
Submitted:
29 January 2025
Posted:
30 January 2025
You are already at the latest version
Abstract
Alternative protein ingredients and vegan products are in demand by consumers due to environmental, animal welfare, and economic reasons associated with consumption of soy and animal proteins. Herein, we describe the development of a marine plant (seaweed) alternative protein ingredient developed from the red seaweed Chondrus crispus Stakehouse and its application as an alternative to egg as an emulsifier ingredient in a vegan fairy cake recipe. Development of alternative proteins faces challenges in recent years including increased costs for production and growing competition. For these reasons, the generation and characterisation of the ingredient for techno-functional attributes is described along with predicted bioactivities and potential health benefits associated with the peptides generated in the hydrolysed protein. A prototype vegan fairy cake made using the C. crispus hydrolysate used as an emulsifier is also described and product proximate qualities are compared to a traditionally manufactured fairy cake. Results suggest that hydrolysis of C. crispus increases the protein content of C.crispus hydrolysate relative to the whole biomass and makes the application of C. crispus in food products more attractive by improving the technofunctional attributes of the protein source, increasing emulsifying properties and enhancing tertiary benefits like bioactive potential beyond basic, nutrition.
Keywords:
Chondrus crispus
; hydrolysis
; bioactive peptides
; emulsifiers
; anti-diabetic potential
; vegan
; baked goods
1. Introduction
Proteins are instrumental in defining the functional characteristics of food products, exerting a profound impact on consumer acceptance. They are pivotal in food processing and product development, as Ogunwolu et al., 2009 highlighted [1]. These proteins are broadly categorized into animal proteins like gelatin and milk and plant proteins like soya, peanut, and wheat. Due to concerns regarding animal protein sources’ availability, cost, and associated health risks, nutritionists are increasingly exploring alternative plant protein sources for both human consumption and feedstock preparation. Plant proteins are favoured in various food applications due to their desirable functional properties, including emulsification, fat and water absorption, texture modification, colour control, and whipping abilities. Despite being more economical than animal protein sources, plant protein supplements often lack certain essential amino acids, energy, and minerals like phosphorus and may contain anti-nutritional factors [2]. The global demand for cost-effective protein sources in value-added food products is escalating, prompting a search for economically viable alternatives, such as marine biomass, considering the susceptibility of plant protein sources to climate change and their reliance on agricultural land. The food industry’s interest in seaweed proteins has surged, driven by the need to enhance product nutritional quality and functionality, especially in developing countries [3].
Seaweeds are widely embraced as a culinary ingredient in coastal cuisines worldwide and have been a traditional dietary staple in Asian cultures. However, they have gained popularity in Norway as animal fodder [4]. Among the approximately 250 seaweed species commercially utilized globally, about 150 are favoured for human consumption, while in Western countries, they serve primarily as sources of polysaccharides like agar, alginates, or carrageenans for the food and pharmaceutical industries. Nutritionally, edible seaweeds offer low-calorie, high-nutrient value due to their rich content of vitamins, proteins, and minerals, including vitamins A, B1, B12, and C, as well as thiamine, riboflavin, β-carotene, tocopherols, and long-chain polyunsaturated essential fatty acids like eicosapentaenoic acid [5]. Their protein and lipid quality, characterized by high levels of essential amino acids and unsaturated fatty acids, surpasses many other vegetables, making them valuable for human and animal consumption. While seaweeds are predominantly utilized for their mineral content and polysaccharide functional properties in food and feed, their nutritional value, particularly regarding proteins, is often overlooked [6].
Recently, there has been growing interest in exploring seaweed as a viable and economical alternative source for proteins. Research indicates that the nutritional potential and protein content of seaweeds differs depending on the environmental factors, geographical location, seasonal variations and species, including red (Rhodophyta), brown (Ochrophyta), and green (Chlorophyta) seaweeds [7,8,9]. Notably, red seaweeds are the most significant protein source, boasting protein levels ranging from 19% to 44% of dry weight (dw) and recognized for their substantial content of readily available protein, making them an appealing protein source [10]. Conversely, green and brown seaweeds exhibit lower protein concentrations, approximately 20% or 10% of dry weight [11]. These protein levels rival or slightly surpass those found in legumes (20–30%), cereals (10–15%), or nuts (20–30%) [12]. Despite this potential, considerable challenges are associated with extracting proteins from seaweed, leading to a scarcity of studies on seaweed protein quality [13]. Surprisingly, there is a notable lack of research focused on the suitability of seaweed protein for human consumption. The extraction of proteins from seaweed presents challenges due to the presence of phenolic compounds [14] and abundant polyanionic cell wall mucilage [15,16]. Phenolic compounds can disrupt native protein structures and can form covalent bonds with them, particularly in oxidative environments [17]. Moreover, the presence of cell wall mucilages leads to the formation of dense solutions, thereby hindering the extraction and purification of proteins [18,19]. In order to optimize the applicability of seaweed proteins as functional ingredients in innovative food formulations, it is imperative to process whole biomass into high-protein products sustainably. One practical approach involves the generation of protein hydrolysates using various enzymes. Hydrolysates are pre-digested proteins broken down into smaller peptides and amino acids, manifesting improved bioavailability and faster absorption of nutrients [20] They often have improved solubility and dispersibility, facilitating their incorporation into diverse food matrices without affecting their texture or taste [21]. Furthermore, hydrolysis can remove undesirable flavours and odours associated with seaweed proteins, resulting in a more palatable product [22]. Lastly, seaweed protein hydrolysates demonstrate enhanced techno-functional properties such as emulsification, foaming, and gelation, making them suitable for a broader range of applications in the food and beverage industry [23].
Chondrus crispus Stakehouse, commonly known as Irish moss or carrageen moss or Mousse d'Irlande, has a rich history of human consumption dating back centuries [24] Chondrus crispus has been employed as a thickening agent in soups, stews, and desserts, imparting a unique texture and a subtle briny flavour [24]. Beyond its culinary applications, this seaweed has also been traditionally utilized for its potential medicinal properties, believed to aid in digestion, soothe respiratory ailments, and even promote skin health [24]. Additionally, it is worth highlighting its protein content, which averages 27.2 ± 1.4%, making it a valuable plant-based protein source for those interested in alternative protein sources [25]. With sustainability at the forefront, the present investigation prioritizes the production of protein hydrolysates from Chondrus crispus, utilizing both carbohydrase and protease enzymes. Subsequently, we meticulously analyze the techno-functional attributes of these hydrolysates, paving the way for a comprehensive evaluation of their potential as emulsifiers in vegan fairy cakes.
2. Materials and Methods
2.1. Biological Materials, Chemical Reagents, and Fairy Cake Ingredients
The red Irish seaweed (Chondrus crispus Stakehouse) was harvested off the West Coast of Ireland and supplied dried and milled by the Irish company SeaLac Ltd, Sligo, Ireland. Samples were freeze-dried, milled and vacuum-preserved until further analysis. To avoid physicochemical modifications of the protein, all reagents used during these experiments were food-grade chemicals. Sunflower, rapeseed, and peanut oils from Musgrave House Ltd. (Cork, Ireland) were purchased for use in this study.
2.2. Chemicals and Reagents
Merck Ltd. (Dublin, Ireland) supplied the enzymes Viscozyme and Alcalase; Hydrochloric acid (HCl) and sodium hydroxide (NaOH). Deionized water obtained from a Millipore nano-water purification and filtration system was supplied by Sigma-Aldrich (Dublin, Ireland). The Stuart SWB6D digital water bath and Stuart shaking incubator were supplied by Cole-Parmer (Saint Neots, England, UK).
The ingredients used to prepare the fairy cake were: Self-raising flour (Shackleton, Ireland), castor sugar (Supervalu, purchased locally, Dublin, Ireland), fresh whole eggs, rapeseed oil, salt (purchased locally,Dublin, Ireland), sodium bicarbonate (Royal, baking powder, bought locally, Dublin, Ireland), and vanilla extract (Dublin, Ireland).
2.3. Preparation, Extraction and Enrichment of Protein Hydrolysates from Chondrus crispus
The hydrolysis procedure for dried and milled Chondrus crispus was conducted independently in triplicate as outlined below: Initially, 50 grams of seaweed were combined with 500 millilitres of MilliQ deionized water. This mixture underwent heat treatment at 95°C for 15 minutes in a Stuart SWB6D digital water bath (St. Neots, UK) to deactivate inherent enzymes. After cooling to room temperature, the pH was adjusted to 5 using 0.1 M NaOH and 0.1 M HCl. Viscozyme was introduced at a concentration of 1% (w/v), followed by a 2-hour incubation in a Stuart shaking incubator (Saint Neots, England, UK) set at 45°C and 220 rpm. Hydrolysis was terminated by subjecting the mixture to 10 minutes of heating at 95°C. The pH of the samples was then adjusted to 8 – 8.5 using 0.1 M NaOH and 0.1 M HCl. Subsequently, the samples underwent a second round of hydrolysis with Alcalase (1% w/v) under shaking incubation conditions at 55°C and 220 rpm for 2 hours. The enzyme was deactivated by heating at 95°C for 10 minutes. The samples were then centrifuged at 8000x g for 30 minutes (Thermofisher Scientific Lynx 6000 centrifuge, Dublin, Ireland), followed by the separation of supernatant and residue fractions. Both the spent biomass and supernatants were frozen at -20°C and later freeze-dried using an FD80GP freeze-drier (Cuddon Freeze Dry, New Zealand). The supernatant and spent biomass fractions were stored at -80°C until further analysis.
2.4. Proximate Analysis
2.4.1. Ash Content
In order to assess the ash content of the whole biomass, protein hydrolysate, and spent biomass of Chondrus crispus, each sample was individually weighed into dried crucibles. Before initiating the ashing process, the samples were subjected to charring on a hotplate for a duration of 3 hours at 100 °C. Subsequently, the pre-charred samples were transferred to a muffle furnace and exposed to ashing in air for 24 hours at 540 °C. Following ashing, the samples were allowed to cool to room temperature within a desiccator. The percentage of ash was then determined utilizing formula 1,
% Ash = [(M2 – M0) / (M1 – M0)] x 100,
M0 is the mass of the dish, M1 is the mass of the dish and test portion, and M2 is the mass of the dish and ash portion.
2.4.2. Protein Content
The protein content assessment of all samples involved the determination of nitrogen content using the Dumas method with a LECO FP328 Protein analyzer (LECO Corp., Saint Joseph, MI, USA), in accordance with the protocols outlined in the Association of Official Analytical Chemists (AOAC) method 992.15 [26]. Prior to analysis, each sample was weighed into foil cups, and their respective weights were recorded. These samples were then analyzed using the LECO FP328 arranged in numbered carousels. The combustion and nitrogen quantification steps were executed utilizing a specific nitrogen conversion factor of 4.59 tailored for Chondrus crispus, established from the literature [27].
2.4.3. Lipid Content
The determination of total fat content followed a gravimetric procedure utilizing an ORACLE Rapid NMR Fat Analyzer (CEM Corp., Charlotte, NC, USA) in accordance with AOAC official method 2008.06 and AOAC PVM 1:2004, with adaptations drawn from the work of [28] All samples were weighed on CEM glass fibre sample pads and subjected to overnight drying in an oven. After recording the weights of the dried samples, the pads containing the samples were enveloped in Trac Film. Subsequently, these wrapped samples were inserted into an ORACLE tube and subjected to heating for 30 minutes at 40°C on a heat block before undergoing fat analysis using the ORACLE instrument.
2.4.4. Moisture Content
The total moisture content was assessed employing a LECO TGM 800 Moisture Analyzer (LECO Corp., Saint Joseph, MI, USA). Individual samples were weighed into foil cups, and their weights were recorded before drying in the LECO TGM 800, arranged in numbered carousels. The decrease in mass of the sample following drying was utilized to calculate the moisture content, following the methodology previously outlined by [29].
2.5. Colour Evaluation
The colour of the seaweed protein hydrolysates and whole biomass was assessed utilizing an UltraScan PRO Spectrophotometer (HunterLab, Germany) equipped with illuminant D65, diffuse 8° observer angle and automated specular component. Calibration was performed using white and black standards (HunterLab, Germany) prior to all measurements. Results were recorded in the CIE Lab* system, where L* indicates lightness, a* denotes redness/greenness, and b* represents yellowness/blueness.
The chroma (C*) and hue (h°) values were calculated using the following equations (2 and 3):
C* = √ (a*2) + (b*2),
h° = (tan-1) (b*/a*) × 180/π,
In order to assess the colour disparity between the hydrolysates and the whole biomass, we computed ΔE*ab using the formula (4). This metric quantifies the spatial gap between two colours within the CIELAB colour space, delineated by their respective L*, a*, and b* coordinates.
∆E*ab = √(L*1 - L*2)2 + (a*1 - a*2)2 + (b*1 - b*2)2,
L*1, a*1, and b*1 represent the colour attributes of the protein hydrolysates, while those of the whole biomass are denoted by L*2, a*2, and b*2 for comparison purposes. A ΔE*ab value exceeding two is typically deemed perceptually significant.
2.6. Determination of pH and Water Activity (aW)
The freeze-dried protein hydrolysate extract was reconstituted in distilled water at a concentration of 1% w/v, and the pH was determined using a pH meter (Orion model 420A, Thermo Orion, Cambridgeshire, UK).
The water activity of the extracted hydrolysate was assessed utilizing an AquaLab Water Activity System (Pullman, Washington, USA). The measurement was conducted at a temperature of 8.17 ± 0.07 °C.
2.7. Technofunctional and Bioactive Properties
2.7.1. Water (WHC) and Oil Holding Capacity (OHC)
The water-holding capacity (WHC) and oil-holding capacity (OHC) of the seaweed protein hydrolysates were determined following the method outlined by Bencini (1986) with minor adjustments [30]. Specifically, 1 gram of protein hydrolysate was combined with 10 mL of either distilled water or sunflower oil using a vortex mixer (Henry Troemner, USA), followed by centrifugation at 2200g for 30 minutes. After centrifugation, the supernatants were carefully decanted, and the centrifuge tubes containing the sediment were weighed. WAC and OAC were quantified as the grams of water or sunflower oil retained per gram of protein hydrolysate, calculated using the provided formula (5).
