
Submitted:
29 January 2025
Posted:
30 January 2025
You are already at the latest version
Abstract
Endometriosis is a pathological condition characterized by the presence of the endometrial tissue, including epithelial and stromal components, outside the uterine cavity. It affects nearby 10% of women of reproductive age and is responsible for infertility, chronic pain and the weakening of the quality of life. Various pathogenetic mechanisms have been suggested, such as retrograde menstruation, coelomic metaplasia, embryological defects, stem cell involvement, and alterations in epigenetic regulation. However, the essential pathogenesis of endometriosis remains insufficiently comprehended. Individuating the exact mechanism of the growth and development of endometriosis is essential for valuable cure. This review reports the foremost pathogenetic theories of endometriosis based on recent investigations with a major emphasis on the potential role of uterine adenogenesis factors.
Keywords:
endometriosis
; endometrium
; uterus
; adenogenesis
; adenomiosys
1. Introduction
Approximately 10% of women of reproductive age are affected by endometriosis. This condition can present symptoms such as intense pain, painful menstruation (dysmenorrhea), painful intercourse (dyspareunia), difficulty urinating (dysuria), infertility, and fatigue [1,2]. The exact cause remains unknown, but it may be linked to the complex development and maturation of the uterus. Regardless of its origin, there is broad consensus that endometriosis is linked to a localized inflammatory response, with vascularization at the site of invasion playing a critical role in its pathogenesis. While it is well established that endometriotic lesions consist of stromal and epithelial glandular cells, the development and specific characteristics of the cell types involved in the disease’s pathogenesis remain poorly understood [3].
In humans, the uterus is not fully developed or functionally differentiated at birth. While its basic structure is present, the uterus undergoes significant postnatal changes in size, morphology, and cellular organization. These developmental processes, influenced by hormonal cues during childhood, puberty, and later life, gradually prepare the uterus for its reproductive functions [4]. Endometriosis is often discovered during evaluations for infertility, but a direct causal relationship has not been definitively established. Surgical visualization remains the gold standard for diagnosis [5], which may lead to an overestimation of its prevalence and incidence among infertile women compared to the general population [6]. Women with otherwise ‘asymptomatic’ endometriosis linked to underlying infertility causes may be disproportionately represented. Without attempts to conceive, these women might not have undergone laparoscopic diagnosis [7]. Moreover, high-quality clinical trials have shown no significant differences in spontaneous pregnancy rates among endometriosis patients receiving hormonal treatments [8]. This persistence may partly explain the lack of improvement in infertility. Additionally, surgical removal of pelvic lesions offers only modest benefits for reproductive outcomes [9]. While current treatments for endometriosis primarily focus on symptom relief, such as pain management, surgical removal, and hormonal suppression, the development of a more targeted treatment, based on a deeper understanding of its pathological mechanisms, remains an urgent need.
The disease is estrogen-dependent, with the expression of estrogen receptor (ER)β significantly higher in EMS compared to normal endometrial tissues. Elevated levels of ERβ suppress ERα, leading to very high ERβ/ERα ratios [10]. ERβ plays a critical role in EMS pathogenesis by modulating apoptosis and the inflammasome. Furthermore, ERβ regulates mitochondrial function through nuclear respiratory factor-1 (NRF1) and helps control the basal expression of superoxide dismutase 2 (SOD2) to minimize oxidative stress [11]. ERβ also influences the expression of cyclooxygenase-2 (COX2), involved in inflammation, and matrix metalloproteinase 1 (MMP1), which contributes to tissue degradation, cell invasion, and growth [12,13]. Although endometriosis is not considered a tumor, many of its features, such as tissue invasion and neovascularization, are like those of malignant tumors. The microenvironment of endometriotic lesions is characterized by chronic inflammation, oxidative stress, and angiogenesis. These factors contribute to the pathogenesis of the disease and may explain the resistance to pharmacological treatments [14]. Given the complexities and gaps in understanding the pathogenesis of endometriosis, this review aims to consolidate and critically evaluate the foremost pathogenetic theories based on recent investigations. Specifically, we focus on the potential role of uterine adenogenesis factors in the development and progression of endometriosis. By shedding light on these mechanisms, we hope to contribute to a deeper understanding that could pave the way for more targeted and effective therapeutic strategies for this condition.
2. Development of the Female Reproductive System
The development of the human female reproductive system is a topic of significant academic and clinical importance. Congenital malformations of this anatomical region often arise from disruptions in normal morphogenetic processes and the molecular mechanisms that support them. Even though relatively rare, many of these congenital anomalies are associated with defects in Müllerian duct (MD) development. Some occur spontaneously, while others are induced by exposure to endocrine-disrupting substances, particularly those with estrogenic activity [15].
Most of the female reproductive tract develops from the Müllerian ducts (MD), which arise as coelomic epithelial invaginations on the urogenital ridges around 5–6 weeks of gestation [16] (Figure 1). These invaginations form the ostia of the uterine tubes. The MDs grow caudally, using the Wolffian ducts (WD) as “guide wires,” as the WDs are essential for the caudal migration of the MDs toward the urogenital sinus (UGS). During migration, the MDs and WDs are initially separated by the mesenchyme of the urogenital ridge. However, further caudally, their epithelia come closer together, eventually sharing a common basement membrane. At the tip of the MDs, direct contact is established with the WD epithelium. The fusion of the MDs with the UGS, critical for forming the uterovaginal canal, occurs by the 8th week of gestation. Failures during this stage, such as unsuccessful fusion, can lead to malformations like vaginal agenesis [17]. During the fusion process, a temporary epithelial septum separates the MD lumina, but this septum disappears by the 9th week, forming a single uterovaginal canal lined by undifferentiated columnar epithelium of Müllerian origin. Mutations in key genes such as Lhx1 and Lhfpl2 can disrupt this process, leading to degeneration of the ducts or malformations of the reproductive tract [18,19].
The uterine tubes, also known as Fallopian tubes, consist of four main sections: the infundibulum, funnel-shaped with fimbriae; the ampulla, where fertilization typically occurs; the isthmus, located near the uterus; and the intramural portion, which traverses the uterine wall and ends at the uterotubal junction. These structures develop from the cranial portions of the MD, while the caudal segments fuse to form the uterovaginal canal. The fimbriae of the tubes arise from the irregular openings of the MD into the abdominal cavity.
