
Submitted:
02 February 2025
Posted:
03 February 2025
You are already at the latest version
Abstract
Efficient farm practices are crucial for livestock health and performance, and cold stress is a major challenge for cattle in winter. This study aimed to investigate the effects of cold and warm water in winter on growth performance, thermal stress indicators, serum hormones and metabolites, rumen fermentation parameters, rumen fiber degrading enzyme and rumen microflora of yellow cattle during winter. Eight Wandong Bulls (Bos taurus) were divided into two groups: group C, which received cold water (6.36 ± 1.99 °C) and group E, which received warm water (32.00 ± 3.12 °C) for 30 d. Results showed that warm-water intake significantly increased ADG (p = 0.024) and DMI (p = 0.046), and decreased (p = 0.047) the ratio of feed intake to weight gain. Furthermore, the heat production, respiratory rate, surface temperature and rectal temperature of cattle did not alter varying with water temperature, but the heat production value of bulls increased (29.64 vs 25.76 MJ/W0.75 h-1) by cold-water intake compared to warm water. The concentrations of thyroxine (p = 0.021), serum urea nitrogen (p = 0.025), and glucose (p = 0.011) increased with bulls drinking cold water compared to warm water. The concentrations of NH3-N (p = 0.048), total VFA (p = 0.010), acetate (p = 0.009), propionate (p = 0.009), cellulase (p < 0.01) and xylanase (p < 0.05) were lower in group C compared to group E. The total bacterial abundance, as well as specific species including Ruminococcus flavus, Ruminococcus albus, and Prevotella ruminicola, were lower (p < 0.05) in group C compared to group E. In conclusion, in comparison with cold water, drinking warm water during the winter enhanced growth performance by influencing energy metabolism, regulating serum hormones and metabolites, and modulating ruminal microflora of bulls.
Keywords:
Energy metabolism
; Growth performance
; Rumen microbiota
; Wandong Bulls
; Water temperature
1. Introduction
The cattle industry is becoming increasingly important in providing meat and dairy products to meet the demands of a growing global population[1]. However, cattle production is influenced by a variety of factors, including environmental conditions, forage availability, genetics, feeding practices, and disease[2,3,4]. Among these, temperature is a key factor that impacts animal health, productivity, and gut microbiota composition[5,6]. Cold stress, in particular, is a temperature-related factor that can lead to a range of health and performance issues. Castellani et al. emphasized that cold exposure adversely affects animal physiology[7]. In mice, cold stress exacerbates lipopolysaccharide-induced intestinal damage, leading to dysbiosis in the gut microbiome [8]. Similarly, in rat offspring, cold stress has been linked to increased anxiety levels [9]. For dairy cows and beef cattle, cold stress not only decreases productivity but also raises their susceptibility to diseases[10]. Wang et al. reported that long-term cold stress decreased growth performance, nutrient digestion and rumen volatile fatty acids (VFA) production[11]. These findings underscore the need for effective environmental management strategies to protect animals from the harmful effects of cold stress.
The microbiota refers to the diverse community of microbes inhabiting the intestinal tract, including bacteria, protozoa, archaea, viruses, and fungi [12]. Often referred to as the "second genome," these microorganisms play vital roles in digestion, metabolism, immunity, and maintaining the gut barrier [13]. The composition of this dynamic microbiota is influenced by a variety of factors, including genetics, nutrition, and environmental conditions[14]. Disruptions to this balance have been linked to a range of diseases, such as inflammatory bowel disease[15], diarrhea[16], and liver disorders[17]. It is reported that water temperature can affect the rumen temperature, and the lower the water temperature, the longer the rumen recovery temperature[18]. In fact, the effect of water temperature on rumen may be more direct due to the direct contact of water with rumen fluid and rumen microorganisms compared with temperature stress. In many regions of China, winters can be harsh, and continuous access to warm water in cowsheds is often unavailable. This is particularly common on smaller farms, where cattle are typically provided with cold water during the winter months. As a result, cattle production and health may be negatively impacted by the cold water. However, there is limited research on how water temperature influences the microbiota of Wandong Bulls during these cold winter months. Therefore, we hypothesized that drinking cold water would reduce growth performance, rumen fermentation, fiber enzyme activity, and microbial function in cattle, while increasing thermoregulation, energy metabolism, and heat production compared to drinking warm water. The objective of this study was to compare the growth performance, thermal stress indicators, serum hormones and metabolites, ruminal fermentation characteristics, fiber enzyme activity, and microbiota of bulls drinking warm versus cold water during the winter months, in order to explore an often overlooked aspect of animal health in response to environmental conditions.
2. Materials and Methods
The animals were treated as approved by the Humane Animal Care and Ethics Committee of the Nanjing Agricultural University, China (SYXK2011-0036).
2.1. Experiment Design
Eight Wandong Bulls (Bos taurus) (initial body weight of 365.45 ± 22.90 kg) in this study were kept on a local cattle farm in Chuzhou, China, and were divided into two groups: group C (cold water) and group E (warm water). In December, when the local temperature was averaged 5.74 ± 2.63 °C, bulls in group C received cold water at 6.36 ± 1.99 °C, while bulls in group E were given warm water at 32.00 ± 3.12 °C. The bulls were individually housed in pens with free access to water and were fed ad libitum a total mixed ration (TMR) with a concentrate-to-forage ratio of 1:9. Diet composition and nutrient levels were shown in Table 1. Cold water is the conventional drinking water for bulls exposed to low temperatures in winter, and actual water temperature is 5.74 ± 2.63 °C. Warm water is supplied by a constant temperature water tank equipped with a temperature control and detection system, and the temperature is set to 30 °C (actual water temperature is 32.00 ± 3.12 °C). The study included a 7-d adaptation period, followed by a 30-d data and sample collection period.
2.2. Data and Sample Collection
The initial and final weights of each bull were recorded before morning feeding to calculate the average daily gain (ADG). The dry matter intake (DMI) of each bull was recorded every day and the ratio of feed intake to weight gain (F:G ratio) was calculated according to DMI and weight gain. Heat production in bulls was measured by a 80 × 80 × 136 cm respiratory metabolic apparatus at d 25, 26 and 27 of collection period, and was calculated according to the formula described by McLean et al[19]:
Heat production = -0.2047 × ∆F(O2) × V × 60 / kg0.75
Where “∆F(O2)” is oxygen concentration difference between intake and exhaust, “V” is exhaust volume per unit time of the breathing chamber under standard conditions. The respiratory rate of cattle was recorded through the breathing cage. Bull surface temperature at different ambient temperature (5-9, 9-11 and 14-16 ℃) was analyzed using hand-held infrared thermometer (59 Mini, Fluke®, USA) base on the weighted average of the body surface temperature of different parts of the bull and the proportion of the skin area of the part to the total body area. Rectal temperature was measured using a hand-held bovine rectal thermometer (29110, Jinan Borui agriculture and animal husbandry products Co., Ltd., China) inserted 15 cm into the anus of bull daily.