WHC / OHC (g of H2O or sunflower oil / g of protein hydrolysate) = [(W2 - W1) / W0] × 100,
W0 is the weight of the dry sample (g), W1 is the weight of the tube plus the dry sample (g), and W2 is the weight of the tube plus the sediment (g).
2.7.2. Emulsifying Activity and Emulsion Stability
The emulsifying activity (EA) of the protein hydrolysates was evaluated following the methodology established by Naczk, Diosady, and Rubin [31], with minor adaptations. Initially, the hydrolysate sample was diluted in Millipore ultrapure water (Millipore, Ireland) to a concentration of 1% w/v, and the pH was adjusted to pH 2, 4, 6, 8 and 10 using either 0.1 M NaOH or 0.1 M HCl. The protein solution underwent homogenization for 30 seconds at 14,000 rpm. Subsequently, to create the emulsion, commercial oils such as sunflower, rapeseed, and peanut oil were introduced into the aqueous phase containing the protein hydrolysate at a ratio of 3 parts oil to 2 parts protein solution. This addition of oil occurred in two stages: firstly, half of the oil volume was added and mixed for 30 seconds at 14,000 rpm, followed by the addition of the remaining oil volume and further mixing for 90 seconds at the same speed. The resulting emulsion was transferred to centrifuge tubes and centrifuged at 1100g for 5 minutes, after which the volume of the emulsion layer was measured. The EA was then determined using the specified formula (6);
EA % = (VE / VT) × 100,
VE is the volume of the emulsion layer after centrifuging, and VT is the total volume inside the tube.
For emulsion stability (ES), the previously prepared emulsions were heated at 85°C for 15 minutes, cooled at room temperature for 10 minutes and centrifuged again at 1100 g for 5 minutes. The ES was expressed as the % of the emulsion layer remaining after centrifugation (formula 7):
ES % = (VEmulsion after heating / VOriginal emulsion) × 100,
2.7.3. Foaming Capacity and Stability
The protein hydrolysates’ foaming capacity (FC) was assessed following the method established by Poole, West, and Walters in 1984, with slight adjustments [32]. Suspensions of hydrolysates were prepared at room temperature with a concentration of 1.5% w/v, and the pH was adjusted to 2, 4, 6, 8, and 10 using 0.1M NaOH or 0.1 M HCl, respectively. The suspensions underwent homogenization using a T 25 digital ULTRA-TURRAX® homogenizer (IKA®, Germany) at 10,000 rpm for 1 minute, and the resulting foam volume was quantified using a graduated cylinder. The foaming capacity was determined by calculating the volume of foam produced as a percentage of the initial solution volume using the provided formula (8);
FC % = [(VF – V0)/ (V0)] × 100,
V0 is the initial volume of protein solution before homogenization, and VF is the volume of foam generated after homogenization.
The foam stability was determined by using the formula (9) that measures the decrease in volume of foam as a function of time up to a period of 15 minutes.
FS % = (VS / VF) × 100,
VS is the volume of protein solution after standing for 15 minutes, and VF is the volume of foam generated after homogenization.
2.7.4. Mass Spectrometry in Tandem Analysis
1 µL of the C. crispus hydrolysate sample was quantified by nanoDrop (A 280 nm, ε= 1mg/mL). The volume corresponding to 190 ng was diluted in 20 µL with 0.1% Formic acid (FA) and loaded in an Evotip pure tip (EvoSep) according to the manufacturers’ instructions. Tandem mass spectrometry analysis (LC–MS/MS) was performed in a Tims TOF fleX mass spectrometer (Bruker). The sample loaded in the Evotip pure was eluted to an analytical column (EvoSep 15 cm x 150 µm, 1.5 µm; Evosep) by the Evosep One system, and solved with the 30 SPD chromatographic method defined by the manufacturer.
The eluted peptides were ionized in a captive Spray with 1700 V at 200 ºC, and analyzed using ddaPASEF mode with the following settings: TIMS settings Mode: custom; 1/K0: 0.7-1.76 V.s/cm2; ramp time: 100 ms; Duty Cylce: 100%; Ramp Rate: 9.42 Hz; Ms Averaging:1; Auto Calibration: off. MS settings: Scan: 100-1700 m/z; Ion Polarity: Positive; Scan Mode: PASEF. MS/MS: Number of PASEF ramps: 4; Total Cycle time: 0.5 s; Charge Minimun:0 (unkwon); Charge maximum: 5; Scheduling: target Intensity: 12500, Intensity Threshold: 1000. Active exclusion: ON. The system sensitivity was controlled with 20 ng of HELA digested proteins. 4646 proteins identified with the 30 SPD gradient.
2.7.5. Protein Identification
MSFragger searches were performed (via FragPipe) for the identification of non-tryptic peptides. Single databases were generated at Rhodophyta and Uniprot-Chondrus. Every sample was analyzed with the adequate database.
2.7.6. In silico Analysis of Identified Peptides to Predict Bioactivities
Following identification of peptides (>2000, Table S1) found within the C. crispus hydrolysate using MS, potential bioactivities were determined using an in silico approach previously used by our group in relation to characterisation of peptides from hydrolysates of the microalga Porphyridium sp., mesopelagic fish [33,34] and the brown seaweed Laminaria digitata [35]. Briefly, the identified peptides (>2000) were assessed for their potential bioactivity (antimicrobial) nature using Peptide Ranker ( - accessed on the 17th of January 2025) based on amino acid charge and peptide structure, as described previously (Mooney et al.,). The anti-inflammatory predictive programme PreAIP -http://kurata14.bio.kyutech.ac.jp/PreAIP/ (accessed on 17th of January 2025) [36] ranked peptides in terms of their potential anti-inflammatory potential by viewing the primary peptide sequence, as well as evolutionary and structural information of the peptide, through a random forest classifier [36]. Scores ≥ 0.468 indicate high confidence that a peptide is anti-inflammatory. Scores between 0.468 and ≥ 0.388 indicate medium confidence that a peptide is anti-inflammatory, while a score of 0.388 to 0.342 indicates that the peptide has a low probability of having anti-inflammatory activity. Peptides were identified as novel by performing a search in the database BIOPEP-UWM. The potential of peptides within the C. crispus hydrolysate to impact taste was determined using Umami-MRNN https://umami-mrnn.herokuapp.com/ (accessed on 17th of January 2025) [37]. The programme AntiDMPpred, available at http://i.uestc.edu.cn/AntiDMPpred/cgi-bin/AntiDMPpred.pl (accessed on 17th of January 2025), was used to identify the anti-diabetes type 2 activity of peptides using a threshold of 0.6 [38]. Values with threshold figures greater than 0.6 have a higher probability of having anti-diabetic benefits.
2.8. Preparation of Fairy Cakes
To prepare 09 vegan fairy cake samples, the cake recipe outlined in Table 1 was adapted;
The oat milk portion in the formulation was substituted with seaweed protein hydrolysates based on their protein content. Oat milk typically contains around 1% protein [39], meaning that 150 ml of oat milk provides approximately 1.5 grams of protein. The protein hydrolysates were included in the formulation based on their protein content. In a large mixing bowl, self-raising flour, seaweed protein hydrolysates, caster sugar, salt, and baking powder were sifted together. A mixture of rapeseed oil and vanilla essence was gradually added to 200 ml of water and then slowly incorporated into the dry ingredients, followed by thorough manual stirring. The resulting cake batter was evenly distributed into cake cases, filling them almost to the top. The cakes were baked in the centre of a preheated conventional oven (Macpan MDBt8, Thiene, Italy) at 200°C (180°C fan) for 20–25 minutes or until golden brown afterwards allowed to cool to room temperature.
For preparing nine fairy cake control samples, the recipe outlined in Table 2 was adapted;
All the ingredients are mixed, followed by thorough manual stirring and even distribution into cake cases, which are filled almost to the top. The cakes were baked in the centre of a preheated conventional oven (Macpan MDBt8, Thiene, Italy) at 200°C (180°C fan) for 20–25 minutes or until golden brown and afterwards allowed to cool to room temperature. The cakes were stored in PP (Polypropylene) containers.
2.9. Cake Analysis
The cake samples were analyzed in triplicate over three consecutive days (Days 0, 1, and 2 post baking).
2.9.1. Baking loss (BL %)
The weight loss during baking was calculated on day 0 as the difference in the weight of the batter relative to the weight of the prepared cake by using the following equation (10) [40]:
BL % = [(WBatter – WCake) / (WBatter)] × 100,
2.9.2. Crust and Crumb Colour
The L*, a* and b* values of the crust and crumb of the cakes were determined using a Chroma meter CR-410 (Konica Minolta, UK), which had been calibrated using a standardized white tile [40]. Three readings were recorded on the surface (on the top region of the cake), and three readings on the crumb colour of the cake. The ΔE values indicate the perceptibility of color differences: a value less than 1 means the difference is not easily visible, between 1 and 3 means it’s not readily noticeable, and above 3 means the difference is apparent [41]. The overall colour difference was calculated as follows (11);
∆E*ab = √(L - LControl)2 + (a - aControl)2 + (b - bControl)2,
2.9.3. Cake Density, Specific Volume and Volume Index
The density of the prepared cake samples was calculated as the ratio between the weight of the cake and its volume (g/cm3) [40]. The volume of the vegan fairy cakes was measured by the rapeseed displacement method [42]. The specific volume of the cake (cm3/g) was determined by measuring the cake volume (cm3) divided by the mass (g) [40]. The volume index of cakes was determined following the AACC method 10–91 [42]. Cakes were vertically sliced through their centres, and the heights of the samples were measured at three designated points (B, C, D) across the cross-sections using a template. The volume index was then calculated using the following equation (12):
Volume index = B + C + D,
Where C is the height of the cake at the centre, and B and D are the heights of the cake sample at the points 2.5 cm away from the centre towards the left and right sides of the cake.
2.9.4. Moisture Content, Water Activity, and pH
Crumb moisture was undertaken according to the AACC 44-15A method on days 0, 1 and 2 post-baking. The crust was removed from the central two slices of the cake. The sample was cut into approximately 1 cm3 cubes and allowed to dry in a Brabender oven (Mollelec Ltd., UK) at 130 °C for 60 minutes [40]. The moisture content was calculated using the formula (13), where W2 is the weight of the sample before drying, and W1 is the weight of the sample after drying.
MC % = [(W2 – W1) / (W1)] x 100,
For water activity, a section of the central region of the two centre slices was crumbed, and the water activity of the cakes was determined using an Aqua Lab Lite (Decagon Devices, WA, USA), [43].
To measure the pH of the cake sample, fifteen grams of small pieces of cake were mixed with 150 mL of distilled water and vortexed for 3 min for cake pH measurements. The mixture was held at room temperature for 30 min to separate solid and liquid phases. The pH of the supernatant was measured using the pH meter mentioned above [44].
2.9.5. Sensory Evaluation
The sensory evaluation of the developed cakes was conducted in terms of their appearance, colour, odour and texture. The samples were scored using the scoresheet described below (Table 3).
3. Results
3.1. Extraction Yield and Proximate Composition of Protein Hydrolysates, Spent Biomass and Whole Biomass Fractions of Chondrus crispus
The objective of this study was to evaluate the feasibility of enzyme-assisted hydrolysis for extracting proteins and peptides from red seaweeds in substantial quantities. This exploration aims to broaden the potential utilisation of seaweeds in food products and as functional ingredients. The amount of protein hydrolysates (supernatant) obtained from 50 grams of dried/milled Chondrus crispus following independent treatment with viscozyme and alcalase was 9.71 ± 0.23 g, respectively, (Table 4). The percentage yield of the protein hydrolysates obtained was 19.42 ± 0.46 % from the whole seaweed when measured on a dry weight basis.
The proximate compositions of Chondrus crispus whole biomass, spent biomass, and recovered protein hydrolysates are outlined in Figure 1. The recovered hydrolysate from Chondrus crispus exhibited a protein content of 8.65%, whereas the protein content for the spent and whole biomass fractions measured 8.47% and 9.67%, respectively. The lipid content of the Chondrus crispus hydrolysate was 1%, and the moisture content was 11%. Additionally, the ash content of the hydrolysate was 15.8%.
3.2. Colour of Protein Hydrolysates and Whole Biomass Fractions of Chondrus crispus
The colour assessment of protein hydrolysates derived from Chondrus crispus, as well as the whole biomasses, was conducted by evaluating the L*, a*, and b* values. L* signifies the extent of colour lightness, where a value of 100 indicates white, and 0 denotes blackness. Redness is indicated by +a*, while −a* values indicate a greenish hue. Yellowness is denoted by +b*, whereas −b* signifies blueness [45]. The Chroma value (C*) signifies the degree of deviation from grey towards a pure chromatic colour, while the hue value (h°) distinguishes one colour from another, such as red, yellow, green, blue, or purple, based on their quality. The colour values of Chondrus crispus protein hydrolysates were determined as L* 68.35 ± 0.25, a* 4.47 ± 0.05, b* 11.60 ± 0.10, C* 12.14 ± 0.11 and h° 69.23 ± 0.17 (Table 5). The ∆E* ab value denotes the difference in the colour of the hydrolysates with respect to the whole biomass and a value of 18.34 indicates that there was a significant difference between the colour perceptual to the human eyes.
3.3. Water Activity and pH of Chondrus crispus Protein Hydrolysates
The aW of the freeze-dried hydrolysates, measured at 8.17 ± 0.07°C, was recorded as 0.45 ± 0.005, while the pH of hydrolysate dispersion was determined to be 6.82 ± 0.01 (Table 6).
