
Submitted:
30 January 2025
Posted:
30 January 2025
You are already at the latest version
Abstract
Inhibin (INH) plays a key role in the regulation of the reproductive performance of geese. It inhibits the follicle-stimulating hormone (FSH) secretion from the anterior pituitary gland to regulate spermatogenesis. Immunization against INH in male geese leads to the production of antibodies to neutralize the INH activity that enhances testicular function and gonadotropin production. The objectives of the present study were to investigate the potential role of immunization against INH on testicular histology, plasma LH, pituitary PRL mRNA, and hypothalamic VIP mRNA expressions in Yangzhou Ganders. A total of 60 birds were selected and divided into control (CON) and INH immunized (INH immunized), having 30 in each group. In this experiment, the ganders were immunized with INH three times, and birds in the CON group were inoculated with bovine serum albumin (BSA). Analyzed data revealed that immunization against inhibin has no significant effects on improving the plasma concentration of LH hormone, germ cell numbers, hypothalamic VIP mRNA, and pituitary PRL mRNA expressions. It is concluded that INH (INH) immunization is a sublime tool to improve reproductive efficiency in Yangzhou ganders, however, INH immunization may have a positive effect on pituitary PRL mRNA and hypothalamic mRNA expressions and LH plasma concentration. The seasonality has played a vital impact on its hypothalamus pituitary gonadal (HPG) axis.
Keywords:
INH immunization
; testicular histoarchitecture
; PRL mRNA
; VIP mRNA
; Yangzhou ganders
1. Introduction
Males, especially male geese, have reproductive physiology that is regulated by the glycoprotein hormone inhibin. It is mostly released by the Sertoli cells and is important for the feedback control of the pituitary gland's release of follicle-stimulating hormone (FSH). There are two primary forms of inhibin, inhibin A and inhibin B. Inhibin B is more common in men and is a marker for spermatogenic status and Sertoli cell function [1]. Spermatogenic activity in the testes is intimately related to inhibin B secretion. According to studies, inhibin B levels have an inverse relationship with FSH levels and a positive correlation with sperm count and Sertoli cell activity [2]. This feedback system is essential for preserving the gonadotropin balance required for healthy spermatogenesis. Age and metabolic state are two variables that affect inhibin B production; decreases in inhibin B levels are frequently indicative of decreased spermatogenesis [3]. The expression of inhibin subunits has been observed in the testes at different developmental stages, suggesting a role in the maturation process of the male reproductive system [4]. Inhibin's role goes beyond simple feedback inhibition, as it also is involved in the paracrine regulation of testicular function, influencing the activity of Leydig cells and the overall hormonal milieu within the testes [5].
Studies on other bird species, including Japanese quail, have shown that inhibin is released from the testes and plays a role in controlling gonadotropin levels, which suggests that inhibin is physiologically significant in male geese [6]. Active immunization against INH improved testes weight and plasma testosterone concentrations in aging broiler breeders [7]. Another very important modulator of fertility is a vasoactive intestinal peptide (VIP) [8]. VIP functions as a neurotransmitter. It is not only produced in the hypothalamus but also distributed in the testis and involved in gametogenesis and testosterone synthesis [9-14]. In seasonal breeders, seasonality has a marginal effect on gander’s reproductive efficiency. Seasonal reproduction in both short and long-breeding birds is regulated by gonadotrophin-releasing hormone (GnRH) and vasoactive intestinal peptide (VIP) in the hypothalamus equivalent to prolactin (PRL) and luteinizing hormone (LH) in the pituitary gland [15,16].
In geese production, the reproductive efficiency of ganders declines with seasonal breeding [17]. After two months of sublime reproductive efficiency, a decline in semen quality, and fertility accompanied by alterations in endocrine parameters and sexual behaviors was observed in ganders [18,19]. In commercial goose production, this evidence raises attention to the importance of high reproductive male breeder stock. Its need of the hour to elaborate reasons of poor reproduction in ganders and remedies explored by poultry endocrinologists and theriogenologists all over the World to improve commercial ganders stock reproduction. In past years, there have been attempts to enhance reproductive efficiency in ganders by changing lighting regimes, breeding stage, dietary supplementation of vitamin E and Selenium, and crossbreeding [20,21]. INH immunization is one of the optimum tools to improve reproductive efficiency in birds and mammals. In birds, both LH and PRL are secreted by the anterior pituitary. LH acts on Leydig cells to produce testosterone (T) while FSH acts on Sertoli cells that secrete inhibin. Inhibin via the feedback loop acts on FSH and Leydig cells also need inhibin for testosterone (T) synthesis as inhibin acts as a cross-link between Leydig and Sertoli cells to maintain spermatogenesis. This makes a cross-link between LH, PRL, and inhibin. FSH is an important modulator in this loop. So, we hypothesized that active immunization against INH may improve pituitary PRL and hypothalamic VIP and thus it may aid in improving reproduction of Yangzhou ganders. Geese breeds in China except Yili geese were developed from wild swan geese (Anser cygonoides) about 6000 years before [22]. Our experimental bird, the Yangzhou goose is a Chinese-origin long-day breeder bird developed using local germplasm resources. It is popular due to its sublime fertility, hatchability, egg and carcass quality. Luteinizing Hormone (LH) in the pituitary and Vasoactive intestinal peptide (VIP) are important players in the hypothalamus-pituitary gonadal axis (HPG axis) and reproductive cycle [23,24]. Vasoactive Intestinal Peptide is secreted in the hypothalamus and inhibits the reproductive performance of young White Leghorn roosters [25]. Similarly, INH inhibits the secretion of FSH, so the activity of these two hormones i.e., VIP and INH are interlinked on the hypothalamus gonadal axis (HPG). In our previous work, we discovered that INH immunization affects spermatogenesis and testicular development in Yangzhou ganders. Also, the Sertoli cell genes (SOX9, Wt1, Dhh, AMH, and FSHR) were downregulated with INH immunization describing that Sertoli cell development had been downregulated. Also, we had enunciated that INH immunization regresses germ cell (spermatogonia, spermatocytes) development in the seminiferous epithelium of Yangzhou ganders. Our previous findings give the notion that INH immunization has its effect on the hypothalamus-pitutary-gonadal axis but whether it affects LH and PRL and PRL mRNA expressions in the pituitary and hypothalamus remains unknown. The Purpose of the present experiment was to elucidate the impact of INH immunization on Luteinizing Hormone (LH), changes in the seminiferous epithelium, and relative mRNA expressions of PRL and VIP in the pituitary and hypothalamus.
