
Submitted:
29 January 2025
Posted:
30 January 2025
You are already at the latest version
Abstract
Background: Microsporidia MB (MB), a promising biological control agent suppresses Plasmodium falciparum transmission in Anopheles mosquitoes. Methods: This study examined the spatial distribution of MB infection in natural populations of An. gambiae s.l. mosquitoes collected in Nigeria and Niger Republic, and its potential modulation of insecticide susceptibility in the Anopheles mosquitoes. Results: The overall prevalence of MB in F₀ mosquitoes was 12.25% (95% CI: 7.76%-16.75%); 25 mosquitoes out of 204 were positive. Geographic variation was observed, with a higher prevalence (5/15 mosquitoes) in Ebonyi State (33.33%, 95% CI: 9.48%-57.19%, Fisher's exact test, p = 0.008). Infection rates were higher in An. coluzzii mosquitoes (21/133 mosquitoes), estimated at 15.79% (95% CI: 9.59%-21.99%) compared to An. gambiae s.s. mosquitoes (4/67), approximately 5.63% (95% CI: 0.27%-11.00%), χ² = 4.44; df = 1, p = 0.035)]. Resistant mosquitoes had a significantly higher prevalence of MB infection at 28.57% (95% CI: 16.74%-40.40%) and OR of 3.33 (95% CI: 1.23-9.03, p = 0.017) compared with susceptible mosquitoes. Conclusion: MB has wide geographic distribution across Nigeria and Niger Republic. An. coluzzii and resistant mosquitoes are more infected with this fungi. Understanding the geographic and species-specific infection patterns and implication in insecticide resistance could guide targeted vector control strategies using this biological agent.
Keywords:
Anopheles
; malaria
; Microsporidia MB
; control
; insecticide resistance
; spatial distribution
1. Introduction
Malaria remains a major global public health issue, particularly in the African continent, where more than 94% of the infection and 95% of malaria-related mortalities occur, disproportionately in children under the age of 5 and pregnant women [1]. Nigeria has the highest burden of malaria in the world, accounting for ~26% of malaria cases and ~30% of malaria-related deaths globally. However, the neighboring Niger also faces significant malaria challenges, with a prevalence of approximately 237 cases per 1,000 inhabitants in 2022 [2], and contributing 5.9% of malaria-related deaths globally, ranking third, only below Nigeria and Democratic Republic of Congo [1].
The major Anopheles species found in Niger and Nigeria belong to the Anopheles gambiae complex and secondarily the Anopheles funestus s.s. [3,4,5,6,7,8]. These species exhibit considerable ecological diversity and wide distribution patterns across different habitats [9,10,11]. Their distribution is often influenced by environmental factors, such as changes in temperature, humidity, and precipitation [11,12]. Studies conducted in Niger and Nigeria have highlighted the abundance and broad distribution of species within the Anopheles gambiae complex and Anopheles funestus group along various ecological zones [4,7,13]. Plasmodium infection status varies across the two countries, with P. falciparum being the major parasite, with parasite rate of between 0.77% and 22.3% in An. coluzzii from several studies conducted in Nigeria [4,14]. From Niger, according to a study published in 2019 the parasite rate was ~ 13% in An. coluzzii [13]. Recent discoveries have also identified Plasmodium vivax from Niger [15].
Excessive use of insecticides for vector control threatens to derail progress in malaria control efforts. Resistance has now been reported to nearly all classes of insecticides used for public health, with pyrethroids, the key ingredients in insecticide-treated bed nets (ITNs) being affected disproportionately [1]. In Niger, pyrethroid resistance towards alpha-cypermethrin, deltamethrin, and permethrin has been documented [13,16,17,18,19]. Also, in Nigeria, widespread pyrethroid resistance has been reported in Anopheles mosquitoes. Studies have shown high resistance levels in southern and northern Nigeria, with significant operational implications for ITN effectiveness [4,20,21,22,23,24]. A comprehensive RNA-seq analysis, including mosquitoes from Niger and Nigeria, has identified molecular drivers of pyrethroid resistance, such as overexpression of detoxification enzymes and target-site insensitivity mutations [25].
In Niger and Nigeria high pyrethroid resistance in Anopheles gambiae is associated with increased activities of metabolic enzymes (P450 + GST) and the West Africa knock down resistance (kdr-w) mechanism [18,19,22,25,26]. While much of research focus has been on these major resistance mechanisms, mechanisms such as the role of gut microbiome in mediating pyrethroid resistance are receiving attention. For example, Dada and colleagues have demonstrated that variations in the Anopheles gut microbiome can influence pyrethroid resistance, suggesting an additional layer of complexity to resistance management [27].
In the face of the resistance challenges associated with synthetic insecticides, efforts are increasingly invested towards alternative, more eco-friendly control measures. For example, Wolbachia, and Bacillus thuringiensis (Bti). However, the recent discovery of Microsporidia MB, an intracellular symbiont in Anopheles mosquitoes, has sparked interest due to its potential for malaria control. The presence of MB in Anopheles mosquitoes negatively impacts their ability to transmit Plasmodium [28]. MB accumulates in mosquito tissues, with sexual and vertical transmission ensuring its persistence. Its prevalence and intensity in tissues during the mosquito lifecycle suggest an adaptation for maximizing transmission while minimizing virulence [29,30]. These traits facilitate its spread within Anopheles populations. The presence of MB has been confirmed in several Anopheles species across Africa, including in Kenya, Ghana, Niger, and Benin [7,28,31,32]. Studies have shown that its prevalence varies by geographic region, habitat type, and climatic conditions. Unfortunately, these studies have been geographically restricted to single countries. For instance, in Kenya, MB prevalence in An. gambiae complex mosquitoes was higher in low-altitude areas with warm temperatures and high rainfall compared to high-altitude areas with cooler temperatures and lower rainfall [28]. Similarly, in Ghana, MB prevalence in An. funestus was higher in forest zones compared to savanna zones [31]. These findings underscore the significant role of environmental conditions in shaping the geographic distribution of MB.
While MB shows promise as a biological control agent [33], its effectiveness may be influenced by fluctuating environmental conditions and inherent variability in transmission rates, complicating its use. Also, its potential to influence mosquito phenotypes, particularly insecticide resistance, remains unexplored. Understanding interspecific and intraspecific differences of MB within Anopheles populations and their potential role in resistance modulation is essential for informing malaria control strategies. This study investigated the spatial distribution of MB along clinal gradient of increasing aridity and assess its ability to influence insecticide resistance phenotype in Anopheles mosquitoes from Nigeria and Niger Republic.
2. Materials and Methods
2.1. Collection of Anopheles mosquitoes Along Gradient of Increasing Aridity
Mosquitoes were collected across twelve sites, spanning contrasting ecological settings: one site in Niger Republic and eleven in Nigeria (Figure 1). These sites cover mangrove swamp, tropical rainforest, Guinea savanna, Sudan savannah, and the Sahel (Table 1). Blood fed female Anopheles mosquitoes (F0) resting indoor were collected using Prokopack electric aspirators (John W. Hook, Gainesville, FL, USA) early in the morning, between 05:00-06:00 am. The mosquitoes were transferred to Bayero University Kano, in paper cups (using cooling boxes) and maintained under standard insectarium condition (25oC and 75% relative humidity, with 12h:12h light:dark cycles), fed with 10 % sugar. Collections were conducted in Nigeria between June-September and in Niger in September 2024.