3.4. Water (WHC) and Oil Holding Capacity (OHC)
The WHC of the protein hydrolysates from Chondrus crispus was determined as 10.14 ± 0.221 g water/g protein hydrolysate. The oil holding capacity (OHC) of Chondrus crispus protein hydrolysates was evaluated with sunflower, peanut, and rapeseed oils, yielding values of 9.226 ± 0.084, 9.175 ± 0.135, and 9.169 ± 0.137g of oil per gram of protein hydrolysates, respectively (Table 7).
3.5. Emulsifying Activity (EA) and Emulsion Stability (ES)
The EA and ES of the protein hydrolysates was assessed with respect to sunflower, peanut and rapeseed oils against a pH range of 2 to 10, (Figure 2(a&b)). The EA ranged from 85.28 ± 1.37% (pH 2) to 5.23 ± 1.31% (pH 10) for sunflower oil, 85.64 ± 1.34% (pH 2) to 13.15 ± 1.82% (pH 10) for peanut oil and 94.33 ± 0.91% (pH 2) to 13.91 ± 0.98% (pH 10) for rapeseed oil.
The ES for the hydrolysates ranged from 72.43 ± 1.28% (pH 2) to 50% (pH 10) for sunflower oil, 73.25 ± 1.01% (pH 2) to 50 (pH 10) for peanut oil, and 59.04 ± 0.85% (pH 2) to 44.44 ± 4.54% (pH 10) for rapeseed oil.
3.6. Foaming Capacity (FC) and Stability (FS)
The foaming properties of the seaweed protein hydrolysates were comprehensively evaluated across a pH spectrum of 2 to 10, as outlined in Figure 3 (a & b). The FC demonstrated a dynamic pH-dependent profile, peaking at 20.20 ± 2.88% at pH 10 and reaching 15.55 ± 1.81% at pH 2, with intermediate values of 10.26 ± 2.09%, 12.12 ± 2.47%, and 9.11% at pH 4, 6, and 8, respectively. Similarly, the FS exhibited significant variation, maintaining high stability with 89.32 ± 1.19% at pH 6, followed by 88.45 ± 0.18% at pH 2, 88.89 ± 2.27% at pH 8, 83.33 ± 1.94% at pH 10, and 79.21 ± 2.93% at pH 4.
The techno-functional properties of Chondrus crispus protein hydrolysates suggest that this novel ingredient could be suitable for use in the formulation of baked products. So, the generated protein hydrolysates were used as an egg replacer (emulsifier) for the preparation of vegan fairy cakes in 3.8.
3.7. Predicted Bioactivities of C. crispus Hydrolysate Peptides
The bioactivities of identified C. crispus hydrolysate peptides with sequence homology greater than 85% with the parent protein were predicted using in silico analysis and results are shown in Table 8. Two peptide sequences, GGPAGGGPAGGDAGLA and AQVPPLPNMPRMPA derived from the proteins Transitional endoplasmic reticulum ATPase (Valosin-containing protein), (accession number R7QAA8_CHOCR) and Prohibitin (accession number R7QRM0_CHOCR) were predicted to be antimicrobial with Peptide Ranker (scores of 0.87 for both peptides), anti-inflammatory (scores of 0.480 and 0.478 respectively for GGPAGGGPAGGDAGLA and AQVPPLPNMPRMPA) with PreAIP and anti-diabetic with AntiDMPpred and a threshold probability of 0.6. In addition, both peptides were identified as having potential to cause Umami flavour. The peptide GIPDEWMGL having sequence homology with the protein Phosphoglycerate kinase (accession number Q8GVE6_CHOCR) was predicted to be very bioactive (0.9) using Peptide Ranker and has predicted anti-inflammatory activity (0.469 value determined for the peptide using PreAIP) but it was not identified as having potential to inhibit enzymes important in the development of T2D. Another peptide, with the sequence VLPSLPFM, having sequence homology with the protein Animal heme peroxidase homologue (accession number R7QJ77_CHOCR), is predicted to be highly bioactive with Peptide Ranker (0.89)and anti-inflammatory (value of 0.478 with PreAIP) but also was not predicted to have T2D prevention qualities. Both peptides are predicted to cause Umami flavour however. The predicted bioactivities determined herein for these peptides, suggest that these peptides are ideal candidates for further study in vitro and in vivo as potential health beneficial ingredients/compounds but chemical synthesis and confirmation of predicted bioactivities is required.
3.8. Baking Loss and Colour of the Developed Vegan Cakes
The baking loss for the control sample was 8.92 ± 0.63%, whereas the vegan fairy cake incorporating protein hydrolysates exhibited a higher baking loss of 11.72 ± 0.13% (Table 8). In terms of crust color, the control sample had L*, a*, and b* values of 49.50 ± 0.33, 17.52 ± 0.15, and 37.54 ± 0.22, respectively. Its crumb displayed L*, a*, and b* values of 68.02 ± 0.42, 2.70 ± 0.09, and 38.03 ± 0.21, with a total color difference (∆E*ab) of 10.16. In contrast, the vegan fairy cake formulated with seaweed protein hydrolysate had a crust with L*, a*, and b* values of 56.23 ± 0.22, 10.03 ± 0.26, and 36.17 ± 0.36. The crumb of this sample showed L*, a*, and b* values of 59.93 ± 0.37, 3.25 ± 0.06, and 29.71 ± 0.30, respectively, with a ∆E*ab of 11.61.
3.9. Density, Specific Volume and Volume Index of the Developed Vegan Cakes
Table 9 outlines relevant data pertaining to cake density, specific volume, and volume index of the developed vegan cakes in comparison to the egg containing contraol cake samples. The control cake had a density of 0.51 ± 0.01 g/cm³, a specific volume of 1.95 ± 0.02 cm³/g, and a volume index of 9.66 ± 0.13 cm. In comparison, the cake with seaweed protein hydrolysates exhibited a density of 0.61 ± 0.02 g/cm³, a specific volume of 1.66 ± 0.04 cm³/g, and a volume index of 9.21 ± 0.18 cm.
3.10. Moisture Content, Water Activity and pH of the Developed Vegan Cakes
Cake samples were analysed in triplicate for moisture content, water activity, and pH over three consecutive days post-baking and the obtained results are outlined in Table 10. On day 0, the moisture content of the control cake was 26.03 ± 0.41%, while for the protein hydrolysate-containing vegan cake was 29.34 ± 0.18%. The water activity (aw) values were 0.810 ± 0.001 for the control and 0.754 ± 0.004 for the vegan cake, with pH values of 7.84 ± 0.01 and 7.25 ± 0.00, respectively. On day 1, the moisture content decreased to 24.63 ± 0.60% in the control and 28.15 ± 0.13% in the vegan cake. The water activity values were 0.817 ± 0.003 for the control and 0.759 ± 0.005 for the vegan cake, while the pH values were 7.85 ± 0.02 and 7.30 ± 0.01, respectively. By day 2, the moisture content further declined to 24.06 ± 0.08% in the control and 27.20 ± 0.07% in the vegan cake. The water activity values reached 0.822 ± 0.003 for the control and 0.762 ± 0.005 for the vegan cake, while the pH values were 7.89 ± 0.00 and 7.31 ± 0.01, respectively.
3.11. Sensory evaluation
Sensory evaluation results (appearance, colour, odour and texture) of the vegan cakes compared with control cakes are summarised in Table 11 and Figure 4 (a & b). The sensory evaluation of cakes revealed that the vegan cake samples scored acceptable scores across all attributes. The appearance, color, odor, and texture of the control cakes were rated at 9.33 ± 0.22, 9.78 ± 0.14, 10.00 ± 0.00, and 9.67 ± 0.16, respectively. In comparison, the protein hydrolysate-containing vegan cakes received comparable scores, with appearance rated at 8.67 ± 0.22, color at 8.56 ± 0.23, odor at 7.56 ± 0.17, and texture at 7.67 ± 0.16.
4. Discussion
4.1. Outcome of Extraction Yield and Proximate Composition of Protein Hydrolysates, Spent Biomass and Whole Biomass Fractions of Chondrus crispus
The protein hydrolysate yield from Chondrus crispus in our study significantly exceeded that reported by [46]. Their work, which involved generating hydrolysates from Palmaria palmata using Chymotrypsin and a Tris–HCl buffer at pH 8 (30 °C for 24 hours), achieved an average yield of just 12.50% on a dry weight basis. In terms of protein content, our findings align closely with a previou [47], where a crude protein content of 13.94 ± 0.94% for Chondrus crispus hydrolysates was repored. The efficiency of protein extraction, however, is influenced by extraction method, harvesting location, prevailing weather conditions, and the maturity stage of the macroalgae [48].
4.2. Colour of Protein Hydrolysates and Whole Biomass Fractions of Chondrus crispus
To the best of our knowledge, no studies have explored the color attributes of Chondrus crispus protein hydrolysates. Interestingly, our findings reveal that the hydrolysates exhibited a perceptually distinct color compared to the whole biomass, with significantly higher values across all measured color attributes. This enhancement in lightness, yellowness, purity, and overall color distinction suggests that hydrolysis plays a crucial role in intensifying the visual characteristics of the resulting hydrolysates.
Color is a key factor in food appearance, directly influencing consumer acceptance [49]. The striking color features observed in Chondrus crispus hydrolysates can be attributed to its unique light-absorbing pigments, which are bound to proteins and function in solar energy capture, leading to the formation of phycobiliproteins (PBPs) [50]. These PBPs—including allophycocyanin (APC), phycocyanin (PC), and phycoerythrin (PE)—are widely present in cyanobacteria and red seaweeds [51,52,53]. Beyond their role in pigmentation, PBPs exhibit bioactive properties, such as anti-inflammatory, hepatoprotective, and antioxidant effects. Additionally, they have shown significant potential in photodynamic therapy for conditions like tumors and leukemia [54,55].
Assessing the color of food ingredients is essential for determining their compatibility in formulations, ensuring minimal impact on the final product's visual appeal. In this study, enzymatic hydrolysis not only enhanced the lightness and yellowness of the hydrolysates but also improved their potential for seamless integration into various food applications, making them a promising functional ingredient with both aesthetic and nutritional benefits.
4.3. Water activity and pH of Chondrus crispus protein hydrolysates
The pH and water activity of the hydrolysates are critical physicochemical properties that determine their stability during storage before further utilisation by the industry and subsequent applications. Higher aW levels in foods accelerate degradation due to biochemical and microbial changes, as water activity directly influences enzymatic and non-enzymatic reactions, microbial spoilage, and lipid oxidation [56]. Maintaining optimal aW is therefore essential for preserving the quality and shelf life of hydrolysates, ensuring their suitability for various food formulations.
4.4. Water (WHC) and Oil Holding Capacity (OHC)
Interactions between proteins, water, and oil are fundamental to food formulation and product stability [57]. The capacity of proteins to bind and retain water and oil plays a crucial role in shaping the texture, consistency, and sensory characteristics of food products [58]. WHC and OHC are dictated by both intrinsic factors—such as amino acid composition, protein conformation, surface polarity, and secondary structure (β-sheet or α-helix)—and extrinsic factors including enzyme type, hydrolysis conditions, and degree of hydrolysis [59].
High WHC is particularly beneficial in baked goods, where it delays retrogradation, preserves moisture, and enhances texture. Conversely, elevated OHC is essential in stabilizing emulsified products like cakes, mayonnaise, and salad dressings, preventing phase separation and improving flavor retention [60]. Proteins with superior WHC and OHC are highly desirable for bakery applications, meat formulations (as emulsifiers and partial meat substitutes), and other processed foods [57].
The WHC values obtained in this study were notably higher than those reported for Chlorella protein hydrolysates prepared with different proteases, which ranged between 1.7 and 5.1 g of water/g of protein hydrolysate [61]. Similarly, Spirulina platensis hydrolysates exhibited WHC values between 1.2 and 5.4 g of water/g of hydrolysate [62]. According to Samsudin et al., enzymatic hydrolysis unfolds the protein structure in a controlled manner, exposing more polar groups (–COOH, –NH₃, and –OH), thereby enhancing interactions with water molecules and increasing WHC [63]. In baked goods, this property is advantageous as it retains moisture without protein dissolution, contributing to improved viscosity, dough thickening, and reduced moisture loss in packaged products [64,65].
The ability of proteins to bind oil is affected by multiple physical and chemical factors, including the oil’s viscosity, molecular weight, degree of saturation, polarity, and external conditions like temperature and pressure [66]. The OHC values of Chlorella protein hydrolysates produced using different proteases ranged from 4.73 to 5.7 g of oil/g of hydrolysate [61] while Spirulina platensis hydrolysates demonstrated OHC values between 5.3 and 7 g of oil/g of hydrolysate, with the highest OHC observed in alcalase-treated samples [62]. Similarly, Xu et al. [67] highlighted the superior OHC of legume protein hydrolysates treated with alcalase and bromelain, attributing this to increased exposure of hydrophobic amino acids. Nonpolar side chains, particularly aromatic and aliphatic groups, enhance fat absorption capacity through hydrophobic interactions [68].
The ability of proteins to retain oil is crucial for flavor retention and improving overall product palatability [69]. Given their high WHC and OHC, protein hydrolysates can serve as multifunctional ingredients in diverse food applications, enhancing both the structural and sensory attributes of formulated products.
4.5. Emulsifying Activity (EA) and Emulsion Stability (ES)
One of the key functional properties of proteins, particularly in food formulations, is their emulsification activity, which stems from their amphiphilic nature. This property is especially valuable in food systems such as sauces, baked goods, and spreads. Enzymatic hydrolysis enhances emulsification by exposing hydrophobic groups within protein molecules, optimizing their ability to interact with oil and water phases. Striking the right balance between the hydrophilic and hydrophobic regions of protein molecules is crucial for optimizing the efficiency of emulsification.
The emulsification process unfolds in two distinct stages: first, the breakdown and deformation of lipid droplets determine the emulsion’s surface area; second, surface-active agents position themselves at the newly formed interface to stabilize the emulsion [70]. Protein adsorption at the oil-water interface follows a diffusion-controlled mechanism, which is more effective at lower protein concentrations. However, higher protein concentrations may activate an energy barrier, hindering diffusion and subsequently reducing emulsifying activity [71,72,73]. Understanding emulsification properties, including emulsifying activity (EA) and emulsion stability (ES), is crucial for expanding the application of Chondrus crispus protein hydrolysates as natural emulsifiers in the food industry, offering a promising alternative to conventional chemical emulsifiers.