Blood samples were collected from the jugular vein of each bull using vacutainer tubes (gel & clot activator) before morning feeding on day 30 of the collection period. The collected samples were then centrifuged at 3,000 rpm for 15 minutes at 4°C to separate the serum, which was stored at -20°C until further analysis.
Rumen fluid samples were collected using a rumen catheter through the mouth on d 28, 29 and 30 of collection period. Fresh rumen fluid samples were measured pH value using pH meter (PHS-3E, INESA Scientific Instrument Co., Ltd., Shanghai, China). The rumen fluid samples were divided into 3 fractions; two portions were mixed with either 25% HPO3 (v/v) or 1% H2SO4 (v/v) and stored at -20 ℃ until laboratory analysis; and the 3rd portion was collected into cryotubes, immediately frozen in liquid nitrogen, and stored at -80 °C for DNA extraction.
2.3. Laboratory Analysis
2.3.1. Nutritional Composition
The samples of TMR were analyzed for dry matter (method 930.15), crude protein (method 990.03), ether extract (method 920.39) and ash (method 942.05) as outlined by AOAC (2005)[20]. The content of organic matter in the TMR is calculated by the difference between 100 % and ash. The contents of NDF (heat-stable α-amylase-treated) and ADF in TMR were determined according to the method described by Van Soest et al. (1991)[21].
2.3.2. Serum Indices
The concentration of triiodothyronine, growth hormone, blood urea nitrogen, glucose and total protein in serum were determined using commercial ELISA kits (Feiya Biology Co., Ltd., Jiangsu, China), and the serum thyroxine concentration was analyzed using commercial ELISA kit (Beijing Rigor Bioscience Development Co., Ltd., Beijing, China). The specific operation steps are performed according to the manufacturer’s instructions. The absorbance of the reaction was measured by a microplate reader (Multiskan Go, Thermo Fisher Scientific Oy, Vantaa, Finland) and the serum hormones and metabolites concentrations were calculated according to the formula on the instructions.
2.3.3. Ruminal Fermentation Characteristics and Cellulases
VFAs in rumen fluid samples from both group C and group E were analyzed using gas chromatography (Shimadzu GC-4800A, Kyoto, Japan) equipped with a 30 m × 0.32 mm × 0.25 mm capillary column and flame ionization detection. The temperatures of injector oven, column oven and detector were set at 230, 160 and 240 ℃, respectively. The NH3-N concentration in the rumen fluid was measured according to the indophenol colorimetric method described by Broderick and Kang (1980)[22]. The rumen fluid was centrifuged at 12000 ×g for 20 min, and 40 μL of the supernatant was mixed with 2.5 mL phenol chromogenic agent and 2 mL hypochlorite solution in turn, and the color was developed in a water bath at 37 ℃ for 30 min. The NH3-N concentration was calculated according to the absorbance of the solution at the wavelength of 550 nm and the standard curve of NH3-N.
The activity of rumen cellulases were measured using bovine-specific cellulase and xylanase ELISA kits provided by Feiya Biology Co., Ltd. and Lvye Biology Co., Ltd. The rumen fluid samples were preliminary treated by ultrasonic disruption to ensure the optimal decomposition of the microbial cell wall to enhance the extraction of enzymes, and then follow the manufacturer's instructions for subsequent operations.
2.3.4. Ruminal Flora Quantitative Analysis
Total bacterial DNA was extracted from rumen fluid samples collected from group C and group E using the APINeasy DNA Kit for feces (Beijing ZEPING Bioscience & Technology Co., Ltd, China). The purity of the extracted DNA was assessed through agarose gel electrophoresis (1.5%), as recommended in a previous study[23]. The concentration of the DNA products was measured using a UV spectrophotometer (NanoDrop 2000, Thermo Scientific, USA). All samples were adjusted to a standardized concentration of 50 ng/µL for subsequent analyses.
To quantify bacterial concentrations, real-time PCR was conducted using specific primers targeting total bacteria, Ruminococcus albus (R. albus), Ruminococcus flavus (R. flavus), Butyrivibrio fibrisolvens (B. fibrisolvens), Fibrobacter succinogenes (F. succinogenes), Methanogens, Succinimonas amylolytica (S. amylolytica), Streptococcus bovis (S. bovis), and Prevotella ruminicola (P. ruminicola), as listed in Table 2. Initially, PCR amplification was performed, and the resulting products were purified using the GenElute Gel Recovery Kit (Sigma-Aldrich, USA). Following purification, the concentrations of the PCR products were assessed, and gradual dilutions were prepared to generate standard curves for real-time PCR quantification. Finally, bacterial concentrations were calculated based on the quantitative analysis methods described Li et al. (2014)[24] to provide details of the microbial community and its response to varying water temperatures in the feed of Eastern Anhui Yellow cattle.
2.4. Statistical Analysis
The valid difference analysis between bulls drinking warm and cold water was performed using t-test in SPSS (version 27.0). All results were presented as means ± standard deviation (SD), with statistical significance recognized at p < 0.05, whereas a tendency was considered at 0.05 ≤ p < 0.10.
3. Results
3.1. Growth Performance
The initial body weight (BW) and final BW did not differ between group C and E (Table 3). However, drinking warm water increased ADG (p = 0.024) and DMI (p = 0.046), and decreased (p = 0.047) the F:G ratio of bulls.
3.2. Thermal Stress Indicators
The heat production, respiratory rate, surface temperature and rectal temperature of cattle were not different with varying water temperature.
3.3. Serum Hormones and Metabolites
The concentrations of triiodothyronine (trend, p = 0.058), thyroxine (p = 0.021), blood urea nitrogen (p = 0.025), glucose (p = 0.011) and total protein (trend, p = 0.063) increased in bulls drinking cold water compared to warm water (Table 5).
3.4. Ruminal Fermentation Characteristics and Fiber Enzyme Activity
The rumen pH of bulls decreased (trend, p = 0.061) in group E than group C. The concentrations of NH3-N (p = 0.048), total VFA (p = 0.010), acetate (p = 0.009), propionate (p = 0.009), butyrate (trend, p = 0.066), isobutyrate (trend, p = 0.074) and valerate (trend, p = 0.091) concentrations were lower in group C compared to group E, whereas the A:P ratio was not changed (Table 6).
