
Submitted:
30 January 2025
Posted:
31 January 2025
You are already at the latest version
Abstract
Present study was designed to investigate the potential role of inhibin immunization on plasma hormone concentration and testicular histoarchitecture of Dezhou donkeys in the non breeding season (November-February). For this purpose, adult Dezhou donkeys (n=30), were equally divided into groups A, B, and C. Group A was actively immunized with 3 mg inhibin, group B with (1.5 mg), and group C was immunized with Bovine serum albumin (BSA) and served as a control. All animals in groups A and B were given a primary dose of inhibin (INH) antigen i.e. 3 mg and 1.5 mg on day 1 and followed by a booster dose on the 23rd day of the experiment. Blood samples were collected on 21st, 28th, 34th, and 40th day of experiment. Primary and booster INH immunization (3 mg and 1.5 mg) slightly elevated plasma hormone concentrations of FSH, LH, Testosterone, and Activin A. The number of spermatogonia significantly increased on 28th day in group A. Inhibin immunization also caused apoptosis in testicular histoarchitecture. In conclusion, Inhibin immunzition can potentially improve the reproductive efficiency of Dezhou donkeys in nonbreeding season by elevating plasma hormone concentrations of FSH, LH, AMH, and Activin A.
Keywords:
Inhibin immunization
; plasma hormone concentrations
; spermatogenesis
; non-breeding seasonality
1. Introduction
China has a rich history of raising donkeys. In past years, the donkey population in China significantly declined. Still now it’s increasing owing to the high nutritional value of meat, milk, and most of all, a very popular traditional Chinese medicine (E-jiao) is prepared from donkey skin that has various health-beneficial properties [1,2]. Among 24 donkey breeds in China, the Dezhou breed is quite popular in commercial farming due to itspredominant black hair with a straight back and waist, arch of chest rib, and round, strong hooves [3,4]. With emerging technologies and advancements in the meat sector, consumption of beef, mutton, and chicken meat has their market value. However, the importance and consumption of donkey meat and milk are indispensable. Dezhou doney is further divided into Sanfen and Wutou breeds. They are covered with black hair, however, the Sanfen donkey has white hair on the eyes, around the nose, and under the belly [1]. Seasonal reproduction is usual in mammals [5]. The effect of seasonality on the reproductive efficiency of donkeys is highly controversial. It is influenced by various factors including breed, environment, nutrition, and health [6]. Long or short days highly influence the reproduction of seasonal breeders like donkeys. With approaching shorter day length having minimal sunlight, the semen quality of male animals also declines. Seasonal alterations affect on hormonal regulation, gametogenesis, FSH, and testosterone secretions [7]. The testicular steroidogenesis, spermatogenesis and endocrine functions of follicle-stimulating hormone (FSH) are linked with inhibin and activin [7]. Based on the available data, we can hypothesize that donkeys experience reproductive quiescence throughout the winter months when sunshine hours are reduced and that Jack's reproductive efficacy is also downregulated during these times.
Inhibin (INH) is a glycoprotein hormone secreted by gonads (Sertoli cells in males and granulose cells in females). Inhibin is one of the major players on the hypothalamus- pituitary gonadal (HPG) axis [8]. Inhibin is a 31-34 kDa heterodimeric glycoprotein, which forms a disulphide-linked dimer that shares a common α-subunit and differs in β-subunit (βA-subunit and βB-subunit), βA in inhibin A (αβA) and βB in inhibin B (αβB). Inhibin is a member of the transforming growth factor β (TGF-β) superfamily and has been proposed as an autocrine/paracrine factor that modulates follicular growth, atresia, gonadotropin responsiveness, and steroidogenesis [8]. Inbibin is a negative feedback regulator of FSH, i.e. it down-regulates FSH secretion in males and females [9]. In most cases, active or passive INH immunization lowers the negative feedback effect of INH and results in the up-regulation of FSH [10]. In past years, immunization against inhibin proved to be a promising tool in improving the reproductive efficiency of animals. Inhibin immunization and progesterone (P4) treatment elevated ovarian follicle development which will subsequently enhance the early embryo developments in Holstein cows [11]. Active immunization against INH improved the fresh and post-thaw semen quality of Beetal bucks [12]. Inhibin immunization at the dose of 0.5 mg greatly improved the diameter and size of pre-ovulatory follicles and ovarian follicles [13]. Immunization against INH improves fertility in cattle [14]. Active immunization against INH up-regulates the spermatogenesis and testicular development in rats [15]. INH immunzation in Yangzhou goose ganders elevated the testicular weights [8]. INH immunization elevates the FSH concentrations without affecting the concentration of LH and testosterone [16]. The potential for INH vaccination to raise FSH and testosterone levels is evident from these data. We hypothesize that semen quality can be enhanced even during non breeding seasons in Dezhou jacks if INH immunization can help increase plasma FSH, LH, and testosterone concentrations in these animals. Therefore, the current study was designed to investigate the potential role of inhibin (INH) immunization during low-breeding seasonality (November-February) on plasma hormone concentrations of FSH, LH, T, P4 Progesterone, AMH, and Activin A.
2. Materials and Methods
2.1. Inhibin Immunogen Preparation
A porcine recombinant inhibin α-subunit protein was expressed in a prokaryotic expression system in E-coli strain BL21 (DE3) and was utilized as the inhibin antigen. The recombinant protein contained 175 amino acid residues, including a 41-residue leading sequence derived from the expression plasmid pRSETA (Invitrogen, Carlsbad, CA, USA). Also, it included 134-residue porcine inhibin α-subunit mature peptide. After purifying recombinant protein, it was homogenized with a mineral oil adjuvant, composed of a 1:2 (v/v) mixture of water and grade 10 white oil for injections (Hangzhou Refinery, Hangzhou, China), at the final concentration of immunogen was 1 mg/mL.
2.2. Experimental Design
This study was approved by the Research Committee of the Animal Policy and Welfare Committee of Liaocheng University (No. LC2019-1). The care and use of laboratory animals fully comply with local animal welfare laws, guidelines, and policies.
Figure 1.
(A) Dezhou donkey body weight at Ist, 38th, 63rd and 78th day of experiment. Each bar represents the mean value of six determinations including the standard error. *, *** indicate statistical significance based on P < 0.05 and P < 0.001 respectively. (B) Anti-Inhibin antibody titers in Inhibin Immunized Group A  , Group B
, Group B  and control Group C
and control Group C  of Dezhou donkeys at 21, 28,34 and 40 days of experiment. Vertical bars represent standard error of mean (SEM). The values with ** indicate the difference (P < 0.001) whereas the values with * indicate difference (P < 0.05) between groups A, B and C. Arrows indicate primary and booster Inhibin (INH) immunziation at Ist and 23rd day of experiment.
of Dezhou donkeys at 21, 28,34 and 40 days of experiment. Vertical bars represent standard error of mean (SEM). The values with ** indicate the difference (P < 0.001) whereas the values with * indicate difference (P < 0.05) between groups A, B and C. Arrows indicate primary and booster Inhibin (INH) immunziation at Ist and 23rd day of experiment.
, Group B and control Group C of Dezhou donkeys at 21, 28,34 and 40 days of experiment. Vertical bars represent standard error of mean (SEM). The values with ** indicate the difference (P < 0.001) whereas the values with * indicate difference (P < 0.05) between groups A, B and C. Arrows indicate primary and booster Inhibin (INH) immunziation at Ist and 23rd day of experiment.
Figure 1.