During development, the fetal tubal epithelium initially consists of a single layer of columnar cells. Over time, mucosal folds develop and become especially prominent in infundibulum and ampulla by the fourteenth week of gestation. Concurrently, the surrounding mesenchyme differentiates into an inner stromal layer and an outer layer of circularly arranged smooth muscle [20]. In adulthood, the tubal epithelium includes ciliated cells, secretory cells, and intercalated (“peg”) cells. During fetal life, epithelium expresses differentiation markers such as keratins 7, 8, and 19, starting from the ninth week. Androgen receptors and ESR1 appear around the fourteenth week, whereas the progesterone receptor is undetectable between the eighth and twenty-first weeks, although it can be artificially induced under experimental conditions [21].
Numerous cellular and molecular mechanisms are involved in the development and differentiation of the MD in the female reproductive organs. Despite significant progress in recent years in understanding the signaling pathways that regulate the development of the female reproductive system, many areas remain poorly understood, particularly regarding the specific processes that coordinate the formation and maturation of reproductive organs [22].
Several signaling pathways have been identified as playing a crucial role in the development of the MDs and uterine tubes, including bone morphogenetic proteins (BMP), the WNT signaling pathway, transforming growth factor-β (TGF-β), the PI3K/Akt pathway, G protein-coupled receptors, and fibroblast growth factors (FGF). These molecular mechanisms are involved in various stages of development, from cell proliferation to differentiation and the formation of the final anatomical structures. In particular, BMP signaling is essential for the proper development and function of the female reproductive system [23]. The BMP pathway regulates several critical aspects, including the formation of the MDs, their migration and fusion, and the subsequent differentiation of reproductive organs. Additionally, BMP signaling interacts with other pathways, such as WNT, to maintain the homeostasis of the reproductive system throughout both embryonic and postnatal development. While much has been learned about these signaling pathways, a complete understanding of how they interact in a coordinated manner remains an area of intense research [24,25,26].
The WNT protein family plays a critical role in numerous developmental processes and in tissue homeostasis. Among these, WNT4 plays a pivotal role in shaping the female phenotype during fetal development and in sustaining the integrity of Müllerian and reproductive tissues [27]. Alterations or dysregulation of WNT4 are associated with conditions such as sex reversal syndromes, highlighting its crucial function in female sex determination. Furthermore, WNT4 is involved in various gynecological pathologies, including uterine fibroids, endometriosis, infertility, and tumors [28]. WNT4 also plays a significant role in processes such as decidualization, implantation, and gestation, which are essential for normal reproduction. However, abnormal activation of WNT4 signaling has been linked to the development of gynecological and breast tumors. Single nucleotide polymorphisms (SNPs) in the WNT4 gene have been strongly associated with these conditions and may represent a point of connection between estrogen signaling and WNT4-mediated pathways, contributing to the upregulation of WNT4 activity. SNPs in the 1p36 region of the human genome, which includes the WNT4 gene, are linked to an increased risk of various gynecological conditions. Genome-wide association studies (GWAS) have identified SNPs in this region as associated with endometriosis (often accompanied by infertility), uterine fibroids, ovarian cancer, and pelvic organ prolapse [29]. Notably, these variants increase the risk for endometriosis and ovarian cancer but appear to have a protective effect against pelvic organ prolapse and are associated with slightly longer gestation durations (approximately two additional days per allele) [30,31,32]. The primary SNP in most studies is located within the WNT4 gene locus, and its associations have been confirmed through multiple independent studies and meta-analyses. These findings are consistent across diverse ethnic and geographic populations, underscoring the robust nature of the link between SNPs in this genomic region and gynecological health outcomes.
In each developmental or pathological context, the expression and activity of WNT4 signaling are regulated by tissue-specific pathways [33].
3. Uterine Adenogenesis
Uterine adenogenesis, the development of endometrial glands, is a critical process for the formation and functionality of the adult uterus [34,35]. In humans, this process begins during fetal life, continues postnatally, and completes at puberty. In other species, such as rodents, sheep, and pigs, adenogenesis primarily occurs postnatally. Endometrial gland formation involves the differentiation and budding of glandular epithelium from the luminal epithelium, followed by invagination, tubular morphogenesis, and branching into the uterine stroma. This process is regulated by both local and systemic factors, including site-specific changes in cell proliferation, extracellular matrix (ECM) remodeling, and paracrine signaling between cells and the ECM [36]. Several hormones are involved in regulating adenogenesis, including prolactin and estradiol-17β, which influence cell proliferation, extracellular matrix remodeling, and paracrine signaling. These hormones play a significant role in regulating both the development and function of endometrial glands in mammals, with receptors for these hormones being crucial for the process. The interaction between these hormones and specific molecular pathways ensures the proper formation of the glands, which are essential for successful implantation and pregnancy [37]. Uterine glands and their secretions are fundamental for blastocyst implantation and decidualization, processes essential for reproduction in rodents and humans. In mice, leukemia inhibitory factor (LIF), produced exclusively by uterine glands, is critical for blastocyst adhesion to the uterine luminal epithelium (LE). Its absence, as observed in Lif knockout mice, results in infertility due to the failure of embryonic implantation. Similarly, in neonatal progesterone-treated mice, which develop aglandular uteri (Progesterone-treated Uterine Gland Knockout: PUGKO), blastocysts hatch but fail to adhere to the LE, and no decidualization is observed, even after artificial stimulation or intrauterine treatment with LIF [38].
During pregnancy, species such as sheep and pigs exhibit hyperplasia and hypertrophy of endometrial glands, necessary for producing histotroph, a critical nutrient source for the conceptus. Alterations or defects in endometrial gland morphogenesis, caused by genetic anomalies, epigenetic influences, or endocrine disorders, can impair uterine functionality and contribute to unexplained peri-implantation embryonic losses in humans and animals. Understanding the mechanisms that regulate uterine adenogenesis is therefore essential for improving reproductive health and addressing infertility and reproductive failures [34,37].
Adenogenesis involves the coordinated expression of several critical genes, including forkhead box A2 (Foxa2), Wnt4, Wnt5a, Wnt7a, and E-cadherin (Cdh1), among others. One of the most well-studied genes in this process is Foxa2, a transcription factor specifically expressed in uterine glands. Conditional deletion of FOXA2 in the uterus leads to a substantial decrease in the number of uterine glands, indicating its crucial role in glandular development and uterine function [39]. The Wnt signaling pathway, particularly Wnt4, Wnt5a, and Wnt7a, is also fundamental for adenogenesis. Studies have shown that the deletion of Wnt7a or Wnt4 in the mouse uterus results in either a complete absence or a significant reduction in the number of uterine glands [40,41]. Additionally, E-cadherin (Cdh1), a key molecule involved in cell-cell adhesion, is essential for proper gland development. Conditional ablation of Cdh1 results in the loss of uterine glands in neonatal mice, highlighting the importance of cell adhesion molecules in maintaining the integrity and functionality of the developing uterine glands [42].