Bulls drinking warm water significantly increased the concentrations of cellulase (p < 0.01) and xylanase (p < 0.05) in rumen fluid compared to cold water (Figure 1).
3.5. Rumen Bacterial Abundance
Rumen bacterial analysis revealed that the total bacterial abundance, as well as specific species including Ruminococcus flavus (R. flavus), Ruminococcus albus (R. albus), and Prevotella ruminicola (P. ruminicola) were significantly lower (p < 0.05) in group C compared to group E. Conversely, there was no difference between the two groups in the abundances of Fibrobacter succinogenes (F. succinogenes), Methanogens,
4. Discussion
4.1. Growth Performance
Cold temperature is a well-documented stressor that can impair antioxidant activity, immune function, and overall growth performance in animals [25,26]. In this study, we observed that bulls provided with cold water had lower weight gain compared to those given warm water. This finding is consistent with previous studies that show similar effects in beef cattle receiving warmer water during cold seasons[25], as well as in weaned piglets[27] and yaks[28].Together, these finding reinforce the conclusion that cold water have negatively impact on the animal performance, likely due to the increased metabolic demands associated with maintaining body temperature. Moreover, the digestion and absorption of feed in the digestive tract determines feed efficiency, and low feed digestibility may reduce feed efficiency and thus affect weight gain[29]. In our study, bulls drinking warm water during the winter showed enhanced cellulase and xylanase activity, suggesting that warm water may promote feed digestion by boosting cellulase activity in the rumen, thereby improving weight gain and feed efficiency. Additionally, cold stress is known to increase feed intake as the animal works to generate more heat [30]. For example, Kennedy et al. reported that cold exposure increased the DMI of sheep but decreased the digestibility of dry matter and organic matter[31]. In our study, bulls drinking warm water during the winter showed enhanced cellulase and xylanase activity, suggesting that warm water may promote feed digestion by boosting cellulase activity in the rumen, thereby improving weight gain and feed efficiency. In addition, cattle reduced feed intake to reduce heat production to adapt to heat stress[30], and on the contrary, cold stress increased feed intake to increase heat production. Kennedy et al. reported that cold exposure increased the DMI of sheep, but decreased the digestibility of dry matter and organic matter[31]. However, in the current study, dulls drinking cold water reduced the feed intake, which may be due to the fact that cold water reduced the activity of microorganisms, cellulase and xylanase in the rumen of bulls, resulting in a decrease in the digestion rate of feed and increasing the retention time of feed in the rumen, thus reducing the feed intake[32]. Furthermore, Serviento et al. reported the water intake of dairy cows reduced when the ambient temperature was reduced from 17.25 ℃ to 8.27 ℃[33]. In fact, water intake of cattle is positively correlated with DMI, the decrease of water temperature in this study may reduce the water intake, thereby reducing the DMI[34].
4.2. Thermal Stress Indicators
Cold-stimulated ruminants adapt to temperature reduction by increasing heat production and reducing heat dissipation[35]. In this study, cold water intake increased heat production (29.64 vs 25.76 MJ/W0.75 h-1), likely as a result of the body’s efforts to maintain thermal balance. In fact, when temperatures rise, cattle can increase the frequency of their breathing to facilitate heat exchange and help regulate body temperature[36]. Conversely, in response to colder temperatures, cattle reduce their respiratory rate to conserve heat. However, in this study, water temperature did not alter respiratory rate of bulls, which may be due to the limited change in energy exchange between bulls and the environment at the same temperature. Furthermore, the study found that cold water intake did not affect the bulls’ body surface or rectal temperature, which may reflect the final outcome of the body’s heat balance regulation. Although heat production, as well as levels of triiodothyronine, thyroxine, and glucose, increased, these changes likely helped elevate body temperature. However, the heat produced was counterbalanced by the cold water intake, which may have prevented any observable changes in the bulls's body surface or rectal temperature.
4.3. Serum Hormones and Metabolites
Cold-stimulated cattle can trigger changes in the concentrations of corticosterone, triiodothyronine, and thyroxine in the bloodstream through the regulation of the hypothalamic-pituitary-adrenal and hypothalamic-pituitary-thyroid axes. These hormonal changes help maintain body temperature stability and enable the cattle to adapt to environmental temperature fluctuations [37]. In this study, the higher concentrations of triiodothyronine and thyroxine in the serum of bulls drinking cold water suggest that cold water exposure imposed additional cold stress, prompting a physiological response to maintain thermal balance during the winter months.
Glucose, as a vital energy source, plays a crucial role in providing heat through energy metabolism[38]. In this study, bulls drinking cold water in winter had elevated serum glucose concentration. This likely indicates that the cold water stimulated the bulls to release more heat in order to maintain body temperature, which in turn resulted in increased glucose concentration. Wang et al. also reported a significant increase in serum glucose concentrations in beef cattle subjected to long-term cold stress [11].
Serum urea nitrogen and total protein levels reflect the protein balance in cattle[39]. It has been reported that long-term cold stress can increase the secretion of glucocorticoids in cattle, thereby enhancing the decomposition of proteins to obtain energy and leading to an increase in serum urea nitrogen concentration[11,40]. Moreover, the synthesis of cold shock proteins in organisms increased to protect and maintain the normal physiological functions of the body when the ambient temperature decreases[41]. Therefore, the serum concentrations of urea nitrogen and total protein were both increased in bulls drinking cold water compared to warm water.
4.4. Ruminal Fermentation Characteristics and Cellulase
Warm water intake during winter may promote the production of beneficial VFAs such as acetate and propionate, which are microbial byproducts that support energy metabolism and gut health [42,43]. The increase in total VFA concentration in the rumen of bulls drinking warm water could be due to the enhanced activity of rumen microorganisms and feed digestion, which in turn boosts VFA production. In addition, VFA as an important energy substance of the body, may be rapidly consumed to maintain body temperature balance, while the intake of cold water consumes the body's heat to maintain body temperature, resulting in a decrease in total volatile acid in the rumen fluid of bulls fed with cold water. Acetate is primarily produced by anaerobic bacteria such as Bacteroides spp and Akkermansia muciniphila[44], which contributes to energy and lipid metabolism, immunity, and stress resilience[45]. Propionate is mostly generated through carbohydrate metabolism, supports lipid metabolism, mitochondrial function, and immune health[46,47]. The higher concentrations of acetate and propionate in bulls drinking warm water further indicate that higher water temperature may facilitate lipid metabolism to enhance resilience and growth performance during cold period. NH3-N is the product of protein degradation in the rumen, which is affected by many factors such as dietary nitrogen content, protein degradation rate, microbial synthesis efficiency, rumen wall absorption and energy level. The intake of warm water in this study increased the NH3-N concentration in rumen fluid, which may be due to the fact that warm water enhanced the activity of proteolytic bacteria such as P. ruminicola in the rumen and promoted the digestion of protein in feed [48].