The emulsification activity of Chondrus crispus protein hydrolysates was evaluated using sunflower, peanut, and rapeseed oils (Figure 2a). The results revealed significant differences in EA among the tested oils and pH levels. Notably, all hydrolysates exhibited the highest emulsification activity at pH 2, with rapeseed oil showing the greatest EA (94.33%), followed by peanut (85.64%) and sunflower oil (85.27%). Significant variations were observed at pH 4, where EA values for sunflower, peanut, and rapeseed oils were 77.36%, 66.27%, and 56.41%, respectively. The lowest EA for all oils was recorded at pH 10, ranging from 5.23% to 13.91%. These findings are in agreement with the study by Liu et al. [74] who reported an EA of 20.11% for Chlorella pyrenoidosa protein hydrolysates in Antarctic krill oil at a native pH of 6.6. Similarly, Mohammadi et al. [75] examined Spirulina platensis protein hydrolysates and documented an EA of 20% for pepsin hydrolysates and 25% for pancreatin hydrolysates at pH 6. The observed differences in EA among Chondrus crispus hydrolysates are likely due to alterations in the hydrophobic-hydrophilic balance of amino acid constituents at varying pH levels [76].
Emulsion stability (ES) was assessed across the same pH range for sunflower, peanut, and rapeseed oils (Figure 2b). Results indicated that ES was significantly influenced by both oil type and pH. Overall, sunflower and peanut oils exhibited comparable and effective ES across all tested pH levels, with emulsions showing higher stability between pH 2 and 6. Similar trends were reported by Mohammadi et al. [75]who observed emulsion stability indices of 75% for pepsin hydrolysates and 25% for pancreatin hydrolysates at pH 6 in Spirulina platensis. The stability of emulsions is influenced by several key factors, such as pH, droplet size, net charge, interfacial tension, viscosity, and the conformation of the proteins involved [77]. In this study, the remarkable stability of emulsions after heating at 85°C for 15 minutes may be attributed to the dissociation of specific proteins, which formed subunits with greater lipophilic properties. This enhanced interaction with the lipid phase, contributing to the improved thermal stability of the emulsions [78].
These findings highlight the potential of Chondrus crispus protein hydrolysates as effective natural emulsifiers, capable of maintaining stable emulsions across a range of food applications. Their ability to enhance emulsification properties without the need for synthetic additives positions them as a valuable ingredient in the development of clean-label, functional food products.
4.6. Foaming Capacity (FC) and Foam Stability (FS)
In the case of Chondrus crispus protein hydrolysates, the foam capacity (FC) was significantly influenced by the pH of the suspension. The highest FC was observed at pH 10 (20.21 ± 2.88%), while the lowest was found at pH 8 (9.09%). Notably, FC at pH values of 2, 4, and 6 showed no significant statistical differences, with values of 15.55 ± 1.81%, 10.25 ± 2.09%, and 12.12 ± 2.47%, respectively (Figure 3(a)). The decreased FC at pH 8 can likely be attributed to the neutral charge on proteins at their isoelectric point [79]. A study by Kandasamy et al. examined the salt and pH-induced changes in the foaming properties of protein concentrates from the green seaweed Enteromorpha species, reporting FC values ranging from 6.4 ± 1.0% to 55.0 ± 2.6% from pH 2 to 12 at zero ionic strength [80]. They proposed that the higher FC observed at alkaline pH could be due to an increase in net protein charges, which weaken hydrophobic interactions and increase protein flexibility, allowing faster diffusion to the air-water interface and improving foam formation [81].
Foam stability (FS), defined as the volume of foam remaining after a set time expressed as a percentage of the initial foam volume, varied significantly across different pH conditions for Chondrus crispus protein hydrolysates. Foam stability showed an upward trend at alkaline pH levels of 6 (89.32 ± 1.92%) and 8 (88.88 ± 2.26%) (Figure 3(b)). This could be attributed to increased charge density, which reduces the coalescence of air bubbles. The enhanced charge density likely stabilized the foam by amplifying electrostatic repulsion, thereby slowing down bubble coalescence [81]. The foam stability observed in this study was notably higher compared to protein concentrates extracted from three Enteromorpha species (E. compressa, E. linza, and E. tubulosa), which had FS values ranging from 5.0 to 37.5%, 4.4 to 39.0%, and 11.7 to 29.2%, respectively, across a pH range from 2 to 12 with zero salt concentration. In foams, two critical factors that contribute to stability are the ability of the protein film to retain water around the air bubbles and the presence of electrostatic repulsion [82].
4.7. Predicted Bioactivities
In total, over 2000 peptides were identified from the C. crispus hydrolysate and Table 8 lists those predicted to be most bioactive using in silico analysis. The peptide AQVPPLPNMPRMPA derived from the protein Prohibitin is predicted to be anti-inflammatory, anti-diabetic and to cause Umami (Table 8). Prohibitin has been described in animals, unicellular eukaryotes, plants and fungi. Prohibitins are positive regulators of cell proliferation in plants. The peptide AQVPPLPNMPRMPA was not found in the literature or BIOPEP-UWM and warrant further research base on predicted bioactivities. GGPAGGGPAGGDAGLA, the peptide corresponding to amino acid sequences of a valosin protein also was predicted to have anti-inflammatory, anti-diabetic and umami benefits. The peptide GIPDEWMGL derived from a Phosphoglycerate kinase protein was also predicted to have anti-inflammatory and umami activities. These peptides warrant further research including chemical synthesis and confirmation of observed bioactivities in vitro but this work is beyond the scope of this paper. All peptides predicted to be anti-inflammtory using PredAIP contain known tripeptides within their sequences. Peptide AQVPPLPNMPRMPA contains the well known anti-hypertensive peptide VPP at position 3-5 [83]. For example, the peptide sequence GLA, found at the C-terminal end of peptide GGPAGGGPAGGDAGLA, predicted to have anti-inflammatory benefits, has sequence homology with the peptide RLARAGLAQ (BIOPEP-UWM entry number 9812), identified by Zlotek and colleagues as having Cyclooxygenase 1 and 2 inhibitory activities, linked to prevention of pain and inflammation [84].
4.8. Baking Loss and Colour of the Developed Vegan Cakes
Chondrus crispus protein hydrolysates were utilized as an egg replacer in the preparation of vegan fairy cakes, and their performance was evaluated alongside control cakes over three consecutive days after baking. One critical factor in cake quality is the baking loss percentage, which represents the weight lost during baking, mainly due to moisture evaporation. This loss is significant for several reasons: it directly influences the cake's texture and moisture content—too much loss can result in a dry, crumbly texture, while too little can lead to a dense, overly moist cake. Additionally, baking loss affects the cake’s volume and density, ensuring proper rise and crumb structure. Moreover, the level of baking loss can impact the shelf life of the cake; excessive loss may lead to faster staling, while insufficient loss could result in a shorter shelf life due to excess moisture. Therefore, understanding and managing baking loss is essential to producing cakes with desirable characteristics and maintaining freshness.
As shown in Table 8, the vegan fairy cake made with Chondrus crispus protein hydrolysates as an emulsifier experienced a baking loss of 11.72 ± 0.13%. In contrast, the control cake, which used egg as the conventional emulsifier, had a lower baking loss of 8.92 ± 0.63%. Silva et al. (2022) explored an egg-less, gluten-free cake made using aquafaba powder, lentil protein, and citric acid as egg replacers, reporting a baking loss range of 12.27% to 15.20% [85]. The control cake with egg as an emulsifier had a baking loss of 10.40 ± 0.80%. A similar comparison was made by Aslan and Ertaş with chickpea aquafaba as an egg replacer, where the baking loss for the egg-less formulation was 13.74%, while the egg-based control had a baking loss of 12.72% [86].
In addition to texture and moisture, the hue of the cake crust and crumb is also important. The color changes in the crust during baking are primarily due to the Maillard reaction and caramelization, while the crumb color is influenced by the specific ingredients used [87]. The crust color of the vegan cakes appeared darker compared to the control cakes. The crumb color of the vegan cakes exhibited a slightly redder tone, which could be attributed to the presence of phycobiliproteins (PBPs) and other phenolic compounds in the hydrolysates. Overall, the color parameters of the eggless cakes were considered highly desirable.
In a similar study, Jarpa-Parra and colleagues evaluated the quality of angel food cakes and muffins developed with lentil protein as an egg/milk replacer and found comparable results [88]. The inclusion of lentil protein led to a significant decrease in the L*, a*, and b* values of the cakes, as well as a notable increase in ΔEab values, which were 18.9 for the crust and 21.2 for the crumb. Mohammadi et al. (2022) also investigated the use of enzymatically generated lentil protein hydrolysates in functional bakery goods and observed a gradual decrease in the L values in the developed muffins [89]. Gani et al. (2015) reported similar findings when enriching cookies with casein protein hydrolysates, which significantly altered the color characteristics, particularly due to the Maillard reaction [90]. Similarly, Majzoobi and colleagues found that incorporating soy protein isolate into sponge cake formulations led to a reduction in the lightness of the crust, which they attributed to reactions between free amino groups and reducing sugars during baking [87].
4.9. Density, Specific Volume and Volume Index of the Developed Vegan Cakes
Density and volume are key quality parameters for consumers when assessing cakes, particularly in eggless varieties. The results indicated that vegan fairy cakes made with seaweed protein hydrolysates exhibited a texture similar to the control cakes, with a slight increase in density, a minor reduction in specific volume, and no significant change in the volume index. This suggests that the inclusion of protein hydrolysates preserved the cake's overall structure, while contributing to a slightly more compact texture. This makes it a promising alternative to traditional formulations, offering a balance between texture and structure in egg-free cakes.
In line with these findings, Anwar et al. (2020) reported similar outcomes in their study on wholemeal chia flour in eggless cakes [91]. They noted a slight increase in cake density and a reduction in specific volume, reinforcing the idea that plant-based protein alternatives can influence cake texture without compromising its overall structure. Similarly, Mohammadi and colleagues observed a 10% decrease in the specific volume of muffins made with lentil protein hydrolysates [89].
During the baking process, the production of steam and CO2 from the leavening agent plays a crucial role in forming air cells that contribute to the cake's spongy texture and low density [92]. The protein hydrolysates in the vegan cakes exhibited a remarkable ability to retain moisture, highlighting their capacity to effectively absorb and hold onto water molecules. While this water retention is beneficial for the cake's moisture content, it may also reduce the water available for steam production during baking, potentially limiting the formation of air cells and contributing to the slight reduction in specific volume.
On the other hand, Lin et al. (2017b) found a significant decrease in the specific volume of eggless cakes made with isolated pea proteins [93]. They attributed this reduction to insufficient aeration during mixing, which hindered the expansion of the batter during baking. The volume index of a cake, which reflects its overall size and internal structure, was comparable between the control and the vegan cakes. A higher volume index typically signals a lighter, more aerated cake, while a lower value suggests a denser, more compact product. In this study, the volume index of the vegan cakes was on par with the control cakes, indicating that the use of protein hydrolysates did not negatively impact the cake's rise.
Anwar et al. (2020) observed similar trends when they used wholemeal chia flour in eggless cakes, noting a slightly denser structure in the flour-based cakes compared to their egg-containing counterparts [91]. These results suggest that plant-based protein alternatives can successfully maintain a cake's volume while subtly affecting its texture. This makes protein hydrolysates a viable and innovative option for creating high-quality, egg-free baked goods without compromising the desired characteristics of traditional cakes.
4.10. Moisture Content, Water Activity and pH of the Developed Vegan Cakes
The cake samples were analyzed in triplicate for moisture content, water activity, and pH over three consecutive days following baking. On day 0, the moisture content of the vegan cakes was found to be 29.34 ± 0.18%, notably higher than the control, which measured 26.03 ± 0.41%. Preserving the freshness and sensory appeal of baked goods relies on reducing the factors that contribute to degradation during storage, particularly retrogradation. This process involves the gradual movement, redistribution, and eventual loss of water within the cake matrix [94,95]. The vegan cakes maintained a higher moisture content, likely due to the superior moisture retention capabilities of the protein hydrolysates used. Similar trends were noted in studies by Lin et al. (2017a), who observed a significant decrease in moisture content in soy protein isolate and polysaccharide-based eggless cakes compared to control samples [40]. Likewise, isolated pea proteins also led to reduced moisture retention in eggless formulations [93].
Water activity (aw), a critical determinant of food shelf life, strongly influences microbial growth and overall product stability [96]. Consistently, the control cakes exhibited a higher water activity than the vegan cakes over the three days. This difference can be attributed to the high water-holding capacity of the protein hydrolysates, which limits the availability of free water. Comparable results were observed in a study by Anwar et al. (2020), where the water activity of a control cake was measured at 0.808 ± 0.007, whereas the eggless version made with wholemeal chia flour showed a reduced water activity of 0.766 ± 0.007 [91].
The pH level of a cake plays a significant role in influencing various chemical reactions that occur during baking, such as leavening, browning, and flavor development. It is a critical factor for ensuring the desired texture, taste, and overall quality of the final product. The pH of the developed vegan cakes was found to be slightly lower than that of the control cakes, which could have impacted the flavor profile and the chemical environment of the cake. Typically, the pH of cakes falls within a neutral to slightly alkaline range, which is ideal for achieving optimal flavor, texture, and quality.
4.11. Sensory Evaluation
The sensory evaluation of the vegan cakes, compared with the control cakes, focused on aspects such as appearance, colour, odour, and texture, as summarized in Table 11. Overall, the vegan cakes were found to be very similar to the control cakes across all sensory attributes. The appearance of the vegan cakes was comparable, with a neat and smooth surface, free from wrinkles, cracks, or collapses. The surface colour was a pleasant yellow, and while the crumb colour was slightly darker than the control, it remained consistent without any noticeable colour variations.
