
Submitted:
04 February 2025
Posted:
04 February 2025
You are already at the latest version
Abstract
Pungent red peppers and chilies are healthy foods or crucial ingredients of modern diets due to their content of bioactive phytochemicals such as carotenoids, tocopherols, capsaicinoids, poly phenols and vitamin C. Production of pungent peppers with outstanding nutritional properties on genetic factors and their interaction with the environments where they are cultivated. This study was conducted to investigate the effects of genotype and climate conditions on the response of capsaicinoids and vitamin C. The extracts of pungent materials and vitamin C were separated and qualitatively determined by HPLC alone or hyphenated with mass spectroscopy. Four genotypes were selected and cultivated under traditional agricultural practices in southern Hungary. The yield properties, and the contents of capsaicinoids and vitamin C were significantly influenced by genetic factors and, to a high extent, by the interaction between genotype x environments. The highest yield of 2.86 ± 0.59 kg/m2 was recorded for the CS variety. The yield was significantly decreased under high precipitation and low air temperature. The highest concentration of capsaicin of 1586-1734 µg/g dwt was found in BHJ variety. The lowest level of 514 µg/g dwt of capsaicin was determined in the CS variety, in which the content significantly increased 772 µg/g dwt with the increase of rainfall and decrease of air temperature in the warmer season 2022. In most cultivars examined capsaicin content negatively responded to change of climate toward higher precipitation and lower temperature except the SZ178, in which the concentration of capsaicinoids stayed insignificantly varied. The amount of vitamin C ranged between 570 and 135 µg/g dwt with the highest content being in the BHJ and the lowest in the REK variety. In most varieties except the REK, vitamin C positively responded to a high and negatively to the reduced levels of precipitation.
Keywords:
capsaicinoids
; vitamin C
; pungent pepper
; chili
; Capsicum sp.
; climate
; genotype
1. Introduction
Plant-based meals rich in bioactive phytochemicals are fundamentally beneficial to human health. Sweet and pungent types of peppers are among the plant-based functional foods or food ingredients due to their considerably high content of bioactive phytochemicals such as carotenoids, tocopherols, poly phenols, capsaicinoids, and vitamin C [1,2,3]. Capsaicinoids are responsible for the pungent flavor of chili and other hot peppers [4]. Capsaicinoids deserve considerable attention for their positive pharmacological effects, but only in the last decades extensive research has been conducted to determine their biological and physiological effects, such as antioxidant [5], anti-inflammatory [6], anticancer [7,8], antibacterial [9], gastric mucosal [10] and fat accumulation prevention [11].The clinical application of capsaicin is hampered by its poor water solubility [12], its high irritant activity, pain and burning sensation on skin and mucous membranes even at low concentrations [13], and its topical application is mostly limited to patches, creams and ointments [14]. Therefore, increasing the solubility and bioavailability of capsaicin has become an important challenge in clinical trials.
Capsaicinoid and vitamin C content has been found to be influenced by genetic and environmental factors [15]. Bae et al. [16] observed highly significant interactions between cultivar, growing season and maturity stage for key bioactive compounds including ascorbic acid, capsaicinoids, flavonoids and phenolics. In a study conducted in northeastern India, Capsicum chinense genotypes had the highest capsaicin content, followed by C. frutescens, Capsicum pubescence, and then C. annuum of different cultivars [17]. The results of the genotypes studied by Souza et al. [18] differed not only in capsaicinoid content but also in the ratio of capsaicin to dihydrocapsaicin.
Vitamin C is an essential micronutrient for humans and its pleiotropic functions are related to its electron donating capacity. It is a powerful antioxidant and cofactor of a family of biosynthetic and gene-regulatory enzymes. It contributes to immune protection by supporting the various cellular functions of the innate and adaptive immune system, as well as supporting epithelial protection against pathogens [19]. Studies by Lekala et al. [20] showed that the interaction of pepper cultivars and growing environmental conditions affected the accumulation of different antioxidant compounds. They found that cultivation in a temperature-controlled foil greenhouse resulted in higher yields and promoted the accumulation of carotenoids, ascorbic acid and vitamin C. Among the pre-harvest factors, light intensity and temperature are the factors most likely to affect final vitamin C content of the crops [21].
Maturity and ripeness have been found to impact on vitamin C and capsaicinoid content of pungent peppers Deepa and co-authors [22], Gnayfeed et al. [23] and Jifon et al. [24] found that genetics, agronomic features, and other factors can change the bioactive qualities of foods. However, little are available on the impact of environmental factors and production practices on the bioactive phytochemicals in pungent or sweet peppers.
The main objective of the present work was to investigate the impact of climate conditions prior to harvest of different four traditional and recently developed varieties of pungent spice peppers using recent approaches in the analysis of vitamin C and capsaicinoids.
2. Materials and Methods
2.1. Experimental Design
In Hungary's Pusztaföldvár (GPS coordinates: 46.535054, 20.800754), open-field, mulch-free growing was the setting for the studies in 2022 and 2023. Chernozem meadow makes up the soil in the area. The results of a soil analysis completed in March 2022 are displayed in the supplemental material (Table S1). Planting was on 13 May in 2022 and 7 May in 2023. The first harvest in 2022 was on 9 September, the second on 20 September. In 2023, the first harvest was on 30 August and the second on 16 September. The four distinct types received the identical treatment in terms of fertilization, irrigation, and plant protection. Drip irrigation was also used to apply nutrients and water supply. Nutrient solution was prepared with Solinure 5 general purpose complex fertilizer, which contains 20% Total nitrogen (N) (4.4% nitrate nitrogen; 3.4% ammonia nitrogen; 12.2% carbamide nitrogen), 20 % phosphorus pentoxide (P2O5), 20.0 % P2O5, 20 % potassium oxide (K2O). Fully water soluble enriched with microelements: 0,01 % boron (B), 0.010 % copper (Cu), 0.06 % iron (Fe), 0.05 % manganese (Mn), 0.005 % molybdenum (Mo), 0.022 % zinc (Zn). Applied from the post-flowering period Haifa MKP fertilizer, which contains 52% P2O5 and 34% K2O. The Trade crop Bentley microelement fertilizer, which contains iron (Fe): 5.9 %, manganese (Mn): 3 %, boron (B): 1.4 %, copper (Cu): 0.47 %, zinc (Zn): 0.7 %, molybdenum (Mo): 0.2 % and cobalt (Co): 0.02 %.was used as a microelement supplement throughout the growing season. As the pepper types are very sensitive to calcium deficiency, Fitohorm Ca 40 calcium solution (N: 12 %, CaO: 21 %) as a foliar fertilizer was applied. Solinure 5 G was applied, as a basic fertilizer, at a dose of 1.5 kg/1000l. Trade Corp Bentley was used as a microelement supplement in an amount of 50g/1000l, as well as 250ml/1000l nitric acid to adjust the pH. During the fruit development period, mono-potassium phosphate (Haifa MKP) was applied as a supplement in the amount of 1 kg/1000 l. The pH of the nutrient solution was on average between 5.5-6, and its EC between 2-3mS/cm. The amount of irrigation was applied according to the needs of the plant and the rainfall. If the average daily temperature is multiplied by 0.2, it gives the daily water requirement in mm, in case of rain-free weather [25]. This amount of irrigation water was between 400 and 600 l for the entire plant population, on occasion.
Four replicates per species and 30 plants per replicate were tested for a total of 480 plants. The row spacing was 90cm and the plant spacing 30 cm. The varieties studied were the Bhut jolokia (Capsicum chinense x Capsicum frutescens) chili pepper, the Cserkó (Capsicum annuum L. var. cerasiforme) cherry pepper, the Szegedi 178 (Capsicum annuum L. var. longum grossum) spice pepper and the Rekord (Capsicum annuum L.) table pepper. All these varieties produce pungent (hot) fruits having red color at biological ripeness.
2.2. Measurements During Vegetation
Daily measurements of solar radiation intensity (Voltcraft LX-10) and rainfall, maximum and minimum temperatures, and solar radiation intensity were made. Every week, the irrigation water, fertilizer solution, pH (Milwaukee PH600), and EC (Adwa AD32) were measured.
2.3. Preparation of samples
The fruit was kept in boxes for a week following the two harvests, which occurred at different periods, before being sliced and dried. This step is necessary for spice peppers to reach the over-ripeness, at which the quality parameters are accomplished. The drying process was performed in a Memmert UF55 drying cabinet for 24 hours at 60°C.The dried samples were vacuum-foiled and milled by a coffee mill immediately before analytical determination to avoid subsequent oxidative deterioration of vitamin C and capsaicinoids.