(A) Dezhou donkey body weight at Ist, 38th, 63rd and 78th day of experiment. Each bar represents the mean value of six determinations including the standard error. *, *** indicate statistical significance based on P < 0.05 and P < 0.001 respectively. (B) Anti-Inhibin antibody titers in Inhibin Immunized Group A , Group B and control Group C of Dezhou donkeys at 21, 28,34 and 40 days of experiment. Vertical bars represent standard error of mean (SEM). The values with ** indicate the difference (P < 0.001) whereas the values with * indicate difference (P < 0.05) between groups A, B and C. Arrows indicate primary and booster Inhibin (INH) immunziation at Ist and 23rd day of experiment.
, Group B and control Group C of Dezhou donkeys at 21, 28,34 and 40 days of experiment. Vertical bars represent standard error of mean (SEM). The values with ** indicate the difference (P < 0.001) whereas the values with * indicate difference (P < 0.05) between groups A, B and C. Arrows indicate primary and booster Inhibin (INH) immunziation at Ist and 23rd day of experiment.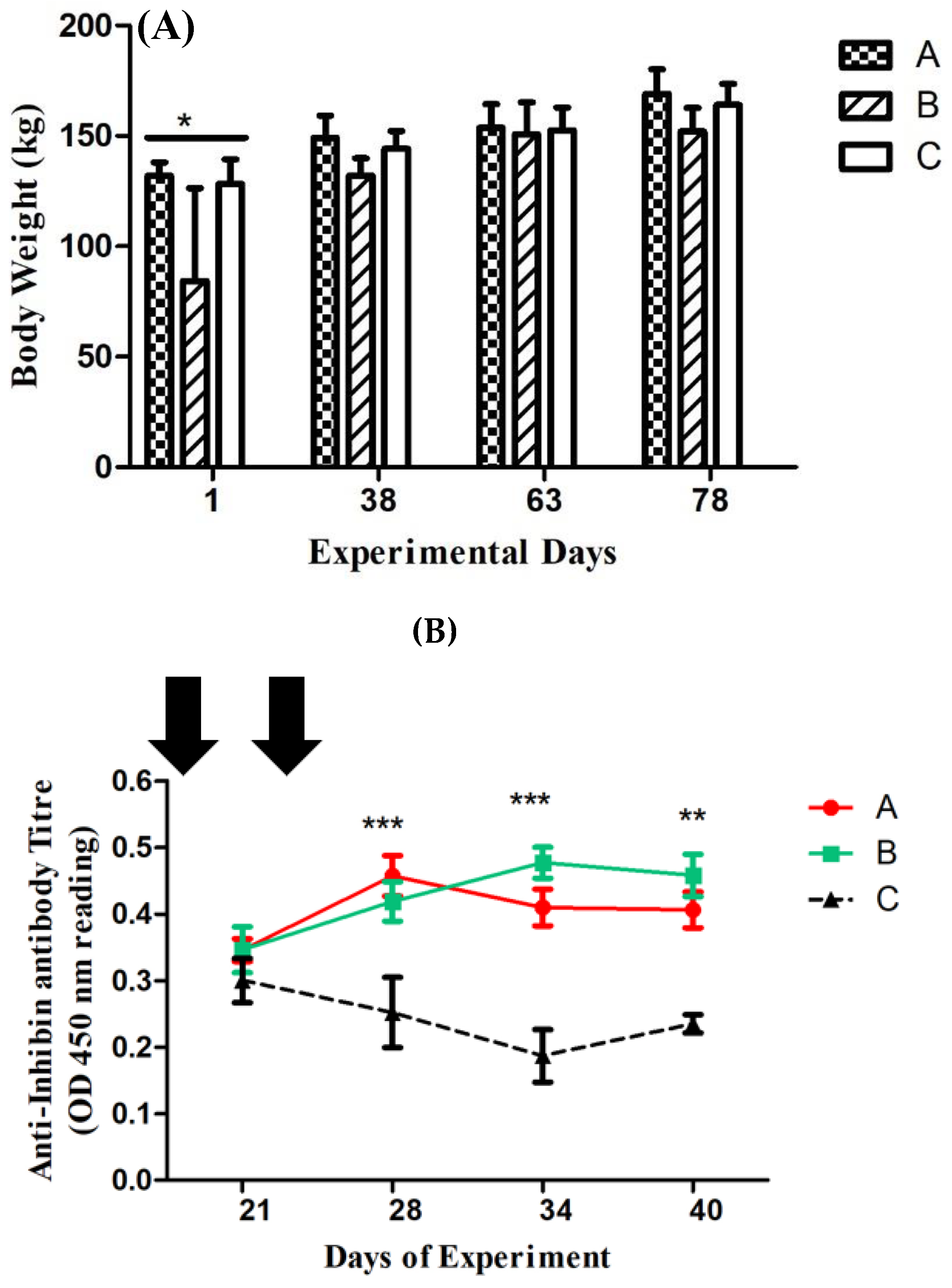
All animals were offered silage and had free access to drinking water. The current study was conducted from 27th November to 27th February (90 days) at Liaocheng Wanshixing Breeding Co. Ltd (E 115º and N 36º), Liaocheng, Shandong Province, China. Adult Dezhou jacks, n=30, having the same genetic origin and an average age of 2.5 + 0.50 years, were randomly divided into three groups i.e. A, B, and C. Each group had 10 animals each. All animals in experimental groups were intramuscularly (i.m) injected with different concentrations of inhibin. Animals in group A were immunized with 3 mg of inhibin immunogen; group B was immunized with 1.5 mg of inhibin and group C kept as the control group received a bovine serum albumin (BSA) injection. Inhibin was injected on the 1st and 23rd days of the experiment in groups A and B.
2.3. Measurement of Body Weight, Blood, and Testes Tissue Collection
Body weights of all animals in groups A, B, and C were measured on 21Ist, 38th, 34th and 40th day of the experiment as shown in figure 1 (A). Blood samples were collected on the 21st, 38th, 34th, and 40th day of the experiment via the jugular vein into heparinized tubes. Within three hours of sample collection, plasma was separated from the blood by centrifugation at 1000 × g and kept at 20 ⁰C until analysis. The testes tissues from groups A, B, and C were taken from the slaughterhouse at the end of the experiment. Testes were frozen in liquid nitrogen and kept at -80 ⁰C as soon as they were collected
2.4. Antibody Titer
Standard ELISA was used to analyze inhibin antibody titer in donkey plasma. The α -inhibin recombinant fusion protein was used to coat a 96-well microtiter plate (0.5 µg/well in 100 µl). Then, a 100 μl plasma sample (1:1200 dilutions with 5% skimmed milk) was added to each well and then incubated at room temperature to bind anti-inhibin antibodies with the coated inhibin fusion protein. The bound antibodies were further labeled by incubation with horseradish peroxidase (HRP) conjugated rabbit anti-bovine antibody (SantaCruzBiotechnology, SantaCruz, CA, USA). Finally, color development was preceded by chromogen tetramethyl Benzedrine (Sigma) solution containing 0.03% H2O2, and terminated after appropriate with the addition of 2% H2SO4. Optical absorbance was taken at 450 nm on the EON Bioteke spectrophotometer to represent inhibin antibodies titer for both control and immunized donkeys.