Regarding odour, the vegan cakes emitted pleasant fragrance, without any undesirable or off-putting smells. The texture of the vegan cakes was also well-received, with a soft and elastic crumb that exhibited a honeycomb-like structure, featuring some larger air pockets that contributed to a pleasing mouthfeel. These results are consistent with findings by Gani et al. (2015), who investigated the sensory properties of cookies fortified with whey and casein hydrolysates [90]. They noted that protein hydrolysates could sometimes lead to a decrease in sensory scores due to the undesirable odours associated with them. Similarly, Anwar and colleagues found that eggless cakes containing wholemeal chia flour were met with negative reactions due to an unpleasant odour [91].
Author Contributions
Conceptualization, M.H. and D.B.; methodology, D.B, M.H., L.M..; software, L.M..; , M. H. validation, M.H., L.M. and D.B.; formal analysis, D.B.; investigation, M.H.; resources, M.H.; data curation, M.H.; writing—original draft preparation, D.B.; writing—review and editing, M.H.; visualization, M.H.; supervision, M.H.; project administration, M.H.; funding acquisition, M.H. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research was funded by the U-Protein project that received funding from the Irish Department of Agriculture, Food and the Marine, under the Food Institutional Research Measure, grant number 2019PROG702.
Data Availability Statement
Data is available from the corresponding author on request.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| WHC | Water Holding Capacity |
| OHC | Oil Holding Capacity |
| EA | Emulsifying Activity |
| ES | Emulsion Stability |
| FC | Foaming Capacity |
| FS | Foaming Stability |
References
- Ogunwolu, S.O.; Henshaw, F.O.; Mock, H.P.; Santros, A.; Awon Orin, S.O. Functional properties of protein concentrates and isolates produced from cashew (Anacardium occidentale L.) nut. Food Chem. 2009, 115, 852–858. [CrossRef]
- Yun, J.H.; Kim, S.W.; Lee, J.H.; Kim, J.S.; Han, I.K. Comparative efficacy of plant and animal protein sources on the growth performance, nutrient digestibility, morphology, and caecal microbiology of early-weaned pigs. Asian-Australas. J. Anim. Sci. 2005, 18, 1285–1293. [CrossRef]
- Wong, K.H.; Cheung, P.C.; Ang, P.O. Nutritional evaluation of protein concentrates isolated from two red seaweeds: Hypnea charoides and Hypnea japonica in growing rats. Hydrobiologia 2004, 512, 271–278. [CrossRef]
- Slaski, R.J.; Franklin, P.T. A review of the status of the use and potential to use micro and macroalgae as commercially viable raw material sources for aquaculture diets. Report commissioned by Scottish Aquaculture Research Forum (SARF), 94 pp. Available online: http://www.sarf.org.uk/cms-assets/documents/29524-222388.sarf077.pdf (accessed on 20 January 2025).
- Kumar, V.; Kaladharan, P. Amino acids in the seaweeds as an alternate source of protein for animal feed. Food Rev. Int. 1989, 5, 101–104.
- Fleurence, J. Seaweed proteins: biochemical, nutritional aspects, and potential uses. Trends Food Sci. Technol. 1999, 10, 25–28. [CrossRef]
- Lorenzo, J.M.; Agregán, R.; Munekata, P.E.S.; Domínguez, R.; Franco, D.; Simal-Gándara, J. Proximate composition and nutritional value of three macroalgae: Ascophyllum nodosum, Fucus vesiculosus, and Bifurcaria bifurcata. Mar. Drugs 2017, 15, 360. [CrossRef]
- Gamero-Vega, G.; Palacios-Palacios, M.; Quitral, V. Nutritional composition and bioactive compounds of red seaweed: A mini-review. J. Food Nutr. Res. 2020, 8, 431–436. [CrossRef]
- Ito, K.; Hori, K. Seaweed: Chemical composition and potential food uses. Food Rev. Int. 1989, 5, 101–104. [CrossRef]
- Pliego-Cortés, H.; Wijesekara, I.; Lang, M.; Bourgougnon, N.; Bedoux, G. Current knowledge and challenges in extraction, characterisation, and bioactivity of seaweed protein and seaweed-derived proteins. In Advances in Botanical Research, 1st ed.; Lang, M., Bourgougnon, N., Bedoux, G., Eds.; Elsevier: Amsterdam, Netherlands, 2020; Volume 95, pp. 289–326.
- Ohanenye, I.C.; Tsopmo, A.; Ejike, C.E.C.C.; Udenigwe, C.C. Germination as a bioprocess for enhancing the quality and nutritional prospects of legume proteins. Trends Food Sci. Technol. 2020, 101, 213–222. [CrossRef]
- Wong, K.H.; Cheung, P.C.K.; Ang, P.O. Nutritional evaluation of protein concentrates isolated from two red seaweeds: Hypnea charoides and Hypnea japonica in growing rats. In Asian Pacific Phycology in the 21st Century: Prospects and Challenges; Ang, P.O., Ed.; Springer: Dordrecht, Netherlands, 2004; pp. 577–582.
- Ragan, M.A.; Glombitza, K.W. Phlorotannins, Brown Algal Polyphenols. Prog. Phycol. Res. 1986, 4, 129–241.
- Fleurence, J.; Le Coeur, C.; Mabeau, S.; Maurice, M.; Landrein, A. Comparison of different extractive procedures for proteins from the edible seaweeds Ulva rigida and Ulva rotundata. J. Appl. Phycol. 1995, 7, 577–582. [CrossRef]
- Jordan, P.; Vilter, H. Extraction of proteins from material rich in anionic mucilages: Partition and fractionation of vanadate-dependent bromoperoxidases from the brown algae Laminaria digitata and L. saccharina in aqueous polymer two-phase system. Biochim. Biophys. Acta. 1991, 1073, 98–106. [CrossRef]
- Loomis, W.D.; Battaile, J. Plant phenolic compounds and the isolation of plant enzymes. Phytochemistry 1966, 5, 423–438. [CrossRef]
- Ochiai, Y.; Katsuragi, T.; Hashimoto, K. Proteins in three seaweeds: “Aosa” Ulva lactuca, “Arame” Eisenia bicyclis, and “Makusa” Gelidium amansii. Bull. Jpn. Soc. Sci. Fish. 1987, 53, 1051–1055.
- Abeer, M.M.; Trajkovic, S.; Brayden, D.J. Measuring the oral bioavailability of protein hydrolysates derived from food sources: A critical review of current bioassays. Biomed. Pharmacother. 2021, 144, 112275. [CrossRef]
- Echave, J.; Fraga-Corral, M.; García-Oliveira, P.; Carpena, M.; Pereira, A.G.; Xiao, J.; Prieto, M.A.; Simal-Gándara, J. Seaweed Protein Hydrolysates and Bioactive Peptides: Extraction, Purification, and Applications. Mar. Drugs 2021, 19, 500. [CrossRef]
- Mirzapour-Kouhdasht, A.; McClements, D.J.; Taghizadeh, M.S.; Niazi, A.; Garcia-Vaquero, M. Strategies for oral delivery of bioactive peptides with focus on debittering and masking. NPJ Sci. Food 2023, 7, 22. [CrossRef]
- Grossmann, L.; Weiss, J. Alternative Protein Sources as Technofunctional Food Ingredients. Annu. Rev. Food Sci. Technol. 2021, 12, 93–117. [CrossRef]
- Chen, L.C.M.; McLachlan, J. The life history of Chondrus crispus in culture. Can. J. Bot. 1972, 50, 1055–1060. [CrossRef]
- O’Connor, K. Seaweed: A Global History; Reaktion Books: London, UK, 2017.
- Rudtanatip, T.; Lynch, S.A.; Wongprasert, K.; Culloty, S.C. Assessment of the effects of sulfated polysaccharides extracted from the red seaweed Irish moss Chondrus crispus on the immune-stimulant activity in mussels Mytilus spp. Fish Shellfish Immunol. 2018, 75, 284–290. [CrossRef]
- Kulshreshtha, G.; Burlot, A.S.; Marty, C.; Critchley, A.; Hafting, J.; Bedoux, G.; Bourgougnon, N. Enzyme-assisted extraction of bioactive material from Chondrus crispus and Codium fragile and its effect on herpes simplex virus (HSV-1). Mar. Drugs 2015, 13, 558–578. [CrossRef]
- Association of Official Analytical Chemists International (AOAC). Official Methods of Analysis of Association of Official Analytical Chemists International; Association of Official Analytical Chemists: Washington, DC, USA, 1995; 1, p. 870.
- López-Hortas, L., Caleja, C., Pinela, J., Petrović, J., Soković, M., Ferreira, I.C.F.R., To M.D. Domínguez, H., Pereira, E., Barros, L. Comparative evaluation of physicochemical profile and bioactive properties of red edible seaweed Chondrus crispus subjected to different drying methods, Food Chemistry, 383, 2022. [CrossRef]
- Samarakoon, Y.M.; Gunawardena, N. Knowledge and self-reported practices regarding leptospirosis among adolescent school children in a highly endemic rural area in Sri Lanka. Rural Remote Health 2013, 13, 8–19.
- Purcell, D., Packer, M. A., & Hayes, M. Angiotensin-I-converting enzyme inhibitory activity of protein hydrolysates generated from the macroalga Laminaria digitata (Hudson) J.V. Lamouroux 1813. Foods 2022, 11, 1792. [CrossRef]
- Bencini, M. C. Functional properties of drum-dried chickpea (Cicer arietinum L.) flours. J. Food Sci. 1986, 51, 1518–1521. [CrossRef]
- Naczk, M., Diosady, L. L., & Rubin, L. J. Functional properties of canola meals produced by a two-phase solvent extraction system. J. Food Sci. 1985, 50, 1685–1688. [CrossRef]
- Poole, S., West, S. I., & Walters, C. L. Protein–protein interactions: Their importance in the foaming of heterogeneous protein systems. J. Sci. Food Agric. 1984, 35, 701–711. [CrossRef]
- Hayes, M.; Aluko, R.E.; Aurino, E.; Mora, L. Generation of Bioactive Peptides from Porphyridium sp. and Assessment of Their Potential for Use in the Prevention of Hypertension, Inflammation and Pain. Mar. Drugs 2023, 21, 422. [CrossRef]
- Hayes, M.; Naik, A.; Mora, L.; Iñarra, B.; Ibarruri, J.; Bald, C.; Cariou, T.; Reid, D.; Gallagher, M.; Dragøy, R.; et al. Generation, Characterisation and Identification of Bioactive Peptides from Mesopelagic Fish Protein Hydrolysates Using In Silico and In Vitro Approaches. Mar. Drugs 2024, 22, 297. [CrossRef]
- Purcell, D., Packer, M. A., Hayes, M. Identification of Bioactive Peptides from a Laminaria digitata Protein Hydrolysate Using In Silico and In Vitro Methods to Identify Angiotensin-1-Converting Enzyme (ACE-1) Inhibitory Peptides. Mar Drugs. 2023 Jan 27;21(2):90. [CrossRef] [PubMed] [PubMed Central]
- Khatun MS, Hasan MM, Kurata H. PreAIP: Computational Prediction of Anti-inflammatory Peptides by Integrating Multiple Complementary Features. Front Genet. 2019 Mar 5;10:129. [CrossRef] [PubMed] [PubMed Central]
- Qi, L., Du, J., Sun, Y., Xiong, Y., Zhao, X., Pan, D, Zhi, Y, Dang, Y., Gao, X. Umami-MRNN: Deep learning-based prediction of umami peptide using RNN and MLP, Food Chemistry, 405, Part A, 2023. [CrossRef]
- Chen, X., Huang, J., He, B. AntiDMPpred: a web service for identifying anti-diabetic peptides. PeerJ. 2022 Jun 14;10:e13581. [CrossRef] [PubMed] [PubMed Central]
- Syed, S. J., Gadhe, K. S., & Shaikh, R. P. Studies on quality evaluation of oat milk. J. Pharmacogn. Phytochem. 2020, 9, 2275–2277.
- Lin, M., Tay, S. H., Yang, H., Yang, B., & Li, H. Replacement of eggs with soybean protein isolates and polysaccharides to prepare yellow cakes suitable for vegetarians. Food Chem. 2017a, 229, 663–673. [CrossRef]
- Baixauli, R.; Salvador, A.; Fiszman, S. M. Textural and colour changes during storage and sensory shelf life of muffins containing resistant starch. Eur. Food Res. Technol. 2008, 226, 523–530. [CrossRef]
- American Association of Cereal Chemists (AACC). Approved Methods of the AACC. Methods 10–91; AACC: St Paul, MN, USA, 1983.