2.4. Analytical Measurements
For the determination of capsaicinoid content, 0.5 g of well homogenized ground powder from each sample was weighed into an Erlenmeyer flask and 50 ml of HPLC grade methanol were added. The flasks were placed in a 300rpm rotating mechanical shaker (GFL 3005) for 20 min followed by a 5-min ultrasound sonication using Raya (Spain) water bath ultrasonic device. The samples were then filtered through a 0.45µm, Whatman no.1 filter paper (Dassel, Germany). The filtered extracts were 10 times diluted with HPLC grade methanol and further cleaned up by passing through Chromofil hydrophobic PTFE syringe filter before injection into HPLC instrument.
The capsainoid extracts were separated to their individual compounds on Purospher Star C18, 2.7µ, 125mm reversed phase column (Merck Life and Science Ltd, Budapest, Hungary) with isocratic elution of 48:52 water-acetonitrile at a flow rate of 0.7 ml/min according to a previous work [26]. Capsaicinoids were detected by fluorescence detector at Ex:285nm and Em: 320 nm. The affluents were identified by comparing their retention time and total ion from a previously described HPLC-MS/MS protocol [26]. Quantification was based on calibration of standard nor-dihydrocapsaicin, capsaicin, and dihydrocapsaicin from Sigma-Aldrich via Merckgroup Life and Science Ltd, Budapest, Hungary.
For the determination of vitamin C content, to 0.5 g of ground powder of each sample 30 ml of cooled 3% metaphosphoric acid were added in a stoppered flask. The mixtures were shaken at 300 rpm by rotating mechanical shaker (GFL 3005) for 20 min. The mixtures were then transferred to centrifuge tubes and centrifuged at 5000 g for 5 min. The supernatants were taken and cleaned up by passing through a 0.22 µ, 25 mm nylon syringe filter before injection into the HPLC instrument.
Separation of L-ascorbic acid (vitamin C) from other organic acid and polar phenolic compounds was performed on Nautilus C18 aqua, 3µ, 150mm column (Machery Nagel, Darmstadt, Germany) with gradient elution of acetonitrile in 0.01Mpotasium dihydrogen phosphate buffer at a flow rate of 0.7 ml/min. Detection was carried out at 244nm using diode array detector (DAD). L-ascorbic acid C was identified by comparing retention time and spectral characteristic of the sample peak with those of standard material (Sgma-Aldrich, via Merckgroup Life Science Ltd, Budapest, Hungary), from which standard curve was made for quantification.
A Hitachi Chromaster instrument consisting of a Model 5440 diode-array detector, a Model 5210 autosampler, a Model 5440 Fluorescence detector and a Model 5110 gradient pump was used. The instrument was operated, and the chromatograms were evaluated using EZChrome Elite Software.
LC-MS/MS analysis was performed on an Agilent 1100 HPLC equipped with a degasser (G1322A), a binary pump (G1312A), an auto sampler (G1313A), a column thermostat (G1316A) and a fluorescent detector (G1314A). The mass spectrometry detection was carried out an Applied Biosytems API 2000 triple quadrupole. Data acquisition and evaluation was performed using Analyst software. Positive electrospray ion source was used with the following parameters: IS 5000, CUR 20, TEM 550, GS1 60, GS2 70, CAD 7. Nitrogen was used as collision gas for collision-induced dissociation (CID), curtain, nebulizing and drying gas. The product ion spectrums were acquired at 25 and 70 eV from 30 to 600 m/z according to Daood and co-workers [26]
2.5. Statistical Evaluation
All analyses were performed using R version 4.2.1 (RStudio Team, 2020). Capsaicinoids and C-vitamins concentrations were assessed across years, harvests, and varieties via General Linear Model (GLM), with treatment efficacy evaluated using Wilks' lambda (λ). Homogeneity of variances was tested with the Box test, and residual normality was confirmed through Shapiro-Wilk and Kolmogorov-Smirnov tests. Graph values show means ± SD. Partial eta-squared (η²) was used to determine effect size, indicating the probability of detecting a significant effect, ranging from 0 (independence) to 1 (deterministic relationship).
3. Results and Discussion
3.1. Temperature and Precipitation in the Two Years Under Study
The two cultivation seasons under study varied substantially in the amount of precipitation and temperature. The average temperatures for the two months of June and September showed significant differences (P<0.05) (Figure 1 and Figure 2). In 2022. the number of days, at which the maximum temperature exceeded 40°C and the average temperature exceeded 30°C was substantially higher than what measured in 2023, particularly 3 weeks before the first and second harvest at each season. The longer sunshine period in 2022 manifested itself in higher intensity of solar irradiation (Figure 3), which may influence the photosynthetic activity of the plants.
As regards precipitation, the least amount (a total of 154mm) between May and September of the growing season of 2022 was measured, while in 2023 there was a total precipitation of 437 mm at the same period (Figure 4). However, in the last month of cultivation, the amount of precipitation was significantly higher in 2022 than in 2023.
3.2. Properties of Pepper Fruits Yield
Table 1 summarizes the changes took place on number of fruits/plant and the average weight of fruit as a function of changes in climate conditions in the two cultivation seasons. The different genotypes varied significantly in the yield properties as well as in the response of the yield to changes in climate conditions. The highest yield was found in CS variety followed by REK and the standard pungent variety SZ178.
In the context of the response to climate factors, it was indicated that the high temperature and lower precipitation amount in 2022 caused a marked increase in the number of fruit/ plant and fruit weight as compared to those properties of fruits harvested in 2023, in which the precipitation and air temperature were opposite. An exception was that the number of fruit / plants of the BHJ variety positively responded to low temperature and high precipitation during the season of 2023. The obtained results demonstrate that the response of pepper fruits is influenced by the interaction between genetic and agroclimatic factors. The better properties, in terms of number and weight of fruits in warmer cultivation season may be attributed the higher solar radiation intensity (from long sunshine period), that has been reported to be beyond better yield (in terms of size, length and weight of fruit) of pungent habanero produced in the open fields as compared to those measure in fruits of the same genotype produced inside greenhouse [27]
3.3. Impact of Climate Conditions on Capsaicinoids
The HPLC protocol used for the determination and identification of capsaicinoids gave excellent separation of 8 capsaicin-derived compounds that contributed to the pungency of such crop (Figure 5). The detected and identified capsaicinoids are nor-nor-dihydrocapsaicin (NNDC), nor-dihydrocapsaicin (NDC), capsaicin (CAP), dihydrocapsaicin (DC), homo-capsaicin (HCAP), isomer of DC (iDC), homo-dihydrocapsaicin-1 (HDC-1) and homo-dihydrocapsaicin-2 (HDC-2). The HPLC profile of capsaicinoids is like that found for most Hungarian pungent spice red peppers [26] and some Italian genotypes Gioffrida et al. 2014) [28]. As a results of genetic factor, the different pungent peppers showed significant variation in the content and proportion of the individual and total capsaicinoids.
The concentration of the dominant CAPS and DC in BHJ, CS and SZ178 is close to that found in highly pungent cultivars such as Corneto sottile and Numex cultivated in Italy in two different locations [29]. In terms of Scoville Hotness Unit (SHU) a range between 36292 and 40293 was obtained for the three cultivars mentioned before. The REK cultivar has a SHU of 16740 that makes such cultivar as a moderately pungent pepper. Usama et al [30] reported similar concentration for the sum of CAPS and DC contents in AVPP0002 and AVPP0805 varieties cultivated in Bangladesh, and Shams et al. [31] determined slightly higher contents for the same capsaicinoids in Maras and Habanero varieties than that determined in REK genotype in the present study. Schmidt et al. [32] studied, among others, Jalapeño (C. annum) and Bhut Jolokia (C. chinense) peppers grown in Austria and found CAP and DC content of the fruit placenta and seeds to be 296 ± 27.3 µg/g and 233 ± 18.3 µg/g dry base, respectively. The concentration of the major capsaicinoids achieved in our study is much lower than that reported for extremely pungent varieties such as Naga King chili, which distributed 2.54 ± 0.58, 0.50 ± 0.17 and 0.02 ± 0.005 g/100 g dry matter of CAPS, DC, and NDC respectively [33]. Duah et al. [34] and Souza et al. [18] investigated the capsaicinoid content in different hybrids including the highly pungent Unijol. None of the varieties investigated in this work showed as high content of all capsaicinoidsas that reported for Unijol.
Table 2 shows the differences between harvest dates and genotypes in the content of the six most important capsaicinoids found in the examined varieties in 2022. The examined types of Capsicum species differed significantly in their capsaicinoid content as well as in the proportion (%) of everyone particularly in that of the major compounds. As regards the content, the REK variety contained the least amount of all capsaicinoids, particularly CAPS, in the season of 2022, while the highest content of CAPS was recorded for BHJ (chili pepper) in both 1st and 2nd harvest. As for DC, the significantly highest level was found in SZ178 followed by CS in both harvesting times of the same season.