2.5. Plasma Hormone Concentrations
Plasma concentrations were performed by ELISA using quantitative kits (MEIMIAN from Jiangsu Meimian Industrial Co., Ltd). Assays were performed using protocols provided by the kit supplier. For FSH, assay sensitivity was 0.075 U/L. Both inter and intra-assay coefficients were below 10%. The detection range was 0.3 U/L-18 U/L. For plasma LH, assay sensitivity was 0.005 ng/ml. Inter and intra-assay co-efficients were below 10%. Detection range was 0.002 ng/ml-0.05 ng/ml. For progesterone (P4), assay sensitivity was 5 pmol/L. Inter and intra-assay coefficients were below 10%. The detection range was 20 pmol/L - 800 pmol/L. For testosterone, assay sensitivity was 0.02 ng/ml. Inter and intra-assay coefficients were below 10%. The detection range was 0.094 ng/ml- 3.77 ng/ml. For AMH, assay sensitivity was 0.05 ng/ml. Inter and intra-assay coefficients were below 10%. The detection range was 0.2 ng/ml – 8.5 ng/ml. For Activin (A), assay sensitivity was 0.4 ng/ml. Inter and intra-assay coefficients were below 10%. The detection range was 1.6 ng/ml – 65 ng/ml.
2.6. Microscopy Performance
To observe the effect of INH immunization on testicular histoarchitecture, testicular biopsy was performed on 21, 28, 34th and 40th day of experiment by randonly selecting two aniamls from each group. Testicular biopsy was performed by following procedure as performed by Mohammadreza Baqerkhani et. al [17]. A slice of left testicular tissue (0.125 cm3) was taken and embedded in a 10% neutral buffered formalin solution for 24 hours to observe histological alterations in seminiferous tubules. Histological analysis was performed with an automated tissue processor (LECIA RM 2235). After fixation, tissues were dehydrated in alcohol of increasing concentrations, i.e., 70%, 80%, 90%, 100% and absolute alcohol. After dehydration, testis tissues were cleared in xylene embedded in paraffin wax. Testis tissues were cut perpendicular in 5 μm thickness to testicular long axis. Slides were then mounted on glass slides and were stained using hematoxylin and eosin (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). Many histomorphometric metrics of the seminiferous epithelium have been measured, such as diameter of seminiferous tubules (ST), ST/field, luminal tubular diameter; as well as number of spermatogonia, spermatocytes, elongated spermatids were calculated. Moreover, epithelium height was determined as described previously [18]. All ST were observed under a bright field light microscope (LEICA Dmi8, Germany) with 40X(25μm) magnifications.
2.7. Statistical Analysis
The data were analyzed by using SPSS (Version 20.0 Armonk NY, USA) and Graph Pad Prism (Version 5.0). The Kolmogorov-Smirnov goodness-of-ft test was applied to determine normality, The data were transformed to logarithms if not normally distributed and then re-tested for normality before analysis. Then, two-way ANOVA was applied to compare mean values. All the values were expressed as mean + standard error of the mean (SEM). The differences across groups at various time points were analyzed Bonferroni post-test. The probability levels P<0.05 or 0.001 were set to determine significant difference among groups.
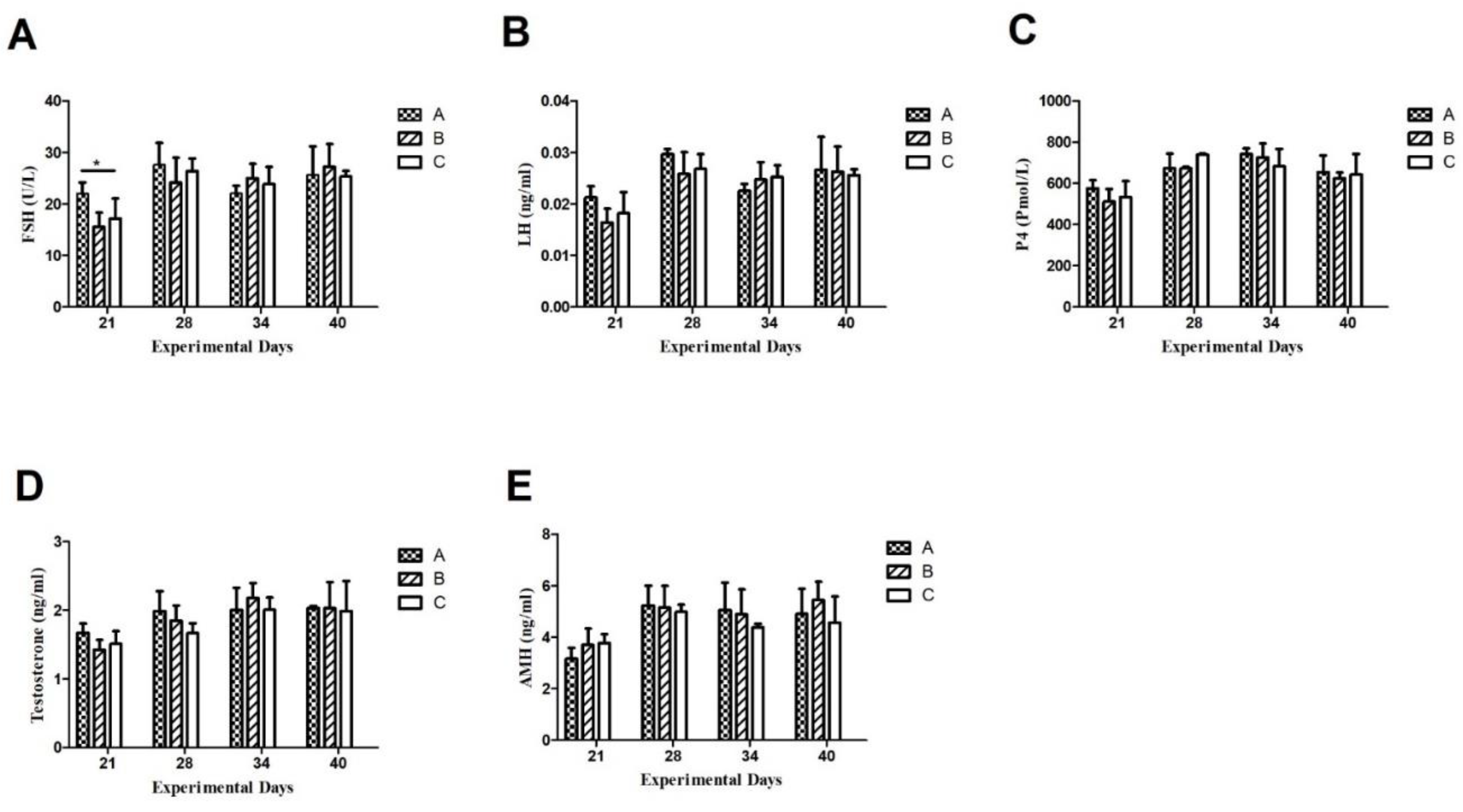
Figure 2.
Plasma concentrations of Follicle Stimulating Hormone (A), Luteining Hormone (B), progesterone (C), testosteterone (D), Anti-Mullerian hormone (E), and activin-A (F). Data are shown as mean values ± standard error of the mean. *, ** and *** indicate difference at P < 0.05, P < 0.01and P < 0.001, respectively between the groups.
Figure 2.
Plasma concentrations of Follicle Stimulating Hormone (A), Luteining Hormone (B), progesterone (C), testosteterone (D), Anti-Mullerian hormone (E), and activin-A (F). Data are shown as mean values ± standard error of the mean. *, ** and *** indicate difference at P < 0.05, P < 0.01and P < 0.001, respectively between the groups.
3. Results
3.1. Body Weights
Throughout the experimental period (on 38th, 63rd, and 78th day of the experiment), body weights in all groups exhibited non-significant difference (as shown in figure 1A) except on day 21, at which body weights of group A and C animals were significantly higher (P<0.05) as compared to group B animals.