2. Materials and Methods
2.1. Ethics Consideration
This experiment was conducted at the Nanjing Agricultural University, China. All experimental designs and procedures were approved regarding ethical conditions by the Animal Care and Use Committee (ACUC) held at Jiangsu Academy of Agriculture Sciences, Nanjing Agricultural University, China Vide reference No: 31572403 and 31402075.
The present study has been conducted using prepared INH protein injected in yangzhou ganders used as an experimental model. The whole study was completely applying blind (masking) evaluation. The method of protecting the study group under evaluation hidden after randomization is usually used to reduce the risk of bias in clinical trials with the study groups.
2.2. Place of Experiment
The current experiment was conducted at Sunlake Swan farm in Henglin Township, Changzhou Jiangsu province, China.
2.3. Selection of Birds and Experimental Design
A total of 60 Yangzhou ganders (having the same genetic origin) were selected for this study and randomly assigned into two groups A and B. These birds were kept under the naturally prevailing climatic conditions and were allowed to acclimatize for one week before the start of the experiment. For identification, tags were attached under the wings of each bird. Each group comprises 30 birds, group A was kept as control (CON) while group B was kept as INH group. Birds in CON were immunized with BSA (bovine serum albumin) while those in the INH group were immunized against INH. Throughout the study, the birds were housed at temperatures ranging from 25°C to 32°C. The ganders had free access to water and were fed a diet containing 12.5% crude protein (CP), which was mixed with green grass. Ad libitum feed was offered in daytime (from sunrise to evening). The mean body weight (BW) of the control and INH immunized group is provided in Table 1 as follows.
2.4. INH Preparation
A recombinant goose INH fusion peptide (142 amino acid residue), plasmid pRSET-A, and 106-residue fragment of goose INH α-subunit mature peptide fragment was expressed in E. coli BL21(DE3) and purified according to the protocol as described by [26]. After the final purified concentration of INH protein was 3 mg/ml, INH protein was then mixed with a mineral oil adjuvant (Solarbio Life Sciences) at a 1:2 (v/v) ratio. The INH protein and mineral oil adjuvant mixture was condensed to achieve a final protein concentration of 1 mg/mL, preparing the INH immunogenic protein. Subsequently, physiological saline was mixed with the mineral oil adjuvant to prepare the bovine serum albumin (BSA) solution. Ganders in the INH group were immunized with the INH protein on days 161, 181, and 227, while birds in the CON group were inoculated with BSA on the same days. The first immunization occurred when the birds were 161 days of age. It typically takes 20 days for the inhibin vaccine to induce an antibody response and produce antibodies. Therefore, the second booster shot was administered at 181 days of age, and the third inhibin shot was given at 227 days of age. Consequently, time points at 181, 200, and 227 days of age were selected for sampling. Additionally, by 161 days of age, Yangzhou ganders begin reaching their optimal body weight (B.W.), which further justified the selection of 181, 200, and 227 days to assess whether age might have any impact on the immune response.
Figure 1.
Graphical abstract showing molecular regulation of inhibin and related hormones on hypothalamus-pituitary-gonadal axis.
Figure 1.
Graphical abstract showing molecular regulation of inhibin and related hormones on hypothalamus-pituitary-gonadal axis.

2.5. LH plasma Concentration Measurement
Plasma luteinizing hormone (LH) concentration was quantified using chicken LH (USDA-cLH-K-3) as the reference standard and USDA-cLH-I-3 as the radiolabeled ligand. Rabbit anti-chicken LH (USDA-AcLH-5) and donkey anti-rabbit antiserum (Abcam ab7080) were used as the primary and secondary antibodies, respectively. The intra-assay and inter-assay coefficients of variation were both below 15%, and the sensitivity of the assay was 0.1 ng/mL. It should be noted that the R-values of the standard curves were consistently above 0.99.
2.6. Isolation of, Synthesize Complementary DNA (cDNA) Synthesis, and Quantitative Real-Time PCR (qRT-PCR)
RNA was extracted from the pituitary and hypothalamus using the Trizol reagent with an RNA extraction kit (Tianjin) following the manufacturer's instructions. Briefly, 0.2 mL of chloroform (Henan GP Chemicals) was added to the Trizol reagent, and the mixture was homogenized by shaking. The sample was then centrifuged at 12,000×g for 15 minutes at 4°C. The aqueous phase (supernatant) was mixed with 0.5 mL isopropanol and centrifuged again at 12,000×g for 10 minutes at 4°C. The resulting RNA pellet was washed with 75% ethanol, air-dried, and re-suspended in 20 µL of sterile treated diethylpyrocarbonate water.
RNA concentration and purity were assessed using a spectrophotometer (NanoDrop 2000c, Thermo Scientific, Germany). For reverse transcription, 5 µg of RNA was used with the Takara PrimeScript™ RT Reagent Kit (RR037, Takara, Japan) to synthesize complementary DNA (cDNA). Gene sequences are listed in Table 2. The entire process of RNA isolation, cDNA synthesis, and quantitative real-time PCR (qRT-PCR) followed the protocol described in [28]. Gene expression levels were calculated using the 2-CT method.