Since the climate factors of the 2nd harvest in 2022 substantially altered from those prior to the 1st one significant variation in the response of the genotypes to climate changes in the two harvesting time was expected (Table 3). A positive response to climate change of CAPS was noticeable in all genotypes investigated, except REK, which showed negative response. In case of NDC and DC, their concentrations tended to increase, but only in REK and SZ178. The minor capsaicinoid isomers in CS and BHJ showed opposite response to that exhibited by CAPS and DC as the climate changed towards the 2nd harvest, while in REC and SZ178 genotypes the minors showed similar response to that of the majors. The first interesting and remarkable alteration was that CAPS concentration extremely dropped in all genotypes tested except in the SZ178 variety, in which CAPS was interestingly stable (unchanged). As regards response of DC, the genotypes showed an interesting variation. In the variety CS, dramatic decrease took place in the content of the isomer DC (decreased from 1133.67 ± 83.22 and 1171.11 ± 103.56 ug/g dry base in 2022 to 289.00 ± 32.97 and 267.50 ± 63.36 ug/g dry base in 2023 at the 1st and 2nd harvest respectively. On contrast, in BHJ and REK varieties, DC responded negatively to the climate changes, and in SZ178 it stayed unchanged. The hydrated capsaicin (HCAPS) in BHJ and CS varieties responded negatively (decreased) as the weather changed in2023, whereas, in REC and SZ-178 its concentration increased, in the later it was 2 folds higher than that in 2022. The changes in the levels of the homo-derivatives were not clearly understood due to their being variable intermediates.
In 2022, the temperature dropped from 42°C before the 1st harvest to 23°C prior at the 2nd harvest, while precipitation increased from 41mm to 67mm (Figure 5). The change in such parameter was opposite in 2023, when the temperature slightly decreased from 32°C to 29°C, and the precipitation dramatically dropped from 87mm to 18mm. Therefore, there was a considerable variation in the content and response of capsaicinoids between the genotypes cultivated and harvested in 2022 and 2023. In 2022, a significant change in the major capsaicinoids was found between the two harvests only in CS variety, while in the others the levels stayed not significantly changed. HCAPS response to change in climate condition was genotype dependent. In BHJ and CS, the content of the capsaicin isomer significantly (P < 0.05-0.01) decreased toward the 2nd harvest with that in BHJ being dramatic. In REC and SZ178, there was a slightly significant increase in the content of HCAS. As concerns HDC-1, its response was not understood in all genotypes examined, whereas the climate change was favorable and had positive impact (significant at P < 0.01) on the content of HDC-2 isomer in all genotypes studied.
In the cultivation year of 2023, the climate factors prior to harvesting time were characterized by relatively higher temperature (29°C) and extremely lower precipitation (18 mm) as compared to those recorded in 2022. The response of capsaicinoids to climate condition few weeks before harvests was influenced mostly by the genetic factors. For instance, NDC in BHJ and SZ178 showed decreasing tendency, while REK exhibited significant increase (P < 0.01) as a response to high temperature and low precipitation. The response of the major compounds (CAPS and DC) was negative in BHJ and CS varieties, and positive in REK.
The effect of seasonal variation as a factor had a strong significant impact on capsaicinoids content (F(6,43) = 292,38; P < 0.001; Wilk's λ = 0.024; partial η2 = 0.97). Similarly, the differences between harvests (F(6,43) = 21,035; P < 0.001; Wilk's λ = 0.25; partial η2 = 0.74) and between varieties were also significant (F(18,122) = 103.59; P < 0.001; Wilk's λ = 0.00037; partial η2 = 0.92). The greatest effects were observed for the differences between years and varieties, with both having the highest partial η2 value.
It seems that the response of pungent materials in peppers to the environment is affected by the interaction between the impacts of two abiotic factors (air temperature and precipitation). The air temperature has been found to influence the expression of capsaicinoid biosynthesis genes in various peppers. A study by González-Zamora et al. [35] on the effect of air temperature on the capsaicinoid content in various varieties, it has been found that with rising temperatures the levels of CAPS and DC increased. Yang and co-workers [36] reviewed several studies, in which high temperature led to increase the content of capsaicinoids in many but not all hot pepper genotypes. Additionally, Naves et al. [37] emphasized the positive effect of high air temperature on the accumulation of capsaicinoids for some varieties, while in other varieties the high temperature had negative impact.
The air temperature in the cultivation location associates with the sunshine period and the intensity of radiation coming from sunlight. Although the effect of light intensities on capsaicinoid content has been demonstrated to be inconsistent, many studies indicated that optional light intensity is required to regulate biosynthesis of capsaicinoids and increase their content in chili peppers [38,39]. Furthermore, exposure to elevated light intensity has been reported to increase significantly the total capsaicinoids in wild genotypes [40]. In the present study, the high concentration of CAPS and DC in 2022 when the maximum temperature before the 1st harvest exceeded 40°C for some days together with high radiation intensity supported the afore-mentioned facts. The other climate variable that may stand beyond the low content of capsaicinoids in 2023 is the high amount of precipitation. It has been demonstrated that in some cases excessive watering or high precipitation with certain genotypes of hot peppers may lead to a reduced capsaicinoid content [37]. Other studies indicated that water deficit or mild drought increased the activity of the enzymes involved in the regulation and biosynthesis of capsaicinoids, while the over watering had the opposite impact on such enzymes [41,42].
One of the analytical parameters that assists in a better understanding of the changes in capsaicinoids as a matter of genetic variation and as a response to climate changes is the ratio of CAPS to DC. Table 4 shows the effects of harvesting time, climate condition and genotype on the CAPS/DC ratio. The highest values (2.4) for the ratio were recorded for BHJ in both cultivation years and harvest. Like such ratio has been reported by Tupoz and Ozdemir [43] for some Capsicum annum cultivars (1245F1, amazon F1, Serademre 8, and Kusak 295 F1) cultivated in Turky. The highest value achieved for the ratio in BHJ is close to the values determined in most conventional varieties of pungent spice red pepper genotypes widely cultivated in Hungary [26]. This revealed that the recently developed BHJ cultivar of Capsicum annum might originate from one traditional Hungarian variety via breeding-based biotechnological protocol. The values determined by Duelund et al. [44] for proportion of CAPS and DC in hot peppers in Danmark were slightly lower than what we found for BHJ. The values found for other varieties ranged between 0.97 and 1.04 are close to those found for some accessions of hot peppers harvested at green or fully ripe stages [45].
The seasonal variation in climate parameters caused the ratio to decrease in BHJ variety from 2.4 in 2022 to 1.08 in 2023. The change in the climate between 1st and 2nd harvest in 2022 did not result in a significant change in the ratio in all genotypes studied, while in 2023 the climate conditions of the 2nd harvest caused the ratio to significantly increase (P < 0.05) in all genotypes examined. The change in the CAP/DC associates rather with the level of precipitation prior to harvesting times than with the changes in the air temperature. Such finding supported the fact that high water supply diminishes the activity of the enzymes regulating biosynthesis of capsaicinods in certain landraces of hot peppers or chilies. Of the enzymes of the biosynthesis of capsaicinods hydrogenases catalyze the synthesis of dihydro derivatives of capsaicin [31].
The drastic dropping in the level of precipitation before the 2nd harvest of 2023 caused the ratio to increase significantly as compared to the 1st harvest conforming the positive effect of mild drought (low water supply) on the activity of the enzymes catalyzing capsaicin synthesis (capsaicin synthase) and diminishes, to some extent, the activity of dihydrocapsain synthase.
3.4. Content and Response of Vitamins C
The recent analytical protocol applied to determine L-ascorbic acid, the abundant biological form of vitamin C in the nature, allowed for efficient resolution of the vitamin from the accompanying organic acids and water-soluble phenolics (Supplement Fig.2).
Figure 6 shows the vitamin C content of pungent peppers as affected by genetic factor, seasonal variation, and climate conditions before harvesting times. The results showed that BHJ variety had the highest vitamin C content (570 mg/100 g dry base) in 2022 as compared to other varieties tested. The lowest concentration of 130 mg /100 g dry base was recorded for REK. The range found between the highest and the lowest concentration determined for the 4 hot peppers under study is in the range reported for vitamin C in several varieties cultivated in Italy [29]. The obtained range of vitamin C in all varieties examined is much higher than that reported by Tupoz and Ozdimer [43] for some cultivars of Capsicum annuum cultivated in Turkey. Furthermore, Kantar et al. [46] studied the vitamin C content of different types of pepper cultivars including some Hungarian-bred varieties like BHJ, Fehérözön and Szegedi óriás, which when cultivated in North America, contained less vitamin C than the varieties investigated in the present work. In another study [47], chili and spice hot pepper contained between 90 and 130 mg/100g vitamin C that is close to the range measured for the 4 varieties of our study. Nagy et al. [48] determined 248.8 and 368.4 mg/ 100g dry matter for Beibiehong and Fire Flame cultivars of chili pepper respectively, which is less than that found in all varieties examined.in this work.