3.2. Anti-Inhibin Antibody Titer
On day 21st, following the first inhibin immunization, OD values in group A initially did not exhibit any significant differences as shown in figure 1(B). It then increased to 0.45 and 0.40 (P<0.001) in groups A and B following the second booster dose of immunization on day 23rd of the experiment, compared to 0.25 in the control group. Antibodies titerrs in groups B and A were substantially greater (P<0.001) than those in the control group on the 34th day of the experiment. Titer of group B was 0.49, but somewhat lower (i.e., 0.40 of OD 450 nm). On day 40th of the experiment groups A and B showed a similar pattern (P<0.001). The OD=450 nm was considered the base noise level of the assay.
3.3. Plasma Hormone Concentrations
3.3.1. Follicle Stimulating Hormone (FSH)
Throughout the experimental period, the plasma FSH concentrations did not change significantly. After 1st Inhibin immunization on day 1, the FSH concentration in group A was at a base level of 22 U/L on the 21st day of the experiment and it was up-regulated to peak levels of 29 U/L on the 28th day of the experiment and remained higher as compared to group B (1.5 mg INH) and C (Control) on 21 and 28th day of experiment i.e 15 U/L and 24 U/L in group B and 17 U/L and 27 U/L, respectively. After booster Inhibin immunization, FSH in group B remained at almost the same level i.e. 22 U/L on the 38th day of the experiment but slightly elevated to 28 U/L on the 40th day of the experiment. Plasma FSH levels remained slightly higher on 21 and 28th day of the experiment i.e. 17 U/L and 27 U/L as compared to group B.
3.3.2. Luteinizing Hormone (LH)
Plasma hormone concentrations of LH (ng/ml) remained higher on the 21st and 28th day of the experiment in group A as compared to B and C i.e. 0.021 (ng/ml) and 0.03 (ng/ml), respectively. Throughout the experimental period, the pattern of LH (ng/ml) concentration remained the same as between groups B and C even after booster immunization with inhibin shot on day 21. There existed non-significant differences among groups.
3.3.3. Progesterone (P4)
Plasma progesterone (Pmol/L) concentrations displayed similar ascending and descending patterns throughout the experimental period with the except of day 34, where P4 (Pmol/L) concentration was slightly elevated as compared to group C.
3.3.4. Testosterone (T)
Plasma hormone concentrations of testosterone (T) remained higher i.e. 1.8 (ng/ml) and 2 (mg/ml) in group A on days 21 and 28th as compared to groups B and C. On the 34th day of the experiment, the plasma T (ng/ml) in group was 2.2 (ng/ml), while it was 2 (mg/ml) in groups A and C. On the 40th day, plasma T (ng/ml) was almost the same i.e. (2 ng/ml) in all groups. Moreover, plasma T (ng/ml) declined to 2 (ng/ml) on the 40th day as compared to 2.2 (ng/ml) in group B.
3.3.5. AntiMullerian Hormone
Plasma AMH concentrations in group A were 3 (ng/ml) on day 21 in group A, while it was 3.85 (ng/ml) in group B. After booster INH immunization, it elevated to 5 ng/ml in group A and was almost 5 ng/ml in group B. On day 34, the AMH concentration was 4.9 ng/ml in group A, while it was slightly lowered in group B i.e. 4.8 ng/ml. On day 40th of the experiment, plasma AMH was elevated to 5.9 (ng/ml) and it remained lower in groups A and C i.e. 4.7 (ng/ml) and 4.2 (ng/ml), respectively. Throughout the experimental period, the plasma AMH concentrations remained lower in group C (control) as compared to groups A and B.
3.3.6. Activin A
Plasma Activin A concentrations showed similar ascending and descending patterns to AMH throughout the experimental period. After 1st INH immunization, plasma Actvin A concentration was 6 (ng/ml) in group A, and 7 (ng/ml) in group B and 8 (ng/ml) in group C. On day 28th, it elevated to 11 (ng/ml) and 9 (ng/ml) in groups A, B and C, respectively. Activin A concentration increased significantly after the 2nd INH immunization on the 28th day as compared to the 21Ist day of the experiment. However, on day 40, their plasma Activin-A concentrations were the same in groups A, B, and C i.e. 9 (ng/ml).
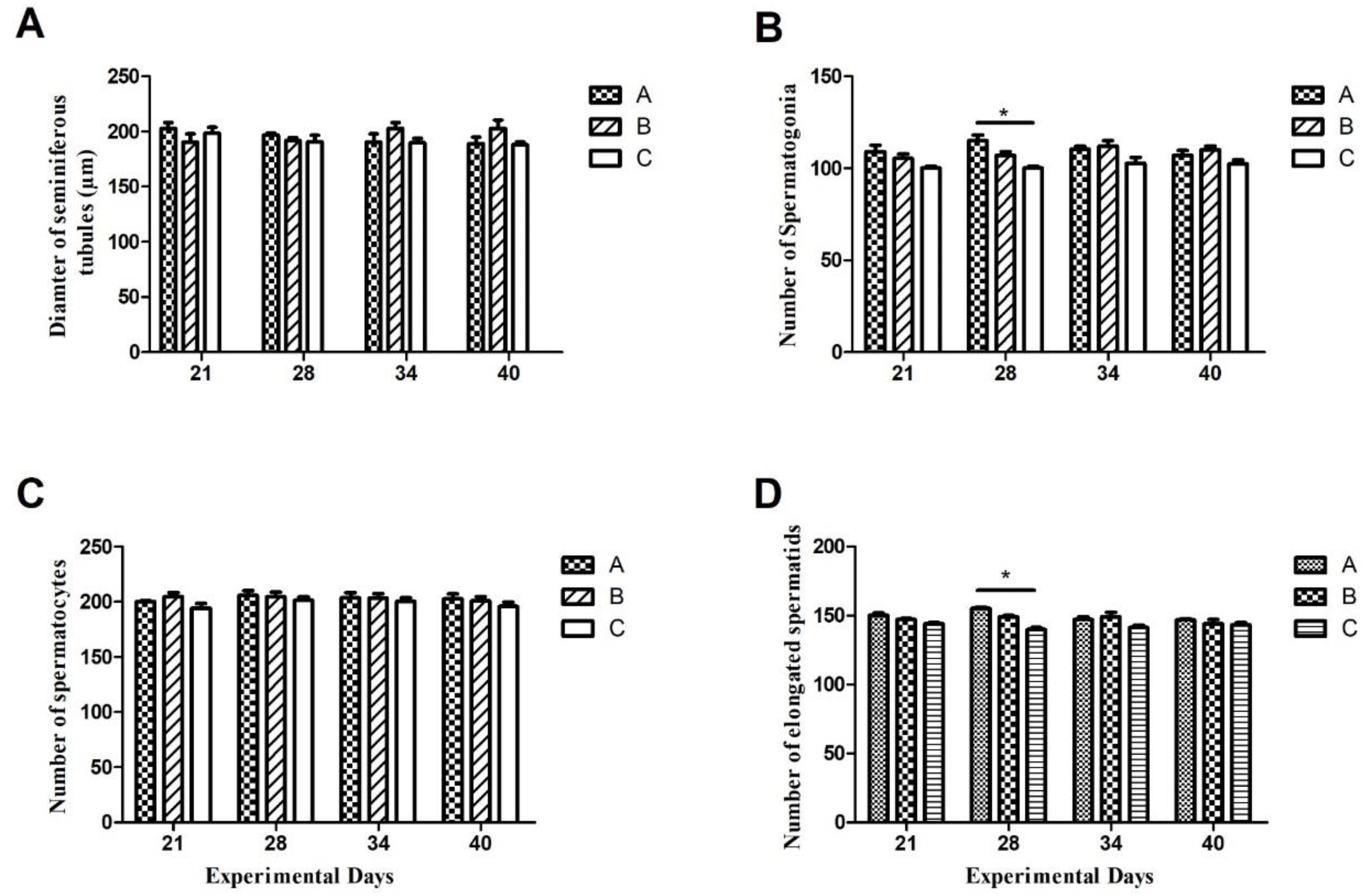
3.4. Germ cells Count and Variations in Seminiferous Epithelium
From figure 4 we can see that, throughout the experimental period, there existed non-significant differences among germ cells (spermatogonia, spermatocytes, and number of elongated spermatids), except on day 28th, on which the number of spermatogonia and number of elongated spermatids were significantly higher in group A as compared to group C. Also, empty lumen and apoptosis were observed in INH immunized groups (A and B) while normal lumen was observed in control group C.