4. Pathogenesis of Endometriosis: Mechanisms and Theories
Endometriosis is characterized by the presence of endometrial-like cells outside the uterus, leading to a chronic inflammatory response. It presents a wide range of clinical manifestations, with common symptoms including chronic abdominal or pelvic pain, painful menstruation (dysmenorrhea), painful intercourse (dyspareunia), and painful defecation (dyschezia) [1,14]. The specific symptoms can vary depending on the organ or site involved: abdominal wall pain, chest pain, hemoptysis, or bladder dysfunctions such as urgency, urinary frequency, and dysuria. In cases of intestinal endometriosis, patients may experience diarrhea, constipation, intestinal cramps, dyschezia, and, in rare cases, rectal bleeding. An increasing number of symptoms is associated with a higher probability of endometriosis diagnosis. In particular, dyschezia and dyspareunia are strongly predictive of deep infiltrating endometriosis (DIE) [43]. However, the clinical presentation is highly variable, ranging from asymptomatic forms to severe symptoms, contributing to the long delay between symptom onset and diagnosis, which can range from 7 to 12 years. One study found that 30% of patients were referred to a gynecologist at least twice before receiving a definitive diagnosis [44,45].
Research on endometriosis has gained significant attention in recent decades, but substantial challenges remain, particularly in achieving timely diagnosis. Factors such as diverse symptom presentation, the lack of standardized diagnostic criteria, and the underestimation of symptoms by both patients and healthcare providers contribute to prolonged diagnostic delays, averaging 4–10 years. These delays can exacerbate symptoms, reduce quality of life, and increase healthcare costs [6]. While historical and recent studies indicate slight improvements in diagnostic timelines, largely due to enhanced awareness and targeted initiatives such as advocacy campaigns, educational programs for clinicians, and public awareness efforts, significant work is still needed. Reducing diagnostic delays is critical for improving patient outcomes and the effective management of endometriosis [46]. Interestingly, recent data from our research group and from others have indicated some molecular markers that could be used as diagnostic tools for endometriosis [47].
The pathogenesis of endometriosis is multifactorial, involving genetic and cellular factors, as well as altered immunity (Table 1). The most widely accepted theory is Sampson’s retrograde menstruation theory, which suggests that endometrial cells flow through the fallopian tubes into the peritoneal cavity [48]. The retrograde menstruation theory proposes that during menstruation, some endometrial tissue flows backward through the fallopian tubes into the peritoneal cavity, causing endometriosis. This explains common sites of endometriosis, like the fallopian tubes and peritoneal wall, and is supported by the increased risk in women with uterine outflow obstruction and the frequent occurrence of blood in the pelvis during menstruation [2,49]. However, this theory fails to explain deep endometriosis, where lesions are deeper in the pelvic organs, or endometriosis in distant sites like the lungs and skin. It also doesn’t apply to endometriosis in adolescents, newborns, women with Mayer-Rokitansky-Küster-Hauser syndrome, or men. Moreover, studies suggest that endometriotic tissue does not resemble an auto-transplant [50,51,52].
In 1942, Gruenwald proposed the theory of coelomic metaplasia, which suggests that the coelomic walls, such as the peritoneal serosa, are embryologically linked to the Müllerian ducts. According to this theory, endometriosis may develop in various coelomic derivatives due to a metaplastic process. Additionally, endocrine-disrupting chemicals might play a role in the transformation or activation of these coelomic cells, as suggested by several studies. This theory could help explain cases where retrograde menstrual flow is not possible [53,54].
The lymphatic and vascular spread theory suggests that endometrial cells could spread through lymphatic or blood vessels. In this model, it is proposed that menstrual endometrium, containing both epithelial and stromal cells, can enter these circulatory systems without disruption and then exit the vessels to implant in the muscular layers of organs. While deep endometriosis shares characteristics with cancer metastasis, such as tissue invasion, these are typical of tumor cells. However, there is currently no scientific evidence to support the notion that menstrual endometrium, originating from benign tissue, can perform such cancer-like functions [50].
The stem cell recruitment theory suggests that endometriosis may arise from stem cells, either from the uterine endometrium or the bone-marrow. The first hypothesis proposes that stem cells from the uterine endometrium, specifically from the basal glands, are responsible for regenerating the endometrial epithelium and could contribute to endometriosis if misplaced. These epithelial stem cells are thought to reside in the basal layer at the endometrial/myometrial interface, although no specific markers have been identified for them [55]. The second hypothesis suggests that bone marrow-derived stem cells are a key source of endometriosis, especially when these cells differentiate into both epithelial and stromal cells. If these stem cells are misplaced and end up in soft tissues instead of returning to the endometrium, endometriosis may develop. Experimental evidence supports this theory, with bone marrow stem cells also implicated in rare cases of endometriosis occurring outside the peritoneal cavity or in men [56,57].
The theory of embryonic cell remnants proposes that remnants of embryonic cells from the Wolffian or Müllerian ducts can develop into endometriotic lesions under the influence of estrogen. This theory suggests that abnormal differentiation or migration of the Müllerian ducts during embryogenesis could lead to the spread of primordial endometrial cells, which remain dormant until puberty, when estrogen activation causes their proliferation. This mechanism could explain the common sites of endometriosis, such as the deep peritoneum and the pouch of Douglas [58].
Recent studies have supported this theory, with findings of ectopic endometrial structures in female fetuses during autopsies. These structures were found in locations typically affected by endometriosis in adults, such as the Douglas pouch and rectovaginal septum. This provides strong evidence for the fetal origin of endometriosis [59,60,61].
Immunohistochemical studies of endometriotic structures have shown that these lesions have characteristics similar to ectopic endometrium, with both epithelial components (expressing estrogen receptor, CA-125, and cytokeratin 7) and stromal cells (positive for CD-10 and estrogen receptor). Interestingly, fetal endometrial cells from these patients showed identical staining patterns. These findings support the theory of embryonic cell remnants as a cause of endometriosis, showing that ectopic endometrial structures can persist outside the uterus during organogenesis [62]. Further supporting this theory, a study by de Jolinière et al. found ectopic endometrial glands in female fetuses, with similar immunohistochemical profiles [63]. In addition, previous studies have once again reported cases of fetal endometriosis, such as a cystic mass in the left ovary diagnosed in a fetus at 35 weeks [64]. The presence of displaced Müllerian tissue during organogenesis has been suggested by various research groups as a potential factor in the development of endometriosis. Additionally, a significant body of research has demonstrated a strong correlation between uterine malformations or Müllerian duct anomalies and the occurrence of endometriosis [65]. Moreover, genomic studies by our research group have shown that the transcriptome of endometriotic tissue expresses numerous genes related to embryogenesis in a manner distinct from normal endometrial tissue. Notably, this altered gene expression occurs independently of the hormonal cycle phase [66,67].