The range between 130 and 570 mg/100 g of dry matter (18-80 mg/100 g fresh weight) may contribute to 20-26% to 88-114% of RDI for men and women respectively. According to Yuni et al. [49] and Olatunji et al. [50] the recent RDI for vitamin C in men and women is 90 and 75 mg/day, respectively.
As concerns the impact of climate conditions at harvesting time, in all genotypes, it was found that there was a significant increase in the concentration of the vitamin as the precipitation significantly increased (P < 0.01). At the 2nd harvest of the first cultivation season particularly with CS variety, in which the increment was approximately 3 folds making the level of vitamin C in such variety to rank second after that of vitamin C in BHJ. In 2023 when the precipitation decreased at the 2nd harvest the vitamin C content tended to significantly decrease for all genotypes under study except REK, in which the level of the vitamin surprisingly increased almost twofold. Such findings confirm that biosynthesis of vitamin C in peppers with a high pungency is significantly affected by increasing water supply via precipitation or irrigation, while in cultivars with low or medium pungency the reduced water supply or mild drought promotes regulation and accumulation of vitamin C. The positive correlation between water supply and content of vitamin C doesn’t agree with inverse relation between water supply and vitamin C level reported previously by Duah et al. [51]. As a matter of fact, the increasing amount of rainfall leads to arise in the moisture content of the soil that has been stated to correlate positively with accumulation of vitamin C in chili peppers [52]
The exposure to high sunlight intensity in 2022, in which the air temperature exceeded 40°C for some days might assist in increasing the biochemical factors that regulate vitamin C synthesis. It has been found that high air temperature caused by high radiation intensity increases the content of vitamin C in several cultivars of sweet peppers [53]
5. Conclusions
In the present study, it was found that the content variation and response to environmental factors of capsaicinoids and vitamin C in chili and pungent spice peppers is associated with the effect of genotype in addition to the significant effect of interaction between genotype and environment (G x E). The findings achieved could highlight how the genotype is ruling aspect to the climate conditions in the variation of capsaicinoid and vitamin C content. It could be concluded that the response of phytochemical in hot peppers to the changes in the climate factors such as high precipitation, high air temperature and intensive radiation of sunlight would be positive or negative depending on the genotype adaptation to the changes in the environmental conditions. Accordingly, the BHJ and CS varieties are recommended as crucial sources of capsaicinoids and vitamin C due to their high adaptation to climate conditions. Also concluded that all varieties examined are a potential source of capsaicinoids and can contribute efficiently to the recommended dietary intake of vitamin C, and good raw materials for pharmaceutical industries even when subjected to unexpected changes in the climate conditions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Ingrid M. Gyalai and Flórián Kovács: Analysis, Methodology, Data curation, Hussein. G. Daood: Conceptualization, Writing original draft, Correspondence, Adrienn Szarvas: Investigation, Statistical Analysis. Lajos Helyes: Validation, Supervision, Ferenc Lantos: supervision, data curtation.
Funding
No funding given to this research work.
Data Availability Statement
All of the data are incorporated in the manuscript.
Acknowledgments
This work was supported by the Flagship Research Groups Programme (2024-2027) of the Hungarian University of Agriculture and Life Sciences.
Conflicts of Interest
The authors declare that no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| BHJ | Bhut jolokia variety |
| CS | Cserkő variety |
| REK | Rekord variety |
| SZ178 | Szegedi178 variety |
| RWI | Recommended Dietary Intake |
References
- Dludla, P.V.; Ilenia Cirilli, I.; Marcheggiani, F.; Silvestri, S.; Orlando, P.; Muvhulawa, N.; Meltdown, M.T.; Nkambule, B.B.; Mazibuko-Mbeje, S.E.; Nokulunga Hlengwa, N.; Hanser, S.; Duduzile Ndwandwe, D.; Jeanine, L.; Marnewick, J.L.; Albertus, K.; Basson, A.K.; Luca Tiano, L. Bioactive Properties, Bioavailability Profiles, and Clinical Evidence of the Potential Benefits of Black Pepper (Piper nigrum) and Red Pepper (Capsicum annum) against Diverse Metabolic Complications. Molecules 2023, 28, 6569. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Pérez, T.; Gómez-García, M.D.R.; Valverde, M.E.; Paredes-López, O. Capsicum annuum (hot pepper): An ancient Latin-American crop with outstanding bioactive compounds and nutraceutical potential. A review. Comp. Rev. Food Sci. Food Safe. 2020, 19, 2972–2993. [Google Scholar] [CrossRef]
- de Sá Mendes, N.; de Andrade Gonçalves, É.C.B. The role of bioactive components found in peppers. Trends Food Sci. Technol. 2020, 99, 229–243. [Google Scholar] [CrossRef]
- Reyes-Escogido, M.L.; Gonzalez-Mondragon, E.G.; Vazquez-Tzompantzi, E. Chemical and Pharmacological Aspects of Capsaicin. Molecules 2011, 16, 1253–1270. [Google Scholar] [CrossRef]
- Chen, K.S.; Chen, P.N.; Hsieh, Y.S.; Lin, C.Y.; Lee, Y.H.; Chu, S.C. Capsaicin protects endothelial cells and macrophage against oxidized low-density lipoproteininduced injury by direct antioxidant action. Chem-Biol. Interact. 2015, 228, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Kawada, T.; Kim, B.S.; Han, I.S.; Choe, S.Y.; Kurata, T.; Yu, R. Capsaicin exhibits anti-inflammatory property by inhibiting IkB-a degradation in LPS-stimulated peritoneal macrophages. Cell. Signal. 2003, 15, 299–306. [Google Scholar] [CrossRef]
- Lin, C.H.; Lu, W.C.; Wang, C.W.; Chan, Y.C.; Chen, M.K. Capsaicin induces cell cycle arrest and apoptosis in human KB cancer cells. BMC Complem. Altern. M. 2013, 13, 1–9. [Google Scholar] [CrossRef]
- Meral, O.; Alpay, M.; Kismali, G.; Kosova, F.; Cakir, D.U.; Pekcan, M.; Yigit, S.; Sel, T. Capsaicin inhibits cell proliferation by cytochrome c release in gastric cancer cells. Tumor Biol. 2014, 35, 6485–6492. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yu, L.; Li, F.; Zhang, G.; Zhou, W.; Jiang, X. Synthesis of amide derivatives containing capsaicin and their antioxidant and antibacterial activities. J. Food Bioch. 2019, 43, e13061. [Google Scholar] [CrossRef]
- Park, J.S.; Choi, M.A.; Kim, B.S.; Han, I.S.; Kurata, T.; Yu, R. Capsaicin protects against ethanol-induced oxidative injury in the gastric mucosa of rats. Life Sci. 2000, 67, 3087–3093. [Google Scholar] [CrossRef]
- Lee, G.R.; Shin, M.K.; Yoon, D.J.; Kim, A.R.; Yu, R.; Park, N.H.; Han, I.S. Topical application of capsaicin reduces visceral adipose fat by affecting adipokine levels in high-fat diet-induced obese mice. Obesity 2013, 21, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Al-Samydai, A.; Aburjai, T.; Alshaer, W.; Azzam, H.; Al-Mamoori, F. Qualitative and Quantitative Analysis of Capsaicin from Capsicum annum Grown in Jordan. Intern. J. Res. Pharm. Sci. 2019. 10, 3768–3774. [CrossRef]
- Bode, A.M.; Dong, Z. The two faces of capsaicin. Cancer Res. 2011, 71, 2809–2814. [Google Scholar] [CrossRef]
- Mou, J.; Paillard, F.; Turnbull, B.; Trudeau, J.; Stoker, M.; Katz, N.P. Efficacy of Qutenza (capsaicin) 8 % patch for neuropathic pain: A meta-analysis of the Qutenza Clinical Trials Database. Pain 2013, 154, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Orellana-Escobedo, L.; Garcia-Amezquita, L.E.; Olivas, G.I.; Ornelas-Paz, J.J.; Sepulveda, D.R. Capsaicinoids content and proximate composition of Mexican chili peppers (Capsicum spp.) cultivated in the State of Chihuahua. CyTA-J. Food 2013, 11, 179–184. [Google Scholar] [CrossRef]
- Bae, H.; Jayaprakash, G.K.; Crosby, K.; Yoo, K.S.; Leskovar, D.I.; Jifon, J.; Patil, B.S. Ascorbic acid, capsaicinoid, and flavonoid aglycone concentrations as a function of fruit maturity stage in greenhouse-grown peppers. J. Food Compos. Anal. 2014, 33, 195–202. [Google Scholar] [CrossRef]
- Dubey, R.K.; Singh, V.; Upadhyay, G.; Pandey, A.K.; Prakash, D. Assessment of phytochemical composition and antioxidant potential in some indigenous chilli genotypes from Northeast India. Food Chem. 2015, 188, 119–125. [Google Scholar] [CrossRef]
- Souza, C.S.; Daood, H.G.; Agyemang, S.D.; Vinogradov, S.; Palotás, G.; Neményi, A.; Helyes, L.; Pék, Z. Stability of carotenoids, carotenoid esters, tocopherols and capsaicinoids in new chili pepper hybrids during natural and thermal drying. LWT – Food Sci. Techno 2022, 163, 1–9. [Google Scholar] [CrossRef]
- Anitra, C.C.; Maggini, S. Vitamin C and immune function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef]
- Lekala, C.S.; Saber, M.K.H.; Phan, A.D.T.; Maboko, M.M.; Fotouo, H.; Soundy, P.; Sultanbawa, Y.; Sivakumar, D. Cultivar-specific responses in red sweet peppers grown under shade nets and controlled-temperature plastic tunnel environment on antioxidant constituents at harvest. Food Chem. 2019, 275, 85–94. [Google Scholar] [CrossRef]
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharv. Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef]
- Deepa, N.; Kaur, C.; George, B.; Singh, B.; Kapoor, H.C. Antioxidant constituents in some sweet pepper (Capsicum annuum L.) genotypes during maturity. LWT- Food Sci. Technol. 2007, 40, 121–129. [Google Scholar] [CrossRef]
- Gnayfeed, M.H.; Daood, H.G.; Biacs, P.; Alcaraz, F. Content of bio-active compounds in pungent spice paprika as affected by ripening and genotype. J. Sci. Fd Agric. 2001, 81, 1580–1585. [Google Scholar] [CrossRef]
- Jifon, J.L.; Laster, G.; Stewart, M.; Crosby, C.; Leskovar, D.I. Fertilizer use and functional quality fruits and vegetables. Chapter 9: Fertilizer Use and Human Health. In: Proceedings of the Conference Name, Conference Location. The International Fertilizer Industry Association (IFA) and the International Plant Nutrition Institute (IPNI), 2012, 21-24.