- Milner, L., Kerry, J. P., O'Sullivan, M. G., & Gallagher, E. Physical, textural, and sensory characteristics of reduced sucrose cakes incorporated with clean-label sugar-replacing alternative ingredients. Innov. Food Sci. Emerg. Technol. 2020, 59, 102235. [CrossRef]
- Shao, Y. Y., Lin, K. H., & Chen, Y. H. Batter and product quality of eggless cakes made of different types of flours and gums. J. Food Process. Preserv. 2015, 39, 2959–2968. [CrossRef]
- Aziah, A. N., & Komathi, C. A. Physicochemical and functional properties of peeled and unpeeled pumpkin flour. J. Food Sci. 2009, 74, S328–S333. [CrossRef]
- Beaulieu, L., Sirois, M., & Tamigneaux, É. Evaluation of the in vitro biological activity of protein hydrolysates of the edible red alga, Palmaria palmata (Dulse), harvested from the Gaspe coast and cultivated in tanks. J. Appl. Phycol. 2016, 28, 3101–3115. [CrossRef]
- Habibie, A., Raharjo, T. J., Swasono, R. D., & Retnaningrum, E. Antibacterial activity of active peptide from marine macroalgae Chondrus crispus protein hydrolysate against Staphylococcus aureus. Pharmacia 2023, 70, 983–992. [CrossRef]
- Vieira, E. F., et al. Seaweeds from the Portuguese coast as a source of proteinaceous material: Total and free amino acid composition profile. Food Chem. 2018, 269, 264–275. [CrossRef]
- Jiménez-Aguilar, D. M.; Ortega-Regules, A. E.; Lozada-Ramírez, J. D.; Pérez-Pérez, M. C. I.; Vernon-Carter, E. J.; Welti-Chanes, J; Color and chemical stability of spray-dried blueberry extract using mesquite gum as wall material. J. Food Compos. Anal. 2011, 24, 889–894. [CrossRef]
- Combet, S.; Pieper, J.; Coneggo, F.; Ambroise, J. P.; Bellissent-Funel, M. C.; Zanotti, J. M.; Coupling of laser excitation and inelastic neutron scattering: Attempt to probe the dynamics of light-induced C-phycocyanin dynamics. Eur. Biophys. J. 2008, 37, 693–700. [CrossRef]
- Jiang, T., Zhang, J. P., Chang, W. R., & Liang, D. C. Crystal structure of R-phycocyanin and possible energy transfer pathways in the phycobilisome. Biophys. J. 2001, 81, 1171–1179. [CrossRef]
- Murray, J. W., Maghlaoui, K., & Barber, J. The structure of allophycocyanin from Thermosynechococcus elongatus at 3.5 Å resolution. Acta Crystallogr. F Struct. Biol. Cryst. Commun. 2007, 63, 998–1002. [CrossRef]
- Shi, F., Qin, S., & Wang, Y. C. The coevolution of phycobilisomes: Molecular structure adapting to functional evolution. Int. J. Genomics 2011, 2011, 1–10. [CrossRef]
- Básaca-Loya, G., Valdez, M., Enríquez-Guevara, E., Gutierrez-Millán, L., & Burboa, M. Extraction and purification of B-phycoerythrin from the red microalga Rhodosorus marinus. Cienc. Mar. 2009, 35, 359–368. [CrossRef]
- Mishra, S. K.; Shrivastav, A.; Maurya, R. R.; Patidar, S. K.; Haldar, S.; Mishra, S. Effect of light quality on the C-phycoerythrin production in marine cyanobacteria Pseudanabaena sp. isolated from Gujarat coast, India. Protein Expr. Purif. 2012, 81, 5–10. [CrossRef]
- Sablani, S., Kasapis, S., & Rahman, M. Evaluating water activity and glass transition concepts for food stability. J. Food Eng. 2007, 78, 266–271. [CrossRef]
- Hemker, A. K., Nguyen, L. T., Karwe, M., & Salvi, D. Effects of pressure-assisted enzymatic hydrolysis on functional and bioactive properties of tilapia (Oreochromis niloticus) by-product protein hydrolysates. LWT 2020, 122, 109003. [CrossRef]
- Suresh Kumar, K., Ganesan, K., Selvaraj, K., & Subba Rao, P. V. Studies on the functional properties of protein concentrate of Kappaphycus alvarezii (Doty) Doty – An edible seaweed. Food Chem. 2014, 153, 353–360. [CrossRef]
- Yu, J., Ahmedna, M., & Goktepe, I. Peanut protein concentrate: Production and functional properties as affected by processing. Food Chem. 2007, 103, 121–129. [CrossRef]
- Ge, J.; Li, C.; Zhang, J.; Wang, Q.; Liu, J. Physicochemical and pH-dependent functional properties of proteins isolated from eight traditional Chinese beans. Food Hydrocoll. 2021, 112, 106288. [CrossRef]
- Gharehbeglou, P.; Sarabandi, K.; Akbarbaglu, Z. Insights into enzymatic hydrolysis: Exploring effects on antioxidant and functional properties of bioactive peptides from chlorella proteins. J. Agric. Food Res. 2024, 10, 101129. [CrossRef]
- Akbarbaglu, Z.; Ayaseh, A.; Ghanbarzadeh, B.; Sarabandi, K. Techno-functional, biological and structural properties of Spirulina platensis peptides from different proteases. Algal Res. 2022, 66, 102755. [CrossRef]
- Samsudin, N. A.; Halim, N. R. A.; Sarbon, N. M. pH levels effect on functional properties of different molecular weight eel (Monopterus sp.) protein hydrolysate. J. Food Sci. Technol. 2018, 55, 4608–4614. [CrossRef]
- Chandi, G. K.; Sogi, D. Functional properties of rice bran protein concentrates. J. Food Eng. 2007, 79, 592–597. [CrossRef]
- Seena, S.; Sridhar, K. Physicochemical, functional and cooking properties of underexplored legumes, Canavalia of the southwest coast of India. Food Res. Int. 2005, 38, 803–814. [CrossRef]
- Chen, C.; Sun-Waterhouse, D.; Zhao, J.; Zhao, M.; Waterhouse, G. I.; Sun, W. Soybean protein isolate hydrolysates-liposomes interactions under oxidation: Mechanistic insights into system stability. Food Hydrocoll. 2021, 112, 106336. [CrossRef]
- Xu, X.; Qiao, Y.; Shi, B.; Dia, V. P. Alcalase and bromelain hydrolysis affected physicochemical and functional properties and biological activities of legume proteins. Food Struct. 2021, 27, 100178. [CrossRef]
- Pires, J. B.; Dos Santos, F. N.; de Lima Costa, I. H.; Kringel, D. H.; da Rosa Zavareze, E.; Dias, A. R. G. Essential oil encapsulation by electrospinning and electrospraying using food proteins: A review. Food Res. Int. 2023, 170, 112970. [CrossRef]
- Kyriakopoulou, K., Keppler, J.K., van der Goot ,A. J. Functionality of Ingredients and Additives in Plant-Based Meat Analogues. Foods. 2021 Mar 12;10(3):600. [CrossRef] [PubMed] [PubMed Central]
- Nalinanon, S.; Benjakul, S.; Kishimura, H.; Shahidi, F. Functionalities and antioxidant properties of protein hydrolysates from the muscle of ornate threadfin bream treated with pepsin from skipjack tuna. Food Chem. 2011, 124, 1354–1362. [CrossRef]
- Wang, Z.; Liu, X.; Xie, H.; Liu, Z.; Rakariyatham, K.; Yu, C.; Shahidi, F.; Zhou, D. Antioxidant activity and functional properties of Alcalase-hydrolyzed scallop protein hydrolysate and its role in the inhibition of cytotoxicity in vitro. Food Chem. 2021, 344, 128566. [CrossRef]
- Lawal, O. S. Functionality of African locust bean (Parkia biglobossa) protein isolate: Effects of pH, ionic strength and various protein concentrations. Food Chem. 2004, 86, 345. [CrossRef]
- Jafari, S. M.; Assadpoor, E.; He, Y.; Bhandari, B. Re-coalescence of emulsion droplets during high-energy emulsification. Food Hydrocoll. 2008, 22, 1191–1202. [CrossRef]
- Liu, Y.; Qi, Y.; Wang, Q.; Yin, F.; Zhan, H.; Wang, H.; Liu, B.; Nakamura, Y.; Wang, J.; Antioxidative effect of Chlorella pyrenoidosa protein hydrolysates and their application in krill oil-in-water emulsions. Mar. Drugs 2022, 20, 345. [CrossRef]
- Mohammadi, M.; Soltanzadeh, M.; Ebrahimi, A. R.; Hamishehkar, H. Spirulina platensis protein hydrolysates: Techno-functional, nutritional and antioxidant properties. Algal Res. 2022, 65, 102739. [CrossRef]
- Sathe, S.; Deshpande, S.; Salunkhe, D. Functional properties of lupin seed (Lupinus mutabilis) proteins and protein concentrates. J. Food Sci. 1982, 47, 491–497. [CrossRef]
- Hung, S.; Zayas, J. Emulsifying capacity and emulsion stability of milk proteins and corn germ protein flour. J. Food Sci. 1991, 56, 1216–1218. [CrossRef]
- Chau, C.-F.; Cheung, P. C.; Wong, Y.-S. Functional properties of protein concentrates from three Chinese indigenous legume seeds. J. Agric. Food Chem. 1997, 45, 2500–2503. [CrossRef]
- Ragab, D. M.; Babiker, E. E.; Eltinay, A. H. Fractionation, solubility and functional properties of cowpea (Vigna unguiculata) proteins as affected by pH and/or salt concentration. Food Chem. 2004, 84, 207–214. [CrossRef]
- Kandasamy, G., Karuppiah, S. K. & Subba Rao, P. V. Salt- and pH-induced functional changes in protein concentrate of edible green seaweed Enteromorpha species. Fish Sci. 2012, 78, 169–176. [CrossRef]
- Aluko, R.; Yada, R. Structure-function relationships of cowpea (Vigna unguiculata) globulin isolate: Influence of pH and NaCl on physicochemical and functional properties. Food Chem. 1995, 53, 259–265. [CrossRef]
- Kinsella, J. E.; Damodaran, S.; German, B. Physicochemical and functional properties of oilseed proteins with emphasis on soy proteins. In New Protein Foods; Alschul, A. M., Wilke, H. L., Eds.; Academic Press: New York, NY, USA, 1985; pp. 107–179.
- Yamaguchi, N., Kawaguchi, K., Yamamoto, N. Study of the mechanism of antihypertensive peptides VPP and IPP in spontaneously hypertensive rats by DNA microarray analysis. European Journal of Pharmacology,2009, 620, 71-77.
- Zlotek, U., Jakubczyk, A., Tkaczyk, K., Cwiek, P., Baraniak, B., Lewicki, S. Characterisation of new peptides GQLGEHGGAGMG, GEHGGAGMGGGQFQPV, EQGFLPGPEESG, RLARAGLAQ, YGNPVGGVG, and GNPVGGVGHGTTGT as inhibitors of enzymes involved in Metabolic Syndrome and Antimicrobial potential. Molecules, 2020, 27, 2492. doi.10.3390/molecules25112492.
- Silva, P. G.; Kalschne, D. L.; Salvati, D.; Bona, E.; Rodrigues, A. C. Aquafaba powder, lentil protein and citric acid as egg replacer in gluten-free cake: A model approach. Appl. Food Res. 2022, 2, 100188. [CrossRef]
- Aslan, M.; Ertaş, N. Possibility of using 'chickpea aquafaba' as egg replacer in traditional cake formulation. Harran Tarım ve Gıda Bilimleri Dergisi 2020, 24, 1–8. [CrossRef]
- Majzoobi, M.; Ghiasi, F.; Habibi, M.; Hedayati, S.; Farahnaky, A. Influence of soy protein isolate on the quality of batter and sponge cake. J. Food Process. Preserv. 2014, 38, 1164–1170. [CrossRef]
- Jarpa-Parra, M.; Bamdad, F.; Tian, Z.; Temelli, F.; Zeng, H.; Shaheen, N.; Chen, L. Quality characteristics of angel food cake and muffin using lentil protein as egg/milk replacer. Int. J. Food Sci. Technol. 2017, 52, 1604–1613. [CrossRef]
- Mohammadi, M.; Salami, M.; Yarmand, M.; Emam-Djomeh, Z.; McClements, D. J. Production and characterization of functional bakery goods enriched with bioactive peptides obtained from enzymatic hydrolysis of lentil protein. J. Food Meas. Charact. 2022, 16, 3402–3409. [CrossRef]
- Gani, A.; Wani, S. M.; Masoodi, F. A.; Hameed, G.; Ashraf, Z. Effect of whey and casein protein hydrolysates on rheological, textural and sensory properties of cookies. J. Food Sci. Technol. 2015, 52, 5718–5726. [CrossRef]
- Anwar, D. A.; Shehita, H. A.; Soliman, H. R. E. S. Development of eggless cake physical, nutritional and sensory attributes for vegetarians by using wholemeal chia (Salvia hispanica L.) flour. Middle East J. Appl. Sci. 2020, 10, 313–329. [CrossRef]
- Rahmati, N. F.; Tehrani, M. M. Influence of different emulsifiers on characteristics of eggless cake containing soymilk: Modelling of physical and sensory properties by mixture experimental design. J. Food Sci. Technol. 2014, 51, 1697–1710. [CrossRef]
- Lin, M.; Tay, S. H.; Yang, H.; Yang, B.; Li, H. Development of eggless cakes suitable for lacto-vegetarians using isolated pea proteins. Food Hydrocoll. 2017b, 69, 440–449. [CrossRef]
- Gray, J. A.; Bemiller, J. N. Bread staling: Molecular basis and control. Compr. Rev. Food Sci. Food Saf. 2003, 2, 1–21. [ 10.1111/j.1541-4337.2003.tb00011.x].
- Wilderjans, E.; Luyts, A.; Brijs, K.; Delcour, J. A. Ingredient functionality in batter type cake making. Trends Food Sci. Technol. 2013, 30, 6–15. [CrossRef]
- Singh, J. P.; Kaur, A.; Singh, N. Development of eggless gluten-free rice muffins utilizing black carrot dietary fibre concentrate and xanthan gum. J. Food Sci. Technol. 2016, 53, 1269–1278. [CrossRef]
Figure 1.
Compositional analysis values obtained for Chondrus crispus whole biomass, spent biomass and supernatant fractions generated following viscozyme and alcalase enzyme hydrolysis and centrifugation steps. (SD, standard deviation, n=3)..
Figure 1.
Compositional analysis values obtained for Chondrus crispus whole biomass, spent biomass and supernatant fractions generated following viscozyme and alcalase enzyme hydrolysis and centrifugation steps. (SD, standard deviation, n=3)..