Based on these observations, it can be hypothesized that endometriosis results from disturbances in uterine embryogenesis during a critical period of morphogenesis. These disruptions could lead to the abnormal displacement of endometrial tissue outside the uterine cavity. However, the specific genetic and epigenetic factors responsible for these disturbances remain unidentified [68]. Estrogen, a key hormone in the development of the female reproductive system, may play a central role in this process. It is possible that altered estrogen levels, acting on a genetic predisposition, could trigger the formation of ectopic endometrial tissue during embryogenesis. Furthermore, there is substantial evidence linking exposure to endocrine disruptors, substances that mimic estrogen, to both uterine abnormalities and endometriosis [69,70,71].
5. Expression Patterns of Different Adenogenesis Factors in Endometriosis and Normal Endometrium
Our previous studies investigated stromal and epithelial samples from deep endometriosis lesions, along with normal endometrial epithelium and stroma used as controls. The analysis revealed no significant morphological differences between samples taken during the proliferative and secretory phases. Endometriotic lesions consistently showed the presence of both endometriotic glands and stroma, regardless of the lesion site. The glands displayed a classical endometrioid appearance, while the stroma closely resembled eutopic endometrial stroma in inactive or proliferative states.
The immunohistochemical analysis focused on the expression of IFN-τ, FGF-7, FGF-10, FGF-23, and HGF in both normal endometrial tissues and deep endometriosis samples. Specific cytoplasmic immunopositivity was detected for all antibodies tested. The findings demonstrated that FGF-7, FGF-10, and HGF exhibited significantly higher expression levels in the epithelium and stroma of normal controls compared to endometriosis samples. In contrast, FGF-23 and IFN-τ showed significantly higher expression in the ectopic endometrial stroma of endometriosis samples relative to eutopic endometrium, whereas no notable differences in epithelial expression were observed between the two tissue types.
Statistical analysis of the normal endometrium during the proliferative and secretory phases revealed no significant differences in the expression of these growth factors within either the epithelial or stromal compartments. Similarly, this pattern was observed in endometriotic tissues, suggesting that fluctuations in estrogen and progesterone levels do not substantially influence the expression of these factors in normal or endometriotic samples, highlighting molecular distinctions between deep endometriosis and normal endometrium. These observations support the notion that endometriotic tissues, including both epithelial and stromal components, exhibit a distinct phenotype compared to the eutopic endometrium. This reinforces the hypothesis that alterations in the molecular mechanisms regulating adenogenesis and the survival of endometrial structures are closely linked to the development and persistence of endometriotic lesions outside the uterus [72].
In a preceding study, we analyzed stromal and epithelial samples from endometriosis lesions and normal endometrium controls during both the proliferative and secretory phases. Immunohistochemistry was used to assess PRL-R and GH hormones, as well as IGF1 and IGF2 expressions in normal endometrium and endometriosis tissues. IGF1 and IGF2 showed significantly higher expression in both epithelium and stroma of controls compared to endometriosis samples. For PRL-R, this pattern was observed only in the epithelium, while GH displayed significantly higher expression in the epithelium of endometriosis samples compared to controls. Statistical analysis further revealed that in normal uterine tissue, GH and PRL-R expression in the epithelium was significantly higher during the secretory phase compared to the proliferative phase. However, this phase-dependent variation was absent in endometriotic tissues. The correlation data generated provides insights into some of the molecular mechanisms involved in the adenogenesis and survival of endometriotic structures outside the uterus, shedding light on the processes that sustain the development and persistence of these lesions [73].
Finally, we analyzed and compared the expression of various proteoglycans and specific glycosaminoglycans (GAGs), key components of the extracellular matrix, between deep endometriotic lesions and normal endometrial tissue. These GAGs play a crucial role in the interaction between the epithelium and stroma, a process essential for proper uterine gland morphogenesis. The study examined the expression of CSPG4 (chondroitin sulfate proteoglycan 4), CS-56 (a chondroitin sulfate), HEP (heparan sulfate), keratan sulfate, and hyaluronic acid in endometriosis-affected tissues compared to normal endometrium. CSPG4 and CS-56 showed significantly higher expression in the epithelium of endometriosis samples, while HEP was more highly expressed in the epithelium of normal samples. Stromal expression for these markers showed no significant differences. Keratan sulfate was largely absent in the epithelium of endometriosis samples, with stromal expression predominantly low compared to the medium or high intensity observed in normal endometrium. Hyaluronic acid expression was consistently lower in both the epithelium and stroma of endometriosis samples. Logistic regression analysis demonstrated that high epithelial expression of CSPG4, CS-56, and HEP increased the likelihood of association with endometriosis, while high expression of keratan sulfate and hyaluronic acid in both the epithelium and stroma correlated with a reduced likelihood of belonging to the endometriosis group.
Although HEP showed higher epithelial expression in normal endometrium, logistic regression analysis identified that relatively higher expression levels of CSPG4, CS-56, and HEP in the epithelium were predictive of endometriosis, while higher keratan sulfate and hyaluronic acid expression in both epithelium and stroma were associated with a reduced likelihood of endometriosis. All the data described in this section are summarized in Table 2.
These findings highlight significant alterations in marker expression, providing insights into endometriosis pathophysiology and suggesting potential diagnostic markers [74].
6. Conclusions
The pathogenesis of endometriosis is complicated and comprises many causes and processes which can occur simultaneously. Many theories have been investigated, but there is no individual theory up to now, which could justify all aspects of endometriosis. Endometriosis growth and progression probably incorporate elements from all the recorded pathogenetic theories. In this context, the role of uterine adenogenetic factors, also based on the data produced about their differential expression in endometrium respect to endometriosis tissue, is a research area to be deeply investigated. An improved comprehension of the pathogenesis of this cryptic but, unfortunately, very common reproductive disorder, can lead to a better treatment and an enhanced quality of life for patients experiencing endometriosis.
Author Contributions
PGS and AB were responsible for conceptualization, formal analysis and revision; MB and RV were responsible for writing and original draft preparation; PGS was responsible for funding acquisition.
Funding
This work was supported by Fondazione Italiana Endometriosi (Grant: “Progetto Genomico”).