- Helyes, L. A paradicsom termesztése (Tomato cultivation); Mezőgazda Kiadó: Budapest, 2000; pp. 75–76. ISBN 9789630053280. [Google Scholar]
- Daood, H.G.; Halasz, G.; Palotás, G.; Palotás, G.; Bodai, Z.; Helyes, L. HPLC Determination of Capsaicinoids with cross-linked C18 column and buffer-free eluent.). J. Chromatogr. Sci. 2015, 53, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Camposeco-Montejo, N.; Flores-Naveda, A.; Ruiz-Torres, N.; Álvarez-Vázquez, P.; Niño-Medina, G.; Xochitl Ruelas-Chacón, X.; Torres-Tapia, M.M.; Rodríguez-Salinas, P.; Villanueva-Coronado, V.; Josué, I.; García-López, G. Agronomic Performance, Capsaicinoids, Polyphenols and Antioxidant Capacity in Genotypes of Habanero Pepper Grown in the Southeast of Coahuila, Mexico. Horticulturae 2021, 7, 327. [Google Scholar] [CrossRef]
- Giuffrida, D.; Dungo, P.; Torre, G.; Bignardi, C.; Cavazza, A.; Corradini, C.; Dugo, G. Evaluatioon of carotenoid and capsaicinoid content in powder of red chili peppers during on year of storage. Food Res. Inter. 2014, 65, 163–170. [Google Scholar] [CrossRef]
- Tripodi, P.; Cardi, T.; Blanchi, G.; Migliori, C.A.; Schiavi, M.; Rotino, G.L.; Scalzo, R.L. Genetic and environmal factors unferlying variation in yield performance and bioactive compound content of hot pepper varieties (Capsicum annuum) cultivated in two contrasing Italian locations. Eur. Food Res. Technol. 2018, 244, 1555–1567. [Google Scholar] [CrossRef]
- Usama, M.G.; Rafii, M.Y.; Ismail, M.R.; Abdul Malek, M.; Abdul Latif, M. Capsaicin and dihydrocapsaicin determination in chili pepper genotypes using ultra-fast liquid chromatography. Molecules 2014, 19, 6474–6488. [Google Scholar] [CrossRef]
- Shams, M.; Yuksel, E.A.; Agar, G.; Ekinci, M.; Kul, R.; Turan, M.; Yildirin, E. Biosynthesis of capsaicioids in pungent peppers under salinity stress. Physiol. Plantar. 2023, 175, e13889. [Google Scholar] [CrossRef]
- Schmidt, A.; Fiechter, G.; Fritz, E.-M.; Mayer, H.K. Quantitation of capsaicinoids in different chilies from Austria by a novel UHPLC method. Journal of Food Compos. Analy. 2017, 60, 32–37. [Google Scholar] [CrossRef]
- Ananthan, R.; Subhash, K.; Longvah, T. Capsaicinoids, amino acid and fatty acid profiles in different fruit components of the world hottest Naga king chilli (Capsicum chinense Jacq). Food Chem. 2018, 238, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Duah, A.S.; Souza, C.S.; Daood, H.G.; Pék, Z.; Neményi, A.; Helyes, L. Content and response to Ɣ-irradiation before over-ripening of capsaicinoid, carotenoid, and tocopherol in new hybrids of spice chili peppers. LWT–Food Sci. Technol. 2021, 147, 111555. [Google Scholar] [CrossRef]
- González-Zamora, A.; Sierra-Campos, E.; Luna-Ortega, J.G.; Pérez-Morales, R.; Ortiz, J.C.; Garcia-Hornández, J.L. Characterization of different Capsicum varieties by evaluation of their capsaicinoids content by high performance liquid chromatography, detection of pungency and effect of high temperature. Molecules 2013, 18, 13471–13486. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, C.; Ye, Q.; Liu, C.; Wan, H.; Ruan, M.; Zhou, G.; Wang, R.; Li, Z.; Diao, M.; Cheng, Y. The influence of different factors on the metabolism of capsaicinoids in pepper (Capsicum annum L.). Plants 2024, 13, 2887. [Google Scholar] [CrossRef] [PubMed]
- Navez, E.R.; de Ávil Silva, L.; Sulpice, R.; Araújo, W.L.; Nunes-Nesi, A.; Peres, L.E.P.; Zsögön, A. Capsaicinoids: Pungency beyond Capsicum. Trend in Plant Science 2019, 24, 2. [Google Scholar] [CrossRef]
- Jeeatid, N.; Techawongstien, S.; Suriham, B.; Bosland, P.W.; Techawongstien, S. Light intensity affects capsaicinoids accumulation in hot peppers (Capsicum Chinese Jscq) Cultgivars. Hortic. Environ. Biotechnol. 2017, 58, 103–110. [Google Scholar] [CrossRef]
- Mawarni Saputri, M.; Qory Oktaria, Q.; Alnia Junaidi, A.; Agis Ardiansyah, M.A. The effect of light intensity and sound intensity on the growth of various types of chili in indoor system. J. Pen. Pend. IPA 2023, 8, 6330–6336. [Google Scholar] [CrossRef]
- Gurung, T.; Techawongstien, S.; Suriharn, B.; Techawongstien, S. Impact of environments on the accumulation of capsaicinoids in Capsicumm spp. HortScience 2011, 461, 1576–1581. [Google Scholar] [CrossRef]
- Sung, Y.; Chang, Y.Y.; Ni-lun, T. Capsaicin biosynthesis in water stressed hot pepper fruits. Bot. Bull. Acad. Sin 2005, 46, 35–42. [Google Scholar]
- Phimohan, P.; et al. Impact of drought stress on the accumulation of capsaicinoids in Capsicum cultivarswith different initial capsaicinoid levels. HortScience 2012, 47, 1204–1208. [Google Scholar] [CrossRef]
- Tupoz, A.; Ozdemir, F. Assessment of carotenoids, capsaicinoidsand ascorbic acid composition of some selected pepper cultivars (Capsicum annuum L) grown in Turkey. J. Food Comp. Anal. 2009, 20, 596–602. [Google Scholar] [CrossRef]
- Duelund, L.; Mouritsen, O.G. Contents of capsaicinoids in chilies grown in Denmark. Food Chem. 2017, 221, 913–918. [Google Scholar] [CrossRef]
- Guijarro-Real, C.; Adalid-Martinez, A.M.; Pires, C.K.; Ribes-Moya, A.M.; Fita, A.; Rodriguez-Burruezo, A. The effect of the varietal, ripening stage, and growing conditions in Capsicum peppers. Plants 2023, 12, 231. [Google Scholar] [CrossRef] [PubMed]
- Kantar, M.B.; Anderson, J.E.; Lucht, S.A.; Mercer, K.; Bernau, V.; Case, K.A.; Le, N.C.; Frederiksen, M.K.; DeKeyser, H.C.; Wong, Z.-Z.; Hastings, J.C.; Baumler, D.J. Vitamin Variation in Capsicum Spp. Provides Opportunities to Improve Nutritional Value of Human Diets. PLoS ONE 2016, 11, e0161464. [Google Scholar] [CrossRef] [PubMed]
- Agostini-Costa, S.T.; Silva Gomes, I.; Melo, L.A.M.P.; Becker Reifschneider, F.J.; Costa Ribeiro, C.S. Carotenoid and total vitamin C content of peppers from selected Brazilian cultivars. J. of Food Compos. Anal. 2017, 57, 73–79. [Google Scholar] [CrossRef]
- Nagy, Z.; Daood, H.G.; Ambrózy, Z.; Helyes, L. Determination of Polyphenols, Capsaicinoids, and Vitamin C in New Hybrids of Chili Peppers. J. Analyt. Meth. Chem. 2015, 5, 102125. [Google Scholar] [CrossRef]
- Yuni, W.; Ana-Rosa, B.; Enny, S.; Raoul, B.J.; Arnaud, B.G. Secondary metabolites of Capsicum species and their importance in the human diet. J. Nat. Prod. 2013, 76, 783–793. [Google Scholar]
- Olatunji, T.L.; Afolayan, A.J. The suitability of chili pepper (Capsicum annuum L.) for alleviating human micronutrient dietary deficiencies: A review. Food Sci Nutr. 2018, 6, 2239–2251. [Google Scholar] [CrossRef]
- Duah, A.S.; Souza, S.C.; Nagy, Z.; Pék, Z.; Neményi, A.; Daood, H.G.; Szergej Vinogradov, S.; Helyes, L. Effect of Water Supply on Physiological Response and Phytonutrient Composition of Chili Peppers. Water 2021, 13, 1284. [Google Scholar] [CrossRef]
- Mahendran, S.; Bandara, D.C. Effects of Soil Moisture Stress at Different Growth Stages on Vitamin C, Capsaicin and P-Carotene Contents of Chilli (Capsicum annuum L.) Fruits and their Impact on Yield. Tropical Agricultural Research 2000, 12, 95–106. [Google Scholar]
- Selahle, K.M.; Sivakumar, D.; Jifon, J.; Soundy, P. Postharvest response of red and yellow sweet peppers grown under photo-selective nets. Food Chem. 2015, 173, 951–956. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Air temperatures measured during the season 2022 in the experiment location.