2.7. Tissue Sample Collection
Hypothalamus and pituitary tissues were collected at 181, 200, and 227 days of age. At 181 days of age, 10 birds from each group were selected and slaughtered for tissue sampling. Similarly, 10 birds from each group were randomly selected and slaughtered on days 200 and 227 of age for tissue collection. The samples were immediately stored at -80°C after collection. Blood samples were collected via brachial venipuncture on days 161, 181, 190, 200, 227, 225, and 227 into heparinized tubes (5 mL, Hangzhou Rollmed Co. Limited, China). The heparinized tubes were centrifuged at 2000 ×g for 3 minutes at 4°C. Within 3 hours of blood collection, the plasma was separated by centrifugation at 1000 ×g and stored at -20°C.
2.8. Microscopy
To observe histological alterations in the seminiferous tubules, a small portion (0.125 cm³) of the left testis tissue was collected and fixed in 10% buffered formalin for 24 hours. Histological evaluation was performed using a tissue processor (LEICA RM 2235). The testis tissues were dehydrated in ascending concentrations of alcohol (70%, 80%, 90%, and 100%), followed by absolute alcohol. After clearing in xylene, the tissues were embedded in melted paraffin wax.
Sections of 5 µm thickness were cut perpendicular to the tissue and mounted onto glass slides. The slides were then stained with hematoxylin and eosin. These tissue sections were analyzed under a light microscope (Olympus BX63) at magnifications of 10× and 40× to examine changes in spermatogonia, spermatocytes, seminiferous tubule (ST) diameter, and elongated spermatids.
2.9. Statistical Analysis
Statistical analysis was performed using SPSS (Version 20.0, Armonk, NY, USA) and GraphPad Prism (Version 5.0). The Kolmogorov-Smirnov test was applied to assess the normality of the data. If the data were not normally distributed, they were log-transformed and re-tested for normality before further analysis. A two-way ANOVA was then conducted to compare the mean values. Results are presented as the mean ± standard error of the mean (SEM). Differences among groups at experimental time points were assessed using the Bonferroni post-test. A probability level of P < 0.05 was considered statistically significant.
3. Results
3.1. Plasma LH Concentrations
The plasma concentration of LH was significantly increased at day 225 in both groups A and B (CON and INH) after treatment of the third booster of INH immunization on day 227. Similarly, the plasma concentration of the CON group showed remains slight elevation till day at 227 of post-immunization treatment however, plasma LH concentration was detected lowest levels in the INH group treated with INH protein. Moreover, plasma concentration of LH in both groups (CON and INH) followed similar patterns of ascending and descending were observed until day 225 of age. Plasma LH concentrations in both groups CON and INH are shown in Figure 3a.
