
Submitted:
05 February 2025
Posted:
06 February 2025
You are already at the latest version
Abstract
This study investigates the potential of incorporating Bergamot juice and olive extracts as functional ingredients in craft beer production, focusing on their effects on key biochemi-cal processes, including cell viability, reactive oxygen species (ROS) levels, and the regula-tion of RNA expression of antioxidant enzymes. The research aims to explore how these botanicals can enhance the health-related qualities of beer, transforming it from a mere beverage into a functional food that supports wellness. The effects of lyophilized beer samples on cellular ROS levels were assessed by treating fibroblast cells with 0.31 and 0.62 µg/µl concentrations of various beer samples for 24 and 48 hours. While basal beers (Blanche and Weiss) did not show significant changes in ROS levels, the addition of Ber-gamot juice in Heraclea and olive extract in Elais resulted in a significant decrease in ROS levels at both incubation times, indicating the potent antioxidant properties of these beers. At higher concentrations (0.62 µg/µl), a decrease in ROS was observed in Weiss beer-treated cells after 48 hours, while no such effect was noted in other beer samples treatment, possibly due to pro-oxidant behavior exhibited under certain conditions. Addi-tionally, gene expression analysis revealed that exposure to oxidative stress induced by t-BHP led to a significant upregulation of SOD2 expression in all beer-treated groups, ex-cept for Elais, suggesting an adaptive cellular response to oxidative damage. These find-ings underscore the complex role of phenolic compounds and flavonoids in modulating oxidative stress and enhancing cellular health. Overall, this study highlights the potential of functional craft beers, enriched with Bergamot and olive extracts, as innovative health-conscious options that provide both flavor and antioxidant benefits, paving the way for the future of wellness-focused brewing.
Keywords:
functional craft beer
; antioxidant properties
; bergamot juice
; olive extract
; oxidative stress
1. Introduction
Beer, one of the oldest and most widely consumed alcoholic beverages globally, has evolved significantly in terms of ingredients and production methods. Historically, beer was crafted using simple ingredients: water, malted barley, hops, and yeast. In recent years, health concerns and a general desire for a healthier lifestyle have led to an increased demand for functional beers, and craft beer production has flourished with an increased emphasis on innovation and experimentation [1]. Modern brewers have explored the use of various botanicals and extracts to enhance the flavor, aroma, and content of bioactive compounds with functional properties in beer, creating new sensory experiences for consumers [2,3]. Among these innovations, incorporating plant-based extracts has received considerable attention due to their potential to enrich beer with unique sensory profiles such as bergamot (Citrus bergamia) and olive (Olea Europaea) [4]. Bergamot, a citrus fruit native to the coastal regions of southern Italy, is best known for its distinctive fragrance. In the past, Bergamot was mainly harvested for its essential oil, while by-products such as juice, pulp, and seeds were considered as industrial waste. However, in recent years, a growing body of research has highlighted the significant nutraceutical potential of these overlooked parts of the fruit [5]. Studies have demonstrated that Bergamot when consumed fresh or as a juice, offers an impressive array of bioactive compounds that contribute to its health benefits. Unlike other fruits of the same genus, Bergamot is particularly rich in a diverse range of phytochemicals and nutraceuticals, including organic acids, limonoids, phenolic acids, and flavonoids. These compounds have been shown to exhibit antioxidant, anti-inflammatory, and antimicrobial properties, suggesting that Bergamot may offer various therapeutic benefits when incorporated into human diets [6,7,8]. Key compounds such as bergapten, flavonoids (e.g., rutin, quercetin), and coumarins have been shown to exhibit significant biological activities, including the modulation of oxidative stress and the inhibition of pro-inflammatory pathways. These bioactive compounds are thought to provide numerous health benefits, including protecting cells from oxidative damage, support for cardiovascular health and potential anticancer effects [9]. The addition of Bergamot extract into beer could offer a novel way to combine the antioxidant potential of the fruit with the functional properties of beer, promoting health-conscious consumption The bio compounds and the essential oils from bergamot cultivated in the province of Reggio Calabria, are of exceptional quality compared to those derived from bergamot cultivated outside this area. The superior quality of Reggio Calabria's Bergamot is largely attributed to the unique microclimate and soil conditions, which provide an ideal growing environment and play a critical role in determining its distinctive qualitative characteristics [10]. Recent studies have confirmed that these environmental factors are key in influencing the chemical composition and therapeutic potential of Bergamot. Similarly, olive extract, derived from the fruit of the olive tree, is well- documented for its wealth of polyphenolic compounds such as oleuropein, hydroxytyrosol, and oleocanthal [11,12]. These compounds have potent antioxidant, anti-inflammatory, and antimicrobial properties, that have been associated with a variety of health benefits, particularly in the context of the Mediterranean diet. Olive extracts are believed to support heart health by improving lipid profiles, reducing blood pressure, and protecting against oxidative stress, while also exhibiting antimicrobial properties that could enhance the safety and preservation of foods and beverages [13]. The addition of olive extract to beer may not only enhance its organoleptic characteristics but also contribute to its functional properties by providing these bioactive benefits. As reported by De Bruno et al (2019), the olive of the cultivar Carolea contained more total phenolic compounds and had higher antioxidant activity expressed as DPPH than other Calabrian cultivars. The superior quality of the cultivar Carolea can be attributed to the ideal growing environment that plays a critical role in determining its distinctive qualitative characteristics [14].
This manuscript aims to explore the use of Bergamot juice extracts and olive extracts as functional ingredients in craft beer production. We will investigate their impact on key biochemical processes including cell viability, reactive oxygen species (ROS) levels, and the regulation of RNA expression related to antioxidant enzymes. By evaluating the effects of these extracts on these molecular pathways, we aim to understand how these botanicals can potentially enhance the health-related qualities of beer, making it not just a beverage but a functional food that supports wellness. Furthermore, we will discuss how the integration of such extracts in brewing can shape the future of craft beer, providing an innovative and health-conscious option for consumers seeking both flavour and potential health benefits.
2. Results and Discussion
2.1. Effect of Antioxidant Compounds
As reported by Muscolo et al [4], Heraclea and Elais both had lower protein content (0.2 g/100ml), than basal Weiss and Blanche (Figure 1). Ideally, beer protein content should approach zero, as proteins can bind to polysaccharides, forming insoluble complexes that lead to turbidity and compromise the stability of the beverage. Total carbohydrate levels were slightly lower in basal Weiss and Blanche than in Heraclea and Elais (Figure 1) but remained within the recommended range of 3.3–4.4 g/100ml. Heraclea and Elais had higher levels of Vitamin C, 20 times higher than those found in Basal Weiss and Blanche. Vitamin C is a powerful antioxidant that protects tissues, lipids and proteins from oxidative damage and plays a vital role in immune system regulation, cell growth, and differentiation. The DPPH and TAC activities were significantly higher in Heraclea and Elais beers, reflecting the greater concentration of antioxidant compounds in these beverages (Figure 1). In terms of bioactive compounds, Elais had the highest total polyphenols, followed by Heraclea. Flavonoids followed a similar trend to polyphenols. The phenolic profiles of the beers analyzed revealed differences among the basal and bergamot and olive added beers (Figure 2).
Coumaric acid although less representative than the other phenolic acids in all the beers, was present in the highest amount in Elais followed by Heraclea. Chlorogenic acid was also mostly present in Elais and Heraclea, syringic and ethyl gallate acids were significantly more abundant in Elais than in Heraclea and basal beers, while the protocatechuic acid was more concentrated in Heraclea than in the other beers. Additionally, p-coumaric acid, a phenolic compound found in various plant sources such as bergamot, has been associated with several health benefits. It is known for its strong antioxidant activity, which may help protect cells from oxidative stress [15]. p-Coumaric acid has been shown to possess anti-inflammatory properties, which may help reduce chronic inflammation linked to several health conditions, including cardiovascular diseases [16]. Research also suggests that p-coumaric acid may play a role in cancer prevention by inhibiting the growth of cancer cells and inducing apoptosis in certain types of cancer [17]. Studies have also suggested that p-coumaric acid may support cardiovascular health by reducing the risk of atherosclerosis and improving blood vessel function [18]. Chlorogenic acid as reported in Kumar et al. [19] is considered an antioxidant, glycemic controller, anti-hypertensive, anti-inflammatory, antimicrobial, neuro-protective and anti-obesity agent. It primarily activates the AMP-activated protein kinase, inhibits 3-hydroxy 3-methylglutaryl coenzyme-A reductase and enhances the activity of carnitine palmitoyl transferase to control obesity. Ethyl gallate and syringic acid both more expressed in Elais beer can be considered markers of the olive extract addition to beer in fact syringic acid as reported by Bartel et al. [20], is present in large amounts in olive fruit and assesses its pivotal effects on oxidative stress and inflammatory parameters. It is also effective on metabolic risk factors as well, including hyperglycemia, high blood pressure, and hyperlipidemia [20]. SA is one of the prominent polyphenolic compounds that may help address health issues related to civilization diseases. Ethyl gallate, which was found by Gotes et al. [21], to inhibit the hydrogen peroxide signaling, may instead represent a valid alternative class of vasopressors before use in septic shock. Osorio-Paz et al. [22], demonstrated that vanillic acid with well-attributed antioxidant, anti-inflammatory, and neuroprotective features, may have beneficial health effects in extending health and lifespan. Protocatechuic acid was instead much more abundant in Heraclea and it could be considered as the marker for the addition of bergamot juice extract to the basal blanche beer. The effects of protocatechuic acid on health are well documented and evidenced by a wide range of pharmacological activities including antioxidant, anti-inflammatory, neuroprotective, antibacterial, antiviral, anticancer, antiosteoporotic, analgesic, antiaging activities with consequent protection against metabolic syndrome and preservation of liver, kidney, and reproductive functions [23]. The beers added with the natural extracts had a greater amount of polyphenols and single phenolic acids even if differed from each other for significant differences in the single phenolic acid concentrations.
The profile of Heraclea was notably richer in both variety and quantity of flavonoids compared to the other beers. Neoeriocitrin, naringin, and neohesperidin were highly expressed in Heraclea mainly because they are the bioactive molecules present in high concentrations in bergamot fruit (Figure 3).
These compounds exhibit a range of beneficial activities, such as antidiabetic, antiatherogenic, antidepressant, immunomodulatory, anticancer, anti-inflammatory, DNA protection, hypolipidemic, antioxidant, and cognitive-enhancing effects [24]. Melitidin and Brutieridin , as far present in Heraclea are unique to bergamot. These molecules are structurally similar to statins (drugs used to lower blood cholesterol and fatty acids), and several studies have suggested their involvement in inhibiting HMG-CoA reductase [25], leading to reduced cholesterol and fatty acids in human blood, contributing to their hypocholesterolemic properties [26]. In short, the results showed substantial and significant differences between the beers enriched with natural extracts and the basal ones. Additionally, significant differences were found between the profiles of enriched beers. When comparing Elais with Heraclea, a substantially lower total phenolic content was observed in Heraclea. Elais had a more complex phenolic profile compared to Heraclea due to the inclusion of olive extract, which is the richest in phenolic compounds. Conversely, Heraclea exhibited a superior qualitative and quantitative flavonoid profile compared to Elais, suggesting a different potentiality of the two beers in inducing health benefits for the possibility of activating/stimulating different metabolic pathways.