Cellulase and xylanase, two key enzymes produced by microbes, play crucial roles in converting lignocellulose into animal feed or fermentable sugars for ethanol production, with widespread applications in food, medicine, and agriculture[49,50]. In this study, bulls provided with warm water showed significantly higher activity of cellulase and xylanase, consistent with findings in beef cattle[25]. In fact, cellulase and xylanase in the rumen are metabolism products of bacteria such as F. succinogenes, R. albus, and R. flavefaciens, and their concentrations are closely related to the activity of rumen microorganisms[51]. He et al. reported that feeding cold water to beef cattle in winter reduced the activity of cellulase and xylanase in the rumen, which was consistent with this study[25]. Current study showed that bulls provided with warm water exhibited a significantly higher abundance of total bacteria, B. fibrisolvens, R. albus, R. flavefaciens, and P. ruminicola, which may contribute to the observed increase in rumen cellulase and xylanase activity.
4.5. Rumen Bacterial Abundance
Microorganisms play a crucial role in feed digestion and overall animal health [52]. B. fibrisolvens, R. albus, R. flavefaciens, and P. ruminicola are known ruminal fiber-degrading bacteria that produce digestive enzymes to break down fiber and contribute to feed digestion in the rumen. Brod et al. reported that the lower (30 to 0 ℃) the intake water temperature of sheep led to a longer time (72 to 108 min) for the rumen to return to normal temperature[18]. Similar studies reported that cold water intake significantly reduced rumen temperature compared to warm water intake[53]. In the present study, cold water intake decreased the abundance of rumen microorganisms, likely due to reduced microbial activity caused by lower rumen temperature[25]. In addition, B. fibrisolvens is a prominent rumen bacterium that ferments polysaccharides, assisting in cellulose breakdown [47], and also acts as a probiotic that promotes the production of beneficial metabolites such as butyrate and lactate[54]. R. albus, another probiotic and cellulolytic bacterium, enhances fiber digestion and oxidative resistance[55]. Similarly, R. flavefaciens is a key fiber-degrading anaerobe associated with improved weight gain, feed digestibility, and fermentation efficiency[56,57]. P. ruminicola (cellulolytic species) has been found in higher abundance in cows supplemented with riboflavin[58] and in high-yielding dairy cattle[59]. These findings suggest that warm water intake during the cold period may enhance cellulose metabolism by promoting the colonization of these beneficial probiotic bacteria in the rumen, ultimately supporting better digestion and improved growth performance.
5. Conclusion
Bulls drinking warm water in winter improved the growth performance of by promoting ADG, DMI, and reducing F:G ratio. This improvement may be attributed to reduced energy loss and increased ruminal VFA concentrations. Additionally, warm water intake alleviated the stress associated with maintaining thermal balance from cold water, as evidenced by lower concentrations of thyroxine, serum urea nitrogen, and glucose in the blood. Furthermore, warm water intake enhanced rumen fermentation by increasing the abundance of total bacteria, fiber-digesting bacteria, and the activity of cellulase and xylanase in the rumen. Therefore, providing warm water to bulls is crucial for improving their growth during the winter months.
Author Contributions
Conceptualization, Peishi Yan; Data curation, Jing Li; Formal analysis, Jing Li and Zhihao Cui; Funding acquisition, Peishi Yan; Investigation, Ming Wei and Chunqing Yin; Resources, Peishi Yan; Supervision, Peishi Yan; Writing-original draft, Jing Li; Writing-review & editing, Peishi Yan.
Funding
This study was supported by the Public Welfare Industry (Agriculture) Scientific Research Projects (201303145) and the National Natural Science Foundation of China (3150930).
Institutional Review Board Statement
The animals were treated as approved by the Humane Animal Care and Ethics Committee of the Nanjing Agricultural University, China (SYXK2011-0036).
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to further scientific use.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gleason, C.B.; White, R.R. BEEF SPECIES-RUMINANT NUTRITION CACTUS BEEF SYMPOSIUM: A role for beef cattle in sustainable U.S. food production1. Journal of animal science, 2019, 97, 4010–4020. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.B.; Fordyce, G.; Mcgowan, M.R.; et al. Perspectives for reproduction and production in grazing sheep and cattle in Australasia: The next 20 years. Theriogenology, 2024, 230, 174–182. [Google Scholar] [CrossRef]
- Sorbolini, S.; Bongiorni, S.; Cellesi, M.; et al. Genome wide association study on beef production traits in Marchigiana cattle breed. Journal of animal breeding and genetics = Zeitschrift fur Tierzuchtung und Zuchtungsbiologie, 2017, 134, 43–48. [Google Scholar] [CrossRef]
- Zachut, M.; Šperanda, M.; DEAlmeida, A.M.; et al. Biomarkers of fitness and welfare in dairy cattle: healthy productivity. The Journal of dairy research, 2020, 87, 4–13. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Rashamol, V.P.; Bagath, M.; et al. Impacts of heat stress on immune responses and oxidative stress in farm animals and nutritional strategies for amelioration. International journal of biometeorology, 2021, 65, 1231–1244. [Google Scholar] [CrossRef]
- Hylander, B.L.; Repasky, E.A. Temperature as a modulator of the gut microbiome: what are the implications and opportunities for thermal medicine? International journal of hyperthermia : the official journal of European Society for Hyperthermic Oncology, North American Hyperthermia Group 2019, 36, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Castellani, J.W.; Tipton, M.J. Cold Stress Effects on Exposure Tolerance and Exercise Performance. Comprehensive Physiology, 2015, 6, 443–469. [Google Scholar] [PubMed]