3.2. Pituitary Prolactin (PRL) mRNA Expression
The quantitative real-time (qRT-PCR) analysis showed that the PRL mRNA expressions were reduced on day 181 as compared CON group of Yangzhou ganders. Statistical analysis of PRL mRNA expressions showed a significantly decreased pattern on day 200 after 2nd dose of immunization in Yangzhou ganders of the INH group. In contrast on day 227 of age, mRNA expressions were increased in the INH group after 3rd dose of immunization (Figure 2b).
3.3. Hypothalamic Vasoactive Intestinal Peptide (VIP) mRNA Expression
The quantitative real-time (qRT-PCR) analysis showed that the level of VIP mRNA expression in the hypothalamus was significantly (P < 0.001) decreased in the INH group, while, significant (P < 0.001) increases were observed in the INH group on day 181 of age after immunization treatment. However, the level of VIP mRNA expression in the hypothalamus was increased in the INH group, and lower expression was observed in the CON group after immunization treatment at the end of the experiment on day 227 of age (Figure 2b).
3.4. Histology of Testes
Towards the end of the study, histological findings of the testes of the INH group were significantly different from theCON group after treatment of immunization at days 181, 200, and 227 of age. Additionally, normal development of germ cells was observed in both groups post-treatment of INH immunization at days 181, 200, and 227. However, the number of germ cells in histological sections in the INH group appeared deficient as compared to CON group.
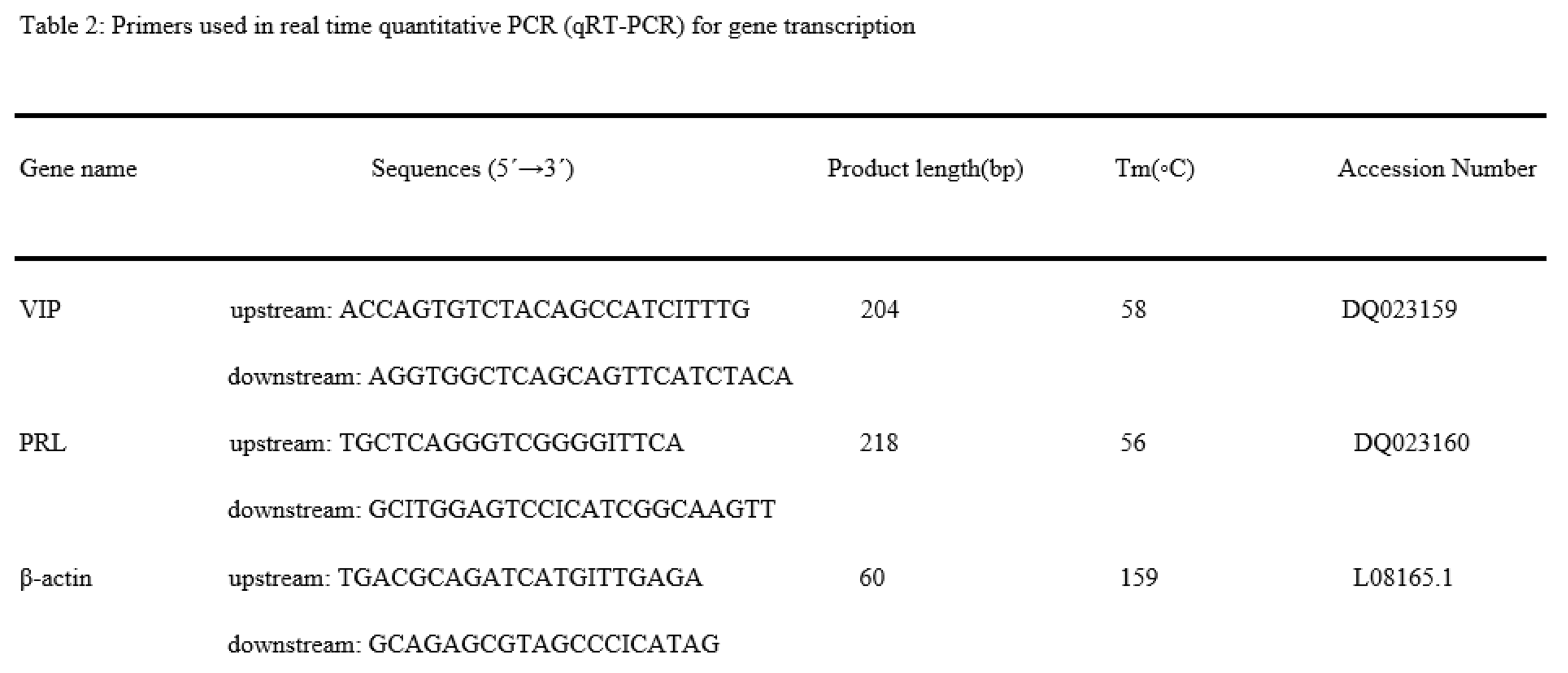
Histologically, testes collected from Yangzhou ganders in CON group showed significant filled with spermatogonia, spermatocytes, and spermatids. However, the histological study of group INH immunized with INH irregular shrinkage of seminiferous tubules with a marked reduction in the number of germ cells includes spermatogonia, spermatocytes, and spermatids. In Figure 3, a histological photomicrograph shows morphological alterations in seminiferous tubules of testes of Yangzhou ganders after post-INH immunization treatment at days 181, 200, and 227.
Figure 2.
Photo-micrographic changes in testicular of Yangzhou ganders at 181, 200, and 227 days of age in Control and INH immunized groups. Arrow with tail showing spermatogonia; arrowhead showing spermatogonia; arrow with spiral tail sowing spermatids. The scale bar represents 20μm at 40 × magnification.
Figure 2.
Photo-micrographic changes in testicular of Yangzhou ganders at 181, 200, and 227 days of age in Control and INH immunized groups. Arrow with tail showing spermatogonia; arrowhead showing spermatogonia; arrow with spiral tail sowing spermatids. The scale bar represents 20μm at 40 × magnification.
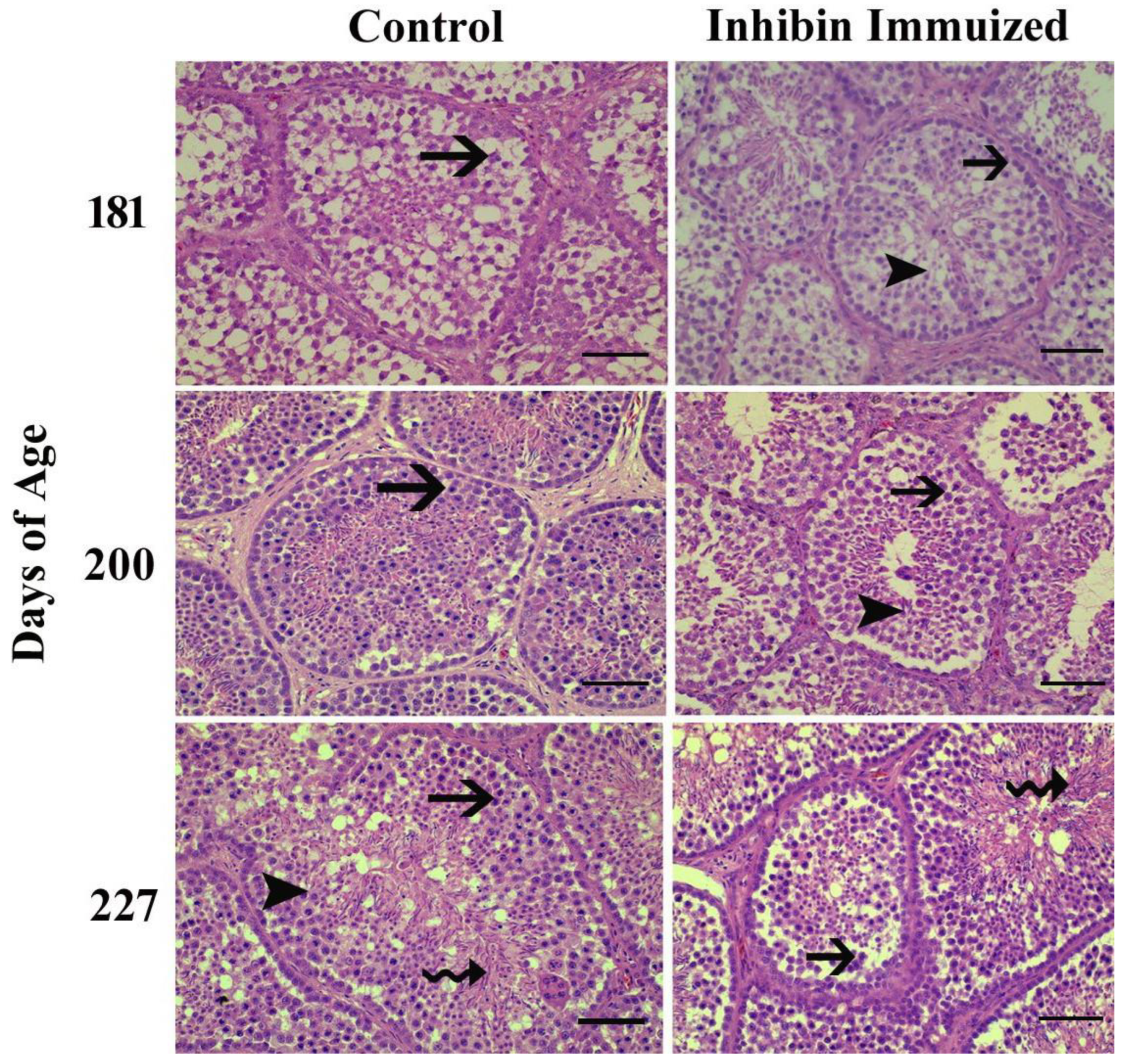
Figure 3.