2.2. Effects of Lyophilized Beer Samples on Cell Viability
To assess the potential effects of the lyophilized beer, highly characterized primary human dermal fibroblasts isolated from a healthy subject have been used as described in Materials and Methods. Cell viability test was used to monitor viable cells with active metabolism after the exposure of cells in culture to the different beers at 24 and 48 hours of treatments following the chemical reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltet-razolium bromide (MTT) by mitochondrial reductases in living cells [27]
As shown in Figure 4, treatment with the different beer samples resulted in a dose- and time-dependent response on cell viability. No toxic effects were observed in the presence of all the beer samples under different treatment conditions. Notably, a significant increase in cell proliferation was observed in cells treated with 0.31 µg/µl of all the lyophilized beer samples after 24 hours of exposure, reaching 35% and 21% in the presence of Blanche and Heraclea, respectively, and 30% in the presence of Weiss and Elais, compared to untreated cells (Figure 4A). This effect persisted and significantly increased in cells treated with 0.31 µg/µl even at the longest incubation time (Figure 4B). Fibroblasts treated for 24 hours with a double concentration (0.62 µg/µl) of beer samples showed an increase in viability of 70% in the presence of Blanche and Heraclea and 100% in the presence of Weiss and Elais (Figure 4C). At the longest incubation time (48 hours), treatment with 0,62 µg/µl of the same beer samples elicited a lower response as compared to 24 hours-treatment (Figure 4D).
As already shown by Ebadi and Fazeli [28], this response could be ascribed to the longer contact time of some phenolic acids with fibroblasts, combined with a higher concentration and exposure time. No differences in cell viability were observed in the presence of enriched beers compared to the respective basal beers.
2.3. Effects of Lyophilized Beer Samples on Reactive Oxygen Species Level
Recently, the potential anti-ageing benefits of polyphenols have attracted increasing scientific interest due to their ability to modulate cellular oxidative damage 62 µg/µl. The effect of beer treatments on cellular ROS levels was then investigated to determine whether the improved vitality observed in the presence of lyophilised beers was related to their antioxidant activity. Specifically, cells were treated with 0.31 and 0.62 µg/µl of beer samples for 24 and 48 hours as previously described. The basal ROS levels of cells treated with basal beers Blanche and Weiss (0.31 62 µg/µl) were not affected at either 24 (Figure 5A) and 48 hours (Figure 5B). However, a significant decrease was observed in the presence of Heraclea (Blanche beer with bergamot juice extract) and Elais (Weiss with olive extract) at both incubation times (Figure 5A, B). Elais and Heraclea have the highest levels of total phenolic acids, compared to basal beers Blanche and Weiss, which could exert a powerful antioxidant action to protect against reactive oxygen species (ROS)/cellular oxidative stress [31]. In the presence of higher concentration of beer (0.62 µg/µl), a decrease in basal ROS levels was observed after 24h of treatment for all beer samples compared to untreated cells (Figure 5A). Nevertheless, a decrease in basal ROS levels after 48h of treatment was observed only in the Weiss beer-treated cells (Figure 5B). The lack of the antioxidant effect of the other beer samples could be ascribed to the possibility that some antioxidants can exhibit pro-oxidant behaviour under certain conditions in the biological systems [32,33,34]. Pro-oxidant behaviour allows cells to respond quickly to oxidative stress by activating various protective mechanisms [35]. To assess the effect of beer exposure under oxidative stress environment, fibroblasts were pre-treated with the beer samples, as described previously, and then incubated in the presence of t-BPH for 45 minutes as described in Matrella et al. [27]. t-BHP cell treatments induce a significant increase in intracellular ROS and mitochondrial depolarization, and reduced mitochondrial viability [36]. As expected, a significant increase in cellular ROS levels was measured in the t-BPH- treated fibroblasts, at 24 (Figure 5C) and 48 hours (Figure 5D). A significant reduction in t-BPH-induced ROS levels was observed after pre-treatment with both low and high concentrations of all beer samples over an incubation period of 24 hours, compared to t-BPH-treated cells, except for pre-treatment with low concentrations of Elais, which resulted in an increased ROS levels comparable to that induced by t-BPH treatment (Figure 5C). After 48 hours of pre-treatment, a significant reduction in t-BPH-induced ROS level was observed with 0.31 µg/µl Heraclea and Elais beer (Figure 5D). It should be noted that this significant reduction of ROS level induced by pre-treatment with enriched beers Heraclea and Elais is also relevant compared to the pre-treatment with the basal beers Blanche and Weiss, respectively (Figure 5D)
2.4. Effects of Lyophilized Beer Samples on RNA Levels of Antioxidant Enzymes
In view of these results, we hypothesised that the antioxidant function of beers may be due not only to a direct action of the beer components, but also to the activation of cellular responses that regulate gene expression levels of antioxidant enzymes. The first line of defence in the cells is antioxidant enzymes such as superoxide dismutases (SODs), catalase (CAT) and glutathione peroxidase (GPxs) involved in the dismutation of superoxide radicals and hydrogen peroxide [37]. The impact of beer treatment was assessed by comparing changes in the transcript levels of genes encoding antioxidant enzymes through real-time quantitative polymerase chain reaction (qPCR) analysis in fibroblast treated for 24 hours with 0.31 µg/µl and 0.62 µg/µl of the distinct beer samples. Under basal conditions, the scavenger genes expression was not affected by the beer samples treatment, at concentration of 0.31 µg/µl and 0.62 µg/µl (Figures A1,2). This evidence indicates that the observed reduction in ROS levels following exposure to 0.31 μg/μl of the Heraclea and Elais and 0.62 µg/µl of the basal and enriched beer samples may be attributed to the intrinsic antioxidant properties of the polyphenols and phytochemicals present in the beer extracts, rather than the activation of antioxidant enzymatic systems for ROS removal.
To assess the gene expression response to an oxidative stress condition, the t-BHP was employed as the inducing agent, as previously described [27]. Following pre-treatment with low concentrations (0.31 µg/µl) of beer samples, a significant fold increase in SOD2 expression was observed in all treated groups, except for Elais, compared to the t-BPH-treated control cells (Figure 6B). The upregulation of SOD2 gene expression, one of the key antioxidant enzymes, in response to oxidative stress induced by t-BHP reflects an adaptive cellular mechanism against oxidative damage, which differs from the outcomes observed under basal conditions. SOD2 is a mitochondrial protein which binds to superoxide byproducts of oxidative phosphorylation and converts them to hydrogen peroxide. Additionally, a significant reduction in GPX4 expression levels was detected in cells pre-treated with Blanche beer (Figure 6D).
Consistent with the effects observed at a concentration of 0.31 μg/μl, treatment with a higher concentration (0.62 μg/μl) of the beer samples resulted in a significant fold increase in SOD2 gene expression for all beer samples, except for Elais, in comparison to t-BHP-treated cells (Figure 7B). The increase in SOD2 expression without the simultaneous increase in catalase and glutathione peroxidase could lead to an increase in hydrogen peroxide contributing to cell signaling [38]. Several studies have shown that increased intracellular levels of reactive oxygen species can stimulate AMPK activity [39,40]. AMPK is the primary sensor of cellular energy which regulates diverse metabolic and physiological processes, stress responses and cell growth [41].
Data of PCA analysis evidenced that Elais and Heraclea beers behavioral differently. Elais correlated with antioxidant compounds (Phenolic acids, DPPH, TAC, Carbohydrates and MTT 0.31 and 0.62 at 48h exposure). Conversely, Heraclea correlated with flavonoids, vitamin C and MTT 0.31 at 24 h exposure. Basal beers Weiss and Blanch, correlated only with ROS both at 24 and 48 h exposure (Figure 8). The results suggested that the basal beers increased the ROS, and didn’t show any possible scavenger activity. These findings are supported by the correlation heatmap that provides valuable insights into the relationships among biochemical compounds, antioxidant activity, cytotoxicity, and oxidative stress parameters (Figure 9). A strong positive correlation (light green) was observed between total phenols, flavonoids, and antioxidant activity (DPPH, TAC), indicating that these compounds significantly contribute to the overall antioxidant potential. Furthermore, several phenolic acids (e.g., ferulic acid, chlorogenic acid, epicatechin) showed a highly significant correlation with TAC and DPPH, suggesting their essential role in free radical scavenging. Conversely, ROS levels exhibited a negative correlation (red) with most phenolic compounds, implying that these bioactive molecules effectively reduce oxidative stress. In particular, flavonoids such as hesperidin, neohesperidin, and naringin displayed strong inverse correlations with ROS, reinforcing their protective role against oxidative damage. Regarding cytotoxicity (MTT assay), higher concentrations and longer exposure times (MTT 0.62 µg/µL at 48h) showed negative correlations with some flavonoids, suggesting potential cytotoxic effects at prolonged exposure. Interestingly, some phenolic acids, including p-coumaric acid and protocatechuic acid, showed mixed correlations with ROS and MTT, suggesting a concentration-dependent or dual role in modulating oxidative stress. Additionally, flavonoids exhibited strong intercorrelations, suggesting possible synergistic effects in modulating antioxidant activity and cellular responses. Overall, these findings highlighted the significant role of phenols and flavonoids in antioxidant defense, cytotoxicity modulation, and oxidative stress regulation, with potential implications for therapeutic applications.
3. Materials and Methods
3.1. Sample Preparation
In this study, we analyzed and compared the chemical and sensory properties of two craft beers produced in Calabria: Heraclea Blanche, enriched with unfiltered and unpasteurized bergamot juice extract, and Elais Weiss, which contained unfiltered and unpasteurized olive extract. Both Calabrian craft beers were compared to their respective base varieties (Blanche and Weiss), which were free of any added extracts, to assess how the natural additions affected the aromatic profiles, flavor characteristics, and functional properties.
Each sample was analyzed in triplicate for each batch and beer variety. Prior to testing, the samples were degassed at 20°C using a magnetic stirrer to ensure the complete release of dissolved gases, thus standardizing the conditions for sensory and chemical analysis. This procedure allowed for a more accurate evaluation of the changes in flavor, aroma, and functional properties induced by the addition of Bergamot and olive extracts.
3.2. Determination of Antioxidant Compounds
3.2.1. Total Phenolic Content (TP)
Total phenolic content (TP) was determined according to Singleton & Rossi [42]. Absorbance was measured at 725 nm, and the concentration of TP was quantified using a tannic acid standard curve. The results were expressed as milligrams of tannic acid (TA) per liter of extract.
3.2.2. Total Flavonoid Content (TF)
The total flavonoid content (TF) was measured using a modified colorimetric method based on the procedure by Chang et al. [43]. Flavonoids react with aluminum chloride to form a flavonoid -Al3+ complex, which was then detected at 510 nm. The results were expressed as milligrams of quercetin (QE) per liter of extract.
3.2.3. Total Carbohydrates and Ascorbic Acid Detection
Total carbohydrates were determined by the anthrone method with minor modifications [44]. Sugars react with anthrone reagent under acidic conditions, resulting in a blue-green color. After mixing the samples with sulphuric acid and anthrone reagent, the solution was boiled, cooled, and the absorbance was measured at 620 nm. A calibration curve was prepared using known concentrations of glucose to quantify the sugar content. This method quantifies both reducing and non-reducing sugars. For ascorbic acid determination, the protocol described by Muscolo et al. [44] was followed.
3.2.4. Antioxidant Activity Assays
The 2,2′-diphenyl-1-picrylhydrazyl (DPPH %) radical scavenging activity was evaluated according to Muscolo et al. [44]. A DPPH concentration was selected to achieve an absorbance of approximately 1.0. The reaction mixture consisted of 10 μL of each extract, 700 μL of DPPH solution, and 95% ethanol to a final volume of 1.0 mL. After incubation at 37°C for 30 minutes, the absorbance was measured at 517 nm. DPPH scavenging activity was expressed as micromolar Trolox (T) equivalents, calculated from a calibration curve (1.0–50 μM Trolox).