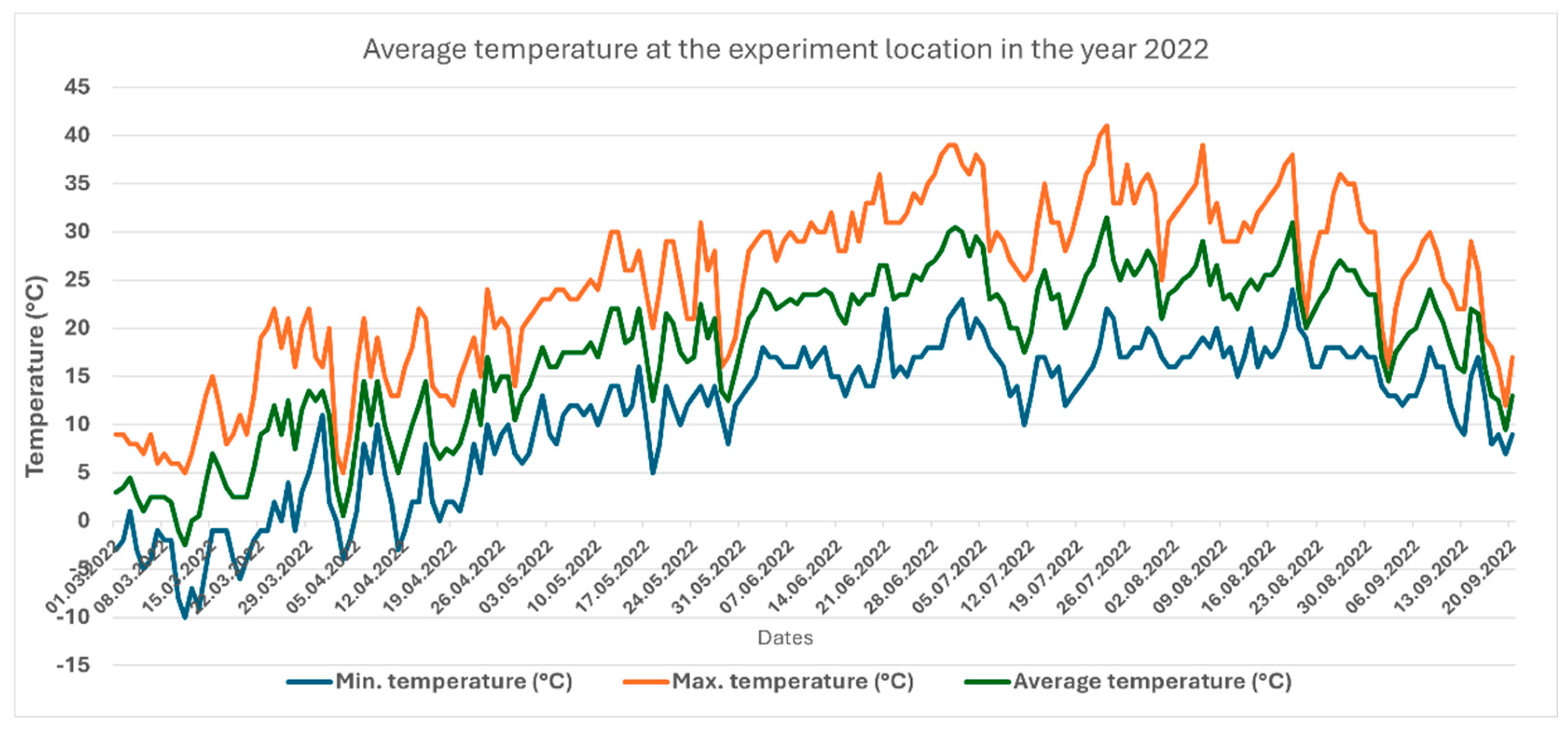
Figure 2.
Air temperatures measured during the season 2023 in the experiment location.
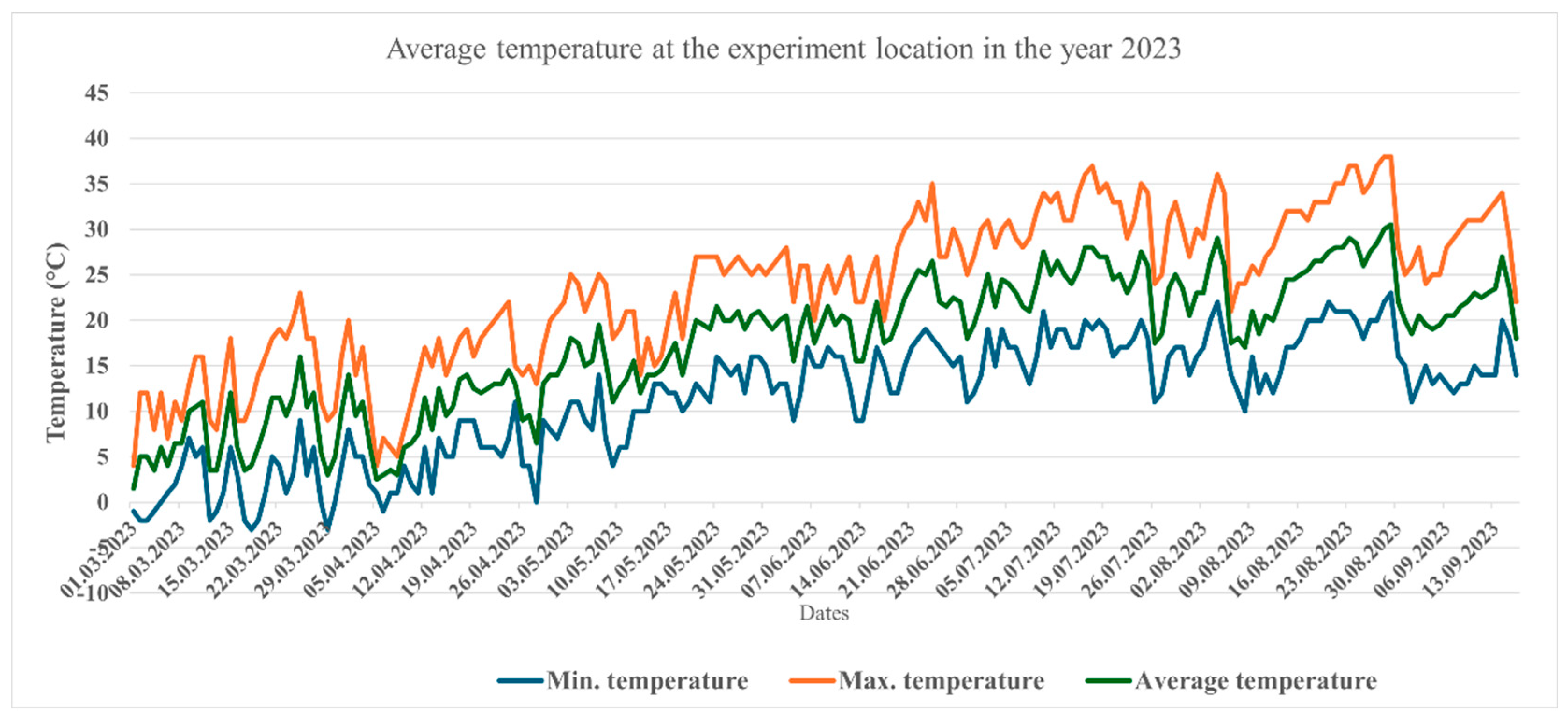
Figure 3.