4. Discussion
In the present study, INH immunization's effects on the levels of FSH, LH, P4, Testosterone, AMH, Activin A, anti-inhibin antibody titer, seminiferous tubule diameter, and the number of germ cells (spermatogonia, spermatocytes, and elongated) spermatids in adult Dezhou donkeys (jacks) have thoroughly described. Results of 1st immunization significantly lower the body weight of animals in group B as compared to groups A and C. Afterwards, throughout the experiment, a non-significant difference was observed among all groups after primary and booster immunizations. However, our data suggests that the inhibin protein did not significantly affect the body weight indicating modulation of inhibin levels at least within the parameters measured in this study does not influence overall body weight.
Previous studies in various species suggest that the inhibin’s role has been predominantly focused on its involvement in reproductive functions and gonadotropin regulation, rather than energy metabolism or growth [19]. Moreover, gonads produce proteins in TGFβ family members (Inhibin A and B) that suppress the secretion of FSH without affecting LH secretion [20]. Conceivably, inhibin is primarily known for its role in inhibiting the secretion of follicle-stimulating hormone (FSH) and regulating the function of the gonads, rather than acting as a direct modulator of body weight or adiposity. Furthermore, inhibin is a glycoprotein hormone that is secreted by gonads (testes in males and ovaries in females) which inhibits the secretion and synthesis of FSH from the anterior pituitary gland [19]. Our findings also reinforce the theory that INH is a key modulator on the hypothalamus-pituitary gonadal axis only, rather than affecting metabolic activities in animals.
In the present study, inhibin antibody titer tended to increase in groups A and B after initial and booster INH immunization, indicating that INH immunization had triggered innate responses in these experimental groups. The reason may be that inhibin antibody titer is associated with testosterone concentrations and follows similar ascending and descending patterns in all groups. In our other study, though it was in Yanghou geese, inhibin antibody titer was associated with plasma Testosterone concentrations [8].
Semen quality in male animals can be improved by enhancing plasma hormone concentrations of FSH. So, we can either enhance its secretion from FSH or mitigate its negative effects on testes. In our study, we had attempted to elevate plasma FSH concentrations. In several studies, inhibin immunization has been proven as a promising tool in enhancing FSH secretion by downregulating negative feedback of endogenous inhibin[8,10,21-25]. In our outcomes, the FSH upregulation on 21Ist and 28th day of the experiment enunciates that exogenous inhibin immunization had suppressed the effect of endogenous inhibin that resulted in elevated FSH secretion in groups A as compared to group B and C. On the hypothalamus-pituitary-testiscualr axis, FSH and testosterone secretion are downregulated or upregulated by inhibin, which plays a vital role in the up or down-regulation of spermatogenesis [26]. Moreover, inhibin immunization reduces the effect of endogenous inhibin and upregulates pituitary activity and FSH [14]. Furthermore, inhibin immunization also elevated the secretion of activin A [10,21,22]. Activin is the antagonist of inhibin, and elevated levels of activin may be due to activin secretion from Sertoli cells [27-29]. Both FSH and activin work together to regulate the function of Sertoli cells in the testes to regulate spermatogenesis [29-31]. Moreover, slightly upregulated FSH and activin may have stimulated Sertoli cell development. Similarly, FSH activates the cAMP/PKA pathway, which promotes the gene expression related to spermatogenesis and Sertoli cell function [32,33]. Ultimately, activin activates the Smad pathway, enhancing proliferation and survival [34,35]. To our knowledge, the present study speculates that both pathways integrate PI3K/Akt and MAPK/ERK signaling for Sertoli cell growth and differentiation.
In our present findings, plasma LH concentrations elevated after the first INH immunization, and after the first booster INH injection, it was still higher among all groups on day 28th. At the same time, Testosterone concentration followed ascending and descending patterns of plasma LH concentrations. This was initially a surprising result in light of other findings in which the LH concentration elevated after INH immunzation in goats [13], dairy cows [21], and sows [36]. However, elevated levels of plasma LH after INH immunization can be species-specific. However, our previous findings elaborate that the LH-β did not increase significantly after INH immunization in Yangzhou ganders [8]. However, on the 34th and 40th day of the experiment, the plasma LH seemed to be slightly lower and upregulated and exhibit non-significant differences. It is also possible that INH immunization did not affect plasma LH concentrations and our results, INH did not improve plasma LH concentrations, significantly. In addition, the LHβ remains unaffected after INH immunization in aging white leghorn roosters[37]. As such, inhibin immunization did not affect the plasma LH concentrations in Shiba male goats [38]. On the hypothalamus pituitary gonadal axis (HPG), LH directly acts on Leydig cells that control T [39-41]. Our findings also reinforce previous outcomes.
Antimullerian hormone (AMH), a crucial marker of Sertoli cells, is a member of the TGF-β superfamily. AMH plays an important role in Sertoli cell development [42]. AMH has a vital function in testes development. According to our current findings, there was no discernible difference between experimental groups A (3 mg INH) and B (1.5 mg) and group C in the impact of INH immunization on plasma AMH.AMH is produced in fetal and postnatal Sertoli cells and its production lowers with puberty in adult males[43,44]. The development of Sertoli cells depends on the Sertoli cell marker Sox9 and it highly regulates the production of [45]. In our previous findings [8], we noticed that INH immunization did not affect gene expression levels of Sox9 and AMH in adult Yangzhou ganders. Our current findings are per previous one in that, in adult animals, INH immunization does not affect AMH. So, we can speculate that if Sox9 regulation remains lower after INH immunization AMH must also remain lower.
Inhibin primarily acts to inhibit the secretion of follicle-stimulating hormone (FSH) and is thought to modulate gonadal function. In males, P4 is produced primarily in the testes by Leydig cells and Sertoli cells[46,47], where it serves as a precursor to other steroid hormones, including testosterone.It is possible that its effects on P4 production in male donkeys were not as pronounced as expected. Our findings suggest that, despite INH immunization, the P4 production pathways may be resistant to changes in FSH levels, or that compensatory mechanisms within the hypothalamic-pituitary-testicular axis may have prevented significant alterations in P4 concentrations.
Figure 3 enunciates alterations in testicular histoarchitecture in control group C and INH immunized groups A and B. In INH immunized groups A and B, there is a clear sign of apoptosis and Sertoli cell vacuolation. According to recent data, the phenomenon of steroidogenesis and breeding seasonality enhances the chance of apoptosis in the normal testis of seasonal breeder animals [48-50]. In our previous work, in which we had immunized Yangzhou gander with INH protein, similar findings were observed, i.e. apoptosis after INH immunization [8,18]. This may be possible due to the immunized group experienceing oxidative stress because of exogenous INH vaccination surpassing indigenous INH, which leads to high FSH levels and causes Sertoli cell vacuolation. It is obvious that hormonal imbalance may cause disruption in testicular histoartichecture, including Sertoli cell vacuolation. Animals in the control group showed that some part of the lumen was empty too but there seemed no Sertoli cell vacuolation. However, signs of apoptotic cells may not be merely by INH immunization. Because they are seasonal breeders, there can also be a change in the testicular architecture that results in Sertoli cell vacuolation and apoptosis. Further, other studies reported that the histology of the testis alters with the breeding stage and age of maturity [18].