Total antioxidant capacity (TAC) was assessed using the Oxygen Radical Absorbance Capacity (ORAC) assay, as described by Papalia et al. [45]. This assay measures the inhibition of peroxyl radical-induced oxidation and provides an estimate of the total antioxidant capacity of the sample
3.2.5. Polyphenolic Profile Determination
Beer samples were degassed by magnetic stirring (500 rpm) for 8 hours and filtered through a 0.45 μm regenerated cellulose filter (Aisino Corporation). The polyphenolic profile was analyzed using Ultra-High-Performance Liquid Chromatography (UHPLC) with a Photo-Diode Array (PDA) detector (Shimadzu, Milan, Italy). The UHPLC system was equipped with a column oven (CTO-20AC), an autosampler (SIL-30AC), and an in-line degasser (DGU-20A5R). The chromatographic separation was performed on a Kinetex C18 column (50 mm × 3 mm × 1.7 μm particle size; Phenomenex), with a Kinetex C18 guard column (Phenomenex) used to protect the analytical column [4].
The optimized chromatographic conditions were as follows: mobile phase A was water with 0.1% formic acid, and mobile phase B was acetonitrile with 0.1% formic acid. The flow rate was set at 0.6 mL/min, and the column temperature was maintained at 40°C. Gradient elution was performed: 1% B for 5 minutes, followed by a 15-minute gradient from 1% to 30% B, and then a 7.5-minute gradient from 30% to 65% B, before washing and reconditioning the system. The total separation time was approximately 28 minutes.
The PDA detector was set with a spectrum resolution of 256 nm, a split width of 8 nm, and a sampling rate of 40 Hz. Data acquisition was performed over a wavelength range of 190–400 nm, and chromatograms were recorded at the optimal absorbance for the compounds of interest [4].
3.2.6. Validation Method for Polyphenols
Seven concentration levels of the polyphenolic standards were prepared with methanol from a 1000 mg/L stock solution with concentration range of 0.5-120 mg/L. Five analyzes were performed for each concentration level with the HPLC-PDA system under optimized chromatographic conditions. Retention time, instrumental recovery and percentage relative standard deviation (RSD%) were determined using the fourth level (n=4) of each calibration curve (Table 1).
3.3. Lyophilized Sample Preparation for Cell Culture Treatments
The lyophilized beer samples were dissolved in phosphate-buffered saline (PBS), filtered through a 0.22 µm membrane filter and sterilized. The final concentration of the stock solution was 33.4 mg/ml for Blanche basal beer (B) and Blanche basal beer with the addition of the bergamot extract Heraclea (H) and 53.82 mg/ml for Weiss basal beer (W) and Weiss basal beer with the addition of olive extract Elais (E). For the cell culture treatments, the concentrations of the beer samples tested were 0.31 and 0.62 µg/µL.
3.4. Cells and Culture Conditions
Primary fibroblasts from a healthy subject obtained by explants from a skin punch biopsy after informed consent [46,47,48] were grown in high-glucose Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% (v/v) fetal bovine serum (FBS), 1% (v/v) L-glutamine, and 1% (v/v) penicillin/streptomycin at 37 C in a humidified atmosphere of 5% CO2. For the treatment conditions, cells were seeded in 96-well plates and grown for 24 h. For treatment conditions, cells were seeded in 96-well plates and grown for 24 h. The media were then removed, and cells were cultured at 37 °C in culture plates for 24 and 48 h in media containing different concentrations of lyophilized beer
3.5. Cell Viability
Cell viability was assessed by the 3-(4,5-dimethylthiazol-2-yl)2,5-diphenyltetrazolium (MTT) assay [49] after 24 and 48 h of exposure of the cells and seeded in 96-well plates to the different concentrations of lyophilized beer samples. After the incubation, 150 µL DMEM and 15 µL of MTT (5 mg/mL) were added to each well. The plates were incubated for 3 h at 37 °C. The media were removed, and formazan crystals were dissolved in 150 µL isopropanol by gentle shaking. Absorbance was measured at 570 nm using a Victor 2030 multi-label reader (PerkinElmer, Waltham, MA, USA).
3.6. Determination of Reactive Oxygen Species (ROS)
The H2O2 levels were measured using the cell-permeant probe 2'-7'-dichlorodihydrofluorescindiacetate (H2DCFDA). Briefly, after 24 and 48 h of exposure to lyophilized beer samples, the media were changed and oxidative stress was induced by treatment with 50 µM tert-butyl hydroperoxide (t-BHP) (Sigma-Aldrich, B2633, St. Louis, MO, USA), as described in [27]. After 45 min of treatment, the cells were incubated with 10 µM H2DCFDA for 20 min at 37 °C in the dark. The cells were then washed and resuspended in 150 µL PBS. The H2O2-dependent oxidation of the fluorescent probe was measured (at 507 nm excitation and 530 nm emission wavelength) using the Victor 2030 Multilabel Reader (PerkinElmer, Waltham, MA, USA).
3.7. RNA Isolation Reverse Transcription and Quantitative PCR
The purification of total RNA from cells was carried out by using Aurum Total RNAMini Kit (Bio-Rad, Hercules,CA, USA) according to the manufacturer’s protocol. Complementary DNA (cDNA) was synthesized by reverse-transcription of total RNA using the iScriptcDNA Synthesis kit (Bio-Rad, Hercules, CA, USA), following the manufacturer’s instructions. Semi-quantitative determination of mRNA levels was performed by Real-Time Quantitative Reverse Transcription PCR (qRT-PCR) using SsoAdvanced Universal SYBRR Green Supermix (Bio-Rad, Hercules, CA, USA). Reactions were performed in a CFX96 Touch Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA, USA) in duplicate for each sample for three independent experiments. Relative quantification was performed using the comparative CT method (ΔΔCT). Quantitative normalization for each sample was performed by using glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as an internal control. Validated primers for semi-qRT-PCR are provided in Table 2.
3.8. Statistical Analysis
Analysis of variance was carried out for all the data sets. One-way ANOVA with Tukey's Honestly. Powerful Statistical Analysis and Graphics Software for Windows 7, was used for all the statistical analyses. Effects were considered significant at p ≤ 0.05.
4. Conclusions
In short, these results suggest that beer extracts, particularly those rich in phenolic compounds, may have a nuanced impact on cellular oxidative stress, with potential therapeutic implications for managing oxidative damage. The differential behavior of the various beer samples particularly the unique antioxidant profiles of Elais and Heraclea points to the importance of the specific composition of bioactive compounds in influencing both antioxidant activity and cytotoxicity. Furthermore, the observed concentration- and time-dependent effects highlight the complexity of beer's interactions with cellular mechanisms, suggesting that both the concentration of bioactive compounds and the duration of exposure are critical factors in modulating oxidative stress responses. Future research could further investigate the role of specific phenolic compounds in the regulation of oxidative stress, as well as the long-term effects of beer consumption on cellular health, to better understand its potential as a functional food with antioxidant benefits.
Author Contributions
Conceptualization, writing and editing Adele Muscolo; methodology, Maria Laura Metrella, Angela Maffia and Francesco Canino; software, Angela Maffia, Maria Laura Metrella; validation, Tiziana Cocco and Adele Muscolo; formal analysis and investigation Francesco Canino and Maria Laura Metrella, data curation, Adele Muscolo, Russo Mariateresa; writing—original draft preparation, Tiziana Cocco, Maria Laura Metrella and Adele Muscolo; writing—review and editing, Mariateresa Russo; supervision, Adele Muscolo, Tiziana Cocco and Maria Teresa Russo; Adele Muscolo; funding acquisition, Adele Muscolo and Russo Mariateresa. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation.
Funding
This research received no external funding” or “This research was funded by NAME OF FUNDER, grant number XXX” and “The APC was funded by XXX”. Check carefully that the details given are accurate and use the standard spelling of funding agency names at https://search.crossref.org/funding. Any errors may affect your future funding.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Poelmans, E.; Swinnen, J.F.M. 1 A brief economic history of beer. In Oxford University Press eBooks; 2011; pp 3–28. [CrossRef]
- Paiva, R. a. M.; Mutz, Y.S.; Conte-Junior, C.A. A review on the obtaining of functional beers by addition of Non-Cereal adjuncts rich in antioxidant compounds. Antioxidants 2021, 10, 1332. [Google Scholar] [CrossRef] [PubMed]
- Habschied, K.; Živković, A.; Krstanović, V.; Mastanjević, K. Functional Beer—A review on possibilities. Beverages 2020, 6, 51. [Google Scholar] [CrossRef]
- Muscolo, A.; Marra, F.; Salafia, F.; Andronaco, P.; Di Sanzo, R.; Carabetta, S.; Russo, Mt. Bergamot and olive extracts as beer ingredients: their influence on nutraceutical and sensory properties. European Food Research and Technology 2022, 248, 2067–2077. [Google Scholar] [CrossRef]
- Maruca, N.G.; Laghetti, N.G.; Mafrica, N.R.; Turiano, N.D.; Hammer, N.K. The Fascinating History of Bergamot (Citrus Bergamia Risso & Poiteau), The Exclusive Essence of Calabria: a review. Journal of Environmental Science and Engineering - A 2017, 6. [Google Scholar] [CrossRef]
- Gattuso, A.; Mafrica, R.; Cannavò, S.; Mafrica, D.; De Bruno, A.; Poiana, M. Quality evaluation of bergamot juice produced in different areas of Calabria region. Foods 2024, 13, 2080. [Google Scholar] [CrossRef]
- Adorisio, S.; Muscari, I.; Fierabracci, A.; Thuy, T.T.; Marchetti, M.C.; Ayroldi, E.; Delfino, D.V. Biological effects of bergamot and its potential therapeutic use as an anti-inflammatory, antioxidant, and anticancer agent. Pharmaceutical Biology 2023, 61, 639–646. [Google Scholar] [CrossRef]
- Russo, M.; Arigò, A.; Calabrò, M.L.; Farnetti, S.; Mondello, L.; Dugo, P. Bergamot ( Citrus bergamia Risso ) as a source of nutraceuticals: Limonoids and flavonoids. Journal of Functional Foods 2015, 20, 10–19. [Google Scholar] [CrossRef]