Acknowledgments
We would like to thank Roberta Riposati and all the staff of Fondazione Italiana Endometriosi for their precious help in the research activity of the Foundation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Guo, J.; Wang, Y.; Chen, G. Causal Relationship between Endometriosis, Female Infertility, and Primary Ovarian Failure through Bidirectional Mendelian Randomization. Int. J. Women’s Health 2024, 16, 2143–2155. [Google Scholar] [CrossRef] [PubMed]
- Signorile, P.G.; Cassano, M.; Viceconte, R.; Marcattilj, V.; Baldi, A. Endometriosis: A Retrospective Analysis of Clinical Data from a Cohort of 4,083 Patients, with Focus on Symptoms. In Vivo 2022, 36, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Boccellino, M.; Quagliuolo, L.; Verde, A.; La Porta, R.; Crispi, S.; Piccolo, M.T.; Vitiello, A.; Baldi, A.; Signorile, P.G. In Vitro Model of Stromal and Epithelial Immortalized Endometriotic Cells. J. Cell. Biochem. 2012, 113, 1292–1301. [Google Scholar] [CrossRef] [PubMed]
- Veth, V.B.; Keukens, A.; Reijs, A.; Bongers, M.Y.; Mijatovic, V.; Coppus, S.F.P.J.; Maas, J.W.M. Recurrence after Surgery for Endometrioma: A Systematic Review and Meta-Analysis. Fertil. Steril. 2024, 122, 1079–1093. [Google Scholar] [CrossRef]
- Pardanani, S.; Barbieri, R.L. The Gold Standard for the Surgical Diagnosis of Endometriosis: Visual Findings or Biopsy Results? J. Gynecol. Tech. 1998, 4, 121–124. [Google Scholar]
- Signorile, P.G.; Baldi, A. Looking for an effective and non-invasive diagnostic test for endometriosis: where are we? Transl. Med. 2018, 6 (Suppl. 2), S106. [Google Scholar] [CrossRef]
- Missmer, S.A.; Hankinson, S.E.; Spiegelman, D.; Barbieri, R.L.; Marshall, L.M.; Hunter, D.J. Incidence of laparoscopically confirmed endometriosis by demographic, anthropometric, and lifestyle factors. Am. J. Epidemiol. 2004, 160, 784–796. [Google Scholar] [CrossRef]
- Alkatout, I.; Mettler, L.; Beteta, C.; Hedderich, J.; Jonat, W.; Schollmeyer, T.; Salmassi, A. Combined surgical and hormone therapy for endometriosis is the most effective treatment: prospective, randomized, controlled trial. J. Minim. Invasive Gynecol. 2013, 20, 473–481. [Google Scholar] [CrossRef]
- Vercellini, P.; Vigano, P.; Somigliana, E.; Fedele, L. Endometriosis: pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261–275. [Google Scholar] [CrossRef]
- Liu, Z.; Gou, Y.; Zhang, H.; Zuo, H.; Zhang, H.; Liu, Z.; Yao, D. Estradiol improves cardiovascular function through up-regulation of SOD2 on vascular wall. Redox Biol. 2014, 3, 88–99. [Google Scholar] [CrossRef]
- Klinge, C.M. Estrogenic control of mitochondrial function. Redox Biol. 2020, 31, 101435. [Google Scholar] [CrossRef] [PubMed]
- Su, E.J.; Lin, Z.H.; Zeine, R.; Yin, P.; Reierstad, S.; Innes, J.E.; Bulun, S.E. Estrogen receptor-beta mediates cyclooxygenase-2 expression and vascular prostanoid levels in human placental villous endothelial cells. Am. J. Obstet. Gynecol. 2009, 200, 427.e1–8. [Google Scholar] [CrossRef] [PubMed]
- Hudelist, G.; Keckstein, J.; Czerwenka, K.; Lass, H.; Walter, I.; Auer, M.; Wieser, F.; Wenzl, R.; Kubista, E.; Singer, C.F. Estrogen receptor beta and matrix metalloproteinase 1 are coexpressed in uterine endometrium and endometriotic lesions of patients with endometriosis. Fertil. Steril. 2005, 84 (Suppl. 2), 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Signorile, P.G.; Baldi, A. New evidence in endometriosis. Int. J. Biochem. Cell Biol. 2015, 60, 19–22. [Google Scholar] [CrossRef]
- Cunha, G.R.; Robboy, S.J.; Kurita, T.; Isaacson, D.; Shen, J.; Cao, M.; Baskin, L.S. Development of the human female reproductive tract. Differentiation 2018, 103, 46–65. [Google Scholar] [CrossRef]
- Chandler, T.M.; Machan, L.S.; Cooperberg, P.L.; Harris, A.C.; Chang, S.D. Mullerian duct anomalies: from diagnosis to intervention. Br. J. Radiol. 2009, 82, 1034–1042. [Google Scholar] [CrossRef]
- Georgas, K.M.; Armstrong, J.; Keast, J.R.; Larkins, C.E.; McHugh, K.M.; Southard-Smith, E.M.; Cohn, M.J.; Batourina, E.; Dan, H.; Schneider, K.; et al. An illustrated anatomical ontology of the developing mouse lower urogenital tract. Development 2015, 142, 1893–1908. [Google Scholar] [CrossRef]
- Huang, C.-C.; Orvis, G.D.; Kwan, K.M.; Behringer, R.R. Lhx1 is required in Müllerian duct epithelium for uterine development. Dev. Biol. 2014, 389, 124–136. [Google Scholar] [CrossRef]
- Zhao, F.; Zhou, J.; Li, R.; Dudley, E.A.; Ye, X. Novel function of LHFPL2 in female and male distal reproductive tract development. Sci. Rep. 2016, 6, 23037. [Google Scholar] [CrossRef]
- Robboy, S.J.; Kurita, T.; Baskin, L.; Cunha, G.R. New insights into human female reproductive tract development. Differentiation 2017, 97, 9–22. [Google Scholar] [CrossRef]