Figure 3.
Histological sections from donkeys were collected at 90th day of experiment and stained with hematoxylin and eosin. Arrow with spiral tail: Spermatogonia, Arrow head: Seroli cell vacuolation, Arrow with tail: Empty lumen, Arrow with thick tail: elongated spermatids. Each group (n=10). All images were obeserved at 75 μm at 20X.
Figure 3.
Histological sections from donkeys were collected at 90th day of experiment and stained with hematoxylin and eosin. Arrow with spiral tail: Spermatogonia, Arrow head: Seroli cell vacuolation, Arrow with tail: Empty lumen, Arrow with thick tail: elongated spermatids. Each group (n=10). All images were obeserved at 75 μm at 20X.
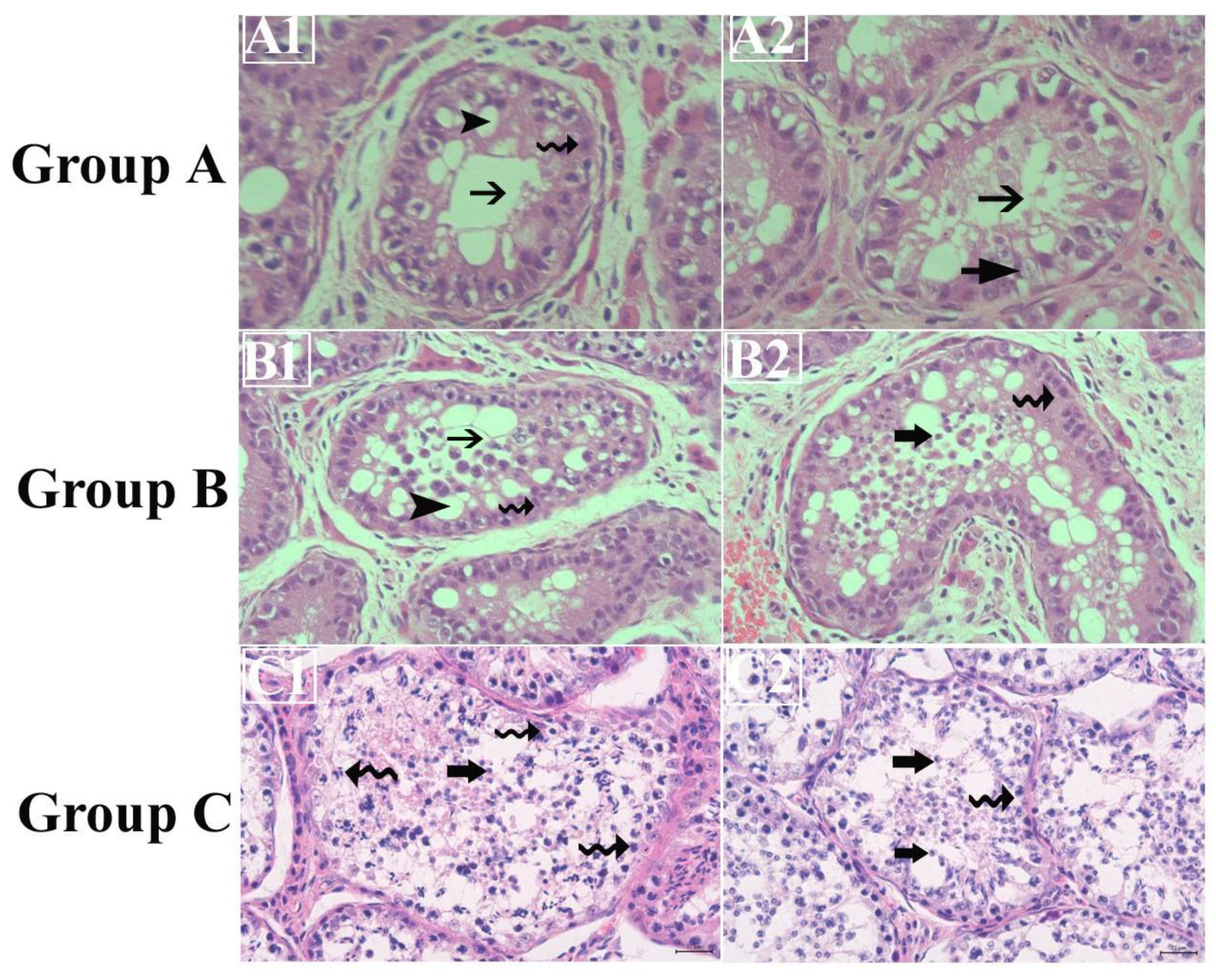
Figure 4.
Histological pictures showing apoptosis of seminiferous epithelium in testes of Dezhou donkeys after 40th day in Inhibin immunized Groups A (A1 and A2), B (B1 and B2). Group C (C1 and C2) shows normal seminiferous epithelium. Scale bar is 25µm at 40X. Black arrow with tail: Empty lumen, Spiral arrow: spermatogonia, Arrow head: Sertoli cell vacuolation, Arrow head with thick tail: round spermatids. Each group n = 4.
Figure 4.
Histological pictures showing apoptosis of seminiferous epithelium in testes of Dezhou donkeys after 40th day in Inhibin immunized Groups A (A1 and A2), B (B1 and B2). Group C (C1 and C2) shows normal seminiferous epithelium. Scale bar is 25µm at 40X. Black arrow with tail: Empty lumen, Spiral arrow: spermatogonia, Arrow head: Sertoli cell vacuolation, Arrow head with thick tail: round spermatids. Each group n = 4.
5. Conclusion
Taken together, inhibin immunization slightly upregulates plasma hormone concentrations of FSH, LH, Testosterone, and AMH in non breeding seasonality in donkeys. Testicular histoarchitecture shows signs of apoptosis and Sertoli cell vacuolation. We speculate that inhibin immunization can lower efficiency of spermatogenesis in donkeys also. The concomitant effect of non breeding season and inhibin immunization imbalanced plasma hormone concentrations that resulted in disrupted testicular histoarchitecture. In contrast, the slight upregulation of FSH, LH, AMH, and Testosterone after inhibin immunization gives the notion that if we slightly enhance the INH antigen dose, it may improve the reproductive efficiency of donkeys in terms of plasma hormone concentrations and semen quality. Semen-quality biomarkers and molecular pathways are needed to be explored for further studies.
Author Contributions
Writing original draft, M.F.A; visualization, M.U, formal analysis, E.A; investigation, L.L; project administration, W.C.
Funding
This research was funded by the National Key R&D Program of China (grant numbers 2022YFD1600103; 2023YFD1302004), the Shandong Province Modern Agricultural Technology System Donkey Industrial Innovation Team (grant no. SDAIT-27), Livestock and Poultry Breeding Industry Project of the Ministry of Agriculture and Rural Affairs (grant number 19211162), the National Nat ural Science Foundation of China (grant no. 31671287), The Open Project of Liaocheng University Animal Husbandry Discipline (grant no. 319312101-14), the Open Project of Shandong Collaborative Innovation Center for Donkey Industry Technology (grant no. 3193308), Doctoral research start-up fee of the Science and Technology Department (grant no. 318/318052367), Excellent foreign young scientists NSFC (grant no. 307/307272201).