- Roy, A.; Khan, A.; Ahmad, I.; Alghamdi, S.; Rajab, B.S.; Babalghith, A.O.; Alshahrani, M.Y.; Islam, S.; Islam, Md. R. Flavonoids a Bioactive Com-pound from Medicinal Plants and Its Therapeutic Applications. BioMed Research International 2022, 2022, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Maria, G.A.; Riccardo, N. Citrus bergamia, Risso: the peel, the juice and the seed oil of the bergamot fruit of Reggio Calabria (South Italy). Emirates Journal of Food and Agriculture 2020, 522. [Google Scholar] [CrossRef]
- Gorzynik-Debicka, M.; Przychodzen, P.; Cappello, F.; Kuban-Jankowska, A.; Gammazza, A.M.; Knap, N.; Wozniak, M.; Gorska-Ponikowska, M. Potential health benefits of olive oil and plant polyphenols. International Journal of Molecular Sciences 2018, 19, 686. [Google Scholar] [CrossRef]
- Bullo, M.; Lamuela-Raventos, R.; Salas-Salvado, J. Mediterranean diet and oxidation: Nuts and olive oil as important sources of fat and antioxidants. Current Topics in Medicinal Chemistry 2011, 11, 1797–1810. [Google Scholar] [CrossRef] [PubMed]
- Selim, S.; Albqmi, M.; Al-Sanea, M.M.; Alnusaire, T.S.; Almuhayawi, M.S.; AbdElgawad, H.; Jaouni, S.K.A.; Elkelish, A.; Hussein, S.; Warrad, M.; El-Saadony, M.T. Valorizing the usage of olive leaves, bioactive compounds, biological activities, and food applications: A comprehensive review. Frontiers in Nutrition 2022, 9. [Google Scholar] [CrossRef]
- De Bruno, A.; Zappia, A.; Piscopo, A.; Poiana, M. Qualitative evaluation of fermented olives grown in Southern Italy (cvs. Carolea, Grossa of Gerace and Nocellara Messinese). Emirates Journal of Food and Agriculture 2019, 587. [Google Scholar] [CrossRef]
- Roychoudhury, S.; Sinha, B.; Choudhury, B.P.; Jha, N.K.; Palit, P.; Kundu, S.; Mandal, S.C.; Kolesarova, A.; Yousef, M.I.; Ruokolainen, J.; Slama, P.; Kesari, K.K. Scavenging properties of Plant-Derived natural biomolecule Para-Coumaric acid in the prevention of oxidative Stress-Induced diseases. Antioxidants 2021, 10, 1205. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.J.; Prince, P.S.M. Preventive effects of p-coumaric acid on cardiac hypertrophy and alterations in electrocardiogram, lipids, and lipoproteins in experimentally induced myocardial infarcted rats. Food and Chemical Toxicology 2013, 60, 348–354. [Google Scholar] [CrossRef]
- Tehami, W.; Nani, A.; Khan, N.A.; Hichami, A. New insights into the anticancer effects of P-Coumaric acid: Focus on colorectal cancer. Dose-Response 2023, 21. [Google Scholar] [CrossRef] [PubMed]
- Ziółkiewicz, A.; Kasprzak-Drozd, K.; Rusinek, R.; Markut-Miotła, E.; Oniszczuk, A. The influence of polyphenols on atherosclerosis development. International Journal of Molecular Sciences 2023, 24, 7146. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Sharma, A.; Iqbal, M.S.; Srivastava, J.K. Therapeutic Promises of Chlorogenic Acid with Special Emphasis on its Anti-Obesity Property. Current Molecular Pharmacology 2019, 13, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Bartel, I.; Mandryk, I.; Horbańczuk, J.O.; Wierzbicka, A.; Koszarska, M. Nutraceutical Properties of Syringic Acid in Civilization Diseases—Review. Nutrients 2023, 16, 10. [Google Scholar] [CrossRef] [PubMed]
- Gotes, J.; Kasian, K.; Jacobs, H.; Cheng, Z.-Q.; Mink, S.N. Benefits of ethyl gallate versus norepinephrine in the treatment of cardiovascular collapse in Pseudomonas aeruginosa septic shock in dogs*. Critical Care Medicine 2011, 40, 560–572. [Google Scholar] [CrossRef]
- Osorio-Paz, I.; Valle-Jiménez, X.; Brunauer, R.; Alavez, S. Vanillic Acid Improves Stress Resistance and Substantially Extends Life Span in Caenorhabditis elegans. The Journals of Gerontology Series A 2023, 78, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; He, Y.; Luo, C.; Feng, B.; Ran, F.; Xu, H.; Ci, Z.; Xu, R.; Han, L.; Zhang, D. New progress in the pharmacology of protocatechuic acid: A compound ingested in daily foods and herbs frequently and heavily. Pharmacological Research 2020, 161, 105109. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.-H.; Jaremko, M. Important flavonoids and their role as a therapeutic agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef]
- Leopoldini, M.; Malaj, N.; Toscano, M.; Sindona, G.; Russo, N. On the Inhibitor Effects of Bergamot Juice Flavonoids Binding to the 3-Hydroxy-3-methylglutaryl-CoA Reductase (HMGR) Enzyme. Journal of Agricultural and Food Chemistry 2010, 58, 10768–10773. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tocmo, R.; Nauman, M.C.; Haughan, M.A.; Johnson, J.J. Defining the Cholesterol Lowering Mechanism of Bergamot (Citrus bergamia) Extract in HepG2 and Caco-2 Cells. Nutrients 2021, 13, 3156. [Google Scholar] [CrossRef]
- Matrella, M.L.; Valletti, A.; Marra, F.; Mallamaci, C.; Cocco, T.; Muscolo, A. Phytochemicals from Red Onion, Grown with Eco-Sustainable Fertilizers, Protect Mammalian Cells from Oxidative Stress, Increasing Their Viability. Molecules 2022, 27, 6365. [Google Scholar] [CrossRef] [PubMed]
- Ebadi, P.; Fazeli, M. Evaluation of the potential in vitro effects of propolis and honey on wound healing in human dermal fibroblast cells. South African Journal of Botany 2021, 137, 414–422. [Google Scholar] [CrossRef]
- Tatullo, M.; Simone, G.M.; Tarullo, F.; et al. Antioxidant and antitumor activity of a bioactive polyphenolic fraction isolated from the brewing process. Scientific Reports. 2016, 6, 36042. [Google Scholar] [CrossRef] [PubMed]
- Nardini, M. An Overview of Bioactive Phenolic Molecules and Antioxidant Properties of Beer: Emerging Trends. Molecules. 2023, 28, 3221. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef]
- Babich, H.; Schuck, A.G.; Weisburg, J.H.; Zuckerbraun, H.L. (2011). Research Strategies in the Study of the Pro-oxidant Nature of Polyphenol Nutraceuticals. J. Toxicol 2011, 1–12. [Google Scholar] [CrossRef]
- Ivonne, M.C.M. 33. Ivonne M.C.M. Rietjens, Marelle G. Boersma, Laura de Haan, Bert Spenkelink, Hanem M. Awad, Nicole H.P. Cnubben, Jelmer J. van Zanden, Hester van der Woude, Gerrit M. Alink, Jan H. Koeman, The pro-oxidant chemistry of the natural antioxidants vitamin C, vitamin E, carotenoids and flavonoids. Environmental Toxicology and Pharmacology 2002, 11, 321–333. [Google Scholar]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant Activity of Polyphenols, Flavonoids, Anthocyanins and Carotenoids: Updated Review of Mechanisms and Catalyzing Metals. Phytother Res. 2016, 30, 1379–91. [Google Scholar] [CrossRef] [PubMed]
- Lecci, R.M.; D'Antuono, I.; Cardinali, A.; Garbetta, A.; Linsalata, V.; Logrieco, A.F.; Leone, A. Antioxidant and Pro-Oxidant Capacities as Mechanisms of Photoprotection of Olive Polyphenols on UVA-Damaged Human Keratinocytes. Molecules. 2021, 26, 2153. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Luo, G.; Giannelli, S.; Szeto, H.H. Mitochondria-targeted peptide prevents mitochondrial depolarization and apoptosis induced by tert-butyl hydroperoxide in neuronal cell lines. Biochem Pharmacol. 2005, 70, 1796–806. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Alomar, S.Y.; Alwasel, S.H.; et al. Several lines of antioxidant defense against oxidative stress: antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Arch Toxicol 2024, 98, 1323–1367. [Google Scholar] [CrossRef]
- Di Marzo, N.; Chisci, E.; Giovannoni, R. The Role of Hydrogen Peroxide in Redox-Dependent Signaling: Homeostatic and Pathological Responses in Mammalian Cells. Cells. 2018, 7, 156. [Google Scholar] [CrossRef] [PubMed]
- Zmijewski, J.W.; Banerjee, S.; Bae, H.; Friggeri, A.; Lazarowski, E.R.; Abraham, E. Exposure to hydrogen peroxide induces oxidation and activation of AMP-activated protein kinase. J Biol Chem. 2010, 285, 33154–33164. [Google Scholar] [CrossRef]
- Auciello, F.R.; Ross, F.A.; Ikematsu, N.; Hardie, D.G. Oxidative stress activates AMPK in cultured cells primarily by increasing cellular AMP and/or ADP. FEBS Lett. 2014, 588, 3361–6. [Google Scholar] [CrossRef]
- Jeon, S.M. Regulation and function of AMPK in physiology and diseases. Exp Mol Med. 2016, 48, e245. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. American Journal of Enology and Viticulture 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total favonoid content in propolis by two complementary colorimetric methods. J Food Drug Anal 2002, 10, 178–182. [Google Scholar]
- Muscolo, A.; Calderaro, A.; Papalia, T.; Settineri, G.; Mallamaci, C.; Panuccio, M.R. Soil salinity improves nutritional and health promoting compounds in three varieties of lentil (Lens culinaris Med.). Food Bioscience 2020, 35, 100571. [Google Scholar] [CrossRef]
- Papalia, T.; Barreca, D.; Panuccio, M. Assessment of Antioxidant and Cytoprotective Potential of Jatropha (Jatropha curcas) Grown in Southern Italy. International Journal of Molecular Sciences 2017, 18, 660. [Google Scholar] [CrossRef] [PubMed]
- Pacelli, C.; De Rasmo, D.; Signorile, A.; Grattagliano, I.; Di Tullio, G.; D’Orazio, A.; Nico, B.; Pietro Comi, G.; Ronchi, D.; Ferranini, E.; Pirolo, D.; Seibel, P.; Schubert, S.; Gaballo, A.; Villani, G.; Cocco, T. Mitochondrial defect and PGC-1α dysfunction in parkin-associated familial Parkinson’s disease. Biochimica Et Biophysica Acta (BBA) - Molecular Basis of Disease 2011, 1812, 1041–1053. [Google Scholar] [CrossRef]
- Vergara, D.; Gaballo, A.; Signorile, A.; Ferretta, A.; Tanzarella, P.; Pacelli, C.; Di Paola, M.; Cocco, T.; Maffia, M. Resveratrol Modulation of protein expression in parkin-Mutant human skin fibroblasts: A proteomic approach. Oxidative Medicine and Cellular Longevity 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Tanzarella, P.; Ferretta, A.; Barile, S.N.; Ancona, M.; De Rasmo, D.; Signorile, A.; Papa, S.; Capitanio, N.; Pacelli, C.; Cocco, T. Increased levels of CAMP by the Calcium-Dependent activation of soluble adenylyl cyclase in Parkin-Mutant fibroblasts. Cells 2019, 8, 250. [Google Scholar] [CrossRef] [PubMed]
- Van De Loosdrecht, A.A.; Beelen, R.H.J.; Ossenkoppele, G.J.; Broekhoven, M.G.; Langenhuijsen, M.M.A.C. A tetrazolium-based colorimetric MTT assay to quantitate human monocyte mediated cytotoxicity against leukemic cells from cell lines and patients with acute myeloid leukemia. Journal of Immunological Methods 1994, 174, 311–320. [Google Scholar] [CrossRef]
Figure 1.
Nutritional values of Elais (a), Heraclea (b), Blanche (c) and Weiss (d). Protein (%) Carbohydrates (%), Phenols (%), flavonoids (%), Vitamin C (%), 2,2-diphenyl-1-picrylhydrazyl (DPPH %), total antioxidant capacity (TAC %).
Figure 1.
Nutritional values of Elais (a), Heraclea (b), Blanche (c) and Weiss (d). Protein (%) Carbohydrates (%), Phenols (%), flavonoids (%), Vitamin C (%), 2,2-diphenyl-1-picrylhydrazyl (DPPH %), total antioxidant capacity (TAC %).