2.2. Morphological and Molecular Identification of Anopheles Mosquitoes to Species Level
Gravid F0 female mosquitoes (4-5 d old after blood meal) were forced to lay eggs inside 1.5 mL Eppendorf tubes. They were morphologically identified [34]. Molecular identification to species level were done using SINE200 PCR [35]. Genomic DNA was extracted using the LIVAK protocol [36]. For each gDNA premix of 14 µl comprised of 1.5 µl of 10x Taq A buffer, 0.75 µl of MgCl2, 0.12 µl of dNTP mixes, 0.51 µl each of the SINE200 F and R primers, 0.12 µl of KapaTaq Polymerase (Kapa Biosciences, Wilmington, MA, USA), and 10.49 µl of ddH20 were constituted. 1 µl each of gDNA were added to the premix. Thermal cycling conditions were an initial denaturation at 95°C for 5 min, followed by 35 cycles each of denaturation at 95°C for 30 s, annealing at 54°C for 30 s, and extension at 72°C for 1 min. A final extension step was performed at 72°C for 5 minutes. PCR products were separated on 1.5% agarose gel stained with pEqGREEN, and visualized using GelDoc, with expected band sizes of 479 bp, 245 bp and 220 bp respectively for An. coluzzii, An. gambiae s.s. and An. arabiensis.
2.3. Investigation of Spatial Distribution of MB Infection in Anopheles Mosquitoes
Subsets of gDNA extracted from the F0 females above (Table 1) were used for PCR identification of MB infection. PCR amplification of the 18S rRNA gene was performed using MB18SF and MB18SR primers according to previously published protocol [28]. Briefly, each 12 μl reaction mix contained 3 μl of 5x master mix (Solis Biodyne, Estonia), comprise of 7.5 mM of MgCl2, 2 mM of each dNTP mixes and HOT FIREPol® DNA polymerase). This was mixed with 0.75 μl each of F and R primer, 3 μl of DNA template, and 7.5 μl of nuclease-free water. Thermal cycling conditions included an initial denaturation at 95°C for 15 mins, followed by 35 cycles each of denaturation at 95°C for 30 s, annealing at 59°C for 45 s, and extension at 72°C for 45 s, PCR products were separated on 1.5% agarose gel stained with pEqGREEN, and visualized using GelDoc for the expected band size of approximately 500 bp characteristic of MB.
2.4. Investigation of the Modulation of Insecticide Resistance by MB
The F1 mosquitoes from six sites in Nigeria, which were phenotyped with pyrethroid insecticides were selected to screen MB infection (Table 1). Subsets each of alive and dead mosquitoes were screened for infection, attempting to even the number of resistant (R) and susceptible (S) mosquitoes. Bioassays were conducted using impregnated papers in WHO tubes according to [37]. Mosquitoes were exposed to 0.05% deltamethrin or 0.75% permethrin for 1 h, transferred to holding tubes and supplied with 10 % sugar. Mortalities were recorded 24 h post-exposure. Details of the insecticide test profiles will be provided in a separate publication (in preparation).
2.5. Data Analysis
Data were analysed using XLSTAT software [38], to investigate the relationships between MB infection status, Anopheles species, locality and insecticide resistance. To evaluate the relationships between variables, a Chi-square test or Fisher’s Exact test of independence was applied to determine whether MB infection status is associated with species, locality and insecticide susceptibility. The odds ratio (OR) was determined to measure the strength of association between variables. P values of p < 0.05 were considered statistically significant.
Multiple Correspondence Analysis (MCA) was done using XLSTAT’s “Factor Analysis” package, a multivariate technique designed for categorical data. The MCA was applied to explore key relationships in the study, including the association between MB infection status and Anopheles species, the relationship between infection status and mosquito susceptibility (resistant vs. susceptible), and the spatial distribution of MB across sampled localities.
3. Results
3.1. MB Infects Anopheles Mosquitoes from Contrasting Ecological Settings Along Clinal Gradient
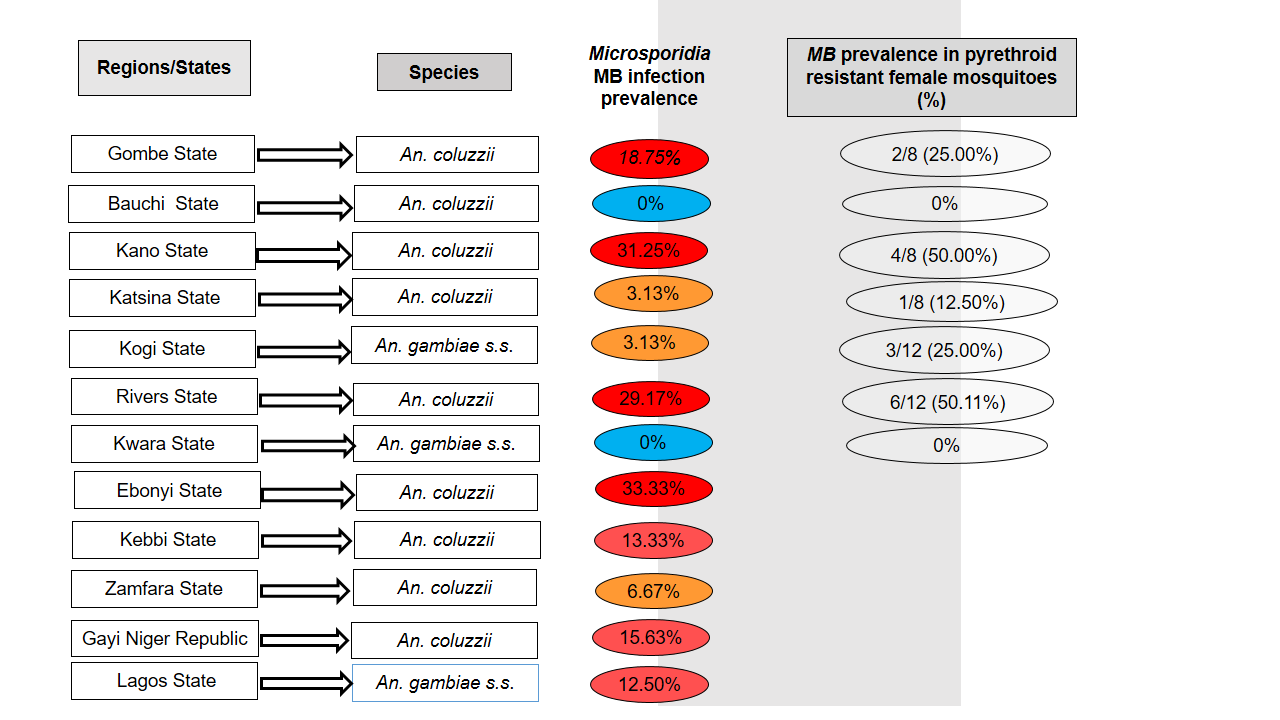
A total of 204 adult female F₀ mosquitoes were initially analysed for MB infection, out of which 25 were infected (gel pictures in File S1) 12.25% prevalence (95% CI: 7.76%-16.75%). In addition F1 samples (from sites where F0 females were not screened for MB infection), which were used for bioassays and screened for MB infection (see below) were lumped in this analysis. MB is widespread across sites with only 2 out of the 12 sites negative (Figure 2). The prevalence of infection varied, ranging from of 0% to 33% (File S2). For F0 females the infection rate was highest in Ebonyi State, where 5 out of 15 mosquitoes were infected (33.33%, 95% CI: 9.48%-57.19%); followed by Rivers State, where 7 out of 24 mosquitoes were positive (29.17%, 95% CI: 10.98%-47.35%). However, mosquitoes from Kwara and Gombe States were not infected. A Fisher’s test confirmed a statistically significant association between MB infection status and localities (p = 0.008).