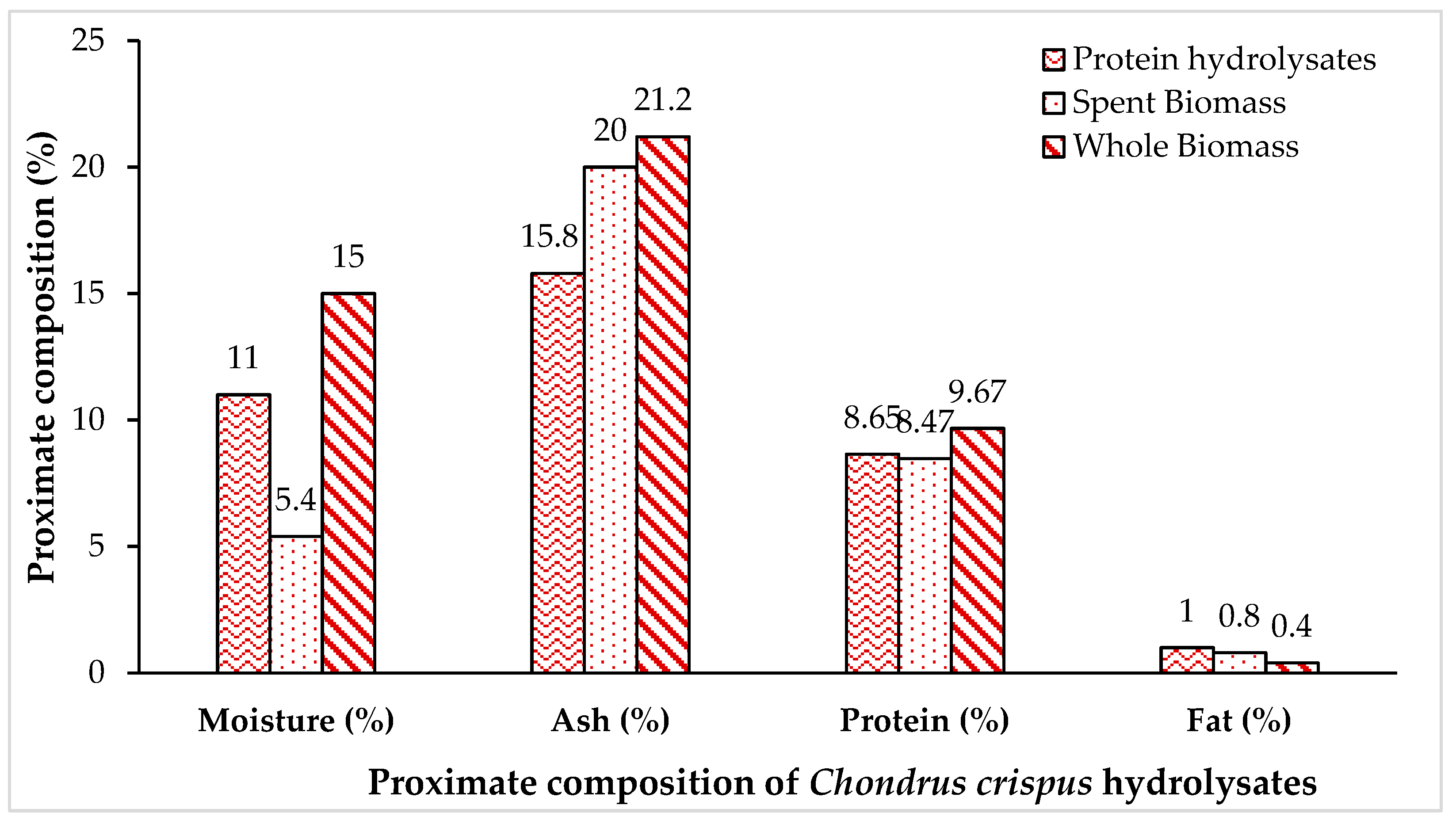
Figure 2.
Emulsion characteristics of Chondrus crispus protein hydrolysates against sunflower, peanut and rapeseed oil. (a) Emulsifying activity of Chondrus crispus protein hydrolysates. (SE, standard error, n=3); (b) Emulsion stability of Chondrus crispus protein hydrolysates. (SE, standard error, n=3).
Figure 2.
Emulsion characteristics of Chondrus crispus protein hydrolysates against sunflower, peanut and rapeseed oil. (a) Emulsifying activity of Chondrus crispus protein hydrolysates. (SE, standard error, n=3); (b) Emulsion stability of Chondrus crispus protein hydrolysates. (SE, standard error, n=3).
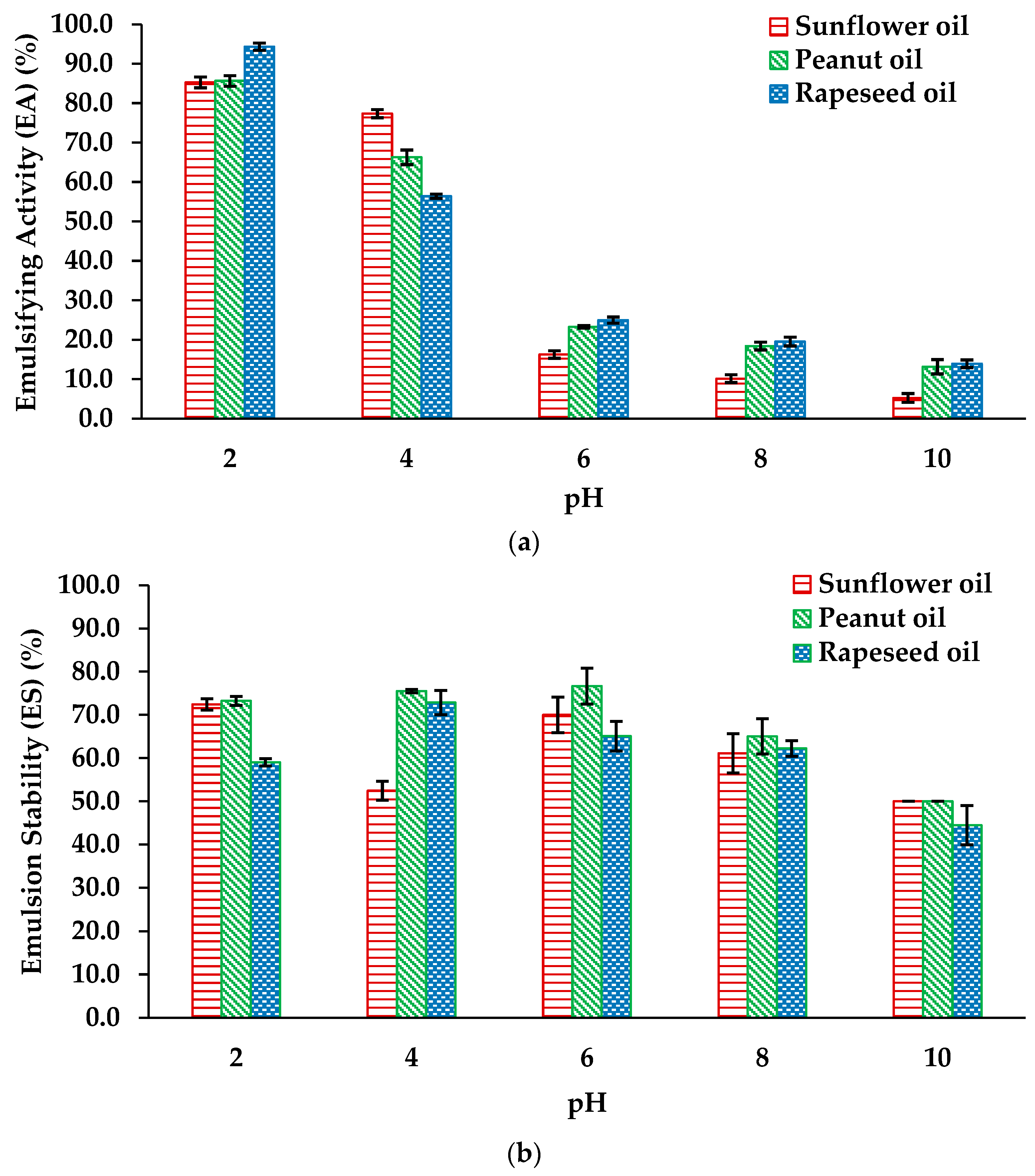
Figure 3.
Foaming characteristics of Chondrus crispus protein hydrolysates across a pH spectrum of 2 to 10. (a) Foaming capacity of Chondrus crispus protein hydrolysates. (SE, standard error, n=3); (b) Foam stability of Chondrus crispus protein hydrolysates. (SE, standard error, n=3).
Figure 3.
Foaming characteristics of Chondrus crispus protein hydrolysates across a pH spectrum of 2 to 10. (a) Foaming capacity of Chondrus crispus protein hydrolysates. (SE, standard error, n=3); (b) Foam stability of Chondrus crispus protein hydrolysates. (SE, standard error, n=3).
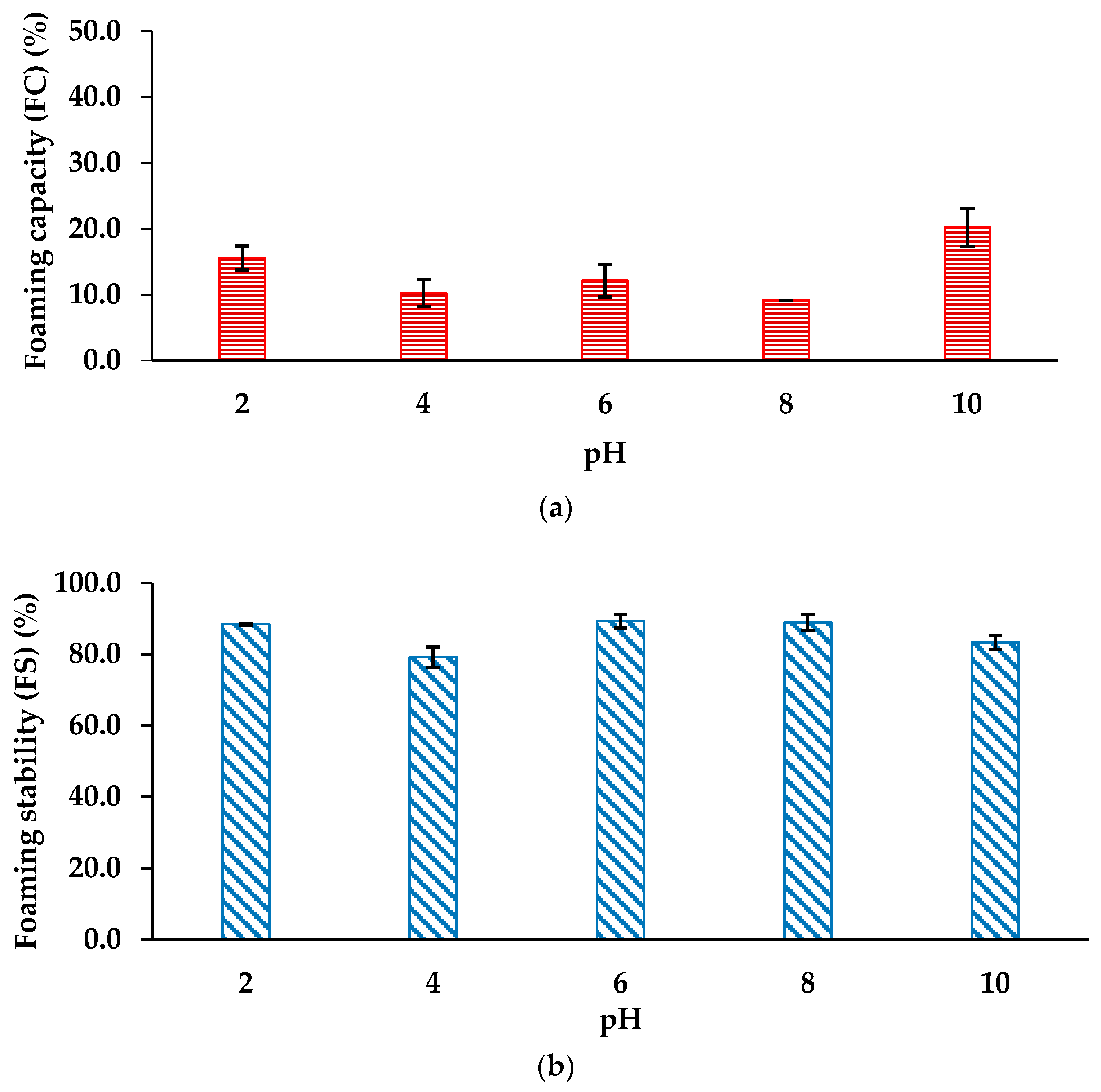
Figure 4.
(a) Vegan fairy cakes containing protein hydrolysates as an emulsifier (b) Control fairy cake.
Figure 4.
(a) Vegan fairy cakes containing protein hydrolysates as an emulsifier (b) Control fairy cake.


Table 1.
Vegan fairy cake recipe.
| Ingredients | Quantity (g or ml) |
| Oat milk | 150 ml |
| Self-raising flour | 150 g |
| Caster sugar | 150 g |
| Rapeseed oil | 60 ml |
| Salt | ½ Pinch |
| Baking powder | Pinch |
| Vanilla extract | ½ tsp |
Table 2.
Control fairy cake recipe.
| Ingredients | Quantity (g or ml) |
| Butter | 150 g |
| Self-raising flour | 150 g |
| Caster sugar | 150 g |
| Eggs | 2 |
| Vanilla extract | ½ tsp |
Table 3.
Sensory evaluation scoresheet for fairy cakes.
| Quality | Standard For Evaluation | Score |
| Appearance | Neat shape, flat surface and no wrinkle, collapse and crack | 8 – 10 |
| Neat shape and surface with minor wrinkle collapse or crack | 5 – 7 | |
| Irregular shape and surface with evident wrinkle, collapse and crack | 0 – 4 | |
| Colour | Gold-yellow surface, milk yellow inside, uniform colour and no spots | 8 – 10 |
| Yellow surface, light yellow inside, uniform colour and a few spots | 5 – 7 | |
| Uneven colour and many spots | 0 – 4 | |
| Odour | Pure fragrance, no peculiar smell | 8 – 10 |
| A little fragrance, no peculiar smell | 5 – 7 | |
| No fragrance, with a peculiar smell | 0 – 4 | |
| Texture | Elastic, uniform honeycomb structure and even pore size | 8 – 10 |
| Elastic, honeycomb structure with some large pore size | 5 – 7 | |
| Non-uniform honeycomb structure with plenty of large holes | 0 – 4 |
Table 4.