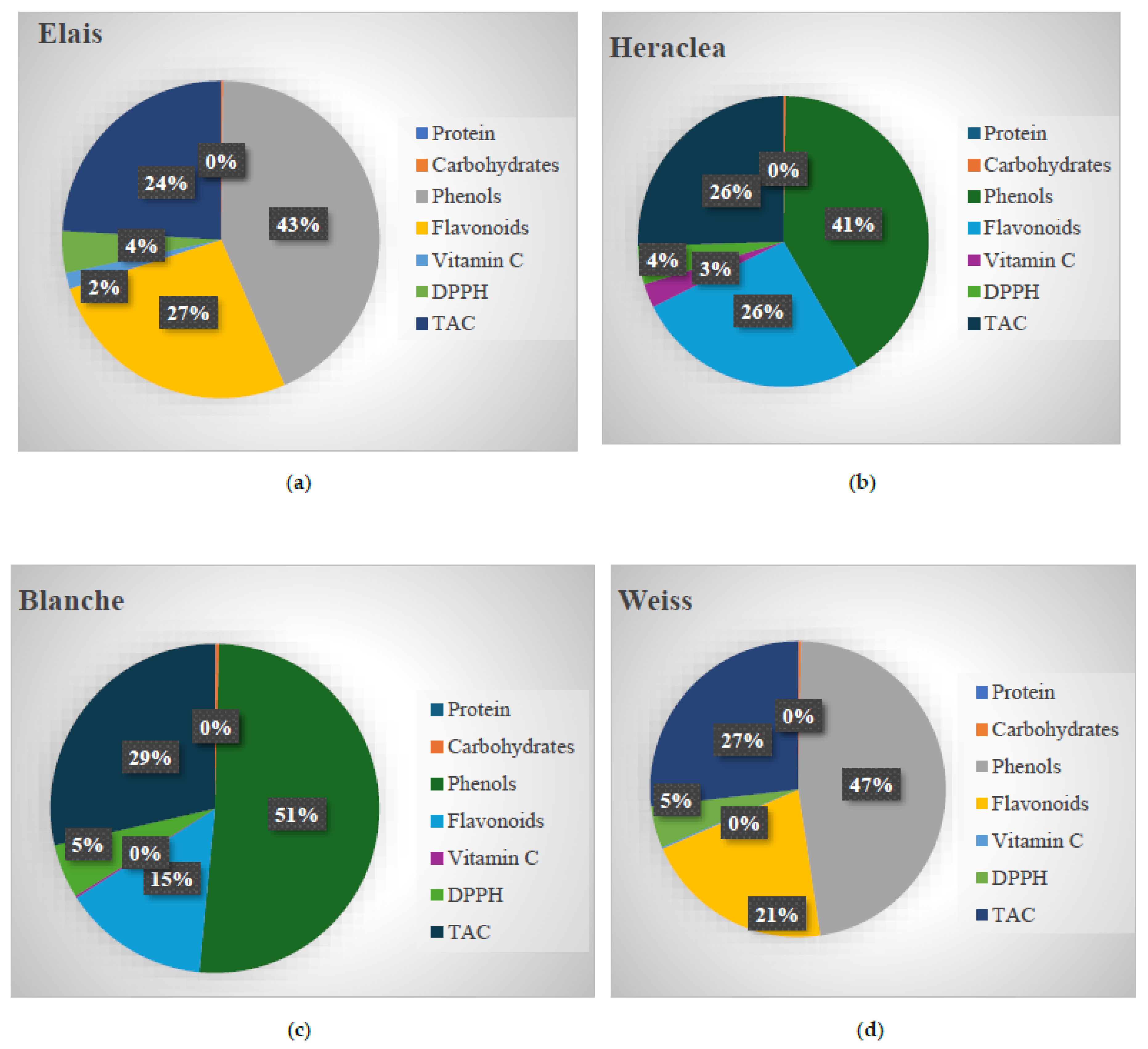
Figure 2.
Phenolic acid content (ng/µL) in different craft beers: Blanche base beer; Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); Weiss base beer, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised). Data are the mean of 3 replications ± standard deviation.
Figure 2.
Phenolic acid content (ng/µL) in different craft beers: Blanche base beer; Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); Weiss base beer, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised). Data are the mean of 3 replications ± standard deviation.
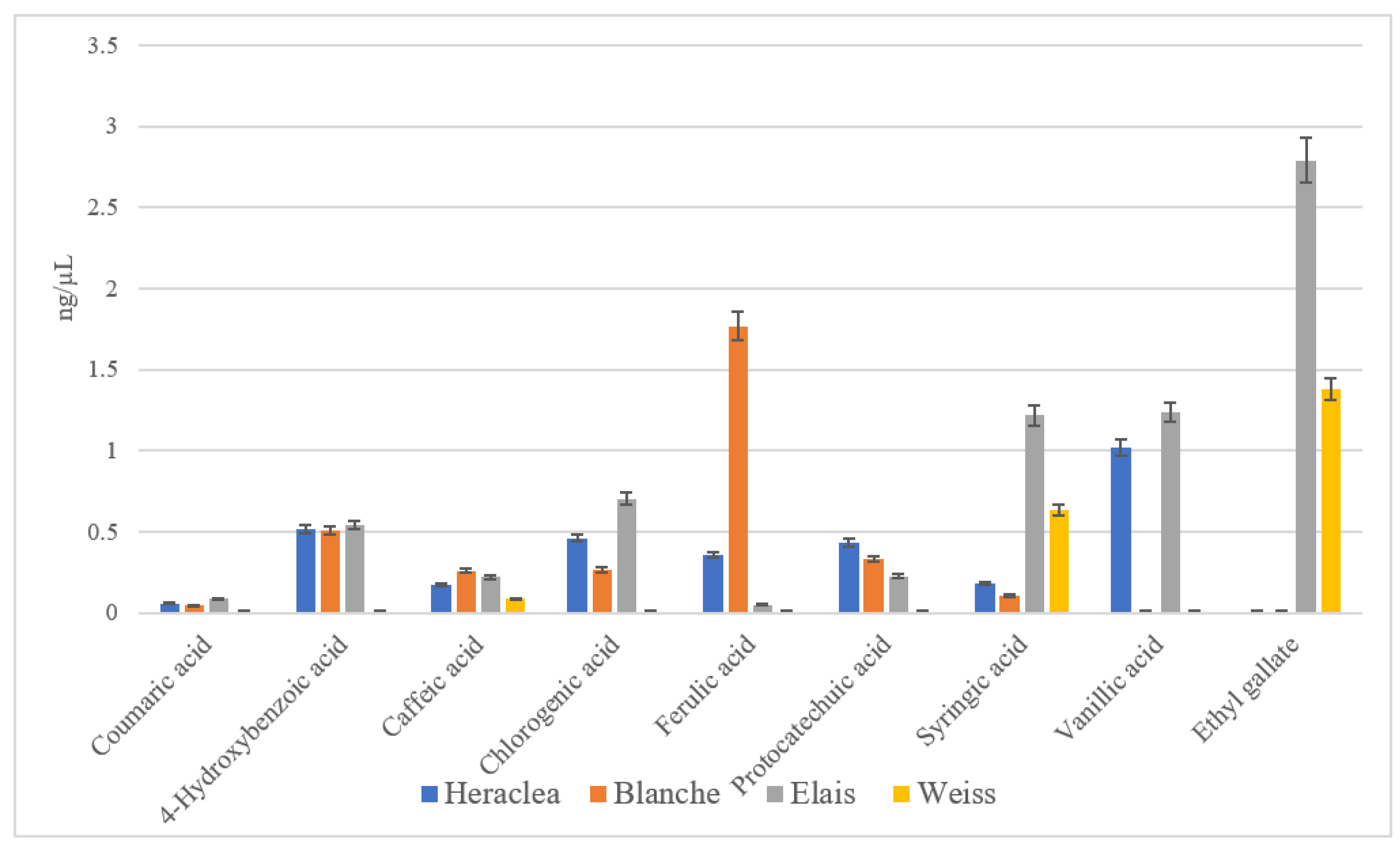
Figure 3.
Flavonoids content (ng/µL) in different craft beers. Blanche base beer; Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); Weiss base beer, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised). Data are the mean of 3 replications ± standard deviation.
Figure 3.
Flavonoids content (ng/µL) in different craft beers. Blanche base beer; Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); Weiss base beer, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised). Data are the mean of 3 replications ± standard deviation.
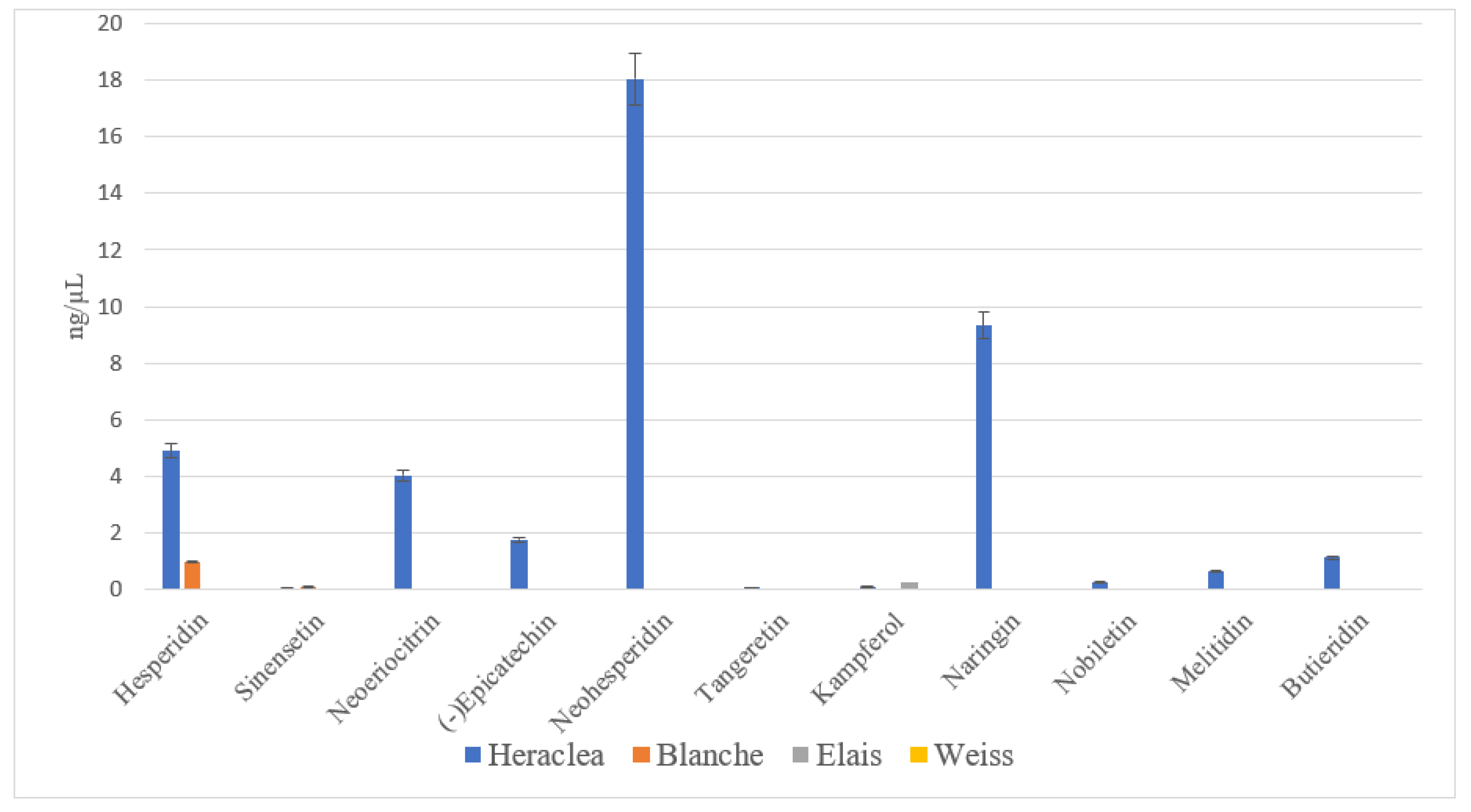
Figure 4.