Infection varied across Anopheles species with An. coluzzii exhibiting a higher infection rate. A total of 21 out of 133 An. coluzzii mosquitoes were infected, translating into prevalence of 15.79% (95% CI: 9.59%-21.99%), compared to only 4 out of 67 for An. gambiae s.s. (prevalence = 5.63%, 95% CI: 0.27%-11.00%). A statistically significant association was observed between MB infection status and mosquito species (χ² = 4.44; df = 1, p = 0.035). Furthermore, the odds ratio of infection with MB was 3.14 (95% CI: 1.03-9.54, p = 0.035) for An. coluzzii, suggesting a positive association, when compared to odds ratio of 0.32 (95% CI: 0.11-0.91) for An. gambiae s.s. Relationship between MB infection status and Anopheles species was also analysed using multiple correspondence analysis (Figure 3).
The MCA plot illustrates the association between MB infection status and Anopheles species by clustering variable categories with similar profiles on the factor map. The two dimensions (F1 and F2) together explain 100% of the variability in the dataset, with F1 accounting for 57% and F2 explaining 43% of the variance. On the map, An. coluzzii and MB+ variables clustered together in the positive quadrant of F1. This clustering indicates that these categories share similar profiles, consistent with the significantly higher prevalence of MB infection observed in An. coluzzii. The strong association along F1 suggests that the dimension captures the variability driven by the high infection rates in An. coluzzii mosquitoes. Conversely, MB- (negative infection status-No) and An. gambiae s.s. cluster in the negative and positive quadrants of F2, respectively. This separation reflects the lower prevalence of infection in An. gambiae s.s., highlighting a distinct profile compared to An. coluzzii. This clear spatial differentiation on the MCA plot corroborates the statistical findings, where An. coluzzii exhibited a significantly higher infection with MB compared to An. gambiae s.s.
3.2. MB Infection Probably Correlates with Pyrethroid Resistance
To test the hypothesis that MB modulate insecticide resistance a subset of F1 female mosquitoes previously exposed to pyrethroids, were PCR-screened to detect MB infection. Out of the 112 females tested with pyrethroid, an equal proportion of 50% susceptible (S) and 50% resistant (R) mosquitoes were utilized. Overall, 16 of 56 resistant (R) mosquitoes (28.57%, 95% CI: 16.74%-40.40%) were infected with MB, compared to 6 of 56 susceptible (S) mosquitoes (10.71%, 95% CI: 2.61%-18.82%). These differences in infection were statistically significant (χ² = 5.65, df = 1, p = 0.017). The odds ratio of likelihood of surviving pyrethroid exposure (R phenotype) when MB infected was significant (OR = 3.33, 95% CI: 1.23-9.03, p = 0.017), compared with the non-significant odds ratio of 0.30 (95% CI: 0.11-0.81) observed in susceptible mosquitoes. The MCA plot in Figure 4 depicts the association between MB infection status and phenotypes (resistant vs. susceptible) by clustering variable categories with similar profiles. The two dimensions (F1 and F2) explain 100% of the variability, with F1 accounting for 61% and F2 explaining 39% of the variance.
Variables Resistant (R) and MB+ (positive infection status) cluster closely in the positive quadrant of F1. This indicates association between the resistance phenotype and MB infection status. In contrast, susceptible (S) phenotype and MB- (negative infection status) cluster in the negative and positive quadrants of F2, respectively. This suggests an association between low MB infection and pyrethroid susceptibility.
4. Discussion
The development and spread of insecticide resistance which is threatening the progress so far made in malaria control targeting mosquito vectors spurs efforts for alternative control measures, including using biological agents such as Wolbachia and B. thuringiensis. The discovery of MB and the recent description of its ability to reduce Plasmodium infection in malaria mosquitoes [28] has catalyzed new efforts to understand the bionomics of this promising fungus.
This primary study generates information about the distribution of MB in Anopheles mosquitoes from contrasting ecological settings and its influence on insecticide resistance. MB is relatively well established in mosquito populations from Nigeria and Niger Republic, with only two mosquito populations out of 12 screened free of its infection. Indeed, previous studies have suggested that local and ecological variations influence presence of MB [30,31,39]. To the best of our knowledge this is the first study to report the presence of MB in Anopheles mosquitoes from Nigeria, and the second study to describe it in mosquitoes from Niger.
Across Africa MB appears to be well established in the main malaria vectors. The 12.25% MB infection prevalence observed in this study was double the 6.8% (17/251) prevalence reported in An. coluzzii, in a study in which no infection was observed in An. gambiae from Zinder city in Niger Republic [7]. However, it is lower than 53.4% prevalence previously reported in Benin Republic [32]. The MB prevalence observed in Ghana (1.8%) [31] and Kenya (1.7%) [39] were much lower than our findings (though fewer mosquito samples were screened in our study).
This study demonstrated a significantly higher prevalence of MB in An. coluzzii, compared to An. gambiae s.s. A similar pattern has been described in a previous study from Benin Republic, with a prevalence of 41.0% in An. gambiae s.s. versus 57.0% in An. coluzzii [32].
Most studies in East Africa indicate that MB infects An. arabiensis [28,29,30,39], while in West Africa it primarily infects An. coluzzii [7,31,32].
The MB infection rates varied significantly across the study sites, reflecting a likely influence of geographic and ecological factors. The highest infection rates were observed in Ebonyi State (a Tropical Rainforest site) and Rivers State (Mangrove Swamp) in the wild-caught mosquitoes, possibly because these localities offer particularly favourable conditions for optimal MB growth, including high humidity, moderate temperatures, and habitats for mosquito vectors. These findings are consistent with a previous study that reported that MB prevalence is influenced by species-specific and ecological factors [28]. Similarly, other studies [39,40] have documented geographic variation in MB prevalence, with higher rates in humid environments.
To the best of our knowledge, this is the first study to explore the hypothesis of possible association between insecticide resistance and MB infection. The higher prevalence of MB in pyrethroid resistant mosquitoes raises important questions about its potential role in modulating insecticide resistance. Several studies have linked microbiome with insecticide resistance showing that pyrethroid resistant mosquitoes harbour distinct microbiota compared to the pyrethroid-susceptible strains [27,41]. For example, in the major malaria vector An. coluzzii, an abundance of Asaia and Serratia bacteria has been associated with deltamethrin resistance [42]. Resistant mosquitoes have been found to be enriched with bacteria and enzymes that can degrade insecticides, suggesting that the microbiota may play a role in resistance [27]. Also, it has been shown that MB infection can influence the phenotype of Anopheles such as adaptation to varying dietary conditions [43], faster larval development and higher adult emergence rates [44], all which suggests modulation of various phenotypes. Our finding of possible association between pyrethroid resistance and MB infection highlights the need for further research into its roles in insecticide resistance, particularly towards the key malaria control tools, such as the long-lasting insecticidal bed nets, and the ingredients in use for indoor residual spraying. Understanding these may strengthen managing resistance and improving malaria control efforts and could lead to novel strategies to use the link with insecticide resistance to facilitate the spread of MB through mosquito populations for sustainable malaria control.