Solar radiation measured between July and September in2022 and 2023 in the experiment location.
Figure 3.
Solar radiation measured between July and September in2022 and 2023 in the experiment location.
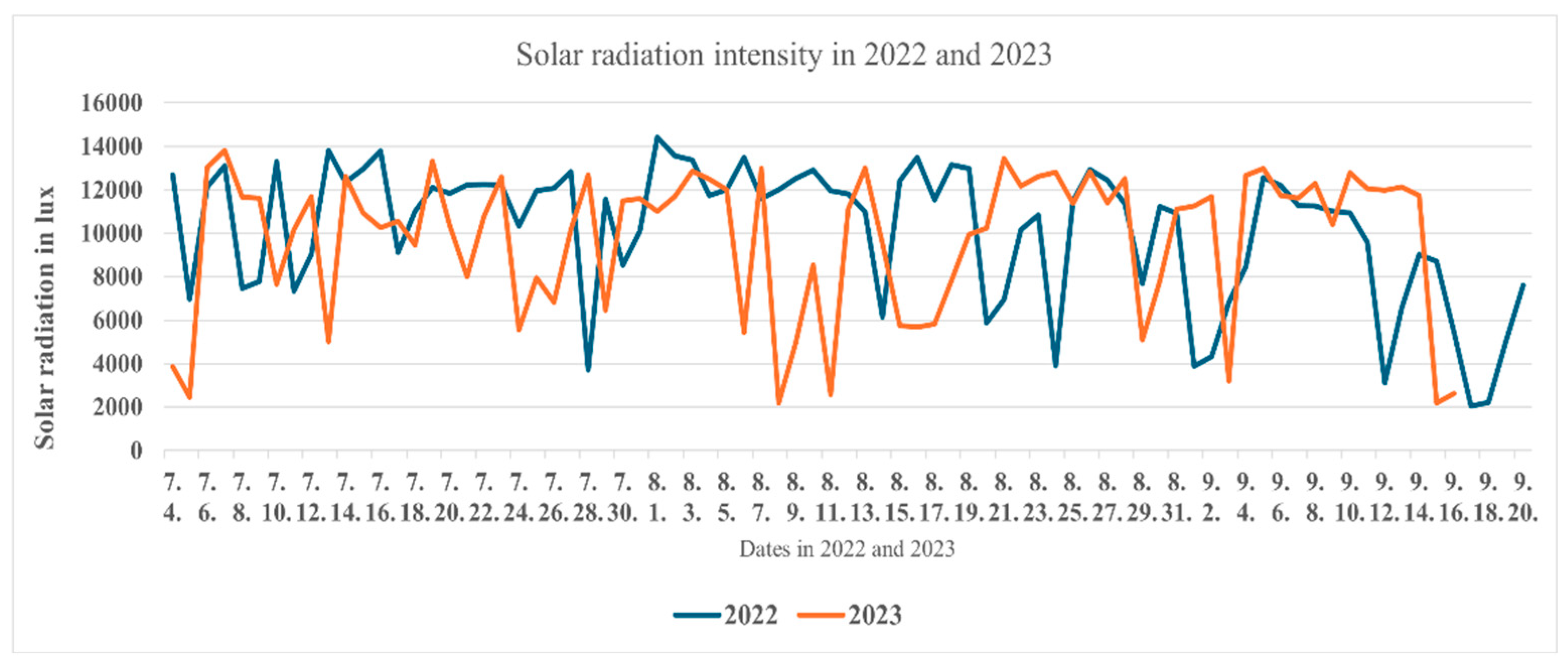
Figure 4.
Amounts of rainfall (mm) measured between March and September in2022 and 2023 in the experiment location.
Figure 4.
Amounts of rainfall (mm) measured between March and September in2022 and 2023 in the experiment location.
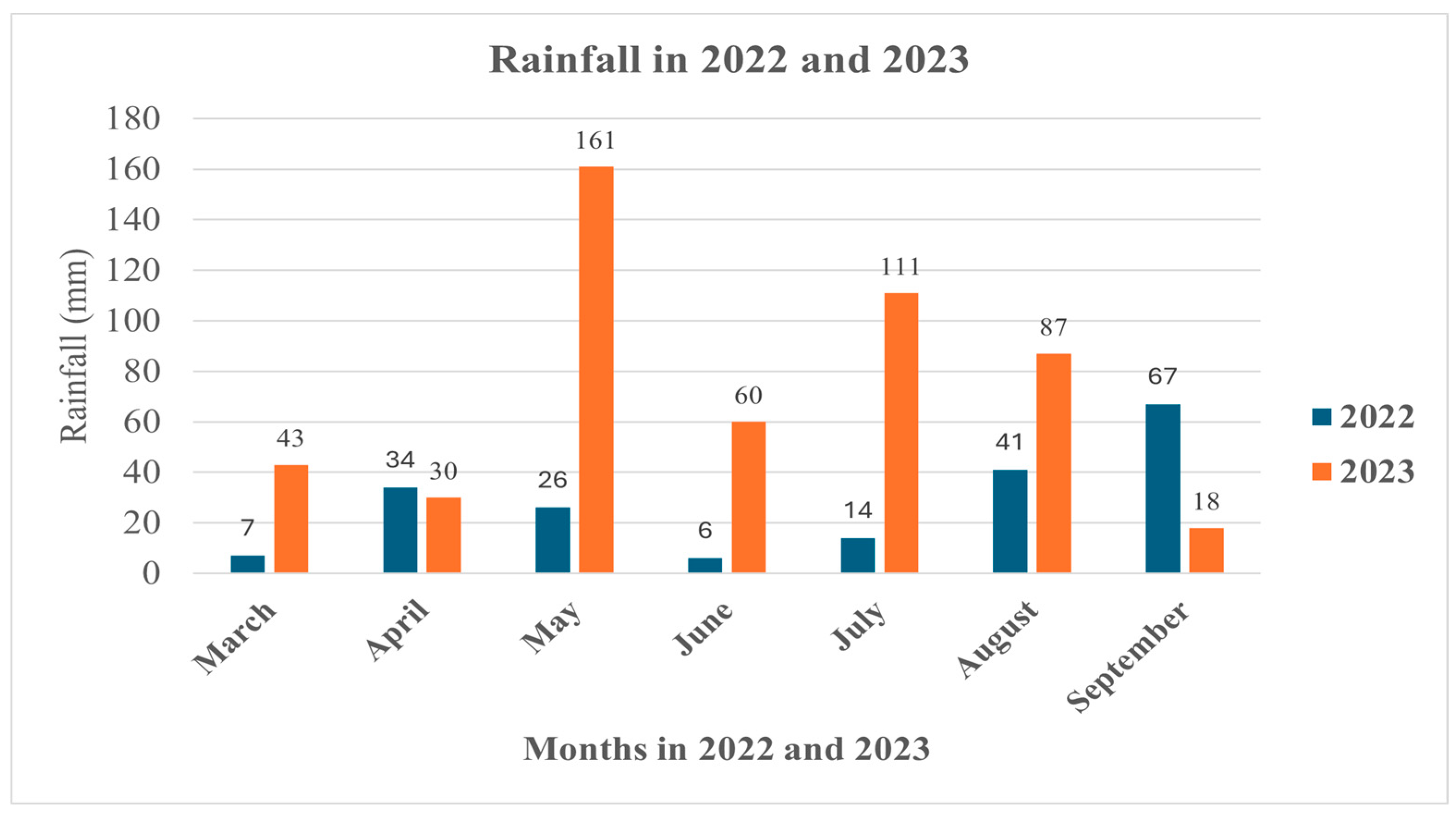
Figure 5.
HPLC profie of capsaicinoids separated on Purospher Star column with isocratic elution of wate-acetonitrile.
Figure 5.
HPLC profie of capsaicinoids separated on Purospher Star column with isocratic elution of wate-acetonitrile.