- Cunha, G.R.; Kurita, T.; Cao, M.; Shen, J.; Robboy, S.J.; Baskin, L. Response of xenografts of developing human female reproductive tracts to the synthetic estrogen, diethylstilbestrol. Differentiation 2017, 98, 35–54. [Google Scholar] [CrossRef] [PubMed]
- Acién, P.; Acién, M. Malformations of the female genital tract and embryological bases. Curr. Women’s Health Rev. 2007, 3, 248–288. [Google Scholar] [CrossRef]
- Greenfeld, H.; Lin, J.; Mullins, M.C. The BMP signaling gradient is interpreted through concentration thresholds in dorsal–ventral axial patterning. PLoS Biol. 2021, 19, e3001059. [Google Scholar] [CrossRef]
- Chan, S.W.; Rizwan, M.; Yim, E.K.F. Emerging methods for enhancing pluripotent stem cell expansion. Front. Cell Dev. Biol. 2020, 8, 70. [Google Scholar] [CrossRef]
- Bier, E.; De Robertis, E.M. BMP gradients: A paradigm for morphogen-mediated developmental patterning. Science 2015, 348, aaa5838. [Google Scholar] [CrossRef]
- Carreira, A.C.O.; Zambuzzi, W.F.; Rossi, M.C.; Filho, R.A.; Sogayar, M.C.; Granjeiro, J.M. Bone morphogenetic proteins. In Vitamins & Hormones; Elsevier: Amsterdam, The Netherlands, 2015; pp. 293–322. [Google Scholar]
- Kossack, M.E.; High, S.K.; Hopton, R.E.; Yan, Y.L.; Postlethwait, J.H.; Draper, B.W. Female sex development and reproductive duct formation depend on Wnt4a in zebrafish. Genetics 2019, 211, 219–233. [Google Scholar] [CrossRef]
- Méar, L.; Herr, M.; Fauconnier, A.; Pineau, C.; Vialard, F. Polymorphisms and endometriosis: a systematic review and meta-analyses. Hum. Reprod. Update 2020, 26, 73–102. [Google Scholar] [CrossRef]
- Edwards, T.L.; Giri, A.; Hellwege, J.N.; Hartmann, K.E.; Stewart, E.A.; Jeff, J.M.; Bray, M.J.; Pendergrass, S.A.; Torstenson, E.S.; Keaton, J.M.; et al. A trans-ethnic genome-wide association study of uterine fibroids. Front. Genet. 2019, 10, 511. [Google Scholar] [CrossRef]
- Gallagher, C.S.; Mäkinen, N.; Harris, H.R.; Rahmioglu, N.; Uimari, O.; Cook, J.P.; Shigesi, N.; Ferreira, T.; Velez-Edwards, D.R.; Edwards, T.L.; et al. Genome-wide association and epidemiological analyses reveal common genetic origins between uterine leiomyomata and endometriosis. Nat. Commun. 2019, 10, 4857. [Google Scholar] [CrossRef]
- Painter, J.N.; Anderson, C.A.; Nyholt, D.R.; Macgregor, S.; Lin, J.; Lee, S.H.; Lambert, A.; Zhao, Z.Z.; Roseman, F.; Guo, Q.; et al. Genome-wide association study identifies a locus at 7p15. 2 associated with endometriosis. Nat. Genet. 2011, 43, 51–54. [Google Scholar]
- Zhang, G.; Feenstra, B.; Bacelis, J.; Liu, X.; Muglia, L.M.; Juodakis, J.; Miller, D.E.; Litterman, N.; Jiang, P.P.; Russell, L.; et al. Genetic associations with gestational duration and spontaneous preterm birth. N. Engl. J. Med. 2017, 377, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Pitzer, L.M.; Moroney, M.R.; Nokoff, N.J.; Sikora, M.J. WNT4 balances development vs disease in gynecologic tissues and women’s health. Endocrinology 2021, 162, bqab093. [Google Scholar] [CrossRef]
- Gray, C.A.; Taylor, K.M.; Ramsey, W.S.; Hill, J.R.; Bazer, F.W.; Bartol, F.F.; Spencer, T.E. Endometrial glands are required for preimplantation conceptus elongation and survival. Biol. Reprod. 2001, 64, 1608–1613. [Google Scholar] [CrossRef] [PubMed]
- Gray, C.A.; Burghardt, R.C.; Johnson, G.A.; Bazer, F.W.; Spencer, T.E. Evidence that absence of endometrial gland secretions in uterine gland knockout ewes compromises conceptus survival and elongation. Reproduction 2002, 124, 289–300. [Google Scholar] [CrossRef]
- Spencer, T.E.; Dunlap, K.A.; Filant, J. Comparative developmental biology of the uterus: insights into mechanisms and developmental disruption. Mol. Cell. Endocrinol. 2012, 354, 34–53. [Google Scholar] [CrossRef]
- Cooke, P.S.; Spencer, T.E.; Bartol, F.F.; Hayashi, K. Uterine glands: development, function and experimental model systems. Mol. Hum. Reprod. 2013, 19, 547–558. [Google Scholar] [CrossRef]
- Kelleher, A.M.; Milano-Foster, J.; Behura, S.K.; Spencer, T.E. Uterine glands coordinate on-time embryo implantation and impact endometrial decidualization for pregnancy success. Nat. Commun. 2018, 9, 2435. [Google Scholar] [CrossRef]
- Kelleher, A.M.; Peng, W.; Pru, J.K.; Pru, C.A.; DeMayo, F.J.; Spencer, T.E. Forkhead box a2 (FOXA2) is essential for uterine function and fertility. Proc. Natl. Acad. Sci. USA 2017, 114, E1018–E1026. [Google Scholar] [CrossRef]
- Franco, H.L.; Dai, D.; Lee, K.Y.; Rubel, C.A.; Roop, D.; Boerboom, D.; et al. WNT4 is a key regulator of normal postnatal uterine development and progesterone signaling during embryo implantation and decidualization in the mouse. FASEB J. 2011, 25, 1176–1187. [Google Scholar] [CrossRef]