Enhanced cellular viability in beer–treated human fibroblasts. Cells were treated with 0.31 µg/µl (A-B) and 0.62 µg/µl (C-D) of different beer samples for 24 (A-C) and 48 hours (B-D). Data mean ± SEM from at least three independent experiments under each condition are expressed as the percentage of vehicle-treated cells (NT). Statistical analyses were performed using Brown-Forsythe and Welch’s one-way analysis of variance. * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001. B, Blanche basal beer; H, Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); W, Weiss basal beer; E, Elais (Weiss with the addition of olive extract not filtered and not pasteurised).
Figure 4.
Enhanced cellular viability in beer–treated human fibroblasts. Cells were treated with 0.31 µg/µl (A-B) and 0.62 µg/µl (C-D) of different beer samples for 24 (A-C) and 48 hours (B-D). Data mean ± SEM from at least three independent experiments under each condition are expressed as the percentage of vehicle-treated cells (NT). Statistical analyses were performed using Brown-Forsythe and Welch’s one-way analysis of variance. * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001. B, Blanche basal beer; H, Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); W, Weiss basal beer; E, Elais (Weiss with the addition of olive extract not filtered and not pasteurised).
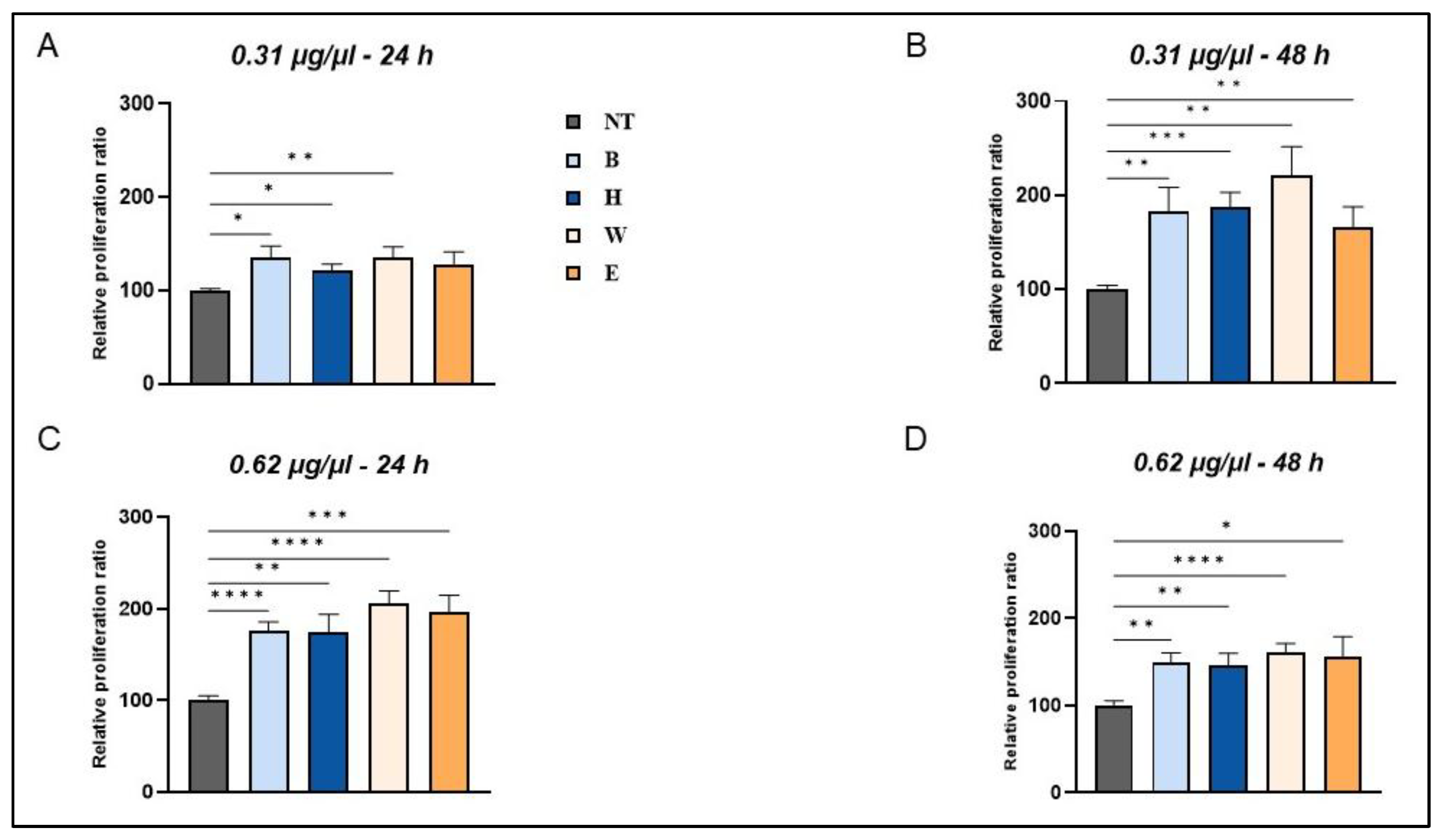
Figure 5.
Reduced H2O2 levels in beer–treated human fibroblasts. Cells were treated with 0.31 µg/µl and 0.61 µg/µl of different beer samples for 24 and 48 hours. ROS levels were measured by the DCF assay under basal conditions (A, B) and after exposure to the exogenous oxidative stress inducer t-BHP (C, D). Data mean ± SEM from at least three independent experiments under each condition are expressed as a percentage of vehicle-treated cells (dotted line) or tert-butyl hydroperoxide (t-BPH) treated cells (red line). Statistical analyses were performed using Brown-Forsythe and Welch's one-way analysis of variance. * p < 0.05, ** p < 0.01 and *** p < 0.001. Asterisks indicate statistically significant differences compared to vehicle or T-BPH-treated cells. Triangles indicate statistical significance (Brown-Forsythe and Welch's one-way analysis of variance) as compared to the respective base beers. ** p < 0.01 and *** p < 0.001. B, Blanche base beer; H, Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); W, Weiss base beer; E, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised).
Figure 5.
Reduced H2O2 levels in beer–treated human fibroblasts. Cells were treated with 0.31 µg/µl and 0.61 µg/µl of different beer samples for 24 and 48 hours. ROS levels were measured by the DCF assay under basal conditions (A, B) and after exposure to the exogenous oxidative stress inducer t-BHP (C, D). Data mean ± SEM from at least three independent experiments under each condition are expressed as a percentage of vehicle-treated cells (dotted line) or tert-butyl hydroperoxide (t-BPH) treated cells (red line). Statistical analyses were performed using Brown-Forsythe and Welch's one-way analysis of variance. * p < 0.05, ** p < 0.01 and *** p < 0.001. Asterisks indicate statistically significant differences compared to vehicle or T-BPH-treated cells. Triangles indicate statistical significance (Brown-Forsythe and Welch's one-way analysis of variance) as compared to the respective base beers. ** p < 0.01 and *** p < 0.001. B, Blanche base beer; H, Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); W, Weiss base beer; E, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised).
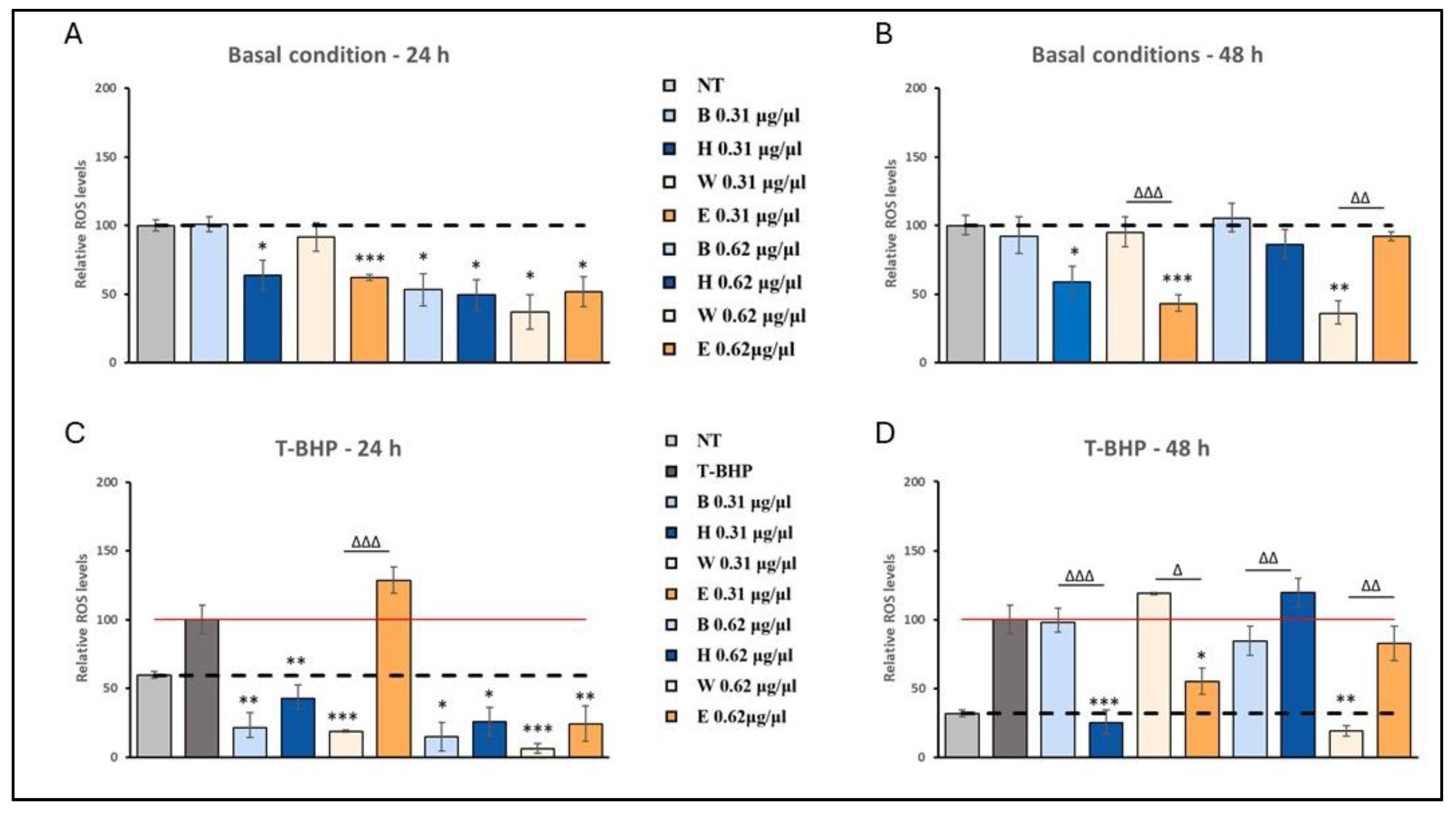
Figure 6.