5. Conclusions
MB is ubiquitous in Anopheles mosquitoes across Nigeria, and predominantly in An. coluzzii in both Nigeria and Niger Republic. Its ability to infect and establish in Anopheles mosquitoes is probably influenced by ecological and biological factors. Understanding the geographic and species-specific infection patterns could guide targeted vector control strategies, particularly in regions with high infection prevalence.
These findings highlight the need to better understand the complexities of MB – mosquito interactions. A more comprehensive understanding of the impact of MB infection on resistance towards the major public health insecticides and malaria control tools will be key to the development of appropriate MB based malaria control strategies. These insights can potentially inform innovative strategies for non-synthetic insecticide approaches for malaria vector control in Africa.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. File S1: Agarose gel of MB and File S2: File S2. Prevalence of MB infection across geographical site.
Author Contributions
Conceptualization, S.S.I., J.K.H., and M.L.M.; methodology, M.L.M., A.H.S.,M.M.M., Y.Y.A., S.A., U.C.N., P.C.O., D.E.N., A.O., M.U.M. and H.K.E.; software, S.S.I. and M.L.M,; validation, S.S.I. and J.K.H.; formal analysis, M.L.M., S.S.I. and M.M.M.; resources, S.S.I.; data curation, S.S.I. and M.L.M; writing—original draft preparation, M.L.M. and S.S.I.; writing—review and editing, J.K.H., P.C.O., D.E.N. and U.C.N.; visualization, M.L.M. and S.S.I.; supervision, S.S.I.; project administration, S.S.I. and M.M.M.; funding acquisition, S.S.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data generated from this study are available in the supplementary files.
Acknowledgments
We thank the technicians of the Biochemistry laboratory of Bayero University of Kano and Sahelo-Saharan Ecology and Biodiversity Laboratory of Université André Salifou who provided technical support to the study both in the field and laboratory.
Conflicts of Interest
The authors declare no conflict of interest. This study is privately funded by SSI and MLM.
References
- WHO World Malaria Report 2024: Addressing Inequity in the Global Malaria Response. 2024.
- National Institute of Statistics Statistical Directories. National health information system, 2023.
- Ibrahim, S.S.; Mukhtar, M.M.; Irving, H.; Riveron, J.M.; Fadel, A.N.; Tchapga, W.; Hearn, J.; Muhammad, A.; Sarkinfada, F.; Wondji, C.S. Exploring the Mechanisms of Multiple Insecticide Resistance in a Highly Plasmodium-Infected Malaria Vector Anopheles Funestus Sensu Stricto from Sahel of Northern Nigeria. Genes (Basel) 2020, 11, 454. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.S.; Mukhtar, M.M.; Datti, J.A.; Irving, H.; Kusimo, M.O.; Tchapga, W.; Lawal, N.; Sambo, F.I.; Wondji, C.S. Temporal Escalation of Pyrethroid Resistance in the Major Malaria Vector Anopheles Coluzzii from Sahelo-Sudanian Region of Northern Nigeria. Sci Rep 2019, 9, 7395. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.S.; Manu, Y.A.; Tukur, Z.; Irving, H.; Wondji, C.S. High Frequency of Kdr L1014F Is Associated with Pyrethroid Resistance in Anopheles Coluzzii in Sudan Savannah of Northern Nigeria. BMC Infect Dis 2014, 14, 441. [Google Scholar] [CrossRef] [PubMed]
- Lamidi, T.B.; Alo, E.B.; Naphtali, R. Distribution and Abundance of Anopheles Mosquito Species in Three Selected Areas of Taraba State, North-Eastern Nigeria. Animal Research International 2017, 14, 2730–2740. [Google Scholar]
- Moustapha, L.M.; Sadou, I.M.; Arzika, I.I.; Maman, L.I.; Gomgnimbou, M.K.; Konkobo, M.; Diabate, A.; Bilgo, E. First Identification of Microsporidia MB in Anopheles Coluzzii from Zinder City, Niger. Parasites Vectors 2024, 17, 39. [Google Scholar] [CrossRef]
- Oduola, A.O.; Adelaja, O.J.; Aiyegbusi, Z.O.; Tola, M.; Obembe, A.; Ande, A.T.; Awolola, S. Dynamics of Anopheline Vector Species Composition and Reported Malaria Cases during Rain and Dry Seasons in Two Selected Communities of Kwara State. Nig. J. Para. 2016, 37, 157. [Google Scholar] [CrossRef]
- Ebenezer, A.; Okiwelu, S.; Agi, P.; Noutcha, Ma.E.; Awolola, T.; Oduola, A. Species Composition of the Anopheles Gambiae Complex across Eco-Vegetational Zones in Bayelsa State, Niger Delta Region, Nigeria. J Vector Borne Dis 2012, 49, 164. [Google Scholar] [CrossRef]
- Thabet, H.S.; TagEldin, R.A.; Fahmy, N.T.; Diclaro, J.W.; Alaribe, A.A.; Ezedinachi, E.; Nwachuku, N.S.; Odey, F.O.; Arimoto, H. Spatial Distribution of PCR-Identified Species of Anopheles Gambiae Senu Lato (Diptera: Culicidae) Across Three Eco-Vegetational Zones in Cross River State, Nigeria. Journal of Medical Entomology 2022, 59, 576–584. [Google Scholar] [CrossRef]
- Adeogun, A.; Babalola, A.S.; Okoko, O.O.; Oyeniyi, T.; Omotayo, A.; Izekor, R.T.; Adetunji, O.; Olakiigbe, A.; Olagundoye, O.; Adeleke, M.; et al. Spatial Distribution and Ecological Niche Modeling of Geographical Spread of Anopheles Gambiae Complex in Nigeria Using Real Time Data. Sci Rep 2023, 13, 13679. [Google Scholar] [CrossRef]
- Mamane Sale, N.; Zamanka Naroua, H.; Zoulkifouly Hounkarin, W.; Labbo, R.; Maman Laminou, I.; Djibo Souley, A.; Soumana, A.; Issa Arzika, I.; Jambou, R.; Doumma, A. Influence of environmental factors on the abundance of anopheles in the different agroecosystems of the city of Niamey. IJAR 2023, 11, 01–15. [Google Scholar] [CrossRef]
- Ibrahim, S.S.; Mukhtar, M.M.; Irving, H.; Labbo, R.; Kusimo, M.O.; Mahamadou, I.; Wondji, C.S. High Plasmodium Infection and Multiple Insecticide Resistance in a Major Malaria Vector Anopheles Coluzzii from Sahel of Niger Republic. Malar J 2019, 18, 181. [Google Scholar] [CrossRef] [PubMed]
- Oduola, A.O.; Obembe, A.; Lateef, S.A.; Abdulbaki, M.K.; Kehinde, E.A.; Adelaja, O.J.; Shittu, O.; Tola, M.; Oyeniyi, T.A.; Awolola, T.S. Species Composition and Plasmodium Falciparum Infection Rates of Anopheles Gambiae s.l. Mosquitoes in Six Localities of Kwara State, North Central, Nigeria. jasem 2022, 25, 1801–1806. [Google Scholar] [CrossRef]
- Garba, M.N.; Moustapha, L.M.; Sow, D.; Karimoun, A.; Issa, I.; Sanoussi, M.K.; Diallo, M.A.; Doutchi, M.; Diongue, K.; Ibrahim, M.L.; et al. Circulation of Non-Falciparum Species in Niger: Implications for Malaria Diagnosis. Open Forum Infectious Diseases 2024, ofae474. [Google Scholar] [CrossRef]