(a) Plasma LH concentrations in INH Immunized (INH) and control group (CON) of Yangzhou goose ganders at age 161, 181, 190, 200, 227, 225, and 227 days. Vertical bars represent the standard error of mean (SEM). The values with ** indicate the difference (P < 0.001). Arrows indicate the age of birds at the time of immunization and following booster shots. (b) Pituitary PRL (c) Hypothalamic VIP mRNA levels in INH (INH) and (CON) group of Yangzhou goose ganders. Data are shown as mean values ± standard error of the mean. *, ** and *** indicate difference at P < 0.05, P < 0.01and P < 0.001, respectively between the groups.
Figure 3.
(a) Plasma LH concentrations in INH Immunized (INH) and control group (CON) of Yangzhou goose ganders at age 161, 181, 190, 200, 227, 225, and 227 days. Vertical bars represent the standard error of mean (SEM). The values with ** indicate the difference (P < 0.001). Arrows indicate the age of birds at the time of immunization and following booster shots. (b) Pituitary PRL (c) Hypothalamic VIP mRNA levels in INH (INH) and (CON) group of Yangzhou goose ganders. Data are shown as mean values ± standard error of the mean. *, ** and *** indicate difference at P < 0.05, P < 0.01and P < 0.001, respectively between the groups.
4. Discussion
The effects of INH immunization on Yangzhou gander’s plasma LH levels, testicular histological alterations, and PRL and VIP mRNA expression in the pituitary and hypothalamus are complex and can be explained by severalhormonal interactions and regulatory systems.
On the hypothalamus-pituitary-gonadal (HPG) axis, INH plays a critical role (Rehman et al., 2021). To increase spermatogenesis and reproductive efficiency, mammalian species have been immunized against INH (Akhtar et al., 2021). In Merino rams, sperm counts began to rise following INH vaccination (Voge & Wheaton, 2007). Our results show a significant difference in LH concentrations between the two groups on day 227, following the third dose of INH immunization. Moreover, the findings of the present study align with those of [27], which found no significant effect of inoculation with the INH-α subunit on plasma LH concentrations in developing cockerels. Similarly, in Shiba goats, passive INH immunization did not affect LH concentrations [28]. Likewise, turkey hens immunized with INH showed no effect on LH-β concentrations [29]. FSH acts on Sertoli cells and seminiferous tubules in initiating spermatogonium division [30], while LH influences Leydig cells to produce testosterone. This suggests that both LH and FSH play crucial roles in spermatogenesis, both inside and outside the seminiferous tubules [31].
The hypothalamic GnRH and gonadal steroid and INH, which either directly or indirectly influence gonadotropins through positive and negative feedback effects, positively regulate LH and FSH on the hypothalamus-pituitary-gonadal axis [32]. In a feedback loop, INH vaccination generates particular antibodies that neutralize endogenous INH, which increases FSH secretion and suppresses LH secretion [33]. In agreement with our findings, it was reported that a significant declining pattern of LH concentrations was observed in the INH group as compared to the rest of the experimental CON group. Moreover, by neutralizing endogenous INH, INH immunization has been suggested to increase FSH concentrations and stimulate follicular development [34]. INH immunization generates specific antibodies to neutralize endogenous INH, which in turn stimulates FSH secretion and inhibits LH secretion in a feedback manner [35]. It shows that only INH immunization doesn't seem to modulate endocrinological changes, and simultaneously, photoperiod has its effect on the modulation of LH. Geographical location affects the annual pattern of seasonality and reproduction in domestic geese [36]. In various avian species, photoperiod is a central factor for seasonal reproduction control in the temperate zone [37,38]. Yangzhou geese are long-day breeding geese species whose breeding period starts in October, peaks between February- March, and ends in June. Age of maturity largely affects the reproductive activities of birds, not only seasonality [39]. Until Yangzhou ganders reach the age of maturity, the major factor, i.e., photoperiod, peaked, modulating LH plasma concentrations followed by similar T concentrations. In contrast, high levels of T have a positive feedback effect on male animal spermatogenesis and maturation, but they also negatively affect LH levels [40].
In our results, testicular histology showed a reduced rate of spermatogenesis in the INH immunized group compared to the control group after INH immunization. Our findings are per [41], in which INH lowered spermatogonial numbers in the testes of mature mice and Chinese hamsters. According to a prior study, Yangzhou gander’s testicular cell counts began to decline following INH immunization; testicular apoptosis may be caused by the combined effects of INH and seasonality [42]. Moreover, seasonality in breeding improves the rate of apoptosis, which is a typical event in proliferating and continuously regenerating tissues and seminiferous epithelium [43]. However, INH immunization disrupted germ cell development, and spermatogenesis and caused apoptosis in seminiferous epithelium [42]. Another study depicted that INH immunization lowered the number of spermatogonia and elongated spermatids in ganders [44]. In contrast, active immunization against INH has been shown to improve ram’s testicular weight and reproductive efficiency [45] and influence spermatogenesis and testicular growth in goose ganders [29]. In cockerels, the size of the testicles has a direct impact on sperm production [46].