- Dunlap, K.A.; Filant, J.; Hayashi, K.; Rucker, E.B., 3rd; Song, G.; Deng, J.M.; Behringer, R.R.; DeMayo, F.J.; Lydon, J.; Jeong, J.W.; et al. Postnatal deletion of Wnt7a inhibits uterine gland morphogenesis and compromises adult fertility in mice. Biol. Reprod. 2011, 85, 386–396. [Google Scholar] [CrossRef]
- Reardon, S.N.; King, M.L.; MacLean, J.A., II; Mann, J.L.; DeMayo, F.J.; Lydon, J.P.; Hayashi, K. Cdh1 is essential for endometrial differentiation, gland development, and adult function in the mouse uterus. Biol. Reprod. 2012, 86, 141, 1–10. [Google Scholar] [CrossRef]
- Ballard, K.D.; Seaman, H.E.; De Vries, C.S.; Wright, J.T. Can symptomatology help in the diagnosis of endometriosis? Findings from a national case–control study—part 1. BJOG: Int. J. Obstet. Gynaecol. 2008, 115, 1382–1391. [Google Scholar]
- Pugsley, Z.; Ballard, K. Management of endometriosis in general practice: the pathway to diagnosis. Br. J. Gen. Pract. 2007, 57, 470–476. [Google Scholar] [PubMed]
- De Corte, P.; Klinghardt, M.; von Stockum, S.; Heinemann, K. Time to diagnose endometriosis: current status, challenges and regional characteristics—a systematic literature review. BJOG: Int. J. Obstet. Gynaecol. 2025, 132, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Signorile, P.G.; Dominici, S.; Viceconte, R.; Baldi, A. Prototype salivary assay for quantification of two biomarkers for in vitro diagnosis of endometriosis. Crit. Rev. Eukaryot. Gene Expr. 2024, 34, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Sampson, J.A. Peritoneal endometriosis due to menstrual dissemination of endometrial tissue into the peritoneal cavity. Am. J. Obstet. Gynecol. 1927, 14, 422–469. [Google Scholar] [CrossRef]
- Signorile, P.G.; Cassano, M.; Viceconte, R.; Spyrou, M.; Marcattilj, V.; Baldi, A. Endometriosis: a retrospective analysis on diagnostic data in a cohort of 4401 patients. In Vivo 2022, 36, 430–438. [Google Scholar] [CrossRef]
- Robboy, S.J.; Bean, S.M. Pathogenesis of endometriosis. Reprod. Biomed. Online 2010, 21, 4–5. [Google Scholar] [CrossRef]
- Wang, Y.; Nicholes, K.; Shi, I.M. The origin and pathogenesis of endometriosis. Annu. Rev. Pathol. 2020, 15, 71–95. [Google Scholar] [CrossRef]
- Dun, E.C.; Khom, K.A.; Morozov, V.V.; Kearney, S.; Zurawin, J.L.; Nezhat, C.H. Endometriosis in adolescents. In Nezhat, C.H., Ed. Endometriosis in Adolescents; Springer Nature: Berlin, Germany, 2020; pp. 129–141. [Google Scholar]
- Ricciardi, E.; Laganà, A.S.; Triolo, O.; Caserta, D. Epigenetic modifications of primordial reproductive tract: a common etiologic pathway for Mayer-Rokitansky-Kuster-Hauser Syndrome and endometriosis? Med. Hypotheses 2016, 90, 4–5. [Google Scholar]
- Gruenwald, P. Origin of endometriosis from the mesenchyme of the celomic walls. Am. J. Obstet. Gynecol. 1942, 44, 470–474. [Google Scholar] [CrossRef]
- Zondervan, K.T.; Becker, C.M.; Koga, K.; Missmer, S.A.; Taylor, R.N.; Viganò, P. Endometriosis. Nat. Rev. Dis. Prime 2018, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Salamonsen, L.A.; Hutchison, J.C.; Gargett, C.E. Cyclical endometrial repair and regeneration. Development 2021, 148, dev199577. [Google Scholar] [CrossRef] [PubMed]
- Parasar, P.; Sacha, C.R.; Ng, N.; McGuirk, E.R.; Chinthala, S.; Ozcan, P.; et al. Differentiating mouse embryonic stem cells express markers of human endometrium. Reprod. Biol. Endocrinol. 2017, 15, 52. [Google Scholar] [CrossRef]
- Figueira, P.G.; Abrao, M.S.; Krikun, G.; Taylor, H.S. Stem cells in endometrium and their role in the pathogenesis of endometriosis. Ann. N.Y. Acad. Sci. 2011, 1221, 10–17. [Google Scholar] [CrossRef]
- Klattig, J.; Englert, C. The mullerian duct: recent insight into its development and regression. Sex Dev. 2007, 1, 271–278. [Google Scholar] [CrossRef]
- Signorile, P.G.; Baldi, F.; Bussani, R.; D’Armiento, M.; De Falco, M.; Baldi, A. Ectopic endometrium in human foetuses is a common event and sustains the theory of müllerianosis in the pathogenesis of endometriosis, a disease that predisposes to cancer. J. Exp. Clin. Cancer Res. 2009, 28, 49. [Google Scholar] [CrossRef]
- Signorile, P.G.; Baldi, F.; Bussani, R.; D’Armiento, M.; De Falco, M.; Boccellino, M.; Quagliuolo, L.; Baldi, A. New evidence of the presence of endometriosis in the human fetus. Reprod. Biomed. Online 2010, 21, 142–147. [Google Scholar] [CrossRef]
- Signorile, P.G.; Baldi, F.; Bussani, R.; Viceconte, R.; Bulzomi, P.; D’Armiento, M.; D’Avino, A.; Baldi, A. Embryologic origin of endometriosis: analysis of 101 human female fetuses. J. Cell. Physiol. 2012, 227, 1653–1656. [Google Scholar] [CrossRef]
- Signorile, P.G.; Viceconte, R.; Baldi, A. New insights in pathogenesis of endometriosis. Front. Med. 2022, 9, 879015. [Google Scholar] [CrossRef]