Institutional Review Board Statement
This study was approved by the Research Committee of the Animal Policy and Welfare Committee of Liaocheng University (No. LC2019-1). The care and use of laboratory animals fully comply with local animal welfare laws, guidelines, and policies.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shi, T.; Hu, W.; Hou, H.; Zhao, Z.; Shang, M.; Zhang, L. Identification and comparative analysis of long non-coding RNA in the skeletal muscle of two dezhou donkey strains. Genes 2020, 11, 508. [Google Scholar] [CrossRef]
- Tian, F.; Wang, J.; Li, Y.; Yang, C.; Zhang, R.; Wang, X.; Ju, Z.; Jiang, Q.; Huang, J.; Wang, C. Integrated analysis of mRNA and miRNA in testis and cauda epididymidis reveals candidate molecular markers associated with reproduction in Dezhou donkey. Livestock Science 2020, 234, 103885. [Google Scholar] [CrossRef]
- Zeng, L.; Dang, R.; Dong, H.; Li, F.; Chen, H.; Lei, C. Genetic diversity and relationships of Chinese donkeys using microsatellite markers. Archives Animal Breeding 2019, 62, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Wu, F.; Zhou, Z.; Li, M.; Gao, Y.; Yin, G.; Yu, J.; Lei, C.; Dang, R. Expression profiles and polymorphic identification of the ACSL1 gene and their association with body size traits in Dezhou donkeys. Archives Animal Breeding 2020, 63, 377–386. [Google Scholar] [CrossRef]
- Clauss, M.; Zerbe, P.; Bingaman Lackey, L.; Codron, D.; Müller, D.W. Basic considerations on seasonal breeding in mammals including their testing by comparing natural habitats and zoos. Mammalian Biology 2021, 101, 373–386. [Google Scholar] [CrossRef]
- Tibary, A.; Sghiri, A.; Bakkoury, M.; Fite, C. Reproductive patterns in donkeys. In Proceedings of the Proceedings of the 9th International Congress of the World Equine Veterinary Association, 2006; pp. 311-319.
- Aissanou, S.; Besseboua, O.; Abdelhanine, A. Some reproductive characteristics in common donkey male (Equus asinus)-A mini review. Turkish Journal of Veterinary Research 2022, 6, 77–84. [Google Scholar] [CrossRef]
- Akhtar, M.F.; Wei, Q.; Zhu, H.; Chen, Z.; Ahmad, E.; Zhendan, S.; Shi, F. The role of active immunization against inhibin α-subunit on testicular development, testosterone concentration and relevant genes expressions in testis, hypothalamus and pituitary glands in Yangzhou goose ganders. Theriogenology 2019, 128, 122–132. [Google Scholar] [CrossRef]
- Woodruff, T.K.; Mather, J.P. Inhibin, activin and the female reproductive axis. Annual review of physiology 1995, 57, 219–244. [Google Scholar] [CrossRef]
- Liu, Y.; Mao, X.; Wei, Y.; Yu, J.; Li, H.; Chen, R.; Shi, Z. Studies on enhancing embryo quantity and quality by immunization against inhibin in repeatedly superovulated Holstein heifers and the associated endocrine mechanisms. Animal reproduction science 2013, 142, 10–18. [Google Scholar] [CrossRef]
- Chen, F.; Lu, J.; Guo, R.; Mei, C.; Guo, B.; Li, W.; Tsigkou, A.; Shi, Z. Rectifying cow infertility under heat stress by immunization against inhibin and supplementation of progesterone. Domestic Animal Endocrinology 2022, 80, 106726. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Ahmad, E.; Sattar, A.; Riaz, A.; Khan, J.A.; Naseer, Z.; Akhtar, M.F.; Abbas, M.; Shi, Z. Long term effects of immunization against inhibin on fresh and post-thawed semen quality and sperm kinematics during low and peak breeding seasons in Beetal bucks. Small Ruminant Research 2021, 201, 106442. [Google Scholar] [CrossRef]
- Rehman, A.; Ahmad, E.; Arshad, U.; Riaz, A.; Akhtar, M.S.; Ahmad, T.; Khan, J.A.; Mohsin, I.; Shi, Z.; Sattar, A. Effects of immunization against inhibin α-subunit on ovarian structures, pregnancy rate, embryonic and fetal losses, and prolificacy rate in goats where estrus was induced during the non-breeding season. Animal Reproduction Science 2021, 224, 106654. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, Z.; Ma, Z.; Ma, J.; Zhao, F. Immunization against inhibin promotes fertility in cattle: A meta-analysis and quality assessment. Frontiers in Veterinary Science 2021, 8, 687923. [Google Scholar] [CrossRef]
- Meng, J.; Feng, J.H.; Xiao, L.; Zhou, W.; Zhang, H.; Lan, X.; Wang, S. Active immunization with inhibin DNA vaccine promotes spermatogenesis and testicular development in rats. Journal of Applied Animal Research 2024, 52, 2360408. [Google Scholar] [CrossRef]
- Lovell, T.M.; Knight, P.G.; Groome, N.P.; Gladwell, R.T. Measurement of dimeric inhibins and effects of active immunization against inhibin α-subunit on plasma hormones and testis morphology in the developing cockerel. Biology of reproduction 2000, 63, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Baqerkhani, M.; Soleimanzadeh, A.; Mohammadi, R. Effects of intratesticular injection of hypertonic mannitol and saline on the quality of donkey sperm, indicators of oxidative stress and testicular tissue pathology. BMC Veterinary Research 2024, 20, 99. [Google Scholar] [CrossRef]
- Akhtar, M.F.; Ahmad, E.; Ali, I.; Shafiq, M.; Chen, Z. The effect of inhibin immunization in seminiferous epithelium of Yangzhou goose ganders: a histological study. Animals 2021, 11, 2801. [Google Scholar] [CrossRef]
- Han, Y.; Jiang, T.; Liu, A.; Liu, L. Role and regulatory mechanism of inhibin in animal reproductive system. Theriogenology 2023, 202, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Bernard, D.J.; Li, Y.; Toufaily, C.; Schang, G. Regulation of gonadotropins. In Oxford Research Encyclopedia of Neuroscience; 2019.