Sertoli cell expansion, as indicated by improved mRNA expression of Sertoli cell genes [47]. In the current investigation, the expression patterns of pituitary PRL mRNA and hypothalamus VIP mRNA was nearly identical, with very few exceptions. A prior study found similar expressions of PRL mRNA and VIP mRNA [48]. Moreover, the INH immunization increased the FSHR mRNA in the large white follicles of patridge shank hens. LH-β mRNA expression remained unaffected after INH immunization [49]. Pituitary PRL mRNA expressions are markedly elevated in this study and completely at odds with the LH plasma concentration. Similar results of the inverse connection between plasma PRL and LH and T concentrations in Zatorska ganders during the first reproductive cycle were previously reported by researchers [50]. We can infer from these results that PRL may influence endocrinological variations in the concentration of LH plasma, which are then reflected in pituitary PRL mRNA expressions, leading to a decreased rate of spermatogenesis. Moreover, PRL appears to influence changes in the first reproductive cycle, such as LH plasma concentration, which may change during the second season of reproductive activity. It is necessary to clarify the direct effect of PRL on testicular morphology.
5. Conclusions
Yangzhou gander’s reproductive activities are regulated on the HPG axis by the breeding season and their maturity age. An important part of the hypothalamus-pituitary-gonadal (HPG) axis is played by daily photoperiod and INH immunization. INH immunization caused disruptions to testicular histology and germ cells, resulting in decreased spermatogenesis efficiency in Yangzhou ganders. However, INH immunization may have a positive impact on pituitary PRL mRNA and hypothalamic mRNA expressions and LH plasma concentration. Moreover, Yangzhou geese are long-day breeding birds, even following INH immunization, the photoperiod has a significant impact on reproductive and endocrinological changes in Yangzhou geese. To gain a deeper understanding, more molecular pathways must be investigated.
Author Contributions
Conceptualization, writing original draft preparation, data curation: Muhammad Faheem Akhtar, supervision, data curation, funding acquisition: Wanf Changfa. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by grants of Jiangsu Innovation of Agricultural Science and Technology CX (15)1008, National Natural Science Foundation of China (31372314), China Agriculture Research System (CARS-40-20), Shandong Province Modern Agricultural Industrial Technology System Project (SDAIT-27), Ministry of Agriculture and Rural Livestock Seed Industry Project "Donkey Camel Species Molecular ID Construction" (19211162), Key research and development project of Shandong Province "Innovation and Demonstration of Key Technologies for Integrated Development of Dong'e Black Donkey Industry" (2021TZXD012).
Institutional Review Board Statement
Experimental protocols were conducted according to the Guide for Care and Use of Laboratory Animals approved by the institutional Animal Care and Use Committee of Nanjing Agricultural University, China (Approval Numbers: 31572403 and 31402075).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets analyzed during the current study are not publicly available due to the individual privacy of Reindeer owners but are available from the authors on reasonable request.
Conflicts of Interest
Conflicts of Interest: Authors declare that there is no conflict of interest.
References
- Kubini, K.; Zachmann, M.; Albers, N.; Hiort, O.; Bettendorf, M.; Wölfle, J.; Bidlingmaier, F.; Klingmüller, D. Basal inhibin B and the testosterone response to human chorionic gonadotropin correlate in prepubertal boys. The Journal of Clinical Endocrinology & Metabolism 2000, 85, 134–138. [Google Scholar]
- Luisi, S.; Florio, P.; Reis, F.M.; Petraglia, F. Inhibins in female and male reproductive physiology: role in gametogenesis, conception, implantation and early pregnancy. Human reproduction update 2005, 11, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Laniyan, D.; Charles-Davies, M.; Fasanmade, A.; Olaniyi, J.; Oyewole, O.; Owolabi, M.; Adebusuyi, J.; Hassan, O.; Ajobo, B.; Ebesunun, M. Inhibin B levels in relation to obesity measures and lipids in males with different numbers of metabolic syndrome components. 2016.
- Weng, Q.; Medan, M.S.; Ren, L.; Watanabe, G.; Arai, K.Y.; Taya, K. Immunolocalization of inhibin/activin subunits in the Shiba goat fetal, neonatal, and adult testes. Journal of Reproduction and Development 2005, 51, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Muttukrishna, S.; Farouk, A.; Sharma, S.; Evans, L.; Groome, N.; Ledger, W.; Sathanandan, M. Serum activin A and follistatin in disorders of spermatogenesis in men. European journal of endocrinology 2001, 144, 425–429. [Google Scholar] [CrossRef]
- Sedqyar, M.; Weng, Q.; Watanabe, G.; Kandiel, M.M.; Takahashi, S.; Suzuki, A.K.; Taneda, S.; Taya, K. Secretion of inhibin in male Japanese quail (Coturnix japonica) from one week of age to sexual maturity. Journal of Reproduction and Development 2008, 54, 100–106. [Google Scholar] [CrossRef]
- Avital-Cohen, N.; Heiblum, R.; Argov, N.; Rosenstrauch, A.; Chaiseha, Y.; Mobarkey, N.; Rozenboim, I. The effect of active immunization against vasoactive intestinal peptide (VIP) and inhibin on reproductive performance of aging White Leghorn roosters. Poultry science 2012, 91, 161–174. [Google Scholar] [CrossRef]