Modulation of antioxidant enzymes expression in beer–treated human fibroblasts. Cells were treated with 0.31 µg/µl of different beer samples for 24 h. Transcript levels of NRF2 (A), SOD2 (B), GPX1 (C), GPX4 (D) and CATA (E) were measured after exposure to the exogenous oxidative stress inducer t-BHP. Relative mRNA levels were evaluated by qRT-PCR and normalized to the housekeeping gene GAPDH. Data expressed as fold-change mRNA expression levels in beer-treated cells, compared to t-BPH treated cells, are means±SEM of two replicates from three independent experiments. Statistical analyses were performed using Brown-Forsythe and Welch's one-way analysis of variance. * p < 0.05, ** p < 0.01 and *** p < 0.001. Asterisks indicate statistically significant differences compared to T-BPH treated cells. B, Blanche base beer; H, Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); W, Weiss base beer; E, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised).
Figure 6.
Modulation of antioxidant enzymes expression in beer–treated human fibroblasts. Cells were treated with 0.31 µg/µl of different beer samples for 24 h. Transcript levels of NRF2 (A), SOD2 (B), GPX1 (C), GPX4 (D) and CATA (E) were measured after exposure to the exogenous oxidative stress inducer t-BHP. Relative mRNA levels were evaluated by qRT-PCR and normalized to the housekeeping gene GAPDH. Data expressed as fold-change mRNA expression levels in beer-treated cells, compared to t-BPH treated cells, are means±SEM of two replicates from three independent experiments. Statistical analyses were performed using Brown-Forsythe and Welch's one-way analysis of variance. * p < 0.05, ** p < 0.01 and *** p < 0.001. Asterisks indicate statistically significant differences compared to T-BPH treated cells. B, Blanche base beer; H, Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); W, Weiss base beer; E, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised).
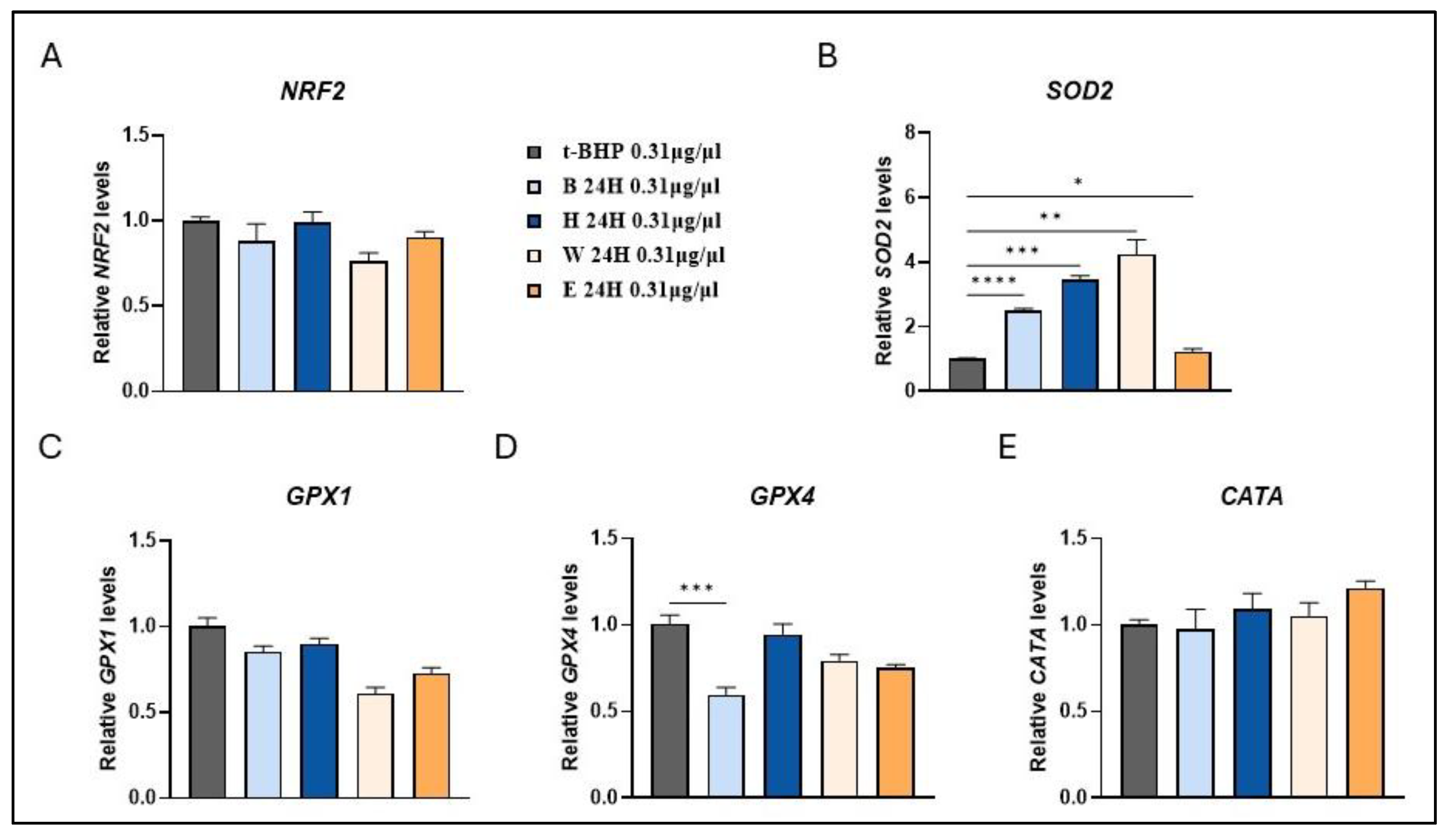
Figure 7.
Modulation of antioxidant enzymes expression by beer samples in t-BHP treated cells. Cells were treated with 0.62 µg/µl of different beer samples for 24 h. Transcript levels of NRF2 (A), SOD2 (B), GPX1 (C), GPX4 (D) and CATA (E) were measured after exposure to the exogenous oxidative stress inducer t-BHP. Relative mRNA levels were evaluated by qRT-PCR and normalized to the housekeeping gene GAPDH. Data, expressed as fold-change mRNA expression levels in beer-treated cells, compared to t-BPH treated cells, are means±SEM of two replicates from three independent experiments. Statistical analyses were performed using Brown-Forsythe and Welch's one-way analysis of variance. * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001. Asterisks indicate statistically significant differences compared to T-BPH treated cells. B, Blanche base beer; H, Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); W, Weiss base beer; E, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised).
Figure 7.
Modulation of antioxidant enzymes expression by beer samples in t-BHP treated cells. Cells were treated with 0.62 µg/µl of different beer samples for 24 h. Transcript levels of NRF2 (A), SOD2 (B), GPX1 (C), GPX4 (D) and CATA (E) were measured after exposure to the exogenous oxidative stress inducer t-BHP. Relative mRNA levels were evaluated by qRT-PCR and normalized to the housekeeping gene GAPDH. Data, expressed as fold-change mRNA expression levels in beer-treated cells, compared to t-BPH treated cells, are means±SEM of two replicates from three independent experiments. Statistical analyses were performed using Brown-Forsythe and Welch's one-way analysis of variance. * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001. Asterisks indicate statistically significant differences compared to T-BPH treated cells. B, Blanche base beer; H, Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); W, Weiss base beer; E, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised).
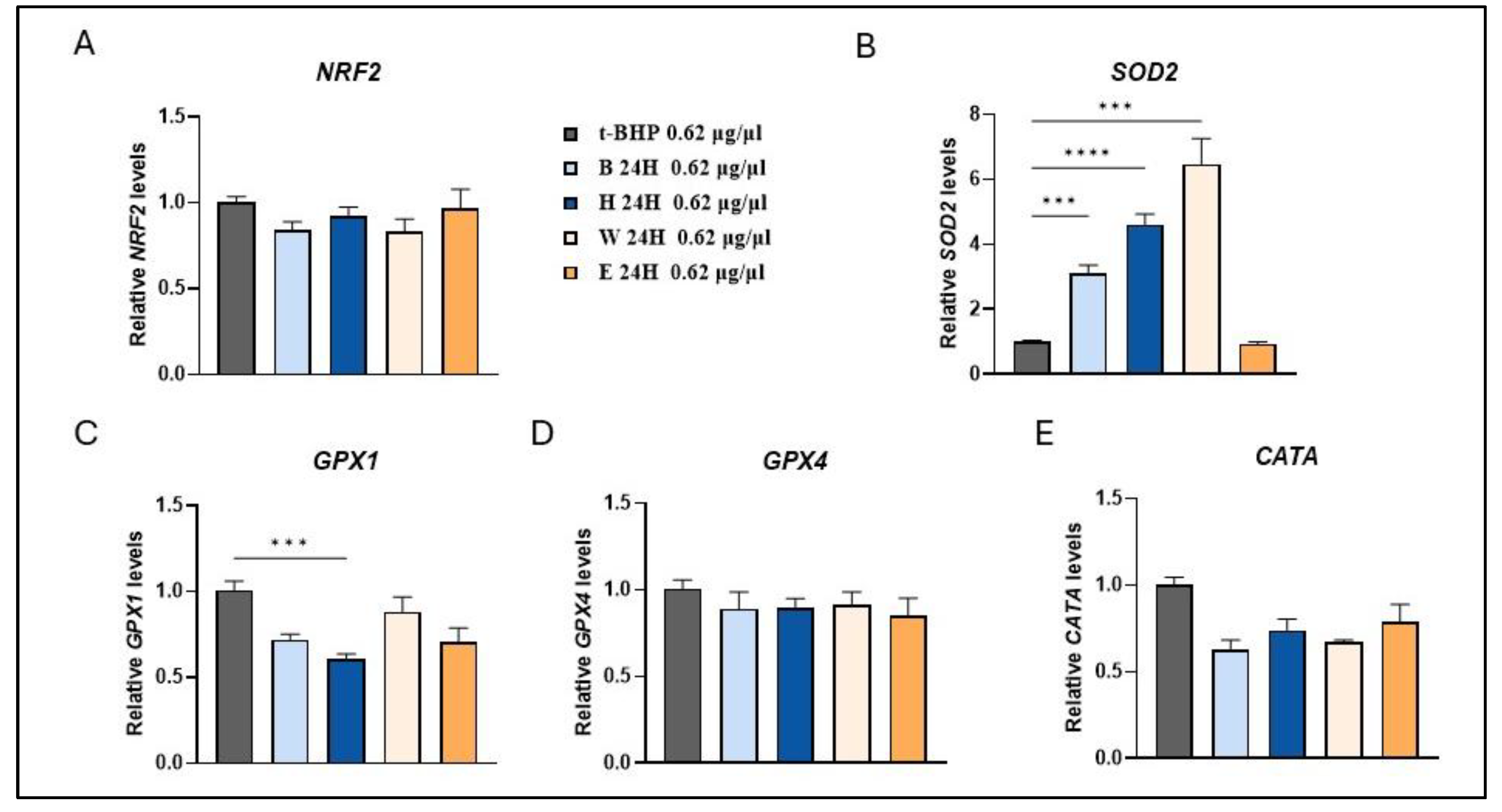
Figure 8.
Principal Component Analysis (PCA) showing how the variables (total phenols, total flavonoids, phenolic acids, flavonoids and vitamins) contained in the different beers, relate to each other, to reactive oxygen species and to cell viability (MTT) at two different concentration (0.31 and 0.62 µg/µL) and exposure time (24 and 48 h). B, Blanche base beer; H, Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); W, Weiss base beer; E, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised).
Figure 8.
Principal Component Analysis (PCA) showing how the variables (total phenols, total flavonoids, phenolic acids, flavonoids and vitamins) contained in the different beers, relate to each other, to reactive oxygen species and to cell viability (MTT) at two different concentration (0.31 and 0.62 µg/µL) and exposure time (24 and 48 h). B, Blanche base beer; H, Heraclea (Blanche with the addition of bergamot juice extract, not filtered and not pasteurised); W, Weiss base beer; E, Elais (Weiss with the addition of olive extract, not filtered and not pasteurised).