- Guo, R.; Chen, F.; Mei, C.; Dai, Z.; Yan, L.; Shi, Z. Conception rate and reproductive hormone secretion in Holstein cows immunized against inhibin and subjected to the ovsynch protocol. Animals 2020, 10, 313. [Google Scholar] [CrossRef]
- Li, D.; Qin, G.; Wei, Y.; Lu, F.; Huang, Q.; Jiang, H.; Shi, D.; Shi, Z. Immunisation against inhibin enhances follicular development, oocyte maturation and superovulatory response in water buffaloes. Reproduction, Fertility and Development 2011, 23, 788–797. [Google Scholar] [CrossRef]
- Anderson, R.; Groome, N.; Baird, D. Inhibin A and inhibin B in women with polycystic ovarian syndrome during treatment with FSH to induce mono-ovulation. Clinical endocrinology 1998, 48, 577–584. [Google Scholar] [CrossRef]
- Medan, M.; Akagi, S.; Kaneko, H.; Watanabe, G.; Tsonis, C.; Taya, K. Effects of re-immunization of heifers against inhibin on hormonal profiles and ovulation rate. Reproduction 2004, 128, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Medan, M.S.; Watanabe, G.; Sharawy, S.; Taya, K. Immunization of goats against inhibin increased follicular development and ovulation rate. Journal of Reproduction and Development 2006, 52, 543–550. [Google Scholar] [CrossRef]
- Lazebny, O.; Kulikov, A.; Butovskaya, P.; Proshakov, P.; Fokin, A.; Butovskaya, M. Analysis of aggressive behavior in young Russian males using 250 SNP markers. Russian Journal of Genetics 2020, 56, 1118–1128. [Google Scholar] [CrossRef]
- O’Donnell, L.; Whiley, P.A.; Loveland, K.L. Activin A and sertoli cells: key to fetal testis steroidogenesis. Frontiers in Endocrinology 2022, 13, 898876. [Google Scholar] [CrossRef]
- Rodriguez, K.F.; Brown, P.R.; Amato, C.M.; Nicol, B.; Liu, C.-F.; Xu, X.; Yao, H.H.-C. Somatic cell fate maintenance in mouse fetal testes via autocrine/paracrine action of AMH and activin B. Nature Communications 2022, 13, 4130. [Google Scholar] [CrossRef]
- Shah, W.; Khan, R.; Shah, B.; Khan, A.; Dil, S.; Liu, W.; Wen, J.; Jiang, X. The molecular mechanism of sex hormones on Sertoli cell development and proliferation. Frontiers in endocrinology 2021, 12, 648141. [Google Scholar] [CrossRef]
- Arato, I.; Grande, G.; Barrachina, F.; Bellucci, C.; Lilli, C.; Jodar, M.; Aglietti, M.C.; Mancini, F.; Vincenzoni, F.; Pontecorvi, A. “In vitro” Effect of Different Follicle—Stimulating Hormone Preparations on Sertoli Cells: Toward a Personalized Treatment for Male Infertility. Frontiers in Endocrinology 2020, 11, 401. [Google Scholar] [CrossRef]
- Santi, D.; Crépieux, P.; Reiter, E.; Spaggiari, G.; Brigante, G.; Casarini, L.; Rochira, V.; Simoni, M. Follicle-stimulating hormone (FSH) action on spermatogenesis: a focus on physiological and therapeutic roles. Journal of Clinical Medicine 2020, 9, 1014. [Google Scholar] [CrossRef]
- Wang, J.-M.; Li, Z.-F.; Yang, W.-X.; Tan, F.-Q. Follicle-stimulating hormone signaling in Sertoli cells: a licence to the early stages of spermatogenesis. Reproductive Biology and Endocrinology 2022, 20, 97. [Google Scholar] [CrossRef]
- Ni, F.-D.; Hao, S.-L.; Yang, W.-X. Molecular insights into hormone regulation via signaling pathways in Sertoli cells: With discussion on infertility and testicular tumor. Gene 2020, 753, 144812. [Google Scholar] [CrossRef]
- Olsen, O.E.; Hella, H.; Elsaadi, S.; Jacobi, C.; Martinez-Hackert, E.; Holien, T. Activins as dual specificity TGF-β family molecules: SMAD-activation via activin-and BMP-type 1 receptors. Biomolecules 2020, 10, 519. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, B.; Qi, Y.; Zhu, L.; Cui, X.; Liu, Z. Antagonistic effects of activin A and TNF-α on the activation of L929 fibroblast cells via Smad3-independent signaling. Scientific Reports 2020, 10, 20623. [Google Scholar] [CrossRef]
- Zeng, X.; Turkstra, J.; Tsigos, A.; Meloen, R.; Liu, X.; Chen, F.; Schaaper, W.; Guo, D.; van de Wiel, D. Effects of active immunization against GnRH on serum LH, inhibin A, sexual development and growth rate in Chinese female pigs. Theriogenology 2002, 58, 1315–1326. [Google Scholar] [CrossRef]
- Avital-Cohen, N.; Heiblum, R.; Argov, N.; Rosenstrauch, A.; Chaiseha, Y.; Mobarkey, N.; Rozenboim, I. The effect of active immunization against vasoactive intestinal peptide and inhibin on reproductive performance of young White Leghorn roosters. Poultry science 2011, 90, 2321–2331. [Google Scholar] [CrossRef] [PubMed]
- Samir, H.; El Sayed, M.A.; Nagaoka, K.; Sasaki, K.; El-Maaty, A.M.A.; Karen, A.; Abou-Ahmed, M.M.; Watanabe, G. Passive immunization against inhibin increases testicular blood flow in male goats. Theriogenology 2020, 147, 85–91. [Google Scholar] [CrossRef]
- Oduwole, O.O.; Huhtaniemi, I.T.; Misrahi, M. The roles of luteinizing hormone, follicle-stimulating hormone and testosterone in spermatogenesis and folliculogenesis revisited. International journal of molecular sciences 2021, 22, 12735. [Google Scholar] [CrossRef]
- Ali, A.; Derar, D.R.; Zeitoun, M.M.; Al-Sobayil, F. Impotentia generandi in male dromedary camels: FSH, LH and testosterone profiles and their association with clinical findings and semen analysis data. Theriogenology 2018, 120, 98–104. [Google Scholar] [CrossRef]
- Swelum, A.A.-A.; Saadeldin, I.M.; Zaher, H.A.; Alsharifi, S.A.; Alowaimer, A.N. Effect of sexual excitation on testosterone and nitric oxide levels of water buffalo bulls (Bubalus bubalis) with different categories of sexual behavior and their correlation with each other. Animal reproduction science 2017, 181, 151–158. [Google Scholar] [CrossRef] [PubMed]
- ur Rehman, Z.; Worku, T.; Davis, J.S.; Talpur, H.S.; Bhattarai, D.; Kadariya, I.; Hua, G.; Cao, J.; Dad, R.; Hussain, T. Role and mechanism of AMH in the regulation of Sertoli cells in mice. The Journal of Steroid Biochemistry and Molecular Biology 2017, 174, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Matuszczak, E.; Hermanowicz, A.; Komarowska, M.; Debek, W. Serum AMH in physiology and pathology of male gonads. International journal of endocrinology 2013, 2013, 128907. [Google Scholar] [CrossRef] [PubMed]
- Rey, R.A.; Grinspon, R.P. Normal male sexual differentiation and aetiology of disorders of sex development. Best Practice & Research Clinical Endocrinology & Metabolism 2011, 25, 221–238. [Google Scholar]
- Hui, H.-B.; Xiao, L.; Sun, W.; Zhou, Y.-J.; Zhang, H.-Y.; Ge, C.-T. Sox9 is indispensable for testis differentiation in the red-eared slider turtle, a reptile with temperature-dependent sex determination. Zoological Research 2021, 42, 721. [Google Scholar] [CrossRef]
- Shima, Y.; Miyabayashi, K.; Haraguchi, S.; Arakawa, T.; Otake, H.; Baba, T.; Matsuzaki, S.; Shishido, Y.; Akiyama, H.; Tachibana, T. Contribution of Leydig and Sertoli cells to testosterone production in mouse fetal testes. Molecular endocrinology 2013, 27, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Wu, J.; Liu, B.; Jiang, Y.; Chen, W.; Li, J.; He, Q.; He, Z. The roles and mechanisms of Leydig cells and myoid cells in regulating spermatogenesis. Cellular and molecular life sciences 2019, 76, 2681–2695. [Google Scholar] [CrossRef]
- Banerjee, S.; Chaturvedi, C.M. Apoptotic mechanism behind the testicular atrophy in photorefractory and scotosensitive quail: Involvement of GnIH induced p-53 dependent Bax-Caspase-3 mediated pathway. Journal of Photochemistry and Photobiology B: Biology 2017, 176, 124–135. [Google Scholar] [CrossRef]
- Jiménez, R.; Burgos, M.; Barrionuevo, F.J. Circannual testis changes in seasonally breeding mammals. Sexual Development 2015, 9, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Beltrán-Frutos, E.; Seco-Rovira, V.; Martínez-Hernández, J.; Ferrer, C.; Serrano-Sánchez, M.I.; Pastor, L.M. Cellular modifications in spermatogenesis during seasonal testicular regression: an update review in mammals. Animals 2022, 12, 1605. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