- Bouquet de Jolinière, J.; Ayoubi, J.M.; Lesec, G.; Validire, P.; Goguin, A.; Gianaroli, L.; Dubuisson, J.B.; Feki, A.; Gogusev, J. Identification of displaced endometrial glands and embryonic duct remnants in female fetal reproductive tract: possible pathogenetic role in endometriotic and pelvic neoplastic processes. Front. Physiol. 2012, 3, 444. [Google Scholar] [CrossRef] [PubMed]
- Schuster, M.; Mackeen, D.A. Fetal endometriosis: a case report. Fertil. Steril. 2015, 103, 160–162. [Google Scholar] [CrossRef] [PubMed]
- Bhamidipaty-Pelosi, S.; Kyei-Barffour, I.; Volpert, M.; O’Neill, N.; Grimshaw, A.; Eriksson, L.; Vash-Margita, A.; Pelosi, E. Mullerian anomalies and endometriosis: associations and phenotypic variations. Reprod. Biol. Endocrinol. 2024, 22, 157. [Google Scholar] [CrossRef]
- Pitot, M.A.; Bookwalter, C.A.; Dudiak, K.M. Müllerian duct anomalies coincident with endometriosis: a review. Abdom. Radiol. 2020, 45, 1723–1740. [Google Scholar] [CrossRef]
- Crispi, S.; Piccolo, M.T.; D’Avino, A.; Donizetti, A.; Viceconte, R.; Spyrou, M.; Calogero, R.A.; Baldi, A.; Signorile, P.G. Transcriptional profiling of endometriosis tissues identifies genes related to organogenesis defects. J. Cell. Physiol. 2013, 228, 1927–1934. [Google Scholar] [CrossRef]
- Signorile, P.G.; Severino, A.; Santoro, M.; Spyrou, M.; Viceconte, R.; Baldi, A. Methylation analysis of HOXA10 regulatory elements in patients with endometriosis. BMC Res. Notes 2018, 11, 722. [Google Scholar] [CrossRef]
- Newbold, R.R.; Jefferson, W.J.; Padilla-Banks, E. Prenatal exposure to bisphenol A at environmentally relevant doses adversely affects the murine female reproductive tract later in life. Environ. Health Perspect. 2009, 117, 879–885. [Google Scholar] [CrossRef]
- Signorile, P.G.; Spugnini, E.P.; Mita, L.; Mellone, P.; D’Avino, A.; Bianco, M.; Diano, N.; Caputo, L.; Rea, F.; Viceconte, R.; et al. Prenatal exposure of mice to bisphenol A elicits an endometriosis-like phenotype in female offspring. Gen. Comp. Endocrinol. 2010, 168, 318–325. [Google Scholar] [CrossRef]
- Signorile, P.G.; Spugnini, E.P.; Citro, G.; Viceconte, R.; Vincenzi, B.; Baldi, F.; Baldi, A. Endocrine disruptors in utero cause ovarian damages linked to endometriosis. Front. Biosci. (Elite Ed.) 2012, 4, 1724–1730. [Google Scholar] [CrossRef]
- Signorile, P.G.; Baldi, A.; Viceconte, R.; Vincenzi, B.; Montella, M. Adenogenesis factors FGF7, FGF10, FGF23, IFN-τ and HGF in endometriosis tissue respect to eutopic endometrium: an immunohistochemical study. Crit. Rev. Eukaryot. Gene Expr. 2023, 33, 85–94. [Google Scholar] [CrossRef]
- Signorile, P.G.; Viceconte, R.; Vincenzi, B.; Baldi, A. Differential expression in endometriosis tissue versus endometrium of the uterine adenogenesis factors PRL-R, GH, IGF1, and IGF2. Crit. Rev. Eukaryot. Gene Expr. 2023, 33, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Signorile, P.G.; Baldi, A.; Viceconte, R.; Carraturo, E.; Boccellino, M.; Fordellone, M.; Montella, M. Glycosaminoglycan adenogenesis factors: immunohistochemical expression in endometriosis tissues compared with the endometrium. Crit. Rev. Eukaryot. Gene Expr. 2025, 35, 67–79. [Google Scholar] [CrossRef]
Figure 1.
Uterine formation.
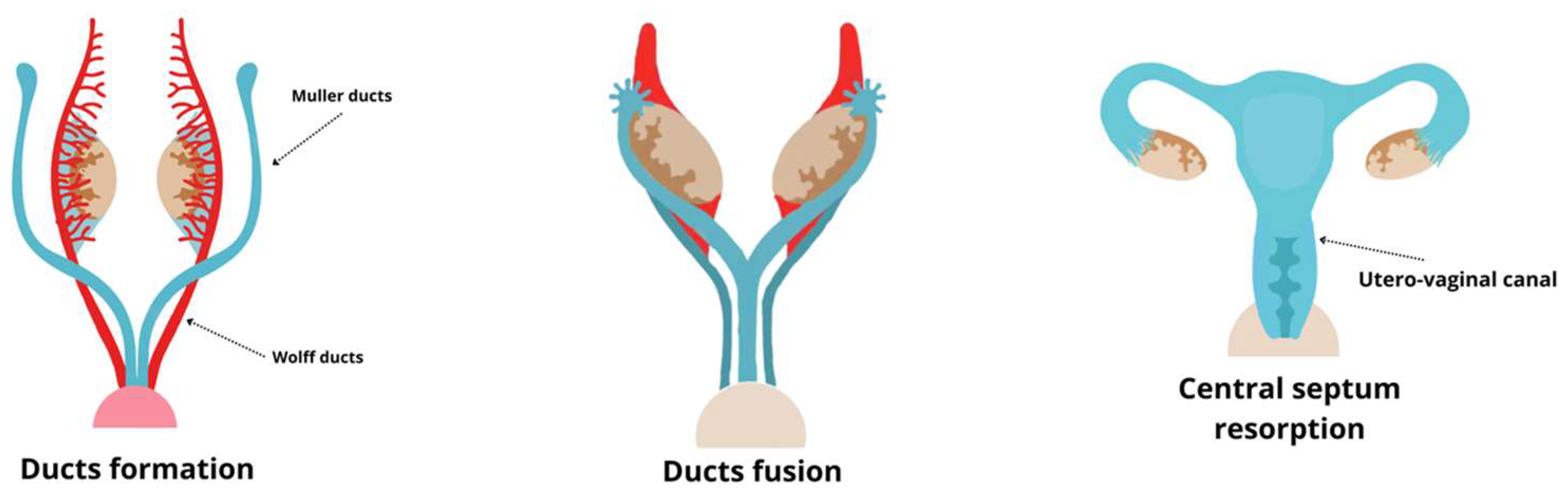

Table 1.
Major theories on the origin of endometriosis.
| Theory | Proposed Mechanism | Reference |
|---|---|---|
| Retrograde Menstruation | Retrograde menstrual flow allows the implantation of endometrial tissue in the peritoneal cavity. |
[47] |
| Coelomic Metaplasia Vascular dissemination Stem cell involvement Embryonic remnants |
Structures derived from the coelomic epithelium may undergo transformation into endometriotic tissue via metaplasia. Endometrial cells spread through blood or lymphatic vessels, colonizing distant anatomical sites. Endometrial or hematopoietic stem cells may differentiate into endometriotic lesions in ectopic locations. Residual embryonic cells from Wolffian or Müllerian ducts may develop into endometriotic lesions under estrogen stimulation. |
[53] [50] [57] [59,60,61,62,63] |
Table 2.
Immunohistochemical comparison between eutopic and ectopic endometrium.
| Marker | Eutopic Epithelium/Stroma | Ectopic Epithelium/Stroma |
|---|---|---|
| FGF7 FGF10 FGF23 IFN-τ HGF PRL-R GH IGF1 IGF2 CSPG4 CS-56 HEP Keratan Hyaluronic acid |
High/High High/High HighLow High/Low High/High High/Low Low/Low High/High High/High High/Low High/Low High/Low Low/Low Low/Low |
Low/Low Low/Low High/High High/High Low/Low Low/Low High/Low LowLow Low/Low High/Low High/Low High/Low Low/Low Low/Low |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