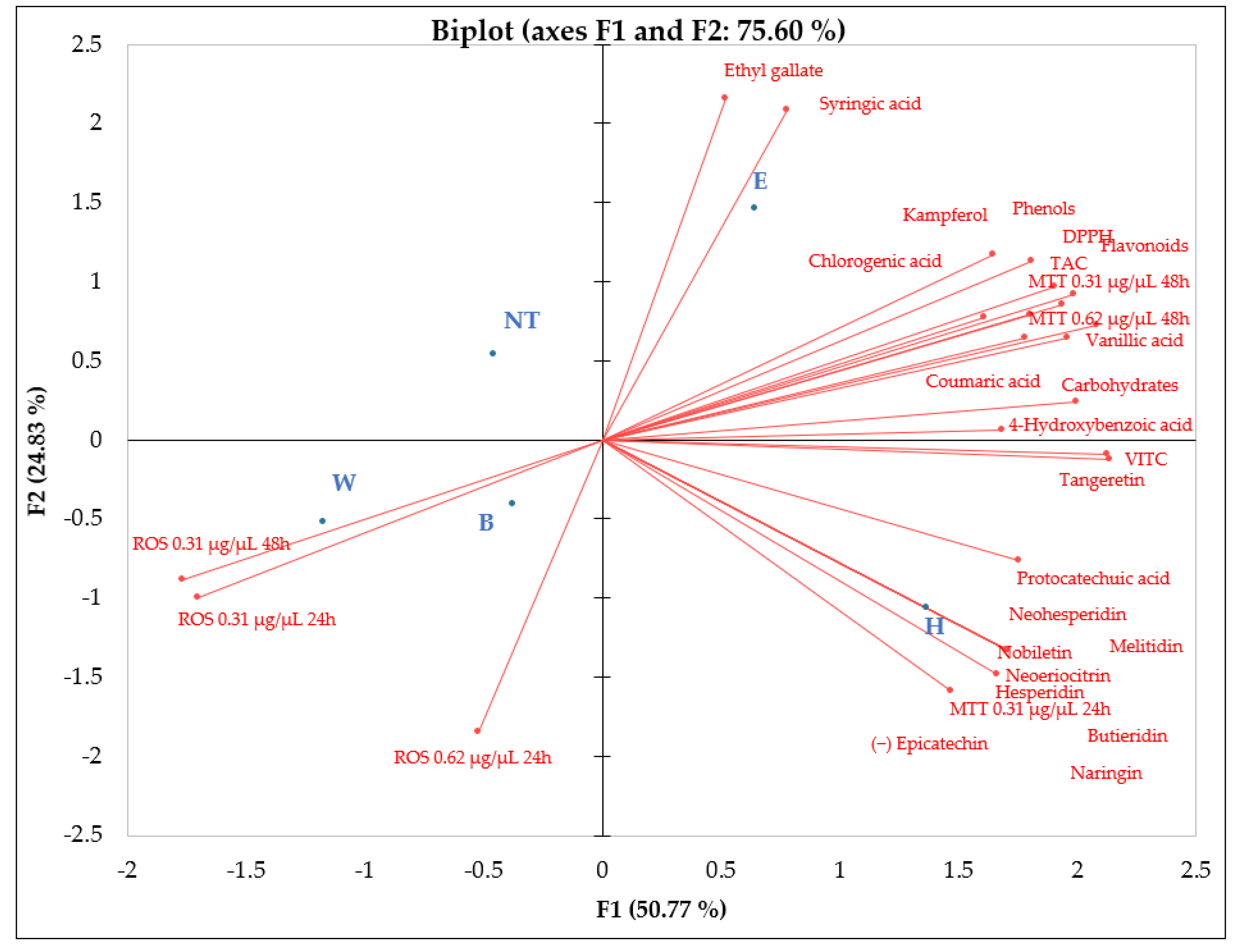
Figure 9.
Correlation matrix (Pearson (n − 1)) of different compounds (total phenols, total flavonoids, phenolic acids, flavonoids and vitamins) contained in the different beers, relate to each other, to reactive oxygen species (ROS) and to cell viability (MTT) at two different concentration Green color and its shades, in the correlation matrix, indicate a positive correlation, signifying that the variables move in the same direction. On the other hand, the red color and its gradients represent an inverse correlation, suggesting that the variables move in the opposite direction.
Figure 9.
Correlation matrix (Pearson (n − 1)) of different compounds (total phenols, total flavonoids, phenolic acids, flavonoids and vitamins) contained in the different beers, relate to each other, to reactive oxygen species (ROS) and to cell viability (MTT) at two different concentration Green color and its shades, in the correlation matrix, indicate a positive correlation, signifying that the variables move in the same direction. On the other hand, the red color and its gradients represent an inverse correlation, suggesting that the variables move in the opposite direction.
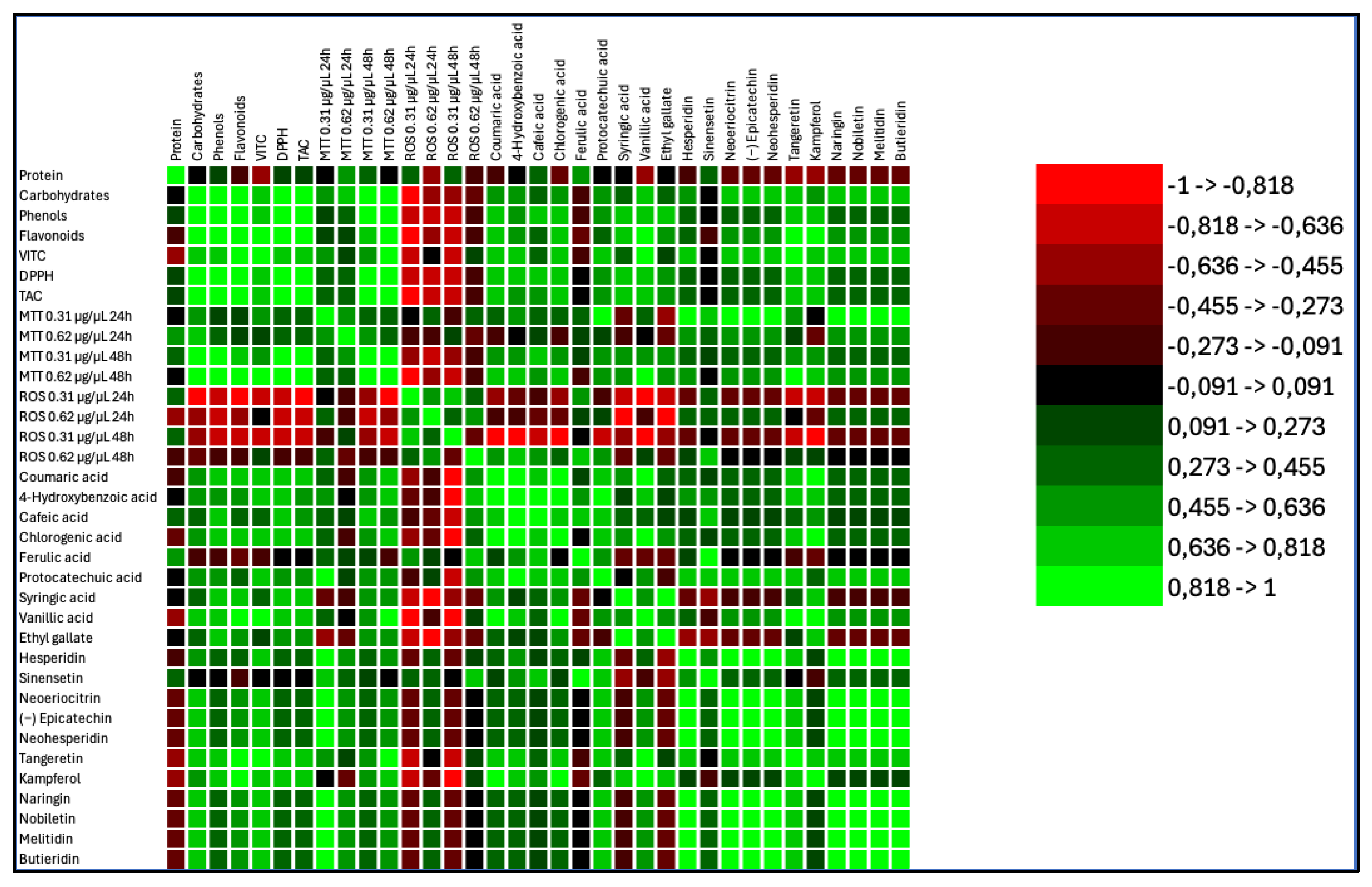

Table 1.
Retention time instrumental recovery and percentage relative standard deviation of polyphenols.
Table 1.
Retention time instrumental recovery and percentage relative standard deviation of polyphenols.
| Compounds | Rt | ±SD | RSD% | Recovery (%) |
|---|---|---|---|---|
| Coumaric acid | 12.396 | 0.008 | 0.496 | 102.1 |
| 4-Hydroxybenzoic acid | 4.060 | 0.014 | 1.310 | 103.8 |
| Caffeic acid | 7.751 | 0.011 | 0.601 | 102.0 |
| Ethylgallate | 9.836 | 0.020 | 1.396 | 102.7 |
| Ferulic acid | 11.323 | 0.007 | 0.258 | 101.9 |
| Kampferol | 18.551 | 0.014 | 0.767 | 100.8 |
| Protocatechuic acid | 2.157 | 0.009 | 0.769 | 102.3 |
| Syringic acid | 8.996 | 0.008 | 0.283 | 97.3 |
| Vanillic acid | 7.344 | 0.015 | 1.172 | 95.9 |
| Chlorogenic acid | 8.637 | 0.015 | 0.837 | 95.9 |
| Naringin | 14.125 | 0.010 | 0.358 | 93.2 |
| Hesperidin | 14.531 | 0.007 | 0.321 | 95.8 |
| Sinensetin | 21.488 | 0.006 | 0.385 | 108.2 |
| Neoeriocitrin | 12.986 | 0.011 | 0.772 | 95.1 |
| (-)Epicatechin | 9.880 | 0.013 | 0.868 | 93.9 |
| Neohesperidin | 14.925 | 0.042 | 0.604 | 100.8 |
| Tangeretin | 23.290 | 0.019 | 0.964 | 104.4 |
| Melitidin | 11.422 | 0.009 | 0.655 | 102.8 |
| Nobiletin | 22.432 | 0.006 | 0.077 | 99.7 |
| Butieridin | 11.611 | 0.011 | 0.101 | 99.6 |
Table 2.
Oligonucleotide sequences for Real-time PCR.
| Gene | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| NRF2 | 5'-AGCCCAGCACATCCAGTCA-3' | 5'-TGTGGGCAACCTGGGAGTAG-3' |
| SOD2 | 5'-CTGGACAAACCTCAGCCCT-3' | 5'-CTGATTTGGACAAGCAGCAA-3' |
| GPX1 | 5'-AGAACGCCAAGAAGCAGAAGA-3' | 5'-CATGAAGTTGGGCTCGAACC-3' |
| GPX4 | 5'-AACTACACTCAGCTGCTGC-3' | 5'-GCAGGTCTTCTCTCATCACC-3' |
| CATA | 5'-TGGAAAGAAGACTCCCATCG-3' | 5'-CCAGAGATCCCAGACCATGT-3' |
| GAPDH | 5'-CAACTTTGGTATCGTGGAAGGAC-3' | 5'-ACAGTCTTCTGGGTGGCAGTG-3' |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