- Li, J.; Cui, Z.; Wei, M.; et al. Omics analysis of the effect of cold normal saline stress through gastric gavage on LPS induced mice. Frontiers in microbiology, 2023, 14, 1256748. [Google Scholar] [CrossRef]
- Lian, S.; Xu, B.; Wang, D.; et al. Possible mechanisms of prenatal cold stress induced-anxiety-like behavior depression in offspring rats. Behavioural brain research, 2019, 359, 304–311. [Google Scholar] [CrossRef]
- Fu, X.; Zhang, Y.; Zhang, Y.G.; et al. Research and application of a new multilevel fuzzy comprehensive evaluation method for cold stress in dairy cows. Journal of dairy science, 2022, 105, 9137–9161. [Google Scholar] [CrossRef]
- Wang, S.; Li, Q.; Peng, J.; et al. Effects of Long-Term Cold Stress on Growth Performance, Behavior, Physiological Parameters, and Energy Metabolism in Growing Beef Cattle. Animals : an open access journal from MDPI 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Jia, D.; Sun, Y.; et al. Microbiota: A key factor affecting and regulating the efficacy of immunotherapy. Clinical and translational medicine, 2023, 13, e1508. [Google Scholar] [CrossRef]
- Jia, X.; Xu, W.; Zhang, L.; et al. Impact of Gut Microbiota and Microbiota-Related Metabolites on Hyperlipidemia. Frontiers in cellular and infection microbiology, 2021, 11, 634780. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: a review. Antonie van Leeuwenhoek, 2020, 113, 2019–2040. [Google Scholar] [CrossRef]
- Nishino, K.; Nishida, A.; Inoue, R.; et al. Analysis of endoscopic brush samples identified mucosa-associated dysbiosis in inflammatory bowel disease. Journal of gastroenterology, 2018, 53, 95–106. [Google Scholar] [CrossRef]
- Li, Y.; Xia, S.; Jiang, X.; et al. Gut Microbiota and Diarrhea: An Updated Review. Frontiers in cellular and infection microbiology, 2021, 11, 625210. [Google Scholar] [CrossRef] [PubMed]
- Szóstak, N.; Figlerowicz, M.; Philips, A. The emerging role of the gut mycobiome in liver diseases. Gut microbes, 2023, 15, 2211922. [Google Scholar] [CrossRef]
- Brod, D.L.; Bolsen, K.K.; Brent, B.E. Effect of water temperature on rumen temperature, digestion and rumen fermentation in sheep. Journal of animal science, 1982, 54, 179–182. [Google Scholar] [CrossRef]
- Mclean, J.; Tobin, G. Animal and human calorimetry; Cambridge university press, 2007. [Google Scholar]
- Berbert, Queiroz D M, Melo E C. AOAC. 2005. Official Methods of Analysis of AOAC International. 18th Ed. USA.
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. Journal of dairy science, 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. Journal of dairy science, 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Gu, Z.; Zhu, Y.; Jiang, S.; et al. Tilapia head glycolipids reduce inflammation by regulating the gut microbiota in dextran sulphate sodium-induced colitis mice. Food Funct, 2020, 11, 3245–3255. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Cao, Y.; Liu, N.; et al. Subacute ruminal acidosis challenge changed in situ degradability of feedstuffs in dairy goats. Journal of dairy science, 2014, 97, 5101–5109. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Long, S.; Yi, G.; et al. Heating Drinking Water in Cold Season Improves Growth Performance via Enhancing Antioxidant Capacity and Rumen Fermentation Function of Beef Cattle. Antioxidants (Basel, Switzerland) 2023, 12. [Google Scholar] [CrossRef]
- Zhao, F.Q.; Zhang, Z.W.; Qu, J.P.; et al. Cold stress induces antioxidants and Hsps in chicken immune organs. Cell stress & chaperones, 2014, 19, 635–648. [Google Scholar]
- Zhang, Z.; Li, Z.; Zhao, H.; et al. Effects of Drinking Water Temperature and Flow Rate during Cold Season on Growth Performance, Nutrient Digestibility and Cecum Microflora of Weaned Piglets. Animals : an open access journal from MDPI 2020, 10. [Google Scholar] [CrossRef]
- Liu, T.; Wang, Q.; Gao, C.; et al. Drinking Warm Water Promotes Performance by Regulating Ruminal Microbial Composition and Serum Metabolites in Yak Calves. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- Callet, T.; Dupont-Nivet, M.; Cluzeaud, M.; et al. Detection of new pathways involved in the acceptance and the utilisation of a plant-based diet in isogenic lines of rainbow trout fry. PloS one, 2018, 13, e0201462. [Google Scholar] [CrossRef]
- Sutter, F.; Borchardt, S.; Schuenemann, G.M.; et al. Evaluation of 2 different treatment procedures after calving to improve harvesting of high-quantity and high-quality colostrum. Journal of dairy science, 2019, 102, 9370–9381. [Google Scholar] [CrossRef]
- Kennedy, P.M.; Milligan, L.P. Effects of cold exposure on digestion, microbial synthesis and nitrogen transformations in sheep. The British journal of nutrition, 1978, 39, 105–117. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, H.; Ran, T.; et al. Influence of yeast culture and feed antibiotics on ruminal fermentation and site and extent of digestion in beef heifers fed high grain rations1. Journal of animal science, 2018, 96, 3916–3927. [Google Scholar] [CrossRef]
- Serviento, A.M.; He, T.; Ma, X.; et al. Modeling the effect of ambient temperature on reticulorumen temperature, and drinking and eating behaviors of late-lactation dairy cows during colder seasons. Animal : an international journal of animal bioscience, 2024, 18, 101209. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.J.; Engle, T.E. Invited Review: Water consumption, and drinking behavior of beef cattle, and effects of water quality. Applied Animal Science, 2021, 37, 418–435. [Google Scholar] [CrossRef]
- Mun, S.; Park, K.; Lee, S. Evaluation of thermal sensitivity is of potential clinical utility for the predictive, preventive, and personalized approach advancing metabolic syndrome management. The EPMA journal, 2022, 13, 125–135. [Google Scholar] [CrossRef]
- Brcko, C.C.; Da Silva JA, R.; Martorano, L.G.; et al. Infrared Thermography to Assess Thermoregulatory Reactions of Female Buffaloes in a Humid Tropical Environment. Frontiers in veterinary science, 2020, 7, 180. [Google Scholar] [CrossRef] [PubMed]
- HUSVéTH F J D U, UNIVERSITY OF WEST HUNGARY, PANNON UNIVERSITY. P3. Physiological and reproductional aspects of animal production. 2011.