- Maiga, A.-A.; Sombié, A.; Zanré, N.; Yaméogo, F.; Iro, S.; Testa, J.; Sanon, A.; Koita, O.; Kanuka, H.; McCall, P.J.; et al. First Report of V1016I, F1534C and V410L Kdr Mutations Associated with Pyrethroid Resistance in Aedes Aegypti Populations from Niamey, Niger. PLoS ONE 2024, 19, e0304550. [Google Scholar] [CrossRef]
- Mamane Salé, N.; Labbo, R.; Laminou, I.M.; Issa Arzika, I.; Djibo Souley, A.; Zoulkifouly Hounkarin, W.; Zamanka Naroua, H.; Soumana, A.; Maiga, A.-A.; Jambou, R.; et al. Insecticide Resistance in Anopheles Gambiae Sensu Lato (Diptera: Culicidae) across Different Agroecosystems in Niamey, Niger. AJTER 2024, 3, 30–40. [Google Scholar] [CrossRef]
- Soumaila, H.; Hamani, B.; Arzika, I.I.; Soumana, A.; Daouda, A.; Daouda, F.A.; Iro, S.M.; Gouro, S.; Zaman-Allah, M.S.; Mahamadou, I.; et al. Countrywide Insecticide Resistance Monitoring and First Report of the Presence of the L1014S Knock down Resistance in Niger, West Africa. Malar J 2022, 21, 385. [Google Scholar] [CrossRef]
- Soumaila, H.; Idrissa, M.; Akgobeto, M.; Habi, G.; Jackou, H.; Sabiti, I.; Abdoulaye, A.; Daouda, A.; Souleymane, I.; Osse, R. Multiple Mechanisms of Resistance to Pyrethroids in Anopheles Gambiae s. l Populations in Niger. Médecine et Maladies Infectieuses 2017, 47, 415–423. [Google Scholar] [CrossRef]
- Busari, L.O.; Raheem, H.O.; Iwalewa, Z.O.; Fasasi, K.A.; Adeleke, M.A. Investigating Insecticide Susceptibility Status of Adult Mosquitoes against Some Class of Insecticides in Osogbo Metropolis, Osun State, Nigeria. PLoS One 2023, 18, e0285605. [Google Scholar] [CrossRef]
- Chukwuekezie, O.; Nwosu, E.; Nwangwu, U.; Dogunro, F.; Onwude, C.; Agashi, N.; Ezihe, E.; Anioke, C.; Anokwu, S.; Eloy, E.; et al. Resistance Status of Anopheles Gambiae (s. l.) to Four Commonly Used Insecticides for Malaria Vector Control in South-East Nigeria. Parasit Vectors 2020, 13, 152. [Google Scholar] [CrossRef]
- Muhammad, A.; Ibrahim, S.S.; Mukhtar, M.M.; Irving, H.; Abajue, M.C.; Edith, N.M.A.; Da’u, S.S.; Paine, M.J.I.; Wondji, C.S. High Pyrethroid/DDT Resistance in Major Malaria Vector Anopheles Coluzzii from Niger-Delta of Nigeria Is Probably Driven by Metabolic Resistance Mechanisms. PLoS One 2021, 16, e0247944. [Google Scholar] [CrossRef]
- Ojianwuna, C.C.; Enwemiwe, V.N.; Esiwo, E.; Mekunye, F.; Anidiobi, A.; Oborayiruvbe, T.E. Susceptibility Status and Synergistic Activity of DDT and Lambda-Cyhalothrin on Anopheles Gambiae and Aedes Aegypti in Delta State, Nigeria. PLoS One 2024, 19, e0309199. [Google Scholar] [CrossRef] [PubMed]
- Omotayo, A.I.; Ande, A.T.; Oduola, A.O.; Adelaja, O.J.; Adesalu, O.; Jimoh, T.R.; Ghazali, A.I.; Awolola, S.T. Multiple Insecticide Resistance Mechanisms in Urban Population of Anopheles Coluzzii (Diptera: Culicidae) from Lagos, South-West Nigeria. Acta Trop 2022, 227, 106291. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.S.; Muhammad, A.; Hearn, J.; Weedall, G.D.; Nagi, S.C.; Mukhtar, M.M.; Fadel, A.N.; Mugenzi, L.J.; Patterson, E.I.; Irving, H.; et al. Molecular Drivers of Insecticide Resistance in the Sahelo-Sudanian Populations of a Major Malaria Vector Anopheles Coluzzii. BMC Biol 2023, 21, 125. [Google Scholar] [CrossRef]
- Awolola, T.S.; Adeogun, A.; Olakiigbe, A.K.; Oyeniyi, T.; Olukosi, Y.A.; Okoh, H.; Arowolo, T.; Akila, J.; Oduola, A.; Amajoh, C.N. Pyrethroids Resistance Intensity and Resistance Mechanisms in Anopheles Gambiae from Malaria Vector Surveillance Sites in Nigeria. PLoS ONE 2018, 13, e0205230. [Google Scholar] [CrossRef]
- Dada, N.; Sheth, M.; Liebman, K.; Pinto, J.; Lenhart, A. Whole Metagenome Sequencing Reveals Links between Mosquito Microbiota and Insecticide Resistance in Malaria Vectors. Sci Rep 2018, 8, 2084. [Google Scholar] [CrossRef]
- Herren, J.K.; Mbaisi, L.; Mararo, E.; Makhulu, E.E.; Mobegi, V.A.; Butungi, H.; Mancini, M.V.; Oundo, J.W.; Teal, E.T.; Pinaud, S.; et al. A Microsporidian Impairs Plasmodium Falciparum Transmission in Anopheles Arabiensis Mosquitoes. Nat Commun 2020, 11, 2187. [Google Scholar] [CrossRef]
- Nattoh, G.; Maina, T.; Makhulu, E.E.; Mbaisi, L.; Mararo, E.; Otieno, F.G.; Bukhari, T.; Onchuru, T.O.; Teal, E.; Paredes, J.; et al. Horizontal Transmission of the Symbiont Microsporidia MB in Anopheles Arabiensis. Front. Microbiol. 2021, 12, 647183. [Google Scholar] [CrossRef]
- Makhulu, E.E.; Onchuru, T.O.; Gichuhi, J.; Otieno, F.G.; Wairimu, A.W.; Muthoni, J.N.; Koekemoer, L.; Herren, J.K. Localization and Tissue Tropism of the Symbiont Microsporidia MB in the Germ Line and Somatic Tissues of Anopheles Arabiensis. mBio 2024, 15, e02192-23. [Google Scholar] [CrossRef]
- Akorli, J.; Akorli, E.A.; Tetteh, S.N.A.; Amlalo, G.K.; Opoku, M.; Pwalia, R.; Adimazoya, M.; Atibilla, D.; Pi-Bansa, S.; Chabi, J.; et al. Microsporidia MB Is Found Predominantly Associated with Anopheles Gambiae s. s and Anopheles Coluzzii in Ghana. Sci Rep 2021, 11, 18658. [Google Scholar] [CrossRef]
- Ahouandjinou, M.J.; Sovi, A.; Sidick, A.; Sewadé, W.; Koukpo, C.Z.; Chitou, S.; Towakinou, L.; Adjottin, B.; Hougbe, S.; Tokponnon, F.; et al. First Report of Natural Infection of Anopheles Gambiae s. s. and Anopheles Coluzzii by Wolbachia and Microsporidia in Benin: A Cross-Sectional Study. Malar J 2024, 23, 72. [Google Scholar] [CrossRef]
- Bukhari, T.; Pevsner, R.; Herren, J. Keith. Microsporidia: A Promising Vector Control Tool for Residual Malaria Transmission. Front. Trop. Dis 2022, 3, 957109. [Google Scholar] [CrossRef]
- Coetzee, M. Key to the Females of Afrotropical Anopheles Mosquitoes (Diptera: Culicidae). Malar J 2020, 19, 70. [Google Scholar] [CrossRef] [PubMed]
- Santolamazza, F.; Mancini, E.; Simard, F.; Qi, Y.; Tu, Z.; della Torre, A. Insertion Polymorphisms of SINE200 Retrotransposons within Speciation Islands of Anopheles Gambiae Molecular Forms. Malaria Journal 2008, 7, 163. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J. Organization and Mapping of a Sequence on the DROSOPHILA MELANOGASTER X and Y Chromosomes That Is Transcribed during Spermatogenesis. Genetics 1984, 107, 611–634. [Google Scholar] [CrossRef]
- WHO Standard Operating Procedure for Testing Insecticide Susceptibility of Adult Mosquitoes in WHO Tube Tests. 2022.
