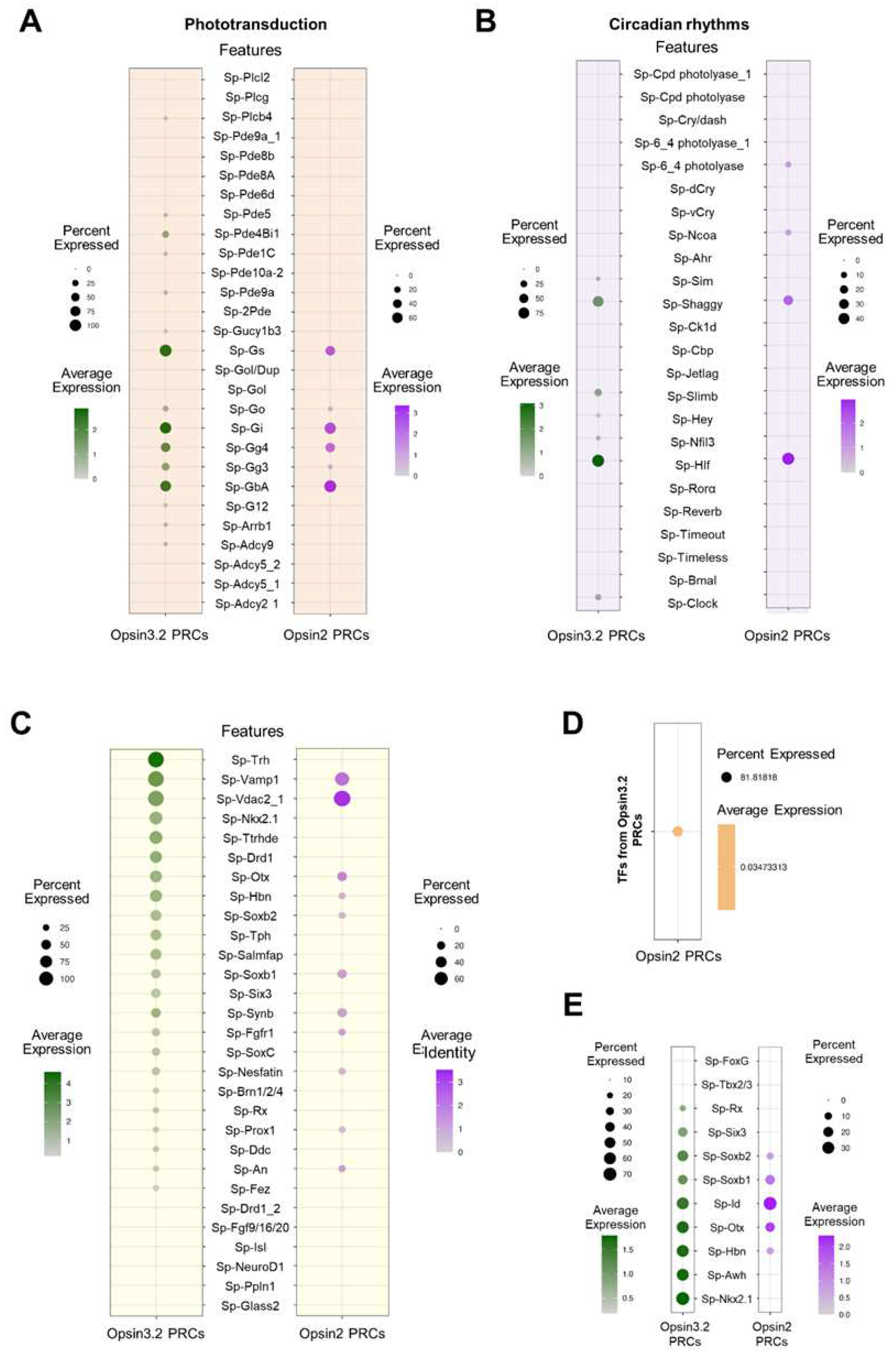
Figure 5.
Opsin3.2 PRCs summary. Schematic reconstruction of the morphology, topology and molecular signature of the Opsin3.2 PRCs.
Figure 5.
Opsin3.2 PRCs summary. Schematic reconstruction of the morphology, topology and molecular signature of the Opsin3.2 PRCs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