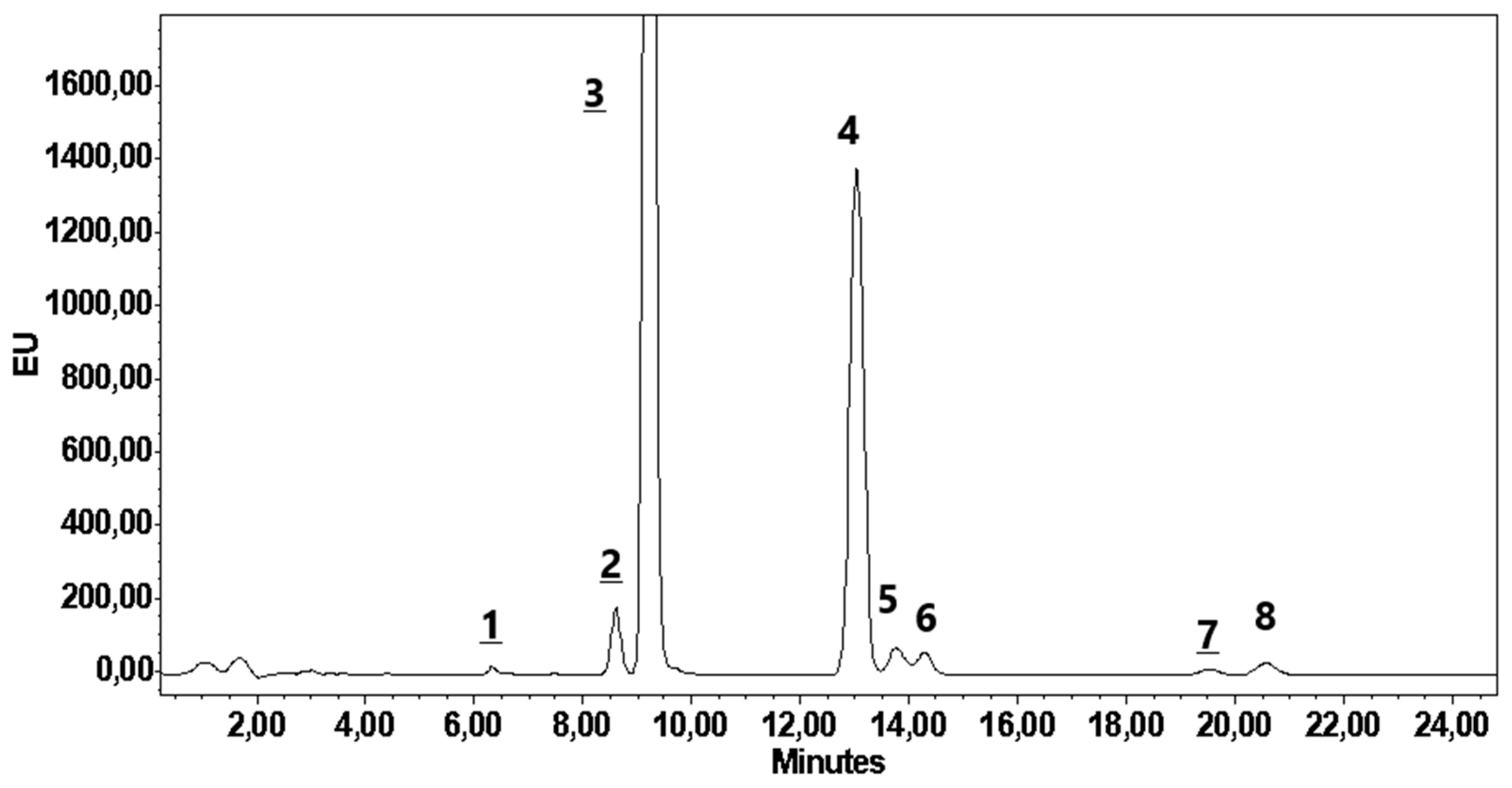

Table 1.
Average fruit number and fruit weight and yield for different genotypes in 2022-2023.
| Year | Variety | Number of fruits/plants | weight (g/fruit) Yield kg/m2 |
|
| 2022 | Bhut jolokia | 18 | 8,7 | 0.579 ± 0.19 AXa |
| 2022 | Cserkó | 24 | 32,3 | 2.86 ± 0.59 AXb |
| 2022 | Szegedi 178 | 17 | 19,8 | 1.24 ± 0.27 AXc |
| 2022 | Rekord | 15 | 22 | 1.21 ± 0.14 AXc |
| 2023 | Bhut jolokia | 22 | 9 | 0.73 ± 0.11AXa |
| 2023 | Cserkó | 21 | 28,2 | 2.19 ± 0.39 AXd |
| 2023 | Szegedi 178 | 15 | 19,1 | 1.06 ± 0.21 AXc |
| 2023 | Rekord | 13 | 15,8 | 0.75 ± 0.12 BXa |
upper case letters (AB) show differences between varieties between harvests; lower case letters indicate differences between varieties in the same years. Means within a column followed by the same letter are not significantly different (Tukey’s HSD test, P < 0.05).
Table 2.
Content of capsaicinoids as affected by genotype and climate conditions at harvesting times in 2022.
Table 2.
Content of capsaicinoids as affected by genotype and climate conditions at harvesting times in 2022.
| Harvest | Variety | N DC | CAPS | DC | HCAPS | HDC-2 |
| 1st | BHJ | 36.11 ± 5.36 AXa | 1586.36 ± 123.35 AXc | 640.13 ± 85.04 AXa | 35.04 ± 4.86 AXb | 5.27 ± 0.52 AXa |
| CS | 246.31 ± 25.12 AXd | 1339.94 ± 73.36 AXbc | 1133.67 ± 83.22 AXb | 9.87 ± 0.90 AXa | 92.25 ± 6.84 AXd | |
| R | 82.39 ± 3.50 AXb | 514.24 ± 38.25 AXa | 522.76 ± 26.40 AXc | 10.37 ± 0.81 AXa | 29.41 ± 1.26 AXb | |
| SZ178 | 195.02 ± 22.33 AXc | 1200.74 ± 124.39 AXb | 1263.53 ± 123.30 AXb | 30.33 ± 4.12 AXb | 65.38 ± 6.30 AXc | |
| 2nd | BHJ | 37.88 ± 1.10 AXa | 1734.01 ± 44.41 AXc | 646.98 ± 7.70 AXa | 10.05 ± 1.08 AYa | 10.16 ± 0.37 BXa |
| CS | 268.84 ± 16.28 AXc | 1319.94 ± 145.89 AXb | 1171.11 ± 103.56 AXb | 8.28 ± 0.70 AYa | 104.98 ± 6.78AXd | |
| R | 129.41 ± 21.22 AXb | 771.99 ± 147.49 AXab | 744.30 ± 125.36 AXab | 14.15 ± 2.10 AXa | 43.10 ± 5.67 BYb | |
| SZ178 | 239.06 ± 32.32 AXc | 1401.93 ± 132.62 AXb | 1448.39 ± 161.53 AXc | 37.91 ± 5.04 AXb | 81.01 ± 10.60 AXc |
Higher case letters show differences between harvest; lower case letters indicate differences between varieties in the same years. Means the same letter are not significantly different (Tukey’s HSD test, P < 0.05.
Table 3.
Content of capsaicinoids as affected by genotype and climate conditions at harvesting times in 2022.
Table 3.
Content of capsaicinoids as affected by genotype and climate conditions at harvesting times in 2022.
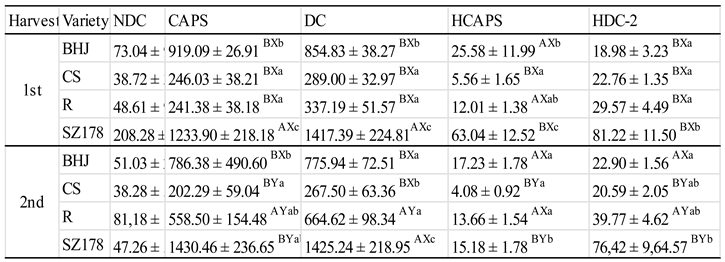 |
Each value represents the mean ± standard deviation (SD) of n=4. Different: upper case letters (AB) show differences between harvests; lower case letters indicate differences between varieties in the same years. Means within a column followed by the same letter are not significantly different (Tukey’s HSD test, P < 0.05).
Table 4.
Caps/DC ratio as affected by genetic and environmental factors 3 weeks before harvess.
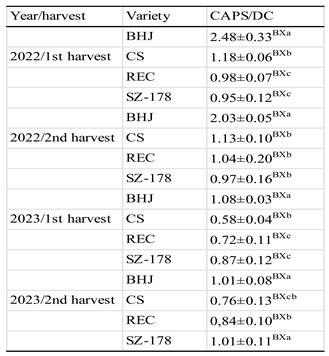 |
Each value represents the mean ± standard deviation (SD) of n=4. Different: upper case letters (AB) show differences between harvests; lower case letters indicate differences between varieties in the same years. Means within a column followed by the same letter are not significantly different (Tukey’s HSD test, P < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