- Larsson, L.I.; Fahrenkrug, J.; Schaffalitzky De Muckadell, O.; Sundler, F.; Håkanson, R.; Rehfeld, J.R. Localization of vasoactive intestinal polypeptide (VIP) to central and peripheral neurons. Proceedings of the National Academy of Sciences of the United States of America 1976, 73, 3197–3200. [Google Scholar] [CrossRef]
- Said, S.I.; Rosenberg, R.N. Vasoactive intestinal polypeptide: abundant immunoreactivity in neural cell lines and normal nervous tissue. Science (New York, N.Y.) 1976, 192, 907–908. [Google Scholar] [CrossRef]
- Rosselin, G.; Maletti, M.; Besson, J.; Rostène, W. A new neuroregulator: the vasoactive intestinal peptide or VIP. Molecular and cellular endocrinology 1982, 27, 243–262. [Google Scholar] [CrossRef]
- Sharp, P.J.; Blache, D. A neuroendocrine model for prolactin as the key mediator of seasonal breeding in birds under long- and short-day photoperiods. Canadian journal of physiology and pharmacology 2003, 81, 350–358. [Google Scholar] [CrossRef]
- Agnese, M.; Rosati, L.; Prisco, M.; Coraggio, F.; Valiante, S.; Scudiero, R.; Laforgia, V.; Andreuccetti, P. The VIP/VPACR system in the reproductive cycle of male lizard Podarcis sicula. General and comparative endocrinology 2014, 205, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Rosati, L.; Prisco, M.; Di Fiore, M.M.; Santillo, A.; Sciarrillo, R.; Valiante, S.; Laforgia, V.; Coraggio, F.; Andreuccetti, P.; Agnese, M. Sex steroid hormone secretion in the wall lizard Podarcis sicula testis: The involvement of VIP. Journal of experimental zoology. Part A, Ecological genetics and physiology 2015, 323, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Agnese, M.; Rosati, L.; Coraggio, F.; Valiante, S.; Prisco, M. Molecular cloning of VIP and distribution of VIP/VPACR system in the testis of Podarcis sicula. Journal of experimental zoology. Part A, Ecological genetics and physiology 2014, 321, 334–347. [Google Scholar] [CrossRef] [PubMed]
- Renthlei, Z.; Yatung, S.; Lalpekhlui, R.; Trivedi, A.K. Seasonality in tropical birds. Journal of experimental zoology. Part A, Ecological and integrative physiology 2022, 337, 952–966. [Google Scholar] [CrossRef]
- Chaiseha, Y.; Halawani, M.E.E. Neuroendocrinology of the Female Turkey Reproductive Cycle. The Journal of Poultry Science 2005, 42, 87–100. [Google Scholar] [CrossRef]
- Gumułka, M.; Rozenboim, I. Effect of breeding stage and photoperiod on gonadal and serotonergic axes in domestic ganders. Theriogenology 2015, 84, 1332–1341. [Google Scholar] [CrossRef]
- Gumułka, M.; Rozenboim, I. Mating activity of domestic geese ganders (Anser anser f. domesticus) during breeding period in relation to age, testosterone and thyroid hormones. Anim Reprod Sci 2013, 142, 183–190. [Google Scholar] [CrossRef]
- Gumułka, M.; Rozenboim, I. Breeding period-associated changes in semen quality, concentrations of LH, PRL, gonadal steroid and thyroid hormones in domestic goose ganders (Anser anser f. domesticus). Anim Reprod Sci 2015, 154, 166–175. [Google Scholar] [CrossRef]
- Shi, Z.D.; Huang, Y.M.; Liu, Z.; Liu, Y.; Li, X.W.; Proudman, J.A.; Yu, R.C. Seasonal and photoperiodic regulation of secretion of hormones associated with reproduction in Magang goose ganders. Domestic animal endocrinology 2007, 32, 190–200. [Google Scholar] [CrossRef]
- Jerysz, A.; Lukaszewicz, E. Effect of dietary selenium and vitamin E on ganders' response to semen collection and ejaculate characteristics. Biological trace element research 2013, 153, 196–204. [Google Scholar] [CrossRef]
- Wen, J.; Li, H.; Wang, H.; Yu, J.; Zhu, T.; Zhang, J.; Li, X.; Jiang, Z.; Ning, Z.; Qu, L. Origins, timing and introgression of domestic geese revealed by whole genome data. Journal of animal science and biotechnology 2023, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Smitz, J. Luteinizing hormone and human chorionic gonadotropin: origins of difference. Molecular and cellular endocrinology 2014, 383, 203–213. [Google Scholar] [CrossRef]
- Acevedo-Rodriguez, A.; Kauffman, A.S.; Cherrington, B.D.; Borges, C.S.; Roepke, T.A.; Laconi, M. Emerging insights into hypothalamic-pituitary-gonadal axis regulation and interaction with stress signalling. Journal of neuroendocrinology 2018, 30, e12590. [Google Scholar] [CrossRef]
- Avital-Cohen, N.; Heiblum, R.; Argov, N.; Rosenstrauch, A.; Chaiseha, Y.; Mobarkey, N.; Rozenboim, I. The effect of active immunization against vasoactive intestinal peptide and inhibin on reproductive performance of young White Leghorn roosters. Poultry science 2011, 90, 2321–2331. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhu, Y.L.; Xue, J.H.; Zhang, S.L.; Ma, Z.; Shi, Z.D. Immunization against inhibin enhances both embryo quantity and quality in Holstein heifers after superovulation and insemination with sex-sorted semen. Theriogenology 2009, 71, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Lovell, T.M.; Knight, P.G.; Groome, N.P.; Gladwell, R.T. Measurement of dimeric inhibins and effects of active immunization against inhibin alpha-subunit on plasma hormones and testis morphology in the developing cockerel. Biol Reprod 2000, 63, 213–221. [Google Scholar] [CrossRef]
- Samir, H.; El Sayed, M.A.; Nagaoka, K.; Sasaki, K.; El-Maaty, A.M.A.; Karen, A.; Abou-Ahmed, M.M.; Watanabe, G. Passive immunization against inhibin increases testicular blood flow in male goats. Theriogenology 2020, 147, 85–91. [Google Scholar] [CrossRef]