- Zhou, L.; Wang, Y.; Zhou, M.; et al. HOXA9 inhibits HIF-1α-mediated glycolysis through interacting with CRIP2 to repress cutaneous squamous cell carcinoma development. Nat Commun, 2018, 9, 1480. [Google Scholar] [CrossRef] [PubMed]
- Geng, A.L.; Zhang, Y.; Zhang, J.; et al. Effects of light regime on the hatching performance, body development and serum biochemical indexes in Beijing You Chicken. Poultry science, 2021, 100, 101270. [Google Scholar] [CrossRef]
- Kocalis, H.E.; Hagan, S.L.; George, L.; et al. Rictor/mTORC2 facilitates central regulation of energy and glucose homeostasis. Molecular metabolism, 2014, 3, 394–407. [Google Scholar] [CrossRef]
- Huang, L.; Shakhnovich, E.I. Is there an en route folding intermediate for Cold shock proteins? . Protein science : a publication of the Protein Society, 2012, 21, 677–685. [Google Scholar] [CrossRef]
- He, J.; Zhang, P.; Shen, L.; et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. International journal of molecular sciences 2020, 21. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De La Fuente, M.K.; Landskron, G.; et al. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Frontiers in immunology, 2019, 10, 277. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; et al. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell, 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Erny, D.; Dokalis, N.; Mezö, C.; et al. Microbiota-derived acetate enables the metabolic fitness of the brain innate immune system during health and disease. Cell metabolism, 2021, 33, 2260–2276.e7. [Google Scholar] [CrossRef]
- Haghikia, A.; Zimmermann, F.; Schumann, P.; et al. Propionate attenuates atherosclerosis by immune-dependent regulation of intestinal cholesterol metabolism. European heart journal, 2022, 43, 518–533. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Liu, W.; Wang, R.; et al. Propionate promotes ferroptosis and apoptosis through mitophagy and ACSL4-mediated ferroptosis elicits anti-leukemia immunity. Free radical biology & medicine, 2024, 213, 36–51. [Google Scholar]
- Luise, D.; Correa, F.; Stefanelli, C.; et al. Productive and physiological implications of top-dress addition of branched-chain amino acids and arginine on lactating sows and offspring. Journal of animal science and biotechnology, 2023, 14, 40. [Google Scholar] [CrossRef]
- Ben Hmad, I.; Mokni Ghribi, A.; Bouassida, M.; et al. Combined effects of α-amylase, xylanase, and cellulase coproduced by Stachybotrys microspora on dough properties and bread quality as a bread improver. International journal of biological macromolecules, 2024, 277, 134391. [Google Scholar] [CrossRef] [PubMed]
- Rabee, A.E.; Forster, R.; Sabra, E.A. Lignocelluloytic activities and composition of bacterial community in the camel rumen. AIMS microbiology, 2021, 7, 354–367. [Google Scholar] [CrossRef]
- Yeoman, C.J.; Fields, C.J.; Lepercq, P.; et al. In Vivo Competitions between Fibrobacter succinogenes, Ruminococcus flavefaciens, and Ruminoccus albus in a Gnotobiotic Sheep Model Revealed by Multi-Omic Analyses. mBio 2021, 12. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, P.; Chen, N.; et al. Effects of housing systems and glucose oxidase on growth performance and intestinal health of Beijing You Chickens. Poultry science, 2021, 100, 100943. [Google Scholar] [CrossRef]
- Petersen, M.K.; Muscha, J.M.; Mulliniks, J.T.; et al. Water temperature impacts water consumption by range cattle in winter. Journal of animal science, 2016, 94, 4297–4306. [Google Scholar] [CrossRef]
- Asanuma, N.; Kawato, M.; Hino, T. Presence of Butyrivibrio fibrisolvens in the digestive tract of dogs and cats, and its contribution to butyrate production. The Journal of general and applied microbiology, 2001, 47, 313–319. [Google Scholar] [CrossRef]
- Park, J.; Lee, J.; Yeom, Z.; et al. Neuroprotective effect of Ruminococcus albus on oxidatively stressed SH-SY5Y cells and animals. Scientific reports, 2017, 7, 14520. [Google Scholar] [CrossRef] [PubMed]
- Præsteng, K.E.; Pope, P.B.; Cann, I.K.; et al. Probiotic dosing of Ruminococcus flavefaciens affects rumen microbiome structure and function in reindeer. Microbial ecology, 2013, 66, 840–849. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; Gado, H.; Anele, U.Y.; et al. Influence of dietary probiotic inclusion on growth performance, nutrient utilization, ruminal fermentation activities and methane production in growing lambs. Animal biotechnology, 2020, 31, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.M.; Zhang, J.; Wang, C.; et al. Effects of riboflavin supplementation on performance, nutrient digestion, rumen microbiota composition and activities of Holstein bulls. The British journal of nutrition, 2021, 126, 1288–1295. [Google Scholar] [CrossRef]
- Lv, X.; Chen, L.; Zhou, C.; et al. Application of different proportions of sweet sorghum silage as a substitute for corn silage in dairy cows. Food science & nutrition, 2023, 11, 3575–3587. [Google Scholar]
Figure 1.
Comparative analysis of rumen cellulase levels in bulls provided with cold versus warm water (*, p < 0.05; **, p < 0.01). C, bulls drinking cold water; E, bulls drinking cold water.
Figure 1.
Comparative analysis of rumen cellulase levels in bulls provided with cold versus warm water (*, p < 0.05; **, p < 0.01). C, bulls drinking cold water; E, bulls drinking cold water.