- Lumivero XLSTAT | Logiciel statistique pour Excel Available online: https://www.xlstat.com/fr/.
- Nattoh, G.; Onyango, B.; Makhulu, E.E.; Omoke, D.; Ang’ang’o, L.M.; Kamau, L.; Gesuge, M.M.; Ochomo, E.; Herren, J.K. Microsporidia MB in the Primary Malaria Vector Anopheles Gambiae Sensu Stricto Is Avirulent and Undergoes Maternal and Horizontal Transmission. Parasites & Vectors 2023, 16, 335. [Google Scholar] [CrossRef]
- Akorli, E.A.; Nana Efua, A.; Egyirifa, R.K.; Dorcoo, C.; Otoo, S.; Tetteh, S.N.A.; Pul, R.M.; Derrick, B.S.; Oware, S.K.D.; Samuel, K.D.; et al. Breeding Water Parameters Are Important Determinants of Microsporidia MB Prevalence in the Aquatic Stages of Anopheles Mosquitoes. Available online: https://www.researchsquare.com (accessed on 2 January 2025).
- Omoke, D.; Kipsum, M.; Otieno, S.; Esalimba, E.; Sheth, M.; Lenhart, A.; Njeru, E.M.; Ochomo, E.; Dada, N. Western Kenyan Anopheles Gambiae Showing Intense Permethrin Resistance Harbour Distinct Microbiota. Malaria Journal 2021, 20, 77. [Google Scholar] [CrossRef]
- Pelloquin, B.; Kristan, M.; Edi, C.; Meiwald, A.; Clark, E.; Jeffries, C.L.; Walker, T.; Dada, N.; Messenger, L.A. Overabundance of Asaia and Serratia Bacteria Is Associated with Deltamethrin Insecticide Susceptibility in Anopheles Coluzzii from Agboville, Côte d’Ivoire. Microbiology Spectrum 2021, 9, e00157-21. [Google Scholar] [CrossRef]
- Boanyah, G.Y.; Koekemoer, L.L.; Herren, J.K.; Bukhari, T. Effect of Microsporidia MB Infection on the Development and Fitness of Anopheles Arabiensis under Different Diet Regimes. Parasites & Vectors 2024, 17, 294. [Google Scholar] [CrossRef]
- Otieno, F.G.; Barreaux, P.; Belvinos, A.S.; Makhulu, E.E.; Onchuru, T.O.; Wairimu, A.W.; Omboye, S.M.; King’ori, C.N.; Mawuko, S.B.; Kebira, A.N.; et al. Temperature Modulates the Dissemination Potential of Microsporidia MB, a Malaria-Blocking Endosymbiont of Anopheles Mosquitoes 2024, 2024.11.15.623820.
Figure 1.
Map of sampling locations in Nigeria and Niger Republic.
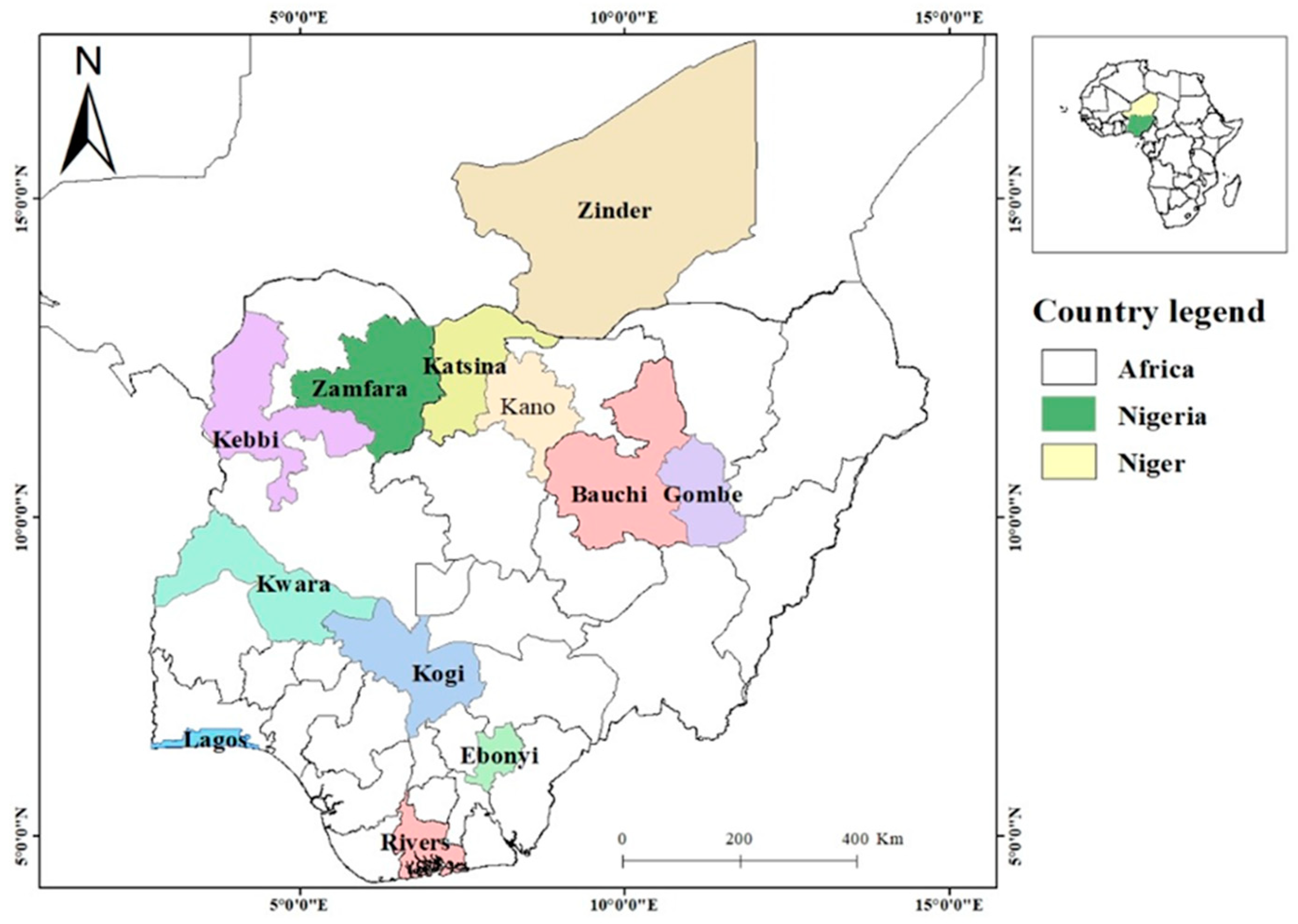
Figure 2.