- Akhtar, M.F.; Shafiq, M.; Ali, I. Improving gander reproductive efficacy in the context of globally sustainable goose production. Animals 2021, 12, 44. [Google Scholar] [CrossRef]
- Han, Y.; Jiang, T.; Shi, J.; Liu, A.; Liu, L. Review: Role and regulatory mechanism of inhibin in animal reproductive system. Theriogenology 2023, 202, 10–20. [Google Scholar] [CrossRef]
- Wang, J.M.; Li, Z.F.; Yang, W.X.; Tan, F.Q. Follicle-stimulating hormone signaling in Sertoli cells: a licence to the early stages of spermatogenesis. Reproductive biology and endocrinology : RB&E 2022, 20, 97. [Google Scholar] [CrossRef]
- Themmen, A.P.; Huhtaniemi, I.T. Mutations of gonadotropins and gonadotropin receptors: elucidating the physiology and pathophysiology of pituitary-gonadal function. Endocrine reviews 2000, 21, 551–583. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.T.; O’Hara, L.; Smith, L.B. Gonadotropin and steroid hormone control of spermatogonial differentiation. The Biology of Mammalian Spermatogonia 2017, 147–178. [Google Scholar]
- Wang, S.-L.; Han, L.; Ahmad, S.; Cao, S.-X.; Xue, L.-Q.; Xing, Z.-F.; Feng, J.-Z.; Liang, A.-X.; Yang, L.-G. Effect of a DNA vaccine harboring two copies of inhibin α (1-32) fragments on immune response, hormone concentrations and reproductive performance in rats. Theriogenology 2012, 78, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.A.; Cui, X.; Schmittling, R.J.; Sanchez-Perez, L.; Snyder, D.J.; Congdon, K.L.; Archer, G.E.; Desjardins, A.; Friedman, A.H.; Friedman, H.S. Monoclonal antibody blockade of IL-2 receptor α during lymphopenia selectively depletes regulatory T cells in mice and humans. Blood, The Journal of the American Society of Hematology 2011, 118, 3003–3012. [Google Scholar] [CrossRef]
- Shi, Z.; Tian, Y.; Wu, W.; Wang, Z. Controlling reproductive seasonality in the geese: a review. World's Poultry Science Journal 2008, 64, 343–355. [Google Scholar] [CrossRef]
- Tolla, E.; Pérez, J.H.; Dunn, I.C.; Meddle, S.L.; Stevenson, T.J. Neuroendocrine Regulation of Seasonal Reproduction. In Oxford Research Encyclopedia of Neuroscience; 2019. [Google Scholar]
- Singh, D.; Reed, S.; Kimmitt, A.; Alford, K.; Stricker, C.; Ketterson, E. Breeding at higher latitude as measured by stable isotope is associated with higher photoperiod threshold and delayed reproductive development in a songbird. bioRxiv 2019, 789008. [Google Scholar]
- Drobniak, S.M.; Gudowska, A. Seasonality, diet, and physiology as a predominant control factors of the moult dynamics in birds–a meta-analysis. 2020.
- Meng, J.; Feng, J.H.; Xiao, L.; Zhou, W.; Zhang, H.; Lan, X.; Wang, S. Active immunization with inhibin DNA vaccine promotes spermatogenesis and testicular development in rats. Journal of Applied Animal Research 2024, 52, 2360408. [Google Scholar] [CrossRef]
- VAN DISSEL-EMILIANI, F.M.; GROOTENHUIS, A.J.; JONG, F.H.D.; ROOIJ, D.G.D. Inhibin reduces spermatogonial numbers in testes of adult mice and Chinese hamsters. Endocrinology 1989, 125, 1898–1903. [Google Scholar] [CrossRef]
- Akhtar, M.F.; Ahmad, E.; Ali, I.; Shafiq, M.; Chen, Z. The Effect of Inhibin Immunization in Seminiferous Epithelium of Yangzhou Goose Ganders: A Histological Study. Animals 2021, 11, 2801. [Google Scholar] [CrossRef]
- Banerjee, S.; Chaturvedi, C.M. Testicular atrophy and reproductive quiescence in photorefractory and scotosensitive quail: involvement of hypothalamic deep brain photoreceptors and GnRH-GnIH system. Journal of Photochemistry and Photobiology B: Biology 2017, 175, 254–268. [Google Scholar] [CrossRef]
- Akhtar, M.F.; Wei, Q.; Zhu, H.; Chen, Z.; Ahmad, E.; Zhendan, S.; Shi, F. The role of active immunization against inhibin α-subunit on testicular development, testosterone concentration and relevant genes expressions in testis, hypothalamus and pituitary glands in Yangzhou goose ganders. Theriogenology 2019, 128, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Voge, J.; Wheaton, J. Effects of immunization against two inhibin antigens on daily sperm production and hormone concentrations in ram lambs. Journal of animal science 2007, 85, 3249–3255. [Google Scholar] [CrossRef] [PubMed]
- Sivinski, J. Sperm in competition. Sperm competition and the evolution of animal mating systems 1984, 85–115. [Google Scholar]
- Cai, K.; Hua, G.; Ahmad, S.; Liang, A.; Han, L.; Wu, C.; Yang, F.; Yang, L. Action mechanism of inhibin α-subunit on the development of Sertoli cells and first wave of spermatogenesis in mice. PLoS One 2011, 6, e25585. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, Z.; Shao, X.; Yu, J.; Wei, C.; Dai, Z.; Shi, Z. Reproductive axis gene regulation during photostimulation and photorefractoriness in Yangzhou goose ganders. Frontiers in zoology 2017, 14, 1–15. [Google Scholar] [CrossRef]
- Mao, D.; Bai, W.; Hui, F.; Yang, L.; Cao, S.; Xu, Y. Effect of inhibin gene immunization on antibody production and reproductive performance in Partridge Shank hens. Theriogenology 2016, 85, 1037–1044. [Google Scholar] [CrossRef]
- Gumułka, M.; Rozenboim, I. Breeding period-associated changes in semen quality, concentrations of LH, PRL, gonadal steroid and thyroid hormones in domestic goose ganders (Anser anser f. domesticus). Animal reproduction science 2015, 154, 166–175. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Mean body weight (BW) of Yangzhou ganders.
| Day of Age | Control (Kg) | INH Immunized (Kg) |
|---|---|---|
| 181 | 4.73 | 4.68 |
| 200 | 4.41 | 4.54 |
| 227 | 4.56 | 4.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