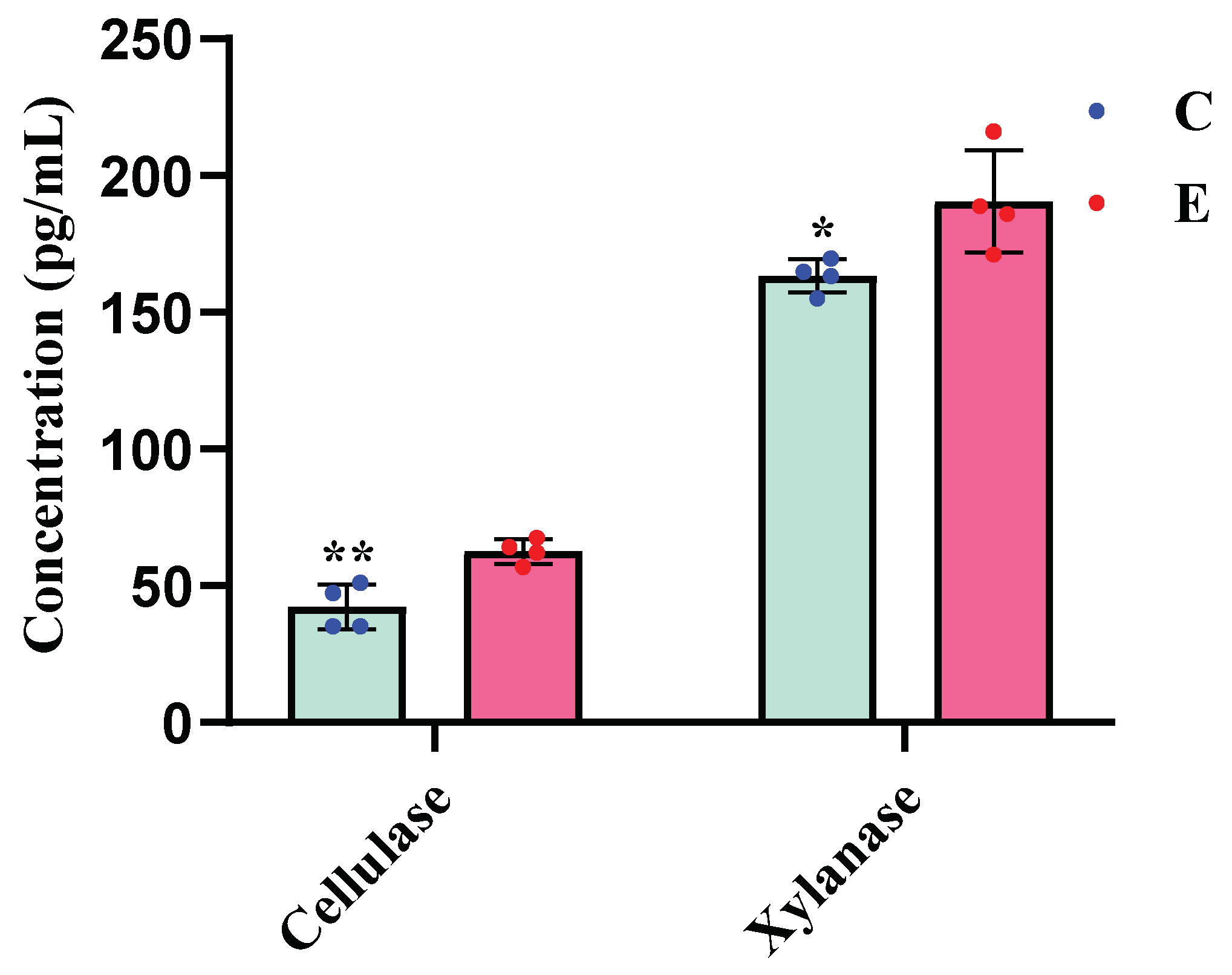
Figure 2.
Comparative analysis of rumen bacterial abundance in bulls provided with cold versus warm water (*, p < 0.05; **, p < 0.01). C, bulls drinking cold water; E, bulls drinking cold water.
Figure 2.
Comparative analysis of rumen bacterial abundance in bulls provided with cold versus warm water (*, p < 0.05; **, p < 0.01). C, bulls drinking cold water; E, bulls drinking cold water.
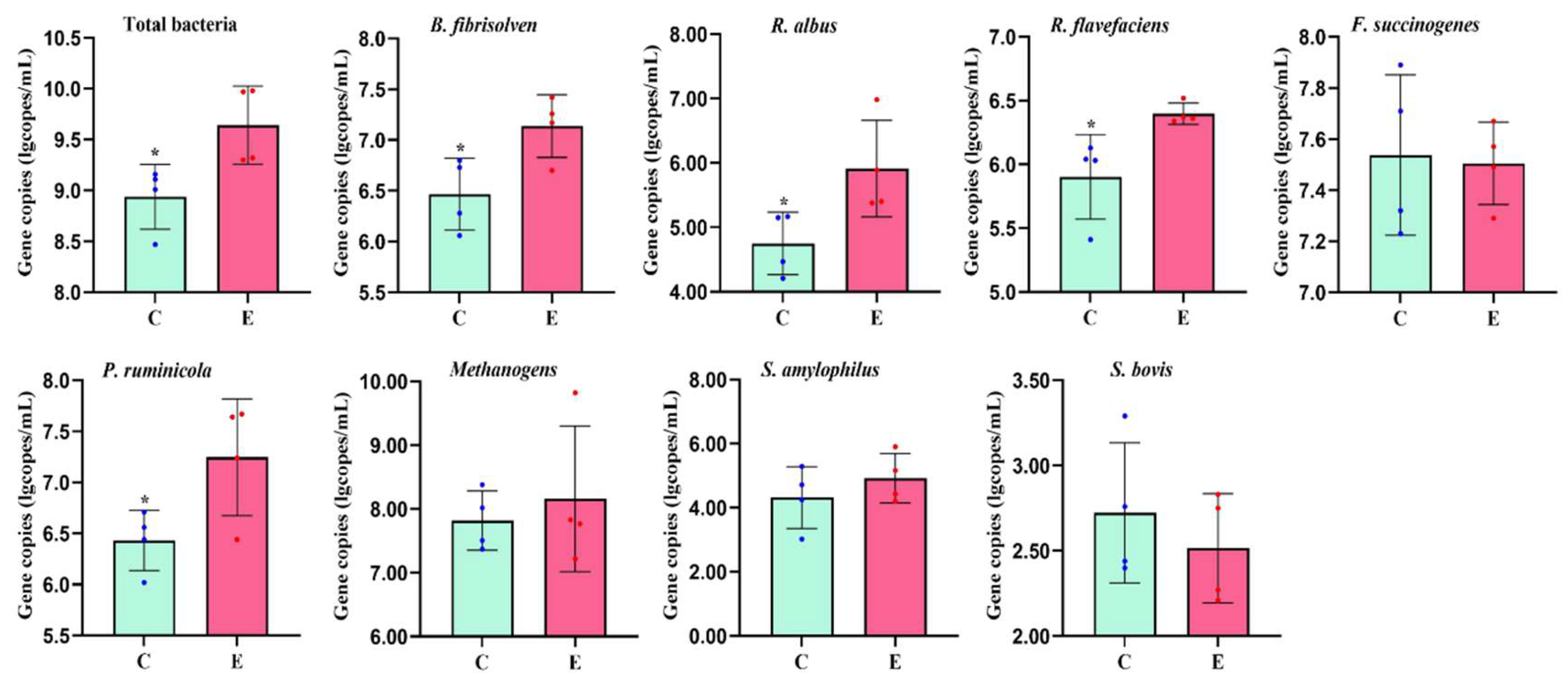

Table 1.