Spatial distribution of MB along clinal gradient of aridity spanning Nigeria and Niger Republic.
Figure 2.
Spatial distribution of MB along clinal gradient of aridity spanning Nigeria and Niger Republic.
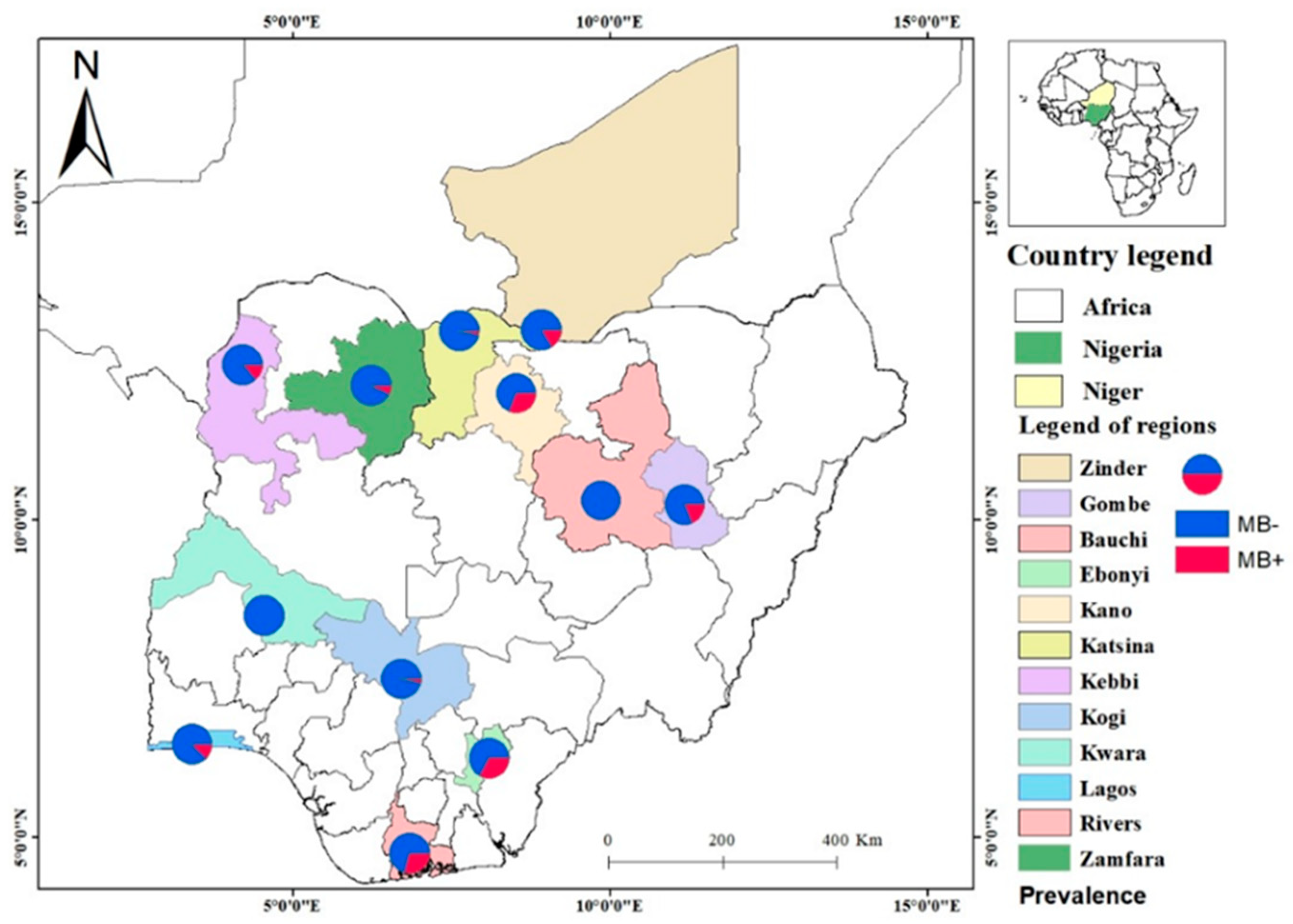
Figure 3.
Spatial association between Microsporidia MB Infection Status and Anopheles species.
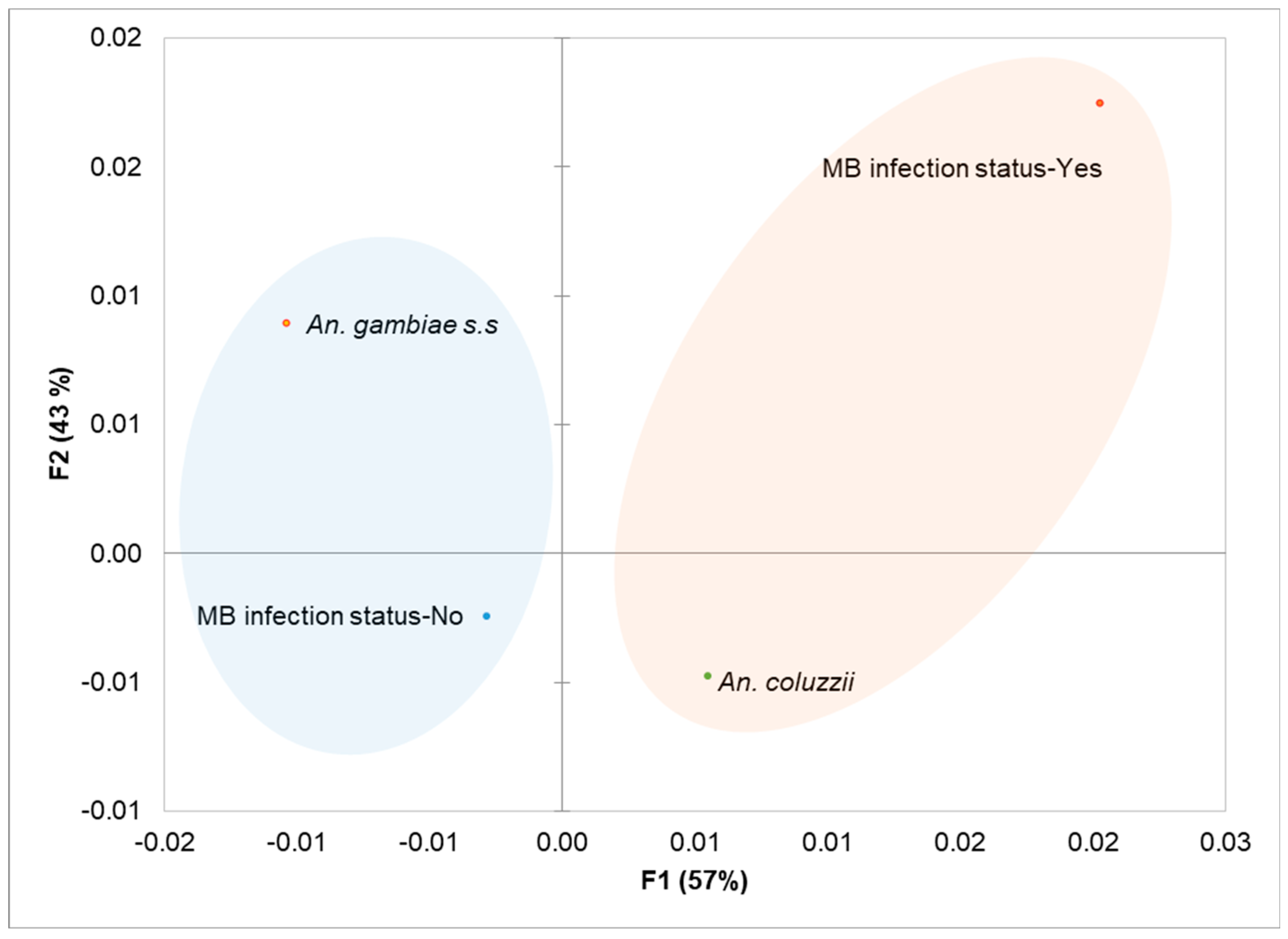
Figure 4.