Extraction yields obtained from independent hydrolysis of Chondrus crispus with viscozyme and alcalase, SD = (standard deviation, n=3).
Table 4.
Extraction yields obtained from independent hydrolysis of Chondrus crispus with viscozyme and alcalase, SD = (standard deviation, n=3).
| Biomass type | Seaweed |
| Chondrus crispus | |
| Whole biomass | 50 ± 0 g |
| Supernatant (protein hydrolysates) | 9.71 ± 0.23 g |
| Protein hydrolysates yield (%) | 19.42 ± 0.46 % |
Table 5.
Colour coordinates of Chondrus crispus protein hydrolysates and whole biomass (SE, standard error, n=3).
Table 5.
Colour coordinates of Chondrus crispus protein hydrolysates and whole biomass (SE, standard error, n=3).
| Colour coordinates | Whole biomass | Protein hydrolysates |
| L* value | 51.02 ± 0.85 | 68.35 ± 0.25 |
| a* (red to green) | 3.08 ± 0.39 | 4.44 ± 0.05 |
| b* (yellow to blue) | 6.37 ± 0.56 | 11.60 ± 0.10 |
| C* | 7.19 ± 0.43 | 12.14 ± 0.11 |
| h° | 62.89 ± 4.41 | 69.23 ± 0.17 |
| ∆E* ab | 18.34 | |
Table 6.
Water activity and pH of Chondrus crispus protein hydrolysates (SD, standard deviation, n=3).
Table 6.
Water activity and pH of Chondrus crispus protein hydrolysates (SD, standard deviation, n=3).
| Water activity (aW) | 0.45 ± 0.005 |
| pH | 6.82 ± 0.01 |
Table 7.
Water Holding Capacity and Oil Holding Capacity of Chondrus crispus protein hydrolysates (SE, standard error, n=3).
Table 7.
Water Holding Capacity and Oil Holding Capacity of Chondrus crispus protein hydrolysates (SE, standard error, n=3).
| (g of water or oil/g of hydrolysate) | ||
| Water Holding Capacity (WHC) | 10.145 ± 0.221 | |
|
Oil Holding Capacity (OHC) |
Sunflower oil | 9.226 ± 0.084 |
| Peanut oil | 9.175 ± 0.135 | |
| Rapeseed oil | 9.169 ± 0.137 |
Table 8.
Peptide sequences identified from C. crispus hydrolysate and in silico analysis predicted bioactivity results for selected peptides identified using mass spectrometry using different programmes, including PeptideRanker http://bioware.ucd.ie/~compass/biowareweb/, accessed on 17th January 2025; PreAIP http://kurata14.bio.kyutech.ac.jp/PreAIP/, accessed on 17th of January 2025; BIO-PEP-UWM https://biochemia.uwm.edu.pl/biopep-uwm/, accessed on 17th of January 2025; AntiDMPpred available at http://i.uestc.edu.cn/AntiDMPpred/cgi-bin/AntiDMPpred.pl, accessed in January 2025 and Umami-MRNN https://umami-mrnn.herokuapp.com/, accessed in January 2025.
Table 8.
Peptide sequences identified from C. crispus hydrolysate and in silico analysis predicted bioactivity results for selected peptides identified using mass spectrometry using different programmes, including PeptideRanker http://bioware.ucd.ie/~compass/biowareweb/, accessed on 17th January 2025; PreAIP http://kurata14.bio.kyutech.ac.jp/PreAIP/, accessed on 17th of January 2025; BIO-PEP-UWM https://biochemia.uwm.edu.pl/biopep-uwm/, accessed on 17th of January 2025; AntiDMPpred available at http://i.uestc.edu.cn/AntiDMPpred/cgi-bin/AntiDMPpred.pl, accessed in January 2025 and Umami-MRNN https://umami-mrnn.herokuapp.com/, accessed in January 2025.
| Peptide Sequence | Protein & accession number | Peptide Ranker Score | Anti-inflammatory prediction with PreAIP RF (combined values) | Anti-diabetic potential prediction with AntiDMPpred (tp 0.6) | Umami prediction with Umami-MRNN | Novelity (found in BIOPEP-UWM) |
| APPPPPPAPF | Uncharacterized protein (R7QQW5_CHOCR) | 0.97 | Negative AIP (0.292) | No | non-umami | Novel |
| DDDPFGPF | Tyrosine-protein kinase ephrin type A/B receptor-like domain-containing protein (R7QEH3_CHOCR) | 0.959 | Negative AIP (0.299) | No | non-umami | Novel |
| VDPSPWPF | Cytochrome c oxidase subunit 3 (COX3_CHOCR) | 0.945 | Low confidence AIP (0.376) | No | non-umami | Novel |
| GMPSDDPFPF | Malate dehydrogenase (R7QPY8_CHOCR) | 0.942 | Low confidence AIP (0.372) | No | umami, predicted threshold: 39.25773mmol/L | Novel |
| SIPDGWMGL | Phosphoglycerate kinase (Q8GVE6_CHOCR) | 0.939 | Medium confidence AIP (0.443) | No | umami, predicted threshold: 31.829782mmol/L | Novel |
| FDPVFIM | Potential copper-zinc superoxide dismutase, CuZn SOD2 R7QRY5_CHOCR) | 0.933 | Medium confidence AIP (0.436) | No | non-umami | Novel |
| SADDDPFGPF | Tyrosine-protein kinase ephrin type A/B receptor-like domain-containing protein (R7QEH3_CHOCR) | 0.932 | Negative AIP (0.299) | No | umami, predicted threshold: 27.236626mmol/L | Novel |
| GGGAPFFPVA | SMP-30/Gluconolactonase/LRE-like region domain-containing protein (R7QCK1_CHOCR) | 0.93 | Low confidence AIP (0.367) | No | non-umami | Novel |
| GPGPGPGPGPGPGPV | Hint domain-containing protein (R7QIA6_CHOCR) | 0.926 | Negative AIP (0.241) | Predicted to be anti-diabetic | umami, predicted threshold: 15.696608mmol/L | Novel |
| VGPPPPPPPPP | C-CAP/cofactor C-like domain-containing protein (R7QP74_CHOCR) | 0.923 | Negative AIP (0.342) | No | non-umami | Novel |
| AMPPFPNPY | Uncharacterized protein (R7QQW5_CHOCR) | 0.916 | Medium confidence AIP (0.410) | No | non-umami | Novel |
| ADMGGMGF | NAD-dependent sugar epimerase (R7QUS9_CHOCR) | 0.913 | Medium confidence AIP (0.434) | No | umami, predicted threshold: 8.370863mmol/L | Novel |
| SPPPPVDPF | Acetohydroxy-acid reductoisomerase (R7QNL4_CHOCR) | 0.912 | Negative AIP (0.317) | No | non-umami | Novel |
| AGPGPGPGPGPGPGPV | Hint domain-containing protein (R7QIA6_CHOCR) | 0.911 | Medium confidence AIP (0.444) | Predicted to be anti-diabetic | umami, predicted threshold: 22.584654mmol/L | Novel |
| DDFDFNPL | catalase (R7QFJ3_CHOCR) | 0.91 | Low confidence AIP (0.349) | No | non-umami | Novel |
| FAGGPKIPF | Ascorbate Peroxidase, APX1(R7QH61_CHOCR) | 0.906 | Medium confidence AIP (0.392) | No | umami, predicted threshold: 38.71417mmol/L | Novel |
| VGGDPSFGPF | NAD-dependent epimerase/dehydratase domain-containing protein (R7Q496_CHOCR) | 0.9 | Low confidence AIP (0.369) | No | umami, predicted threshold: 26.573849mmol/L | Novel |
| GIPDEWMGL | Phosphoglycerate kinase (Q8GVE6_CHOCR) | 0.9 | High confidence AIP (0.469) | No | umami, predicted threshold: 36.3166mmol/L | Novel |
| FDPVFIMN | Potential copper-zinc superoxide dismutase, CuZn SOD2 (R7QRY5_CHOCR) | 0.9 | Medium confidence AIP (0.436) | No | non-umami | Novel |
| VLPSLPFM | Animal heme peroxidase homologue (R7QJ77_CHOCR) | 0.89 | High confidence AIP (0.478) | No | umami, predicted threshold: 34.225975mmol/L | Novel |
| SSPQFGFPT | SMP-30/Gluconolactonase/LRE-like region domain-containing protein (R7QCK1_CHOCR) | 0.89 | Negative AIP (0.335) | No | umami, predicted threshold: 36.78789mmol/L | Novel |
| ALEPPPPVGPPGL | Ubiquitin carboxyl-terminal hydrolase (R7QGF5_CHOCR) | 0.89 | Medium confidence AIP (0.402) | Predicted to be anti-diabetic | umami, predicted threshold: 26.183607mmol/L | Novel |
| GVSAPFDPF | Translation initiation factor eIF1 (R7QJC0_CHOCR) | 0.88 | Low confidence AIP (0.363) | No | umami, predicted threshold: 33.65273mmol/L | Novel |
| YAMPPYAF | Photosystem I P700 chlorophyll a apoprotein A2 (M5DES3_CHOCR) | 0.88 | Low confidence AIP (0.379) | No | non-umami | Novel |
| SPDMPGYVGF | Uncharacterized protein (R7QQW5_CHOCR) | 0.88 | Low confidence AIP (0.364) | No | non-umami | Novel |
| DSDSDLFFG | Gal-2,6-Sulfurylases II (R7Q2Y4_CHOCR) | 0.88 | Low confidence AIP (0.361) | No | umami, predicted threshold: 29.827625mmol/L | Novel |
| GGPAGGGPAGGDAGLA | Transitional endoplasmic reticulum ATPase (Valosin-containing protein), (R7QAA8_CHOCR) | 0.87 | High confidence AIP (0.480) | Predicted to be anti-diabetic | umami, predicted threshold: 13.955362mmol/L | Novel |
| AQVPPLPNMPRMPA | Prohibitin (R7QRM0_CHOCR) | 0.87 | High confidence AIP (0.478) | Predicted to be anti-diabetic | umami, predicted threshold: 37.8484mmol/L | Novel |
| DSDSDLFFGR | Gal-2,6-Sulfurylases II (R7Q2Y4_CHOCR) | 0.87 | Low confidence AIP (0.384) | No | umami, predicted threshold: 8.039184mmol/L | Novel |
Table 8.
Baking loss (%) and colour (L*, a*, b* and ∆E*ab) of control and formulated vegan fairy cakes containing protein hydrolysates (± SE, standard error, n=9).
Table 8.
Baking loss (%) and colour (L*, a*, b* and ∆E*ab) of control and formulated vegan fairy cakes containing protein hydrolysates (± SE, standard error, n=9).
| Samples | Baking loss (%) | Colour characteristics | ||||
| L* | a* | b* | ∆E*ab | |||
| Control cakes | 8.92 ± 0.63 | Crust | 49.50 ± 0.33 | 17.52 ± 0.15 | 37.54 ± 0.22 | Crust (10.16)Crumb (11.61) |
| Crumb | 68.02 ± 0.42 | 2.70 ± 0.09 | 38.03 ± 0.21 | |||
| 11.72 ± 0.13 | Crust | 56.23 ± 0.22 | 10.03 ± 0.26 | 36.17 ± 0.36 | ||
| Vegan cakes | Crumb | 59.93 ± 0.37 | 3.25 ± 0.06 | 29.71 ± 0.30 | ||
Table 9.
Cake density (g/cm3), specific volume (cm3/g), and volume index (cm) of control and formulated vegan fairy cakes containing protein hydrolysates (± SE, standard error, n=9).
Table 9.
Cake density (g/cm3), specific volume (cm3/g), and volume index (cm) of control and formulated vegan fairy cakes containing protein hydrolysates (± SE, standard error, n=9).
| Cake density (g/cm3) | Specific volume (cm3/g) | Volume index (cm) | |
| Control cake | 0.51 ± 0.01 | 1.95 ± 0.02 | 9.66 ± 0.13 |
| Vegan cake | 0.61 ± 0.02 | 1.66 ± 0.04 | 9.21 ± 0.18 |
Table 10.
Moisture content (%), water activity, and pH of control and formulated vegan fairy cakes containing protein hydrolysates (SE, standard error, n=3).
Table 10.
Moisture content (%), water activity, and pH of control and formulated vegan fairy cakes containing protein hydrolysates (SE, standard error, n=3).
| Days | Moisture content (%) | Water activity (aW) | pH | |||
| Control | Vegan | Control | Vegan | Control | Vegan | |
| 0 | 26.03 ± 0.41 | 29.34 ± 0.18 | 0.810 ± 0.001 | 0.754 ± 0.004 | 7.84 ± 0.01 | 7.25 ± 0.00 |
| 1 | 24.63 ± 0.60 | 28.15 ± 0.13 | 0.817 ± 0.003 | 0.759 ± 0.005 | 7.85 ± 0.02 | 7.30 ± 0.01 |
| 2 | 24.06 ± 0.08 | 27.20 ± 0.07 | 0.822 ± 0.003 | 0.762 ± 0.005 | 7.89 ± 0.00 | 7.31 ± 0.01 |
Table 11.
Sensory evaluation of control and formulated vegan fairy cakes containing protein hydrolysates (SE, standard error, n=3).
Table 11.
Sensory evaluation of control and formulated vegan fairy cakes containing protein hydrolysates (SE, standard error, n=3).
| Days | Appearance | Colour | Odour | Texture |
| Control cake | 9.33 ± 0.22 | 9.78 ± 0.14 | 10.00 ± 0.00 | 9.67 ± 0.16 |
| Vegan cake | 8.67 ± 0.22 | 8.56 ± 0.23 | 7.56 ± 0.17 | 7.67 ± 0.16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