Ingredients and chemical composition of experimental diets.
| Item | Content |
| Ingredients, % | |
| Corn silage | 85.50 |
| Dry straw | 4.50 |
| Ground corn | 7.00 |
| Soybean meal | 2.50 |
| Calcium carbonate | 0.04 |
| Calcium hydrogen phosphate | 0.06 |
| Sodium bicarbonate | 0.10 |
| Sodium chloride | 0.08 |
| Limestone | 0.13 |
| Premix1 | 0.09 |
| Chemical composition, % of DM | |
| Dry matter | 87.26 |
| Organic matter | 94.47 |
| Crude protein | 8.14 |
| Ether extract | 6.53 |
| Neutral detergent fiber | 61.95 |
| Acid detergent fiber | 34.24 |
| Calcium | 0.83 |
| Phosphorus | 0.27 |
| Metabolic energy (MJ kg-1)2 | 7.30 |
1 Supplied per kilogram of dietary DM: VA 264 IU, VD 34 IU, VE 2.6 IU, Cu 1.2 mg, Mn 4.8 mg, Fe 6.0 mg, I 0.03 mg, Co 0.012 mg, Se 0.036 mg. 2 Metabolic energy was a calculated value.
Table 2.
Specific primers used for real-time PCR in this study.
| Target gene | Primers | Primer sequence(5’→3’) | Size(bp) |
| Total bacteria | F | CGGCAACGAGCGCAACCC | 130 |
| R | CCATTGTAGCACGTGTGTAGCC | ||
| Ruminococcus albus | F | CCCTAAAAGCAGTCTTAGTTCG | 176 |
| R | CCTCCTTGCGGTTAGAACA | ||
| Ruminococcus flavus | F | CGAACGGAGATAATTTGAGTTTACTTAGG | 132 |
| R | CGGTCTCTGTATGTTATGAGGTATTACC | ||
| Butyrivibrio fibrisolvens | F | GCCTCAGCGTCAGTAATCG | 121 |
| R | GGAGCGTAGGCGGTTTTAC | ||
| Fibrobacter succinogenes | F | GTTCGGAATTACTGGGCGTAAA | 121 |
| R | CGCCTGCCCCTGAACTATC | ||
| Methanogens | F | TTCGGTGGATCDCARAGRGC | 232 |
| R | GBARGTCGWAWCCGTAGAATCC | ||
| Succinimonas amylolytica | F | CGTTGGGCGGTCATTTGAAAC | 139 |
| R | CCTGAGCGTCAGTTACTATCCAGA | ||
| Streptococcus bovis | F | ATGTTAGATGCTTGAAAGGAGCAA | 127 |
| R | CGCCTTGGTGAGCCGTTA | ||
| Prevotella ruminicola | F | GAAAGTCGGATTAATGCTCTATGTTG | 102 |
| R | CATCCTATAGCGGTAAACCTTTGG |
Table 3.
Growth performance in bulls provided with cold water versus warm water.
| Group2 | |||
| Item1 | C | E | P-value |
| IBW, kg | 359.34±10.62 | 371.57±14.56 | 0.523 |
| FBW, kg | 399.98±8.54 | 420.08±16.11 | 0.313 |
| ADG, kg/d | 0.95±0.05 | 1.15±0.03 | 0.024 |
| DMI, kg/d | 6.45±0.92 | 6.76±0.08 | 0.046 |
| F:G, ratio | 6.71±0.26 | 5.89±0.22 | 0.047 |
1 IBW = initial body weight, FBW = final body weight, ADG = average daily gain, DMI = dry matter intake, F:G ratio = the ratio of feed intake and weight gain. 2 C, bulls drinking cold water; E, bulls drinking cold water.
Table 4.
Thermal stress indicators in bulls provided with cold water versus warm water.
| Group1 | |||
| Item | C | E | P-value |
| Heat production, MJ/W0.75 h-1 | 29.64±6.95 | 25.76±4.52 | 0.385 |
| Respiratory rate, min-1 | 8.94±2.38 | 10.44±2.25 | 0.395 |
| Body surface temperature, ℃ | |||
| Spatial temperature 5-9℃ | 15.97±1.49 | 16.19±1.83 | 0.746 |
| Spatial temperature 9-11℃ | 20.67±1.64 | 21.06±0.43 | 0.673 |
| Spatial temperature 14-16℃ | 23.00±1.89 | 23.96±1.12 | 0.416 |
| Rectal temperature, ℃ | 38.00±0.50 | 38.24±0.23 | 0.417 |
1 C, bulls drinking cold water; E, bulls drinking cold water.
Table 5.
Serum hormones and metabolites in bulls provided with cold water versus warm water.
| Group1 | |||
| Item | C | E | P-value |
| Triiodothyronine, ng/mL | 1.53±0.09 | 1.22±0.09 | 0.058 |
| Thyroxine, ng/mL | 134.28±1.04 | 128.99±1.35 | 0.021 |
| Growth hormone, ng/mL | 5.26±0.15 | 5.38±0.26 | 0.688 |
| Serum urea nitrogen, mmol/L | 6.79±0.40 | 5.55±0.13 | 0.025 |
| Glucose, mmol/L | 4.85±0.09 | 4.40±0.08 | 0.011 |
| Total protein, g/L | 58.82±3.61 | 54.07±2.45 | 0.063 |
1 C, bulls drinking cold water; E, bulls drinking cold water.
Table 6.
Rumen pH and fermentation parameters in bulls provided with cold water versus warm water.
| Group2 | |||
| Item | C | E | P-value |
| Rumen pH | 7.27±0.04 | 7.19±0.03 | 0.061 |
| NH3-N, mg/dL | 4.67±0.19 | 5.23±0.11 | 0.048 |
| Total VFA, mM | 60.52±2.19 | 78.61±4.34 | 0.010 |
| Acetate, mM | 41.92±1.62 | 55.14±3.04 | 0.009 |
| Propionate, mM | 10.26±0.40 | 13.49±0.75 | 0.009 |
| Butyrate, mM | 4.53±0.18 | 5.30±0.29 | 0.066 |
| Iso-butyrate, mM | 0.88±0.08 | 1.09±0.06 | 0.074 |
| Valerate, mM | 1.19±0.11 | 1.47±0.08 | 0.091 |
| Iso-valerate, mM | 1.25±0.17 | 2.11±0.21 | 0.139 |
| A:P, ratio1 | 4.36±0.08 | 4.48±0.17 | 0.539 |
1 A:P = the ratio of acetate and propionate. 2 C, bulls drinking cold water; E, bulls drinking cold water.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