Spatial depiction of potential correlation between MB infection and pyrethroid resistance.
Figure 4.
Spatial depiction of potential correlation between MB infection and pyrethroid resistance.
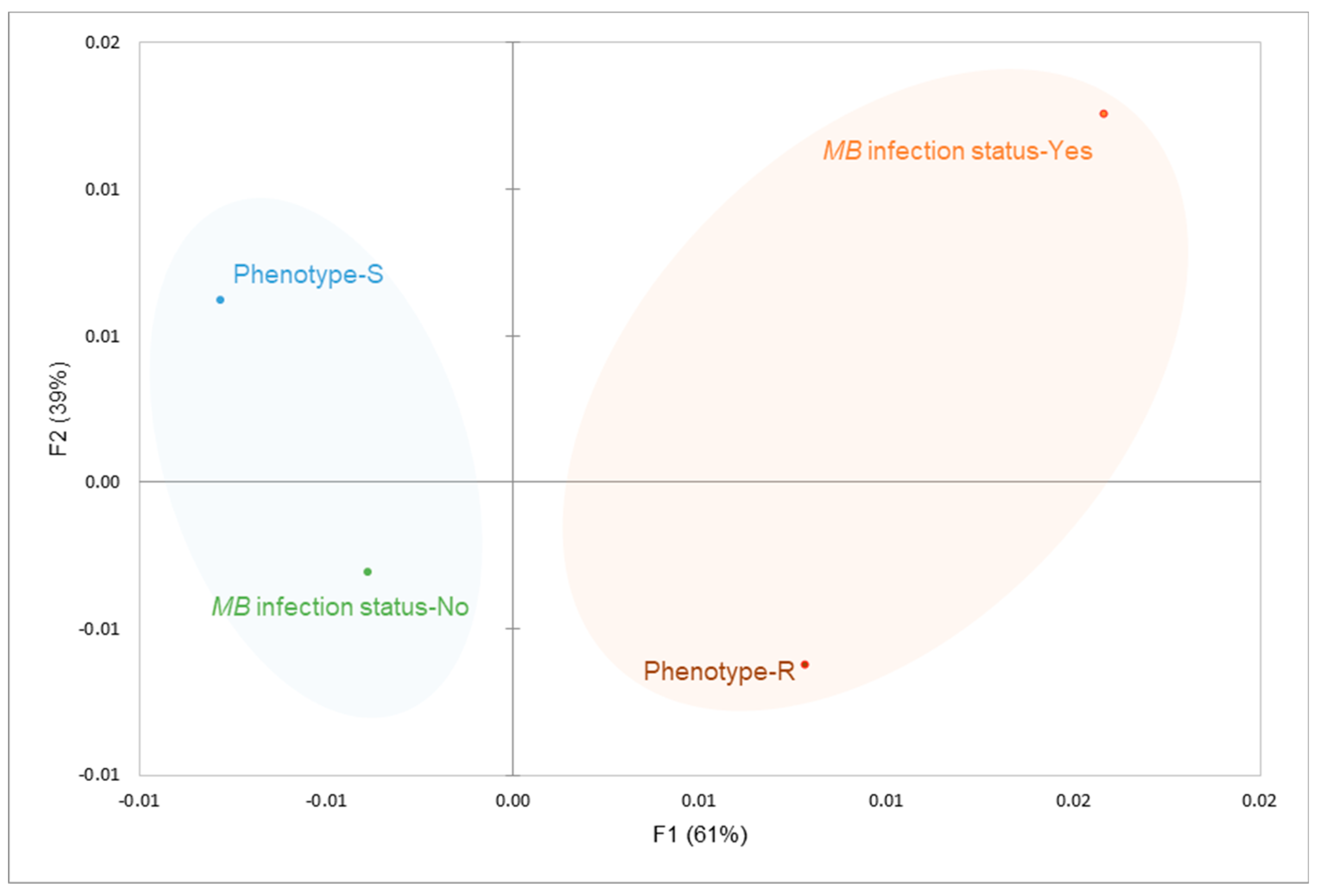

Table 1.
Sampling design and localities for Anopheles mosquito collection and bioassays.
| Locality | n | Species | Bioassay | Ecological setting |
|---|---|---|---|---|
| F0 Parents caught indoor using Prokopack aspirators | ||||
| Gayi Niger, Magaria | 32 | An. coluzzii | - | Sahel |
| Gajerar Giwa, Katsina State | 32 | An. coluzzii | Sahel | |
| Gaa-Bolorunduro, Kwara State | 15 | An. gambiae s.s. | - | Guinea Savanna |
| Zariagi, Lokoja, Kogi State | 32 | An. gambiae s.s. | - | Guinea Savanna |
| Gotomo, Kebbi State | 15 | An. coluzzii | - | Sudan Savanna |
| Tsunami, Gusau, Zamfara State | 15 | An. coluzzii | - | Sudan Savanna |
| Ugalaba, Ezza North, Ebonyi State | 15 | An. coluzzii | - | Tropical Rainforest |
| Obio/Akpor LGA, Rivers State | 24 | An. coluzzii | - | Mangrove Swamp |
| Badagry, Lagos State | 24 | An. gambiae s.s. | - | Mangrove Swamp |
| F1 progenies used for bioassays | ||||
| Gajerar Giwa, Katsina State | 16 | An. coluzzii | Permethrin | Sahel |
| Dadin Kowa, Gombe State | 16 | An. coluzzii | Deltamethrin | Guinea Savanna |
| Zariagi, Lokoja, Kogi State | 24 | An. gambiae s.s. | Deltamethrin | Guinea Savana |
| Gamjin Bappa, Karaye, Kano State | 16 | An. coluzzii | Deltamethrin | Guinea Savana |
| Gadau, Bauchi State | 16 | An. coluzzii | Deltamethrin | Sudan Savanna |
| Obio/Akpor LGA, Rivers State | 24 | An. coluzzii | Permethrin | Mangrove Swamp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



